
ojo biolo uda
20
Mecha nosign aling to the Cell Nucleus and Gene Regulation G.V. Shivashankar Mechanobiology Institute & Department of Biological Sciences, National University of Singapore, 117543 Singapore; email: [email protected] Annu. Rev. Biophys. 2011. 40:361–78 First published online as a Review in Advance on March 8, 2011 The Annual Review of Biophysics is online at biophys.annualreviews.org This article’s doi: 10.1146/annurev-biophys-042910-155319 Copyright c 2011 by Annual Reviews. All rights reserved 1936-122X/11/0609-0361$20.00 Keywords physicochemical signals, cytoplasmic-nuclear links, nuclear architecture, chromosome assembly, transcription compartments Abstract Cells integrate physicochemical signals on the nanoscale from the lo- calmicroe nvi ron men t, resulti ng in altere d functi ona l nuc lear lan dsc ape and gene expression. These alterations regulate diverse biological pro- cesses including stem cell differentiation, establishing robust develop- men tal gen etic pro gra ms and cell ular homeostatic con tro l sys tems. The mechanisms by which these signals are integrated into the 3D spa- tiotemporal organiz ation of the cell nucleus to elicit differ ential gene expression programs are poorly understood. In this review I analyze our current understanding of mechanosignal transduction mechanisms to the cell nucleus to induce differential gene regulation. A descrip- tion of both physical and chemical coupling, resulting in a prestressed nuclear organization, is emphasized. I also highlight the importance of spatial dimension in chromosome assembly, as well as the temporal filtering and stochastic processes at gene promoters that may be im- portant in understanding the biophysical design principles underlying mechanoregulation of gene transcription. 361 A n n u . R e v . B i o p h y s . 2 0 1 1 . 4 0 : 3 6 1 - 3 7 8 . D o w n l o a d e d f r o m w w w . a n n u a l r e v i e w s . o r g b y P o n t i f i c i a U n i v e r s i d a d J a v e r i a n a o n 1 0 / 1 8 / 1 1 . F o r p e r s o n a l u s e o n l y . Click here for quick links to Annual Reviews content online, including: • Other articles in this volume • Top cited articles • Top downloaded articles • Our comprehensive search Further ANNUAL REVIEWS
-
Upload
miltonlafebre -
Category
Documents
-
view
226 -
download
0
Transcript of ojo biolo uda

8/3/2019 ojo biolo uda
http://slidepdf.com/reader/full/ojo-biolo-uda 1/20
Mechanosignaling to the CeNucleus and Gene Regulatio
G.V. Shivashankar
Mechanobiology Institute & Department of Biological Sciences, National UniversitSingapore, 117543 Singapore; email: [email protected]
Annu. Rev. Biophys. 2011. 40:361–78
First published online as a Review in Advance on March 8, 2011
The Annual Review of Biophysics is online at
biophys.annualreviews.org
This article’s doi:10.1146/annurev-biophys-042910-155319
Copyright c 2011 by Annual Reviews. All rights reserved
1936-122X/11/0609-0361$20.00
Keywords
physicochemical signals, cytoplasmic-nuclear links, nucleararchitecture, chromosome assembly, transcription compartments
Abstract Cells integrate physicochemical signals on the nanoscale from th
cal microenvironment, resulting in altered functional nuclear landsand gene expression. These alterations regulate diverse biological
cesses including stem cell differentiation, establishing robust dev
mental genetic programs and cellular homeostatic control systemsmechanisms by which these signals are integrated into the 3D
tiotemporal organization of the cell nucleus to elicit differential expression programs are poorly understood. In this review I an
our current understanding of mechanosignal transduction mechanto the cell nucleus to induce differential gene regulation. A des
tion of both physical and chemical coupling, resulting in a prestrnuclear organization, is emphasized. I also highlight the import
of spatial dimension in chromosome assembly, as well as the tempfiltering and stochastic processes at gene promoters that may be
portant in understanding the biophysical design principles under
mechanoregulation of gene transcription.
361
Click here for quick links to
Annual Reviews content online,
including:
• Other articles in this volume
• Top cited articles
• Top downloaded articles
• Our comprehensive search
FurtherANNUAL
REVIEWS

8/3/2019 ojo biolo uda
http://slidepdf.com/reader/full/ojo-biolo-uda 2/20
Contents
INTRODUCTION . . . . . . . . . . . . . . . . . . 362
MECHANOSIGNALGENERATION FOR GENE
REG U LAT IO N . . . . . . . . . . . . . . . . . . . 3 6 3CHEMICAL TRANSDUCTION:
TRANSCRIPTION FACTORS AS MOLECULAR
INTERMEDIATES.. . . . . . . . . . . . . . 364PHYSICAL TRANSDUCTION OF
MECHANOSIGNALS VIA CYTOPLASMIC-NUCLEAR
L I N K S . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 6 5SPATIAL DIMENSION TO
NUCLEAR ORGANIZATION . . . 366
TEMPORAL DYNAMICS AND FILTERING
BY TRANSCRIPTIONF A C T O R I E S . . . . . . . . . . . . . . . . . . . . . . 3 6 7
MECHANICAL ACCESSIBILITY AND CHEMICAL SPECIFICITY
FOR GENE TRANSCRIPTIONIN LIVING CELLS: PUTTING
THE PIECES TOGETHER . . . . . . 370IMPLICATIONS . . . . . . . . . . . . . . . . . . . . 372
INTRODUCTION
Biological cells are mechanical in origin andsense their local microenvironment using cell
surface receptors (31). In addition to soluble
signals, cells are sensitive to extracellular matrixrigidity, to the geometry or local curvature
of the matrix, and to the applied stress orstrain (119). These physical signals are an
integral component of cellular behavior duringdifferentiation (24), during development (1,
11), or in disease phenotypes (45). A variety of molecular intermediates describing cell matrix
interactions and the remodeling of cytoskeletalnetworks in response to extracellular matrix
signals have now been elucidated. Although thecytoskeleton is a well-appreciated critical com-
ponent of cellular morphology (28), emerging
evidence suggests that it may also haveimportant consequences for maintenance of
functional nuclear architecture and its mechaical properties (23, 121). Work from a numb
of laboratories, including mine, is beginning suggest an elaborate physicochemical netwo
of protein assemblies coupling the cytoskelettothenucleus(71,110).Thiscouplingresults
a prestressed nuclear organization in living ce
that balances contractile forces of thecytoskelton and condensation forces of the chromat
(75–77). In addition, the prestressed nuclear achitecture could perhaps serve as a substrate f
transducing mechanical signals to the nucleu The regulation of gene expression
response to cellular matrix and geometric curequires mechanisms that act at a distance (1
120, 121). A number of canonical signalipathways are activated in response to su
mechanical signals converging on transcriptiofactors—such as NFκB—that translocate
the nucleus upon activation (79, 126). Althou
these soluble factors translocate to the nucle via diffusive processes, recent evidence a
highlights the physical transmission of actistresses via cytoplasmic-nuclear connectio
to remodel chromatin assembly (121). Tphysicochemical signals targeted to the nucle
have to be further sorted to appropriate prmoter sequences within the 3D architectu
of the cell nucleus to bring about changes gene expression programs (22, 61). Althou
the location of promoter sequences along t1D contour of the DNA polymer is know
from genome sequencing, their 3D locatio
when folded into chromatin via histone anonhistone proteins within the nucleus
largely unknown (83). Further, a number essential posttranslational modifications
histone proteins (50) result in specificity anaccessibility of promoter sequences to initia
gene transcription. The regulation of gene expression program
depends on genome organization, posttranlational modifications of histone proteins, an
assembly of transcription apparatus at ge
promoters. Recent work in the literature hshown that genome organization is nonrando
(21) and perhaps specific to cell type (93). Wofrom several laboratories, including mine,
362 Shivashankar

8/3/2019 ojo biolo uda
http://slidepdf.com/reader/full/ojo-biolo-uda 3/20
beginning to show that chromosomes arehighly plastic in stem cells (4, 74, 81, 92) and
that gene-rich chromosomes in differentiatedcells may be spatially clustered (29) and are
correlated with the transcriptome network of the cell (33). In addition, gene transcription
requires transcription machinery enriched in
dynamic compartments within the eukaryoticcell nucleus (113). The spatiotemporal assem-
bly of transcription apparatus onto promotersites, bound with the appropriate initiation
signals derived through cellular mechanicalcues, would finally result in the desired genetic
outputs. However, it remains to be seen howsuch spatially coded maps of chromosome
positioning in the nucleus and dynamic tran-scription machines enable efficient sorting
of mechanical signals and initiate gene tran-scription in a highly regulated manner. The
following sections elaborate on the modularity
in mechanoregulation of gene transcription, asoutlined above in eukaryotic cells.
MECHANOSIGNALGENERATION FOR GENE REGULATION
Cells have evolved a plethora of sensing mech-
anisms in their local microenvironment. Suchsensing could be physical and/or chemical in
nature (24, 66, 116, 122). For example, whensoluble factors such as EGF bind to EGF
receptors, downstream signaling cascades im-
pinge on gene expression. More importantly,the fact that cells can detect their local mechan-
ical environment physically suggests that theremust be distinct modularity in cellular sensing
mechanisms to sample local geometry or sub-strate rigidity to which cells are adhered (31,
119). In addition, cells discriminate shear fromcompressive forces (62) and the direction they
are applied from. Physical sensing modes areelicited, for example, through either stretch-
activated receptors or integrin-like transmem-
brane receptors that attach to extracellular ma-trix proteins (53, 85), thereby sensing local
rigidity of the matrix at the focal adhesion sitesto transduce signals to the nucleus. Further, a
numberofothermolecularsensingmechanisms via receptor clustering are also being uncov-
ered. These physicochemical signals integrateto alter cell behavior and function by modulat-
ing the cytoskeleton and gene expression—thetheme of this review. Mechanisms that regulate
the assembly of focal adhesion complexes me-
diated via the actomyosin machinery and thesubsequent activation of mechanosensing pro-
teins that impinge on cytoskeleton remodeling(48, 66, 116) and alter diverse cellular behav-
iors, including transmigration (18), are beyondthe scope of this review. The reader is directed
to recent comprehensive reviews of this aspectof mechanosignaling (31, 49, 94, 119).
Stem cells sample both physical and chem-ical signals, leading them to differentiate into
multiple cell types, suggesting that modules of mechanosensing mechanisms used to alter gene
expression programs are functional in these
cells (17, 26, 55). Interestingly, lineage-specifictranscription programs stabilize these differen-
tiating cells. The cells are optimized to sensematrix in the context of epithelial cells or to
sense soluble signals in the context of blood lin-eages. In the context of dedifferentiation (70,
128), reprogramming fibroblasts, for example,to a pluripotent cell or to a blood cell lin-
eage suggests that modules of mechanosens-ing via feedback from the local microenviron-
ment must be conserved, whereas specialized(differentiated) cells have the ability to use ei-
ther one or the other mechanosensing module
in a context-dependent manner.Cellular mechanosensing at the plasma
membrane has to be transduced to the nucleus,a few microns away from sensing sites in mam-
malian systems, to elicit differential gene ex-pression programs. Typical diffusion constants
of membrane-bound proteins are in the rangeof less than 1 μ m−2 s−1—perhaps determining
the timescales in early events in mechanosens-ing. Transduction of mechanosignals from
the inner leaflet of the plasma membrane viaa number of protein conformational switches
to phosphorylate buried amino acid residues
and thereby transduce second-messengercalcium flux transport occurs in milliseconds to
www.annualreviews.org • Mechanosignaling to the Cell Nucleus and Gene Regulation 363

8/3/2019 ojo biolo uda
http://slidepdf.com/reader/full/ojo-biolo-uda 4/20
seconds. Although these signals are generated within seconds to minutes, the early gene
transcriptional events are detected only aftera few minutes, providing a temporal window
on gene regulatory processes. Matrix-relatedgenes are upregulated in tens of minutes (16,
120), whereas stem-cell-specific genes are
modulated with matrix or geometric cues (26,55) in a few hours to days, suggesting the need
for integrating sustained versus short-timeinput signals required for mechanoregulation
of gene expression. In the next sections Iexplore how input mechanical signals result in
both physical remodeling of cytoskeleton andnucleus and transfer of signal intermediates via
distinct canonical signaling pathways, such as MAPK/ERK or JAK-STAT (10).
CHEMICAL TRANSDUCTION: TRANSCRIPTION FACTORS AS MOLECULAR INTERMEDIATES
The transduction of mechanical signals even-tually converges on transcription factors and
cofactors that bind to regulatory DNA sitesto determine which genes are upregulated or
downregulated. For this, proteins activated,for example, via phosphorylation by kinases at
the inner leaflet of the plasma membrane aretransmitted either directly to the nucleus or by
secondary mechanisms. Upon application of mechanical force, proteins such as paxillin (an
essential linker betweenintegrins andactin) and
zyxin, which are usually resident at focal adhe-sion complexes, shuttle between the cytoplasm
and nucleus (123). Although these proteinscontain the Lim domain (52), which is involved
in diverse cellular processes including cell ad-hesion and transcription control, the functional
roles of these focal adhesion–resident proteinsin the nucleus remain to be elucidated. Simi-
larly, mechanosignal-induced phosphorylationof β-catenin bound to E-cadherins, present
at the cell-cell junctions, results in β-catenin
translocation into the nucleus (124). Whereas some proteins can be translocated
directly fromfocaladhesion or cell-cell junctionto the nucleus, veryoften downstream signaling
processes activate transcription-relevant fators either in the cytoplasm or in the nucleu
For example, activation of Notch recepto which are transmembrane proteins, results
proteolytic cleavage of the intracellular Notdomain, which is then targeted to the nucle
(42). In contrast, transcription factors such
NFκ
B (114), STAT3 (10), and NFAT (40) thare sequestered in the cytoplasm upon a pho
phorylation step translocate to the nucleuOther examples such as MAL (118), which
bound to actin in the cytoplasm, are transferrfrom F-actin to G-actin, resulting in th
translocation to the nucleus. Formins (8nucleators for actin polymerization, exhib
cytoplasmic to nuclear shuttling. In generalthough transcription factors and cofacto
are activated in the cytoplasm upon stimulati
and then translocated to the nucleus, there iclass of transcription-relevant factors th
reside in the nucleus. These factors are presein an inactive form, but when stimulated, th
target specific regulatory sites on the DNFor example, EGR1 (67), a transcription fact
stimulated by serum-responsive factors, activated in the nucleus, resulting in binding
DNA target sites to regulate gene expressioincluding that of matrix-related genes.
Most signals targeted to the nucleus shoucontain nuclear localization signals (NLS) th
are then recognized by the nuclear import
machinery. The sequence of events that follotransduction of chemical signals to the nucle
to regulate gene expression is highly variabFor example, transcription factors and cofacto
bind regulatory sites and then further recruicomplex assembly of transcription machines
regulate gene expression (56). In the cytoplas whether some proteins translocate direc
to the nucleus or whether others requisecondary and tertiary relay mechanisms upo
mechanical stimulation is currently uncleHowever, early-response genes use a modula
ity in which transcriptional cues are target
directly to the nucleus, whereas genes thencode for transcription factors and cofacto
and thereby regulate a whole set of additiongenes often are connected with sequent
364 Shivashankar

8/3/2019 ojo biolo uda
http://slidepdf.com/reader/full/ojo-biolo-uda 5/20
relay mechanisms for signal transduction. Inaddition, most proteins that shuttle to the
nucleus are short-lived and are exported via thenuclear export signal (NES) that is recognized
by the nuclear export machinery (19) or they are degraded by ubiquitination. Proteins com-
posed of NLS or NES sequences are part of the
protein family that shuttles to and from the nu-cleus (39), with a typical turnover timescale of
a few minutes, suggesting kinetic mechanismsto actively access genetic information.
PHYSICAL TRANSDUCTION OF MECHANOSIGNALS VIA CYTOPLASMIC-NUCLEAR LINKS
Although chemical signals are targeted to the
nucleus via soluble transcription intermediates,recent evidence reveals transduction of active
stresses between thecytoplasm andnucleus thatare inherent in biological cells (44, 76, 77, 105).
The cytoskeleton, composed primarily of actin,
microtubules, and intermediate filaments, hasdynamic links to the nuclear membrane and
chromatin assembly (117). The actin filamentsthat originate at the focal adhesion points also
polymerize into long filaments (stress fibers)that have been implicated in transmitting
stress signals throughout the cytoskeleton(28, 89, 94) and perhaps to the nucleus. In
addition, microtubules that originate fromthe microtubule-organizing centers are highly
filamentous structures that provide structural
integrity to the cellular architecture (9) andmay act as cables to transmit stresses from a dis-
tance. Further, the intermediate filaments (38)that form a dense structure in the cytoplasm
provide a scaffold through which physicalcues could be transmitted. These cytoskeletal
filaments are highly cross-linked, providinga gel-like architecture to the cytoskeleton.
In addition, the cytoskeletal structures act assubstrates to sequester both soluble signaling
intermediates and tracks for transporting
signal-enriched vesicular cargos via the motorsthat run on them.
Recent work has revealed that the cytoskele-tal structures bind to a number of nuclear
membrane proteins (104, 110). In particular,the Sun and Kash domain proteins, residing
on the outer and inner nuclear membrane, re-spectively, connect to cytoskeleton on one side
and to nuclear lamina and chromatin assembly on the other side, bridging the internuclear
membrane space via physical links (36, 37).
Outer nuclear membrane proteins, such asNesprin (132), interact with actin, while
Klaroid binds to microtubules and plectin with intermediate filaments (117). The nuclear
morphology is therefore determined as a resultof these physical links (8, 100). For example,
the nuclear volume in an adherent cell ismuch larger than the nucleus devoid of the
cytoskeleton (76). Experiments have revealedthat inhibiting myosin motors by blebbistatin
or actin polymerization by cytochalasin D
results in a dramatic decrease in nuclear size of adherent cells, whereas inhibiting microtubule
polymerization by nocodazole results in anincrease in nuclear size (77). These results
suggest that the eukaryotic cell nucleus is heldunder tensile load by the actomyosin links,
whereas the microtubules offer a compressiveloading on the nucleus, suggesting a dynamic
prestressed nuclear organization. The dynamicloading is further balanced by the intermediate
filaments, and the degree of nuclear prestressis fine-tuned depending on cellular adhesion,
with increased nuclear prestress in adherent
cells. The degree of nuclear prestress has anumber of important biological consequences.
For example, it determines (a) the plasticity of the stem cell nucleus, (b) the ability of the
nucleus to rotate, (c ) the cytoplasmic-nuclearlinks mediating cell migration, and (d ) the
degree of mechanical unfolding of individualproteins or proteinaceous scaffolds including
chromatin assembly and its remodeling.In stem cells, the reduced cytoplasmic-
to-nuclear ratio results in the lack of nu-clear prestress and thus a soft, highly plas-
tic, and mechanically pliable cell nucleus (4,
92). The emergence of cell differentiationprograms leads to the dynamic reorganiza-
tion of cytoskeleton to nuclear membranes,progressively leading to a stiffer nucleus as
www.annualreviews.org • Mechanosignaling to the Cell Nucleus and Gene Regulation 365

8/3/2019 ojo biolo uda
http://slidepdf.com/reader/full/ojo-biolo-uda 6/20
lineage-committed cells emerge (77, 92). Onthe other hand, the proximity of microtubule
filaments to the nuclear membrane causes therotation of the nucleus by dynein motors (64).
These nuclear rotations are influenced by ge-ometric constraints imposed by cytoplasmic-
nuclear links. While in interphase cells, dynein
motors bind to nuclear membrane proteins;their binding is controlled by the organization
of microtubules, such that the nuclear rota-tions are highly regulated. Although these geo-
metric constraints are far from clear, evidenceof nuclear rotation suggests dynamic links be-
tween the cytoskeleton and the nucleus. Theselinks are essential mechanical intermediates for
cell migration (69) and for defining nuclearmechanical integrity via the perinuclear actin
cap (54). In support of these links, a numberof molecules currently have been revealed by
monitoring the alterations in nuclear morphol-
ogy upon ablation of specific genes in imaging-based RNAi screens.
In the context of this review, the cytoskeletalfilaments that provide nuclear prestress might
transmit active stresses from the plasma mem-brane to the nuclear membrane. This transduc-
tion mechanism has two functions. First, it actsas a mechanosensor with which to unfold pro-
teins (51) linked to the cytoskeleton for phos-phorylation events. Second, it may provide a
means of unfolding chromatin assembly from adistance. The timescales of these force-induced
unfolding processes depend directly on the di-
rect transmission of stresses or stress waves lim-ited by the acoustic transmission rates (121).
The ability to alter chromatin structure from adistance may influence chromatin-remodeling
enzymes (e.g., histone acetylase transferasesand histone deacetylase transferases) by shifting
their dynamic equilibrium toward the boundor unbound forms, which eventually remodels
chromatin structure to regulate gene expres-sion. In addition, mechanical links to the nu-
cleus could modulate theopeningand closing of
nuclear pores (111). This could provide mech-anisms to enhance the transduction of soluble
chemical intermediates via the nuclear pores ina ligand-dependentnuclear translocation that is
influenced by secondary forces exerted by tcytoplasm on the prestressed nucleus. The
processes, however, would require highly reulated control of cytoskeletal structure and d
namics within living cells via Rho GTPases (2and other signaling intermediates. In the ne
section I describe how these physiochemic
signals may be integrated into the 3D architeture of the nucleus and transcription process
SPATIAL DIMENSION TO NUCLEAR ORGANIZATION
Mechanochemical signals that integrate inthe nucleus have to be sorted within nucle
3D architecture to elicit gene expression. Teukaryotic nucleus is a highly crowded env
ronment, and its 3D architecture is determinby compaction of DNA (2), which has a typic
contour length of ∼1 m. Double-strandDNA is a good example of a flexible polym
chain with a persistence length of ∼50 nm,
revealed in DNA micromanipulation experments (13). For this polymer chain, the typic
radius of gyration is ∼200 μ m—the size of tentropic polymer configuration. This entrop
coil is compacted within the cell nucleus (72about 10 to 50 μ m in diameter, dependin
on the cell type. For compaction, histoand nonhistone proteins bind to the DN
stabilized by electrostatic interactions, formia nucleosomal array 11 nm in diameter, an
are folded into a dynamic 30-nm chromat
fiber by the interaction of linker histoproteins (3, 30, 68). Further packaging in
higher-order chromatin structures involvforming condensed DNA structures call
the heterochromatin structure (35), whiis stabilized by heterochromatin bindi
proteins and noncoding RNA (131) and loosepacked euchromatin structure (99, 125). Th
differential packaging is highly regulated a number of epigenetic modifications on th
amino acid residues of histone and nonhisto
proteins and their variants (50, 101). Fexample, increased acetylation of core histo
amino acid tail residues results in more opand flexible chromatin structure; alternative
366 Shivashankar

8/3/2019 ojo biolo uda
http://slidepdf.com/reader/full/ojo-biolo-uda 7/20
methylation of amino acid residues of corehistone proteins facilitates the binding of
heterochromatin binding proteins to formcondensed heterochromatin structure. A 1-m-
long 1D sequence of DNA in any given chro-mosome is thus condensed into a chromosome
∼1–2 μ m in diameter in an interphase nucleus.
In addition to packaging DNA into a chro-mosome, the next level of packaging involves
positioning chromosomes (for example, 23pairs of chromosomes in human cells) within
the nucleus (5). Because the contour lengthand folding fraction of each chromosome are
different, their eventual volumes are variable.In effect, if each chromosome is treated as
a flexible particle (sizes vary within a fewmicrons), 40 such particles must be assembled
within the cell nucleus. A number of scaffoldproteins, comprising a family of intermediate
filaments (lamin proteins) residing in the
nucleus, form a structural scaffold to anchorchromosomes impinging on gene expression
(36, 73, 104, 107). In particular, lamin proteinsare organized as a sheet-like structure anchored
to the inner nuclear membrane proteins andto condensed heterochromatin structures via
the lamin receptor proteins. However, duringmitosis, with the breakdown of the structural
scaffolds, chromosomes are further condensedinto metaphase chromosomes that regain their
excluded volume and perhaps spatial position-ing in the two daughter nuclei (7, 41). The
nuclei of interphase cells (Figure 1) therefore
have to balance the outward entropic forces of the DNA fiber and the inward condensation
forces of a metaphase chromosome, imposingmechanical constraints on the nucleus (75–77).
Recent experiments have revealed a nonran-dom positioning of chromosomes within the
nucleus (5, 25). Therefore, the arrangement of 40 chromosomes in mouse cells or 46 chro-
mosomes in human cells requires spatial posi-tioning mechanisms within the 3D architecture
of the nucleus. These mechanisms are further
augmented by the fact that interphase chromo-somes, depending on their size, appear to have
preferred 3D radial positions when mappedfrom the center of the nucleus (87, 112). For
example, gene-inactive chromosomes appear tobe located in the nuclear periphery, whereas
gene-active chromosomes arelocatedin thenu-clear interior in the same cell type (33, 96).
Evidence also suggests that cells from differenttissues may comprise distinct cell-type-specific
positional ordering of chromosomes (93). Such
positional coding requires physical anchoringof chromosomes by chromosome-chromosome
interactions, anchoring to nuclear scaffold pro-teins, or both. Cytoplasmic-nuclear links and
nuclear scaffold proteins may act as a sub-strate to position chromosomes to their respec-
tive 3D positional coordinates in differentiatedcells (86). However, such structural integrity
is lacking in the stem cell nuclei, resulting intheir plasticityand enhanced collisionsbetween
chromosome interfaces, whereas well-defined
chromosome interfaces (Figure 2) areobservedin differentiated cells (78, 98). How such cell-
type-specific chromosome positioning and pre-cision in their positional inheritance through
mitosis are derived is still far from clear.
TEMPORAL DYNAMICS AND FILTERING BY TRANSCRIPTION FACTORIES
The spatial organization of chromosomes within the 3D architecture of the cell nucleus
imposes physical constraints to accessing of genetic information. A typical gene (∼2 kbp
in linear length) folded around histone and
nonhistone proteins is ∼50 nm in diameter. Whereas the position of genes on the linear
genome sequence is known, their location when folded into a 3D chromosome structure
is still unknown (83, 88). However, chro-mosome surfaces have a largely euchromatin
structure—composed of gene sequences—although there are a number of exceptions (32,
57). Thus, spatial positioning of chromosomes within the cell nucleus results in chromosome
interfaces rich in coding sequences. Studies
using chromosome capture assays have re- vealed that the interchromosome spacing or
territories may contain intermingled flexibleeuchromatin loops, allowing for easy access to
www.annualreviews.org • Mechanosignaling to the Cell Nucleus and Gene Regulation 367
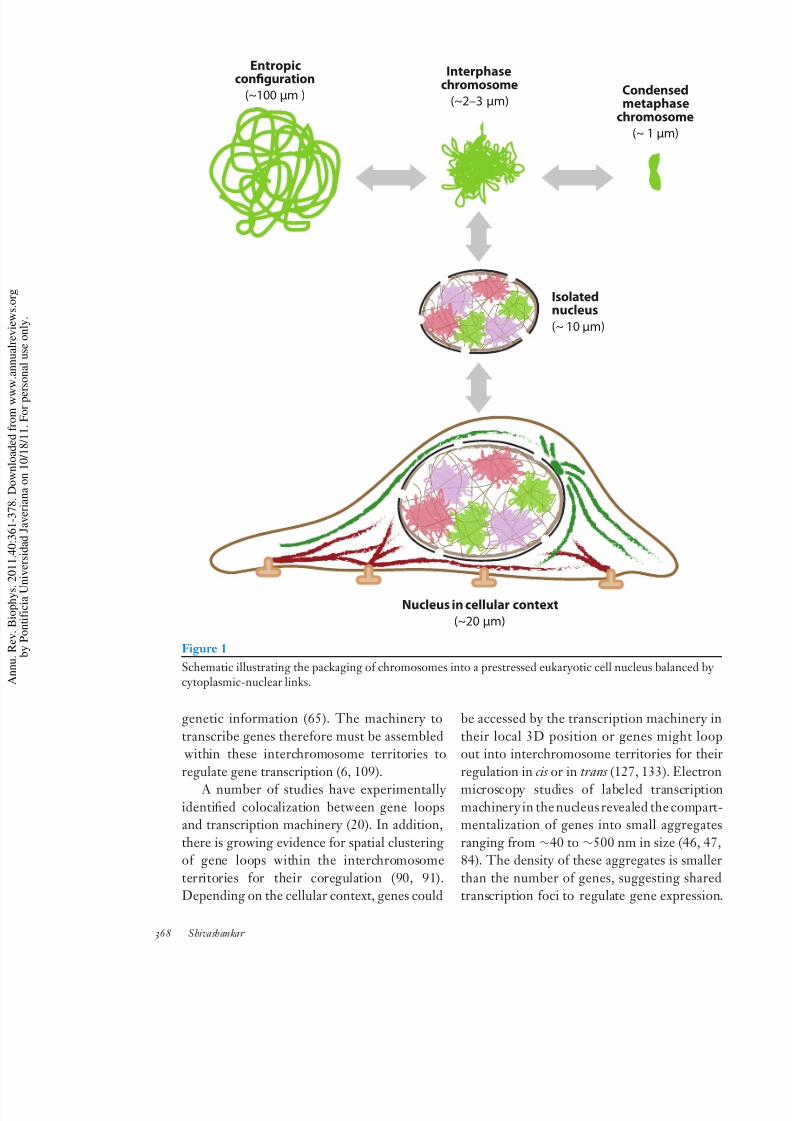
8/3/2019 ojo biolo uda
http://slidepdf.com/reader/full/ojo-biolo-uda 8/20
Entropicconfguration
(~100 µm ) Condensedmetaphase
chromosome
(~ 1 µm)
Isolatednucleus
(~ 10 µm)
Interphasechromosome
(~2–3 µm)
Nucleus in cellular context
(~20 µm)
Figure 1
Schematic illustrating the packaging of chromosomes into a prestressed eukaryotic cell nucleus balanced bcytoplasmic-nuclear links.
genetic information (65). The machinery to
transcribe genes therefore must be assembled within these interchromosome territories to
regulate gene transcription (6, 109).
A number of studies have experimentally identified colocalization between gene loopsand transcription machinery (20). In addition,
there is growing evidence for spatial clusteringof gene loops within the interchromosome
territories for their coregulation (90, 91).Depending on the cellular context, genes could
be accessed by the transcription machinery
their local 3D position or genes might loout into interchromosome territories for the
regulation in cis or in trans (127, 133). Electr
microscopy studies of labeled transcriptimachinery in the nucleus revealed the compamentalization of genes into small aggregat
ranging from ∼40 to ∼500 nm in size (46, 484). The density of these aggregates is small
than the number of genes, suggesting shartranscription foci to regulate gene expressio
368 Shivashankar
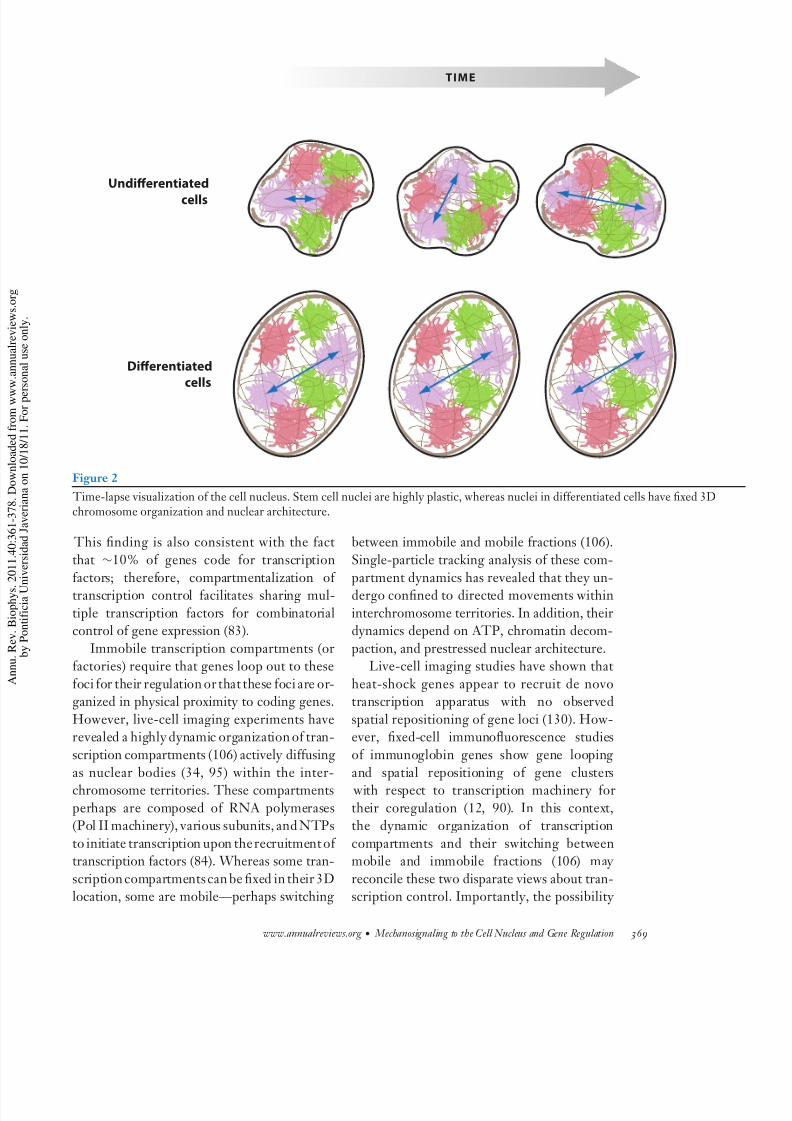
8/3/2019 ojo biolo uda
http://slidepdf.com/reader/full/ojo-biolo-uda 9/20
Undierentiated
cells
Dierentiated
cells
TIME
Figure 2
Time-lapse visualization of the cell nucleus. Stem cell nuclei are highly plastic, whereas nuclei in differentiated cells have fixed 3Dchromosome organization and nuclear architecture.
This finding is also consistent with the fact
that ∼10% of genes code for transcription
factors; therefore, compartmentalization of transcription control facilitates sharing mul-
tiple transcription factors for combinatorialcontrol of gene expression (83).
Immobile transcription compartments (orfactories) require that genes loop out to these
foci for their regulation or that these foci are or-ganized in physical proximity to coding genes.
However, live-cell imaging experiments haverevealed a highly dynamic organization of tran-
scription compartments (106) actively diffusing
as nuclear bodies (34, 95) within the inter-chromosome territories. These compartments
perhaps are composed of RNA polymerases(Pol II machinery), various subunits, and NTPs
to initiate transcription upon therecruitment of transcription factors (84). Whereas some tran-
scription compartments can be fixed in their 3Dlocation, some are mobile—perhaps switching
between immobile and mobile fractions (106).
Single-particle tracking analysis of these com-
partment dynamics has revealed that they un-dergo confined to directed movements within
interchromosome territories. In addition, theirdynamics depend on ATP, chromatin decom-
paction, and prestressed nuclear architecture.Live-cell imaging studies have shown that
heat-shock genes appear to recruit de novotranscription apparatus with no observed
spatial repositioning of gene loci (130). How-ever, fixed-cell immunofluorescence studies
of immunoglobin genes show gene looping
and spatial repositioning of gene clusters with respect to transcription machinery for
their coregulation (12, 90). In this context,the dynamic organization of transcription
compartments and their switching betweenmobile and immobile fractions (106) may
reconcile these two disparate views about tran-scription control. Importantly, the possibility
www.annualreviews.org • Mechanosignaling to the Cell Nucleus and Gene Regulation 369

8/3/2019 ojo biolo uda
http://slidepdf.com/reader/full/ojo-biolo-uda 10/20
of a dynamic conversion between immobileand mobile fractions of transcription foci could
indeed provide a temporal mechanical filteringto access gene regulatory sites within the 3D
architecture of the nucleus.
MECHANICAL ACCESSIBILITY
AND CHEMICAL SPECIFICITY FOR GENE TRANSCRIPTION IN LIVING CELLS: PUTTING THEPIECES TOGETHER Biochemical and structural evidence for tran-
scription initiation at the gene promoter hasrevealed important elements of eukaryotictran-
scription control (56). In summary, context-dependent transcription factors are targeted to
promoter sites, resulting in remodeling of localchromatin structure at theregulatory sites. This
follows the recruitment of transcription ma-chinery, dynamic removal of the core histones
by the chromatin-remodeling machinery in thecoding region, and an efficient repacking of
the local DNAstructure incorporating histones
and histone variants marking the transcriptionstate. However, these processes are highly dy-
namic, and our understanding of how this isinitiated in the context of a living cell nucleus
is just beginning. In addition, we know very lit-tle about the spatiotemporal ordering of these
events. However, the posttranslational mod-ifications of core histones, primarily histone
acetylation induced via histone acetyl trans-ferases to loosen chromatin structure at the
promoter sites and the recruitment and core-
cruitment of transcription factors, chromatinremodeling, and transcription machinery, are
a necessary step to regulate gene transcription. There are a number of examples in which this
recruitment and initiation are regulated via anenhancer sequence and its binding protein, re-
sulting in a distal chromatin looping of a fewhundred bases to a few kilobase pairs between
thepromoter region andthe enhancer region toregulate gene transcription (63). The geomet-
ric scale of this looping ranges from a few tens
to hundreds of nanometers at any given spatialposition of the gene within the 3D architecture
of the nucleus.
How then do mechanical signals that aexerted on the cell membrane, a few micro
away, influence gene transcription within thighly crowded nuclear environment? Ho
does the transcriptional apparatus recruit agiven gene in a context-dependent manne
Although the chemical signals generated at t
plasma membrane are translocated via the trascription factor intermediates, diffusing eith
passively or actively in search of their regultory sites within the 3D nuclear architectur
the physical integration via the cytoplasminuclear links perhaps modulates exposure
buried regulatory sequences to facilitate tsearch process of these promoter sites f
dynamic recruitment of the transcriptional aparatus. The modularity in gene transcriptio
and transcription-dependent compartme
talization within the nucleus perhaps alprovides a mechanical regulation of the d
namic movement of transcription machinethrough the interchromosome separation v
the modulation of prestressed cytoplasminuclear links. In contrast, the plasticity
chromatin organization and nuclear architeture in stem cells (58, 80, 108) may provide
means to enhance interactions between distinchromosome locations for gene clustering an
their coregulation for lineage commitment. There is a need to reconcile a number
facts about gene transcription regulation. F
example, while all the regulatory machineryin place, the positioning of the gene within th
nucleus also appears to play a critical role in reulating its expression (60). When all the reg
latory signaling modules are similar, and whthey are positioned at the nuclear periphery
in the interior, genes elicit distinct expressiostates.On different cell types,identical mecha
ical forces express different genes, whereas the same cell type, altering the amplitude
force or geometry results in differential genexpression patterns (15, 43, 102, 115, 120, 12
In addition, cells experiencing compressive
shear forces exhibit distinct gene expressiprograms. Soft nuclei such as those in stem ce
express distinct lineage-specific geneexpressipatterns based on the substrate stiffness or th
370 Shivashankar
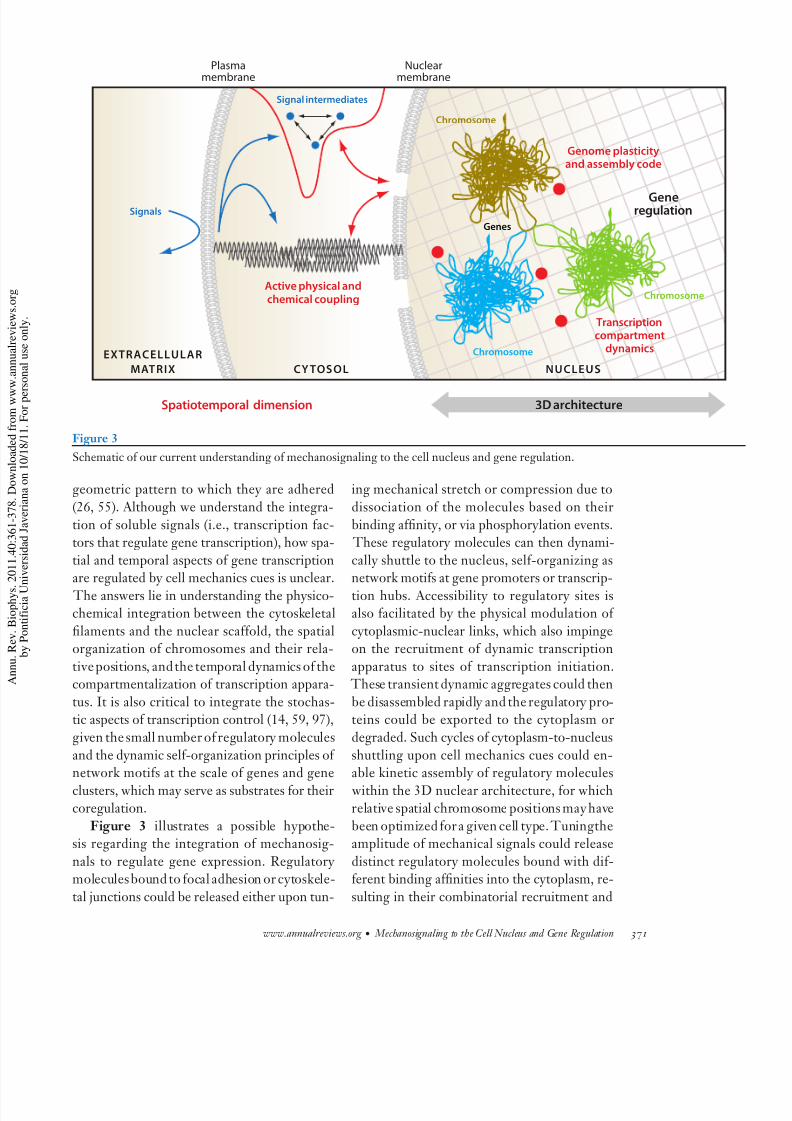
8/3/2019 ojo biolo uda
http://slidepdf.com/reader/full/ojo-biolo-uda 11/20
Plasmamembrane
Signals
Genes
Active physical andchemical coupling
Genome plasticityand assembly code
Transcriptioncompartment
dynamics
Spatiotemporal dimension
Generegulation
Signal intermediates
EXTRACELLULAR
MAT RIX CY TOSOL NUCLEUSUCLEUSNUCLEUS
Nuclearmembrane
Chromosome
Chromosome
Chromosome
3D architecture
Figure 3
Schematic of our current understanding of mechanosignaling to the cell nucleus and gene regulation.
geometric pattern to which they are adhered
(26, 55). Although we understand the integra-
tion of soluble signals (i.e., transcription fac-tors that regulate gene transcription), how spa-
tial and temporal aspects of gene transcriptionare regulated by cell mechanics cues is unclear.
The answers lie in understanding the physico-chemical integration between the cytoskeletal
filaments and the nuclear scaffold, the spatialorganization of chromosomes and their rela-
tive positions, andthe temporal dynamics of thecompartmentalization of transcription appara-
tus. It is also critical to integrate the stochas-tic aspects of transcription control (14, 59, 97),
given the small number of regulatory molecules
and the dynamic self-organization principles of network motifs at the scale of genes and gene
clusters, which may serve as substrates for theircoregulation.
Figure 3 illustrates a possible hypothe-sis regarding the integration of mechanosig-
nals to regulate gene expression. Regulatory moleculesbound to focal adhesion or cytoskele-
tal junctions could be released either upon tun-
ing mechanical stretch or compression due to
dissociation of the molecules based on their
binding affinity, or via phosphorylation events. These regulatory molecules can then dynami-
cally shuttle to the nucleus, self-organizing asnetwork motifs at gene promoters or transcrip-
tion hubs. Accessibility to regulatory sites isalso facilitated by the physical modulation of
cytoplasmic-nuclear links, which also impingeon the recruitment of dynamic transcription
apparatus to sites of transcription initiation. These transient dynamic aggregates could then
be disassembled rapidly and the regulatory pro-teins could be exported to the cytoplasm or
degraded. Such cycles of cytoplasm-to-nucleus
shuttling upon cell mechanics cues could en-able kinetic assembly of regulatory molecules
within the 3D nuclear architecture, for whichrelative spatial chromosome positions may have
been optimized for a given cell type. Tuningtheamplitude of mechanical signals could release
distinct regulatory molecules bound with dif-ferent binding affinities into the cytoplasm, re-
sulting in their combinatorial recruitment and
www.annualreviews.org • Mechanosignaling to the Cell Nucleus and Gene Regulation 371

8/3/2019 ojo biolo uda
http://slidepdf.com/reader/full/ojo-biolo-uda 12/20
induction of differential gene expression pro-grams in differentiated cell types. However, in
stem cells, given the reduced cytoplasmic-to-nuclear ratio and the hyperdynamic nature of
cytoskeletal filaments and chromatin assembly,an increased flux of regulatory molecules and
enhanced collisions between chromosome ter-
ritories could provide a mechanism to keep anactive transcriptome. Matrix rigidity or geo-
metric cues that stiffen the cytoplasmic-nuclearlinks could then provide cytoskeletal substrates
to spatially localize regulatory molecules basedon their binding affinities, resulting in lineage-
specific gene expression programs. While thisis purely speculative, the assumptions on which
this hypothesis stands are beginning to garnerstrong support from a number of scattered ex-
periments in the field.
IMPLICATIONS
In conclusion, we are just beginning to undestand the effect of the cellular microenviro
ment (matrix rigidity or its geometry) on nclear mechanics and genome regulation (10
In addition, the mechanical integrity of the c
nucleus and nuclear mechanical signaling pr
foundly influence cellular homeostatic controby driving cells toward differentiation, proleration, or apoptosis. Further, diseases su
as cancer are hypothesized to originate at thsingle-cell level in its local mechanical enviro
ment. Therefore, understanding the mechancal control of gene function in living cells an
the modularity of their control mechanisms whave important implications in understandi
cellular behavior in diverse functional contex
SUMMARY POINTS
1. Mechanical signals are transduced to the nucleus both by soluble regulatory factors via
nuclear pores and by active stresses via the prestressed cytoplasmic-nuclear links.
2. Integration of physicochemical signals from the cytoplasm and dynamic recruitment of
compartmentalized transcription machinery at regulatory sites are important for chro-matin remodeling and mechanotranscription of genes.
3. Having a stable 3D organization of genes, gene clusters, and chromosome position iscritical for regulating the initiation of gene transcription.
4. Thereexists a spatial andtemporal windowin lengthand timescales between mechanosig-nal generation at the plasma membrane and gene transcription in the nucleus, giving rise
to modularity in mechanosignaling to the nucleus via regulatory molecules and activestresses.
5. Chromosomes are positioned in distinct spatial locations and are mechanically inter-connected with the nuclear lamina in differentiated cells. In addition, the eukaryotic
transcription apparatus is compartmentalized, switching between immobile and mobilefractions.
FUTURE ISSUES1. Do the physicochemical links to the prestressed nucleus imposed by cellular geometry
impinge on the dynamic integration of transcription factors/compartments and activestresses to determine the accessibility and specificity of gene transcription?
2. Is there a coupling betweenfunctional genetic networks andthe 3D nonrandomorganiza-tion of chromosomes to facilitate mechanical regulation of gene transcription programs?
372 Shivashankar

8/3/2019 ojo biolo uda
http://slidepdf.com/reader/full/ojo-biolo-uda 13/20
3. Does the chromatin and nuclear plasticity in undifferentiated cells (stem cells) play anessential role in increasing collisions between interchromosome territories and thereby
regulate mean variability in gene transcription programs?
4. Can one formulate an input (geometry, matrix rigidity)—output (gene transcription
programs) function based on modularity in mechanosignaling to functional nucleararchitecture?
DISCLOSURE STATEMENT
The author is not aware of any affiliations, memberships, funding, or financial holdings that mightbe perceived as affecting the objectivity of this review.
ACKNOWLEDGMENTS
The list of references is not exhaustive and I apologize for omitting many key contributions
from various laboratories in this emerging area of research. I thank members of my laboratory
and a number of collaborators who have contributed to our ongoing research program on nu-
clear mechanics and genome regulation. I also thank the following agencies for generously sup-porting our research program: the Mechanobiology Institute and the Department of BiologicalSciences, National University of Singapore; the National Center for Biological Sciences, TIFR-
Bangalore, India; and the Swarnajayanti Fellowship and the National Nanoscience Initiative of theDepartment of Science and Technology, India.
LITERATURE CITED
1. Adamo L, Naveiras O, Wenzel PL, McKinney-Freeman S, Mack PJ, et al. 2009. Biomechanical forces
promote embryonic haematopoiesis. Nature 459:1131–352. Banerjee B, Bhattacharya D, Shivashankar GV. 2006. Chromatin structure exhibits spatio-temporal
heterogeneity within the cell nucleus. Biophys. J. 91:2297–3033. Bhattacharya D, Mazumder A, Miriam SA, Shivashankar GV. 2006. EGFP-tagged core and linker
histones diffuse via distinct mechanisms within living cells. Biophys. J. 91:2326–364. Bhattacharya D, Talwar S, MazumderA, Shivashankar GV. 2009. Spatio-temporal plasticityin chromatin
organization in mouse cell differentiation and during Drosophila embryogenesis. Biophys. J. 96:3832–395. Bolzer A, Kreth G, Solovei I, Koehler D, Saracoglu K, et al. 2005. Three-dimensional maps of all
chromosomes in human male fibroblast nuclei and prometaphase rosettes. PLoS Biol. 3:e1576. Branco MR, Pombo A. 2006. Intermingling of chromosome territories in interphase suggests role in
translocations and transcription-dependent associations. PLoS Biol. 4:e1387. BrancoMR, Pombo A. 2007. Chromosome organization: newfacts,new models. Trends Cell Biol. 17:127–
348. Brandt A, Papagiannouli F, Wagner N, Wilsch-Brauninger M, Braun M, et al. 2006. Developmental
control of nuclear size and shape by Kugelkern and Kurzkern. Curr. Biol. 16:543–52
9. Brangwynne CP, MacKintosh FC, Kumar S, Geisse NA, Talbot J, et al. 2006. Microtubules can bearenhanced compressive loads in living cells because of lateral reinforcement. J. Cell Biol. 173:733–41
10. Brivanlou AH, Darnell JE Jr. 2002. Signal transduction and the control of gene expression. Science
295:813–1811. Brouzes E, Farge E. 2004. Interplay of mechanical deformation and patterned gene expression in devel-
oping embryos. Curr. Opin. Genet. Dev. 14:367–7412. Brown JM, Leach J, Reittie JE, Atzberger A, Lee-Prudhoe J, et al. 2006. Coregulated human globin
genes are frequently in spatial proximity when active. J. Cell Biol. 172:177–87
www.annualreviews.org • Mechanosignaling to the Cell Nucleus and Gene Regulation 373

8/3/2019 ojo biolo uda
http://slidepdf.com/reader/full/ojo-biolo-uda 14/20
13. Bustamante C, Bryant Z, Smith SB.2003. Tenyears of tension: single-moleculeDNA mechanics. Natu
421:423–27
14. Chang HH, Hemberg M, Barahona M, Ingber DE, Huang S. 2008. Transcriptome-wide noise contr
lineage choice in mammalian progenitor cells. Nature 453:544–47
15. Chiquet M, Gelman L, Lutz R, Maier S. 2009. From mechanotransduction to extracellular matrix ge
expression in fibroblasts. Biochim. Biophys. Acta 1793:911–20
16. Chiquet-Ehrismann R, Tannheimer M, Koch M, Brunner A, Spring J, et al.1994. Tenascin-C expressi
by fibroblasts is elevated in stressed collagen gels. J. Cell Biol. 127:2093–101
17. Chowdhury F, Na S, Li D, Poh YC, Tanaka TS, et al. 2010. Material properties of the cell dictastress-induced spreading and differentiation in embryonic stem cells. Nat. Mater. 9:82–88
18. Cinamon G, Shinder V, Alon R. 2001. Shear forces promote lymphocyte migration across vascul
endothelium bearing apical chemokines. Nat. Immunol. 2:515–22
19. Cook AG, Conti E. 2010. Nuclear export complexes in the frame. Curr. Opin. Struct. Biol. 20:247–52
20. Cook PR. 2010. A model for all genomes: the role of transcription factories. J. Mol. Biol. 395:1–10
21. Cremer M, von Hase J, Volm T, Brero A, Kreth G, et al. 2001. Non-random radial higher-ord
chromatin arrangements in nuclei of diploid human cells. Chromosome Res. 9:541–67
22. Cremer T, Cremer C. 2001. Chromosome territories, nuclear architecture and gene regulation in ma
malian cells. Nat. Rev. Genet. 2:292–301
23. Dahl KN, Ribeiro AJ, Lammerding J. 2008. Nuclear shape, mechanics, and mechanotransductio
Circ. Res. 102:1307–18
24. Discher DE,Mooney DJ, Zandstra PW. 2009. Growth factors, matrices, and forces combine and contstem cells. Science 324:1673–77
25. Duan Z, Andronescu M, Schutz K, McIlwain S, Kim YJ, et al. 2010. A three-dimensional model of t
yeast genome. Nature 465:363–67
26. EnglerAJ, SenS, Sweeney HL,Discher DE.2006. Matrixelasticity directs stem cell lineage specificatio
Cell 126:677–89
27. Etienne-Manneville S, Hall A. 2002. Rho GTPases in cell biology. Nature 420:629–35
28. Fletcher DA, Mullins RD. 2010. Cell mechanics and the cytoskeleton. Nature 463:485–92
29. Fraser P, Bickmore W. 2007. Nuclear organization of the genome and the potential for gene regulatio
Nature 447:413–17
30. Gasser SM. 2002. Visualizing chromatin dynamics in interphase nuclei. Science 296:1412–16
31. Geiger B, Spatz JP, Bershadsky AD. 2009. Environmental sensing through focal adhesions. Nat. R
Mol. Cell Biol. 10:21–3332. Gilbert N, Boyle S, Fiegler H, Woodfine K, Carter NP, Bickmore WA. 2004. Chromatin architectu
of the human genome: Gene-rich domains are enriched in open chromatin fibers. Cell 118:555–66
33. GoetzeS, Mateos-LangerakJ, Gierman HJ,de Leeuw W, Giromus O,et al.2007. Thethree-dimension
structure of human interphase chromosomes is related to the transcriptome map. Mol. Cell Biol. 27:447
87
34. Gorisch SM, Wachsmuth M, Ittrich C, Bacher CP, Rippe K, Lichter P. 2004. Nuclear body moveme
is determined by chromatin accessibility and dynamics. Proc. Natl. Acad. Sci. USA 101:13221–26
35. Grewal SI, Jia S. 2007. Heterochromatin revisited. Nat. Rev. Genet. 8:35–46
36. Gruenbaum Y, Margalit A, Goldman RD, Shumaker DK, Wilson KL. 2005. The nuclear lamina com
of age. Nat. Rev. Mol. Cell Biol. 6:21–31
37. Haque F, Lloyd DJ, Smallwood DT, Dent CL, Shanahan CM, et al. 2006. SUN1 interacts with nucl
lamin A and cytoplasmic nesprins to provide a physical connection between the nuclear lamina and tcytoskeleton. Mol. Cell Biol. 26:3738–51
38. HerrmannH, BarH, Kreplak L,StrelkovSV, Aebi U.2007.Intermediatefilaments: from cell architectu
to nanomechanics. Nat. Rev. Mol. Cell Biol. 8:562–73
39. Hervy M, Hoffman L, Beckerle MC. 2006. From the membrane to the nucleus and back again: bifun
tional focal adhesion proteins. Curr. Opin. Cell Biol. 18:524–32
40. Hogan PG, Chen L, Nardone J, Rao A. 2003. Transcriptional regulation by calcium, calcineurin, an
NFAT. Genes Dev. 17:2205–32
374 Shivashankar

8/3/2019 ojo biolo uda
http://slidepdf.com/reader/full/ojo-biolo-uda 15/20
41. Hubner MR, Spector DL. 2010. Chromatin dynamics. Annu. Rev. Biophys. 39:471–89
42. Hurlbut GD, Kankel MW, Lake RJ, Artavanis-Tsakonas S. 2007. Crossing paths with Notch in the
hyper-network. Curr. Opin. Cell Biol. 19:166–75
43. IlliB, Nanni S, Scopece A, Farsetti A, Biglioli P, et al. 2003. Shear stress-mediated chromatin remodeling
provides molecular basis for flow-dependent regulation of gene expression. Circ. Res. 93:155–61
44. Ingber DE. 1993. Cellulartensegrity:definingnew rulesof biologicaldesign that govern the cytoskeleton.
J. Cell Sci. 104(Pt. 3):613–27
45. Jaalouk DE, Lammerding J. 2009. Mechanotransduction gone awry. Nat. Rev. Mol. Cell Biol. 10:63–73
46. Jackson DA,Hassan AB, Errington RJ,Cook PR.1993. Visualization of focal sites of transcription withinhuman nuclei. EMBO J. 12:1059–65
47. Jackson DA, Iborra FJ, Manders EM, Cook PR. 1998. Numbers and organization of RNA polymerases,
nascent transcripts, and transcription units in HeLa nuclei. Mol. Biol. Cell 9:1523–36
48. Jamora C, Fuchs E. 2002. Intercellular adhesion,signalling and thecytoskeleton. Nat. Cell Biol. 4:E101–8
49. Janmey PA. 1998. The cytoskeleton and cell signaling: component localization and mechanical coupling.
Physiol. Rev. 78:763–81
50. Jenuwein T, Allis CD. 2001. Translating the histone code. Science 293:1074–80
51. Johnson CP, Tang HY, Carag C, Speicher DW, Discher DE. 2007. Forced unfolding of proteins within
cells. Science 317:663–66
52. Kadrmas JL, Beckerle MC. 2004. The LIM domain: from the cytoskeleton to the nucleus. Nat. Rev. Mol.
Cell Biol. 5:920–31
53. Katsumi A, Orr AW, Tzima E, Schwartz MA. 2004. Integrins in mechanotransduction. J. Biol. Chem.279:12001–4
54. Khatau SB, Hale CM, Stewart-Hutchinson PJ, Patel MS, Stewart CL, et al. 2009. A perinuclear actin
cap regulates nuclear shape. Proc. Natl. Acad. Sci. USA 106:19017–22
55. Kilian KA, Bugarija B, Lahn BT, Mrksich M. 2010. Geometric cues for directing the differentiation of
mesenchymal stem cells. Proc. Natl. Acad. Sci. USA 107:4872–77
56. Kornberg RD. 2007. The molecular basis of eukaryotic transcription. Proc. Natl. Acad. Sci. USA
104:12955–61
57. Kosak ST, Groudine M. 2004. Gene order and dynamic domains. Science 306:644–47
58. Kosak ST, Scalzo D, Alworth SV, Li F, Palmer S, et al. 2007. Coordinate gene regulation during
hematopoiesis is related to genomic organization. PLoS Biol. 5:e309
59. Krishna S, Banerjee B, Ramakrishnan TV, Shivashankar GV. 2005. Stochastic simulations of the origins
and implications of long-tailed distributions in gene expression. Proc. Natl. Acad. Sci. USA 102:4771–7660. Kumaran RI, Thakar R, Spector DL. 2008. Chromatin dynamics and gene positioning. Cell 132:929–34
61. Lanctot C, Cheutin T, Cremer M, Cavalli G, Cremer T. 2007. Dynamic genome architecture in the
nuclear space: regulation of gene expression in three dimensions. Nat. Rev. Genet. 8:104–15
62. Leipzig ND, Athanasiou KA. 2008. Static compression of single chondrocytes catabolically modifies
single-cell gene expression. Biophys. J. 94:2412–22
63. Levine M. 2010. Transcriptional enhancersin animal development and evolution. Curr. Biol. 20:R754–63
64. Levy JR, Holzbaur EL. 2008. Dynein drives nuclear rotation during forward progression of motile
fibroblasts. J. Cell Sci. 121:3187–95
65. Lieberman-Aiden E, van Berkum NL, Williams L, Imakaev M, Ragoczy T, et al. 2009. Comprehensive
mapping of long-range interactions reveals folding principles of the human genome. Science 326:289–93
66. Liu B, Kim TJ, Wang Y. 2010. Live cell imaging of mechanotransduction. J. R. Soc. Interface
7(Suppl. 3):S365–7567. Liu C, Yao J, Mercola D, Adamson E. 2000. The transcription factor EGR-1 directly transactivates
the fibronectin gene and enhances attachment of human glioblastoma cell line U251. J. Biol. Chem.
275:20315–23
68. Luger K, Hansen JC. 2005. Nucleosome and chromatin fiber dynamics. Curr. Opin. Struct. Biol. 15:188–
96
69. Luxton GW, Gomes ER, Folker ES, Vintinner E, Gundersen GG. 2010. Linear arrays of nuclear
envelope proteins harness retrograde actin flow for nuclear movement. Science 329:956–59
www.annualreviews.org • Mechanosignaling to the Cell Nucleus and Gene Regulation 375

8/3/2019 ojo biolo uda
http://slidepdf.com/reader/full/ojo-biolo-uda 16/20
70. MacArthur BD, Ma’ayan A, Lemischka IR. 2009. Systems biology of stem cell fate and cellular repr
gramming. Nat. Rev. Mol. Cell Biol. 10:672–81
71. Maniotis AJ, Chen CS, Ingber DE. 1997. Demonstration of mechanical connections between integri
cytoskeletal filaments, and nucleoplasm that stabilize nuclear structure. Proc. Natl. Acad. Sci. USA 94:84
54
72. MarenduzzoD, Micheletti C, CookPR. 2006. Entropy-drivengenome organization. Biophys. J. 90:371
21
73. Mattout A, Dechat T, Adam SA, Goldman RD,Gruenbaum Y. 2006. Nuclear lamins, diseases and agin
Curr. Opin. Cell Biol. 18:335–4174. Mattout A, Meshorer E. 2010. Chromatin plasticity and genome organization in pluripotent embryo
stem cells. Curr. Opin. Cell Biol. 22:334–41
75. Mazumder A, Roopa T, Basu A, Mahadevan L, Shivashankar GV. 2008. Dynamics of chromatin deco
densation reveals the structural integrity of a mechanically prestressed nucleus. Biophys. J. 95:3028–3
76. Mazumder A, Shivashankar GV. 2007. Gold-nanoparticle-assisted laser perturbation of chromatin
sembly reveals unusual aspects of nuclear architecture within living cells. Biophys. J. 93:2209–16
77. Mazumder A, Shivashankar GV. 2010. Emergence of a prestressed eukaryotic nucleus during cellu
differentiation and development. J. R. Soc. Interface 7(Suppl. 3):S321–30
78. Meaburn KJ, Misteli T. 2007. Cell biology: chromosome territories. Nature 445:379–81
79. Mercurio F, Manning AM. 1999. NF-κB as a primary regulator of thestress response. Oncogene 18:616
71
80. Meshorer E, Misteli T. 2006. Chromatin in pluripotent embryonic stem cells and differentiatio Nat. Rev. Mol. Cell Biol. 7:540–46
81. Meshorer E, Yellajoshula D, George E, Scambler PJ, Brown DT, Misteli T. 2006. Hyperdynam
plasticity of chromatin proteins in pluripotent embryonic stem cells. Dev. Cell 10:105–16
82. Miki T, Okawa K, Sekimoto T, Yoneda Y, Watanabe S, et al. 2009. mDia2 shuttles between the n
cleus and the cytoplasm through the importin-α /β- and CRM1-mediated nuclear transport mechanis
J. Biol. Chem. 284:5753–62
83. Misteli T. 2007. Beyond the sequence: cellular organization of genome function. Cell 128:787–800
84. Mitchell JA, Fraser P. 2008. Transcription factories are nuclear subcompartments that remain in t
absence of transcription. Genes Dev. 22:20–25
85. Moore SW, Roca-Cusachs P, Sheetz MP. 2010. Stretchy proteins on stretchy substrates: the importa
elements of integrin-mediated rigidity sensing. Dev. Cell 19:194–206
86. Muller I, Boyle S, Singer RH, Bickmore WA, Chubb JR. 2010. Stable morphology, but dynamic interreorganisation, of interphase human chromosomes in living cells. PLoS One 5:e11560
87. Nagele RG, Freeman T, McMorrow L, Thomson Z, Kitson-Wind K, Lee H. 1999. Chromosom
exhibit preferential positioning in nuclei of quiescent human cells. J. Cell Sci. 112(Pt. 4):525–35
88. Naumova N, Dekker J. 2010. Integrating one-dimensional and three-dimensional maps of genom
J. Cell Sci. 123:1979–88
89. Orr AW, Helmke BP, Blackman BR, Schwartz MA. 2006. Mechanisms of mechanotransduction. D
Cell 10:11–20
90. Osborne CS, Chakalova L, Brown KE, Carter D, Horton A, et al. 2004. Active genes dynamica
colocalize to shared sites of ongoing transcription. Nat. Genet. 36:1065–71
91. Osborne CS, Chakalova L, Mitchell JA, Horton A, Wood AL, et al. 2007. Myc dynamically and pref
entially relocates to a transcription factory occupied by Igh. PLoS Biol. 5:e192
92. Pajerowski JD, Dahl KN, Zhong FL, Sammak PJ, Discher DE. 2007. Physical plasticity of the nuclein stem cell differentiation. Proc. Natl. Acad. Sci. USA 104:15619–24
93. Parada LA, McQueen PG, Misteli T. 2004. Tissue-specific spatial organization of genomes. Geno
Biol. 5:R44
94. Parsons JT, Horwitz AR, Schwartz MA. 2010. Cell adhesion: integrating cytoskeletal dynamics a
cellular tension. Nat. Rev. Mol. Cell Biol. 11:633–43
95. Platani M, Goldberg I, Lamond AI, Swedlow JR. 2002. Cajal body dynamics and association with chr
matin are ATP-dependent. Nat. Cell Biol. 4:502–8
376 Shivashankar

8/3/2019 ojo biolo uda
http://slidepdf.com/reader/full/ojo-biolo-uda 17/20
96. Pombo A, Branco MR. 2007. Functional organisation of the genome during interphase. Curr. Opin.
Genet. Dev. 17:451–55
97. Raj A, van Oudernaarden A. 2009. Single-molecule approaches to stochastic gene expression. Annu. Rev.
Biophys. 38:255–70
98. Rajapakse I, Perlman MD, Scalzo D, Kooperberg C, Groudine M, Kosak ST. 2009. The emergence
of lineage-specific chromosomal topologies from coordinate gene regulation. Proc. Natl. Acad. Sci. USA
106:6679–84
99. Roopa T, Shivashankar GV. 2006. Direct measurement of local chromatin fluidity using optical trap
modulation force spectroscopy. Biophys. J. 91:4632–37100. Rowat AC, Lammerding J, Ipsen JH. 2006. Mechanical properties of the cell nucleus and the effect of
emerin deficiency. Biophys. J. 91:4649–64
101. Sarma K, Reinberg D. 2005. Histone variants meet their match. Nat. Rev. Mol. Cell Biol. 6:139–49
102. Shaik SS, Soltau TD, Chaturvedi G, Totapally B, Hagood JS, et al. 2009. Low intensity shear stress
increases endothelial ELR+CXC chemokine production via a focal adhesion kinase-p38-MAPK-NFkB
pathway. J. Biol. Chem. 284:5945–55
103. Shivashankar GV, ed. 2010. Methods in Cell Biology. Vol. 98: Nuclear Mechanics and Genome Regulation,
p. xiii. New York: Elsevier
104. Shumaker DK, Kuczmarski ER, Goldman RD. 2003. The nucleoskeleton: Lamins and actin are major
players in essential nuclear functions. Curr. Opin. Cell Biol. 15:358–66
105. Sims JR, Karp S, Ingber DE. 1992. Altering the cellular mechanical force balance results in integrated
changes in cell, cytoskeletal and nuclear shape. J. Cell Sci. 103(Pt. 4):1215–22106. Sinha DK, Banerjee B, Maharana S, Shivashankar GV. 2008. Probing the dynamic organization of
transcription compartments and gene loci within the nucleus of living cells. Biophys. J. 95:5432–38
107. Spann TP, Goldman AE, Wang C, Huang S, Goldman RD. 2002. Alteration of nuclear lamin organi-
zation inhibits RNA polymerase II-dependent transcription. J. Cell Biol. 156:603–8
108. Spivakov M, Fisher AG. 2007. Epigenetic signatures of stem cell identity. Nat. Rev. Genet. 8:263–71
109. Sproul D, Gilbert N, Bickmore WA. 2005. The role of chromatin structure in regulating the expression
of clustered genes. Nat. Rev. Genet. 6:775–81
110. Stewart CL, Roux KJ, Burke B. 2007. Blurring the boundary: The nuclear envelope extends its reach.
Science 318:1408–12
111. Strambio-De-Castillia C, Niepel M, Rout MP. 2010. The nuclear pore complex: bridging nuclear trans-
port and gene regulation. Nat. Rev. Mol. Cell Biol. 11:490–501
112. Sun HB, Shen J, Yokota H. 2000. Size-dependent positioning of human chromosomes in interphasenuclei. Biophys. J. 79:184–90
113. Sutherland H, Bickmore WA. 2009. Transcription factories: gene expression in unions? Nat. Rev. Genet.
10:457–66
114. Tay S, Hughey JJ, Lee TK, Lipniacki T, Quake SR, Covert MW. 2010. Single-cell NFkB dynamics
reveal digital activation and analogue information processing. Nature 466:267–71
115. Thomas CH, Collier JH, Sfeir CS, Healy KE. 2002. Engineering gene expression and protein synthesis
by modulation of nuclear shape. Proc. Natl. Acad. Sci. USA 99:1972–77
116. Tzima E, Irani-Tehrani M, Kiosses WB, Dejana E, Schultz DA, et al. 2005. A mechanosensory complex
that mediates the endothelial cell response to fluid shear stress. Nature 437:426–31
117. Tzur YB, WilsonKL, GruenbaumY. 2006. SUN-domain proteins:‘Velcro’that links thenucleoskeleton
to the cytoskeleton. Nat. Rev. Mol. Cell Biol. 7:782–88
118. Vartiainen MK, Guettler S, Larijani B, Treisman R. 2007. Nuclear actin regulates dynamic subcellularlocalization and activity of the SRF cofactor MAL. Science 316:1749–52
119. Vogel V, Sheetz M. 2006. Local force and geometry sensing regulate cell functions. Nat. Rev. Mol. Cell
Biol. 7:265–75
120. Wang JH, Thampatty BP, Lin JS, Im HJ. 2007. Mechanoregulation of gene expression in fibroblasts.
Gene 391:1–15
121. Wang N, Tytell JD, Ingber DE. 2009. Mechanotransduction at a distance: mechanically coupling the
extracellular matrix with the nucleus. Nat. Rev. Mol. Cell Biol. 10:75–82
www.annualreviews.org • Mechanosignaling to the Cell Nucleus and Gene Regulation 377

8/3/2019 ojo biolo uda
http://slidepdf.com/reader/full/ojo-biolo-uda 18/20
122. Wang Y, Botvinick EL, Zhao Y, Berns MW, Usami S, et al. 2005. Visualizing the mechanical activati
of Src. Nature 434:1040–45
123. Wang Y, Gilmore TD. 2003. Zyxin and paxillin proteins: Focal adhesion plaque LIM domain prote
go nuclear. Biochim. Biophys. Acta 1593:115–20
124. Whitehead J, Vignjevic D, Futterer C, Beaurepaire E, Robine S, Farge E. 2008. Mechanical facto
activate beta-catenin-dependent oncogene expression in APC mouse colon. HFSP J. 2:286–94
125. Woodcock CL. 2006. Chromatin architecture. Curr. Opin. Struct. Biol. 16:213–20
126. Xu J, Zutter MM, Santoro SA, Clark RA. 1998. A three-dimensional collagen lattice activa
NF-kappaB in human fibroblasts: role in integrin alpha2 gene expression and tissue remodelin J. Cell Biol. 140:709–19
127. Xu M, Cook PR. 2008. Similar active genes cluster in specialized transcription factories. J. Cell Bi
181:615–23
128. Yamanaka S, Blau HM. 2010. Nuclear reprogramming to a pluripotent state by three approaches. Natu
465:704–12
129. Yang G, Im HJ, Wang JH. 2005. Repetitive mechanical stretching modulates IL-1beta induced COX
MMP-1 expression, and PGE2 production in human patellar tendon fibroblasts. Gene 363:166–72
130. Yao J, Munson KM, Webb WW, Lis JT. 2006. Dynamics of heat shock factor association with nati
gene loci in living cells. Nature 442:1050–53
131. Zaratiegui M, Irvine DV, Martienssen RA. 2007. Noncoding RNAs and gene silencing. Cell 128:763–
132. Zhang Q, Ragnauth CD, Skepper JN, Worth NF, Warren DT, et al. 2005. Nesprin-2 is a multi-isome
protein that binds lamin and emerin at the nuclear envelope and forms a subcellular network in skelemuscle. J. Cell Sci. 118:673–87
133. Zink D, Amaral MD, Englmann A, Lang S, Clarke LA, et al. 2004. Transcription-dependent spat
arrangements of CFTR and adjacent genes in human cell nuclei. J. Cell Biol. 166:815–25
378 Shivashankar

8/3/2019 ojo biolo uda
http://slidepdf.com/reader/full/ojo-biolo-uda 19/20
Annual Review
Biophysics
Volume 40, 20Contents
Respice, Adspice, and Prospice
Harold A. Scheraga p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 1
Equilibrium Sampling in Biomolecular SimulationsDaniel M. Zuckerman p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p41
Decision Making in Living Cells: Lessons from a Simple System Ido Golding p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p63
High-Pressure Protein Crystallography and NMR to ExploreProtein Conformations
Marcus D. Collins, Chae Un Kim, and Sol M. Gruner p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p81
Nucleosome Structure(s) and Stability: Variations on a Theme
Andrew J. Andrews and Karolin Luger p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p99
Molecular Mechanisms of Ubiquitin-Dependent Membrane Traffic James H. Hurley and Harald Stenmark p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 119
The Cyanobacterial Circadian System: From Biophysics
to Bioevolution
Carl Hirschie Johnson, Phoebe L. Stewart, and Martin Egli p p p p p p p p p p p p p p p p p p p p p p p p p p p p
143 Actin Structure and Function
Roberto Dominguez and Kenneth C. Holmes p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 169
Molecular Origin of the Hierarchical Elasticity of Titin: Simulation,
Experiment, and Theory
Jen Hsin, Johan Str¨ umpfer, Eric H. Lee, and Klaus Schulten p p p p p p p p p p p p p p p p p p p p p p p p p p p 187
Proton-Pumping Mechanism of Cytochrome c Oxidase
Shinya Yoshikawa, Kazumasa Muramoto, and Kyoko Shinzawa-Itoh p p p p p p p p p p p p p p p p p p 205
SAXS Studies of Ion–Nucleic Acid Interactions Lois Pollack
p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p225
P-Type ATPases Michael G. Palmgren and Poul Nissen p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 243
Kinesin Assembly and Movement in Cells
Kristen J. Verhey, Neha Kaul, and Virupakshi Soppina p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 267
v

8/3/2019 ojo biolo uda
http://slidepdf.com/reader/full/ojo-biolo-uda 20/20
Stochastic Conformational Pumping: A Mechanism for Free-Energy
Transduction by Molecules
R. Dean Astumian p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 2
Protein Self-Organization: Lessons from the Min System
Martin Loose, Karsten Kruse, and Petra Schwille p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 3
Protein Folding at the Exit Tunnel
Daria V. Fedyukina and Silvia Cavagnerop p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p
3
Mechanosignaling to the Cell Nucleus and Genome RegulationG.V. Shivashankar p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 3
Amphipols From A to Z
J.-L. Popot, T. Althoff, D. Bagnard, J.-L. Baneres, P. Bazzacco, E. Billon-Denis,
L.J. Catoire, P. Champeil, D. Charvolin, M.J. Cocco, G. Cr´ emel, T. Dahmane,
L. de la Maza, C. Ebel, F. Gabel, F. Giusti, Y. Gohon, E. Goormaghtigh,
E. Guittet, J.H. Kleinschmidt, W. K¨ uhlbrandt, C. Le Bon, K.L. Martinez,
M. Picard, B. Pucci, J.N. Sachs, C. Tribet, C. van Heijenoort, F. Wien, F. Zito,
and M. Zoonens p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p
3
Index
Cumulative Index of Contributing Authors, Volumes 36–40 p p p p p p p p p p p p p p p p p p p p p p p p p p p 4
Errata
An online log of corrections to Annual Review of Biophysics articles may be found at
http://biophys.annualreviews.org/errata.shtml


















