
J.A. Boninsegna*, R. Villalba*, Amarilla**, J.Ocampo**
9
IAWA Bulletin n.s., Vol. 10 (2),1989: 161-169 STUDIES ON TREE RINGS, GROWfH RATES AND AGE-SIZE RELATIONSHIPS OF TROPICAL TREE SPECIES IN MISIONES, ARGENTINA by J.A. Boninsegna*, R. Villalba*, L. Amarilla**, and J.Ocampo** Summary Wood samples of 13 tree species from three sites in the Selva Misionera (Misiones Province, Argentina) were analysed macro- and microscopically for occurrence and for- mation of growth rings. Well-defined annual tree rings were found in Cedrelafissilis VeIl., Parapiptadenia rigida Benth., Cordia tricho- toma VeIl. and Chorisia speciosa St. Hil. In 15 trees of Cedrela fissilis VeIl., the growth rate, the current and the mean annual increment (CAl and MAl) and the diameter/ age relationship were estimated using incre- ment borer samples. The estimated mean cul- mination age of the basal MIA was 152 years, while the same parameter measured on individual trees shows a wide range from 61 to 180 years, probably representing different social positions of the trees. The method proposed is discussed as a way to obtain a quick, reliable, and inexpen- sive estimation of parameters which are use- ful in forest management. Key words: Tropical trees, growth rate, tree rings. Introduction One of the most frequent uses of tree ring information is in the determination of the age of individual trees or stands of trees. Age de- termination requires that a researcher is able to recognise rings and also that the trees regularly produce one and only one ring each year. When a species is shown to have iden- tifiable annual rings, a ring count can nor- mally provide age estimates that are suffi- ciently accurate for many purposes (Duever & McCollom 1987). Several studies have been carried out on the wood anatomy of tropical trees in order to identify the growth layers and their temporal sequence and several methods have been used to reach this objective (Mariaux 1967; Catinot 1970; Tomlinson & Craighead 1972; Eckstein et al. 1981; Lieberman et al. 1985; Villalba 1985; Worbes 1985, 1986). Growth rates of woody plants as indicated by their annual ring widths are always to some degree a function of both natural and anthropogenic conditions (Fritts 1976). Com- parisons of ring widths in the same species over time or in different places can provide valuable information on how woody plant growth varies temporally or spatially as a function of various environmental conditions. In addition, it is possible to evaluate the re- lative productivity of species and sites, or how a species' growth is affected by various management practices. The Selva Misionera represents the south- ernmost part of the Amazon pluvial forest, extending over Misiones Province, Argen- tina, from 25° to 27° L.S. (Fig. 1). Precipi- tation is distributed throughout the year reaching a total average of more than 1700 mm. Mean annual temperature ranges be- tween 19 to 20°C. Frosts occasionally occur, mainly in June or July. Lateritic soils, in combination with high temperature and pre- cipitation allow the growth of subtropical evergreen forest, called Selvas Mixtas (Ca- brera 1976). The most important families are Leguminosae, Lauraceae, Meliaceae, Boragi- naceae and Bignoniaceae (Takao Inoue et al. 1984). * Laboratorio de Dendrocronologfa, Centro Regional de Investigaciones Cientfficas y T6cno- l6gicas, CONICEf, Casilla de Correo 330, 5500 Mendoza, Argentina. ** Facultad de Ciencias Forestales, Universidad de Nacional de Misiones, 3380 Eldorado, Misiones, Argentina. Downloaded from Brill.com01/16/2022 01:03:05PM via free access
Transcript of J.A. Boninsegna*, R. Villalba*, Amarilla**, J.Ocampo**

IAWA Bulletin n.s., Vol. 10 (2),1989: 161-169
STUDIES ON TREE RINGS, GROWfH RATES AND AGE-SIZE RELATIONSHIPS OF TROPICAL TREE SPECIES IN MISIONES, ARGENTINA
by
J.A. Boninsegna*, R. Villalba*, L. Amarilla**, and J.Ocampo**
Summary Wood samples of 13 tree species from
three sites in the Selva Misionera (Misiones Province, Argentina) were analysed macroand microscopically for occurrence and formation of growth rings. Well-defined annual tree rings were found in Cedrelafissilis VeIl., Parapiptadenia rigida Benth., Cordia trichotoma VeIl. and Chorisia speciosa St. Hil.
In 15 trees of Cedrela fissilis VeIl., the growth rate, the current and the mean annual increment (CAl and MAl) and the diameter/ age relationship were estimated using increment borer samples. The estimated mean culmination age of the basal MIA was 152 years, while the same parameter measured on individual trees shows a wide range from 61 to 180 years, probably representing different social positions of the trees.
The method proposed is discussed as a way to obtain a quick, reliable, and inexpensive estimation of parameters which are useful in forest management. Key words: Tropical trees, growth rate, tree
rings.
Introduction One of the most frequent uses of tree ring
information is in the determination of the age of individual trees or stands of trees. Age determination requires that a researcher is able to recognise rings and also that the trees regularly produce one and only one ring each year. When a species is shown to have identifiable annual rings, a ring count can normally provide age estimates that are sufficiently accurate for many purposes (Duever & McCollom 1987).
Several studies have been carried out on the wood anatomy of tropical trees in order to identify the growth layers and their temporal sequence and several methods have been used to reach this objective (Mariaux 1967; Catinot 1970; Tomlinson & Craighead 1972; Eckstein et al. 1981; Lieberman et al. 1985; Villalba 1985; Worbes 1985, 1986).
Growth rates of woody plants as indicated by their annual ring widths are always to some degree a function of both natural and anthropogenic conditions (Fritts 1976). Comparisons of ring widths in the same species over time or in different places can provide valuable information on how woody plant growth varies temporally or spatially as a function of various environmental conditions. In addition, it is possible to evaluate the relative productivity of species and sites, or how a species' growth is affected by various management practices.
The Selva Misionera represents the southernmost part of the Amazon pluvial forest, extending over Misiones Province, Argentina, from 25° to 27° L.S. (Fig. 1). Precipitation is distributed throughout the year reaching a total average of more than 1700 mm. Mean annual temperature ranges between 19 to 20°C. Frosts occasionally occur, mainly in June or July. Lateritic soils, in combination with high temperature and precipitation allow the growth of subtropical evergreen forest, called Selvas Mixtas (Cabrera 1976). The most important families are Leguminosae, Lauraceae, Meliaceae, Boraginaceae and Bignoniaceae (Takao Inoue et al. 1984).
* Laboratorio de Dendrocronologfa, Centro Regional de Investigaciones Cientfficas y T6cnol6gicas, CONICEf, Casilla de Correo 330, 5500 Mendoza, Argentina.
** Facultad de Ciencias Forestales, Universidad de Nacional de Misiones, 3380 Eldorado, Misiones, Argentina.
Downloaded from Brill.com01/16/2022 01:03:05PMvia free access

162
~ Sample site
C=:J Su btropical evergreen forest of Brasil
_ Araucaria angustifolia forest
IAWA Bulletin n.s., Vol. 10 (2),1989
.uJJ Mesopotamico park· like forest
_ Parana fluvial forest
I " d Pampas grassland
Fig. 1. Location map of northeastern Argentina and surrounding areas showing the sample site positions.
In this report, 13 species of the Selva Misionera are analysed in order to determine their dendrochronological characteristics. On one of them, Cedrela fissilis VeIl., a preliminary study of the age and growth rate was undertaken on the basis of its growth ring widths.
Wood anatomy and tree rings Anatomical studies of wood samples of 13
tree species from different places in Misiones Province were carried out with special attention to the growth zones. The main anatomical structures responsible for the visibility of growth rings are:
1. Ring-porous or semi-ring-porous structure: a. The earlywood consists of large ves
selsembedded in a mass of initial parenchyma cells:
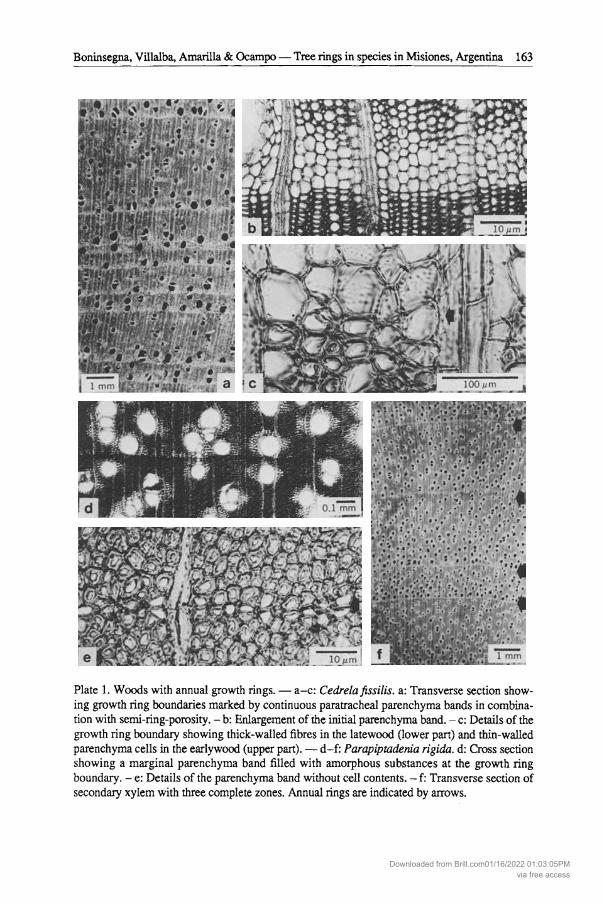
Cedrelafissilis (Plate la-c) b. The latewood is made up of small and
medium-sized vessels and thick- walled fibres: Cordia triclwtoma (Plate 2a, b)
2. Boundaries with uni- or multi seriate marginal parenchyma bands. The parenchyma cells at the boundaries are often filled with crystalline substances:
Parapiptadenia rigida (Plate Id-t) Peltoplwrum dubium (Plate 2d)
Myrocarpus frondosus (Plate 2e, t)
Downloaded from Brill.com01/16/2022 01:03:05PMvia free access
Boninsegna, Villalba, Amarilla & Ocampo - Tree rings in species in Misiones, Argentina 163
Plate 1. Woods with annual growth rings. - a-c: Cedrelafissilis. a: Transverse section showing growth ring boundaries marked by continuous paratracheal parenchyma bands in combination with semi-ring-porosity. - b: Enlargement of the initial parenchyma band. - c: Details of the growth ring boundary showing thick-walled fibres in the latewood (lower part) and thin-walled parenchyma cells in the earlywood (upper part). - d-f: Parapiptadenia rigida. d: Cross section showing a marginal parenchyma band filled with amorphous substances at the growth ring boundary. - e: Details of the parenchyma band without cell contents. - f: Transverse section of secondary xylem with three complete zones. Annual rings are indicated by arrows.
Downloaded from Brill.com01/16/2022 01:03:05PMvia free access

164 IAWA Bulletin n.s., Vol. 10 (2),1989
Plate 2. Woods with annual growth rings (continued). - a, b: Cordia trichotomfl. a: Tree rings defined by a narrow initial parenchyma band. In b, the parenchyma band is associated with semi-ring-porosity (arrows). - c: Chorisia speciosa showing an annual ring consisting almost solely of fibres in the earlywood. - d-f: Species with growth rings, not necessarily aTlnual. -d: Peitophorum dubium with irregular zonate bands of apotracheal parenchyma (arrows). -e, f: Myrocarpusfrondosus. e: Narrow parenchyma bands completely surrounding the trunk.f: Details of the 1-4-seriate parenchyma band.
---+ Plate 3. Species with less distinct, ± irregular growth increments. - a: Aspidosperma poiyneuron, annual ring defined by a narrow band of fibres representing the first formed earlywood. -Nectandra sa/igna (b) and Ocotea puberuia (c) showing both gradual and abrupt changes in radial diameter and wall thickness of the fibres. - d-f: Enteroiobium contortisiliquum.
Downloaded from Brill.com01/16/2022 01:03:05PMvia free access

Boninsegna, Villalba, Amarilla & Ocampo - Tree rings in species in Misiones, Argentina 165
d: Two interannual parenchyma bands (indicated by arrows) formed during the same year of growth. Growth rings defined by a reduction in radial diameter of the last formed fibres (e) or by a parenchyma band (t). - Species with indistinct growth rings, g: Balfourodendron riedelianum, and h: Cabralea oblongifolia.
Downloaded from Brill.com01/16/2022 01:03:05PMvia free access

166 IAWA Bulletin n.s., Vol. 10 (2),1989
3. Growth zone boundaries conslsttng of several rows of fibre cells with a short radial diameter and thick walls.
The patterns described here may occur in various combinations within the growth layer of some species. Thus, thin bands of parenchyma beside flattened fibre cells occur in the same growth layers of Enterolobium contortisiliquum (plate 3d-f).
a. Zones of thicker-walled fibres are present at the beginning of growth layers:
Chorisia speciosa (Plate 2c) Aspidosperma polyneuron (plate 3a)
The abrupt change from the late- to the earlywood with relatively low and high fibre percentages respectively, marks the tree ring limit. Moreover, the tissue percentage taken up by fibres gradually diminishes along the annual ring.
b. Thicker fibre zones are found at the end of growth layers. In this case, the transition of growth zones can be gradual or abrupt in the same tree:
Ocotea puberula (plate 3b) Nectandra saligna (plate 3c)
4. Periodical recurring patterns of parenchyma along the growth layers:
Cabralea oblongifolia (plate 3h)
Dendrochronological studies on the end grains of disks and increment borer samples of the same material analysed microscopically, have allowed to establish a preliminary classification in relation to visibility of growth layers, continuity along the cross section, and annual nature of growth zones. The annual nature was established taking into account the studies performed on other species of the same genus (Villalba et al. 1985) and the local rate of tree growth according to Maradei (1982), Maradei et al. (1982), Gartland et al. (1969), Fernandez Rodriguez (1963), and Mangieri (1957). These observations have been summarised in Table 1.
Table 1. Dendrochronological characteristics of some tropical trees growing in Misiones Province, Argentina.
Growth layers well defined
Continuous over the entire cross section
Representing one year of growth (plates 1a-f, 2a-c)
Cedrelafissilis Vell. (Meliaceae) Parapiptadenia rigida Benth. (Legum.-Mimosoideae) Cordia trichotoma Vell. (Boraginaceae) Chorisia speciosa St. Hil. (Bombacaceae)
Not representing annual growth (Plate 2d-f)
Peltophorum dubium Spreng. (Legum.-Caesalpinioideae) Myrocarpus frondosus Fr. Allem. (Legum. -Papilionoideae)
Not continuous over the entire cross section
Representing one year of growth (Plate 3a)
Aspidosperma polyneuron Mull. (Apocynaceae)
Sometimes, but not always representing one year of growth (Plate 3b-f)
Ocotea puberu/a Nees. et Mart. (Lauraceae) Nectandra sa/igna Nees et Mart. (Lauraceae) Enterolobium contortisiliquum VeIl. (Legum. -Mimosoideae)
Growth layers vague or absent (plate 3g, h)
Tabebuia ipe Mart. (Bignoniaceae) Balfourodendron riedelianum Eng!. (Rutaceae) Cabralea oblongifolia C. DC. (Meliaceae)
Downloaded from Brill.com01/16/2022 01:03:05PMvia free access

Boninsegna, Villalba, Amarilla & Ocampo - Tree rings in species in Misiones, Argentina 167
em 40 a
em
40
em
30 60 90 120 150 180 210 age (years)
c
30 60 90 120 150 age (years)
30 e
20 MAl
10 CAl
30 60 90 120 150 age (years)
em
b 80
60
40
20
50 100 150 200
m2
0.4
age (years)
d
30 60 90 120 150 age (years)
f 40
30
20
10
Ot:;.J...J~L....L..JL....L..J....LJ....LJ...J....1...J....1 30 60 90 120 150
age (years)
MAl
CAl
Fig. 2. Growth rate of Cedrela fissilis. a: Cumulative radial increment of individuals, mean curve = broken line. b: Age-diameter relationship. c & d: Mean cumulative radial and basal increment Mean annual increment (MIA) and current annual increment (CAl) from radial (e) and basal (f) data, respectively.
Downloaded from Brill.com01/16/2022 01:03:05PMvia free access

168
Age and growth rate of CedreIa fissilis A total of 15 trees of CedreLafissilis Vell.
were cored twice at breast height Cored samples were processed following Stokes and Smiley (1968). The rings were observed under a microscope and the estimated age of each one was recorded, starting from the pith which was assigned the age of 1. Ring width measurements were obtained for every growth year to the nearest one hundredth of a millimetre.
The arithmetic mean of the values for each ring was obtained and the series was smoothed by a cubic spline filter of 0.65 N pass, in which N indicates the number of elements of the series.
Based on the ring width' of the corresponding age and the previous radius, basal area increment (cm2) was estimated for each growth year according to the equation:
Ba (t) = 21t r (t-1) Ir (t)
in which Ba(t) is the basal area at age t, r(t-l) is the radius reached by the tree at age t-l and Ir (t) is the ring width of the t-aged ring.
Data analysis was twofold, involving the comparison of radial and basal area growth and the determination of the following parameters: mean annual radial, mean annual basal, current annual radial and current annual basal increment. The mean annual increment (MAl) is defined as the average annual increment that occurs up to a given age as calculated by dividing the accumulated increment by the age. The current annual increment (CAl) is the amount of growth that occurs at a specified age. The culmination age is the age at which the MAl reaches its maximum value (Assman 1970).
The results are summarised in Figure 2a-f. Cross dating was not carried out on this material due to the low circular uniformity observed in cross section and the fact that it was not needed in the analysis of the data.
The differences in culmination age for radial increment (c. 63 years) and basal area increment (c. 153 years), observed in the mean curves, are particularly striking.
The analysis of the culmination age of the basal MAl made on individual trees, showed a wide variation, ranging from 61 to 180 years, independent of tree diameter. Some
IAWA Bulletin n.s., Vol. 10 (2),1989
trees have not reached the culmination age yet. This result could indicate differences in social position between the trees in the forest.
A more or less linear relationship could be established between age and diameter, with a high correlation coefficient (r = 0.87). These results are of a preliminary nature because they are based on a very limited number of trees.
Conclusions Several tropical trees are capable of form
ing growth rings. Out of 13 species, macroand microscopically analysed, from the Selva Misionera, 10 species show well-defined growth rings, whereas only 3 species have vague growth rings or lack them. The principal anatomical structures involved in their formation are initial parenchyma cells, terminal parenchyma cells and radially flattened and/or thick-walled fibres at the beginning or at the end of tree rings. The anatomical structure responsible for the visibility of growth rings is similar within species of the same genus. CedreLafissilis from the Selva Misionera as well as C. angustifolia Sesse Moc. and C. Lilloi C. DC., growing in the Tucumano-oranense forest, show tree rings defined by an initial band of parenchyma (Villalba et aL. 1985). This rule is not valid for species of the same family. Even though CabraLea obLongifolia belongs to the same family as CedreLa fissilis (Meliaceae), the growth rings in the former species are vague or not well defined. However, the occurrence of certain ring types is predominant in some families. The Leguminosae species analysed mainly have bands of terminal parenchyma, even when in some species these bands are not necessarily annual. In Leguminosae species in Central Amazonia, Worbes (1985) found that marginal parenchyma bands are the most common anatomical structure in relation to growth ring demarcation.
The dendrochronological studies on Cedrela fissilis enable a test of the applicability of the tree ring width measurements in determining age and growth rate in a tropical tree. One important advantage of the method is that it permits an insight into the dynamics of growth of each individual and to a certain
Downloaded from Brill.com01/16/2022 01:03:05PMvia free access

Boninsegna, Villalba, Amarilla & Ocampo - Tree rings in species in Misiones, Argentina 169
degree into the life history of the tree. Although the age values have some errors due to the different time at which individual treelets reach breast height, the determination of age and growth rate on the basis of tree ring widths is a very economic, fast and reliable method that could be applied in tropical regions where there are no other sources of information.
References Assman, E. 1970. The principles of forest
yield study (ed. P.W. Davis). Pergamon Press, New York.
Cabrera, A. C. 1976. Regiones Fitogeograficas Argentinas. In: Encidopedia Argentina de Agricultura y Jardinerfa 2. Edit. ACME,
Buenos Aires. Catinot, R. 1970. Premiers reflexions sur
une possibilite d'explication physiologique des rhytmes annuels d'accroissement chez les arbres de la foret tropicale africaine. Bois et Forets Tropiques 131: 3-14.
Duever, M. & 1. McCollom. 1987. Trade offs between the use of tree ring counting and dendrochronology in ecological studies. Proc. International Symp. Ecological Aspects of Tree Ring Analysis, Marymount College, Tarrytown, New York: 611-621.
Eckstein, D., J. Odgen, G.C. Jacoby & J. Ash. 1981. Age and growth rate determination in tropical trees: The application of dendrochronological methods. In: Age and growth rate of tropical trees: new directions for research (eds. F.H. Bormann & G. Berlyn): 83-106. Yale Univ., New Haven.
Fernandez Rodriguez, M. 1963. Multiplicaci6n espontanea del Peteribi (Cordia trichotoma) previa eliminaci6n del bosque original en la provincia argentina de Misiones. Rev. For. Arg. 7 (4): 11-14.
Fritts, H.C. 1976. Tree rings and climate. Acad. Press, New York.
Gartland, H.M. & C.M. Volkart. 1969. Determinaci6n del crecimiento de una plantaci6n de Cordia tricotoma en la provincia de Misiones, Argentina. Actas 1er. Congreso Forestal Argentino: 223-225.
Hueck, K. 1978. Los bosques de Sudamerica. Soc. Alemana de Coopemci6n. Tecnica, Eschborn.
Lieberman, D., M. Lieberman, G. Hartshorn & R. Peralta. 1985. Growth rate and agerelationships of tropical wet forest trees in Costa Rica. J. Trop. Ecol. 1: 97-109.
Mangieri, H.R. 1957. Notas sobre caracterfsticas y cultivo del Cedro misionero (Cedrela tubifiora). Rev. For. Arg. 1 (3): 112-114.
Maradei, D. 1982. Cultivo de especies de la Selva Misionera I. Resefia Bibliografica. Actas Primeras Jornadas Tecnicas sobre Bosques Implantados en el Noreste Argentino, Un. Nac. de Misiones: 105-111.
- , A. Morales, J.R. Ruiz & J.E. Torres. 1982. Ibid. II. Comportamiento inicial bajo cubierta de pinos. Ibid.: 111-113.
Mariaux, A. 1967. Les cernes dans les bois tropicaux africains, nature et periodicite I: Peuvent-ils reveler l'age des arbres? II: Periodicite des cernes, methodes d' etude et premiers resultats. Bois et Forets des Tropiques No. 113: 3-14 & No. 114: 23-37.
Stokes, M.A. & T.L. Smiley. 1968. An introduction to tree ring dating. Univ. Chicago Press, Chicago.
Takao Inoue, M., C. V. Roderjan & Y. S. Kuniyoshi. 1984. Projeto Madeira de Parana. Curitiba, Fundaya5 de Pesquisas Horestais do Parana.
Torillinson, P.B. & F.C. Craighead. 1972. Growth-ring studies on the native trees of subtropical Horida. In: Research trends in plant anatomy (eds. A.K.M. Ghouse & M. Yunus): 39-51. McGraw Hill, New Delhi.
Villalba, R. 1985. Xylem structure and cambial activity in Prosopis flexuosa. IAWA Bull. n.s. 6: 119-130.
- ,1. Boninsegna & R. Holmes. 1985. Cedrela angustifolia and Juglans australis: two new tropical species useful in dendrochronology. Tree Ring Bull. 45: 25-35.
Worbes, M. 1985. Structural and other adaptations to long-term flooding by trees in Central Amazonia. Amazoniana 9: 459-484.
- 1986. Lebensbedingungen undHolzwachstum in zentralarnazonischen Uberschwemmungswaldern. Scripta Geobotanica 17: 1-112.
Downloaded from Brill.com01/16/2022 01:03:05PMvia free access


















