
THE JOURNAL OF Vol 269, No 40, Issue pp 24854-24863, 1994 ... · DNA Sequence and Functions of the...
10
THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1994 by The American Society for Biochemistry and Molecular Biology, Inc Vol . 269, No . 40, Issue of October 7, pp , 24854-24863, 1994 Printed in U.S.A. DNA Sequence and Functions of the actVI Region of the Actinorhodin Biosynthetic Gene Cluster of Streptomyces coelicolor A3(2)* (Received forpublication, April 15, 1994, and in revised form, July 18, 1994) Miguel A. Fernandez-Morenol, Eduardo Martinez$, Jose L. CaballeroPn, Koji IchinoseQ, David A Hopwood§, and Francisco Malpartidalll From the SCentro Nacional de Biotecnologia, Consejo Superior de Investigaciones Cientificas, Campus Universidad Autonoma de Madrid, Cantoblanco 28049, Madrid, Spain and the §John Znnes Institute, John Znnes Center, Colney Lane, Norwich NR4 7UH, United Kingdom Six open reading frames (ORFs) were identified by DNA sequencing of 5.7 kilobase pairs at the left end of the act cluster (the so-called “actVI region”), in the or- der: ORFB, ORFA, ORF1, ORF2, ORF3,ORF4.ORF1-4 are transcribed rightward and inthe same direction as the ORFs of the actVA region which lies to the right of the actVI region, whereas ORFA and ORFB run in the opposite direction. By complementation of mutants and gene disruption of the wild type strain, the two previ- ously genetically characterized actVI mutations were assigned to ORF1. Although disruption of ORFB and ORF4, using +C3l derivatives,did not cause any obvious change in actinorhodin production, defects in actino- rhodin synthesis were obtained by insertional inactiva- tion of ORFA, ORF1, ORF2, or ORF3. RNA analysis within the ORFUORFA intergenic region showed over- lapping divergent promoters, at least one of which is under the control of the actII-ORF4 gene product, the transcriptional activator of the act cluster. Data base searches with the deduced products of ORFB and ORF3 failed to show any significant similarities with other known proteins. The deduced product of ORFA strongly resembles those of genes of unknown function from Sac- charopolyspora hirsuta and Streptomycesroseofulvus, located within polyketide synthase clusters. The ORFl product strongly resembles P-hydroxyacyl-CoA dehy- drogenases of bacteria and mammals and the ORF2 and ORF4 products resemble each other and enoyl reduc- tases from bacteria, animals, and plants, with a highly conserved cofactor-binding domain. These findings strongly suggest that the actVI region is involved in catalyzing reduction processes that determine the two stereochemical configurations at C-3/C-15 during acti- norhodin biosynthesis. A scheme is proposed for the middle steps of the biosynthesis, that is formation of the CICYT (BI0090-0760 and BI093-1181), EC (BIOT-CT91-0255, SMA) * This research was supported by grants (to F. M.) from the Spanish and Proyecto Concertado Smith Kline and Beecham, S. A./CSIC (CDTI, number 930139) and by grants-in-aid (to D. A. H.) to the John Innes Institute from the Agricultural and FoodResearch Council and the John Innes Foundation and Grant BIOT-CT91-0255 from the EC BRIDGE Programme. The costs of publication of this article were de- frayed in part by the payment of page charges. This article must there- fore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. to the GenBankTM/EMBL Data Bank with accession number(s1 X62373. The nucleotide sequence(s) reported in this paper has been submitted ultad de Ciencias, 14071 Cordoba, Spain. ll Present address: Dept. de Bioquimica y Biologia Molecular, Fac- Biotecnologia, CSIC, Campus UniversidadAut6noma de Madrid, Canto 11 To whom correspondence should be addressed: Centro Nacional de Blanco, 28049 Madrid, Spain. Tel.: 34-1-5854548; Fax: 34-1-5854506; E-mail: [email protected]. pyran ring, leading to the benzoisochromanequinone structure. Since the first report of the chemical structure of actino- rhodin (1) (see Fig. 2) (1) as a polyketide compound (2) which can act as a pH indicator (red at acid pH and blue at basic pH) and the characterization of mutations in the biosynthetic genes (3, 41, the act cluster of Streptomyces coelicolor A3(2) has be- come a favored model for analyzing the genetic of antibiotic biosynthesis. Chemical characterization of intermediates and shunt productsof the actinorhodin pathway (5, 6) hasled to a scheme for its biosynthesis in which each half of the dimeric molecule comes from 8 acetate units (7). Following the cloning of the whole biosynthetic pathway as a continuous segment of chromosomal DNA (a), the positions of the actgenes recognized by mutations were determined on the cloned DNA (9). The cluster of act genes (Fig. 1) has been considered for historical reasons as a set of five “regions”: (a) the act11 region, carrying a well documented positive regulator of the act genes and genes for antibiotic export (10); (b) the act111 gene involved in polyketide reduction (7, 11); (e) the act& actVII, actIV, and actVB regions, which turned out to contain genes for the earliest biosynthetic steps (polyketide chain as- sembly: act1 and actVII) and some subsequent steps (putative dehydrase that would act on the nascent cyclized polyketide chain (actIV) and a “dimerase” that would function perhaps as the penultimate biosynthetic step (actVB) (12); (d) the actVA region, containing a further putative export gene and a series of genes for oxidationhydroxylation at middle to late stages in the pathway (13); and (e) the actVI region which is the subject of this paper. The actVI region was located, by genetic mapping, near the left-hand end of the act cluster (9), around PstI site 2 (Fig. 1); this region represents one end of the cloned act DNA and seems to control middle steps of actinorhodin biosynthesis. Based on chemical characterization of the major metabolite produced by the two available actVI mutants, aloesaponarinI1 (2) (see Fig. 2) (7), it was postulated thatone or more proteins, encoded by the actVI region, would recognize the product of the reaction catalyzed by the ActIV protein and, by a dehydration and re- duction at C-3/C-15, would form the heterocyclic intermediate of actinorhodin biosynthesis. Thisintermediate (3) (Fig. 2) would later undergoC-6 and C-8 oxidationhydroxylation (Fig. 2) by the action of the actVA gene products (13). Here we report the sequence of the DNA that includes the position to which the actVI mutations were previously mapped (9) and discuss the involvement of the putative gene products in a proposed scheme for the biosynthetic pathway. With the DNA sequence reported in this work the sequence of the act cluster is complete. 24854
Transcript of THE JOURNAL OF Vol 269, No 40, Issue pp 24854-24863, 1994 ... · DNA Sequence and Functions of the...

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1994 by The American Society for Biochemistry and Molecular Biology, Inc
Vol . 269, No . 40, Issue of October 7, pp , 24854-24863, 1994 Printed in U.S.A.
DNA Sequence and Functions of the actVI Region of the Actinorhodin Biosynthetic Gene Cluster of Streptomyces coelicolor A3(2)*
(Received for publication, April 15, 1994, and in revised form, July 18, 1994)
Miguel A. Fernandez-Morenol, Eduardo Martinez$, Jose L. CaballeroPn, Koji IchinoseQ, David A Hopwood§, and Francisco Malpartidalll From the SCentro Nacional de Biotecnologia, Consejo Superior de Investigaciones Cientificas, Campus Universidad Autonoma de Madrid, Cantoblanco 28049, Madrid, Spain and the §John Znnes Institute, John Znnes Center, Colney Lane, Norwich NR4 7UH, United Kingdom
Six open reading frames (ORFs) were identified by DNA sequencing of 5.7 kilobase pairs at the left end of the act cluster (the so-called “actVI region”), in the or- der: ORFB, ORFA, ORF1, ORF2, ORF3, ORF4. ORF1-4 are transcribed rightward and in the same direction as the ORFs of the actVA region which lies to the right of the actVI region, whereas ORFA and ORFB run in the opposite direction. By complementation of mutants and gene disruption of the wild type strain, the two previ- ously genetically characterized actVI mutations were assigned to ORF1. Although disruption of ORFB and ORF4, using +C3l derivatives, did not cause any obvious change in actinorhodin production, defects in actino- rhodin synthesis were obtained by insertional inactiva- tion of ORFA, ORF1, ORF2, or ORF3. RNA analysis within the ORFUORFA intergenic region showed over- lapping divergent promoters, at least one of which is under the control of the actII-ORF4 gene product, the transcriptional activator of the act cluster. Data base searches with the deduced products of ORFB and ORF3 failed to show any significant similarities with other known proteins. The deduced product of ORFA strongly resembles those of genes of unknown function from Sac- charopolyspora hirsuta and Streptomyces roseofulvus, located within polyketide synthase clusters. The ORFl product strongly resembles P-hydroxyacyl-CoA dehy- drogenases of bacteria and mammals and the ORF2 and ORF4 products resemble each other and enoyl reduc- tases from bacteria, animals, and plants, with a highly conserved cofactor-binding domain. These findings strongly suggest that the actVI region is involved in catalyzing reduction processes that determine the two stereochemical configurations at C-3/C-15 during acti- norhodin biosynthesis. A scheme is proposed for the middle steps of the biosynthesis, that is formation of the
CICYT (BI0090-0760 and BI093-1181), EC (BIOT-CT91-0255, SMA) * This research was supported by grants (to F. M.) from the Spanish
and Proyecto Concertado Smith Kline and Beecham, S. A./CSIC (CDTI, number 930139) and by grants-in-aid (to D. A. H.) to the John Innes Institute from the Agricultural and Food Research Council and the John Innes Foundation and Grant BIOT-CT91-0255 from the EC BRIDGE Programme. The costs of publication of this article were de- frayed in part by the payment of page charges. This article must there- fore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
to the GenBankTM/EMBL Data Bank with accession number(s1 X62373. The nucleotide sequence(s) reported in this paper has been submitted
ultad de Ciencias, 14071 Cordoba, Spain. ll Present address: Dept. de Bioquimica y Biologia Molecular, Fac-
Biotecnologia, CSIC, Campus UniversidadAut6noma de Madrid, Canto 11 To whom correspondence should be addressed: Centro Nacional de
Blanco, 28049 Madrid, Spain. Tel.: 34-1-5854548; Fax: 34-1-5854506; E-mail: [email protected].
pyran ring, leading to the benzoisochromanequinone structure.
Since the first report of the chemical structure of actino- rhodin (1) (see Fig. 2) (1) as a polyketide compound (2) which can act as a pH indicator (red at acid pH and blue a t basic pH) and the characterization of mutations in the biosynthetic genes (3, 41, the act cluster of Streptomyces coelicolor A3(2) has be- come a favored model for analyzing the genetic of antibiotic biosynthesis. Chemical characterization of intermediates and shunt products of the actinorhodin pathway (5, 6) has led to a scheme for its biosynthesis in which each half of the dimeric molecule comes from 8 acetate units (7).
Following the cloning of the whole biosynthetic pathway as a continuous segment of chromosomal DNA (a), the positions of the act genes recognized by mutations were determined on the cloned DNA (9). The cluster of act genes (Fig. 1) has been considered for historical reasons as a set of five “regions”: ( a ) the act11 region, carrying a well documented positive regulator of the act genes and genes for antibiotic export (10); ( b ) the act111 gene involved in polyketide reduction (7 , 11); (e) the act& actVII, actIV, and actVB regions, which turned out to contain genes for the earliest biosynthetic steps (polyketide chain as- sembly: act1 and actVII) and some subsequent steps (putative dehydrase that would act on the nascent cyclized polyketide chain (actIV) and a “dimerase” that would function perhaps as the penultimate biosynthetic step (actVB) (12); ( d ) the actVA region, containing a further putative export gene and a series of genes for oxidationhydroxylation a t middle to late stages in the pathway (13); and (e) the actVI region which is the subject of this paper.
The actVI region was located, by genetic mapping, near the left-hand end of the act cluster (9), around PstI site 2 (Fig. 1); this region represents one end of the cloned act DNA and seems to control middle steps of actinorhodin biosynthesis. Based on chemical characterization of the major metabolite produced by the two available actVI mutants, aloesaponarin I1 (2) (see Fig. 2) (7), it was postulated that one or more proteins, encoded by the actVI region, would recognize the product of the reaction catalyzed by the ActIV protein and, by a dehydration and re- duction at C-3/C-15, would form the heterocyclic intermediate of actinorhodin biosynthesis. This intermediate (3) (Fig. 2) would later undergo C-6 and C-8 oxidationhydroxylation (Fig. 2) by the action of the actVA gene products (13). Here we report the sequence of the DNA that includes the position to which the actVI mutations were previously mapped (9) and discuss the involvement of the putative gene products in a proposed scheme for the biosynthetic pathway. With the DNA sequence reported in this work the sequence of the act cluster is complete.
24854

DNA Sequence and Functions of the actV1 Region 24855
actVI actVA act11 act111 act1 actVII actIV actVB
1 kb 1 kb
< ORFB I (ORFAI I ORFl 1 ORF2 } [x>[ ORF4 ) 0ME22 - 0AB25.2 U 0A024 u
0ME23 U @ME28 - 0ME29 - 0ME12 I I
0ME11
0ME25
0ME24
0ME20
0ME21
022 B159 ComDlementation
5% 4%
95% 88%
>958 >95%
>95% >95%
>95% >95%
organization of the six ORFs in the actVI region as revealed by DNA sequencing. Restriction sites are numbered as reported previously (9); decimal FIG. 1. Restriction map of the whole act cluster. Restriction map of the act cluster with the approximate locations of act mutations and
numbers define further sites assigned in the present work. The solid bar shows the extent of the sequenced DNA fragment; ORFs are indicated by large open arrows; thick lines represent the fragments cloned in the +C31-derived KC515 and PM1 vectors for gene disruption and comple- mentation studies. All the fragments, except BamHI-PstI (sites 1-21, were first cloned into pIJ2925 and then rescued from the resulting plasmid with BglII and ligated to BglII-digested KC515 or PM1. The complementation results for strains B22 and B159 with the set of recombinant phages are indicated as percentages of lysogens with wild type pigmentation out of the total number of transductants.
MATERIALS AND METHODS Bacterial Strains-The Escherichia coli strains were JMlOl (14),
XL1-blue (151, and JMllO (14). The S. coelicolor A3(2) strains were 51501 ( h i d l , uruAI, strAI, pgl, SCPl-, SCP2-) (161, B22 ( h i d , u r d l , strAl, SCP1-, SCPB’, act(VI)l22) (41, and B159 (hisAl, u r d l , s t r A l ,
TK21 (str-6 SLP2‘ SLP3-) (171. SCP1-, SCPB’, act(VI)259) (4). The Streptomyces liuidans 66 strain was
Plasmids and Bacteriophages-The E. coli plasmids were pUC19 (141, pIJ2925 (181, and pSU19 (19). E. coli M13 derivative phages mp18 and mp19 (14) were used for DNAsequencing. The Streptomyces phage vectors were the +C31 derivatives KC515 (20) and PM1 (9).
Media, Culture Conditions, and Microbiological Procedures-Strep- tomyces manipulations were as described previously (21). Thiostrepton (a gift of S. J. Lucania, Bristol-Myers Squibb Research Institute, Prince- ton, NJ) was used a t a concentration of 50 pg/ml in agar medium and 10 pg/ml in broth cultures. Hygromycin B (Sigma, catalog number H2638) was used at 200 pg/ml in solid media and 50 pg/ml in liquid media. E. coli strains were grown on L agar or in L broth (22).
DNA Sequencing-DNA sequencing was by the dideoxy-chain termi- nation method (231, and 96% of the sequence was determined from both strands; we used the 7-deaza-dGTP reagent kit from U. S. Biochemical Corp. (catalog number 707501, following the manufacturer’s recommen- dations. Convenient DNA fragments were previously cloned in either M13mp18 or M13mp19 vectors from suitable restriction fragments or generated by Ex0111 digestion (24). Identification of DNA sequences in
method (251. S1 protection experiments was carried out by the Maxam and Gilbert
Computer Analysis of Sequences-The DNA sequence was analyzed using the software programs of the University of Wisconsin Genetic
Computer Group (Version 7.3-AXP) (26): analysis for open reading frames used CODONPREFERENCE with a codon usage table made from 100 Streptomyces genes (27); comparisons of sequences were made against the EMBL nucleic acid data base (daily updated) and the Swissprot data base (Release 27.0, October 19931, using FASTA, TFASTA, and BESTFIT. Protein alignments were made using PILEUP and PRETTY.
Gene Disruption and Mutant Assignments-For gene disruption and mutant assignments, we used insert-directed recombination (28), using either S. coelicolor act+ strain 51501 or the two S. coelicolor actVI mutants as hosts.
DNA and RNA Manipulations-Isolation, cloning, and manipulation of nucleic acids were as described for Streptomyces (211 and E. coli (22).
Polymerase chain reactions (29) were carried out using Thermostase according to the manufacturer’s recommendations.
For high-resolution S1 mapping, the method of Murray (30) was used. For ORFA’ mRNA mapping, the probe was made as follows: the 312-base pair NaeI-SmaI fragment (nucleotides 1917-22281 (see Fig. 7) was previously cloned in the HincII site of pIJ2925 to yield pMF2001.2; the resulting plasmid was digested with EagI (nucleotide 1927), the 5’ ends were labeled with Lr-”PlATP, and the probe was rescued by a second digestion from the vector as a 335-base pair fragment, labeled at nucleotide position 1928 of the strand complementary to the ORFA mRNA. For the ORFl probe, a synthetic oligonucleotide (5 ’ CTGAA- CAGGTTGATCCATCCCAGG 3’) was labeled at the 5’ end (nucleotide 2144 in the template strand) and used to amplify by polymerase chain reaction on pMF2001.2, a 279-base pair DNA fragment labeled in the
_____ The abbreviation used is: ORF, open reading frame.

24856 DNA Sequence and Functions of the actVI Region
1 Acetate + 7 Malonates
FIG. 2. Putative biosynthetic path- way for actinorhodin. Reactions are based on references (5 , 7); the structures of unnamed intermediates are hypotheti- cal; aloesaponarin I1 is a shunt product of the actVI mutants (7).
template strand. RNA was extracte d from sh:
0
COlH HO
OH 1
act VI1
3HzO actIV
H20d act VI [ 01
~H
I I / OH
C02H 0
aclVA intermediate (3) Aloesaponarin I1 (2)
I act VI, VA, VB
u Actinorhodin (1)
%ken SY liquid cultures (31)grown for 2 days using pregerminated spores (21).
RESULTS
DNA Sequence of the aetVI Region Starting at BarnHI site 1 (9) (Fig. l ) , we sequenced 5.67 kb of
the DNA, covering the whole actVI region and overlapping the adjacent actVA region previously sequenced (13). Computer- assisted analysis of the sequence revealed six ORFs, which
were named from left to right: ORFB, ORFA, ORF1, ORFZ, ORF3, and ORF4. The last four ORFs are transcribed right- ward, toward the actVA region, whereas ORFs A and B run divergently from ORFl toward the left-hand end of the act cluster (Fig. 1); the most relevant features, deduced from the DNA sequence, are summarized in Table I.
The translation start point for each ORF was tentatively lo- cated using two initial criteria: the overall distribution of GC content in the third position of the codons (32) and the codon

DNA Sequence and Functions of the actVI Region 24857 TABLE I
Relevant features ofthe 5.67-kb actVI region deduced from its DNA sequence ”No” below “rbs” (ribosome-binding site) entry indicates absence of significant similarity between the region upstream of the putative start codon
Notice that ORFB and ORFA run divergently to the others. and the 16 S rRNA sequence (35). Numbers below “StadStop codons” shows the first and last nucleotide of the start and stop codons respectively.
ORF rbs StartiStoD codons Amino acidsIM. Predicted function
ORFA ORFB No
No ORFl No ORF2 2990 AGGAGGAGA 2998 ORF3 4046 AGGAGA 4051 ORF4 4728 GGAGA 4732
FIG. 3. Alignments of the putative p a p fold that binds the ADP moiety of NAD or FAD of ActVI-ORF1 with
j3-hydroxybutyryl-CoA dehydrogenase other proteins. Proteins aligned are:
from C. acetobutylicum (CABHBD), enoyl-CoA hydratase:3-hydroxyacyl-CoA
(ECH), L-3-hydroxyacyl-CoA dehydrogen- dehydrogenase from rat peroxisomes
ase from pig heart muscle (HCDH), 3-hy- droxyacyl-CoA dehydrogenase from E. coli fadB (FADB), and enoyl-CoA hy- dratase:3-hydroxyacyl-CoA dehydrogen- ase from P fragi (FAOA). Amino acids are numbered according to their original positions in the proteins.
AGT 1213/ 29 GTA AGT 2009/1569 GTA ATG 2083/3006 TGA GTG 3003/3992 TGA ATG 4059/4703 TAG ATG 4737/5657 TGA
ORF 1 3
CABHBD 3
ECH 2 97
HCDH 17
FADB 315
FAOA 316
usage within the putative ORFs (27). For ORFA and ORFl there are several putative start codons and so an additional criterion was used: the similarities observed between the pu- tative ORFA product with two other deduced products of genes in polyketide synthase clusters and for ORFl product with other known proteins (see below).
ORFl stop codon overlaps the putative start codon for ORF2 (GTGA), suggesting translational coupling, as in many other Streptomyces genes (12, 33, 34). Similarly ORF4 stop codon overlaps the start codon of the adjacent actVA-ORF1 gene, the putative second resistance gene in the actVA region (13).
The start codons for ORF2, ORF3, and ORF4 (but not those for ORFA, ORFB, and ORFl) are preceded by a good potential ribosome binding site (see Table I) (35) at a suitable distance, representing an additional criterion in assigning the transla- tional start codon for these ORFs.
Mutant Assignment and Gene Disruption In order to assign functions to the deduced ORFs, recombi-
nant phages containing DNA fragments from the actVI region of the wild type strain (Fig. 1) were constructed for use to disrupt each of the deduced ORFs by insertional inactivation or to complement the available genetically characterized actVI mutants (4). After construction of the recombinants by trans- fection of S. lividans protoplasts, and infection of Act+ S. coeli- color, lysogens of the latter were obtained by replica-plating to thiostrepton-containing plates (selecting the tsr marker of the phage vector). The expected structures of all lysogens were confirmed by Southern hybridization. These experiments also revealed that BamHI site 1 (Fig. 1) of the act cluster as origi- nally cloned is not present in the chromosome of wild type S. coelicolor; probably it was generated during the cloning after ligation of a Sau3Afragment into the BamHI site of pIJ922 (8).
All the lysogens carrying +ME22 (HincII fragment, sites 1.1 to 1.3, internal to the coding region of ORFB) and +ME29 (SphI-HincII fragment, sites 3.3 to 4.1, internal to ORF4) were Act+. In contrast, lysogens carrying 4AB25.2 (a 266-base pairs ApaLI-NaeI fragment, nucleotides 1650-1916, internal to ORFA), +ME23 (XhoI fragment, sites 1.6 to 2.1, internal to ORFl), +ME28 (BclI-PuuII fragment, sites 2.4 to 2.6, internal
394/41,243 146/16,113
Unknown
307/33,115 Unknown 3-Hydroxyacyl-CoA dehydrogenase
329/34,079 214123,105
Alcohol dehydrogenase Unknown
306/31,552 Alcohol dehydrogenase
TVTVIGAGTIGLGWINLFSARGLTVRVNSRRPD
KIFVLGAGTMGAGIVQAFAQKGYEVIVRDIKDE
SVGVLGLGTMGRGIAISFARVGISWAVESDPK
HVTVIGGGLMGAGIAQVAAATGHTWLVDQTED
QAAVLGAGIMGGGIAYQSAWKGVPVVMKDINDK
QAAVLGAGIMGGGIAYQSASKGTPILMKDINEH - 1 1-
P a P
35
35
330
50
348
349
to ORF2), +AB24 (partial BssHII-ApaI fragment, nucleotides 4141-4629, internal to ORF3), and +ME12 (PstI-BamHI frag- ment, sites 2 to 3, containing the carboxyl terminus of ORF1, the complete ORF2 and the amino terminus of ORF3) were affected in actinorhodin (blue pigment) production. 4AB25.2 lysogens (ORFA) showed an obvious reduction in blue pigment production compared with the wild type strain. +ME23 lyso- gens (ORF1) showed a light brown phenotype indistinguishable from that of the actVI mutants originally described (4). +ME28 lysogens (ORF2) produced a brownish pigment resembling that produced by actVA mutants (4). +ME12 and +AB24 (ORF3) lysogens produced a reddish pigment which, after exposure to ammonia fumes, became first yellow/greenish and finally blue after 24-48 h. These results clearly indicate that at least ORFA, ORF1, ORF2, and ORF3 are involved in actinorhodin biosynthesis, with the last three controlling specific biosyn- thetic steps, whereas disruption of ORFB and ORF4 did not prevent actinorhodin production.
Two actV1 mutations were originally isolated after ultravio- let irradiation of an Act+ S. coelicolor strain; both of them were phenotypically classified as producers of a “light brown” diffus- ible pigment (4). To localize the actVI mutations the following +C31-derived recombinant phages were constructed as speci- fied in the caption to Fig. 1: +ME11 (carrying the BamHI-PstI fragment, sites 1 to 21, +ME20 (carrying the BamHI-BclI frag- ment, sites 1 to 2.4), and +ME21 (carrying the BamHI frag- ment, sites 1 to 3). The recombinant phages were introduced into the actVI mutants B22 and B E 9 by insert-directed recom- bination. Recombination between the cloned fragment and the genome can take place either upstream or downstream of the mutation. In either case, the wild type phenotype should be restored only if the cloned fragment carries an intact copy of the mutated gene. The results (Fig. 1) show that essentially all the lysogens (>95%) carrying +ME21 and +ME20 were Act+, whereas with +ME11 only around 5% were Act+. With +ME24 (an ExoIII-derived clone, carrying a DNA fragment from nucle- otides 1 to 3041 and, therefore, the complete ORFl), and +ME25 (an ExoIII-derived clone, carrying nucleotides 1-2964, and so lacking 13 amino acids of ORF1) the frequencies were more than 95% or slightly below 95% Act’, respectively. These

24858 DNA Sequence and Functions of the actVI Region
01 oa 03 04 0 5 06 07 08 09 10
C
NAD(H)-biading motif
100 I PWLGG DWGRITAVG PGVTALRPGD RVG.AFTLYG AYA..EQVW
1 1 3 9
PQIIGN EFAGWERTG PEATGFAPGD EVL.GFAVLQ SGTELLAVGA
PSGLGT EAAGIVSWG SGVKHIKAGD RWYAQSALG AYSSVHNIIA PVTLGH EIAGKIEEVG DEWGYSKGD LVAVNPWQGE GNCYYCRIGE
EPQLGM DFVGWTAVG EGV?GHQ.VG DRVGGFSEGG CWRTFLTCDA WLDMGT EAAGWTAVG PDVDAFA.PG DRVLGLFQGA .FAPIAVTDH
PDAIPG KWASRDCMLG MEFSGRDKCG RRVMGLVPAE GLATSVLLSP PYTPGS DVAGVIEAVG DNASAFKKGD RVFTSSTISG GYAEYALAAD
PVIAGH EAAGIVESIG EGVTTKQPGD KVI.PLFTPQ CGKCRVCKHP PMVPGH EWGEVLEVG SNVTRFKVGE IVGVGLLVGC CKSCRACDSE p".Q- B--Q-V-."O ......".. "...."" ~.."~....
NADP(H)-biading moiif
r 2 2 4 I I I ' I
KGETVLIHAA AGGVGHIAVQ LAFWAGAGLI IGTAGAAAKR E F AFW.... T GADAWDHRS AHWADEVREL TGGRGPDLIL 303
PGDTVLVHAA AGGVGSLAVQ LW7ERGGT.A IGSAG.EHNH DYLRS . . . . L GALPVAY..G PGFADRVRAL ARTASTPPLD PTKTLLWGA GGGLGTMAVQ IAKAVSGATI IGVDVREEAV EAAKR .... A GADWINASM QDPLAEIRRI TESKGVDAVI PDEQFLFHAA AGGVGLIACQ WAKALGAKL. IGTVGTAQKA QSALK . . . . A GAWQVINYRE EDLVERLKEI TGGKKVRWY AGQSVLIHRA AGGVGMAAVA LARRAGAEV. LATAG.PAKH GTLRAL..GL DDEHIASSRE TGFARKFRER lGGRGVDWL AGDKVLIHSA TGGVGQAAIS IARAKGAEI. FATAGNPAKR AMLRDM..GV ..EHVYDSRS VEFAEQIRRD TEYGVDIVL
HGETVLIHSG SGGVGQAAIS IALSLGCRV. FTTVGSAEKR AYLQARFPQL DDTSFANSRD TSFEQHVLLH EGKGVDLVL AGESVLVHGA SGGVGLAACQ IARAYGLKI. LGTAGTEEGQ KIVLQ . . . . N GAHEVFNHRE VNYIDKIKKY VGEKGIDIII
QGSTCAVFGL .GGVGLSVIMGCKAAGAAFS IGVDINKDKF AKAKE . . . . V GATECVNPQ. .... D..... . . _ . . . . . . . PGLRGGILGL .GGVGHMGVKVAKALGHHVT VISSSDKKKK EALED . . . . L GADNYLVSS. . . . . DTVGMQ EAADSLDYII -Q".LTa-A .OOVO"AV. -A.A.Q"- ""Q".K. ..._.~.... ".V"". """D.V.
Zinc-eootaining motif
FIG. 4. Alignments of the actVI-ORF2 and actVI-ORF4 gene products with other proteins. The compared proteins are: ActVI-ORF2 (01 1, ActVI-ORF4 (02), S. solfataricus NAD(+)-dependent alcohol dehydrogenase (55) (03), E. coli quinone oxidoreductase (04) (53), S. erythraea (05) 6-deoxyerythronolide B synthase (49), M. tuberculosis var. bovis BCG mycocerosic acid synthase (06) (50), Homo sapiens c-crystallidquinone reductase (07) (54), rat fatty acid synthase (08) (511, horse alcohol deydrogenase (09) (52); and Medicago sativa cinnamyl alcohol dehydrogenase (10) (J. Van Doorsselaere; EMBL accession number: 219573). Conditions: plurality = 7; threshold = 1; average weight = 1; average match = 0.54; average mismatch = 0.4; C = consensus. Only part of the sequence is shown for each protein. The amino acid numbering has been given by the computer program; the amino acid position is that below thelast digit.
results suggest that ORFl corresponds to the mutated gene in the actVI mutants, because the Act+ phenotype observed with +ME20, +ME21, and +ME24 seems to be independent of the position of crossing over, as expected if all the recombinant phages carry a whole copy of the wild type gene. With +ME11, the low frequency of "complementation" observed with the two actVI mutants suggests that recombination must take place in ORFl (the only partial ORF within the cloned fragment) and that both mutations would lie near the 3' end of the cloned fragment. For +ME25, the observed frequencies of complemen- tation are higher than expected; ORFl is incomplete (lacking 13 amino acids from the COOH terminus) and so the lysogens generated by recombination upstream of the mutation, ex- pected to be the most frequent, would generate a truncated protein in one side of the integrated recombinant phage and the mutated ORF on the other; perhaps the truncated ActVI pro- tein is still enzymatically active.
Deduced Functions of the actVI Region Gene Products ORFB and ORF3"Data base searches with the translated
products of ORFB and ORF3 gave no similarities with other known proteins and therefore no clue as to their possible functions.
ORFA-Data base comparison of the actVI-ORFA product showed a strong resemblance (55% identity, 75% similarity) and an end-to-end alignment with a gene product (ORF1) with unknown function found within a gene cluster encoding a Type I1 polyketide synthase from Saccharopolyspora hirsuta (36) and with a similar gene product (ORFX) in the frenolicin polyketide synthase gene cluster of Streptomyces roseofuluus (37). Although actVI-ORFA seems to play a role in actinorhodin biosynthesis (as deduced by the insertional inactivation re- sults) its function is unknown.
ORF1-The actVI-ORF1 product clearly controls an essen- tial step in actinorhodin biosynthesis, and, unlike ORFA, its deduced gene product shows similarities with other proteins of known function, such as: P-hydroxybutyryl-CoAdehydrogenase from Clostridium acetobutylicum (38) (26% identity, 52% sim- ilarity); A-crystallin from rabbit lens (39) (32% identity, 56% similarity); L-3-hydroxyacyl-CoA dehydrogenase from pig heart muscle (40) (28% identity, 50% similarity); E. coli FadB protein (41, 42) (23% identity, 49% similarity, with the putative 3-hy- droxyacyl-CoA dehydrogenase domain); the bifunctional rat peroxisomal enoyl-CoA hydratase:3-hydroxyacyl-CoA dehydro- genase (43) (24% identity, 52% similarity with the putative 3-hydroxyacyl-CoA dehydrogenase domain); and the Pseudo- monas fragi FaoA protein (44) (25% identity, 49% similarity with the putative 3-hydroxyacyl-CoA dehydrogenase domain).
40 69 01 .FPWLGGDV VGRITAVGPG VTALRPGDRV 02 EFPQIIGNEF AGWERTGPE ATGFAPGDEV 03 .FPVILGHEA AGWESVGEK VTLLKPGDAV 04 .FPIILGHEA AGVIESVGEK VTSLKPGDAV 05 .LPVIAGHEA AGIVESIGEG VTTVRPGDKV 06 .LPVILGHEA AGIVESIGEG VTTVKPGDKV 07 .LPMILGHEA AGWESTGEG VTSLKPGDKV 08 .FPRILGHEA AGIVESVGEG VTDLAPGDHV 09 .FPRIFGHEA GGIVESVGEG VTDLAPGDHV 10 .FPRILGHEA GGIVESVGEG VTDVAPGDHV 11 NYPWPGHEI AGWTAVGSE VTKYRQGDRV 12 .FPWLGHEG AGIVESVGPG VTEFQPGEKV 13 .FPCVLGHEG AGIVESVGDD VITVKPGDHV 14 KLPLVGGHEG AGVWATGDL VNEFEVGDHA 15 KMPLIGGHEG AGWKVGAG VTRLKIGDRV 16 GLTV..GHEP VGIIEKLGSA VTGYREGQRV 17 K..ADMGTEA AGWTAVGPD VDAFAPGDRV 18 LLPYTPGSDV AGVIEAVGDN ASAFKKGDRV 19 SLPSGLGTE. AGIVSKVGSG VKHIKAGDRV
Consensus -PP-I-G-E- -GIV--VG-- V----- OD-V
Zinc-containing GHEx xGxxxxxGxx V Fingerprint (T) ( A r c )
FIG. 5. Alignments at the NH, terminus of the ActVI-ORF2 and ActVI-ORF4 proteins with alcohol dehydrogenases. 01, ActVI-
genase a chain (69); 04, chicken alcohol dehydrogenase I (70); 05, horse ORF2; 02, ActVI-ORF4; 03, Coturnix coturnixjaponica alcohol dehydro-
alcohol dehydrogenase (52); 06, rabbit alcohol dehydrogenase I (71); 07, Alligator mississippiensis alcohol dehydrogenase (72); 08, Lgcopersicon esculentum (tomato) alcohol dehydrogenase (73); 09, Oryza sativa (rice) alcohol dehydrogenase (74); 10, Zea mays (maize) alcohol dehydrogen- ase (75); 11, M. tuberculosis var. bovis BCG mycocerosic acid synthase (50); 12, Gadus callarias alcohol dehydrogenase (76); 13, Saccharomy- ces cerevisiue alcohol dehydrogenase (77); 14, Aspergillus nzger alcohol
pombe alcohol dehydrogenase (78); 16, Alcaligenes eutrophus alcohol dehydrogenase (Nucleic Patents N70814); 15, Schizosaccharomyces
dehydrogenase (79); 17,s. erythraea 6-deoxyerythronolide B synthase (49); 18, Homo sapiens 6-crystallinlquinone reductase (54); 19, E. coli quinone oxidoreductase (53). A putative zinc-containing motif for alco- hol dehydrogenases is indicated below the sequences, based on the consensus from PROSITE data base (60). Residues within parentheses indicate alternative amino acids. Conditions used were: plurality, 15; threshold, 1.00; average weight, 1.00; average match, 0.54. Some of the mismatches were manually introduced in order to maximized the con- served motif. Amino acid numbering and positioning are given as in Fig. 4.
Comparisons of the ORFl product with these proteins revealed highly conserved stretches; when the whole proteins are com- pared, 24 residues are conserved across the six proteins (the A-crystallin protein (39) was excluded in the comparison, be-

DNA Sequence and Functions of the actVI Region 24859
FIG. 6. High resolution S1 mapping of transcription start points in the actVI intergenic region. A, transcrip- tion initiation point for actVI-ORF1 poly- cistronic transcript. B , transcription ini- tiation point for the actVI-ORFA transcript. Each start point is indicated by asterisks within the sequence, as de- rived from Maxam and Gilbert reactions. Arrows point in the direction of transcrip- tion.
ORFl
mRNA c
3' 5' C G C G C G G C
T A C G
G C G C C G A T
C G C G
G C G C * C G
T
c# ' W
c. c.
2.4
B 5 '
mRNA
I ORFA
NaeI GCCGGCGAAGTCACGGCCGTCCAGCTTCTGCATCTGGTGGCCGTAGAACGACAGGACCTCGGCGTACAGGCCGGTGGGAAGCGCGGT
CGGCCGCTTCAGTGCCGGCAGGTCGAAGACGTAGACCACCGGCATCTTGCTGTCCTGGAGCCGCA~TCCGGCCACCCTTCGCGCCA G A F D R G D L K Q M Q H G Y F S L V E A Y L G T P L A T
1 9 1 4 - - - - - - -+- - - - - - - - -+- - - - - - - - -+- - - - - - - - -+- - - - - - - - -+- - - - - - - - -+- - - - - - - - -+- - - - - - - - -+- - - - - - - - -+ 2000
mRNA ****+ G A T G G T C A T A C G G C C A C G G T G C C C G G T G G C C T C G A A G T T G G C G C T G C C A G G C T C G T C G C C A T G A G C A C
2 0 0 1 "-"-"-+---"""+""""-+""""-+""""-+""""-+""""-+""""-+---"--"+ 2 0 9 0 CTACCAGTATGCCGGTGCCAGCGGGCCACCGGAGCTTCAAGGGAGCTCGGCCC~GCTACCCGCGACGGTCCGAGCAGCGGTACTCGTG
**** mRNA (ORF1) M S T I T M (ORFA)
CGTGACAGTGATGGGGCGGGCACGATCGGCCTGGGATGGATAACCTGTTCAGCGCCCGCGGGCTGACCGTACGCGTCAACAGCCGTCG
GCACTGTCACTAGCCCCGCCCGTGCTAGCCGGACCCTACCTAGT~GACAAGTCGCGGGCGCCCGACTGGCATGCGCAGTTGTCGGCAGC V T V I G A G T I G L G W I N L F S A R G L T V R V N S R R
2 0 9 1 - - - - - - - - -+- - - - - - - - -+- - - - - - - - -+- - - - - - - - -+- - - - - - - - -+- - - - - - - - -+- - - - - - - - -+- - - - - - - - -+- - - - - - - - -+ 2180
GCCCGACGTCCGGCGCGTCGTGCACGAGGCGCTCGAGCTGTTCTCCCCGGG
CGGGCTGCAGGCCGCGCAGCACGTGCTCCGCGAGCTCGACAAGAGGGGCCC P D V R R V V H E A L E L F S P G
SmaI
2 1 8 1 - - - - - - - - -+- - - - - - - - -+- - - - - - - - -+- - - - - - - - -+- - - - - - - - -+-
FIG. 7. DNAsequence ofNaeI-SmaI restriction fragment containing the ORF'NORFl intergenic regi0n.Asterisk.s show the nucleotides corresponding to the 5' end of the mRNAs as suggested by nuclease S1 digestion. Thick arrows indicate the direction of transcription for both genes. Amino acids are represented as single letter code below the DNA sequence. Numbering starts at BamHI (position 1) (see Fig. 1)
cause its biochemical function is not clear). I t is notable that, of these 24 highly conserved residues, 13 are glycines, whose small size allows them to occupy positions where space limita- tions preclude alternative residues (38, 45), and five are acidic residues, whose conservation is apparently more critical than that of basic ones (45). These kinds of conserved residues are considered important for correct maintenance of the secondary structure and the catalytic properties of alcohol dehydrogen- ases (45). Apart from the A-crystallin protein (39), the best agreement found for ORFl was with P-hydroxybutyryl-CoA de- hydrogenase, concerned with butanol biosynthesis (38). The end-to-end similarity provides additional support for choosing the ATG (position 2083) as the translational start codon for ORF1; only a few gaps are needed to make the best alignment between the two proteins.
The most relevant similarities were found near the NH, ter- minus of the ORFl product. Fig. 3 shows an alignment of this
segment with regions of the five proteins, where all have a "fingerprint" characteristic of P (Y P folds that can bind the ADP moiety of NAD or FAD (43, 46). Using the scoring method of Wierenga et al. (461, the sequence of 33 amino acids in the fingerprint of ORFl would score 8 out of 11, including 3 gly- cines and 1 acidic residue, which are requirements of the P a /3 fold; if we consider the possible variation of the length in the fingerprint internal loop (46, 47), Asp35 of the ORFl protein could be the acidic residue postulated as required for the j3 a P fold. The distance of this residue from the glycine core is not only greaterin several NAD(H)/F'AD(H)-dependent dehydrogen- ases, but can be absent, with other acidic residues close to positions 64 and 87 being responsible for hydrogen bonding to the coenzyme (48); ORFl contains other acidic residues that could be candidates (Asp64, Asps1, and Asps2 and Glug2). In ad- dition, 7 out of the 33 amino acids within the fingerprint are identical, whereas others are equivalent, to residues in four
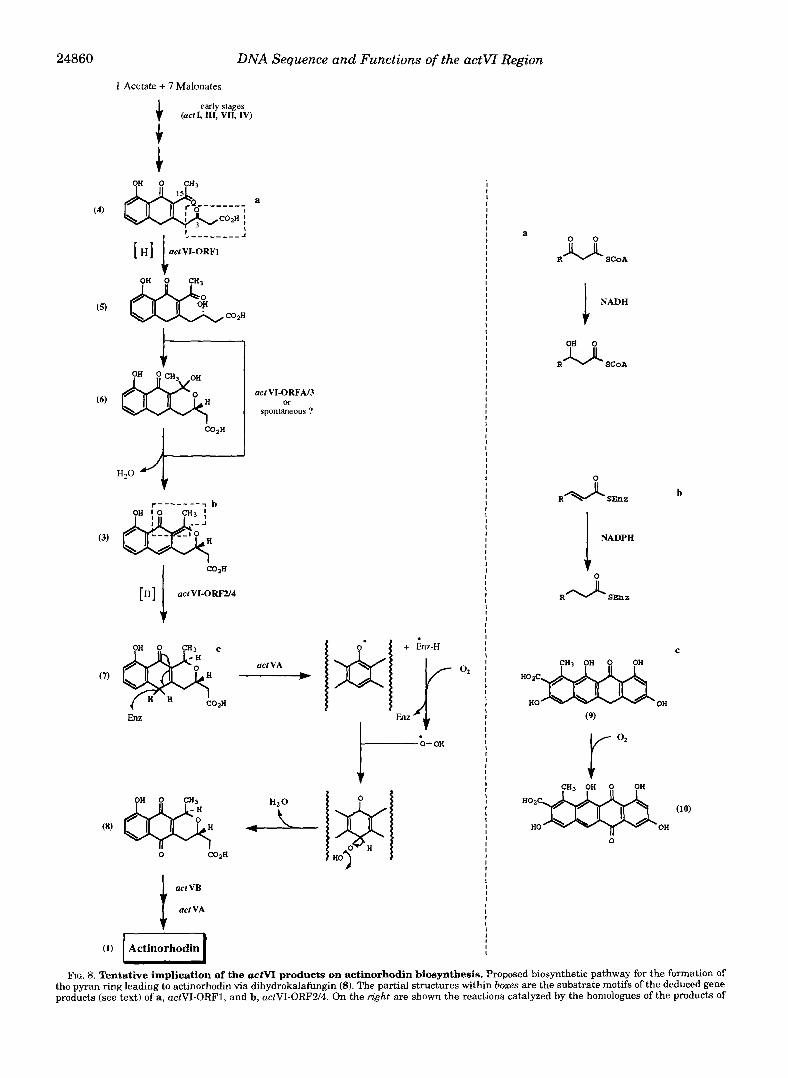
DNA Sequence and Functions of the actVI Region
1 Acetate + 7 Malonates
'""""J
acf VI-ORFA13
spontaneous ? or
r------- - b
0 C02H
acr VB
acf VA
+ Enz-H
0- OH
I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I 1 I I I I I I I I I I I l
a
R SCoA
1 NADH
OH 0
R SCoA
b
I NADPH
E
Y O2 CH3 OH 0 OH
0
FIG. 8. Tentative implication of the actvL products on actinorhodin biosynthesis. Proposed biosynthetic pathway for the formation of the pyran ring leading to actinorhodin via dihydrokalafungin (8). The partial structures within boxes are the substrate motifs of the deduced gene products (see text) of a, actVI-ORF1, and b, actVI-ORF2/4. On the right are shown the reactions catalyzed by the homologues of the products of

DNA Sequence and Functions of the actVI Region 24861
known dehydrogenases (46). ORFZ and ORF4-The actVI-ORF2 and actVI-ORF4 prod-
ucts resemble each other (35% identity, 51% similarity), with a good end-to-end alignment. Data base searches revealed high similarity with other gene products. Some of the most notable were: 6-deoxyerythronolide B synthase from Saccharopoly- sporu erythruea (49) (36% identity, 61% similarity); mycocerosic acid synthase from Mycobacterium tuberculosis var. bovis (50) (37% identity, 58% similarity); fatty acid synthase from rat (51) (25% identity, 49% similarity); alcohol deydrogenase from horse liver (52) (29% identity, 49% similarity); quinone oxidoreduc- tase from E. coli (53) (22% identity, 57% similarity); [-crystal- lidquinone reductase from human liver (54) (33% identity, 57% similarity); NAD(+)-dependent alcohol dehydrogenase from Sulfolobus solfuturicus (55) (32% identity, 53% similarity); and many other alcohol dehydrogenases. The best alignments were centered on the enoyl reductase domain (in the case of gene products corresponding to multifunctional proteins) and also within the putative coenzyme-binding motif of some alcohol dehydrogenases (Fig. 4).
The amino acid sequence of the ActVI-ORF2 protein reveals two putative cofactor-binding domains. One of them is close to the amino terminus of the protein, at positions 104-135 (Fig. 4). It shows 9 matches out of the 11 invariant residues follow- ing Wierenga et d ’ s method for NAD(H)/“AD(H)-dependent dehydrogenases (46). One of the two mismatches occurs within the nucleotide-binding fingerprint (being GXGXXA in- stead of GXGXXG), as in many NADP(H)-dependent dehydro- genases (56, 57) (exceptions to this motif have been reported for some NAD(H)/FAD(H)-dependent dehydrogenases (58)); the second mismatch is Gly’”, a position in which equivalent residues are present also in other NAD(H)/FAD(H)-dependent dehydrogenases (59).
A second putative cofactor-binding domain in the ActVI- ORF2 protein is that from Hisz3’ to Ala259 (Fig. 4) (present also in the ActVI-ORF4 product), with a score of 9 out of 11 highly conserved residues (46); the alignment within this region of the ActVI-ORF214 proteins against other dehydrogenases shows several structural features reported to be typical of the NADP(H)-binding motifs in some dehydrogenases (56): 1) the substitution of alanine instead of the third glycine in the core GXGXXG, and the presence of an additional alanine, with a fingerprint of 236GXGXK4XXXA245 (46, 56,571; 2) the existence of an acidic residue in the right end of the fingerprint, which typically forms the hydrogen bond with the 2”hydroxyl of the ADP moiety (46, 57) is not present; its position is preceded by two basic amino acids (LysZ6’ and and its consensus position is occupied by a hydrophobic residue (alanine) as in the well characterized NADP(H)-dependent glutathione reductase (46, 56). This change is expected to increase interactions be- tween the negatively charged 2’-phosphate of NADP and the corresponding amino acid residue of the fingerprint in those enzymes known to bind NADP(H); (3) the presence of the do- main RxxxxxR downstream of the core GXGXXAXXXG (posi- tions 291-297) which might confer specificity for NADP(H) (571, especially in enzymes belonging to the flavoprotein oxi- doreductase family (57). The best alignments observed with portions of this region of both the ActVI-ORF2 and ActVI-ORF4 proteins (Fig. 4) are with other proteins known to have enoyl reductase activities.
An additional interesting feature is the existence within the ActVI-ORF4 protein (and to a much lesser extent in the ActVI- ORF2 protein) near the NH, terminus of a stretch having high
resemblance with that of the zinc-containing alcohol dehydro- genases (as defined in the Prosite Data Base (60)). The consen- sus pattern G-H-E-X(2)-G-X(5)-G-X(2)-(VAC) is highly con- served, with only a conservative substitution of the histidine (known as the ligand of the catalytic zinc atom in this class of alcohol dehydrogenases) by asparagine within the fingerprint. The multiple alignments between this short region of the ActVI-ORF2/4 proteins and other known dehydrogenases are shown in Fig. 5. Whether or not the observed similarities in the amino acid sequence also reflect the catalytic properties of the ActVI-ORF2/4 proteins needs to be confirmed by enzymatic characterization.
Dunscription Analysis of the actVZ Region The transcription start points for actVI-ORF1 and actVI-
ORFA were determined by S1 mapping (see “Materials and Methods” for details of the probes) (Fig. 6). The start point of each transcript lies on the respectives start codons as four contiguous bases, one of them more prominent, with no room for a ribosome binding site, as in many other Streptomyces genes (61). The -35 region of the two promoters must overlap (Fig. 7).
DISCUSSION
The finding of an Act’ phenotype after directed mutation of ORFB, at the left end of the actVI region, would be consistent with its not forming part of the act cluster, although it does not exclude it, since mutation of other act genes (10) did not pre- vent blue pigment production. Certainly actVI-ORFB is not essential for the biosynthesis of actinorhodin and it may well be that actVI-ORFA defines the left end of the act cluster.
Complementation studies revealed that the two genetically characterized actVI mutations both lie in ORF1, and inser- tional inactivation confirmed the involvement of ORFl in the actinorhodin pathway. Three other ORFs (ORFA, ORF2, and ORF3) play different roles in actinorhodin production, because of the distinct phenotypes of each class of insertional mutant. ORFl,ORF2, and ORF3 are probably cotranscribed, since only insertions through internal fragments of transcripts can mu- tagenize, as +ME12 does. RNA analysis showed overlapping divergent promoters for this polycistronic mRNA and for the divergent ORFA transcript. The overlap of their -35 regions suggests that regulation of expression from both promoters is interrelated. The dependence of actVI-ORF1 transcription on the actII-ORF4 (activator) gene product (62) therefore probably implies dependence also of the ORFA promoter on actII-ORF4. Furthermore, when the fragment containing the ORFA-ORF1 intergenic region (Fig. 7) is introduced, on a high copy number plasmid, into the S. coelicolor wild type, the strain loses the ability to produce actinorhodin, and this effect is overcome when the copy number of uctII-ORF4 is increased.z
The deduced functions of some of the actVI ORFs, as well as previous knowledge of the uctVAregion (13), allow a scheme for the middle steps of actinorhodin biosynthesis to be proposed (Fig. 8). The early steps of chain assembly and ring closure, catalyzed by the uctI, uctIII, uctVII, and uctN gene products, would produce the substrate for the actVI gene products (4). This would be reduced stereospecifically at C-3, prior to cycli- zation. Based on the similarity of the partial structure (a) of (4)
a M. A. Ferndndez-Moreno, unpublished observation.
these actVI ORFs (a, p-hydroxyhutyryl-CoA dehydrogenase; b, enoyl reductase), whereas c shows the production of tetracenomycin D3 (10) from tetracenomycin F l ( 9 ) catalyzed by the monooxygenase encoded by tcrnH, the homologue of actVA-ORFG. (For simplicity, all the intermediates are shown as free acids, hut some might exist enzyme-bound as thioesters: RCOS-Enz. ).

24862 DNA Sequence and Functions of the actVI Region
with the known substrate motif of the C. acetobutylicum p-hy- droxybutyryl-CoA dehydrogenase, we propose that this reduc- tion would be catalyzed by the actVI-ORF1 gene product. The resultant secondary alcohol (5) would undergo cyclization to the hemiketal (61, followed by dehydration to afford the actVA intermediate (31, which is known to be a precursor of actino- rhodin (5). These two reactions might well be expected to pro- ceed spontaneously, but perhaps one or both of them is assisted by the product of actVI-ORFA or, even more likely, actVI-ORF3. This suggestion is prompted by the reduction in the level of actinorhodin biosynthesis caused by disruption of ORFA and the delayed production of blue pigment from a yellow precursor in the ORF3 disruptant.
The partial structure (b) included in the actVA intermediate (31, is a substrate motif of enoyl reductases. Assuming a 1,4- reduction, the product would be the a$-unsaturated ketone (71, which is an ideal substrate for subsequent oxidation (see be- low). However, this transformation might perhaps occur in two coupled steps, with reduction of the double bond of the (b) partial structure to yield a molecule with a p,y double bond, which would then be isomerized to the a$ double bond. It is therefore reasonable to speculate that the essential actVI- ORF2 would encode the reductase at C-15, with the actVI- ORF4 product perhaps assisting in the chemically favored isomerization.
Shen and Hutchinson (63) characterized the monooxygenase involved in formation of the naphthacenequinone tetracenomy- cin D3 (10) from the naphthacenone tetracenomycin F1 by Streptomyces glaucescens (9). This enzyme is encoded by tcmH (641, whose product resembles (39% identity) the actVA-ORF6 product (131, implicating this enzyme also in oxidation using molecular oxygen (65). The proposed intermediate (7 ) produced by the enoyl reduction could therefore undergo oxidation a t C-6 by a similar mechanism (Fig. 8) to afford dihydrokalafungin (8). Perhaps this oxidation step could be coupled with the reaction catalyzed by the actVI-ORF2 (and possibly ORF4) product, be- cause mutations in the actVA region (13) caused accumulation of 3, not 7 (5). However, another possibility is that 7 was too unstable to be isolated from the actVA mutant and only its proposed precursor (3) was characterized instead (5).
Dimerization of dihydrokalafungin (8), catalyzed by the a c t W product (12), followed by hydroxylation a t C-8, controlled also by the actVA region (13, 66), would afford actinorhodin. Thus the whole biosynthetic pathway to actinorhodin, includ- ing the two stereochemical features a t C-3/C-15, could be ex- plained by the proposed scheme.
The availability of the sequence of the entire act cluster will now guide chemicalhiochemical approaches to understand in detail the biosynthesis of actinorhodin, particularly the forma- tion of the pyran ring, followed by oxidation, as a model mem- ber of the benzoisochromanequinone family. A fascinating fea- ture of the family is the opposite stereochemistry at C-3 and C-15 in different members. Actinorhodin represents the 3S,15R-type dihydrogranaticin (67), produced by Streptomyces violaceoruber TU22, the 3R,15S-type, whereas S. roseofulvus makes members of both types, frenolicin B and nanaomycins, respectively (68). The polyketide synthase genes (gra and fren) from these two organisms have been cloned and sequenced (34, 37), and so gra and fren homologues of the actVA and actVI ORFs should be found in the vicinity of the polyketide synthase genes, opening possibilities for the generation of actlgra and actlfren hybrid gene clusters to help to pinpoint the determi- nants of C-3/C-15 chiralities. Such experiments, together with the synthesis of some proposed intermediates followed by feed- ing experiments, and the development of in vitro biosynthesis
using overexpressed gene products, are particularly attractive and are currently in progress.
Acknowledgments-We thank M. J. Bibb for making available the Streptomyces codon usage table; S. J.Lucania for the gift of thiostrepton; and K. F. Chater, C. Khosla, and W. P. Revill for helpful comments on the manuscript.
1.
2.
3. 4. 5.
6.
7.
8.
10. 9.
11.
12.
13.
14. 15. 16.
17.
18. 19.
20. 21.
22.
23.
24. 25. 26.
27. 28. 29.
30. 31.
33. 32.
34.
35.
36.
37.
38.
39.
40.
41. 42.
43
44
45 46
47
REFERENCES Brockman, H., Zeek, A., Van der Merve, K., and Muller, W. (1966) Justus
Gorst-Allman, C. P., Rudd, B. A. M., and Floss, H. G. (1981) J. Org. Chem. 46,
Wright, L. F., and Hopwood, D. A. (1976) J. Gen. Microbiol. 96,289-297 Rudd, B. A,, and Hopwood, D. A. (1979) J. Gen. Microbiol. 114, 3 5 4 3 Cole, S. P., Rudd, B. A,, Hopwood, D. A,, Chang, C. J., and Floss, H. G. (1987)
Zhang, H., He, X. G., Adefarati, A., Galluci, J., Cole, S. P., Beale, J. M., Keller, J. Antibiot. (Tokyo) 40,340-347
Bartel, P. L., Zhu, C. B., Lampel, J. S., Dosch, D. C., Connors, N. C., Strohl, W. P. J., Chang, C., and Floss, H. G. (1990) J. Org. Chem. 66, 1682-1684
Malpartida, F., and Hopwood, D. A. (1984) Nature 309,462-464 R., Beale, J. J., and Floss, H. G. (1990) J. Bacteriol. 172, 48164826
Malpartida, F., and Hopwood, D. A. (1986) Mol. & Gen. Genet. 206, 66-73 Fernsndez-Moreno, M. A,, Caballero, J. L., Hopwood, D. A,, and Malpartida, F.
Hallam, S. E., Malpartida, F., and Hopwood, D. A. (1988) Gene (Amst.) 74,
Fernhdez-Moreno, M. A,, Martinez, E., Boto, L., Hopwood, D. A., and Mal-
Caballero, J. L., Martinez, E., Malpartida, F., and Hopwood, D. A. (1991) Mol.
Yanisch-Perron, C., Vieira, J., and Messing, J. (1985) Gene (Amst.) 33,103-119 Bullock, W. O., Fernhndez, J. M., and Short, J. M. (1987) Biolkchniques 6,376 Chater, K. F., Bruton, C. J., King. A. A,, and Suarez. J. E. (1982) Gene (Amst.)
Liebigs Ann. Chem. 698,3575-3579
455456
(1991) Cell 66,769-780
305-320
partida, F. (1992) J. Biol. Chem. 267,19278-19290
& Gen. Genet. 230,401-412
-.
Hopwood, D. A,, Kieser, T., Wright, H. M., and Bibb, M. J. (1983) J. Gen.
Janssen, G. R., and Bibb, M. J. (1993) Gene (Amst.) 124, 133-134 Bartolome, B., Jubete, Y., Martinez, E., and de la Cruz, F. (1991) Gene (Amst.)
19, 21-32,
Microbiol. 129, 2257-2269
102.75-78 Rodicio, M. R., Bruton, C. J., and Chater, K. F. (1985) Gene (Amst.) 34,283-292 Hopwood, D. A,, Bibb, M. J., Chater, K. F., Kieser, T., Bruton, C. J., Kieser, H.
M., Lydiate, D. J., Smith, C. P., Ward, J. M., and Schrempf, H. (1985) Genetic Manipulation of Streptomyces, A Laboratory Manual, The John Innes Foun-
Maniatis, T., Fritsch, E. F., and Sambrook, J. (1982) Molecular Cloning: A dation, Norwich, United Kingdom
Laboratory Handbook Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, hT
Sanger, F., Nicklen, S., and Coulson, A. R. (1977) Proc. Natl. Acad. Sci. U. S. A.
Henikoff, S. (1984) Gene (Amst.) 28, 351-359 Maxam, A. M., and Gilbert, W. (1980) Methods Enzymol. 66,499-560 Devereux, J., Haeberli, P., and Smithies, 0. (1984) Nucleic Acids Res. 12,
74, 5463-5467
387395 Wright, F., and Bibb, M. J. (1992) Gene (Amst.) 113, 55455 Chater, K. F., and Bruton, C. J. (1983) Gene (Amst.) 26, 67-78 Saiki, R. K., Gelfand, D. H., Stoffel, S., Scharf, S. J., Higuchi, R., Horn, G. T.,
Mullis, K. B., and Erlich, H. A. (1988) Science 239, 487491 Murray, G. M. G. (1986) Annu. Rev. Biochem. 168, 165-170 Oki, T., Matsuzawa, Y., Kiyoshima, I t , Yoshimoto, A,, Naganawa, H., Takeu-
Bibb, M. J. , Findlay, P. R., and Johnson, M. W. (1984) Gene (Amst.) 30,157-166 Bibb, M. J., Bird, S., Motamedi, H., Collins, J . F., and Hutchinson, C. R. (1989)
chi, T., and Umezawa, H. (1981) J. Antibiot. (Tokyo) 34, 783-790
Sherman, D. H., Malpartida, F., Bibb, M. J., Kieser, H. M., Bibb, M. J., and EMBO J. 8,2727-2736
Honwood. D. A. (1989) EMBO J. 8.2717-2725 BibblkJ.<1986) inBiologica1, Biochemical and Biomedical Aspects of acti-
nomycetes, Part A (Szabb, G., Birb, S., and Goodfellow, M., eds) pp. 25-34,
Legouill, C., Desmarais, D., and Dery, C. V (1993) Mol. & Gen. Genet. 240, Akademiai Kiadb, Budapest
Bibb. M. J.. Sherman. D. H.. Omura. S., and Hopwood, D. A. (1994) Gene 146-150
~ ~~
(Amst.) 142,31-39 ' , . - .
Youngleson, J. S., Jones, D. T., and Woods, D. R. (1989) J. Bacteriol. 171,
Mulders, J. W. M., Hendriks, W., Blankesteijn, W. M., Bloemendal, H., and de
Bitar, K. G., Perez-Aranda, A,, and Bradshaw, R. A. (1980) FEBS Lett. 116,
DiRusso, C. (1990) J. Bacteriol. 172, 64594468 Yang, X.-Y. H., Schulz, H., Elzinga, M., andYang, S.-Y. (1991) Biochemistry 30,
Ishii, N., Hijikata, M., Osumi, T., and Hashimoto, T. (1987) J. Biol. Chem. 262,
Sato, S., Havashi, M., Imamura, S., Ozeki, Y., and Kawaguchi, A. (1992) J.
6860-6807
Jong, W. W. (1988) J. Biol. Chem. 263, 15462-15466
196-198
67884795
8144-8150
Jornvall, H., Persson, B., and Jeffery, J. (1987) Eur. J. Biochem. 167,195-201 Wierenga, R. K., Terpstra, P., and Hol, W. G. J. (1986) J. Mol. Biol. 187,
Biochem. i lbkyo) 111, &I5
Young, I. G., Rogers, B. L., Campbell, H. D., Jaworowski, A,, and Shaw, D. C. 101-107
(1981) Eur. J. Biochem. 116, 165-170

DNA Sequence and Functions of the actVI Region 48. Persson, B., Krook, M., and Jomvall(1991) Eur. J. Biochem. 200,537-543 49. Bevitt, D. J., CortBs, J., Haydock, S. F., and Leadlay, P. F. (1992) Eur. J.
50. Mathur, M., and Kolattukudy, P. E. (1992) J. Biol. Chem. 267,19388-19395 51. Schweizer, M., Takabayashi, K., Lam, T., Beck, K . 3 , and Schreglmann, R.
52. Park, D. H., and Plapp, B. V. (1991) J. Biol. Chem. 266, 1329613302 53. Nakayama, N., Arai, N., Bond, M. W., Kaziro, Y., and Arai, K. (1984) J. Biol.
54. GonzBlez, P., Rao, P. V., and Zigler, J. J. (1993) Biochem. Biophys. Res. Corn.
55. Ammendola, S., Raia, C. A,, Camso, C., Camardella, L., DAuria, S., De, R. M.,
56. Hanukoglu, I., and Gutfinger, T. (1989) Eur. J. Biochem. 180,479434 57. Scmtton, N. S., Berry, A., and Perham, R. N. (1990) Nature 343, 38-43 58. Brown, N. L., Ford, S. J., Pridmore, R. D., and Fritzinger, D. C. (1983) Bio.
59. Wierenga, R. K., and Hol, W. G. (1983) Nature 302,842-844 60. Bairoch, A. (1992) Nucleic Acids Res. 20, (suppl.) 2013-2018 61. Strohl, W. R. (1992) Nucleic Acids Res. 20, 961-974 62. Gramajo, H. C., Takano, E., and Bibb, M. J. (1993) Mol. Microbiol. 7,837-845 63. Shen, B., and Hutchinson, C. R. (1993) Biochemistry 32,6656-6663 64. Summers, R. G., Wendtienkowski, E., Motamedi, H., and Hutchinson, C. R.
65. Fujii, I., Chen, Z.-G., Ebizuka, Y., and Sankawa, U. (1991) Biochern. Int. 26,
Biochem. 204,39-49
(1989) Nucleic Acids Res. 17, 567-586
Chem. 269,97-101
mun. 191,902-907
and Rossi, M. (1992) Biochemistry 31, 12514-12523
chemistry 22,4089-4095
(1993) J. Baeteriol. 176, 7571-7580
1043-1049
24863 66. Hopwood, D. A., Malpartida, F., Kieser, H. M., Ikeda, H., Duncan, J., Fujii, I.,
67. Snipes, C. E., Chang, C . J . , and Floss, H. G. (1979) J. Am. Chem. Soc. 101,
68. Tsuzuki, K., Iwai, Y., Omura, S., Shimizu, H., and Kitajima, N. (1986) J.
69. Kaiser, R., Nussrallah, B., Dam, R., Wagner, F. W., and Jornvall, H. (1990)
70. Estonius, M., Karlsson, C., Fox, E. A., Hoog, J. O., Holmquist, B., Vallee, B. L.,
71. Hoog, J. O., Vagelopoulos, N.,Ylp, P. K., Keung, W. M., and Jornvall, H. (1993)
72. Persson, B., Bergman, T., Keung, W. M., Waldenstrom, U., Holmquist, B.,
73. van Der Straeten, D., Rodrigues-Pousada, R., Gielen, J., andVan, M. M. (1991)
74. Xie, Y., and Wu, R. (1989) Plant Mol. Biol. 13, 53-68 75. Dennis, E. S., Sachs, M. M., Gerlach, W. L., Finnegan, E. J., and Peacock, W.
76. Danielsson, O., Eklund, H., and Jornvall, H. (1992) Biochemistry 31, 3751-
77. Wehner, E. P., Rao, E., and Brendel, M. (1993)Mol. & Gen. Genet. 237,351-358 78. Russell, P. R., and Hall, B. D. (1983) J. Biol. Chem. 288, 143-149 79. Jendrossek, D., Steinbiichel, A,, and Schlegel, H. G. (1988) J. Bacteriol. 170,
Rudd, B. A,, Floss, H. G., and Omura, S. (1985) Nature 314, 642-644
701-706
Antibiot. (Tokyo) 39, 1343-1345
Biochemistry 29,8365-8371
Davidson, W. S., and Jomvall, H. (1990) Eur: J . Biochem. 194,593-602
Eur. J. Biochem. 213,3148
Vallee, B. L., and Jornvall, H. (1993) Bur. J. Biochem. 216.49-56
FEBS Lett. 295,3942
J. (1985) Nucleic Acids Res. 13, 727-743
3759
5248-5256


















