
Polypeptide components of Drosophila small nuclear ......Polypeptide components of Drosophila small...
6
© 1991 Oxford University Press Nucleic Acids Research, Vol. 19, No. 21 5877-5882 Polypeptide components of Drosophila small nuclear ribonucleoprotein particles Trevor Paterson, Jean D.Beggs, David J.Finnegan and Reinhard Luhrmann 1 Institute of Cell and Molecular Biology, University of Edinburgh, Mayfield Road, Edinburgh EH9 3JR UK and 1 lnstitut fur Molekularbiologie und Tumorforschung, Klinicum der Philipps-Universitat Marburg, Emil-Mannkopff-Strasse 2, D-3550 Marburg, FRG Received August 20, 1991; Accepted October 14, 1991 ABSTRACT In eukaryotes splicing of pre-mRNAs is mediated by the spliceosome, a dynamic complex of small nuclear ribonucleoprotein particles (snRNPs) that associate transiently during spliceosome assembly and the splicing reaction. We have purified snRNPs from nuclear extracts of Drosophila cells by affinity chromatography with an antibody specific for the trimethylguanosine (m 3 G) cap structure of snRNAs U1-U5. The polypeptide components of Drosophila snRNPs have been characterized and shown to consist of a number of proteins shared by all the snRNPs, and some proteins which appear to be specific to individual snRNP particles. On the basis of their apparent molecular weight and antigenicity many of these common and particle specific Drosophila snRNP proteins are remarkably conserved between Drosophila and human spliceosomes. By probing western blots of the Drosophila snRNP polypeptides with a number of antisera raised against human snRNP proteins, Drosophila polypeptides equivalent to many of the HeLa snRNP-common proteins have been identified, as well as candidates for a number of U1, U2 and U5-specific proteins. INTRODUCTION In eukaryotes the excision of intervening sequences from pre- mRNA is catalyzed in a large ribonucleoprotein complex, the spliceosome. The spliceosome is composed of at least 30 distinct proteins and the five major small nuclear RNA (snRNA) molecules Ul, U2, U4, U5 and U6. The U snRNAs can be recovered from nuclear extracts as four individual small nuclear ribonucleoprotein (snRNP) complexes, the monomeric Ul, U2 and U5 snRNP particles and the dimeric U4/U6 snRNP in which the U4 and U6 snRNAs are found base paired (1,2). The protein composition of the snRNP complexes has been studied in detail in human and rodent nuclear extracts (3,4). Eight distinct polypeptides, B, B', Dl, D2, D3, E, F and G, with apparent molecular weights of between 9 and 29 (M r xl0~ 3 ), are common to all four snRNPs and form a common snRNP core. In addition, individual snRNPs have been shown to contain characteristic particle-specific proteins, denoted 70K, A and C for Ul; A' and B" for U2; and 15, 40, 52, 100, 102, 116 and 200K for U5. The U5 snRNP has a particularly complex protein composition and sediments through glycerol gradients more rapidly than the other snRNPs (5). An even larger tripartite U4/U5/U6 complex can be isolated under low salt conditions and contains at least four further polypeptides (4,6 — 8). Antisera from patients with a variety of connective tissue diseases recognize many of the snRNP polypeptides. Anti-RNP autoantibodies, from patients with mixed connective tissue disease (MCTD), react with the Ul-specific proteins A, C, and 70K. Anti-Sm sera from patients with systemic lupus erythematosus (SLE) immunoreact with the core (Sm) proteins B', B and D (9,10), with variable reactivity to Dl, D2 and D3 (11), and less frequently with E, F and G (12). These sera have been used to study the composition of snRNPs from a variety of species by immunoprecipitation and immunoblotting. These studies indicate that the protein composition of snRNPs is remarkably conserved throughout eukaryotes (reviewed 3) as indeed are the sequences of the U snRNAs (reviewed 13). Purification of snRNPs has been facilitated by the use of the human autoantibodies, but more successfully by the use of mono and polyclonal antisera raised against modified nucleosides peculiar to the snRNAs. In particular antibodies which recognize the highly methylated nucleoside 2,2,7-trimethylguanosine (m 3 G), which is found as a 5'-cap on Ul, U2, U4 and U5 snRNAs, allow a single step purification of m 3 G-capped snRNPs from nuclear extracts by immunoaffinity chromatography. Native snRNPs can be recovered by desorption with an excess of m 3 G (or the cross-reactive nucleoside 7-methylguanosine, m 7 G) or by elution with salt (14-17). The Drosophila U snRNAs closely resemble those of vertebrates in size, structure and nucleotide sequence (18, reviewed 13). Anti-Sm and anti-RNP antisera have identified a number of low molecular weight potential snRNP polypeptides (19,20). In this study we have used a monoclonal antibody which recognizes the 5'-nucleotide cap of snRNAs (16) to immunoaffinity purify snRNPs from nuclear extracts of Drosophila cells in order to identify their protein components and to compare them to those of HeLa cells.
Transcript of Polypeptide components of Drosophila small nuclear ......Polypeptide components of Drosophila small...

© 1991 Oxford University Press Nucleic Acids Research, Vol. 19, No. 21 5877-5882
Polypeptide components of Drosophila small nuclearribonucleoprotein particles
Trevor Paterson, Jean D.Beggs, David J.Finnegan and Reinhard Luhrmann1
Institute of Cell and Molecular Biology, University of Edinburgh, Mayfield Road, Edinburgh EH9 3JRUK and 1lnstitut fur Molekularbiologie und Tumorforschung, Klinicum der Philipps-Universitat Marburg,Emil-Mannkopff-Strasse 2, D-3550 Marburg, FRG
Received August 20, 1991; Accepted October 14, 1991
ABSTRACT
In eukaryotes splicing of pre-mRNAs is mediated by thespliceosome, a dynamic complex of small nuclearribonucleoprotein particles (snRNPs) that associatetransiently during spliceosome assembly and thesplicing reaction. We have purified snRNPs fromnuclear extracts of Drosophila cells by affinitychromatography with an antibody specific for thetrimethylguanosine (m3G) cap structure of snRNAsU1-U5. The polypeptide components of DrosophilasnRNPs have been characterized and shown to consistof a number of proteins shared by all the snRNPs, andsome proteins which appear to be specific to individualsnRNP particles. On the basis of their apparentmolecular weight and antigenicity many of thesecommon and particle specific Drosophila snRNPproteins are remarkably conserved between Drosophilaand human spliceosomes. By probing western blots ofthe Drosophila snRNP polypeptides with a number ofantisera raised against human snRNP proteins,Drosophila polypeptides equivalent to many of theHeLa snRNP-common proteins have been identified, aswell as candidates for a number of U1, U2 andU5-specific proteins.
INTRODUCTION
In eukaryotes the excision of intervening sequences from pre-mRNA is catalyzed in a large ribonucleoprotein complex, thespliceosome. The spliceosome is composed of at least 30 distinctproteins and the five major small nuclear RNA (snRNA)molecules Ul, U2, U4, U5 and U6. The U snRNAs can berecovered from nuclear extracts as four individual small nuclearribonucleoprotein (snRNP) complexes, the monomeric Ul, U2and U5 snRNP particles and the dimeric U4/U6 snRNP in whichthe U4 and U6 snRNAs are found base paired (1,2).
The protein composition of the snRNP complexes has beenstudied in detail in human and rodent nuclear extracts (3,4). Eightdistinct polypeptides, B, B', Dl, D2, D3, E, F and G, withapparent molecular weights of between 9 and 29 (M rxl0~3),are common to all four snRNPs and form a common snRNP core.
In addition, individual snRNPs have been shown to containcharacteristic particle-specific proteins, denoted 70K, A and Cfor Ul; A' and B" for U2; and 15, 40, 52, 100, 102, 116 and200K for U5. The U5 snRNP has a particularly complex proteincomposition and sediments through glycerol gradients morerapidly than the other snRNPs (5). An even larger tripartiteU4/U5/U6 complex can be isolated under low salt conditions andcontains at least four further polypeptides (4,6 — 8).
Antisera from patients with a variety of connective tissuediseases recognize many of the snRNP polypeptides. Anti-RNPautoantibodies, from patients with mixed connective tissue disease(MCTD), react with the Ul-specific proteins A, C, and 70K.Anti-Sm sera from patients with systemic lupus erythematosus(SLE) immunoreact with the core (Sm) proteins B', B and D(9,10), with variable reactivity to Dl, D2 and D3 (11), and lessfrequently with E, F and G (12). These sera have been used tostudy the composition of snRNPs from a variety of species byimmunoprecipitation and immunoblotting. These studies indicatethat the protein composition of snRNPs is remarkably conservedthroughout eukaryotes (reviewed 3) as indeed are the sequencesof the U snRNAs (reviewed 13).
Purification of snRNPs has been facilitated by the use of thehuman autoantibodies, but more successfully by the use of monoand polyclonal antisera raised against modified nucleosidespeculiar to the snRNAs. In particular antibodies which recognizethe highly methylated nucleoside 2,2,7-trimethylguanosine(m3G), which is found as a 5'-cap on Ul, U2, U4 and U5snRNAs, allow a single step purification of m3G-cappedsnRNPs from nuclear extracts by immunoaffinitychromatography. Native snRNPs can be recovered by desorptionwith an excess of m3G (or the cross-reactive nucleoside7-methylguanosine, m7G) or by elution with salt (14-17).
The Drosophila U snRNAs closely resemble those ofvertebrates in size, structure and nucleotide sequence (18,reviewed 13). Anti-Sm and anti-RNP antisera have identified anumber of low molecular weight potential snRNP polypeptides(19,20). In this study we have used a monoclonal antibody whichrecognizes the 5'-nucleotide cap of snRNAs (16) toimmunoaffinity purify snRNPs from nuclear extracts ofDrosophila cells in order to identify their protein componentsand to compare them to those of HeLa cells.
5878 Nucleic Acids Research, Vol. 19, No. 21
MATERIALS AND METHODSCell growth and preparation of nuclear extractsThe Drosophila melanogaster Schneider 2 cell line waspropagated at 22 °C in suspension in Schneider's (modified)Drosophila Medium (purchased from Imperial Laboratories, UK)supplemented with 5 % foetal calf serum, lOOU/ml penicillin and100/tg/ml streptomycin. Nuclear extracts were prepared by themethod of Dignam et al., (21) without homogenization of cellsand with the addition of 0.5% v/v NP40 to disrupt the cytoplasmicmembrane. HeLa cell nuclear extracts and purified snRNPs werekindly provided by Silke Borner.
Anti-m3G immunoaffinity chromatography of snRNPsAffinity purification of m3G-capped snRNPs from nuclearextracts was performed by using mAb H-20 bound covalentlyto CNBr-activated Sepharose 4B essentially as described (16).Nuclear extract prepared as above was passed over a 5mlH-20-Sepharose column and bound RNA-protein complexes weredesorbed with 20mM m7G.
70K-
A —B -D —
E U W F L
-5S
-US
Anti-seraRabbit: anti-FP8.1 raised against a portion of yeast U5 snRNPspecific protein PRP8 fused to j3-galactosidase and affinity-purified against the same portion of PRP8 fused to the trpEprotein (22).
Monoclonal antibodies: Y12 anti-Sm type (9); 7.13 anti-HeLaDl (23); H-20 anti-m3G (16); 4G3 anti-HeLa U2B' (24); H i l lanti-HeLa Ul 70K, H304 anti-HeLa Ul A and H386 anti-HeLaUl 70K and U5 100K (8,25).
Patient 7, E74 and Plasma D anti-Sm sera from patients withsystemic lupus erythematosus were kindly provided by Prof.H.Peter, Freiburg.
Alkaline phosphatase-conjugated secondary antibodies (againstrabbit, human or mouse IgG or mouse IgM antibodies) werepurchased from Promega and Bio-Rad.
RNA and protein analysisSamples were extracted with 1 volume of PC A (phenol,chloroform, isoamylalcohol 50:50:1) with 0.5% (w/v) SDS.Following phase separation by centrifugation RNAs wererecovered from the aqueous phase by ethanol precipitation.Proteins were precipitated from the organic phase with 5 volumesof acetone.
RNAs were fractionated by electrophoresis through 10%polyacrylamide gels containing 7M urea, lOmM Tris-BoratepH8.2 and 2mM EDTA (TBE). RNA was visualized by stainingwith ethidium bromide (0.5/tg/ml) or with silver (26). RNA wastransferred to nylon membrane (Amersham) by electrophoresisin 0.5 XTBE for 3h at 45V and fixed by UV irradiation. UsnRNA-specific oligonucleotides were 5'-end labelled usingpolynucleotide kinase and hybridized to pre-blocked filters in6XSSC (3M NaCl, 3M trisodium citrate), 5xDenhardt'ssolution, 100/tg/ml denatured salmon sperm DNA for 2h at 4°Cand washed briefly in 6xSSC at 20°C before autoradiography.Oligonucleotides were synthesized by OSWEL DNA Service,Edinburgh University, and were directed against: Drosophila U5
H D D
200 =116-
100/2=
7 0 -
A -
- 2 0 5-116- 97
- 6 6
-US
-31
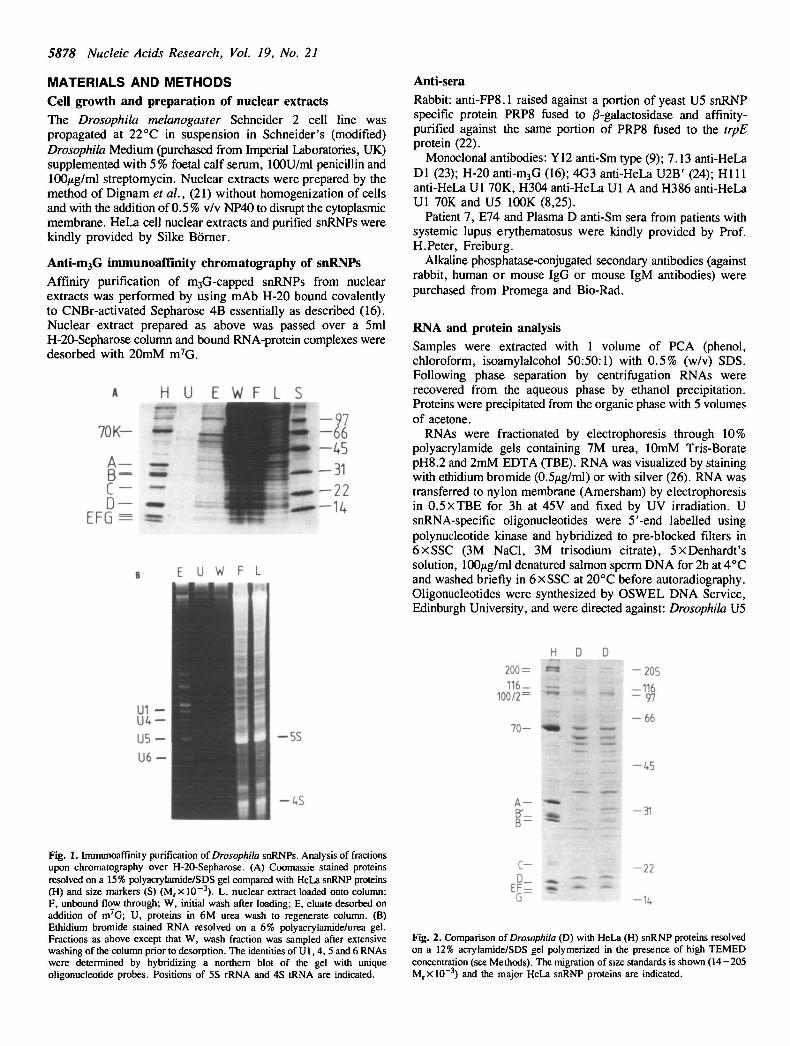
Fig. 1. Immunoaffinity purification of Drosophila snRNPs. Analysis of fractionsupon chromatography over H-20-Sepharose. (A) Coomassie stained proteinsresolved on a 15% polyacrylamide/SDS gel compared with HeLa snRNP proteins(H) and size markers (S) (M rx 10~3). L, nuclear extract loaded onto column;F, unbound flow through; W, initial wash after loading; E, eluate desorbed onaddition of m7G; U, proteins in 6M urea wash to regenerate column. (B)Ethidium bromide stained RNA resolved on a 6% polyacrylamide/urea gel.Fractions as above except that W, wash fraction was sampled after extensivewashing of the column prior to desorption. The identities of U1, 4, 5 and 6 RNAswere determined by hybridizing a northern blot of the gel with uniqueoligonucleotide probes. Positions of 5S rRNA and 4S tRNA are indicated.
c- -22
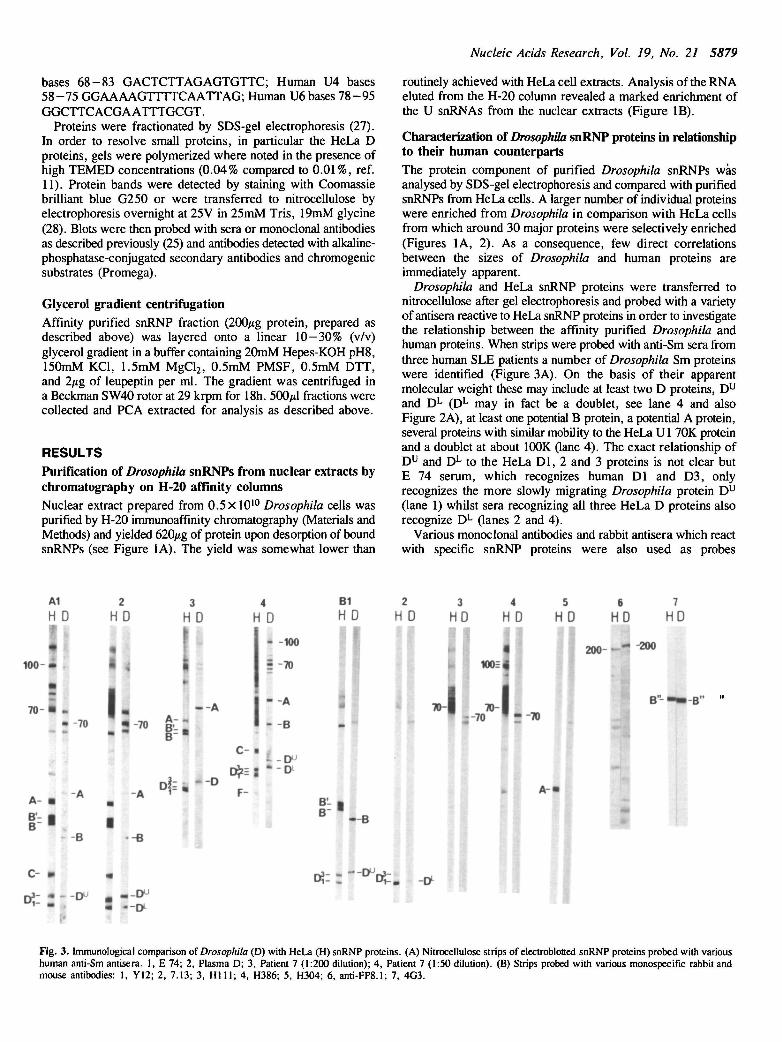
Fig. 2. Comparison of Drosophila (D) with HeLa (H) snRNP proteins resolvedon a 12% acrylamide/SDS gel polymerized in the presence of high TEMEDconcentration (see Methods). The migration of size standards is shown (14-205M rxl0" 3 ) and the major HeLa snRNP proteins are indicated.
Nucleic Acids Research, Vol. 19, No. 21 5879
bases 68-83 GACTCTTAGAGTGTTC; Human U4 bases58-75 GGAAAAGTTTTCAATTAG; Human U6 bases 78-95GGCTTCACGAATTTGCGT.
Proteins were fractionated by SDS-gel electrophoresis (27).In order to resolve small proteins, in particular the HeLa Dproteins, gels were polymerized where noted in the presence ofhigh TEMED concentrations (0.04% compared to 0.01%, ref.11). Protein bands were detected by staining with Coomassiebrilliant blue G250 or were transferred to nitrocellulose byelectrophoresis overnight at 25V in 25mM Tris, 19mM glycine(28). Blots were then probed with sera or monoclonal antibodiesas described previously (25) and antibodies detected with alkaline-phosphatase-conjugated secondary antibodies and chromogenicsubstrates (Promega).
Glycerol gradient centrifugationAffinity purified snRNP fraction (200/tg protein, prepared asdescribed above) was layered onto a linear 10—30% (v/v)glycerol gradient in a buffer containing 20mM Hepes-KOH pH8,150mM KC1, 1.5mM MgCl2, 0.5mM PMSF, 0.5mM DTT,and 2/ig of leupeptin per ml. The gradient was centrifuged ina Beckman SW40 rotor at 29 krpm for 18h. 500/il fractions werecollected and PCA extracted for analysis as described above.
RESULTSPurification of Drosophila snRNPs from nuclear extracts bychromatography on H-20 affinity columnsNuclear extract prepared from 0.5 x 1010 Drosophila cells waspurified by H-20 immunoaffinity chromatography (Materials andMethods) and yielded 620/ig of protein upon desorption of boundsnRNPs (see Figure 1A). The yield was somewhat lower than
routinely achieved with HeLa cell extracts. Analysis of the RNAeluted from the H-20 column revealed a marked enrichment ofthe U snRNAs from the nuclear extracts (Figure IB).
Characterization of Drosophila snRNP proteins in relationshipto their human counterpartsThe protein component of purified Drosophila snRNPs wasanalysed by SDS-gel electrophoresis and compared with purifiedsnRNPs from HeLa cells. A larger number of individual proteinswere enriched from Drosophila in comparison with HeLa cellsfrom which around 30 major proteins were selectively enriched(Figures 1A, 2). As a consequence, few direct correlationsbetween the sizes of Drosophila and human proteins areimmediately apparent.
Drosophila and HeLa snRNP proteins were transferred tonitrocellulose after gel electrophoresis and probed with a varietyof antisera reactive to HeLa snRNP proteins in order to investigatethe relationship between the affinity purified Drosophila andhuman proteins. When strips were probed with anti-Sm sera fromthree human SLE patients a number of Drosophila Sm proteinswere identified (Figure 3A). On the basis of their apparentmolecular weight these may include at least two D proteins, Du
and DL (DL may in fact be a doublet, see lane 4 and alsoFigure 2A), at least one potential B protein, a potential A protein,several proteins with similar mobility to the HeLa Ul 70K proteinand a doublet at about 100K (lane 4). The exact relationship ofD u and DL to the HeLa Dl, 2 and 3 proteins is not clear butE 74 serum, which recognizes human Dl and D3, onlyrecognizes the more slowly migrating Drosophila protein D u
(lane 1) whilst sera recognizing all three HeLa D proteins alsorecognize DL (lanes 2 and 4).
Various monoclonal antibodies and rabbit antisera which reactwith specific snRNP proteins were also used as probes
A1
H D
I'100- •
7 0 - •
A-B-
C- m
2HD
2 ,I.
3HD
4H D
- A
• -70 JJ -70
1 -' -:'•'
-A -A: •
i •-B -B
A -
- -100
= -70
- A
- B
B1HD
II- - D
i
C- •
F-B'-
- B
ii
2H D
3
HD4
HD5
HD
!f I1U 100=|
I200
6HD
_ _ - -200
7
HD
- 7 0 ^ - ^
i-B"
1
II
Fig. 3. Immunological comparison of Drosophila (D) with HeLa (H) snRNP proteins. (A) Nitrocellulose strips of electroblotted snRNP proteins probed with varioushuman anti-Sm antisera. 1, E 74; 2, Plasma D; 3, Patient 7 (1:200 dilution); 4, Patient 7 (1:50 dilution). (B) Strips probed with various monospecific rabbit andmouse antibodies: 1, Y12; 2, 7.13; 3, H i l l ; 4, H386; 5, H304; 6, anti-FP8.1; 7, 4G3.

5880 Nucleic Acids Research, Vol. 19, No. 21
(Figure 3B). These antibodies confirmed the existence of at leasttwo distinct D proteins D u and DL (lanes 1 and 2). Again therelationship of D u and DL to Dl , D2 and D3 could not beunambiguously assigned since in this case antibodies raisedagainst Dl (7.13) recognized DL (lane 2). The mobility of theD proteins was clearly affected by the composition of theacrylamide gel used (Materials and Methods, 11). In a 12%acrylamide gel polymerized with high concentrations of TEMED,D u and DL ran with similar mobility to HeLa D proteins(Figure 3A lanes 1,2; Figure 3B lanes 1,2), but in a 14%
Table 1. Comparison of human and Drosophila snRNP polypeptides
HELA snRNP Proteins Equivalent Drosophila proteinsDesignation Apparent Designation Apparent Comments(a) molecular molecular
weight(a,b) weight
Common proteinsB'BD3D2DlEFG
Ul snRNP70KAC
U2 snRNPB'
U5 snRNP200(+)K116K102K100K52K40K
29281816.51612119
proteins70(62)34
asproteins
28.5
proteins>2001161021005240
Bu
BL
0 u&
EFG
70KA
cB"
2OOK/PRP8120K105K
54K42K
282718.517
12.5119
563814.5
28
240120105
5442
may be doublet
others of 54 & 61?
may be doublet
(a) From Luhrmann et al.,(4). (b) M r x l 0 - 3
acrylamide, low TEMED gel the Drosophila D proteins migratedmore slowly than the HeLa counterparts (Figure 3A lanes 3,4).
As was observed with the Human Sm antisera only a singleDrosophila B protein was detected by the anti-Sm monoclonalantibody Y12 (lane 1). The antibodies Hi l l and H386 (lanes3,4) define a unique Drosophila band related to the HeLa Ul70K protein whereas the human anti-Sm sera (Figure 3A)indicated that there may be multiple post-translationally modifiedvariants of this protein as has been observed in HeLa cells.Antiserum raised against a portion of the yeast U5 snRNP specificprotein PRP8 (22) recognized large 200-260kDa proteins in bothHeLa and Drosophila snRNPs (lane 6) as previously reported(29—31). Antisera raised against other regions of the yeast protein(22) also recognized both the HeLa and Drosophila protein (30,results not shown). A monoclonal (4G3) raised against the HeLaU2-specific B' protein (24) recognized a similarly sizedDrosophila protein (lane 7).
A monoclonal antibody specific to the HeLa A protein (H304,lane 5) and rabbit antipeptide antisera to the HeLa C and Gproteins (results not shown) failed to recognize equivalentDrosophila proteins, perhaps reflecting lack of conservation ofthese epitopes between Drosophila and humans. Results fromthese and the later analyses are summarized in Table 1.
Resolution of the Drosophila U5 snRNP particle bysedimentation through glycerol gradientsThe immunoaffinity purified Drosophila snRNP fraction wassubjected to sedimentation analysis in a linear 10-30% (v/v)glycerol gradient in order to investigate whether rapidly andslowly sedimenting particles could be resolved as has beendemonstrated for HeLa snRNP particles (5). When the proteincontent of the glycerol gradient fractions was analysed by gelelectrophoresis a pattern remarkably similar to that observed withHeLa extracts was found (Figure 4A). The snRNP particlesappear to have sedimented in two different size ranges; the rapidlysedimenting particles (fractions 14 — 17) appear to correspond tothe 20S U5 snRNPs of HeLa (5) whilst the more slowlysedimenting particles (fractions 4-10) resemble in composition
L i 3 4 i • 7 S S « 1 1 1 2 Q W K T 7 « ? 1 2 3 2 5 2 7
I
- 2 0 5-116
- 6 6
- 4 5
-31 U1
-22-14
U5H
U6 JL4
m
16
• • •
• i8 1012
•
i14
•
m16
•
»4<1820 22
c200-
100-
70-
A -
C -
D =
EF=G -
H L 4-10 14-17
Fig. 4. Sedimentation of Drosophila snRNP particles in a linear 10-30% glycerol gradient. (A) Protein fractions 1 (top) to 27 (bottom of gradient) resolved ona 12% acrylamide/SDS gel polymerized in the presence of high TEMED concentration. L, sample of snRNP fraction loaded onto gradient. (B) RNA prepared fromglycerol gradient fractions resolved on a 6% acrylamide/urea gel, transferred to nylon membrane and independently probed with Ul, 5 and 6 specific oligonucleotideprobes. (C) Composition of rapidly sedimenting particles (fractions 14—17) compared to slowly sedimenting particles (fractions 4-10) and total HeLa (H) and Drosophila(L) snRNP proteins. The major HeLa proteins are indicated (G-200) and proteins discussed in the text are highlighted.

Nucleic Acids Research, Vol. 19, No. 21 5881
the smaller snRNPs, including the Ul snRNP, with a value ofaround 10S.
When the RNA present in the gradient fractions was analysedby urea/acrylamide electrophoresis and northern blot a lessdistinct pattern of separation was found. Whilst a peak of U5snRNA indeed co-sedimented with the 20S particles (fractions14 and 16, Figure 4B) all of the U snRNAs examined appearedto smear throughout the gradient, perhaps due to the loss ofproteins from the snRNPs. However, the Ul and U6 snRNAswere more concentrated further up the gradient than U5 (fractions6-12, Figure 4B). The protein composition of the two resolvedparticle types is shown in greater detail in Figure 4C. Commoncore proteins present in both particle types include two candidateB proteins, with apparent molecular weights marginally smallerthan HeLa B and B' (the faster migrating species, BL,corresponds to the B protein recognized by anti-Sm sera;Figure 3A lanes 1,2 and 4); the two D proteins Du and DL
identified immunologically above, and two proteins with similarsizes to the HeLa core proteins E and F. Whilst a protein ofsimilar size to Hela G is present in the total Drosophila snRNPfraction (arrowed in lane L), this band is not discernible in eitherof the glycerol gradient fractions.
The more slowly migrating complexes (fractions 4-10) appearto contain the Ul snRNP specific proteins A and 70K. Againa number of proteins with apparent size of about 70 000 can bedistinguished, perhaps indicating that multiple post-translationallymodified variants of this protein are found in Drosophila as inHeLa. Alternatively this protein could have suffered proteolyticdegradation during purification. Of the three candidate 70Kproteins in Drosophila with Mr 54 000, 56 000 and 61 000 itappears to be the 56 000 species which is recognized by theanti-70K monoclonal antibodies. Whilst there is no protein ofsimilar molecular weight to the HeLa C protein (Mr 22 000) aprominent polypeptide of Mr 14 500 may be the equivalentDrosophila protein. The Drosophila equivalent of the U2-specificprotein B' (Mr 28 000) was shown to be present in this fractionby western blotting and probing with the antibody 4G3 (resultnot shown). The Drosophila B' protein identified by western blotcomigrates with the larger of the two candidate B proteins, Bu ,which is present in both fractions of snRNPs in Figure 4C.
The rapidly sedimenting particles in fractions 14-17 resembleHeLa U5 snRNPs in protein composition. The high molecularweight 240-260K protein which is recognized by antibodiesraised against the yeast U5-specific protein PRP8 is present, asare at least two proteins of Mr 105-120 000 which mightcorrespond to the 100, 102 and 116K proteins in HeLa. Twoproteins of similar size to the HeLa 40K and 52K proteins alsoco-sediment with these Drosophila U5 snRNP particles.
Probing western blots of these glycerol gradient fractions withthe human anti-Sm antisera confirmed the presence of thecommon proteins BL, Du and DL in both fractions 4 -10 and14—17, whilst the Drosophila equivalents of the Ul-specific 70Kand A, and U2-specific B' proteins where restricted to fractions4 -10 , and the U5-specific 100 and 200K proteins to fractions14—17 (results not shown).
DISCUSSION
D. melanogaster provides an ideal model higher eukaryote inwhich to study the mechanism and regulation of pre-mRNAsplicing. An increasing number of genes have been describedwhere splicing of their transcripts is regulated in a developmental,
sex or tissue-specific manner (see references 32-37 forexamples) and a number of genes that regulate splicing have beenidentified, several of which encode proteins with RNA bindingmotifs resembling those found in mammalian snRNP proteins(38—41). Furthermore, nuclear extracts prepared from embryosor cultured Drosophila cells have been shown to accurately splicemRNA precursors by a similar mechanism to that observed inyeast and mammals (42).
In order to characterize the Drosophila splicing machinery wehave purified snRNP particles from cultured Drosophila cellextracts by immunoaffinity chromatography and analysed theirprotein composition. The polypeptide components of DrosophilasnRNPs appear remarkably similar to those characterized in HeLacells. On the basis of molecular weight, antigenicity and particledensity-sedimentation we have identified Drosophila proteinscorresponding to most of the characterized HeLa polypeptides(summarized in Table 1).
Eight proteins have been identified in HeLa extracts which arecommon to all snRNP particles: B, B', Dl, D2, D3, E, F andG. On the basis of molecular weight Drosophila has proteinscorresponding to most if not all of these, and at least three proteinsBL, D u and DL are immunologically related to the equivalentHeLa proteins. Indeed the Drosophila B and D proteins identifiedin this study probably correspond to proteins previously identifiedby cross-reactivity with human anti-Sm and anti-snRNP antiserawith reported molecular weights of 18 and 26 000 (19) and 14and 26 000 (20).
Proteins which are specific to U1, U2 and U5 snRNPs are alsoconserved between humans and Drosophila. A counterpart to theHeLa U5-specific 200K protein which cross reacts with antibodiesraised against the yeast PRP8 protein (29-31) is found inDrosophila U5-containing particles, as are at least two proteinsof about Mr 100-120 000 and other proteins potentiallycorresponding to the less well characterized 40 and 52K HeLaU5 proteins.
Drosophila proteins corresponding to the HeLa Ul snRNPspecific 70K and A proteins were recognizable on the basis oftheir size and antigenicity, but no protein of equivalent molecularweight to the HeLa C protein could be detected. An abundantand potentially Ul-specific protein of Mr 14 500 was, however,resolved by density centrifugation and may be the Drosophilacounterpart to C (Figures 4A,C).
A protein of similar size to the HeLa U2-specific B' proteinwas recognized by an antibody raised against the human protein(24). This antibody also recognizes a protein of Mr 27 000present in plant nuclear extracts and the gene encoding B' hasrecently been cloned from potato and shown to be highlyconserved between the animal and plant kingdoms (43). Acandidate gene coding for either the U2-specific A' or B' proteinshas recently been cloned from Drosophila (44).
As in HeLa extracts, a number of electrophoretically distinctspecies of around 70 000 were detected. The most abundant ofthese had an apparent molecular weight similar to that of the mostabundant HeLa 70K protein (61 -62 000 in this system, Figures2; 4A). However, the species recognized by monoclonalantibodies raised against the HeLa 70K protein had an apparentmolecular weight of 56—58 000 (Figure 3B). A further speciesof 52 -54 000 was also detected (Figure 2) and all three or morebands, perhaps representing modified or partially degradedversions of 70K, were recognized by some anti-Sm sera(Figure 3A). The gene encoding the Drosophila 70K protein hasbeen cloned and found to encode a protein of predicted Mr

5882 Nucleic Acids Research, Vol. 19, No. 21
53 000, the amino-terminal region of which is 68% identical tothe human and Xenopus 70K proteins (45). Whilst there is noevidence in Drosophila that alternative RNA processing pathwaysgive rise to 70K variants it is possible that some different formsof 70K may be generated by variable post translationalmodifications as has been reported for the mouse and human 70Kproteins (46).
Until now the biochemical purification of snRNP particles fortheir characterization has largely been restricted to rodent andhuman cell types. Studies on a disparate range of species,primarily using autoimmune antibodies, have identified proteinscorresponding to D, E, F and G in most systems (reviewed 3).This study shows that in Drosophila the conservation of theproteins involved in the mechanism of splicing extends muchfurther. The striking degree of similarity between the snRNPproteins found in Drosophila and humans is perhaps notunexpected, given the evolutionary constraints on maintainingan accurate and efficient splicing apparatus.
ACKNOWLEDGEMENTS
This work was funded by project grants from the UK MRC (DJF)the Deutsche Forschungsgemeinschaft (RL) and an SERCpostdoctoral research fellowship (TP). JDB is a Royal SocietyE.P.A. Cephalosporin Fund Senior Research Fellow.Collaboration was assisted by a travel grant from the Society forGeneral Microbiology. Antibodies were generously provided byJ.A. Steitz (Y12), S.O. Hoch (7.13), W.J. van Venrooij (4G3)and H. Peter (SLE). The western blot shown in Figure 3B lane7 was performed by Helen Kulesza. TP would like to thank allthe members of the Luhrmann laboratory, especially SilkeBorner, for their assistance and hospitality.
REFERENCES1. Steitz J.A, Black D.L., Gerke V., Parker K.A., Kramer A., Frendewey
D. and Keller W. (1988) In Birnsteil MX. (ed.) Structure and function ofmajor and minor small nuclear ribonucleoprotein particles. Springer-Verlag,Berlin, ppl l5-154.
2. Guthrie C. and Patterson B. (1988) Ann. Rev. Genet. 22, 387-419.3. Luhrmann R. (1988) In Birnsteil M.L. (ed.) Structure and function of major
and minor small nuclear ribonucleoprotein particles. Springer-Verlag, Berlin,pp71-99.
4. Luhrmann R., Kastner B. and Bach M. (1990) Biochem. Biophys. Acta 1087,265-292.
5. Bach M., Winkelmann G. and Luhrmann R. (1989) Proc. Natl. Acad. Sci.USA 86, 6038-6042.
6. Konarska M.M. and Sharpe P.A. (1987) Cell 49, 763-774.7. Cheng S.-C. and Abelson J. (1987) Genes Dev. 1, 1014- 1027.8. Behrens S.-E. and Luhrmann R. (1991) Genes Dev. (In Press).9. Lerner E.A., Lemer M.R., Janeway C.A. and Steitz J.A. (1981) Proc. Natl.
Acad. Sci. USA 78, 2737-2741.10. Habets W.J., Berden J.H.M., Hoch S.O. and van Venrooij W.J. (1985)
Eur. J. Immunol. 15, 992-997.11. Lehmeier T., Foulaki K. and Luhrmann R. (1990) Nucl. Acids Res. 18,
6475-6484.12. Reuter R., Rothe S., Habets W., van Venrooij W.J. and Luhrmann R. (1990)
Eur. J. Immunol. 20, 437-440.13. Reddy R. and Busch H. (1988) In Birnsteil M.L. (ed.) Structure and function
of major and minor small nuclear ribonucleoprotein particles. Springer-Verlag,Berlin, pp l -37 .
14. Bringmann P., Rinke J., Appel B., Reuter R. and Luhrmann R. (1983) TheEMBO J. 2, 1129-1135.
15. Bringmann P., Reuter R., Rinke J., Appel B., Bald R. and Luhrmann R.(1983) J. Biol. Chem. 258, 2745-2747.
16. Bochnig P., Reuter R., Bringmann P. and Luhrmann R. (1987) Eur. J.Biochem. 168, 461-467.
17. Krainer A.R. (1988) Nucl. Acids Res. 16, 9415-9429.
18. Myslinski E., Branlant C , Wieben E.D. and Pederson T. (1984) J. Mol.Biol. 180, 927-945.
19. Wooley J.C., Cone R.D., Tartof D. and Chung S.-Y. (1982) Proc. Natl.Acad. Sci. USA 79, 6762-6766.
20. Wieben E.D. and Pederson T. (1982) Mol. Cell. Biol 2, 914-920.21. Dignam J.D., Lebovitz R.M. and Roeder R.G. (1983) Nucl. Acids Res.
11, 1475-1489.22. Lossky M., Anderson G.J., Jackson S.P. and Beggs J.D. (1987) Cell 51,
1019-1026.23. Billings P.B., Barton J.R. and Hoch S.O. (1985) J. Immunol. 135,428-432.24. Habets W.J., Hoet M.H., De Jong B.A.W., Van Der Kemp A. and van
Venrooij W.J. (1989) J. Immunol. 143, 2560-2566.25. Reuter R. and Luhrmann R. (1986) Proc. Natl. Acad. Sci. USA 83,
8689-8693.26. Merril C.R., Goldman D., Sedman S.A. and Ebert M.H. (1981) Science
211, 1437-1438.27. Laemmli U.K. (1970) Nature 227, 680-685.28. Towbin H., Stachelin T. and Gordon J. (1979) Proc Natl. Acad. Sci. USA
76, 4350-4354.29. Anderson G.J., Buch M., Luhrmann. R. and Beggs J.D. (1989) Nature 342,
819-821.30. Anderson G.J. (1989) PhD Thesis, University of Edinburgh.31. Garcia-Blanco M.A., Anderson G.J., Beggs J.D. and Sharp P.A. (1990)
Proc. Natl. Acad. Sci. USA 87, 3082-3086.32. Baker B.S. (1989) Nature 340, 521-524.33. Bernstein S.I., Hansen C.J., Becker K.D., Wassenberg D.R., Roche E.S.,
Donady, J.J. and Emerson Jr. C.P. (1986) Mol. Cell. Biol. 6, 2511-2519.34. Falkenthal S., Parker V.P. and Davidson N. (1985) Proc. Natl. Acad. Sci.
USA 82, 449-453.35. Bingham, P.M., Chou T.-B., Mims I. and Zachar Z. (1988) Trends Genet.
4, 134-138.36. Laski F.A., Rio D.C. and Rubin G.M. (1986) Cell 44, 7 -19 .37. Chou T.-B., Zachar Z. and Bingham P.M. (1987) The EMBO J. 6,
4095-4104.38. Bell L.R., Maine E.M., Schedl P. and Cline T.W. (1988) Cell 55,
1037-1046.39. GoralskiT.J., Edstrom J.-E. and Baker B.S. (1989) Cell 56, 1011-1018.40. Zachar Z., Chou T.-B. and Bingham P.M. (1987) The EMBO J. 6,
4105-4111.41. Hedley M. and Maniatis T. (1991) Cell 65, 579-586.42. Rio D. (1988) Proc. Natl. Acad. Sci. USA 85, 2904-2908.43. Simpson G.G., Vaux P., Clark G., Waugh R., Beggs J.D. and Brown J.W.S.
(1991) Submitted.44. Harper D.S., Fresco L.D. and Keene J.D. (1990) Mol. Biol. Reports 14, 163.45.M ancebo R., Lo P.C.H. and Mount S.M. (1990). Mol. Cell. Biol. 10,
2492-2502.46. Woppman A., Patschinsky T., Bringmann P., Godt F. and Luhrmann R.
(1990) Nucl. Acids Res. 18, 4427-4438.


















