
L* Michael Berger (Center for Brain Research, Medical University Vienna, Austria): Ligand/Receptor.
165
L* http://cwx.prenhall.com/horton/medialib/media_portfolio/09.html Michael Berger (Center for Brain Research, Medical University Vienna, Austria): Ligand/Receptor Interaction
-
date post
19-Dec-2015 -
Category
Documents
-
view
213 -
download
0
Transcript of L* Michael Berger (Center for Brain Research, Medical University Vienna, Austria): Ligand/Receptor.

L*
http://cwx.prenhall.com/horton/medialib/media_portfolio/09.html
Michael Berger (Center for Brain Research, Medical University Vienna, Austria):
Ligand/Receptor Interaction

Wenn Du mit anderen ein Schiff bauen willst,
Antoine de Saint Exupery

Wenn Du mit anderen ein Schiff bauen willst,beginne nicht, mit Ihnen Holz zu sammeln,
Antoine de Saint Exupery

Wenn Du mit anderen ein Schiff bauen willst,beginne nicht, mit Ihnen Holz zu sammeln,
sondern wecke in Ihnen die Sehnsuchtnach dem großen weiten Meer.
Antoine de Saint Exupery

What is a receptor?

What is a receptor?
A physical target mediating the physiological effect of a drug.

What is a receptor?
A physical target mediating the physiological effect of a drug.
What is a ligand?

What is a receptor?
A physical target mediating the physiological effect of a drug.
What is a ligand?A substance that (strongly) binds to a tissue.

What is a receptor?
A physical target mediating the physiological effect of a drug.
What is a ligand?A substance that (strongly) binds to a tissue.
What is an agonist?

What is a receptor?
A physical target mediating the physiological effect of a drug.
What is a ligand?A substance that (strongly) binds to a tissue.
What is an agonist?
A substance that causes an effect, an active change in the target tissue.

What is a receptor?
A physical target mediating the physiological effect of a drug.
What is a ligand?A substance that (strongly) binds to a tissue.
What is an agonist?
A substance that causes an effect, an active change in the target tissue.
What is an antagonist?

What is a receptor?
A physical target mediating the physiological effect of a drug.
What is a ligand?A substance that (strongly) binds to a tissue.
What is an agonist?
A substance that causes an effect, an active change in the target tissue.
What is an antagonist?
A substance that blocks the effect of an agonist

What is a receptor?
A physical target mediating the physiological effect of a drug.
What is a ligand?A substance that (strongly) binds to a tissue.
What is an agonist?
A substance that causes an effect, an active change in the target tissue.
What is an antagonist?
A substance that blocks the effect of an agonist
What is a transmitter?

What is a receptor?
A physical target mediating the physiological effect of a drug.
What is a ligand?A substance that (strongly) binds to a tissue.
What is an agonist?
A substance that causes an effect, an active change in the target tissue.
What is an antagonist?
A substance that blocks the effect of an agonist
What is a transmitter?
A natural agonist released by a cell and acting on a neighboring cell.
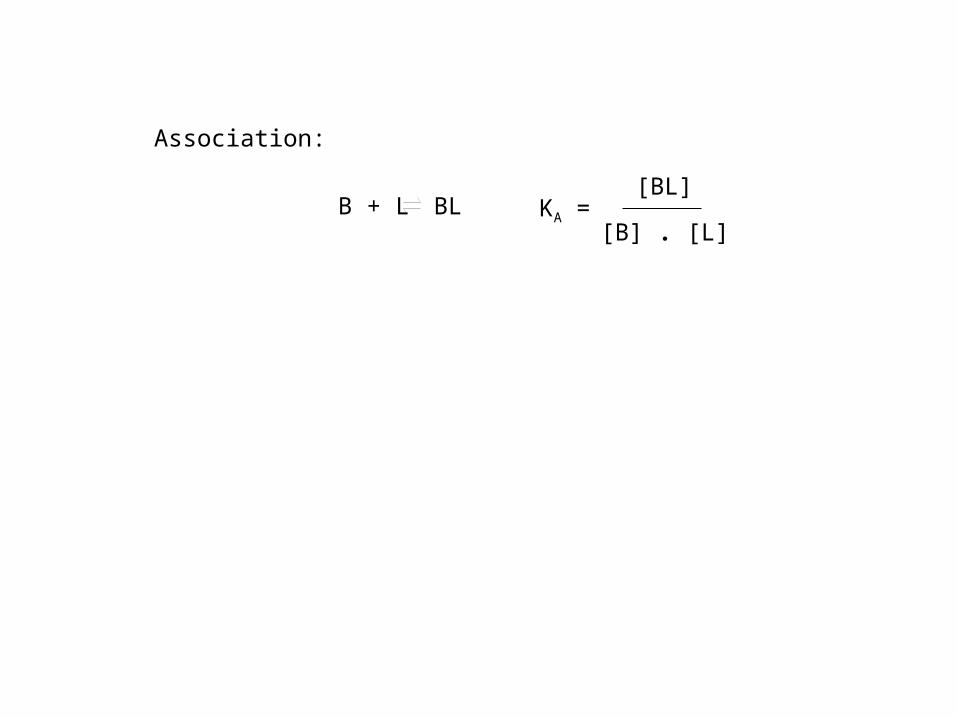
B + L BL
Association:
KA =[BL]
[B] . [L]

B + L BL
Association:
KA =[BL]
[B] . [L]
KA: association equilibrium constant

B + L BL
Association:
KA =[BL]
[B] . [L]
KA: association equilibrium constant
BL B + L
Dissociation:
KD =[BL]
[B] . [L]
KD: dissociation equilibrium constant

B + L BL
Association:
KA =[BL]
[B] . [L]
KA: association equilibrium constantdimension: (concentration)-1
BL B + L
Dissociation:
KD =[BL]
[B] . [L]
KD: dissociation equilibrium constant

B + L BL
Association:
KA =[BL]
[B] . [L]
KA: association equilibrium constantdimension: (concentration)-1
BL B + L
Dissociation:
KD =[BL]
[B] . [L]
KD: dissociation equilibrium constantdimension: concentration

B + L BL BL B + L
Association: Dissociation:

B + L BL BL B + L
Association: Dissociation:
Strong binding: equilibrium is
on right side on left side

B + L BL BL B + L
Association: Dissociation:
Strong binding: equilibrium is
on right side on left side
KA =[BL]
[B] . [L]>> 1 KD =
[BL]
[B] . [L]<< 1

B + L BL BL B + L
Association: Dissociation:
Strong binding: equilibrium is
on right side on left side
KA =[BL]
[B] . [L]>> 1 KD =
[BL]
[B] . [L]<< 1
ln KA positiv ln KD negativ

Van't Hoff: ΔGo = - RT . ln KA = + RT . ln KD
ΔGo: change in free enthalpy (Gibbs energy)R: universal gas constant, 1.987 cal/(Mol . °K) or 8.314 J/(Mol
. °K)T: absolute temperature
B + L BL BL B + L
Association: Dissociation:
Strong binding: equilibrium is
on right side on left side
KA =[BL]
[B] . [L]>> 1 KD =
[BL]
[B] . [L]<< 1
ln KA positiv ln KD negativ

The Van‘t Hoff equation allows the calculation of the free enthalpy change of a reaction from the reaction‘s equilibrium constant:
ΔGo (20 °C)
KA KD kcal/Mol kJ/Mol
107 M-1 10-7 M -9.4 -39.2
108 M-1 10-8 M -10.7 -44.8
109 M-1 10-9 M -12.0 -50.3
Van't Hoff: ΔGo = - RT . ln KA = + RT . ln KD
ΔGo: change in free enthalpy (Gibbs energy)R: universal gas constant, 1.987 cal/(Mol . °K) or 8.314 J/(Mol
. °K)T: absolute temperature

Examples for the change in free enthalpy Go in various reactions
Glucose + 6 O2 6 CO2 + 6 H2O -686 H2 + ½ O2 H2O -46 ATP ADP + Pi -7.3
ΔGo (kcal/Mol)

Examples for the change in free enthalpy Go in various reactions
Glucose + 6 O2 6 CO2 + 6 H2O -686 H2 + ½ O2 H2O -46 ATP ADP + Pi -7.3
ΔGo (kcal/Mol)
In these reactions, Go is reduced (exergonic processes)

Examples for the change in free enthalpy Go in various reactions
Glucose + 6 O2 6 CO2 + 6 H2O -686 H2 + ½ O2 H2O -46 ATP ADP + Pi -7.3
ΔGo (kcal/Mol)
Bond dissociation energies
HO-H HO· + ·H 118CH3CH2-H CH3CH2· + ·H 101CH3-CH3 CH3· + ·CH3 90
In these reactions, Go is reduced (exergonic processes)

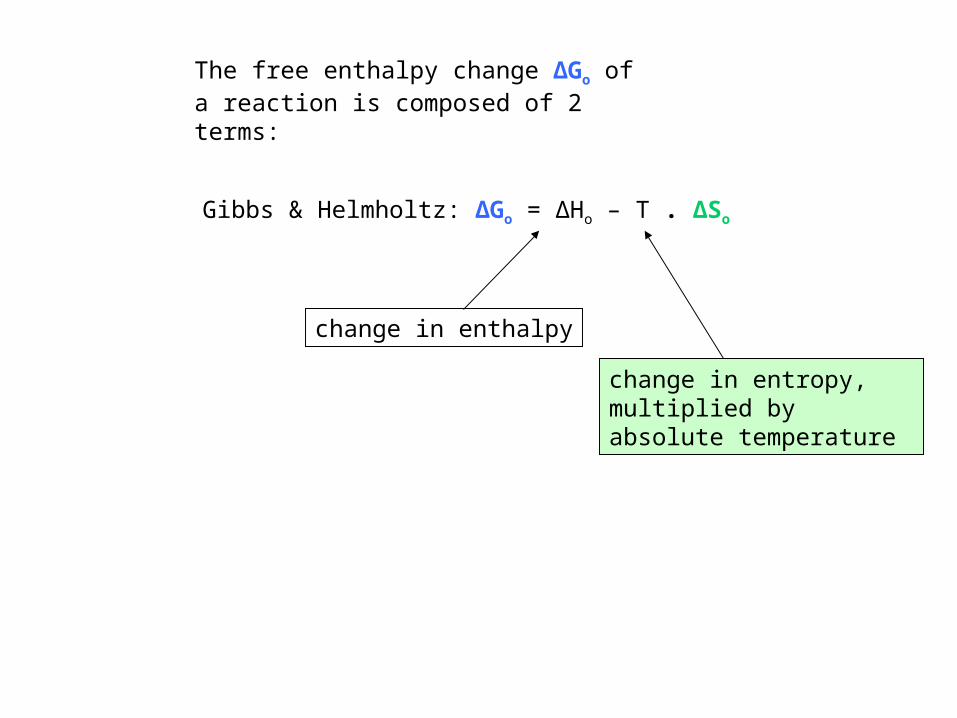
Gibbs & Helmholtz: ΔGo = ΔHo – T . ΔSo
The free enthalpy change ΔGo of a reaction is composed of 2 terms:

Gibbs & Helmholtz: ΔGo = ΔHo – T . ΔSo
The free enthalpy change ΔGo of a reaction is composed of 2 terms:
change in enthalpy
Gibbs & Helmholtz: ΔGo = ΔHo – T . ΔSo
The free enthalpy change ΔGo of a reaction is composed of 2 terms:
change in enthalpy
change in entropy, multiplied by absolute temperature

Gibbs & Helmholtz: ΔGo = ΔHo – T . ΔSo
The free enthalpy change ΔGo of a reaction is composed of 2 terms:
change in enthalpy
change in entropy, multiplied by absolute temperature
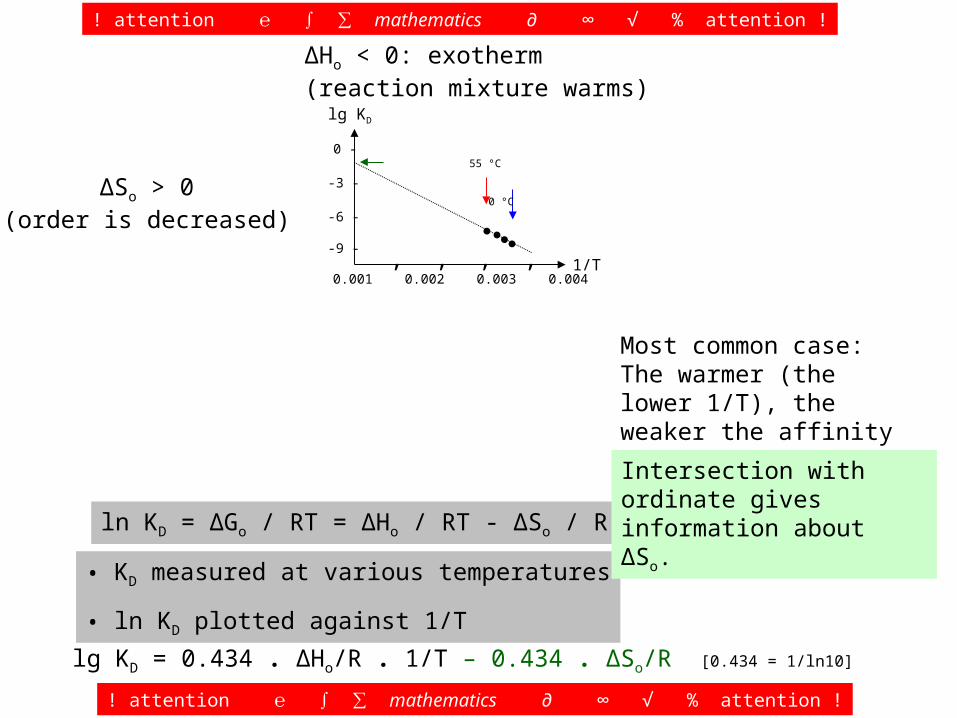
ln KD = ΔGo / RT = ΔHo / RT - ΔSo / R
• KD measured at various temperatures
• ln KD plotted against 1/T
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !

ln KD = ΔGo / RT = ΔHo / RT - ΔSo / R
• KD measured at various temperatures
• ln KD plotted against 1/T
ΔHo < 0: exotherm(reaction mixture warms)
ΔSo > 0(order is decreased)
lg KD
1/T-9 -
-6 -
-3 -
0 -
‚‚ ‚‚0.001 0.002 0.003 0.004
●●●●
Most common case: The warmer (the lower 1/T), the weaker the affinity (the less negative lg KD).
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
55 °C 0 °C
lg KD = 0.434 . ΔHo/R . 1/T – 0.434 . ΔSo/R [0.434 = 1/ln10]
ln KD = ΔGo / RT = ΔHo / RT - ΔSo / R
• KD measured at various temperatures
• ln KD plotted against 1/T
ΔHo < 0: exotherm(reaction mixture warms)
ΔSo > 0(order is decreased)
lg KD
1/T-9 -
-6 -
-3 -
0 -
‚‚ ‚‚0.001 0.002 0.003 0.004
●●●●
Most common case: The warmer (the lower 1/T), the weaker the affinity (the less negative lg KD).
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
55 °C 0 °C
lg KD = 0.434 . ΔHo/R . 1/T – 0.434 . ΔSo/R [0.434 = 1/ln10]
Intersection with ordinate gives information about ΔSo.

ln KD = ΔGo / RT = ΔHo / RT - ΔSo / R
• KD measured at various temperatures
• ln KD plotted against 1/T
ΔHo < 0: exotherm(reaction mixture warms)
ΔSo > 0(order is decreased)
lg KD
1/T-9 -
-6 -
-3 -
0 -
‚‚ ‚‚0.001 0.002 0.003 0.004
●●●●
Most common case: The warmer (the lower 1/T), the weaker the affinity (the less negative lg KD).
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
55 °C 0 °C
lg KD = 0.434 . ΔHo/R . 1/T – 0.434 . ΔSo/R [0.434 = 1/ln10]
Intersection with ordinate gives information about ΔSo.
Slope allows access to ΔHo.
ln KD = ΔGo / RT = ΔHo / RT - ΔSo / R
• KD measured at various temperatures
• ln KD plotted against 1/T
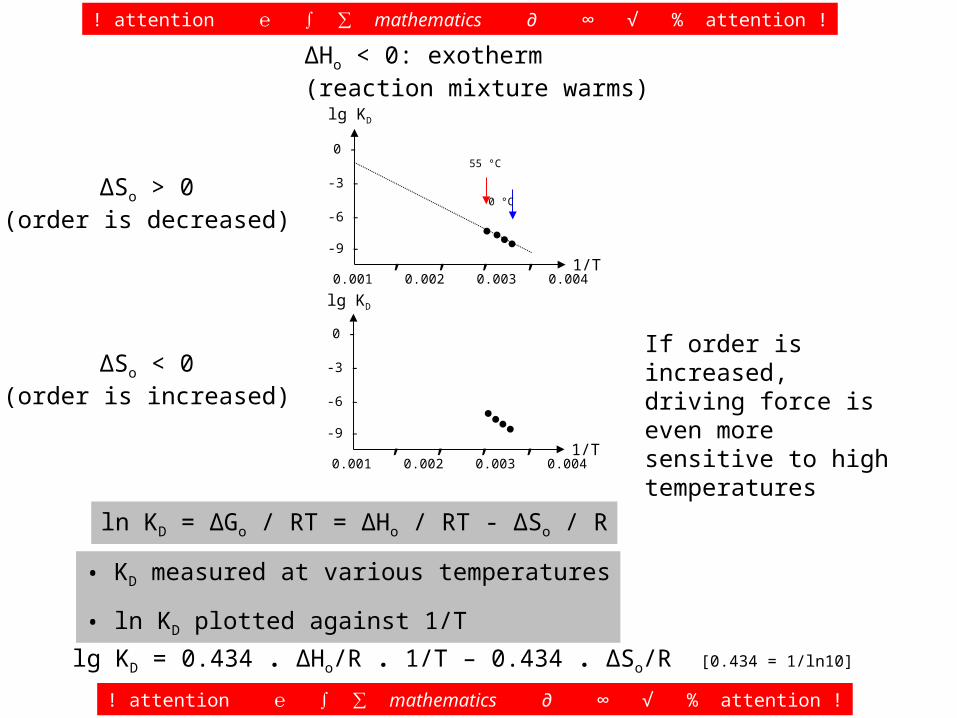
ΔSo > 0(order is decreased)
ΔSo < 0(order is increased)
lg KD
1/T-9 -
-6 -
-3 -
0 -
‚‚ ‚‚0.001 0.002 0.003 0.004
lg KD
1/T-9 -
-6 -
-3 -
0 -
‚‚ ‚‚0.001 0.002 0.003 0.004
If order is increased, driving force is even more sensitive to high temperatures
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
lg KD = 0.434 . ΔHo/R . 1/T – 0.434 . ΔSo/R [0.434 = 1/ln10]
ΔHo < 0: exotherm(reaction mixture warms)
●●●●
55 °C 0 °C
●●●●

ln KD = ΔGo / RT = ΔHo / RT - ΔSo / R
• KD measured at various temperatures
• ln KD plotted against 1/T
ΔSo > 0(order is decreased)
ΔSo < 0(order is increased)
lg KD
1/T-9 -
-6 -
-3 -
0 -
●●●●
‚‚ ‚‚0.001 0.002 0.003 0.004
lg KD
1/T-9 -
-6 -
-3 -
0 -
‚‚ ‚‚0.001 0.002 0.003 0.004
If order is increased, driving force is even more sensitive to high temperatures
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
lg KD = 0.434 . ΔHo/R . 1/T – 0.434 . ΔSo/R [0.434 = 1/ln10]
ΔHo < 0: exotherm(reaction mixture warms)
●●●●
55 °C 0 °C
It may be difficult to obtain solid data that allow to decide, if ΔSo is > or < 0.

ln KD = ΔGo / RT = ΔHo / RT - ΔSo / R
• KD measured at various temperatures
• ln KD plotted against 1/T
ΔSo > 0(order is decreased)
ΔSo < 0(order is increased)
lg KD
1/T-9 -
-6 -
-3 -
0 -
‚‚ ‚‚0.001 0.002 0.003 0.004
lg KD
1/T-9 -
-6 -
-3 -
0 -
‚‚ ‚‚0.001 0.002 0.003 0.004
lg KD
1/T-9 -
-6 -
-3 -
0 -
●●●●
‚‚ ‚‚0.001 0.002 0.003 0.004
ΔHo > 0: endotherm(reaction mixture cools)
Endotherm binding is driven by decrease in order only; here, driving force increases with temperature.
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
lg KD = 0.434 . ΔHo/R . 1/T – 0.434 . ΔSo/R [0.434 = 1/ln10]
ΔHo < 0: exotherm(reaction mixture warms)
●●●●
55 °C 0 °C
●●●●
Mechanisms contributing to ligand/receptor interaction:
1. Ionic interaction2. Hydrogen bonds3. Hydrophobic interaction4. Cation/ interaction5. Van der Waals interaction

attraction between 2 charges depends on
e1 . e2
D . r2
ionic interaction
r ... distanceD ... dielectric constant

attraction between 2 charges depends on
vacuum ...1.0
hexane ...1.9
H2O ...78
e1 . e2
D . r2
ionic interaction
r ... distanceD ... dielectric constant

attraction between 2 charges depends on
vacuum ...1.0
hexane ...1.9
H2O ...78
e1 . e2
D . r2
ionic interaction
r ... distanceD ... dielectric constant
In water, ionic interaction is hindered
by shells of water molecules surrounding
each ion.

hydrogen bondsB + L BL
Formation of a hydrogen bond is highly exergonic, yields 3-7 kcal/mol

hydrogen bonds
BH2O + LH2O BL + H2OH2O
B + L BL
Formation of a hydrogen bond is highly exergonic, yields 3-7 kcal/mol
However, enthalpy balance is poor:

hydrogen bonds
BH2O + LH2O BL + H2OH2O
B + L BL
Formation of a hydrogen bond is highly exergonic, yields 3-7 kcal/mol
However, enthalpy balance is poor:
1. Break this bond.

hydrogen bonds
BH2O + LH2O BL + H2OH2O
B + L BL
Formation of a hydrogen bond is highly exergonic, yields 3-7 kcal/mol
However, enthalpy balance is poor:
1. Break this bond.
2. Break this bond.

hydrogen bonds
BH2O + LH2O BL + H2OH2O
B + L BL
Formation of a hydrogen bond is highly exergonic, yields 3-7 kcal/mol
However, enthalpy balance is poor:
1. Break this bond.
2. Break this bond.
3. Form this bond.

hydrogen bonds
BH2O + LH2O BL + H2OH2O
B + L BL
Formation of a hydrogen bond is highly exergonic, yields 3-7 kcal/mol
However, enthalpy balance is poor:
1. Break this bond.
2. Break this bond.
3. Form this bond.
4. Form this bond.

hydrogen bonds
BH2O + LH2O BL + H2OH2O
B + L BL
Formation of a hydrogen bond is highly exergonic, yields 3-7 kcal/mol
However, enthalpy balance is poor:
1. Break this bond.
2. Break this bond.
3. Form this bond.
4. Form this bond.
Hydrogen bond formation mainly driven by increase in entropy, since the water molecules “get more freedom“ (2 kcal per mol of water).
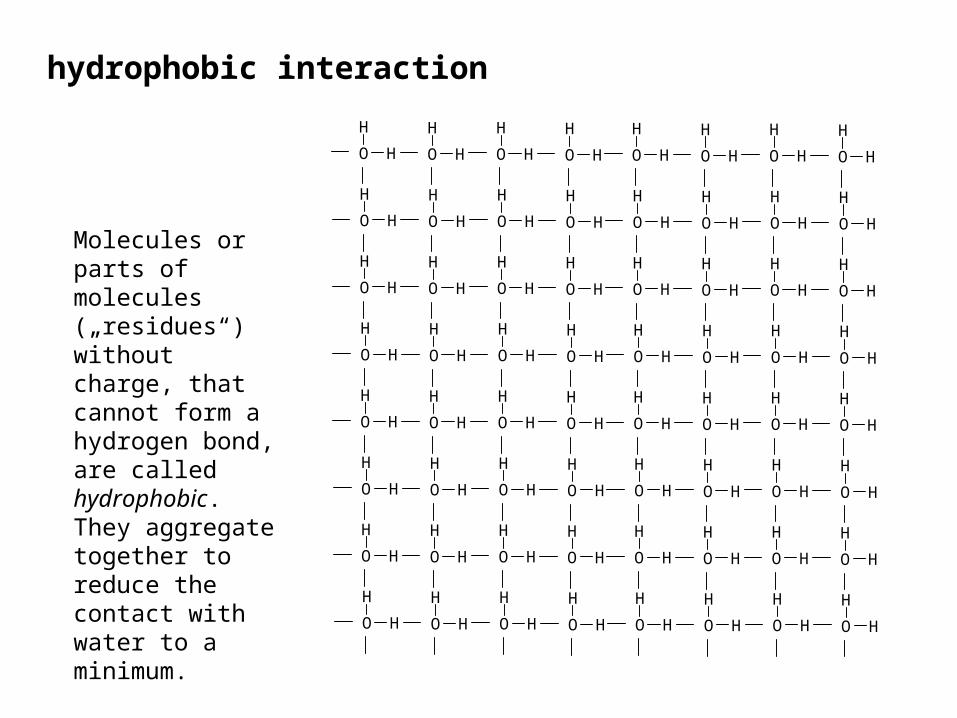
hydrophobic interaction
Molecules or parts of molecules („residues“) without charge, that cannot form a hydrogen bond, are called hydrophobic. They aggregate together to reduce the contact with water to a minimum.
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H

HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
1 2
3
4567
8
9 10
11
12131415
16
hydrophobic interaction
Molecules or parts of molecules („residues“) without charge, that cannot form a hydrogen bond, are called hydrophobic. They aggregate together to reduce the contact with water to a minimum.

HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
HO
H
1
23 4
5
6
7
8910
11
12
hydrophobic interaction
Molecules or parts of molecules („residues“) without charge, that cannot form a hydrogen bond, are called hydrophobic. They aggregate together to reduce the contact with water to a minimum.

hydrophobic interaction
• This example is nice, but wrong.
• Hydrogene bonds are never left open.
• In contact with an inert partner, water molecules are highly ordered.
• Reduction of contact area leads to reduced order.

• This example is nice, but wrong.
• Hydrogene bonds are never left open.
• In contact with an inert partner, water molecules are highly ordered.
• Reduction of contact area leads to reduced order.
• Reduction of Go by hydrophobic interaction is always due to the entropy term T . ΔS
hydrophobic interaction
Gibbs & Helmholtz: ΔGo = ΔHo – T . ΔSo

• This example is nice, but wrong.
• Hydrogene bonds are never left open.
• In contact with an inert partner, water molecules are highly ordered.
• Reduction of contact area leads to reduced order.
• Reduction of Go by hydrophobic interaction is always due to the entropy term T . ΔS
• Empirical rule: Δ Go = -0.03 x area hidden from water (in Ǻ2).
hydrophobic interaction
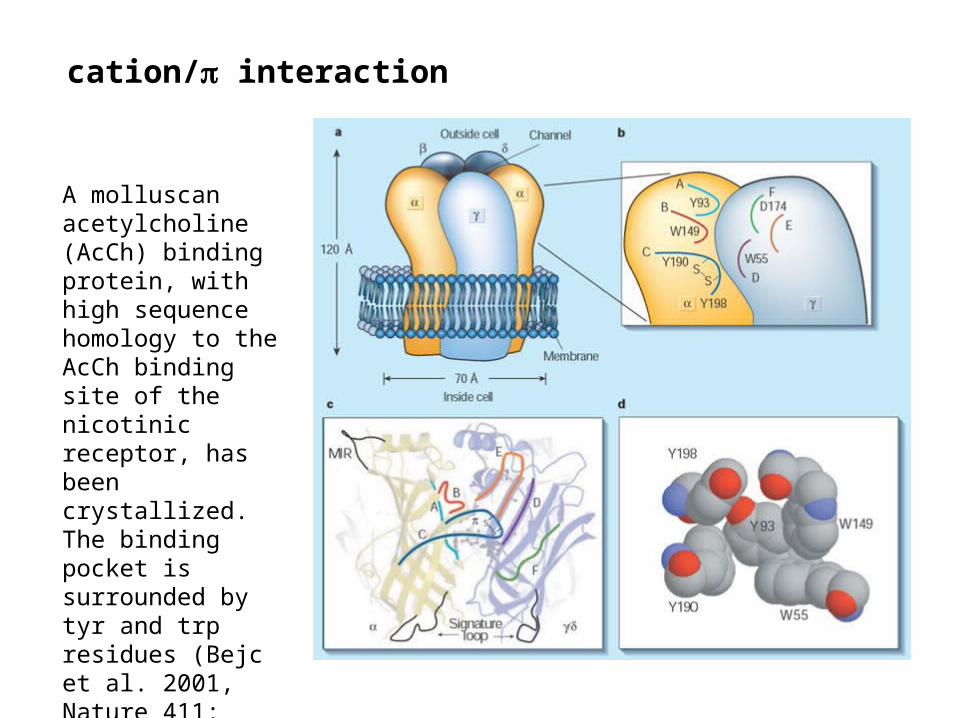
cation/ interaction
A molluscan acetylcholine (AcCh) binding protein, with high sequence homology to the AcCh binding site of the nicotinic receptor, has been crystallized. The binding pocket is surrounded by tyr and trp residues (Bejc et al. 2001, Nature 411: 269)
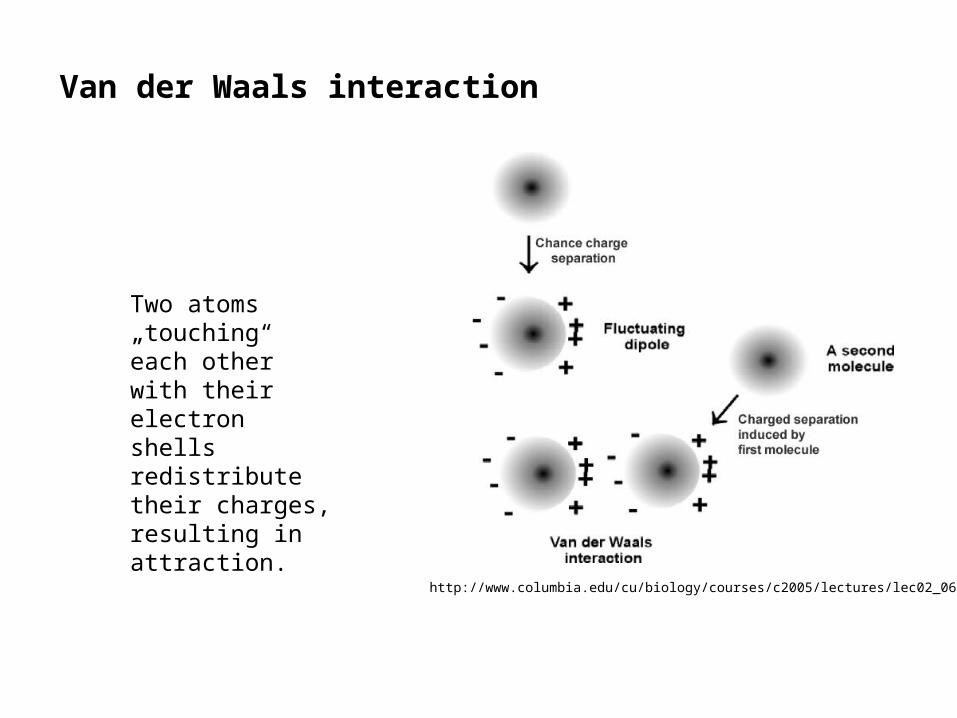
Van der Waals interaction
http://www.columbia.edu/cu/biology/courses/c2005/lectures/lec02_06.html
Two atoms „touching“ each other with their electron shells redistribute their charges, resulting in attraction.
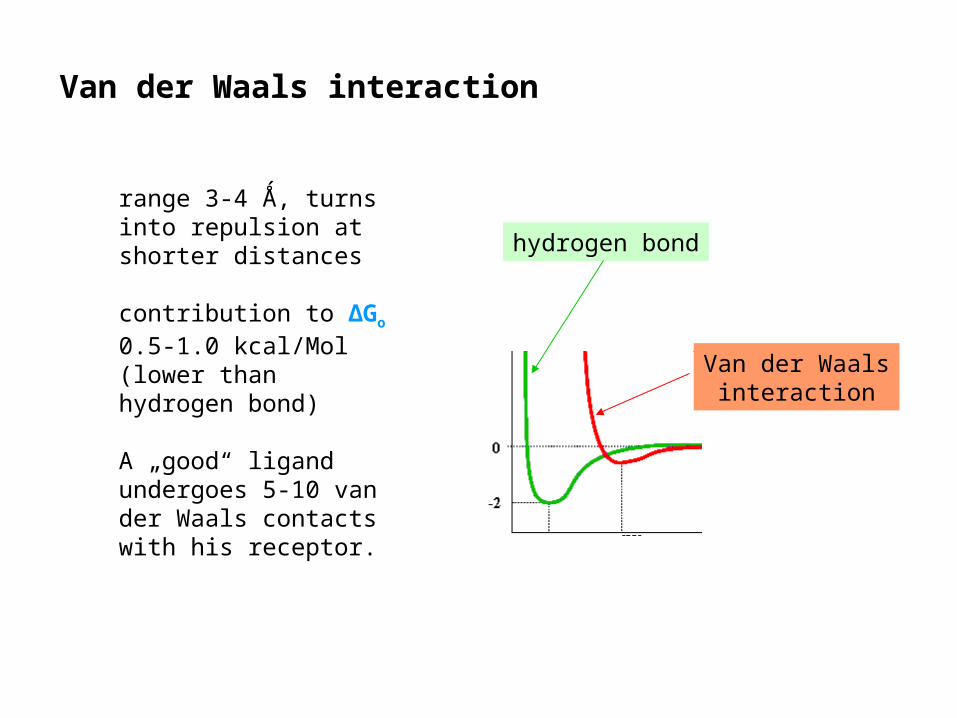
Van der Waals interaction
hydrogen bond
Van der Waals interaction
range 3-4 Ǻ, turns into repulsion at shorter distances
contribution to ΔGo 0.5-1.0 kcal/Mol (lower than hydrogen bond)
A „good“ ligand undergoes 5-10 van der Waals contacts with his receptor.

1998 Leif Saul
Van der Waals interaction
The ensemble of van der Waals interactions is responsible for the key/lock nature of ligand/receptor interaction.


Example for the interaction of a hypothetical ligand with its receptor:
formation of a hydrogen bond ... - 5.0 loss of hydrogen bond with H2O... + 5.0
kcal/mol

Example for the interaction of a hypothetical ligand with its receptor:
formation of a hydrogen bond ... - 5.0 loss of hydrogen bond with H2O... + 5.0
kcal/mol
preliminary balance: ± 0

Example for the interaction of a hypothetical ligand with its receptor:
formation of a hydrogen bond ... - 5.0 loss of hydrogen bond with H2O ... + 5.02 H2O set free … - 4.0Hydrophobic interaction … - 2.08 van der Waals contacts … - 4.7
kcal/mol

Example for the interaction of a hypothetical ligand with its receptor:
formation of a hydrogen bond ... - 5.0 loss of hydrogen bond with H2O ... + 5.02 H2O set free … - 4.0Hydrophobic interaction … - 2.08 van der Waals contacts … - 4.7
kcal/mol
balance: -10.7

ΔGo (20 °C)
KA KD kcal/Mol kJ/Mol
107 M-1 10-7 M -9.4 -39.2
108 M-1 10-8 M -10.7 -44.8
109 M-1 10-9 M -12.0 -50.3
Example for the interaction of a hypothetical ligand with its receptor:
formation of a hydrogen bond ... - 5.0 loss of hydrogen bond with H2O ... + 5.02 H2O set free … - 4.0Hydrophobic interaction … - 2.08 van der Waals contacts … - 4.7
kcal/mol
balance: -10.7

How many receptors do we expect in a responsive tissue?
Which analytical tools will be necessary to detect them?

How many receptors do we expect in a responsive tissue?
• Theoretical assumption: the tissue consists of cubes 10 µm x 10 µm x 10 µm
• Then, 1 mg tissue would consist of 100 x 100 x 100 = 106 cells

• Theoretical assumption: the tissue consists of cubes 10 µm x 10 µm x 10 µm
• Then, 1 mg tissue would consist of 100 x 100 x 100 = 106 cells
• If each cell bears 1 binding site, this would result in 106 binding sites / mg tissue
• 1 fMol = 6 x 1023-15 = 6 x 108 molecules
• 106 molecules = 1/600 fMol
How many receptors do we expect in a responsive tissue?
Josef Loschmidt (1821-1895)

• The most common binding sites occur at densities of 10 to several 100 fMol/mg tissue.
• This is much more than 1/600 fMol/mg tissue.• Thus, receptor-bearing cells have not only 1, but several thousands of
binding sites.
How many receptors do we expect in a responsive tissue?
Freeze-fracture analysis of AMPA receptors labelled with immuno gold antibodies (5 nm)
at the postsynaptic site on cerebellar
Purkinje cells (climbing fiber
input). Tanaka et al (2005) J
Neurosci 25:799

Labelling: Replacement of one or more protons by tritium (3H; molecule practically unchanged)
Marie & Pierre Curie
Which analytical tools will be necessary to detect them?

Marie & Pierre Curie
Which analytical tools will be necessary to detect them?
Radioactivity measured in
• Curie (Ci, mCi, µCi)
(the radioactivity of 1 g radium)

Henry Becquerel
Which analytical tools will be necessary to detect them?
Radioactivity measured in
• Curie (Ci, mCi, µCi)
• Becquerel (Bq, decays / s)
• dpm (decays / min)
1 Bq = 60 dpm

Which analytical tools will be necessary to detect them?
Radioactivity measured in
• Curie (Ci, mCi, µCi)
• Becquerel (Bq, decays / s)
• dpm (decays / min)
1 µCi = 2 220 000 dpm1 nCi = 2 220 dpm1 pCi = 2.22 dpm
Henry Becquerel

t ½ 1 10 100 1000 fMol
14C 5 730 y
3H 12.4 y
35S 87 d
131I 8 d
The shorter the half-life, the hotter the radioligand.
Which analytical tools will be necessary to detect them?
Comparison of 3H with other nuclides (1 radioactive atom / molecule)

Which analytical tools will be necessary to detect them?
How many dpm can be expected from 1 fMol 3H?

A rule of thumb is a principle with broad application that is not intended to be strictly accurate or reliable for every situation. (Wikipedia)
Which analytical tools will be necessary to detect them?
How many dpm can be expected from 1 fMol 3H?

Which analytical tools will be necessary to detect them?
How many dpm can be expected from 1 fMol 3H?• 1 fMol = 10-15 x 6 . 1023 = 6 . 108 molecules

Which analytical tools will be necessary to detect them?
How many dpm can be expected from 1 fMol 3H?• 1 fMol = 10-15 x 6 . 1023 = 6 . 108 molecules
• t½ = 12.3 y = 4 500 d = 108 000 h = 6.48 . 106 min

Which analytical tools will be necessary to detect them?
How many dpm can be expected from 1 fMol 3H?• 1 fMol = 10-15 x 6 . 1023 = 6 . 108 molecules
• t½ = 12.3 y = 4 500 d = 108 000 h = 6.48 . 106 min
General idea: Since we know that half of the radioactive nuclei will decay in 6.48 million minutes, we might obtain the number of nuclei decaying in 1 minute simply by dividing half of the number of nuclei by 6.48 millions.

Which analytical tools will be necessary to detect them?
How many dpm can be expected from 1 fMol 3H?• 1 fMol = 10-15 x 6 . 1023 = 6 . 108 molecules
• t½ = 12.3 y = 4 500 d = 108 000 h = 6.48 . 106 min
• dpm = 6 . 108 . 0.5 / 6.48 . 106 = 46
General idea: Since we know that half of the radioactive nuclei will decay in 6.48 million minutes, we might obtain the number of nuclei decaying in 1 minute simply by dividing half of the number of nuclei by 6.48 millions.

Which analytical tools will be necessary to detect them?
How many dpm can be expected from 1 fMol 3H?• 1 fMol = 10-15 x 6 . 1023 = 6 . 108 molecules
• t½ = 12.3 y = 4 500 d = 108 000 h = 6.48 . 106 min
• dpm = 6 . 108 . 0.5 / 6.48 . 106 = 46
0.5 would be correct, if the decay rate would be the same for the whole decay period.
0 5 10 15 200
20
40
60
80
100
linear decay
t1/2
= 12.3 years
%
orig
inal
act
ivity
years

Which analytical tools will be necessary to detect them?
How many dpm can be expected from 1 fMol 3H?• 1 fMol = 10-15 x 6 . 1023 = 6 . 108 molecules
• t½ = 12.3 y = 4 500 d = 108 000 h = 6.48 . 106 min
• dpm = 6 . 108 . ln2 / 6.48 . 106 = 64
0.5 would be correct, if the decay rate would be the same for the whole decay period. However, radioactive decay follows an exponential law; therefore, 0.5 must be replaced by ln2 = 0.69.
0 5 10 15 200
20
40
60
80
100
exponential decay
linear decay
t1/2
= 12.3 years
%
orig
inal
act
ivity
years

Why the natural logarithm of 2?

Which analytical tools will be necessary to detect them?
A … number of radioactive nucleik … decay constant
dA/dt = -k . A∫(1/A)dA = -k . ∫dtln(A/Ao) = -k . Δt
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !

Which analytical tools will be necessary to detect them?
A … number of radioactive nucleik … decay constant
dA/dt = -k . A∫(1/A)dA = -k . ∫dtln(A/Ao) = -k . Δt
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
A = Ao . e-k.Δt

Which analytical tools will be necessary to detect them?
A … number of radioactive nucleik … decay constant
dA/dt = -k . A∫(1/A)dA = -k . ∫dtln(A/Ao) = -k . Δt
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
A = Ao . e-k.Δt
k is related to t½: ln(½) = -k . t½
k = ln2 / t½

Which analytical tools will be necessary to detect them?
A … number of radioactive nucleik … decay constant
dA/dt = -k . A∫(1/A)dA = -k . ∫dtln(A/Ao) = -k . Δt
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
A = Ao . e-k.Δt
k is related to t½: ln(½) = -k . t½
k = ln2 / t½
for 1 min (Δt = 1): -ΔA = k . A . 1 = ln2 / t½ . A

Which analytical tools will be necessary to detect them?
Therefore, it can be expected that 6 . 108 tritium nuclei (1 fMol) will emit
6 . 108 . ln2 / 6.48 . 106 = 64 electrons / min.
A molecule labeled with one single 3H has a specific radioactivity (short: specific activity) of 64 dpm / fMol.

1 µCi = 2 220 000 dpm1 nCi = 2 220 dpm1 pCi = 2.22 dpm
Which analytical tools will be necessary to detect them?
A molecule labeled with one single 3H has a specific radioactivity (short: specific activity) of 64 dpm / fMol.
Remember:
64 dpm / fMol = 28.8 pCi / fMol = 28.8 Ci / mMol
Therefore, it can be expected that 6 . 108 tritium nuclei (1 fMol) will emit
6 . 108 . ln2 / 6.48 . 106 = 64 electrons / min.

64 dpm / fMol = 28.8 pCi / fMol = 28.8 Ci / mMol
Which analytical tools will be necessary to detect them?

64 dpm / fMol = 28.8 pCi / fMol = 28.8 Ci / mMol
Which analytical tools will be necessary to detect them?

t ½ 1 10 100 1000 fMol
14C 5 730 y 0.14 1.4 14 140
3H 12.4 y 64 640 6.4*103 64*103
35S 87 d 3.3*103 33*103 330*103 3.3*106
131I 8 d 36*103 360*103 3.6*106 36*106
dpm / mg tissue
Comparison of 3H with other nuclides (1 radioactive atom / molecule)
Which analytical tools will be necessary to detect them?

t ½ 1 10 100 1000 fMol
14C 5 730 y 0.14 1.4 14 140
3H 12.4 y 64 640 6.4*103 64*103
35S 87 d 3.3*103 33*103 330*103 3.3*106
131I 8 d 36*103 360*103 3.6*106 36*106
most common experimental condition
dpm / mg tissue
Which analytical tools will be necessary to detect them?
Comparison of 3H with other nuclides (1 radioactive atom / molecule)

Properties of 3H
• can replace 1H present in every organic molecule
• does not change the properties of the labeled molecule (no isotope effect)
• t½ 12.4 y
• decay (emits electrons)
• radiation reaches in air 6 mm, in liquid and tissue 6 µm
• relatively safe to work with (no shielding required)
• the only risk is incorporation of > 1 mCi
• only reliable method of counting:

Properties of 3H
• can replace 1H present in every organic molecule
• does not change the properties of the labeled molecule (no isotope effect)
• t½ 12.4 y
• decay (emits electrons)
• radiation reaches in air 6 mm, in liquid and tissue 6 µm
• relatively safe to work with (no shielding required)
• the only risk is incorporation of > 1 mCi
• only reliable method of counting: liquid scintillation

Analytical techniques
equilibrium dialysis
non-equilibrium techniques for receptors in solution
• gel filtration• charcoal adsorption• precipitation• adsorption to glass fiber filters
non.-equilibrium techniques for particulate receptors
• centrifugation• filtration• slice autoradiography
B + L* BL*

Analytical techniques
equilibrium dialysis
non-equilibrium techniques for receptors in solution
• gel filtration• charcoal adsorption• precipitation• adsorption to glass fiber filters
non.-equilibrium techniques for particulate receptors
• centrifugation• filtration• slice autoradiography
BL*
B
L*
L*
B + L* BL*

Analytical techniques
equilibrium dialysis
non-equilibrium techniques for receptors in solution
• gel filtration• charcoal adsorption• precipitation• adsorption to glass fiber filters
non.-equilibrium techniques for particulate receptors
• centrifugation• filtration• slice autoradiography
B + L* BL*

Analytical techniques
equilibrium dialysis
non-equilibrium techniques for receptors in solution
• gel filtration• charcoal adsorption• precipitation• adsorption to glass fiber filters
non.-equilibrium techniques for particulate receptors
• centrifugation• filtration• slice autoradiography
B + L* BL*

Analytical techniques
equilibrium dialysis
non-equilibrium techniques for receptors in solution
• gel filtration• charcoal adsorption• precipitation• adsorption to glass fiber filters
non.-equilibrium techniques for particulate receptors
• centrifugation• filtration• slice autoradiography
B + L* BL*
polyethylene glycoln = 6 000 – 8 000

Analytical techniques
equilibrium dialysis
non-equilibrium techniques for receptors in solution
• gel filtration• charcoal adsorption• precipitation• adsorption to glass fiber filters
non.-equilibrium techniques for particulate receptors
• centrifugation• filtration• slice autoradiography
B + L* BL*

Analytical techniques
equilibrium dialysis
non-equilibrium techniques for receptors in solution
• gel filtration• charcoal adsorption• precipitation• adsorption to glass fiber filters
non.-equilibrium techniques for particulate receptors
• centrifugation• filtration• slice autoradiography
B + L* BL*

Analytical techniques
equilibrium dialysis
non-equilibrium techniques for receptors in solution
• gel filtration• charcoal adsorption• precipitation• adsorption to glass fiber filters
non.-equilibrium techniques for particulate receptors
• centrifugation• filtration• slice autoradiography
B + L* BL*

glass
coating
L*
L* L*L*
L*
L*
L*
L*
L*
L*
L*L*
Analytical techniques
equilibrium dialysis
non-equilibrium techniques for receptors in solution
• gel filtration• charcoal adsorption• precipitation• adsorption to glass fiber filters
non.-equilibrium techniques for particulate receptors
• centrifugation• filtration• slice autoradiography
B + L* BL*

Saturability: a radioligand can only be displaced if the target density is low.
Other examples for saturability: Langmuir isotherme (mono-molecular layer on a surface), enzyme reaction rate (Michaelis-Menten).
http://www.steve.gb.com/science/membranes.html
Saturation & non-specific binding
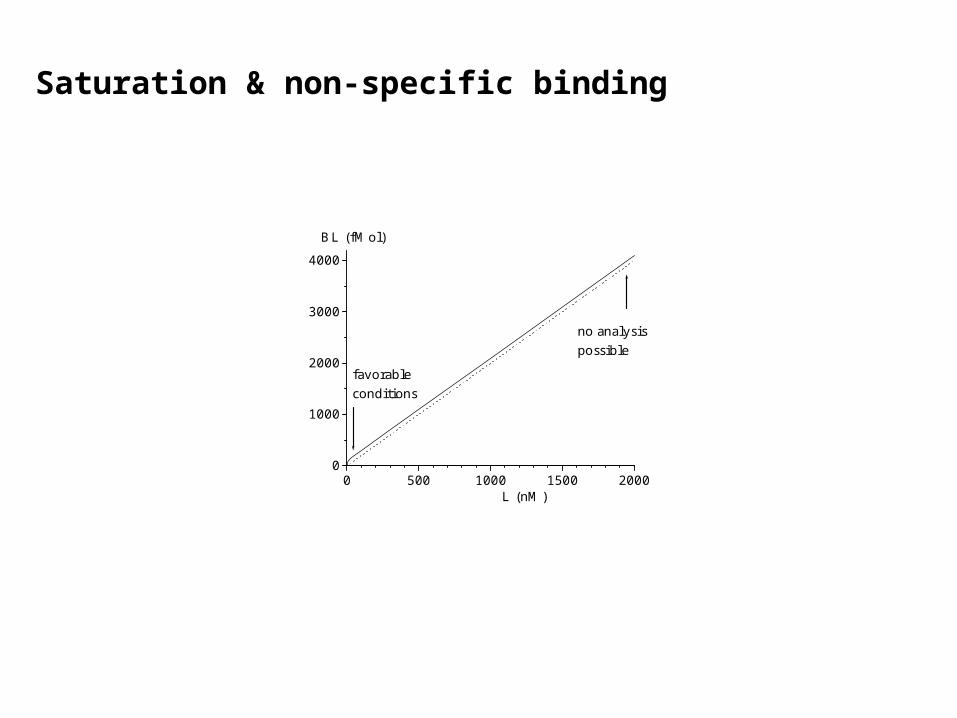
0 500 1000 1500 20000
1000
2000
3000
4000
favorableconditions
no analysispossible
To Save this template,C hooseFile:Template:Template Save.
L (nM)
BL (fMol)
Saturation & non-specific binding

0 500 1000 1500 20000
1000
2000
3000
4000
favorableconditions
no analysispossible
To Save this template,C hooseFile:Template:Template Save.
L (nM)
BL (fMol)
At low nM concentrations,most of the radioligand Lis bound to saturablehigh affinity sites.
Saturation & non-specific binding

0 500 1000 1500 20000
1000
2000
3000
4000
favorableconditions
no analysispossible
To Save this template,C hooseFile:Template:Template Save.
L (nM)
BL (fMol)
At low nM concentrations,most of the radioligand Lis bound to saturablehigh affinity sites.
At high concentrations,the linearly rising
non-specific bindingwill dominate, and
specific bindingcan no longerbe detected.
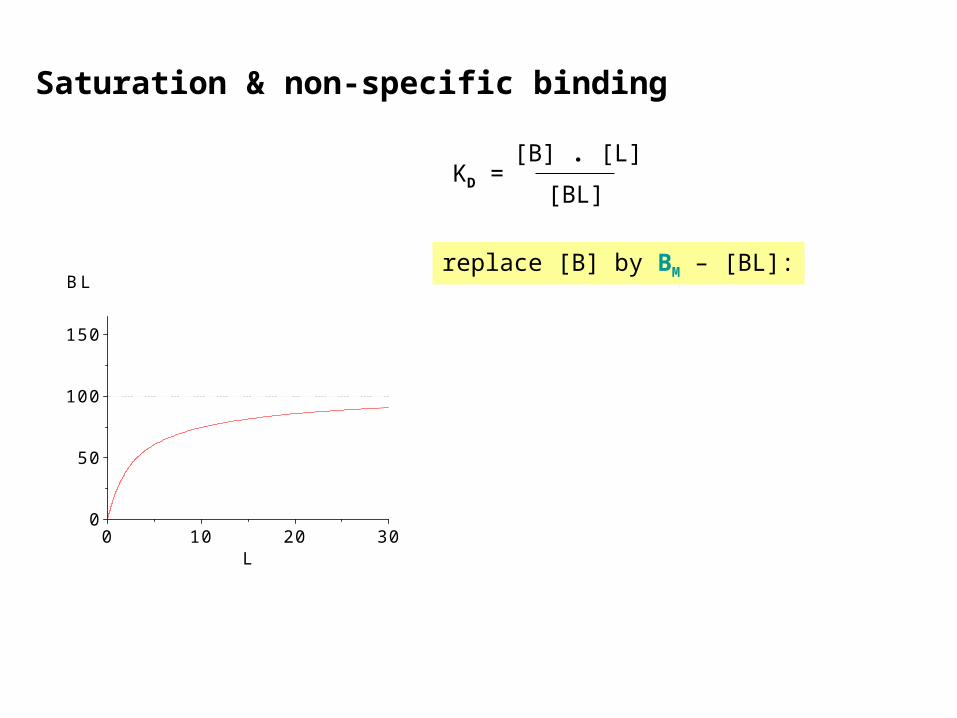
Saturation & non-specific binding

BL B + L KD =[BL]
[B] . [L]
KD: dissociation equilibrium constant
Saturation & non-specific binding
With increasing [L] more binding sites are occupied (BL) and free sites (B) are lost. The sum
BL + B = BM
remains constant.0 10 20 300
50
100
150
BL
L
Saturation & non-specific binding
KD =[BL]
[B] . [L]
replace [B] by BM – [BL]:
0 10 20 300
50
100
150
BL
L
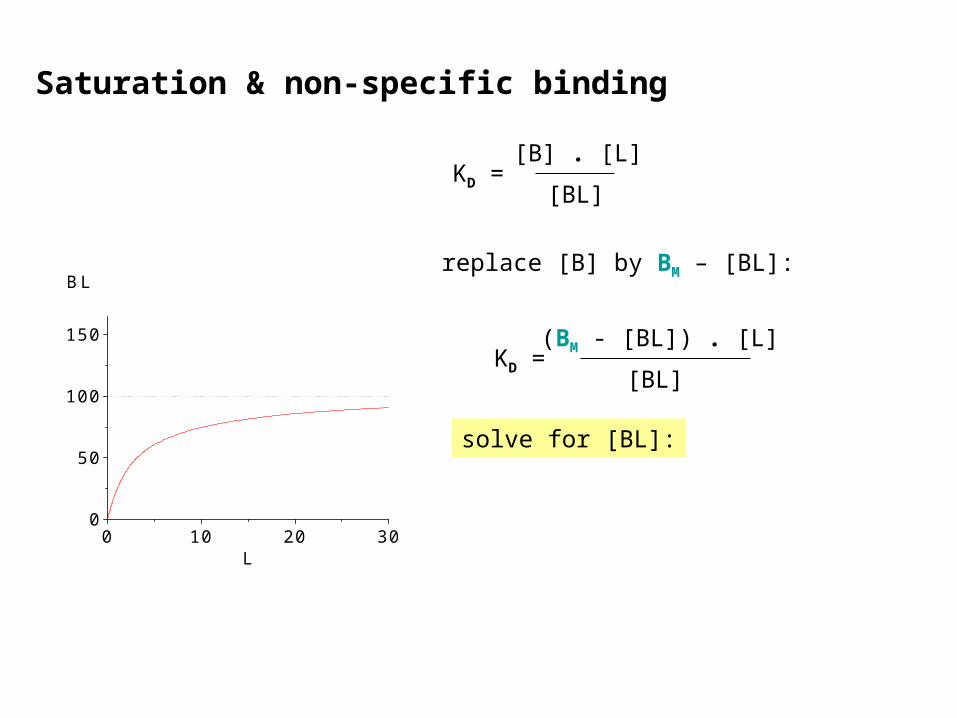
Saturation & non-specific binding
KD =[BL]
[B] . [L]
replace [B] by BM – [BL]:
KD =[BL]
(BM - [BL]) . [L]
solve for [BL]:
0 10 20 300
50
100
150
BL
L

Saturation & non-specific binding
KD =[BL]
[B] . [L]
replace [B] by BM – [BL]:
KD =[BL]
(BM - [BL]) . [L]
solve for [BL]:
0 10 20 300
50
100
150
BL
L
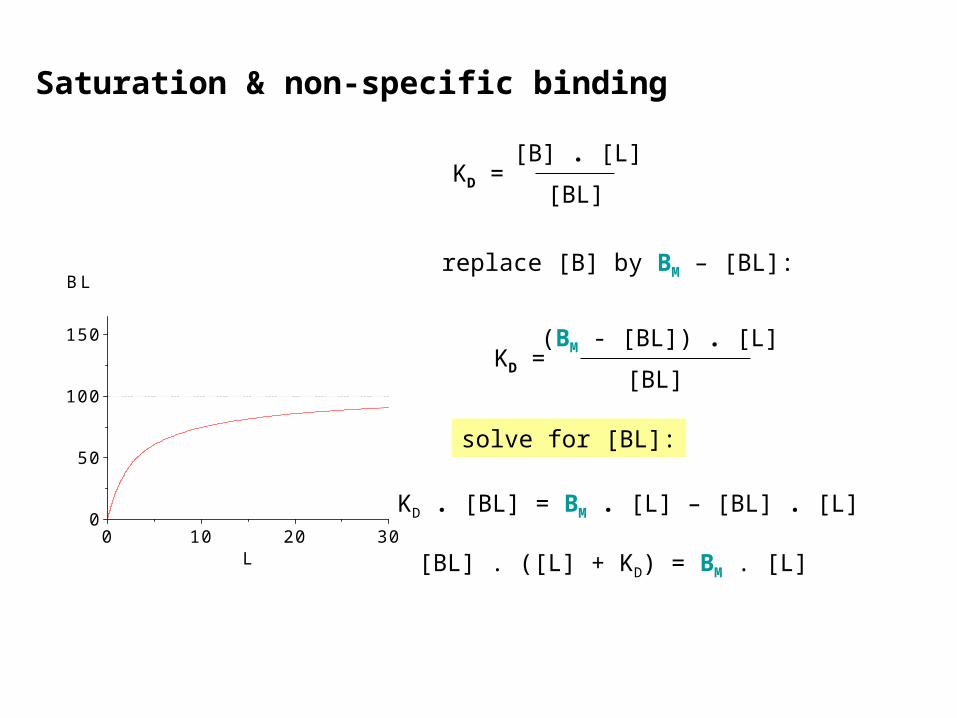
KD . [BL] = BM . [L] – [BL] . [L]
Saturation & non-specific binding
KD =[BL]
[B] . [L]
replace [B] by BM – [BL]:
KD =[BL]
(BM - [BL]) . [L]
solve for [BL]:
0 10 20 300
50
100
150
BL
L
KD . [BL] = BM . [L] – [BL] . [L]
[BL] . ([L] + KD) = BM . [L]

Saturation & non-specific binding
KD =[BL]
[B] . [L]
replace [B] by BM – [BL]:
KD =[BL]
(BM - [BL]) . [L]
solve for [BL]:
[BL] = BM .[L]
[L] + KD Langmuir isotherm
0 10 20 300
50
100
150
BL
L
KD . [BL] = BM . [L] – [BL] . [L]
[BL] . ([L] + KD) = BM . [L]

Saturation & non-specific binding
KD =[BL]
[B] . [L]
replace [B] by BM – [BL]:
KD =[BL]
(BM - [BL]) . [L]
solve for [BL]:
[BL] = BM .[L]
[L] + KD Langmuir isotherm
0 10 20 300
50
100
150
BL
L
KD . [BL] = BM . [L] – [BL] . [L]
[BL] . ([L] + KD) = BM . [L]
Irving Langmuir1881-1957
Nobel price 1932

Saturation & non-specific binding
KD =[BL]
[B] . [L]
replace [B] by BM – [BL]:
KD =[BL]
(BM - [BL]) . [L] wrong!
0 10 20 300
50
100
150
BL
L
solve for [BL]:
[BL] = BM .[L]
[L] + KD Langmuir isotherm
KD . [BL] = BM . [L] – [BL] . [L]
[BL] . ([L] + KD) = BM . [L]
Irving Langmuir1881-1957
Nobel price 1932

Saturation & non-specific binding
KD =[BL]
[B] . [L]
replace [B] by BM – [BL]:
KD =[BL]
(BM - [BL]) . (Lo – [BL]) correct
0 10 20 300
50
100
150
BL
L

Saturation & non-specific binding
KD =[BL]
[B] . [L]
replace [B] by BM – [BL]:
KD =[BL]
(BM - [BL]) . (Lo – [BL])
solve for [BL]:
KD . [BL] = BM . Lo – BM . [BL] – [BL] . Lo + [BL]2
0 10 20 300
50
100
150
BL
L
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !

Saturation & non-specific binding
KD =[BL]
[B] . [L]
replace [B] by BM – [BL]:
KD =[BL]
(BM - [BL]) . (Lo – [BL])
solve for [BL]:
KD . [BL] = BM . Lo – BM . [BL] – [BL] . Lo + [BL]2
0 10 20 300
50
100
150
BL
L
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
[BL]2 – [BL] . (BM + Lo + KD) + BM . Lo = 0

Saturation & non-specific binding
KD =[BL]
[B] . [L]
replace [B] by BM – [BL]:
KD =[BL]
(BM - [BL]) . (Lo – [BL])
solve for [BL]:
0 10 20 300
50
100
150
BL
L
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
Sweet memories…
KD . [BL] = BM . Lo – BM . [BL] – [BL] . Lo + [BL]2
[BL]2 – [BL] . (BM + Lo + KD) + BM . Lo = 0

Saturation & non-specific binding
KD =[BL]
[B] . [L]
replace [B] by BM – [BL]:
KD =[BL]
(BM - [BL]) . (Lo – [BL])
solve for [BL]:
[BL] = ½ . {BM + Lo + KD - [(BM + Lo + KD)2 – 4 . BM . Lo]½}
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
0 10 20 300
50
100
150
BL
L
Sweet memories…
KD . [BL] = BM . Lo – BM . [BL] – [BL] . Lo + [BL]2
[BL]2 – [BL] . (BM + Lo + KD) + BM . Lo = 0

Saturation & non-specific binding
KD =[BL]
[B] . [L]
replace [B] by BM – [BL]:
KD =[BL]
(BM - [BL]) . (Lo – [BL])
solve for [BL]:
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
In this case, quantities Lo and KD are not entered as concentrations, but as moles in the respective volume chosen, in the same units as BM.
0 10 20 300
50
100
150
BL
L
Sweet memories…
[BL] = ½ . {BM + Lo + KD - [(BM + Lo + KD)2 – 4 . BM . Lo]½}
KD . [BL] = BM . Lo – BM . [BL] – [BL] . Lo + [BL]2
[BL]2 – [BL] . (BM + Lo + KD) + BM . Lo = 0

Saturation & non-specific binding
KD =[BL]
[B] . [L]
replace [B] by BM – [BL]:
KD =[BL]
(BM - [BL]) . (Lo – [BL])
solve for [BL]:
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
In this case, quantities Lo and KD are not entered as concentrations, but as moles in the respective volume chosen, in the same units as BM.
0 10 20 300
50
100
150
BL
L
3 times more ligand than receptors at KD concentration (8% loss)
Sweet memories…
[BL] = ½ . {BM + Lo + KD - [(BM + Lo + KD)2 – 4 . BM . Lo]½}
KD . [BL] = BM . Lo – BM . [BL] – [BL] . Lo + [BL]2
[BL]2 – [BL] . (BM + Lo + KD) + BM . Lo = 0

Saturation & non-specific binding
KD =[BL]
[B] . [L]
replace [B] by BM – [BL]:
KD =[BL]
(BM - [BL]) . (Lo – [BL])
solve for [BL]:
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
! attention ℮ ∫ ∑ mathematics ∂ ∞ √ % attention !
In this case, quantities Lo and KD are not entered as concentrations, but as moles in the respective volume chosen, in the same units as BM.
0 10 20 300
50
100
150
BL
L
3 times more receptor than ligand at KD concentration (57% loss)
Sweet memories…
[BL] = ½ . {BM + Lo + KD - [(BM + Lo + KD)2 – 4 . BM . Lo]½}
KD . [BL] = BM . Lo – BM . [BL] – [BL] . Lo + [BL]2
[BL]2 – [BL] . (BM + Lo + KD) + BM . Lo = 0

Saturation & non-specific binding
A realistic saturation function is a composite of 2 simultaneous processes:
0 10 20 300
50
100
150
*
1
BL
L
1: non-specific binding
It is sufficient to measure 2 points; extrapolation of L 0 results in the blank of the measuring method (*).

Saturation & non-specific binding
A realistic saturation function is a composite of 2 simultaneous processes:
1: non-specific binding
It is sufficient to measure 2 points; extrapolation of L 0 results in the
blank of the measuring method (*).
0 10 20 300
50
100
1502
*
1
BL
L
2: specific binding
... Is sitting on the non-specific binding, obtained as difference between total and non-specific binding.

Saturation & non-specific binding
A realistic saturation function is a composite of 2 simultaneous processes:
1: non-specific binding
It is sufficient to measure 2 points; extrapolation of L 0 results in the
blank of the measuring method (*).2: specific binding
... Is sitting on the non-specific binding, obtained as difference between total and non-specific binding (†).
0 10 20 300
50
100
150
†
2
*
1
BL
L
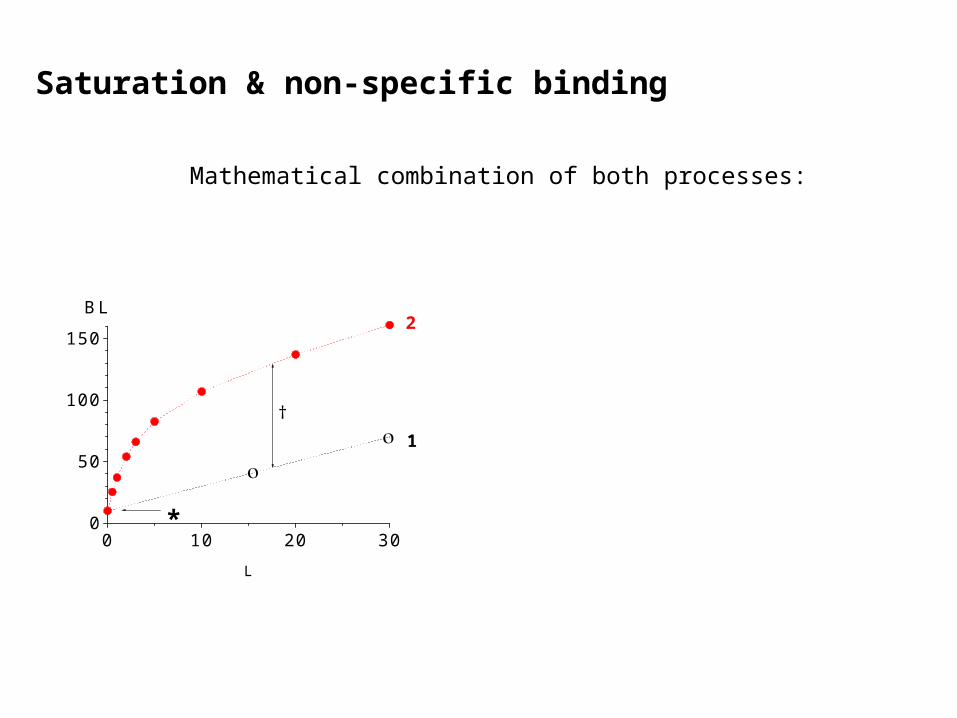
Saturation & non-specific binding
0 10 20 300
50
100
150
†
2
*
1
BL
L
Mathematical combination of both processes:

Saturation & non-specific binding
0 10 20 300
50
100
150
†
2
*
1
BL
L
Mathematical combination of both processes:
1: non-specific binding
[BL] = BU .[L]
[L] + KU

2: specific binding
Saturation & non-specific binding
0 10 20 300
50
100
150
†
2
*
1
BL
L
Mathematical combination of both processes:
1: non-specific binding
[BL] = BU .[L]
[L] + KU
[BL] = BS .[L]
[L] + KS

2: specific binding
Saturation & non-specific binding
0 10 20 300
50
100
150
†
2
*
1
BL
L
Mathematical combination of both processes:
1: non-specific binding
[BL] = BU .[L]
[L] + KU
[BL] = BS .[L]
[L] + KS
KU (~mM) >> Ks (nM)

2: specific binding
Saturation & non-specific binding
0 10 20 300
50
100
150
†
2
*
1
BL
L
Mathematical combination of both processes:
1: non-specific binding
[BL] = BU .[L]
[L] + KU
[BL] = BS .[L]
[L] + KS
KU (~mM) >> Ks (nM)
At reasonable ligand concentrations, [L] + KU ~ KU and non-specific binding is a linear function of [L]:
[BL] = BS .[L]
[L] + KS
BU
KU
. [L]+
-9 -8 -7 -6 -50
50
100
NB
BL
log[I]
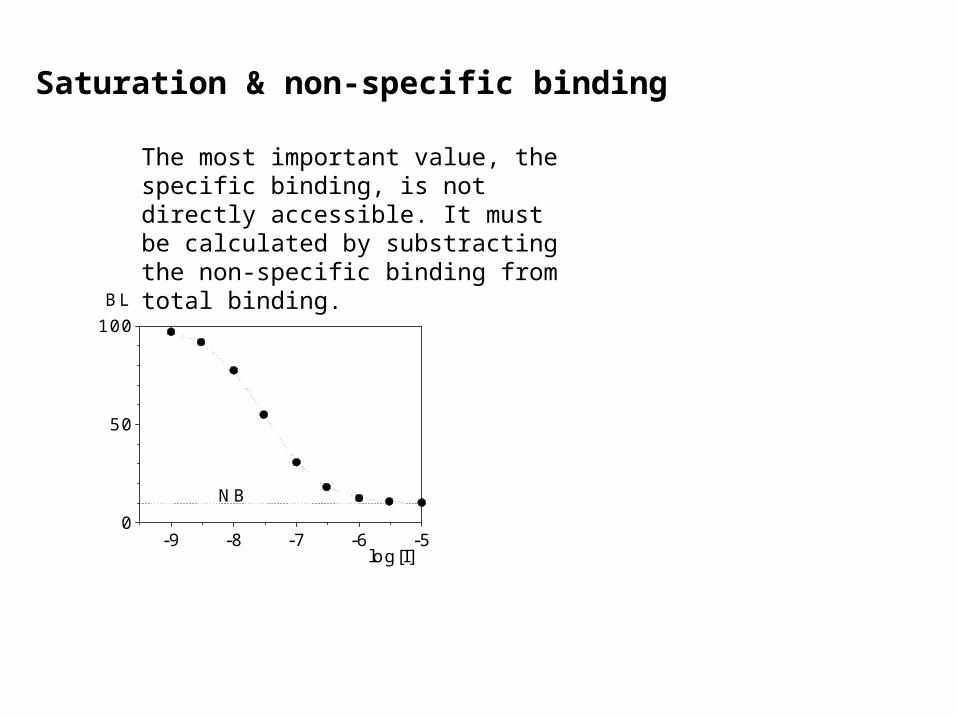
Saturation & non-specific binding
The most important value, the specific binding, is not directly accessible. It must be calculated by substracting the non-specific binding from total binding.
-9 -8 -7 -6 -50
50
100
NB
BL
log[I]
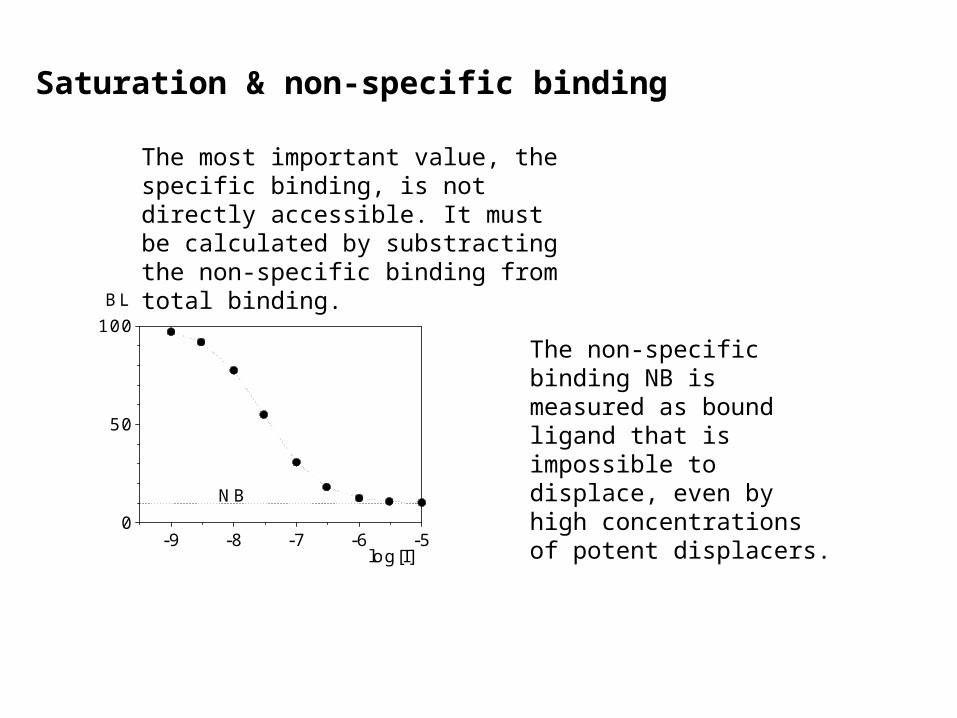
Saturation & non-specific binding
The most important value, the specific binding, is not directly accessible. It must be calculated by substracting the non-specific binding from total binding.
The non-specific binding NB is measured as bound ligand that is impossible to displace, even by high concentrations of potent displacers.

Saturation & non-specific binding
Strategies to keep non-specific binding low:

Saturation & non-specific binding
Strategies to keep non-specific binding low:
• choose a biological source with a high density of high-affinity binding sites

Saturation & non-specific binding
Strategies to keep non-specific binding low:
• choose a biological source with a high density of high-affinity binding sites
• select a radioligand concentration around the expected KD ( a few hundred to a few thousand dpm will be sufficient as result)

Saturation & non-specific binding
Strategies to keep non-specific binding low:
• choose a biological source with a high density of high-affinity binding sites
• select a radioligand concentration around the expected KD ( a few hundred to a few thousand dpm will be sufficient as result)
• use a clean radioligand; if necessary, any radioligand can be purified easily by thin layer chromatography

Saturation & non-specific binding
Strategies to keep non-specific binding low:
• choose a biological source with a high density of high-affinity binding sites
• select a radioligand concentration around the expected KD ( a few hundred to a few thousand dpm will be sufficient as result)
• use a clean radioligand; if necessary, any radioligand can be purified easily by thin layer chromatography
• If you filter your samples and if you use a radioligand with an amino group, pre-treat the glass fiber filters with polyethylene imine

Saturation & non-specific binding
Strategies to keep non-specific binding low:
• choose a biological source with a high density of high-affinity binding sites
• select a radioligand concentration around the expected KD ( a few hundred to a few thousand dpm will be sufficient as result)
• use a clean radioligand; if necessary, any radioligand can be purified easily by thin layer chromatography
• If you filter your samples and if you use a radioligand with an amino group, pre-treat the glass fiber filters with polyethylene imine
• optimise the rinsing procedure of pellets and filters, respectively

Classification of glutamate receptors
ionotropic receptors metabotropic receptors
Radioligands forexcitatory amino acid (EAA) receptors

Radioligands forexcitatory amino acid (EAA) receptors
Classification of glutamate receptors
ionotropic receptors metabotropic receptors
NMDAreceptors
non-NMDAreceptors
Group I Group II Group III

Radioligands forexcitatory amino acid (EAA) receptors
Classification of glutamate receptors
ionotropic receptors metabotropic receptors
NMDAreceptors
non-NMDAreceptors
AMPAreceptors
kainatereceptors
Group I Group II Group III

Radioligands forexcitatory amino acid (EAA) receptors
Classification of glutamate receptors
ionotropic receptors
NMDAreceptors
non-NMDAreceptors
AMPAreceptors
kainatereceptors
Schmid et al (2009) PNAS 106:10320
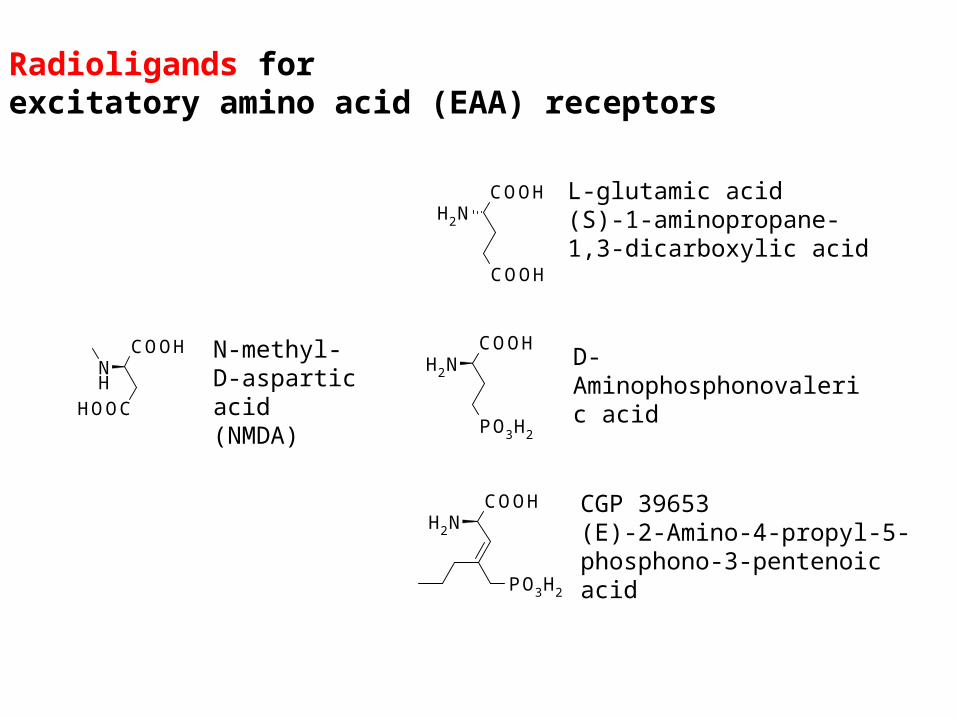
Radioligands forexcitatory amino acid (EAA) receptors
COOH
COOH
NH2
L-glutamic acid(S)-1-aminopropane-1,3-dicarboxylic acid
COOH
PO3H2
NH2
COOH
PO3H2
NH2D-Aminophosphonovaleric acid
CGP 39653(E)-2-Amino-4-propyl-5-phosphono-3-pentenoic acid
COOHNH
HOOC
N-methyl-D-aspartic acid (NMDA)

Mg2+
Mg2+
-70 mV
Na+
Ca2+
Mg2+
Mg2+
outside
membrane
inside
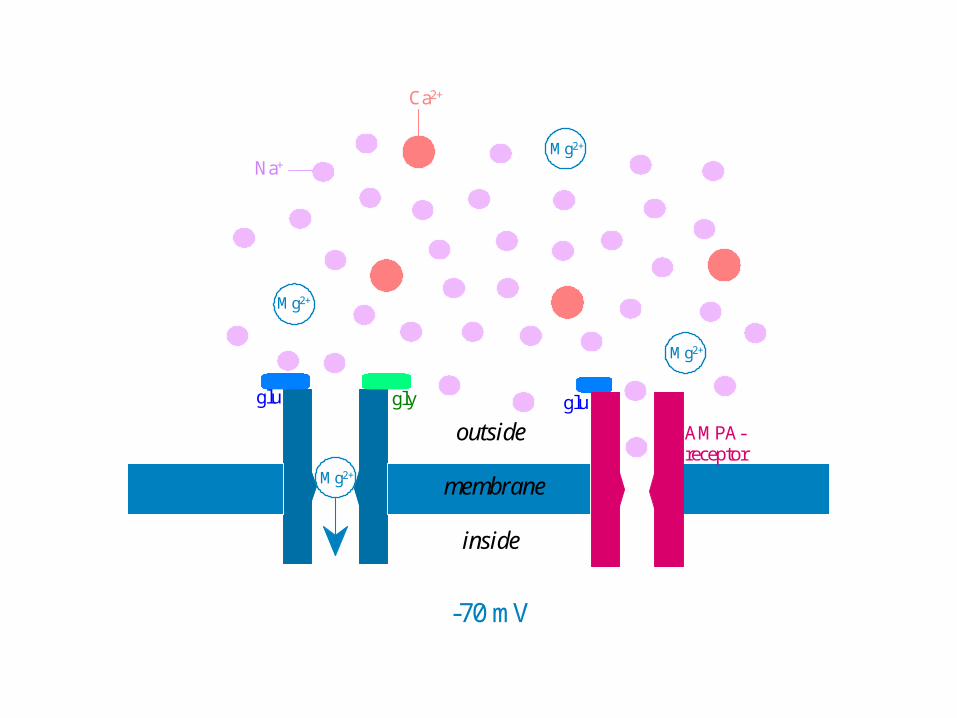
Mg2+
Mg2+
-70 mV
Na+
Ca2+
Mg2+
Mg2+
outside
membrane
inside
glu gly glu
AMPA-receptor

Mg2+
-30 mV
Na+
Ca2+
Mg2+
Mg2+
outside
membrane
inside
glu gly gluMg2+

-10 mV
Na+
Ca2+
Mg2+
Mg2+
outside
membrane
inside
glu gly glu
Mg2+
Mg2+

0 mV
Na+
Ca2+
Mg2+
Mg2+
outside
membrane
inside
glu gly glu
Mg2+
Mg2+
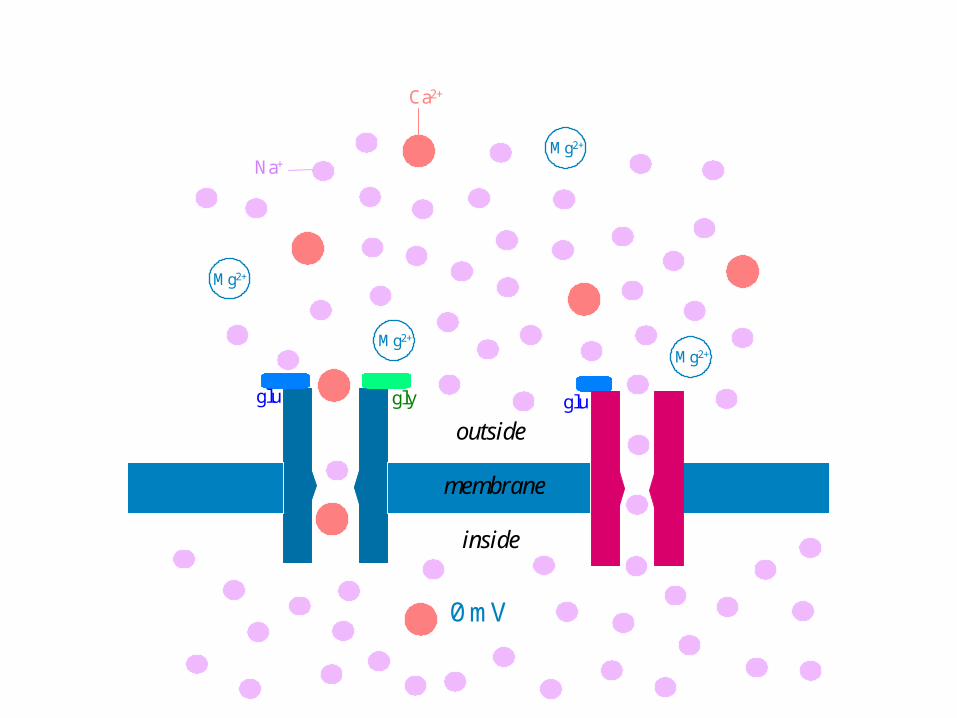
0 mV
Na+
Ca2+
Mg2+
Mg2+
outside
membrane
inside
glu gly glu
Mg2+
Mg2+
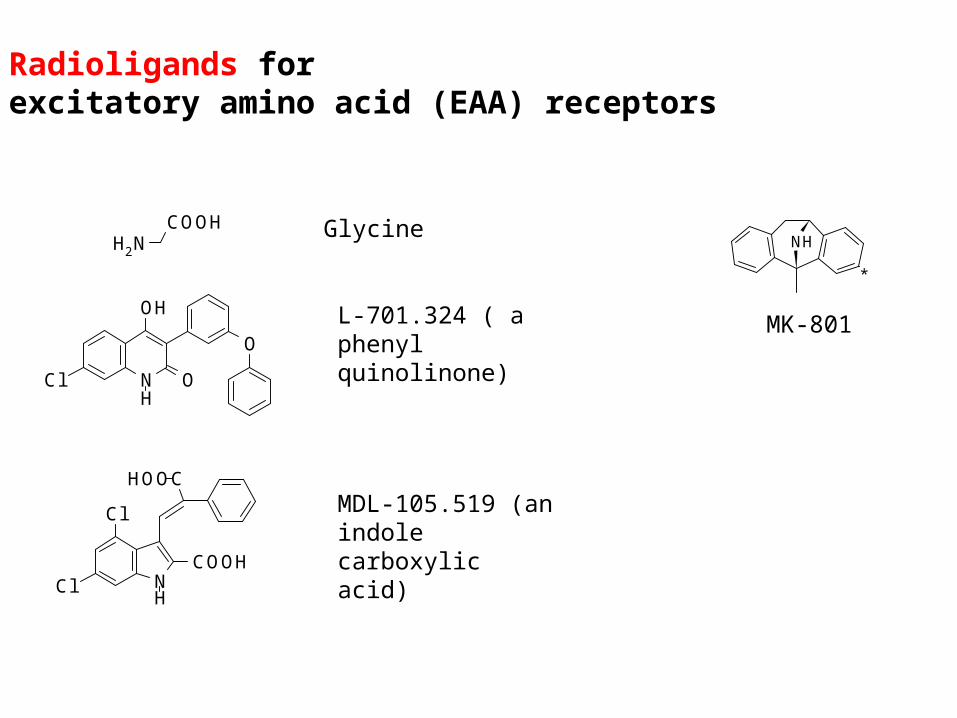
Radioligands forexcitatory amino acid (EAA) receptors
COOHNH2
Glycine
NH
O
OH
O
Cl
L-701.324 ( a phenyl quinolinone)
NCOOH
CHOO
H
Cl
Cl
MDL-105.519 (an indole carboxylic acid)
MK-801
NH
*

Radioligands forexcitatory amino acid (EAA) receptors
COOHNH2
Glycine
NH
O
OH
O
Cl
L-701.324 ( a phenyl quinolinone)
NCOOH
CHOO
H
Cl
Cl
MDL-105.519 (an indole carboxylic acid)
MK-801
NH
*
N
O
N*
[3H]GSK-931.145radioligand for the glycine transporter GlyT-1
(Herdon et al 2010 Neuropharmacol 59:558)
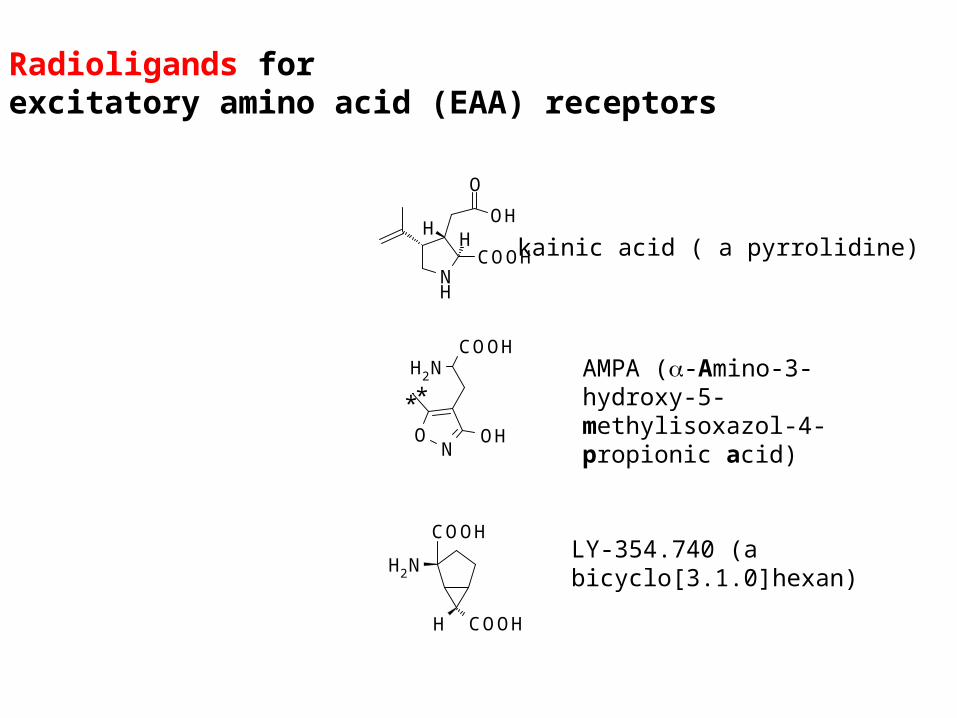
Radioligands forexcitatory amino acid (EAA) receptors
OH
O
NH
COOHH
Hkainic acid ( a pyrrolidine)
COOHNH2
ON
OH
**
AMPA (-Amino-3-hydroxy-5-methylisoxazol-4-propionic acid)
H COOH
COOH
NH2
LY-354.740 (a bicyclo[3.1.0]hexan)

Radioligands forexcitatory amino acid (EAA) receptors
OH
O
NH
COOHH
Hkainic acid ( a pyrrolidine)
COOHNH2
ON
OH
**
AMPA (-Amino-3-hydroxy-5-methylisoxazol-4-propionic acid)
H COOH
COOH
NH2
LY-354.740 (a bicyclo[3.1.0]hexan)
Grant et al (2010) Neurotox Terat 32:132
HOOC
H
OH
O
NH
COOHH
H

Radioligands forexcitatory amino acid (EAA) receptors
OH
O
NH
COOHH
Hkainic acid ( a pyrrolidine)
COOHNH2
ON
OH
**
AMPA (-Amino-3-hydroxy-5-methylisoxazol-4-propionic acid)
H COOH
COOH
NH2
LY-354.740 (a bicyclo[3.1.0]hexan)
NO
COOH
OH
NH2
NO OH
NH2
Muscimol Ibotensäure

Radioligands forexcitatory amino acid (EAA) receptors
OH
O
NH
COOHH
Hkainic acid ( a pyrrolidine)
COOHNH2
ON
OH
**
AMPA (-Amino-3-hydroxy-5-methylisoxazol-4-propionic acid)
H COOH
COOH
NH2
LY-354.740 (a bicyclo[3.1.0]hexan)O
H COOH
COOH
NH2S
H COOH
COOH
NH2
O
O
LY-404.039 LY-379.268

The most important binding techniques
... are all non-equilibrium techniques for particulate receptor preparations:
• Centrifugation• Filtration over glass fiber filters• Slice autoradiography
B + L* BL*

The most important binding techniques
... are all non-equilibrium techniques for particulate receptor preparations:
• Centrifugation• Filtration over glass fiber filters• Slice autoradiography
B + L* BL*
... applied to weak ligands (KD > 20 nM) ● you need a high speed refrigerated certrifuge ● plastic vials must support 40 000 x g ● after centrifugation, pellet and inner wall needs rinsing ● scintillation cocktail added directly to the rinsed incubation vials.

The most important binding techniques
... are all non-equilibrium techniques for particulate receptor preparations:
• Centrifugation• Filtration over glass fiber filters• Slice autoradiography
B + L* BL*
... Can only be applied to high affinity ligands (KD < 20 nM) ● you need a vacuum filter box

The most important binding techniques
... are all non-equilibrium techniques for particulate receptor preparations:
• Centrifugation• Filtration over glass fiber filters• Slice autoradiography
B + L* BL*
... Can only be applied to high affinity ligands (KD < 20 nM) ● you need a vacuum filter box or better a harvester ● for radioligands with amino group, the glass fiber filter must be soaked in 0.3% polyethylenimine ●

The most important binding techniques
... are all non-equilibrium techniques for particulate receptor preparations:
• Centrifugation• Filtration over glass fiber filters• Slice autoradiography
B + L* BL*
... Can only be applied to high affinity ligands (KD < 20 nM) ● you need a vacuum filter box or better a harvester ● for radioligands with amino group, the glass fiber filter must be soaked in 0.3% polyethylenimine ● for best results, filter should be shaken in scintillation cocktail for 30 min.
L* L*L* L*
scintillator

The most important binding techniques
... are all non-equilibrium techniques for particulate receptor preparations:
• Centrifugation• Filtration over glass fiber filters• Slice autoradiography
B + L* BL*
glass
coating
L*
L* L*L*
L*
L*
L*
L*
L*
L*
L*L*
... applied to frozen slices prepared in a cryostat / microtom (10-20 µm) ● tissue must be shock-frozen (-40 °C) in dry ice / isopentane ● slices taken up to coated glass slides ● for incubation, you can use..

The most important binding techniques
... are all non-equilibrium techniques for particulate receptor preparations:
• Centrifugation• Filtration over glass fiber filters• Slice autoradiography
B + L* BL*
... applied to frozen slices prepared in a cryostat / microtom (10-20 µm) ● tissue must be shock-frozen (-40 °C) in dry ice / isopentane ● slices taken up to coated glass slides ● for incubation, you can use a jar or...

The most important binding techniques
... are all non-equilibrium techniques for particulate receptor preparations:
• Centrifugation• Filtration over glass fiber filters• Slice autoradiography
B + L* BL*
... applied to frozen slices prepared in a cryostat / microtom (10-20 µm) ● tissue must be shock-frozen (-40 °C) in dry ice / isopentane ● slices taken up to coated glass slides ● for incubation, you can use a jar or simply a droplet on the slide ● expose dried slices to film or phosphoscreen ● evaluation by co-exposure of stripes containing known amounts of radioactivity.


















