
Drug-Free Induction of a Chloramphenicol Acetyltransferase Gene ...
7
JOURNAL OF BACTERIOLOGY, Sept. 1987, p. 4235-4241 Vol. 169, No. 9 0021-9193/87/094235-07$02.00/0 Copyright © 1987, American Society for Microbiology Drug-Free Induction of a Chloramphenicol Acetyltransferase Gene in Bacillus subtilis by Stalling Ribosomes in a Regulatory Leader ELIZABETH J. DUVALL, NICHOLAS P. AMBULOS, JR., AND PAUL S. LOVETT* Department of Biological Sciences, University of Maryland, Baltimore County, Catonsville, Maryland 21228 Received 27 April 1987/Accepted 16 June 1987 The plasmid gene cat-86 is induced by chloramphenicol in Bacillus subtilis, resulting in the synthesis of the gene product chloramphenicol acetyltransferase. Induction is due to a posttranscriptional regulatory mecha- nism in which the inducer, chloramphenicol, activates translation of cat-86 mRNA. We have suggested that chloramphenicol allows ribosomes to destabilize a stem-loop structure in cat-86 mRNA that sequesters the ribosome-binding site for the coding sequence. In the present report we show that cat-86 expression can be activated by stalling ribosomes in the act of translating a regulatory leader peptide. Stalling was brought about by starving host cells for specific leader amino acids. Ribosomal stalling, which led to cat-86 expression, occurred upon starvation for the amino acid specified by the leader codon located immediately 5' to the RNA stem-loop structure and was independent of whether that codon specified lysine or tyrosine. These observations support a model for chloramphenicol induction of cat-86 in which the antibiotic stalls ribosome transit in the regulatory leader. Stalling of ribosomes in the leader can therefore lead to destabilization of the RNA stem-loop structure. cat genes from gram-positive bacteria are inducibly ex- pressed, and chloramphenicol is the inducer (31). The cat gene product is chloramphenicol acetyltransferase (CAT) (29). Hence, the inducer for cat genes is also the substrate for the corresponding enzyme, which inactivates the antibi- otic activity of chloramphenicol (29). Induction of cat genes was proposed to represent a regulatory event that primarily modulates translation of cat mRNA (13). It was suggested that chloramphenicol modifies ribosomal function, permit- ting ribosomes to destabilize a stem-loop structure in cat mRNA which sequesters the cat ribosome-binding site (RBS) (13, 19). Evidence that is consistent with this post- transcriptional regulatory model has been provided for the cat-86 gene, as well as for inducible cat genes resident on the Staphylococcus plasmids pC194 and pUB112 (1, 2, 4, 7, 8, 11, 12, 19, 22, 27). However, despite efforts in several laboratories, there is no experimental evidence to support models proposed to explain how a ribosome might disrupt a cat RNA stem-loop structure (7, 8, 19). Deletion studies have shown that a short leader region 5' to the RNA stem-loop structure is essential for inducible expression of cat-86 and the pUB112 cat gene (2, 7). The leader region consists of an RBS and an open reading frame of nine codons. Translation of the proposed leader peptide would deliver ribosomes into the left stem of the RNA stem-loop structure. One mechanism through which chlor- amphenicol might cause destabilization of the RNA stem- loop structure involves the stalling of translating ribosomes at a specific location in the leader. To test this hypothesis, we attempted to stall translation at different locations in the leader by starving host cells for leader amino acids. In the present study, we showed that starving Bacillus subtilis for an amino acid specified by the codon located immediately 5' to the RNA stem-loop structure activates expression of cat-86. * Corresponding author. MATERIALS AND METHODS Bacterial strains and plasmids. The strains of B. subtilis used in this study were derivatives of B. subtilis 168. These included BR151 (trpC2 lys-3 metB10) and IS1 (trpC2 thr-5) from I. Smith and 1A206 (trpC2 tyrA) from the Bacillus Genetic Stock Center, Ohio State University, Columbus. Plasmid pPL703 (Fig. 1) and the mutant derivatives pPL703- Thr and pPL703-Tyr were used exclusively. pPL703 (23) is a 5,032-base-pair (bp) plasmid maintained in B. subtilis at a copy number of about 35. The plasmid contains two drug resistance genes. The pUB110 moiety of pPL703 harbors a gene that specifies kanamycin and neomycin resistance. The second drug resistance gene on the plasmid is a promoter- deficient version of cat-86, a gene previously cloned from B. pumilus (38), which specifies chloramphenicol-inducible CAT when the gene is activated by inserting a promoter at a site 144 bp 5' to the cat-86 coding sequence. In derivatives of pPL703 used in the current work, cat-86 was activated by inserting the P4 promoter fragment, from phage SP02, 5' to cat-86 (23). Most of the experiments reported were also performed with plasmids in which cat-86 was activated with the Spac promoter (39). Regardless of the choice of pro- moter, the results were comparable. Plasmid isolation, growth media, and growth conditions were as previously described (20, 23). Assay for CAT specific activity. CAT was assayed by the colorimetric procedure described by Shaw (30). Protein was measured by the method of Bradford (6). CAT specific activity was the amount (in micromoles) of chloramphenicol acetylated per minute per milligram of protein at 25°C. Amino acid starvation experiments. pPL703-P4 or mutant derivatives were transformed into B. subtilis strains having specific nutritional requirements. Cells were grown to log phase at 37°C in Spizizen minimal medium (34) supple- mented with all 20 naturally occurring amino acids at 50 pg/mt. Cells taken from log phase were concentrated by centrifugation, washed twice with cold minimal medium (no 4235
Transcript of Drug-Free Induction of a Chloramphenicol Acetyltransferase Gene ...

JOURNAL OF BACTERIOLOGY, Sept. 1987, p. 4235-4241 Vol. 169, No. 90021-9193/87/094235-07$02.00/0Copyright © 1987, American Society for Microbiology
Drug-Free Induction of a Chloramphenicol Acetyltransferase Genein Bacillus subtilis by Stalling Ribosomes in a Regulatory Leader
ELIZABETH J. DUVALL, NICHOLAS P. AMBULOS, JR., AND PAUL S. LOVETT*Department of Biological Sciences, University of Maryland, Baltimore County, Catonsville, Maryland 21228
Received 27 April 1987/Accepted 16 June 1987
The plasmid gene cat-86 is induced by chloramphenicol in Bacillus subtilis, resulting in the synthesis of thegene product chloramphenicol acetyltransferase. Induction is due to a posttranscriptional regulatory mecha-nism in which the inducer, chloramphenicol, activates translation of cat-86 mRNA. We have suggested thatchloramphenicol allows ribosomes to destabilize a stem-loop structure in cat-86 mRNA that sequesters theribosome-binding site for the coding sequence. In the present report we show that cat-86 expression can beactivated by stalling ribosomes in the act of translating a regulatory leader peptide. Stalling was brought aboutby starving host cells for specific leader amino acids. Ribosomal stalling, which led to cat-86 expression,occurred upon starvation for the amino acid specified by the leader codon located immediately 5' to the RNAstem-loop structure and was independent of whether that codon specified lysine or tyrosine. These observationssupport a model for chloramphenicol induction of cat-86 in which the antibiotic stalls ribosome transit in theregulatory leader. Stalling of ribosomes in the leader can therefore lead to destabilization of the RNA stem-loopstructure.
cat genes from gram-positive bacteria are inducibly ex-pressed, and chloramphenicol is the inducer (31). The catgene product is chloramphenicol acetyltransferase (CAT)(29). Hence, the inducer for cat genes is also the substratefor the corresponding enzyme, which inactivates the antibi-otic activity of chloramphenicol (29). Induction of cat geneswas proposed to represent a regulatory event that primarilymodulates translation of cat mRNA (13). It was suggestedthat chloramphenicol modifies ribosomal function, permit-ting ribosomes to destabilize a stem-loop structure in catmRNA which sequesters the cat ribosome-binding site(RBS) (13, 19). Evidence that is consistent with this post-transcriptional regulatory model has been provided for thecat-86 gene, as well as for inducible cat genes resident on theStaphylococcus plasmids pC194 and pUB112 (1, 2, 4, 7, 8,11, 12, 19, 22, 27). However, despite efforts in severallaboratories, there is no experimental evidence to supportmodels proposed to explain how a ribosome might disrupt acat RNA stem-loop structure (7, 8, 19).
Deletion studies have shown that a short leader region 5'to the RNA stem-loop structure is essential for inducibleexpression of cat-86 and the pUB112 cat gene (2, 7). Theleader region consists of an RBS and an open reading frameof nine codons. Translation of the proposed leader peptidewould deliver ribosomes into the left stem of the RNAstem-loop structure. One mechanism through which chlor-amphenicol might cause destabilization of the RNA stem-loop structure involves the stalling of translating ribosomesat a specific location in the leader. To test this hypothesis,we attempted to stall translation at different locations in theleader by starving host cells for leader amino acids. In thepresent study, we showed that starving Bacillus subtilis foran amino acid specified by the codon located immediately 5'to the RNA stem-loop structure activates expression ofcat-86.
* Corresponding author.
MATERIALS AND METHODS
Bacterial strains and plasmids. The strains of B. subtilisused in this study were derivatives of B. subtilis 168. Theseincluded BR151 (trpC2 lys-3 metB10) and IS1 (trpC2 thr-5)from I. Smith and 1A206 (trpC2 tyrA) from the BacillusGenetic Stock Center, Ohio State University, Columbus.Plasmid pPL703 (Fig. 1) and the mutant derivatives pPL703-Thr and pPL703-Tyr were used exclusively. pPL703 (23) is a5,032-base-pair (bp) plasmid maintained in B. subtilis at acopy number of about 35. The plasmid contains two drugresistance genes. The pUB110 moiety of pPL703 harbors agene that specifies kanamycin and neomycin resistance. Thesecond drug resistance gene on the plasmid is a promoter-deficient version of cat-86, a gene previously cloned from B.pumilus (38), which specifies chloramphenicol-inducibleCAT when the gene is activated by inserting a promoter at asite 144 bp 5' to the cat-86 coding sequence. In derivatives ofpPL703 used in the current work, cat-86 was activated byinserting the P4 promoter fragment, from phage SP02, 5' tocat-86 (23). Most of the experiments reported were alsoperformed with plasmids in which cat-86 was activated withthe Spac promoter (39). Regardless of the choice of pro-moter, the results were comparable. Plasmid isolation,growth media, and growth conditions were as previouslydescribed (20, 23).
Assay for CAT specific activity. CAT was assayed by thecolorimetric procedure described by Shaw (30). Protein wasmeasured by the method of Bradford (6). CAT specificactivity was the amount (in micromoles) of chloramphenicolacetylated per minute per milligram of protein at 25°C.Amino acid starvation experiments. pPL703-P4 or mutant
derivatives were transformed into B. subtilis strains havingspecific nutritional requirements. Cells were grown to logphase at 37°C in Spizizen minimal medium (34) supple-mented with all 20 naturally occurring amino acids at 50pg/mt. Cells taken from log phase were concentrated bycentrifugation, washed twice with cold minimal medium (no
4235

4236 DUVALL ET AL.
A.
Hind III
Xboter
Bgl Il/BomH
B.
RBS-25' * * |GAAAGGA|
AA
sU
cGCGUAJUGAUUGUG~j~UGAAAACAGAUAAAAUAAAAf2!---J
FIG. 1. Diagram of plasmid pPL703 and the RNA stem-loopstructure. (A) Plasmid pPL703, which consists of a 1,250-bp PstI-BglII fragment of B. pumilus NCIB8600 DNA inserted between theEcoRI and BamH4I sites of pUB110 by use of a 21-bp EcoRI-PstIfragment from M13mp7 (23). The promoterless gene cat-86 resideswithin the 1,250-bp fragment, and the gene is followed by an efficienttranscription termination signal designated ter (24). The pUB110portion of pPL703 provides an origin of replication (Ori) and aneomycin resistance (Neot) gene (15, 17). cat-86 specifies CATwhen the gene is transcriptionally activated by insertion of apromoter into any of four unique restriction sites 5' to cat-86:EcoRI, BamHI, Sall, or PstI. RBS-1, RBS-2, and RBS-3 designatethe approximate locations ofRBSs identified by their homology with16S rRNA (2). P1 and P3 are transcripts in the plasmid that do notenter cat = 86. (B) Secondary structure of the RNA in the vicinity ofRBS-3.amino acid supplements), and suspended in Spizizen mini-mal medium containing all amino acids except for the one forwhich the cells were to be starved; e.g., BR151 requireslysine, and therefore lysine starvation was achieved byomitting lysine from the second growth medium. Aftersuspension, the cells were placed at 37°C and a sample wasimmediately taken to assay for CAT (zero-time sample).Samples were taken every 20 min thereafter for 2 h andassayed for CAT and total protein. In some experiments,CAT was assayed in cells before washing and immediatelyafter washing and no change in CAT specific activity (<5%difference) was detected. Mock induction was performedonly when starvation for a specific amino acid was shown toactivate cat-86 expression. For example, starvation forlysine activates expression of the wild-type cat-86 gene inBR151 (pPL703-P4). Mock starvation was performed exactlyas for lysine starvation, with the single exception that lysine(100 ,ug/ml) was present in the second growth medium. Mockstarvation consistently failed to activate cat-86 expression.
Site-directed mutagenesis. Two codons in the cat-86 leader2 region were converted to codons specifying other aminoacids (Fig. 2). Lysine codon 1, AAA, was converted to thethreonine codon ACA, generating cat-86-Thr, which is the
single mutation in the plasmid we designate pPL703-Thr.Lysine codon 2, AAA, was separately changed to TAT,which specifies tyrosine. This mutation generates cat-86-Tyrin pPL703-Tyr.To insert the nucleotide changes into the leader 2 region,
a 637-bp EcoRI-HindlIl fragment was cloned from pPL703into M13mpl9. The insert caused the M13 replicative form togenerate white plaques when transfected into Escherichiacoli KK2186 and plated on YT medium containing 5-bromo-4-chloro-3-indolyl ,-D-galactopyranoside (X-Gal) at 50,ug/ml (40). Single-stranded DNA from progeny phage wasannealed with a 20-mer that had been synthesized to achieveinsertion of the desired nucleotide change(s) (40). The single-stranded circle was converted to a duplex circle by extend-ing the 20-mer with the Klenow fragment of DNA polymer-ase I in the presence of dATP, dTTP, dGTP, and thethiolated nucleotide dCTPaS. All steps were as specified inthe product literature for the Amersham mutagenesis kit.The duplex circle was sealed with T4 DNA ligase, nickedwith NciI, and digested for 0.5 h with exonuclease III(Amersham product literature and reference 35). The duplexcircle was resynthesized with the Klenow fragment and thefour non-sulfur-containing nucleotides and sealed with T4DNA ligase. This mixture was transfected into Escherichiacoli. Five plaques were selected, and single-stranded DNAfrom each was used to run a single-lane screen to determinewhether the desired mutation had been inserted. In bothcases, phage from at least four of the plaques contained thedesired mutation. DNA from one phage was subjected to thefull sequencing reaction. After confirmation that the onlychange in the sequence was the desired mutation, thesingle-stranded phage DNA was annealed with the universalsequencing primer, and the entire insert in M13 was con-verted to a duplex state by DNA synthesis with the Klenowfragment. The duplex insert was cut out as a 616-bp PstI-HindlIl fragment. This fragment was inserted into a receptorversion of pPL703 in which the cat-86 gene is transcribedbecause of the presence of the P4 promoter fragment in theupstream linker. However, the single BclI site (located in thecat-86 coding sequence 5' to the HindIII site; Fig. 1) hadbeen previously gap filled. The gap filling at the BclI siteinactivated CAT activity. Successful replacement of thePstI-HindIII region with the mutant fragment from M13 wasdetected because it restored cat-86 expression. Subsequentto this construction, the PstI-HindIII fragment was reclonedfrom the mutant pPL703 into M13 to confirm by sequencingthat the mutation was indeed present in the plasmid. Se-quencing was by the method of Sanger et al. (28).
RESULTS
Experimental rationale. Inducible regulation of cat-86 isdue to sequences that intervene between the cat-86 codingsequence and the promoter for the gene (22). This tran-scribed region 5' to cat-86 in promoter-containing versions
Thr TyrC T T
I 1
...GAAAGGTGATTGTGrGTG,GTG AAAATCTCCTCCTGAAT....RBS-2 Met Val Lys Thr Asp Lys le Ser Ser *
1 2
FIG. 2. Mutations inserted into the leader 2 region of cat-86 bysite-directed mutagenesis. Lysine codons 1 and 2 were separatelymutagenized to codons for threonine (ACA) or tyrosine (TAT). Thehatched region is conserved region I. The horizontal arrow showsthe left member of the inverted repeats.
J. BACTERIOL.

cat-86 INDUCTION BY AMINO ACID STARVATION 4237
ConservedRegion
I 1I
cat-86 5'--- GAAAGGATGATTGTGGTGGTGAAACAGATAAAATCTCCTCCTGAAzACAGTAAATCACATTCAGGAGGAGATAAAATTG ---
RBS-2 Met Vol Lys Thr Asp Lys lIe Ser Ser * RBS-3 Met (cot)
c at-66 5'--- GAAAGGATGATTGTGGTGGTGAAAACAGATAAAGTCTCCTCCTGAATACAGTAAATCACACATTCAGGAGGAGATAAAAAATG ---
RBS-2 Met Val Lys Thr Asp Lys Val Ser Ser * RBS-3 Met (cot)
cat-57 5'--- GAAAGGATGATTGTGGTGGTGAAAGCAGATAAAGTTTCCTCCTGAATACAGTGTATCACATTCAGGAGGAATAAAAATG ---
RBS-2 Met Val Lys Ala Asp Lys Val Ser Ser * RBS-3 Met(cat)
pC2 21cat 5' --- GGGGTGACATAACGTATGAAAAATCAGAGGATTATTCCCCTAAATATAAAGATTTAAAATTTAGGAGGAAATTATATATG ---
RBS-1 Met Lys Lys Ser Glu Asp Tyr Ser Scr * RBS-2 Met (cot)
pUB112cat 5' --- GAGGTGATATAACGTATGAAAAAATCAGAGGATTATTCCTCCTAMATACAAAGATTTAAAATTTAGGAGGAATTTATATATG ---
RBS-1 Met Lys Lys Ser Glu Asp Tyr Ser Ser RBS-2 Met(cot)
PC19 4cat 5' --- AAAGGATTGATTCTAATGAAGAAAGCAGACAAGTMGCCTCCTAAATTCACTTTAGATAAAAATTTAGGAGGCATATCAAATG ---
RBS-1 Met Lys Lys Ala Glu Lys * Ala Ser * RBS-2 Met (cot)
FIG. 3. Leader regions 5' to the coding sequences for six inducible cat genes. Conserved regions I and II are found in all cat leaders, andthe distance between these regions is 8 nucleotides. The two small vertical arrows designate the bases in conserved regions I and II that varyamong the leaders. The sequences are from references 2, 7, 13, 14, 16, and 32. The leader sequence shown for cat-86 and related genes (cat-66and cat-57) is referred to as leader 2 since in cat-86 another leader sequence (leader 1) is located 5' to leader 2. Leader 1 is not essential tochloramphenicol induction of cat-86 (2). Regulatory regions for cat genes on pC221, pC194, and pUB112 contain only the leader shown. Theasterisks indicate translational stop codons; the horizontal arrows show inverted-repeat sequences.
of pPL703 consists of 144 bp, and chloramphenicol-inducibleexpression is retained after removal of the first 60 bp (2).Removal of 82 bp, however, blocks induction (2). The A60mutant, therefore, reveals the essential sequence informa-tion 5' to cat-86 needed to achieve induction. This sequenceincludes RBS-2 and leader 2 in addition to the sequences thatdictate the formation of the RNA stem-loop structure (Fig.3). Sequences analogous to RBS-2 and leader 2 occur 5' tothe RNA stem-loop structure for all of the inducible catgenes examined (Fig. 3). Furthermore, two regions in leader2 (conserved regions I and II) are highly conserved in all catleaders, and in theory these might dictate the site whereribosomes translating the leader are stalled by chloramphen-icol. However, in the present study we wished to determinewhether translating ribosomes which are stalled in leader 2can indeed destabilize the RNA stem-loop structure andthereby induce translation of cat-86. We therefore per-formed amino acid starvation experiments. The reasoningbehind these experiments is that an uncharged tRNA (e.g.,lysyl tRNA) binding to ribosomes at a lysine codon in leader2 might stall ribosomes at that site (33). Depending on thelocation of the stall site, a stalled ribosome may thenfacilitate destabilization of the RNA stem-loop structure andallow translation of the cat-86 coding sequence if chargedlysyl tRNAs become available to allow insertion of lysineresidues into the CAT structural protein. It was thought thatfree lysine residues, as well as residues of other amino acids,would arise in the cell from protein turnover during starva-tion.
Starvation of B. subtilis for lysine activates cat-86 expres-sion. Two lysine codons (AAA) exist in leader 2. One is inconserved region I and one is at the base of the RNAstem-loop structure (Fig. 2). Starvation of B. subtilis BR151harboring cat-86 (on plasmid pPL703-P4) for lysine in-creased cat-86 expression twofold during the 2-h starvationperiod (Fig. 4). Mock starvation in BR151 did not inducecat-86 (Fig. 4), and starvation for the nonleader amino acidstryptophan (by use of BR151; data not shown) and tyrosine(by use of strain 1A206; Fig. 5) also did not induce cat-86expression. Similarly, starvation for the leader amino acidthreonine by use of a cat-86-containing derivative of strainIS1 did not activate cat-86 expression (data not shown).
Lysine codon 2 in leader 2 is the site of stalling essential tocat-86 induction. To determine whether activation of cat-86by lysine starvation was due to stalling at one of the twolysine codons in leader 2, each of the codons was separatelychanged to a codon for a different amino acid by site-directedmutagenesis. Lysine codon 1 (AAA) was changed to a codonfor threonine (ACA) (Fig. 2). When this mutant gene,designated cat-86-Thr, was tested for induction by lysinestarvation, expression of the gene was higher than wasexpression of wild-type cat-86 during lysine starvation (Fig.4). Mock starvation in BR151 did not induce cat-86-Thr, andcat-86-Thr was not induced when plasmid-containing cells ofstrain IS1 were starved for threonine (data not shown).
In a separate experiment, we changed lysine codon 2 to acodon for tyrosine, TAT (Fig. 2). This change had twoeffects. (i) cat-86-Tyr was unable to be induced by starvationfor lysine in strain BR151 (data not shown). (ii) cat-86-Tyrwas induced when host cells (strain 1A206) were starved fortyrosine (Fig. 5). However, mock starvation in strain 1A206did not induce cat-86-Tyr. Induction of cat-86-Tyr by tyro-sine starvation consistently resulted in CAT levels that wereabout half of the CAT level resulting from activation ofcat-86-Thr by lysine deprivation. The basis for this differ-ence is not known.The data suggest that the ribosomal stall site that leads to
cat-86 induction is at or in the vicinity of lysine codon 2 inleader 2. Consistent with this interpretation is our observa-tion that changing lysine codon 1 in the regulatory leader toa codon for threonine allowed lysine starvation to inducecat-86 expression above the level obtained when both thefirst and second lysine codons were intact. It is likely thatwhen both lysine codons are intact there is an equal proba-bility of ribosomal stalling at either codon during lysinestarvation. Since the first and second lysine codons areseparated by only 6 nucleotides, it is not possible for tworibosomes to stall simultaneously at the two codons becausean individual ribosome reportedly spans about 35 nucleo-tides in mRNA (25). Changing the first lysine codon to athreonine codon therefore allowed all ribosomal stalling inthe leader due to lysine starvation to occur at the locationwhich leads to activation of cat-86 expression.
Induction of cat-86-Thr and cat-86-Tyr by antibiotics. We
VOL. 169, 1987
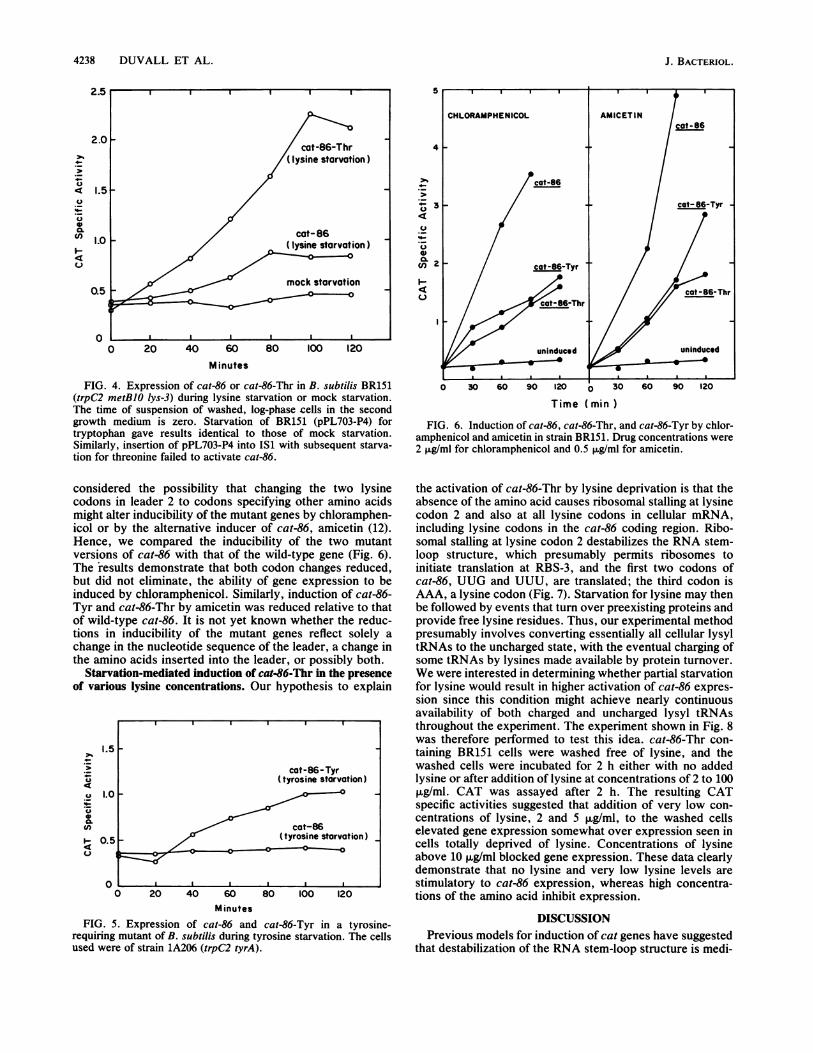
4238 DUVALL ET AL.
(lysine starvation)
4 1.5 -
.2
i_ 1.0 cat -o86(lysine starvation)
mock starvation0.5 AC0o
0 20 40 60 80 100 120
M inutes
FIG. 4. Expression of cat-86 or cat-86-Thr in B. subtilis BR151(trpC2 metB10 Iys-3) during lysine starvation or mock starvation.The time of suspension of washed, log-phase cells in the secondgrowth medium is zero. Starvation of BR151 (pPL703-P4) fortryptophan gave results identical to those of mock starvation.Similarly, insertion of pPL703-P4 into IS1 with subsequent starva-tion for threonine failed to activate cat-86.
considered the possibility that changing the two lysinecodons in leader 2 to codons specifying other amino acidsmight alter inducibility of the mutant genes by chloramphen-icol or by the alternative inducer of cat-86, amicetin (12).Hence, we compared the inducibility of the two mutantversions of cat-86 with that of the wild-type gene (Fig. 6).The results demonstrate that both codon changes reduced,but did not eliminate, the ability of gene expression to beinduced by chloramphenicol. Similarly, induction of cat-86-Tyr and cat-86-Thr by amicetin was reduced relative to thatof wild-type cat-86. It is not yet known whether the reduc-tions in inducibility of the mutant genes reflect solely achange in the nucleotide sequence of the leader, a change inthe amino acids inserted into the leader, or possibly both.
Starvation-mediated induction of cat-86-Thr in the presenceof various lysine concentrations. Our hypothesis to explain
1.5
4
1.0
0.
cn
i- 0.54u I
0
0 20 40 60 80 100 120Minutes
FIG. 5. Expression of cat-86 and cat-86-Tyr in a tyrosine-requiring mutant of B. subtilis during tyrosine starvation. The cellsused were of strain 1A206 (trpC2 tyrA).
cat-86
3 ~~~~~~~~~~~~~~cat-86-Tyr
Cr 2 at-.6-Tyr
C) cat-86-Thrca-8Th
uninduced uninduced
0 30 60 90 120 o 30 60 90 120
Time (min )
FIG. 6. Induction of cat-86, cat-86-Thr, and cat-86-Tyr by chlor-amphenicol and amicetin in strain BR151. Drug concentrations were2 ,ug/ml for chloramphenicol and 0.5 ,ug/ml for amicetin.
the activation of cat-86-Thr by lysine deprivation is that theabsence of the amino acid causes ribosomal stalling at lysinecodon 2 and also at all lysine codons in cellular mRNA,including lysine codons in the cat-86 coding region. Ribo-somal stalling at lysine codon 2 destabilizes the RNA stem-loop structure, which presumably permits ribosomes toinitiate translation at RBS-3, and the first two codons ofcat-86, UUG and UUU, are translated; the third codon isAAA, a lysine codon (Fig. 7). Starvation for lysine may thenbe followed by events that turn over preexisting proteins andprovide free lysine residues. Thus, our experimental methodpresumably involves converting essentially all cellular lysyltRNAs to the uncharged state, with the eventual charging ofsome tRNAs by lysines made available by protein turnover.We were interested in determining whether partial starvationfor lysine would result in higher activation of cat-86 expres-sion since this condition might achieve nearly continuousavailability of both charged and uncharged lysyl tRNAsthroughout the experiment. The experiment shown in Fig. 8was therefore performed to test this idea. cat-86-Thr con-taining BR151 cells were washed free of lysine, and thewashed cells were incubated for 2 h either with no addedlysine or after addition of lysine at concentrations of 2 to 100jig/ml. CAT was assayed after 2 h. The resulting CATspecific activities suggested that addition of very low con-centrations of lysine, 2 and 5 jig/ml, to the washed cellselevated gene expression somewhat over expression seen incells totally deprived of lysine. Concentrations of lysineabove 10 ,ug/ml blocked gene expression. These data clearlydemonstrate that no lysine and very low lysine levels arestimulatory to cat-86 expression, whereas high concentra-tions of the amino acid inhibit expression.
DISCUSSIONPrevious models for induction of cat genes have suggested
that destabilization of the RNA stem-loop structure is medi-
cat-86-Tyr(tyrosine starvation)
cat-86(tyrosine storvation)-_~~~~~~~-O----1---
I I I I .-I
J. BACTERIOL.

cat-86 INDUCTION BY AMINO ACID STARVATION 4239
RBS-2 2 ca1-8-CD -GUG- GAU*AAA GUUGU AAA
Leader 2 cat-86
RBS-2
FIG. 7. Model for destabilization of the cat-86 RNA stem-loopstructure by ribosomal stalling at lysine codon 2. The asteriskindicates the location of the UGA codon that terminates leader 2.The oval structures are ribosomes.
ated by a ribosome that is stalled in the regulatory leaderregion because of the presence of chloramphenicol. Thepresent stuldy demonstrated that induction of cat-86 can beachieved by physiologically stalling ribosomes in the leaderin the absence of any antibiotic. Our data suggest thatribosome transit must be blocked at a relatively specificlocation in the leader for activation of gene expression. Thisis based on the observation that lysine starvation inducedcat-86 expression only when the lysine codon 2 in leader 2was intact. Since lysine codons 1 and 2 in the leader areseparated by only 6 nucleotides, the actual position of thestalled ribosome relative to the RNA stem-loop structureappears to be of importance for destabilization of the RNAsecondary structure. This interpretation is consistent withthe observation that starvation for one other leader aminoacid, threonine, failed to induce cat-86 detectably. However,the latter negative result must be viewed with some cautionsince these experiments were not adequately controlled. Forinstance, it was not shown that threonine starvation canactivate cat-86 when a threonine codon is used to replacelysine codon 2.
It is possible that induction of cat-86 by starvation forlysine mimics the events occurring during induction bychloramphenicol; perhaps both methods stall ribosomes atthe same approximate location in leader 2. Experiments totest this are under way. One of the interesting interpretationsof our data is that chloramphenicol is apparently not neededto destabilize the RNA stem-loop structure. Rather, chlor-amphenicol may only block ribosome transit at a specificlocation in the regulatory leader, which then allows theribosome and associated factors to facilitate localized melt-ing of the RNA secondary structure. Recent in vitro trans-
lation studies by Narayanan and Dubnau (26) revealed thatchloramphenicol can indeed stall ribosomes at a discretelocation in mRNA.
It has been suggested that drug-induced ribosomal stallingregulates gene expression by altering mRNA secondarystructure and that this is the mechanism through whicherythromycin inducibly regulates expression of erm genes ingram-positive bacteria (9, 37). erm genes confer resistance toerythromycin. Induction by erythromycin activates ermmRNA translation, and this depends upon changes in theconformation of the mRNA in the vicinity of the erm RBS.Inducible expression of erm requires translation of a regula-tory leader peptide, as does expression of inducible catgenes (6a, 10, 21, P. S. Lovett, unpublished data). Erythro-mycin is thought to stall ribosomes at a specific region in theregulatory leader (26), and it would therefore be of interest todetermine whether erm induction can also be achieved bystarvation of host cells for selected erm leader amino acids.The regulatory mechanisms that control inducible expres-
sion of erm and cat genes have been described as transla-tional attenuation because of apparent similarity with (tran-scriptional) attenuation seen in amino acid biosyntheticoperons (7, 9, 18). In classical (transcriptional) attenuation,ribosomal stalling is achieved by starvation for a specificleader amino acid, and a stalled ribosome prevents formationof a transcription termination signal in the mRNA. Hence,ribosomal stalling facilitates readthrough transcription intodownstream sequences (18). Leader mRNA can theoreti-cally assume. either of two conformations. The thermody-namically favored conformation in vitro, and apparently invivo, causes transcription termination. Ribosomal stalling ata precise location in the leader transiently favors the confor-mation of mRNA that is not active in transcription termina-tion. In the ermC system, three conformations of leadermRNA are possible, and two of these sequester the ermRBS. Erythromycin-mediated stalling of ribosomes in theerm leader is thought to prevent formation of the mRNAconformations that block translation by favoring an alterna-tive conformation of the mRNA. In contrast with the above
5.0
4.0
I-
;1.0
.2tt
Wu 2.0cn)
<r 1.0
00 2 S 10 20 30 40 50 60 70 ' 100
Lysine Concentration (pg/ml)
FIG. 8. CAT activity expressed by BR151 cells harboringpPL703-Thr-Spac after 2 h of incubation in various lysine concen-trations. The experiment followed a typical lysine starvation proto-col, with the following exception. After log-phase cells were washedfree of lysine and suspended in minimal medium, a sample wasremoved to assay for CAT. These cells were then distributed amongseven flasks. Lysine was added to six of the flasks to a finalconcentration of 2, 5, 10, 20, 50, or 100 ,ug/ml. After 2 h ofincubation at 37°C, the cells were harvested and the specificactivities of CAT were determined. The CAT specific activity shownin the figure represents the increase in CAT after 2 h of incubation.
9 I IV I
I I It ij
I -41
VOL. 169, 1987

4240 DUVALL ET AL.
systems, the nucleotide sequence of cat leaders predicts thatonly a single stem-loop structure is possible in the mRNA,and this sequesters the RBS for the cat coding sequence.Thus, although the principles of cat and erm regulationappear similar, the mechanism of destabilization of cat RNAstem-loop structures by ribosomes may differ somewhatfrom that proposed for erm regulation. Direct in vivo evi-dence that destabilization of the cat-86 RNA stem-loopstructure occurs during chloramphenicol induction is lack-ing. However, we have shown that the cat-86 RNA stem-loop structure does serve as a site of partial transcriptiontermination which is relieved, albeit weakly, during induc-tion (3, 5). Hence, we presume that induction is concomitantwith destabilization of the RNA secondary structure.
Induction of cat genes by chloramphenicol likely takesadvantage of the molecular mechanism through which chlor-amphenicol blocks protein synthesis in bacteria (36). Webelieve that identification of the stall site in leader 2 maydemonstrate those sequences in mRNA which most effec-tively allow chloramphenicol to block peptide elongation.
ACKNOWLEDGMENTS
Un Jin Kim, Skorn Mongkolsuk, and Keith McKenny are thankedfor advice on site-directed mutagenesis.
This investigation was supported by Public Health Service grantAI-21350 from the National Institutes of Health.
LITERATURE CITED
1. Ambulos, N. P., Jr., J. H. Chow, S. Mongkolsuk, L. H. Preis,W. R. Volimar II, and P. S. Lovett. 1984. Constitutive variantsof the pC194 cat gene exhibit DNA alterations in the vicinity ofthe ribosome binding site sequence. Gene 28:171-176.
2. Ambulos, N. P., Jr., E. J. Duvall, and P. S. Lovett. 1986.Analysis of the regulatory sequences needed for induction of thechloramphenicol acetyltransferase gene cat-86 by chloramphen-icol and amicetin. J. Bacteriol. 167:842-849.
3. Ambulos, N. P., Jr., E. J. Duvall, and P. S. Lovett. 1987. ThemRNA for an inducible chloramphenicol acetyltransferase geneis cleaved into discrete fragments in Bacillus subtilis. J. Bacte-riol. 169:967-972.
4. Ambulos, N. P., Jr., S. Mongkolsuk, J. D. Kaufman, and P. S.Lovett. 1985. Chloramphenicol-induced translation of cat-86mRNA requires two cis-acting regulatory regions. J. Bacteriol.164:696-703.
5. Ambulos, N. P., Jr., S. Mongkolsuk, and P. S. Lovett. 1985. Atranscription termination signal immediately precedes the cod-ing sequence for the chloramphenicol-inducible gene cat-86.Mol. Gen. Genet. 199:70-75.
6. Bradford, M. M. 1976. A rapid and sehsitive method forquantitation of microgram quantities of protein ttilizing theprinciple of protein-dye binding. Anal. Biochem. 72:248-252.
6a.Bruckner, R., T. Dick, and H. Matzura. 1987. Dependence ofexpression of an inducible Staphylococcus aureus cat gene onthe translation of its leader sequence. Mol. Gen. Genet. 207:486-491.
7. Bruckner, R., and H. Matzura. 1985. Regulation of the induciblechloramphenicol acetyltransferase gene of the Staphylococcusaureus plasmid pUB112. EMBO J. 4:2295-2300.
8. Byeon, W.-H., and B. Weisblum. 1984. Post-transcriptionalregulation of chloramphenicol acetyltransferase. J. Bacteriol.188:543-550.
9. Dubngu, D. 1984. Translational attenuation: the regulation ofbacterial resistance to the macrolide-lincosamide-streptograminB antibiotics. Crit. Rev. Biochem. 16:103-132.
10. Dubnau, D. 1985. Induction of ermC requires translation of theleader peptide. EMBO J. 4:533-537.
11. Duvall, E. J., and P. S. Lovett. 1986. Chloramphenicol induces
translation of the mRNA for a chloramphenicol-resistance genein Bacillus subtilis. Proc. Natl. Acad. Sci. USA 83:3939-3943.
12. Duvall, E. J., S. Mongkolsuk, U. J. Kim, P. S. Lovett, T. M.Henkin, and G. H. Chambliss. 1985. Induction of the chloram-phenicol acetyltransferase gene cat-86 through the action of theribosomal antibiotic amicetin: involvement of a Bacillus subtilisribosomal component in cat induction. J. Bacteriol. 161:665-672.
13. Duvall, E. J., D. M. Williams, P. S. Lovett, C. Rudolph, N.Vasantha, and M. Guyer. 1983. Chloramphenicol inducible geneexpression in Bacillus subtilis. Gene 24:171-177.
14. Duvall, E. J., D. M. Williams, S. Mongkolsuk, and P. S. Lovett.1984. Regulatory regions that control expression of two chlor-amphenicol-inducible cat genes cloned in Bacillus subtilis. J.Bacteriol. 158:784-790.
15. Gryczan, T. J., S. Contente, and D. Dubnau. 1978. Character-ization of Staphylococcus aureus plasmids introduced by trans-formation into Bacillus subtilis. J. Bacteriol. 134:318-323.
16. Horinouchi, H., and B. Weisblum. 1982. Nucleotide sequenceand functional map of pC194, a plasmid that specifies induciblechloramphenicol resistance. J. Bacteriol. 150:815-825.
17. Keggins, K. M., P. S. Lovett, and E. J. Duvall. 1978. Molecularcloning of genetically active fragments of Bacillus DNA inBacillus subtilis and properties of the vector plasmid pUB110.Proc. Natl. Acad. Sci. USA 75:1423-1427.
18. Kolter, R., and C. Yanofsky. 1982. Attenuation in amino acidbiosynthetic operons. Annu. Rev. Genet. 16:113-134.
19. Lovett, P. S. 1985. Antibiotic-inducible regulation of a plasmidgene encoding chloramphenicol acetyltransferase in Bacillussubtilis, p. 397-400. In L. Leive, P. F. Bonventre, J. A. Morel-lo, S. Schlesinger, S. D. Silver, and H. C. Wu (ed.), Micro-biology-1985. American Society for Microbiology, Washing-ton, D.C.
20. Lovett, P. S., and K. M. Keggins. 1979. Bacillus subtilis as a hostfor molecular cloning. Methods Enzymol. 68:342-357.
21. Mayford, M., and B. Weisblum. 1985. Messenger RNA fromStaphylococcus aureus that specifies macrolide-lincosamide-streptogramin resistance. Demonstration of its conformationsand of the leader it encodes. J. Mol. Biol. 185:769-780.
22. Mongkolsuk, S., N. P. Ambulos, Jr., and P. S. Lovett. 1984.Chloramphenicol-inducible gene expression in Bacillus subtilisis independent of the chloramphenicol acetyltransferase struc-tural gene and its promoter. J. Bacteriol. 160:1-8.
23. Mongkolsuk, S., Y.-W. Chiang, R. B. Reynolds, and P. S.Lovett. 1983. Restriction fragments that exert promoter activityduring postexponential growth of Bacillus subtilis. J. Bacteriol.155:1399-1406.
24. Mongkolsuk, S., E. J. Duvall, and P. S. Lovett. 1985. Transcrip-tion termination signal for the cat-86 indicator gene in a Bacillussubtilis promoter cloning plasmid. Gene 37:83-90.
25. Narayanan, C. S., and D. Dubnau. 1985. Evidence for thetranslational attenuation model: ribosome binding studies andstructural analysis with an in vitro run off transcript of ermC.Nucleic Acids Res. 20:7307-7328.
26. Narayanan, C., and D. Dubnau. 1987. Demonstration of eryth-romycin-dependent stalling of ribosomes on the ermC leadertranscript. J. Biol. Chem. 262:1766-1771.
27. Nicholson, W. L., G. H. Chambliss, L. Buckbinder, N. P.Ambulos, Jr., and P. S. Lovett. 1985. Isolation and expression ofa constitutive variant of the chloramphenicol-inducible plasmidgene cat-86 under control of the Bacillus subtilis 168 amylasepromoter. Gene 35:113-120.
28. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequenc-ing with chain terminating inhibitors. Proc. Natl. Acad. Sci.USA 74:5463-5467.
29. Shaw, W. V. 1967. The enzymatic acetylation of chloramphen-icol by extracts of R factor resistant Escherichia coli. J. Biol.Chem. 242:687-693.
30. Shaw, W. V. 1975. Chloramphenicol acetyltransferase fromchloramphenicol-resistant bacteria. Methods Enzymol. 43:737-755.
31. Shaw, W. V. 1983. Chloramphenicol acetyltransferase: enzy-mology and molecular biology. Crit. Rev. Biochem. 14:1-43.
J. BACTERIOL.

VOL. 169, 1987 cat-86 INDUCTION BY AMINO ACID STARVATION
32. Shaw, W. V., D. G. Brenner, S. F. J. LeGrice, S. E. Skinner, andA. R. Hawkins. 1985. Chloramphenicol acetyltransferase geneof staphylococcal plasmid pC221. FEBS Lett. 179:101-106.
33. Smith, I. 1982. The translational apparatus of Bacillus subtilis,p. 111-145. In D. Dubnau (ed.), The molecular biology of thebacilli, vol. 1. Academic Press, Inc., New York.
34. Spizizen, J. 1958. Transformation of biochemically deficientstrains of Bacillus subtilis by deoxyribonucleate. Proc. Natl.Acad. Sci. USA 44:1072-1078.
35. Taylor, J. W., J. Ott, and F. Eckstein. 1985. The generation ofoligonucleotide-directed mutations at high-frequency using phos-phorothioate-modified DNA. Nucleic Acids Res. 13:8765-8785.
36. Vazquez, D. 1979. Inhibitors of protein synthesis. Mol. Biol.Biochem. Biophys. 30:108-112.
37. Weisblum, B. 1983. Inducible resistance to macrolides, linco-samides, and streptogramin B type antibiotics: the resistancephenotype, its biological diversity, and structural elements thatregulate expression, p. 91-121. In J. Beckwith, J. Davis, andJ. A. Gallent (ed.), Gene function in procaryotes. Cold SpringHarbor Laboratory, Cold Spring Harbor, N.Y.
38. Williams, D. M., E. J. Duvall, and P. S. Lovett. 1981. Cloningrestriction fragments that promote expression of a gene inBacillus subtilis. J. Bacteriol. 146:1162-1165.
39. Yansura, D. G., and D. J. Henner. 1984. Use of the Escherichiacoli lac repressor and operator to control gene expression inBacillus subtilis. Proc. Natl. Acad. Sci. USA 81:439-443.
40. Zoller, M. J., and M. Smith. 1983. Oligonucleotide-directedmutagenesis of DNA fragments cloned into M13 vectors. Meth-ods Enzymol. 100:468-500.
4241


















