
Lecture 3 Specification of the second lineage, primitive endoderm and stem cell lines from early...
21
Lecture 3 Specification of the second lineage, primitive endoderm and stem cell lines from early mouse embryos ould understand anisms governing specification of the primitive endoderm lineage is an ES cell cell lines from early mouse embryos and their relationship to earl ages.
-
Upload
marco-mathers -
Category
Documents
-
view
218 -
download
2
Transcript of Lecture 3 Specification of the second lineage, primitive endoderm and stem cell lines from early...

Lecture 3
Specification of the second lineage, primitive endoderm and stem cell lines from early mouse embryos
You should understand
• Mechanisms governing specification of the primitive endoderm lineage• What is an ES cell• Stem cell lines from early mouse embryos and their relationship to early lineages.

1. Oct4/Pou5f1; uniformly expressed in cleavage stages. Switched off in trophectoderm of blastocyst. Knockout fails to develop ICM.
2. Cdx2; stochastically expressed from 8-cell stage. Progressively restricted to outer TE cells of blastocyst. Knockout fails to develop trophectoderm.
3. Nanog; stochastically expressed from 8-cell stage. Switched off in TE. Expressed in salt and pepperpattern in ICM eventually restricted to primitive ectoderm at d4. Knockout fails to develop ICM.
4. Gata6 (+Gata4); stochastically expressed from 8-cell stage. Switched off in TE. Expressed in salt and pepper pattern in ICM eventually restricted to primitive endoderm at d4. Double knockout fails to develop PE.
Four master transcription factors for early lineage determinationin preimplantation development
Trophectoderm (TE) Primitive endoderm (PE)
Inner cell mass (ICM)
Zona pelucidaBlastocoel cavity
Blastomere
Primitive ectoderm (PrEct)
Day 3.0 Day 3.5 Day 4.0
Morula BlastocystCleavage

Day 3.0 Day 3.5 Day 4.0
High NanogLow GATA6
Low NanogHigh GATA6
• Reciprocal salt and pepper pattern of Nanog and GATA6 in ICM cells of mid-stage blastocysts
Trophectoderm (TE) Primitive endoderm (PE)
Inner cell mass (ICM)
Blastocoel cavity
Primitive ectoderm (PrEct)
Specification of primitive endoderm lineage
Chazaud et al (2006) Dev Cell 10 p615-24.
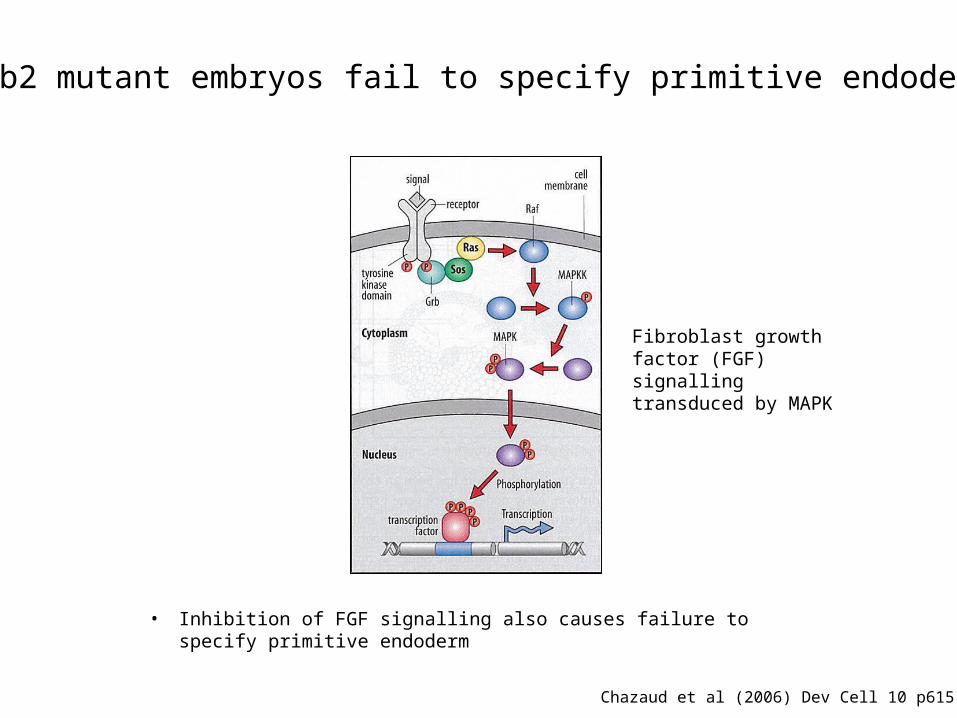
Fibroblast growth factor (FGF) signalling transduced by MAPK
Grb2 mutant embryos fail to specify primitive endoderm
Chazaud et al (2006) Dev Cell 10 p615-24.
• Inhibition of FGF signalling also causes failure to specify primitive endoderm

Grb2
Fgf2r
Gata6NanogGata6
Nanog
Fgf4
Fgf4
Mapk
Fgf4 high Fgf2r high
• Only Nanog expressing ICM cells seen in Grb2 knockout or with disruption of FGF signalling
• Negative feedback by Gata6 on Nanog and vice versa?
Fibroblast growth factor (FGF) signalling regulates primitive endoderm to primitive ectoderm switching
Primitive ectoderm(PrEct) cell
Primitive endoderm(PE) cell
• Cell sorting mechanism?
• FGF4 gene is activated by Oct4
Chazaud et al (2006) Dev Cell 10 p615-24.
Cell sorting

Embryonic Stem (ES) CellsStem cells and progenitors;
Terminology for differentiative capacity of stem cells/progenitors;
• Totipotent; capable of giving rise to all differentiated cell types of the organism, including extraembryonic lineages e.g. morula cells
• Pluripotent; capable of giving rise to cell types of the three germ layers, ectoderm, mesodermand endoderm eg primitive ectoderm cells of the blastocyst.
• Multipotent – capable of giving rise to a limited number of differentiated cell types, e.g.adult stem cells and progenitors
Stem cell; unlimited capacity to self-renew and produce differentiated derivatives
Progenitor cell; limited capacity to self-renew and produce differentiated derivatives
Terminally differentiated cell

Embryonal carcinoma (EC) cells
• Teratocarinomas are malignant tumours derived from germ cells and comprising multiple cell types from all three germ layers, indicating the presence of a pluripotent stem cell population.
• Occur at high frequency in 129 strain of mouse or can be produced by injecting early embryo cells into testis or kidney capsule of syngeneic host.
• Pluripotent stem cell tissue culture cell lines derived from teratocarcinomas are termed embryonal carcinoma (EC) cells. They have an abnormal karyotype and express high levels of alkaline phosphatase.
• EC cells can self-renew indefinitely and can undergo lineage differentiation in vitro and in vivo, following transfer into recipient blastocysts. Cannot contribute to germline
Teratoma
Martin and Evans (1974), Cell 2, p163-172

ES cells
• Contribute to the germ-line of chimeric animals (blastocyst injection) and can therefore be transmitted to subsequent generations.
• Derived from blastocyst stage embryos
• Grow as ‘clumps’ or ‘colonies’ by culturing with fetal calf-serum (FCS) on layer of inactivated primary embryonic fibroblast cells (PEFs).
• Have stable normal karyotype
Alkaline phosphatase positive
Evans and Kaufman (1984) Nature 292, p154-6
• Contribute to all three germ layers (but not trophectoderm) when differentiated in vitro or when transferred to recipient blastocyst – pluripotent.
• Efficient at homologous recombination allowing development of gene knockout technology.

Transcription factor circuitry in ES cells
Availability of unlimited quantity of ES cells grown in vitro has facillitated genome wideanalysis. Key findings include;
• Oct, Nanog and Sox2 participate in negative regulatory loops to block expression of core transcription factors of trophectoderm and primitive endoderm lineages.
• Core transcription factors Oct4, Nanog and Sox2 co-occupy a large proportion of target genes
• Other target genes can be either activated or repressed (recruitment of co-activators or co-repressors).
• Repressed target genes are associated with differentiation into different lineages and are held in a‘poised’ configuration by epigenetic mechanisms (e.g. Polycomb).
• Oct4, Nanog and Sox2 participate in positive feedback loops with themselves and one another to stably maintain the pluripotent state
Boyer et al (2005) Cell 122, p947-56

What is an ES cell?
• No self-renewing pool of embryonic precursors in ICM or epiblast – ES cells are ‘synthetic’.
• Single cell transcriptomics suggest closest to ICM (primitive ectoderm) cells of the blastocyst.

Homogeneous expression of Oct4 but not Nanog
Oct4 Nanog
Immunostaining of ES cell colonies

DNA methylation in ES cells
Somatic cells ES cells
% meCpG 70-80 70-80
Santos et al (2002) Dev Biol 241, 172-182.

Signalling pathways regulating self-renewal and differentiation of mouse ES cells
• Recent evidence suggests LIF +BMP blocks autostimulation of differentiation by FGF4
FGFsVia ERK1/2 pathway
LIF/STAT3 (JAK/STAT)and BMP/Smad/Id
LIF/STAT3 andBMP/Smad/Id
2i - Small moleculeinhibitors of ERK
GSK inhibition(wnt?)
GSK inhibition(wnt?)
Ying et al (2008) Nature 453, p519-23

Without 2i With 2i
Somatic cells ES cells 2i ES cells
% meCpG 70-80 70-80 25-30%
• Due to reduced levels of de novo DNA methyltransferases (Dnmt3a, Dnmt3b), and enhanced levels of enzymes that actively remove methyl groups from CpG.
LIF+ serum
2i
Ground state pluripotency
2i ES cells more closely resemble embryo precursors of the blastocyst
Leitch et al (2013) Nat Struct Mol Biol 20, p311-6

Stem cell types isolated from early mouse embryosDay 4.0 Day 5.5Day 3.5
+LIF +BMP
Polar Trophectoderm
Mural Trophectoderm
Primitive ectodermPrimitive endodermICM
Polar Trophectoderm
Mural Trophectoderm
+FGF4 -LIF+ feeders +FGF
+Activin
Extraembryonicectoderm
Epiblast
Visceralendoderm
Parietalendoderm
ES cell TS cell XEN cell EpiSC(Trophoblaststem cell)
(Extraembryonicendoderm cell)
(Epiblast stem cell)
ChimeraContribution
Germ layersGerm line
TrophectodermPrimitive endoderm
Germ layersGerm line
TrophectodermPrimitive endoderm
Germ layersGerm line
TrophectodermP endoderm
Germ layersGerm line
TrophectodermPrimitive endoderm
In vitrodifferentiation
(-LIF/-BMP)Germ layersGerm cells
Primitive endoderm
(-FGF)Trophoblast giant
cells)
(-FGF)Parietal endoderm like
(-FGF/Activin)Germ layers
+FGF4 +LIF + feeders
Tanaka et al (1998) Science 282, p2072-5; Brons et al (2007) Nature 448, p191-5;Kunath et al (2005), Development, 132, p1649-61
Epigenome(X inactivation Pre-X inactivation
Imprinted-X inactivation
Imprinted-X inactivation
Random-X inactivation

Interconversion of embryo stem cell types
TSES
EpiSC
XEN
+GATA6 and/or+OCT4
+FGF4+LIF
+CDX2and/or-OCT4
+FGF4- LIF
+LIF+2iOr+KLF4
+FGF2+Activin
+serum freemedium
Niwa (2007) Development 134, p635-46

ES cells from other species.
• Pluripotent human ESCs (Thomson et al, 1998, Science 282, p1145-47) derived from blastocysts explanted onto mouse feeder cells. Addition of bFGF improves maintenance.
• Human ESCs resemble mouse EpiSCs e.g. post X inactivation and dependent on Fgf4 and nodalsignals from feeder cells. Not ground state.
• Considered to have significant potential in regenerative therapies.
• 2i method has opened up the possibility of obtaining ESCs from any mouse strain and from otherspecies, notable success being rat (Buehr et al, 2008, Cell 135, p1287-98).

Hatching
Four days after fertilization the blastocyst hatches from the zona pellucida as a precursor to implantation in the uterine wall.

Development of the egg cylinder
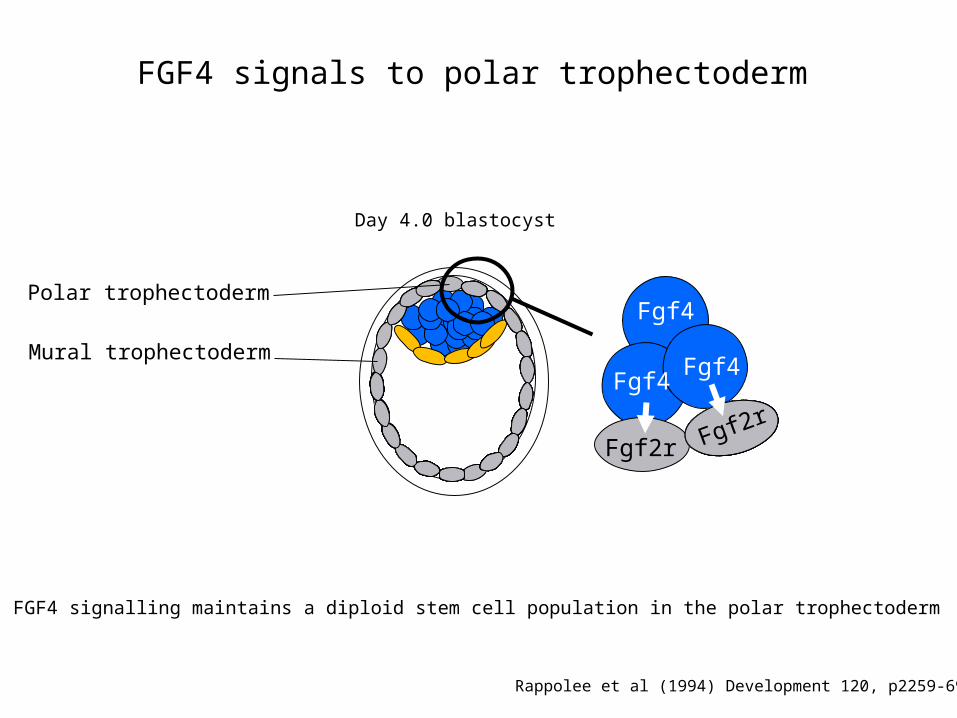
FGF4 signals to polar trophectoderm
Day 4.0 blastocyst
Mural trophectoderm
Polar trophectoderm
Fgf4Fgf4
Fgf4
Fgf2r Fgf2r
• FGF4 signalling maintains a diploid stem cell population in the polar trophectoderm
Rappolee et al (1994) Development 120, p2259-69

Overview of gastrulation
What determines the site of initiation of the primitive streak?
Brachyury expression marksthe primitive streak


















