
Predictors of N-Acetyltransferase Activity: Should...
8
Vol. 5, 449-455, June 1996 Cancer Epidemiology, Biomarkers & Prevention 449 Predictors of N-Acetyltransferase Activity: Should Caffeine Phenotyping and NAT2 Genotyping Be Used Interchangeably in Epidemiological Studies?’ LoIc Le Marchand,2 Lakshmi Sivaraman,3 Adrian A. Franke, Laurie J. Custer, Lynne R. Wilkens, Alan F. Lau, and Robert V. Cooney Etiology Program. Cancer Research Center of Hawaii, University of Hawaii. Honolulu, Hawaii 96813 Abstract To determine whether NAT2 genotyping could be used interchangeably with caffeine phenotyping in assessing N-acetyltransferase activity in epidemiological studies, sources of interindividual variability in N- acetyltransferase activity were assessed among 90 subjects of various ethnic backgrounds in Hawaii. Forty-three subjects were patients with in situ coborectal cancer treated by polypectomy, and 47 were healthy population controls. Subjects were administered a lifestyle questionnaire and were evaluated for N-acetyltransferase activity by caffeine phenotyping. NAT2 genotype was also assessed by PCR amplification of peripheral leukocyte DNA for the Ml, M2, and M3 variant alleles. Fifty-four % of the overall variation in acetylation activity was explained by the three genotype categories (homozygous variant, heterozygous, and homozygous wild-type). This proportion was reduced to 42% when genotype was modeled using only two categories (“slow” being homozygous variant; “rapid” being all others). Use of gout medications (probenecid or allopurinol), consumption of heavily browned fish, and P450IA2 activity (also measured by caffeine phenotyping), together explained another 11% of the variance. No association was found between acetylation activity and sex; race; age; education; smoking; physical activity; weight; consumption of coffee, alcohol, red meat, processed meat, and cruciferous vegetables; or use of menopausal estrogens, after taking genotype into account. Results were similar for coborectal cancer patients and controls. Considerable variation in acetylation activity was Received 9/6/95; revised 1/16/96; accepted 2/5/96. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. I This research was supported in part by NIH Grants POl-CA-33619 and ROl- CA-60987 and contract NOl-CN-05223 from the National Cancer Institute. 2 To whom requests for reprints should be addressed, at Cancer Research Center of Hawaii, University of Hawaii. 1236 Lauhala Street. Suite 407, Honolulu, HI 96813. 4 Current address: Department of Cell Biology. Baylor College of Medicine, Houston, TX 77030. observed within the homozygous wild-type group. This study suggests that the use of genotyping, instead of phenotyping, to assess the association of acetylation with cancer risk is unlikely to introduce major misclassification or bias, especially when the three genotype categories are modeled and the sample size is large. However, when the rapid acetylation phenotype is the at-risk group (e.g., when studying colon cancer), phenotyping appears judicious given the variability in acetylation activity within this group. Introduction The recent associations of the slow N-acetylation phenotype with bladder cancer in arylamine-exposed workers ( 1) and of the rapid acetylator phenotype with colorectal cancer (2) ur- gently need confirmation in large population-based studies be- cause this genetic polymorphism may be useful in characteriz- ing individual cancer risk. Because the world-wide distribution of the slow acetylation phenotype varies greatly, from 5% in Canadian Eskimos, to 10-20% in Japanese, 50-60% in Cau- casians, and 90% in Northern Africans (3), these associations may also clarify unexplained risk patterns, such as the high colorectab cancer risk of Japanese Americans (4). N-acetyltransferase is under autosomal dominant genetic control, and its activity is coded for by two distinct genes, NAT! and NAT2, located on chromosome 8 (5). The latter gene exhibits polymorphisms arising from point mutations in both coding and noncoding regions that can result in decreased expression, low activity, or enzyme instability. The presence of two germline copies of any of several mutations in the NAT2 gene produces a slow-acetybator phenotype. Although NAT1 has also been shown to be polymorphic in humans (6), the investigation of its association with disease has only recently begun (7). Most human studies of N-acetylation have used metabolic phenotyping methods using drugs, such as isoniazid, sulfameth- azine and dapsone, or, more recently, caffeine (8). A urine sample is collected a few hours after dosage and analyzed by HPLC.4 The ratio of urine metabolite(s) to the unmetabolized parent compound permits an assignment to the phenotype. The caffeine method is attractive because the sampling time and dose do not need to be very precise, and the metabolic ratio appears to be independent of urine flow and kidney function. However, although the enzyme is generally regarded as 4 The abbreviations used are: HPLC, high-performance liquid chromatography; AFMU, 5-aeetylamino-6-formylamino-3-methyluracil; MET. metabolic equiva- lent; HAA. heterocyclic aromatic amine; PAH. polycyclic aromatic hydrocarbon. on June 16, 2018. © 1996 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from
-
Upload
truongthuan -
Category
Documents
-
view
220 -
download
1
Transcript of Predictors of N-Acetyltransferase Activity: Should...

Vol. 5, 449-455, June 1996 Cancer Epidemiology, Biomarkers & Prevention 449
Predictors of N-Acetyltransferase Activity: Should Caffeine
Phenotyping and NAT2 Genotyping Be UsedInterchangeably in Epidemiological
Studies?’
LoIc Le Marchand,2 Lakshmi Sivaraman,3Adrian A. Franke, Laurie J. Custer, Lynne R. Wilkens,Alan F. Lau, and Robert V. Cooney
Etiology Program. Cancer Research Center of Hawaii, University of Hawaii.
Honolulu, Hawaii 96813
Abstract
To determine whether NAT2 genotyping could be used
interchangeably with caffeine phenotyping in assessingN-acetyltransferase activity in epidemiologicalstudies, sources of interindividual variability in N-
acetyltransferase activity were assessed among 90 subjectsof various ethnic backgrounds in Hawaii. Forty-threesubjects were patients with in situ coborectal cancertreated by polypectomy, and 47 were healthy populationcontrols. Subjects were administered a lifestylequestionnaire and were evaluated for N-acetyltransferaseactivity by caffeine phenotyping. NAT2 genotype was alsoassessed by PCR amplification of peripheral leukocyteDNA for the Ml, M2, and M3 variant alleles. Fifty-four% of the overall variation in acetylation activity wasexplained by the three genotype categories (homozygousvariant, heterozygous, and homozygous wild-type). Thisproportion was reduced to 42% when genotype wasmodeled using only two categories (“slow” beinghomozygous variant; “rapid” being all others). Use ofgout medications (probenecid or allopurinol),consumption of heavily browned fish, and P450IA2activity (also measured by caffeine phenotyping), togetherexplained another 1 1 % of the variance. No associationwas found between acetylation activity and sex; race; age;education; smoking; physical activity; weight;consumption of coffee, alcohol, red meat, processed meat,and cruciferous vegetables; or use of menopausalestrogens, after taking genotype into account. Resultswere similar for coborectal cancer patients and controls.
Considerable variation in acetylation activity was
Received 9/6/95; revised 1/16/96; accepted 2/5/96.The costs of publication of this article were defrayed in part by the payment of
page charges. This article must therefore be hereby marked advertisement in
accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
I This research was supported in part by NIH Grants POl-CA-33619 and ROl-
CA-60987 and contract NOl-CN-05223 from the National Cancer Institute.
2 To whom requests for reprints should be addressed, at Cancer Research Center
of Hawaii, University of Hawaii. 1236 Lauhala Street. Suite 407, Honolulu, HI
96813.
4 Current address: Department of Cell Biology. Baylor College of Medicine,
Houston, TX 77030.
observed within the homozygous wild-type group. Thisstudy suggests that the use of genotyping, instead ofphenotyping, to assess the association of acetylationwith cancer risk is unlikely to introduce majormisclassification or bias, especially when the threegenotype categories are modeled and the sample size islarge. However, when the rapid acetylation phenotype isthe at-risk group (e.g., when studying colon cancer),phenotyping appears judicious given the variability in
acetylation activity within this group.
Introduction
The recent associations of the slow N-acetylation phenotypewith bladder cancer in arylamine-exposed workers ( 1 ) and of
the rapid acetylator phenotype with colorectal cancer (2) ur-gently need confirmation in large population-based studies be-cause this genetic polymorphism may be useful in characteriz-
ing individual cancer risk. Because the world-wide distribution
of the slow acetylation phenotype varies greatly, from 5% inCanadian Eskimos, to 10-20% in Japanese, 50-60% in Cau-
casians, and 90% in Northern Africans (3), these associationsmay also clarify unexplained risk patterns, such as the highcolorectab cancer risk of Japanese Americans (4).
N-acetyltransferase is under autosomal dominant genetic
control, and its activity is coded for by two distinct genes, NAT!
and NAT2, located on chromosome 8 (5). The latter geneexhibits polymorphisms arising from point mutations in bothcoding and noncoding regions that can result in decreasedexpression, low activity, or enzyme instability. The presence oftwo germline copies of any of several mutations in the NAT2
gene produces a slow-acetybator phenotype. Although NAT1has also been shown to be polymorphic in humans (6), the
investigation of its association with disease has only recently
begun (7).Most human studies of N-acetylation have used metabolic
phenotyping methods using drugs, such as isoniazid, sulfameth-azine and dapsone, or, more recently, caffeine (8). A urine
sample is collected a few hours after dosage and analyzed byHPLC.4 The ratio of urine metabolite(s) to the unmetabolized
parent compound permits an assignment to the phenotype. Thecaffeine method is attractive because the sampling time anddose do not need to be very precise, and the metabolic ratioappears to be independent of urine flow and kidney function.However, although the enzyme is generally regarded as
4 The abbreviations used are: HPLC, high-performance liquid chromatography;
AFMU, 5-aeetylamino-6-formylamino-3-methyluracil; MET. metabolic equiva-
lent; HAA. heterocyclic aromatic amine; PAH. polycyclic aromatic hydrocarbon.
on June 16, 2018. © 1996 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from

450 Predictors of N-Acetyltransferase Activity
noninducible ( 1 ), it is possible that the disease process, itstreatment, or some xenobiotics, such as medications or alcohol(9), may distort the phenotyping. In studies of its associationwith disease, this may result in increased variability (i.e. . loss of
statistical power), or in bias if the distorting factors are alsoassociated with the disease. Thus, the phenotyping approach
may present logistical difficulties in population studies.PCR-based methods have recently been developed for
detecting the main mutations associated with the slow acetyla-
tion phenotype in DNA from blood or fixed-tissue (10, 11).Initially, three different point mutations (MI, M2, and M3) in
the NAT2 gene were described. The genotyping method isrelatively straightforward, and the results are independent fromthe disease status of the subject and not influenced by externalvariables. Moreover, in epidemiological investigations, it is
usually easier to obtain biological material for genotyping anal-ysis than it is to perform lengthy phenotyping protocols. Indeed,recent studies have used the genotyping to investigate associ-
ations of NAT2 polymorphism with bladder and breast cancer(12, 13).
Several reports have shown an excellent concordance be-tween the phenotyping and genotyping methods used in the
classification of slow and rapid acetylators. This concordance
has been 90-98% in Japanese (14), Caucasians (10, 1 1, 15, 16),and Chinese (17). However, it has been shown that, depending
on the prevalence of the at-risk genotype, even a small rate ofgenotyping misclassification may result in a substantial atten-uation of the risk estimates ( I 8). Also, it remains unclearwhether the genotype, a categorical variable, carries as much
information as the phenotype, a continuous variable, which hasthe advantage of integrating both genetic and epigenetic influ-ences on the activity of the enzyme.
We report here on studies aimed at characterizing the main
constitutional, lifestyle, and laboratory predictors of N-acetyl-transferase activity to determine whether NAT2 genotyping canbe used instead of caffeine phenotyping in assessing cancer riskin epidemiological studies.
Materials and Methods
Subjects. Participants were described previously (19) and in-cluded patients with in situ adenocarcinoma of the large boweland healthy population controls. Eligible patients were all those
diagnosed with this condition in the main medical centers ofOahu between July 1989 and October 1991 and whose treat-ment did not include colectomy. The controls were selected
from among the participants in a population-based survey con-ducted by the Hawaii State Department of Health and werematched to each patient on sex, age (within 5 years), and
ethnicity. Overall. 72.4% of the eligible patients and 7 1 .6% ofthe eligible controls were interviewed. A total of 43 cases and47 controls (73.4% of the eligible interviewed subjects) agreed
to the biological component of the study, of whom 45 wereJapanese, 15 Caucasian, 12 Filipino, 12 Hawaiianlpart-Hawai-ian, and 6 Chinese. The mean age of the participants was 65.6
years.
Questionnaire Data. A detailed questionnaire on demograph-ics. diet, and lifestyle was administered at the participants’homes by trained interviewers. The dietary information cob-lected consisted of a quantitative diet history (20) focusing on
usual intake during the year before diagnosis for cases andduring the year preceding the interview for controls. It alsoincluded questions on browning of meat and fish surfaces 5
years before interview, as well as frequency of consumption of
cruciferous vegetables (cabbage, broccoli, Brussels sprouts, and
watercress) during the week before the caffeine test. Daily
tobacco, alcohol, coffee, and soda consumption, as well asmedication and supplement use, was ascertained for the 2-weekperiod preceding the caffeine test. Subjects were also asked tospecify the time spent at various levels of physical activity
during a typical workday and weekend day (21). These data
were summarized over an average 24-h period and converted
into METs (2 1 ). Weight was measured in light clothing.
Dosing Regimen and Specimen Collection. Subjects were
asked to refrain from consuming any foods or beverages (ex-cept water) from 10:00 pm to the time of the blood draw thefollowing morning. After the blood draw, the subjects drank a
cup of coffee made of two packets of Maxwell House instantcoffee (57 mg caffeine/packet). Then they voided 4 h aftercoffee consumption and provided a 1-h urine specimen at theend of the 5th hour after dosing. No other source of caffeinewas consumed during the 5-h period. The urine samples were
acidified with ascorbic acid, aliquoted, and stored at -70#{176}C
until analysis.
Intraindividual Variability Study. An additional and inde-pendent sample of 16 healthy, nonsmoking individuals of var-ious ethnic backgrounds participated in the reproducibility por-
tion of the study. Eight male and eight female volunteersrecruited from among the staff of our institution were admin-
istered caffeine as described above once weekly for 4 weeks.No recommendation was given with regard to diet or lifestyle;however, we varied the day of the week for the dosing tooptimize the possible effect of daily variations in lifestyle.
Phenotyping. We quantified urinary levels of AFMU/l-meth-ybxanthine (IX) to determine the N-acetyltransferase pheno-
type, as well as the urinary ratios of [1,7-dimethyburic acid(17U) + l,7-dimethybxanthine (17X)J/caffeine (l37X) to as-
sess P450IA2 activity. Urine samples were quickly thawed andextracted (200 pi) according to a slight modification of the
method of Butler et a!. (22) by adding 120 mg ammoniumsulfate and 120 p1 4-acetamido-phenol (160 �g/ml) as an
internal standard to the urine in a 15-mb centrifuge tube fob-
lowed by mixing for 2 mm. Caffeine and its metabobites werepartitioned by the addition of 6 ml chloroform:isopropanol( 19: 1 ). The organic phase was removed after centrifugation,
evaporated to dryness, then resuspended in 100 �.d of the HPLCmobile phase A consisting of 0.045% aqueous acetic acid
containing 9% methanol. Chromatography was carried out witha Supelcosil LC-18 5j.tm (4.6 X 250 mm) reverse-phase HPLCcolumn using a Beckman System Gold chromatograph. Sam-pies were eluted at I . 1 mI/mm using linear gradients of mobile
phases A and B (methanol) as follows: 0-3 mm (0% B); 3-5mm (2% B); 5-6 mm, (2-20% B); 6-16 mm, (20% B); 16-21
mm (60% B); 21-35 mm (0% B). The identification of themetabolites was done by comparing their retention times and
absorption spectra (Beckman Model 168 diode array detector)with those of authentic standards.
NAT2 Genotyping. Polymorphisms in the NAT2 gene weredetermined using the DNA purified from lymphocytes obtained
from the subjects. Lymphocytes were digested with SDS (0.5%final) and proteinase K (200 p.g/ml) overnight at 37#{176}C,fol-bowed by extraction with phenol and chloroforrnliso-amyb al-cohol (24: 1) the next day (23). The DNA was precipitated using
isopropanob, and the final DNA pellet was dissolved in TE(10 msi Tris.HC1 pH 8.0, 1 mrvi EDTA pH 8.0). The amount of
DNA was quantified spectrophotometricably by measurementof absorbance at 260 nm. The ratio of absorbance at 260 and280 nm indicated its purity.
The NAT2 gene was PCR amplified using primers N4:
on June 16, 2018. © 1996 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from

Fig. 1. Probit plot of the metabolic ratio
AFMU/I X.
0
C/)
ci
0a:0.
0
0.99
0.95
0.90
0.80
0.700.600.500.40
0.30
0.20
0.10
0.05
0.01
F,.
I I
0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6 1.8 2.0 2.2 2.4
Cancer Epidemiology, Biomarkers & Prevention 45/
LOG [(AFMU /1-X) + 1]
5 ‘TCTAGCATGAATCACTCTGC3’ and N5: 5’GGAACAA-ATTGGACTTGG3’. The Ampliwax TM PCR GEM100 beads
and protocol for Hot Stan PCR from Perkin-Elmer Cetus wereused in this amplification. A 109l-bp fragment was obtained
using 500 ng of genomic DNA, 50 pmol of each primer, 200�mob of each dNTP, 1 unit of AmpliTaq DNA polymerase, anda buffer ( 1 .25X PCR buffer II, Perkin-Elmer Cetus) composedof 12.5 mM Tris.HCI (pH 8.3), 3 mrvi MgCl2, and 62.5 mr�i KCI
in a total volume of 80 pA. In a Perkin-Elmer Cetus PCR 480
system thermal cycler, the reaction was denatured at 94#{176}Cfor4 mm and then subjected to 30 cycles at 94#{176}Cfor 30 s, 58#{176}Cfor
30 s, and 72#{176}Cfor I .5 mm. A final extension at 72#{176}Cfor 5 mmwas performed. After PCR, 20-�l abiquots were subjected to
digestion overnight with restriction enzymes KpnI, TaqI, andBamHI (Promega) according to the manufacturer’ s recommen-
dations. These digestions are diagnostic for the alleles MI, M2,
and M3, respectively (I 1). Taq digests were electrophoresed on4% 3: 1 NuSieve agarose, and the KpnI and BamHI digests wereresolved on a 3% 3: 1 NuSieve agarose (FMC BioProducts,Inc.).
Data Analysis. The objective of the analysis was to study thevariability of N-acetyltransferase activity, as assessed by
the caffeine test. The metabolic ratio was log transformed, asbog(x + 1), for its distribution to approximate normality. In theintraindividual variability study, the coefficient of variation,
averaged across individuals, was taken as a measure of varia-bility, as was the intraclass correlation coefficient, computed
from a one-way random-effects ANOVA model (24). The co-
efficient of variation was also used to measure variability inlaboratory measurement.
A probit model and a probit plot (25) of the AFMU/lXratio were used to determine a cutoff point for the distributionsof slow and rapid metabobizers. Lastly, analysis of covariancewas used to examine how much variability was accounted for
by the NAT2 genotype and other covariates. A stepwise proce-dure was executed to determine the importance of other van-
abbes after genotype was controlled for. The potential covari-
ates selected for the regression were those shown to affectlevels of N-acetyltransferase or other xenobiotic metabolizing
enzymes and/or to be associated with cancer risk. These vari-
abbes have the potential not only to increase the backgroundvariability in N-acetybtransferase activity but also to act asconfounders of an association between acetylation and cancer.
Results
Intraindividual and Laboratory Variability. Among the 16subjects phenotyped with caffeine once a week for 4 weeks, nomisclassification occurred upon repeated measurement when
the subjects were classified as “slow” or “rapid” acetylators(see below for definition). However, when treated as a contin-
uous variable, the mean coefficient of variation for the urinarymetabolic ratio (AFMU/1X) was 24% (range, 9-55%), and theintraclass correlation coefficient for the four repeated measure-
ments was 0.73. Analyses of duplicate samples in our labora-tory resulted in good reproducibility of the measurement ofAFMU/bX, with intra- and interassay coefficients of variationof 7.9 and 12.8%, respectively. These intraindividual and lab-
oratory variations are comparable to published data ( I 7, 22,26, 27).
Acetylator Phenotype. No caffeine metabolites could be
detected in the urine of 2 of the 90 subjects. They were ex-cluded from subsequent analyses involving phenotype. Thedistribution of N-acetyltransferase activity, expressed as
log(AFMU/1X + 1), among the 88 remaining subjects pheno-typed with caffeine is presented in a probit plot in Fig. I . Visualinspection and probit analysis suggested that a cutoff point of0.4 [0.34 on a bog(x + 1 ) scale] could be used to distinguish
slow acetylators (14.9%) from rapid acetylators (85.1%). Thiscutoff point for our multiethnic population was intermediatebetween that of 0.3 found for Japanese (28) and Chinese (29)and that of 0.6 observed in a mainly Caucasian populationfrom Arkansas (22). In agreement with past studies (28), the
proportion of slow acetylators (AFMU/1X < 0.4) was 9.1%
on June 16, 2018. © 1996 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from

Table I Acetylation activity by sex, age, and lifestyle among in situcolorectal cancer cases and population controls
Geometric Mean AFMU/IX”
Cases Controls All(,,=41) (,i=47) (n88)
Sex
Table 1 Continued
Geometric Mean AFMU/1X”
Cases Controls All
(n 41) (‘1 = 47) (ii = 88)
Physical activity (METS)
rt50.O 1.44 1.16 1.24
50.1-61.7 1.64 1.38 1.44
�6l.8 1.27
P = 0.69
1.39
P = 0.85
1.36
P = 0.79
Gout medications
Nonusers 1 .43 1 .35 1.39
Users 0.43
P = 0.07
0.53
P = 0.09
0.56
P = 0.03
P450IA2 activity I) I 7U+ I7X)/l37Xl
rt3.44 1.45 1.08 1.22
3.45-6.89 1.27 1.24 1.34
�6.90 1 .48
P = 0.89
1 .66
P = 0.36
1.52
P = 0.55
Male 1.43 1.18 1.44
Female 1.39 1.47 1.30
P = 0.92 P = 0.40 P - 0.37
Age (years)
36-62 1.38 1.24 1.32
63-71 1.50 1.38 1.44
72-85 1.39 1.35 1.34
P = 0.96 P = 0.93 P = 0.92
Years of schooling
1-12 1.36 1.26 1.29
13-19 1.47 1.37 1.43
P = 0.75 P = 0.77 P = 0.55
Smoking (cigarettes/day)
0 1.36 1.29 1.34
1-40 2.44
P�0.l2
1.58
P=O.64
1.65
P=O.45
Alcohol (drinks/day)
0 1.63 1.49 1.54
1-10 1.20
P = 0.21
0.87
P = 0.08
1.02
P = 0.03
Coffee (cups/day)
0-0.5 1.26 1.47 1.40
0.6-10 1.62
P = 0.31
1.24
P = 0.51
1.34
P = 0.77
Red meat (g/day)
0-39 1.33 1.04 0.12
40-95 1.31 1.74 1.49
�96 1.57
P = 0.80
1.17
P = 0.28
1.39
P = 0.43
Chicken and fish (g/day)
(3-30 0.91 1.54 1.14
31-90 1.95 1.26 1.67
rt9l 1.54 1.11 1.28
P=O.02 P=O.61 P=O.l2
Processed meat (g/day)
0-5.5 1.23 1.38 1.29
5.6-24.0 1.47 1.30 1.33
�24.l 1.60
P = 0.70
1.29
P = 0.98
1.45
P = 0.85
Cruciferous vegetables
(times in past week)
0 1.19 1.36 1.34
>1 1.77 1.20 1.44
P=0.Il P=0.69 P=0.69
Color of meat surface
Did not eat 2.18 1.88
Lightly brown 1.51 1.12 1.30
Moderately brown 1.58 1.34 1.42
Heavily brown 1.32 1.30 1.32
P=0.8l P=O.89 P=O.94
Color of fish surface
Did not eat I .73 1.53
Lightly brown 0.71 0.88 0.78
Moderately brown 1 .29 1 .25 1.30
Heavily brown 1 .65 1 .55 1.69
P = 0.05 P = 0.43 P = 0.03
Weight (kg)
42.2-58.4 1.24 1.40 1.32
58.5-73.7 1.56 1.37 1.46
�73.8 1.33
P - 0.68
1.23
P = 0.90
1.32
P = 0.86
a Adjusted by covariance analysis for race, and case-control status when appro-
priate.
in Japanese. The corresponding proportion for Caucasians(35.7%) was somewhat bow compared to previous studies
(22), in part because of the lower cutoff point used for oursample. There was no statistically significant difference in
the proportion of rapid acetylators (P = 0.23) or in race-adjusted mean metabolic ratio (P = 0.64) between cases andcontrols. The crude odds ratio for in situ coborectal cancerfor rapid compared to slow acetybators was 2.1 (95% confi-
dence interval, 0.6-7.4).The association of sex; age; education; smoking; alcohol,
coffee, red meat, chicken and fish, processed meat, and cruci-ferous vegetable intakes; color of meat and fish surfaces;weight; physical activity; gout medications; and P4501A2 ac-tivity with N-acetybtransferase activity is shown in Table 1. Theresults were similar between the case and control groups inshowing associations with alcohol, consumption of fish with
browned surfaces, and use of gout medications (probenecid or
allopurinol). In this univariate analysis, alcohol and gout med-
ications were associated with a bower metabolic ratio, andconsumption of browned fish was associated with a higherratio. No association was also found between the metabolic
ratio and use of vitamin A, vitamin E, or /3-carotene supple-ments; use of aspirin, menopausal estrogens, acetaminophen, ornonsteroidal antiinflammatory agents; family history of cob-rectal cancer; or personal history of diabetes (data not shown).
Comparison of Genotype and Phenotype. Using the crite-rion that both alleles in an individual must be mutated toproduce the slow phenotype, there was a 94% overall concord-ance between genotype and phenotype, corresponding to asensitivity of 100% and specificity of 93.2% for the genotype
in correctly classifying slow acetybators. Five individuals had a
rapid acetylator phenotype despite being homozygous variants.These “misclassified” subjects were all homozygous with twodifferent mutations (M1/M2 or M2/M3) and were either Cau-casian (n = 2), Filipino (n = 2), or Japanese (n = 1).
Samples for these five subjects were reanalyzed using thesame protocol by an independent laboratory, which confirmedthe genotypes of four of the five subjects. The genotype of oneFilipino subject was reclassified from M2/M3 to WT/M3, for anew overall concordance rate of 95% between phenotype and
genotype. In the subsequent analyses, this individual was clas-sified as WT/M3.
452 Predictors of N-Acetyltransferase Activity
on June 16, 2018. © 1996 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from

Cancer Epidemiology, Biomarkers & Prevention 453
Table 2 Distribution of th e NA72 genotypes
Genotype.
No. of Subjects Frequency (%)Geometric mean
AFMU/lxb
Homozygous wild-type 23 (25.1) 25.6 2.75
�7/WT
Heterozygous
All alleles combined 49 (44.9) 54.4 1.56
WJ’IM/ 6(10.5) 6.7 1.22
WT/M2 28(22.1) 31.1 1.58
WT/M3 15 (12.2) 16.7 1.75
Homozygous mutant
All alleles combined 1 8 (20. 1 ) 20.0 0.37
MI/MI 4 (1 . 1) 4.4 0.21
M2/M2 3 (4.9) 3.3 0.30
M3/M3 1(1.5) 1.1 0.50
MI/M2 4(4.7) 4.4 0.31
M1/M3 2 (2.6) 2.2 0.04
M2/M3 4(5.4) 4.4 0.63
a Number of patients predicted by application of the Hardy-Weinberg law shown
in parentheses: � = 4.85; p 0.85.
b Adjusted for race and case-control status by covariance analysis.
NAT2 Genotype. The distribution for all detected NAT2 gen-
otypes in the 90 genotyped subjects is presented in Table 2 andwas consistent with the Hardy-Weinberg equilibrium
(P = 0.85). Table 3 shows the prevalence of the Ml, M2, andM3 alleles in the ethnic groups studied. These results are
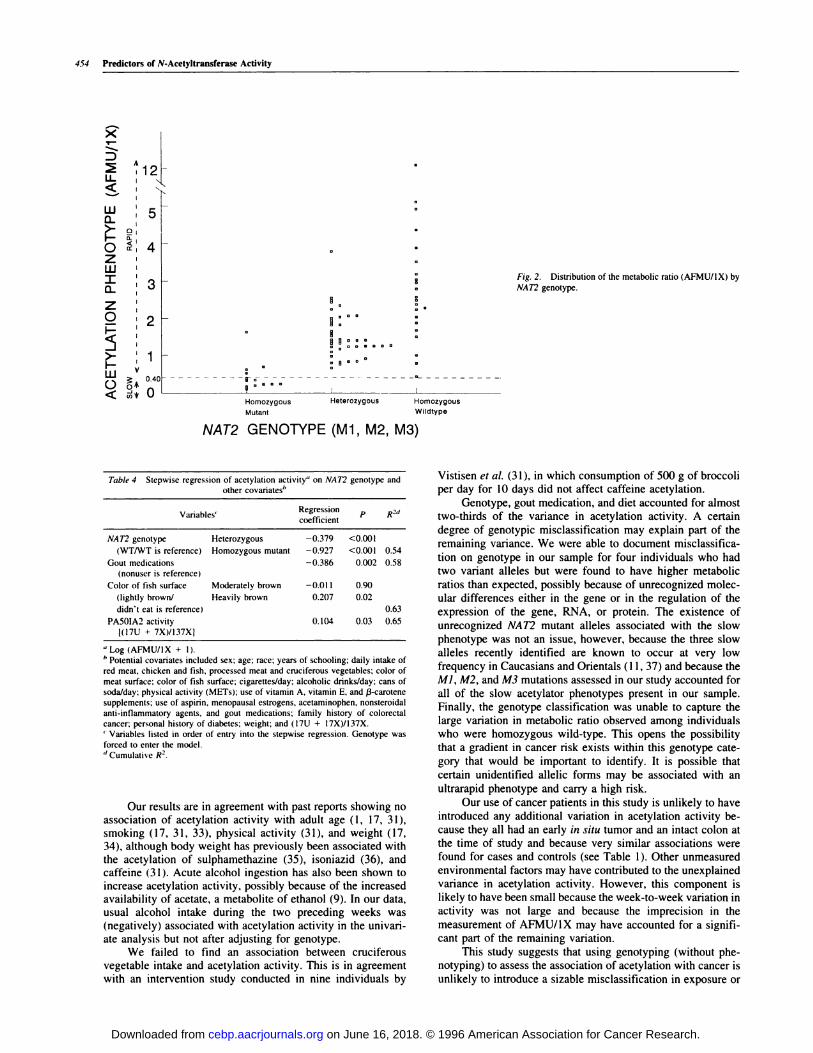
comparable to those reported by Lin et a!. (30) for Japanese,Caucasians, Filipinos, Samoans (who, like Hawaiians, arePobynesians) and Chinese. Individuals with the three genotypes(homozygous wild-type, heterozygous, and homozygous mu-tant) had clearly different acetybation activity (Table 2; Fig. 2;
P < 0.001). However, there was no statistically significant
difference in metabolic ratio by specific allele combinationswithin each of the three genotype categories (Table 2). Asshown in Fig. 2, there was a substantial overlap in metabolic
ratio between the heterozygous group and the homozygouswild-type group. As in other populations (1 1, 15-17), the batter
group showed considerable variation (30-fold) in acetylation
activity.
Predictors of N-Acetyltransferase Activity. The results ofthe stepwise regression showed that the three genotype catego-
ries considered above explained 54% of the variation in theacetybation ratio (Table 4). This proportion was reduced to 42%when modeled using two categories (homozygous mutant andother). The reduction in explained variance reinforces the po-
tential usefulness of contrasting all three genotype categorieswhen exploring disease associations.
Among the covariates competing in the stepwise regres-
sion (Table 4), only three were selected for inclusion into amodel that already contained the three NAT2 genotype catego-ries. Use of gout medications was negatively associated with
acetylation activity, whereas color of the fish surface andP4501A2 activity were positively associated. Together, thesethree variables explained another 1 1% of the variance inN-acetyltransferase activity. Thus, only 35% of the variationremained unexplained by our model.
Discussion
In this study of 90 individuals with various ethnic backgrounds,54% of the variability in N-acetyltransferase activity measured
by the caffeine test was accounted for by NAT2 genotype. Thiscompares well to the 59% variability in dapsone acetybation
Table 3 Frequency (%) of NAT2 alleles by ethnic group
Japanese Caucasian Filipino Hawaiian Chinese(n 90)” (,i = 30) (vi 24) (ii 24) (a = 12)
Wild-type 63.3 26.7 50.0 50.0 50.0
MI 3.3 50.0 0.0 8.3 0.0
M2 17.8 16.7 33.3 29.2 50.0
M3 15.6 6.7 16.7 12.5 0.0
a Total number of alleles.
explained by the NAT2 genotype in the study by Rothman et a!.
(17) among 84 Chinese men. Thus, consistent with the notionthat N-acetybtransferase activity is chiefly under geneticcontrol, genotype explains the majority of its interindividualvariance.
Following the model of the phenotyping studies, the few
genotyping studies reported to date have compared disease riskamong two genotyping categories corresponding to the slow(homozygous variant) and rapid (heterozygous and homozy-
gous wild-type) acetybator phenotypes (12, 13). Our data sug-gest that a better assessment of disease risk may be achievedwith three genotype categories because such a model explained
12% more of the variance in acetylation activity. Moreover, itwould allow for the examination of a genetic dose-response.
Consistent with the commonly accepted notion thatN-acetybtransferase is largely noninducible, few environmental
factors among the many assessed in this study were found tocorrelate with its activity. Only use of medications for gout and
consumption of browned fish were found to be correlated withcaffeine metabolism. After taking genotype into account, thesefactors were responsible for a modest 9% of additional variationin activity. The association of N-acetyltransferase activity withP4501A2 activity probably reflects the fact that, along themetabolic pathway of caffeine, the steps involving P450IA2
take place earlier than those involving N-acetyltransferase (31).To our knowledge, this is the first time that probenecid and
abbopurinol, two drugs used in the treatment of gout, were found
to be related to caffeine metabolism. Because these results werebased on a small number of users (n = 7) and were obtained in
a cross-sectional study, they need confirmation with a largersample size and a prospective design. Nevertheless, they sug-gest that certain drugs may distort the caffeine phenotyping ofN-acetyltransferase. An alternative interpretation of our findingis that slow acetylators may be more susceptible to gout.
We also found that an indicator of past browning of fish
(color of fish surface) was directly associated with acetybationactivity, after adjusting for NAT2 genotype and P4501A2 ac-
tivity. Dietary factors other than alcohol (see below) have notpreviously been associated with N-acetyltransferase activity.Procarcinogens, such as HAAs and PAHs, are formed on thesurface of protein-rich foods, such as meat and fish, when they
are cooked at high temperatures. Thus, our finding is suggestiveof an association between a diet rich in these procarcinogens
and acetylation activity. This is in contrast with the lack ofchange in N-acetybtransferase activity observed by Sinha et a!.(32) in subjects who consumed meat cooked at high tempera-
tures containing high amounts of HAAs for 1 week. In theirstudy, exposure to PAHs did not increase with the interventiondiet. Thus, the possibility that N-acetyltransferase is induced by
PAHs but not HAAs would be consistent with the interventionstudy, as well as ours. The most likely explanation, however, is
that our findings have no biological basis and merely reflect a
chance statistical correlation between the rapid acetylation phe-notype and this panicubar diet in our population.
on June 16, 2018. © 1996 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from
><
U.. i
<I
LU I
1-� a.<I#{188}�) �i
zLU I
a- I
z:Q12-
�L:� � 0.40
#{188}.) O�
< d� 0
I
I� ,
. . 8.’. I- . .-
,� I _ 1
I
IHI
Fig. 2. Distribution of the metabolic ratio (AFMU/1X) by
NAT2 genotype.
“Log(AFMU/lX+ I).5 Potential covariates included sex; age; race; years of schooling; daily intake of
red meat, chicken and fish, processed meat and cruciferous vegetables; color of
meat surface; color of fish surface; cigarettes/day; alcoholic drinks/day; cans of
soda/day; physical activity (METs); use of vitamin A, vitamin E, and a-carotene
supplements; use of aspirin, menopausal estrogens, acetaminophen, nonsteroidal
anti-inflammatory agents, and gout medications; family history of colorectalcancer; personal history of diabetes; weight; and (I7U + 17X)/l37X.
‘ Variables listed in order of entry into the stepwise regression. Genotype was
forced to enter the model.
,! Cumulative R2.
454 Predictors of N-Acetyltransferase Activity
Homozygous Heterozygous Homozygous
Mutant Wildtype
NAT2 GENOTYPE (Ml, M2, M3)
Table 4 Stepwise regression of acetylation activ ity” on NAT2 genotype and
other covariates”
.Variables
Regression.
coefficientP
�,R
NAT2 genotype Heterozygous -0.379 <0.001
(WT/WT is reference) Homozygous mutant -0.927 <0.001 0.54
Gout medications -0.386 0.002 0.58
(nonuser is reference)
Color of fish surface Moderately brown -0.01 1 0.90
(lightly hrown/ Heavily brown 0.207 0.02
didn’t eat is reference) 0.63
PA5OIA2 activity 0.104 0.03 0.65
[(l7U + 7X)/l37Xl
Our results are in agreement with past reports showing no
association of acetybation activity with adult age (1, 17, 31),smoking (17, 31, 33), physical activity (31), and weight (17,34), although body weight has previously been associated withthe acetylation of sulphamethazine (35), isoniazid (36), andcaffeine (3 1). Acute alcohol ingestion has also been shown toincrease acetybation activity, possibly because of the increasedavailability of acetate, a metabolite of ethanol (9). In our data,usual alcohol intake during the two preceding weeks was
(negatively) associated with acetylation activity in the univari-ate analysis but not after adjusting for genotype.
We failed to find an association between cruciferousvegetable intake and acetybation activity. This is in agreement
with an intervention study conducted in nine individuals by
Vistisen et a!. (31), in which consumption of 500 g of broccoli
per day for 10 days did not affect caffeine acetylation.
Genotype, gout medication, and diet accounted for almost
two-thirds of the variance in acetylation activity. A certain
degree of genotypic misclassification may explain part of the
remaining variance. We were able to document misclassifica-
lion on genotype in our sample for four individuals who had
two variant alleles but were found to have higher metabolicratios than expected, possibly because of unrecognized molec-
ular differences either in the gene or in the regulation of the
expression of the gene, RNA, or protein. The existence of
unrecognized NAT2 mutant alleles associated with the slow
phenotype was not an issue, however, because the three slowalleles recently identified are known to occur at very low
frequency in Caucasians and Orientals ( 1 1 , 37) and because theMI, M2, and M3 mutations assessed in our study accounted for
all of the slow acetylator phenotypes present in our sample.Finally, the genotype classification was unable to capture the
large variation in metabolic ratio observed among individualswho were homozygous wild-type. This opens the possibility
that a gradient in cancer risk exists within this genotype cate-
gory that would be important to identify. It is possible that
certain unidentified allelic forms may be associated with an
ultrarapid phenotype and carry a high risk.
Our use of cancer patients in this study is unlikely to have
introduced any additional variation in acetylation activity be-
cause they all had an early in situ tumor and an intact colon at
the time of study and because very similar associations were
found for cases and controls (see Table 1). Other unmeasuredenvironmental factors may have contributed to the unexplainedvariance in acetylation activity. However, this component is
likely to have been small because the week-to-week variation in
activity was not barge and because the imprecision in the
measurement of AFMU/1X may have accounted for a signifi-cant pan of the remaining variation.
This study suggests that using genotyping (without phe-
notyping) to assess the association of acetylation with cancer is
unlikely to introduce a sizable misclassification in exposure or
on June 16, 2018. © 1996 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from

Cancer Epidemiology, Biomarkers & Prevention 455
bias in the risk estimates, especially when all three genotypecategories are modeled and the sample size is barge. Assessingcertain dietary factors (i.e. , browning of meat and fish surfaces)and therapeutic drugs (i.e., gout medications) may modestly
improve the precision of the risk estimates. However, as pre-viously pointed out by Rothman et a!. (17), when the rapid
acetybation phenotype is the at-risk category (e.g., when study-ing colon cancer), it would seem judicious to phenotype be-
cause of the large variability in acetylation existing within thisgroup and the importance of identifying a potential subgroup ofindividuals at very high risk.
Acknowledgments
We thank the Hawaii Tumor Registry and the following hospitals for their support
of the study: Castle Medical Center, Kaiser-Permanente Medical Center, Kuakini
Medical Center, Queen’s Medical Center, Straub Clinic and Hospital. St. Francis
Medical Center, Tripler Medical Center, and Wahiawa General Hospital. We are
indebted to Dr. Douglas A. Bell (National Institute of Environmental Health
Sciences) for help in setting up the NAT2 genotyping assay in our laboratory and
for retesting samples that were found discordant with the phenotype in this study.
We also thank Sau Ling Hicks for her programming assistance.
References
I. Caporaso, N., Landi, M. T., and Vineis, P. Relevance of metabolic polymor-
phisms to human carcinogenesis: evaluation of epidemiologic evidence. Pharma-
cogenetics, 1: 4-19, 1991.
2. Ilett, K. F., David, B. M., Detchon, P., Castleden, W. M.. and Kwa, R.Acetylation phenotype in colorectal carcinoma. Cancer Res., 47: 1466-1469.
1987.
3. Ellard, G. A. Variations between individuals and populations in the acetylation
of isoniazid and its significance for the treatment of pulmonary tuberculosis. Clin.
Pharmacol. & Ther., 19: 610-625, 1976.
4. Le Marchand, L., and Kolonel, L. N. Cancer in Japanese migrants to Hawaii:
interaction between genes and environment (French). Rev. Epidemiol. Sante
Publique, 40: 425-430, 1992.
5. Blum, M., Grant, D. M., McBride. W.. Heim, M., and Meyer, U. A. Human
arylamine N-acetyltransferase genes: isolation, chromosomal localization, and
functional expression. DNA Cell Biol., 9: 193-203, 1990.
6. Weber, W. W., and Vatsis, K. P. Individual variability in p-aminobenzoic acid
N-acetylation by human N-acetyltransferase (NAT1 ) of peripheral blood. Phar-
macogenetics, 3: 209-12, 1993.
7. Bell, D. A., Stephens, E. A., Castranio, T., Umbach, D. M., Watson, M.,
Deakin, M., Elder, J., Hendrickse, C., Duncan, H., and Strange. R. C. Polyadenyl-
ation polymorphism in the acetyltransferase I gene (NATI) increases risk of
colorectal cancer. Cancer Res., 55: 3537-3542, 1995.
8. Grant, D. M., Tang, B. K., and Kalow, W. A simple test for acetylator
phenotype using caffeine. Br. J. Clin. Pharmacol., /7: 459-464, 1984.
9. Olsen, H., and Morland, J. Ethanol-induced increase in procainamide acety-
lation in man. Br. J. Clin. Pharmacol.. /3: 203-208, 1982.
10. Blum, M., Demierre, A., Grant, D. M., Heim, M.. and Meyer, U. A. Molec-
ular mechanism of slow acetylation of drugs and carcinogens in humans. Proc.
NatI. Acad. Sci. USA, 88: 5237-5241, 1991.
I I . Bell, D. A., Taylor, J. A., Butler. M. A., Stephens, E. A., Wiest, J., Brubaker,
L. H., Kadlubar, F. F., and Lucier, G. W. Genotype/phenotype discordance for
human arylamine N-acetyltransferase (NAT2) reveals a new slow-acetylator
allele common in African-Americans. Carcinogenesis (Land.), 14: 1689-1692,1993.
12. Hayes, R. B., Bi. W.. Rothman, N.. Broly, F., Caporaso, N., Feng. P.. You.X.. Yin, S., Woosley, R. L., and Meyer. U. A. N-Acetylation phenotype and
genotype and risk of bladder cancer in benzidine-exposed workers. Carcinogen-
esis (Lond.). 14: 675-678, 1993.
13. Ambrosone, C. B.. Freudenheim, J. L., Marshall, J. R., Graham. S., Vena, J.E., Brasure, J. R., Michalek, A. M., Bowman, E. D., Harrington, A. M., and
Shields, P. G. N-Acetyltransferase (NAT), cigarette smoking, and breast cancer
risk. Proc. Am. Assoc. Cancer Rca., 36: 283, 1995.
14. Deguchi. T., Mashimo, M., and Suzuki, T. Correlation between acetylator
phenotypes and genotypes of polymorphic arylamine N-acetyltransferase in hu-
man liver. J. Biol. Chem., 265: 12757-12760, 1990.
15. Hickman, D., and Sim. E. N-Acetyltransferase polymorphism. Comparison of
phenotype and genotype in humans. Biochem. Pharmacol., 42: 1007-1014, 1991.
16. Graf, T., Broly, F., Hoffmann. F., Probst, M., Meyer. U. A.. and Howald, H.
Prediction of phenotype for acetylation and for debrisoquine hydroxylation byDNA-tests in healthy human volunteers. Eur. J. Clin. Pharmacol., 43: 399-403.
1992.
17. Rothman, N., Hayes, R. B., Bi, W., Caporaso. N.. Broly. F., Woosley, R. L.,
Yin, S., Feng, P., You, X., and Meyer, U. A. Correlation between N-acetyltrans-
ferase activity and NAT2 genotype in Chinese males. Pharmacogenetics. 3:
250-255, 1993.
18. Rothman, N.. Stewart, W. F., Caporaso, N. E., and Hayes. R. B. Misclassi-fication of genetic susceptibility biomarkers: Implications for case-control studies
and cross-population comparisons. Cancer Epidemiol.. Biomarkers & Prey., 2:
299-303, 1993.
19. Sivaraman. L., Leatham, M. P., Yee, J., Wilkens, L. R., Lau. A. F., and Le
Marchand, L. CYP1AI genetic polymorphisms and in situ colorectal cancer.
Cancer Res., 54: 3692-3695, 1994.
20. Hankin, J. H., Wilkens, L. R., Kolonel, L. N., and Yoshizawa, C. N.Validation of a quantitative diet history method in Hawaii. Am. J. Epidemiol..
133: 616-628, 1991.
21. Sallis. J. F.. Haskell, W. L., Wood, P. D., Fortmann, S. P., Rogers, T., Blair.
S. N., and Paffenbarger, R. S.. Jr. Physical activity assessment methodology in the
Five-City Project. Am. J. Epidemiol., 121: 91-106, 1985.
22. Butler, M. A., Lang, N. P., Young. J. F., Caporaso, N. E., Vineis, P., Hayes.
R. B., Teitel, C. H., Massengill. J. P.. Lawsen. M. F., and Kadlubar, F. F.
Determination of CYPIA2 and NAT2 phenotypes in human populations by
analysis of caffeine urinary metabolites. Pharmacogenetics. 2: 1 16-127, 1992.
23. Ausubel, F. M.. Brent, R., Kingston. R. E.. Moore, D. D., Seidman, J. G..
Smith J. A., and Struhl, K. Current Protocols in Molecular Biology. Vol. 1 . JohnWiley & Sons. Inc., 1994.
24. Snedecor, G. W., and Cochran, W. G. Statistical Methods, Ed. 6. Ames. IA:
Iowa State University Press. 1967.
25. Finney, D. J. Probit Analysis. Cambridge. United Kingdom: Cambridge
University Press, I 971.
26. Hardy, B. G., Lemieux, C., Walker, S. E., and Bartle, W. R. Interindividual
and intraindividual variability in acetylation: characterization with caffeine. Clin.
Pharmacol. Ther., 44: 152-157, 1988.
27. McQuilkin. S. H., Nierenberg. D. W.. and Bresnick. E. Analysis of within-
subject variation of caffeine metabolism when used to determine cytochrome
P4501A2 and N-acetyltransferase-2 activities. Cancer Epidemiol.. Biomarkers &
Prey.. 4: 139-146, 1995.
28. Hashiguchi, M., and Ebihara, A. Acetylation polymorphism of caffeine in a
Japanese population. Clin. Pharmacol. Ther., 52: 274-276, 1992.
29. Horai. Y., Zhou, H-H., Zhang, L-M., and Ishizaki, T. N-Acetylation pheno-
typing with dapsone in a mainland Chinese population. Br. J. Clin. Pharmacol.,
25: 81-87. 1988.
30. Lin, H. J., Han, C-Y., Lin, B. K., and Hardy, S. Ethnic distribution of slow
acetylator mutations in the polymorphic N-acetyltransferase (NAT2) gene. Phar-macogenetics. 4: 125-134, 1994.
31. Vistisen, K., Poulsen, H. E.. and Loft, S. Foreign compound metabolism
capacity in man measured from metabolites of dietary caffeine. Carcinogenesis(Lond.), 13: 1561-1568, 1992.
32. Sinha, R., Rothman, N., Brown, E. D., Mark, S. D., Hoover, R. N., Caporaso.
N. E., Levander. 0. A., Knize, M. G.. Lang, N. P., and Kadlubar, F. F. Pan-fried
meat containing high levels of heterocyclic aromatic amines but low levels of
polycyclic aromatic hydrocarbons induces cytochrome P4501A2 activity in hu-
mans. Cancer Res., 54: 6154-6159, 1994.
33. Miller, M. E., and Cosgriff, J. M. No effect of smoking on sulfamethazine
acetylation. Drug Intell. Clin. Pharm.. /8: 620-622, 1984.
34. Weber. W. W. The Acetylator Genes and Drug Response, pp. 164-165. NewYork: Oxford University Press, 1987.
35. Chapron, D. J., Kramer, P. A., and Mercik, S. A. Potential influence of body
weight on the clearance of polymorphically acetylated drugs. J. Clin. Pharmacol.,
22: 271-275. 1982.
36. Evans. D. A. P., Storey. P. B., and McKusick, V. A. Further observations on
the determination of the isoniazid inactivator phenotype. Johns Hopkins Bull.,
108: 60-66, 1961.
37. Hickman. D., Risch, A., Camilleri, J. P., and Sim, E. Genotyping in human
polymorphic arylamine N-acetyltransferase: identification of new slow allotypic
variants. Pharmacogenetics, 2: 217-226. 1992.
on June 16, 2018. © 1996 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from

1996;5:449-455. Cancer Epidemiol Biomarkers Prev L Le Marchand, L Sivaraman, A A Franke, et al. epidemiological studies?phenotyping and NAT2 genotyping be used interchangeably in Predictors of N-acetyltransferase activity: should caffeine
Updated version
http://cebp.aacrjournals.org/content/5/6/449
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cebp.aacrjournals.org/content/5/6/449To request permission to re-use all or part of this article, use this link
on June 16, 2018. © 1996 American Association for Cancer Research. cebp.aacrjournals.org Downloaded from


















