
Journal of Biomaterials Applications
27
http://jba.sagepub.com/ Applications Journal of Biomaterials http://jba.sagepub.com/content/25/7/743 The online version of this article can be found at: DOI: 10.1177/0885328209360425 2011 25: 743 originally published online 17 March 2010 J Biomater Appl R. Nieto-Aguilar, D. Serrato, I. Garzón, A. Campos and M. Alaminos Cells in a Novel Fibrin-agarose Scaffold Pluripotential Differentiation Capability of Human Adipose-derived Stem Published by: http://www.sagepublications.com can be found at: Journal of Biomaterials Applications Additional services and information for http://jba.sagepub.com/cgi/alerts Email Alerts: http://jba.sagepub.com/subscriptions Subscriptions: http://www.sagepub.com/journalsReprints.nav Reprints: http://www.sagepub.com/journalsPermissions.nav Permissions: http://jba.sagepub.com/content/25/7/743.refs.html Citations: What is This? - Mar 17, 2010 OnlineFirst Version of Record - Apr 1, 2011 Version of Record >> at Biblioteca Universitaria de Granada on November 20, 2012 jba.sagepub.com Downloaded from
Transcript of Journal of Biomaterials Applications

http://jba.sagepub.com/Applications
Journal of Biomaterials
http://jba.sagepub.com/content/25/7/743The online version of this article can be found at:
DOI: 10.1177/0885328209360425
2011 25: 743 originally published online 17 March 2010J Biomater ApplR. Nieto-Aguilar, D. Serrato, I. Garzón, A. Campos and M. Alaminos
Cells in a Novel Fibrin-agarose ScaffoldPluripotential Differentiation Capability of Human Adipose-derived Stem
Published by:
http://www.sagepublications.com
can be found at:Journal of Biomaterials ApplicationsAdditional services and information for
http://jba.sagepub.com/cgi/alertsEmail Alerts:
http://jba.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
http://jba.sagepub.com/content/25/7/743.refs.htmlCitations:
What is This?
- Mar 17, 2010 OnlineFirst Version of Record
- Apr 1, 2011Version of Record >>
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

Pluripotential DifferentiationCapability of Human
Adipose-derived Stem Cells in aNovel Fibrin-agarose Scaffold
R. NIETO-AGUILAR, D. SERRATO, I. GARZON, A. CAMPOS
AND M. ALAMINOS*Faculty of Medicine, Department of Histology (Tissue Engineering Group)
University of Granada, Avenida de Madrid 11, E18012 Granada, Spain
ABSTRACT: The potentiality of adipose-derived stem cells (ASCs) cultured on2D systems has been previously established. Nevertheless, very little is known sofar about the differentiation potentiality of ASCs in 3D culture systems usingbiomaterials. In this work, we have evaluated the transdifferentiation capabilitiesof ASCs cultured within a novel fibrin-agarose biomaterial by histologicalanalysis, histochemistry and immunofluorescence. Our results showed that 3Dfibrin-agarose biomaterial is highly biocompatible and supports the transdiffer-entiation capabilities of ASCs to the osteogenic, chondrogenic, adipogenic, andneurogenic lineages.
KEY WORDS: fibrin-agarose, biomaterials, scaffolds, adipose-derived stromalcells, transdifferentiation.
INTRODUCTION
Management of the different conditions that affect the skeletaland neural tissues usually has several anatomic, physiological,
and aesthetic implications. The demand of corrective procedures withthese tissues increases every year due to congenital or acquired diseases,
*Author to whom correspondence should be addressed. E-mail: [email protected] 1–5 appear in color online: http://jba.sagepub.com
JOURNAL OF BIOMATERIALS APPLICATIONS Vol. 25 — March 2011 743
0885-3282/11/07 0743–26 $10.00/0 DOI: 10.1177/0885328209360425� The Author(s), 2010. Reprints and permissions:http://www.sagepub.co.uk/journalsPermissions.nav
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

including trauma, tumors, infections, and cosmetic corrections [1,2].In most of the cases, surgical treatment of the above mentionedpathologies requires the use of autologous or heterologous tissue grafts[3]. In general, these procedures are associated to different degrees oforganic, metabolic and immune complications such as graft rejection,infection, dehiscence, etc. [4], which can impair the results of thesurgical procedure.
In this context, novel techniques have focused on the efficientgeneration of autologous artificial tissues and organs developed bytissue engineering. Due to its autologous origin, these bioengineeredhuman tissues would not be submitted to the possibility of immunerejection once implanted in the host patient [5]. The construction ofartificial organs by tissue engineering is one of the research fields thathave experienced major progress during recent years [6]. By using tissueengineering techniques and biomaterials, different researchers havedeveloped well-organized artificial substitutes of different organs andtissues for therapeutic use, including, among others, human skin [7],cornea [8,9], bone [10], blood vessels [11], and oral mucosa [12,13].However, autologous generation of these tissues in the laboratory ishighly dependent on the availability of a proper source of adult cells thatare usually obtained from tissue biopsies from the same origin [12–15].
Ideally, for the generation of complex structures consisting on severaltissue types, it should be possible to obtain all these tissue types from asingle and accessible cell source. In this regard, one of the mostpromising sources of human adult stem cells is the adipose tissue, whichis very rich in stromal adipose-derived stem cells (ASCs) with highproliferative and differentiation capabilities [16,17]. So far, ASCs havebeen used for several ex vivo applications, including some of potentialusefulness in regenerative medicine such as the generation ofbioengineered bone [18,19], cartilage [20], neurons [21], and adiposetissue [22]. The clinical need of new sources of human tissues suitablefor skeletal, connective and neuronal reconstruction and repair supportsfurther research of novel methods and techniques based on tissueengineering and cell therapy.
In general, bioengineered tissues consist of 3D structures and stemcells that are grown on different biomaterials used as scaffolds. In thissense, we recently developed a novel biomaterial consisting in a mixtureof human fibrin and agarose [12–14]. Although several types of epithelialand connective cells isolated from the human cornea and oral mucosahave shown proper attachment and differentiation on this biomaterial,the influence of fibrin-agarose matrices on the multilineage differentia-tion capabilities of human ASCs have not been explored to the date.
744 R. NIETO-AGUILAR ET AL.
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

In this work, we have efficiently isolated and cultured ASCs in a 2Dculture system as well as immersed in a 3D scaffold of fibrin and agaroseand we have induced them to differentiate to adult cells of bone, cartilage,adipose, and neuron tissue phenotypes. Then, we have carried out ahistological and genetic study of the differentiated cells in order to estimatethe cell changes that take place during the differentiation process.
MATERIALS AND METHODS
Human Tissue Samples
In this work, biopsies of complete thickness of skin were obtained underlocal anesthesia. Average size of the samples used were approximately3� 2.5� 2 cm3. All tissues corresponded to healthy donors who weresubmitted to minor aesthetic surgical procedures at Division of PlasticSurgery of the University Hospital Virgen de las Nieves of Granada. Rightafter the excision, all tissues were submerged in Dulbecco’s modifiedEagle’s medium (DMEM) (Sigma-Aldrich Inc. St. Luis, MO, USA)supplemented with antibiotics and antimycotics (100 U/mL of penicillinG, 100 mg/mL of streptomycin and 0.25 mg/mL of B amphotericin) (Sigma-Aldrich) and stored at 48C until the moment of use.
As controls, normal human tissues corresponding to bone, cartilage,adipose tissue, and brain were used.
All donors provided their written consent for the use of the tissues inthis work, and this research was approved by the local Research andEthics committees at the University of Granada (Spain).
Generation of Primary Cultures of ASCs
On arriving to the laboratory, samples were washed in phosphatebuffered saline (PBS) and the adipose tissue was mechanically isolatedand fragmented into small pieces. Then, the tissue explants wereenzymatically digested in a 3% type I collagenase solution (Gibco BRLLife Technologies, Karlsruhe, Germany) for 8 h at 378C. Finally, isolatedstem cells were harvested by centrifugation and cultured in DMEMsupplemented with 10% fetal bovine serum (Sigma-Aldrich) andantibiotics-antimycotics (100 U/mL of penicillin G, 100 mg/mL of strep-tomycin and 0.25 mg/mL of B amphotericin) to generate 2D ASCs cellcultures (2D-ACC).
The culture medium was renewed every 3 days. Once the cells reachedsubconfluence, the cultures were trypsinized and subcultured on culturechamber slides (NuncTM, Roskilde, Denmark). All cells used in this work
Pluripotential Differentiation Capability of ASCs 745
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

belonged to the first four passages. As controls human oral mucosafibroblasts were cultured on same culture chamber slides.
Generation of Fibrin-agarose 3D Hydro-gels with ASCsImmersed Within (3D-FAH)
Once cultured ASCs reached subconfluence, the cells were trypsinizedand 175,000 cells per mL were seeded on fibrin-agarose scaffolds usingTranswell� permeable supports (Corning incorporated, NY, USA)following previously published protocols [12–14]. In short, the 3Dstromal substitute was generated by using a mixture of human plasmaand 0.1% of type VII agarose with tranexamic acid and calcium chloride.Once the hydrogel polymerized, the artificial constructs were suppliedwith basal medium.
As controls, fibrin-agarose scaffolds were seeded with 175,000 humanoral mucosa fibroblasts per mL.
Induction of Transdifferentiation to the Osteogenic,Chondrogenic, Adipogenic, and Neurogenic Lineagesin 2D-ACC and 3D-FAH
In order to induce both 2D-ACC and 3D-FAH cell cultures to theosteogenic, chondrogenic, adipogenic, and neurogenic phenotypes,samples were cultured in different inductive media. All these mediaconsisted of a basal DMEM medium 10% of FBS and antibiotic-antimycotics that was supplemented with different growth factors andinductive reagents.
For the induction to the osteoblastic lineage, the basal medium wassupplemented with 100 nM of dexametasone, 10 mM b-glycerol phos-phate and of 50 mM of L-ascorbic acid. For condrogenic induction, weadded 40 mg/mL L-proline, 100 mg/mL piruvate, 10 ng/mL TGF-b, 50mML-ascorbic acid and insulin-transferrin-selenium (ITS) 5 mg/mL. For theadipogenic differentiation, the basal medium was supplementedwith 1mM dexametasone, 0.5 mM 3-Isobutyl-1-methylxanthine and ITS5 mg/mL. Finally, for the neurogenic induction, basal medium wassupplemented with 20 nM valproic acid, 1 mM of butylated hydroxynisole(BHA), 50 mM potassium chloride, 10 nM forskolin, 1 nM hydrocortisoneand 5 mg/mL ITS.
All cells and bioengineered tissues were cultured in the differentinductive media for 20 days and the transdifferentiation processwas analyzed after 24 h, 10 days and 20 days of the addition of theinductive media.
746 R. NIETO-AGUILAR ET AL.
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

As noninduced controls, 2D-ACC and 3D-FAH were cultured in basalnoninductive media. As positive controls of cell differentiation, tissuesections of human bone, cartilage, adipose tissue, and brain were used.
Histological Analysis
After the different induction periods, all samples were washed in PBSand fixed in 4% formaldehyde (Sigma-Aldrich). Staining and analysis of2D-ACC samples was carried out directly on the culture chambers,whereas tissue sections of paraffin-embedded tissues were used for3D-FAH.
For detection of the osteoblastic phenotype and to identify any signs ofmineralization, samples were stained with 2% Alizarin Red S for 5 minat room temperature. Chondrogenesis was confirmed by mucopolysac-charide staining using Alcian Blue solution (1% alcian blue 8GX and 3%glacial acetic acid, pH adjusted to 2.5). Adipogenic differentiation wasidentified by Oil Red O staining (0.7 mg in 100 mL of propylene glycol)for 5 min at 608C. Morphological analysis of all samples was carried outby conventional hematoxylin and eosin staining (H&E). Finally, allsamples were analyzed using a light microscope, and the percentage ofcells stained per field was determined using NIS-Elements imagingsoftware (Nikon, Tokyo, Japan).
As controls, tissue sections of formaldehyde-fixed, paraffin-embeddedhuman bone, cartilage, adipose tissue, and brain were processed andstained following the same protocols (Figure 1).
Statistical comparison of the percentage of positive cells between 2D-ACC and 3D-FAH was performed using the exact test of Fisher,considering all p values below 5% as statistically significant.
Immunofluorescence
After 2D-ACC were induced in the slide chambers, the cells were washedfor 10 min in PBS and fixed in 70% ethanol for 30 min. For immunofluor-escence, the cells were washed in distilled water, preincubated in 2.5% horseserum and hybridized for 2 h with one of the following primary antibodies:anti-alkaline phosphatase (1 : 200 dilution, Sigma-Aldrich), anti-collagentype II (1 : 200 dilution, Santa Cruz Biotechnology, CA, USA), anti-leptin-OB(1 : 200 dilution, Sigma-Aldrich) and anti-nestin (1 : 250 dilution, abcam,Cambridge, UK). Then, samples were washed in PBS and incubated for30 min in specific anti-mouse FITC-conjugated secondary antibodies (anti-collagen type II, anti-leptin-OB), anti-goat Cy3-conjugated secondaryantibodies (anti-alkaline phosphatase) or anti-rabbit Cy3-conjugated
Pluripotential Differentiation Capability of ASCs 747
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

secondary antibodies (anti-nestin). Finally, samples were washed in PBS,coversliped with DAPI mounting medium (Vector Laboratories,Peterborough, UK) and analyzed using fluorescence light microscopy.
On the other hand, control human tissues and transdifferentiated3D-FAH were washed in PBS and fixed in metacarn
Figure 1. Human native tissues as positive controls: (A) and (B) human bone tissue.
(A) Alizarin red S stain, (B) alkaline phosphatase protein by immunofluoresecence. (C) and(D) Human native cartilage. (C) alcian blue stain. (D) type II collagen. (E) and (F) Human
adipose tissue controls. (E) H&E staining. (F) leptin marker. (G) and (H) Native human
brain tissue. (G) H&E staining. (H) nestin immunofluorescence.
748 R. NIETO-AGUILAR ET AL.
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

(methanol : chloroform : glacial acetic acid, 60 : 30 : 10) for 6 h at 48C.Then, samples were washed twice in ethanol 100%, dehydrated,paraffin-embedded and sectioned to 5 mm and mounted on microscopeslides.
After this, all samples were deparaffinized in xylene, rehydrated inethanol series (100%, 95%, 70%, 50%) and distilled water, preincubatedin 2.5% horse serum and hybridized for 2 h with one of the followingprimary antibodies: anti-alkaline phosphatase (1 : 200 dilution, Sigma-Aldrich), anti-collagen type II (1 : 200 dilution, Santa Cruz Antibodies,USA), anti-leptin-OB (1 : 200 dilution, Sigma-Aldrich), anti-nestin(1 : 250 dilution, abcam, UK). Then, samples were washed in PBS andincubated for 30 min in specific anti-mouse FITC-conjugated secondaryantibodies: anti-collagen type II and anti-leptin-OB (1 : 500 dilution,Sigma-Aldrich) or anti-goat Cy3-conjugated secondary antibodies: anti-alkaline phosphatase (1 : 500 dilution, Sigma-Aldrich) or anti-rabbit Cy3-conjugated secondary antibodies: anti-nestin (1 : 500 dilution, Sigma-Aldrich). Finally, samples were washed in PBS, coversliped with DAPImounting medium and analyzed using fluorescence light microscopy(Nikon Co. Eclipse i90, Japan). As positive controls, sections of normalhuman bone, cartilage, adipose tissue, and brain were processedfollowing the same immunofluorescence protocols (Figure 1).
Statistical comparison of the percentage of positive cells between 2D-ACC and 3D-FAH was performed using the exact test of Fisher,considering all p values below 5% as statistically significant.
Genome-wide Gene Expression Analysis of 2D-ACC usingOligonucleotide Microarrays
Total RNA corresponding to control noninduced ASCs and induced2D-ACC was extracted using the Qiagen RNeasy System (Qiagen,Mississauga, Ontario, Canada), according to the manufacturers’recommendations. RNA concentration was determined by absorbencyat 260 nm, and quality was verified by using a Bioanalyzer (Agilent).Total cDNA was synthesized with a T7-polyT primer and reversetranscriptase (Superscript II, Life Technologies, Inc., Carlsbad, CA)before in vitro transcription with biotinylated UTP and CTP (EnzoDiagnostics, Farmingdale, NY). Labeled nucleic acid target washybridized (458C for 16 h) to Affymetrix Human Genome U133 plus2.0 oligonucleotide arrays. After automated washing and staining,absolute values of expression were calculated and normalized from thescanned array by using Affymetrix Microarray Suite.
Pluripotential Differentiation Capability of ASCs 749
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

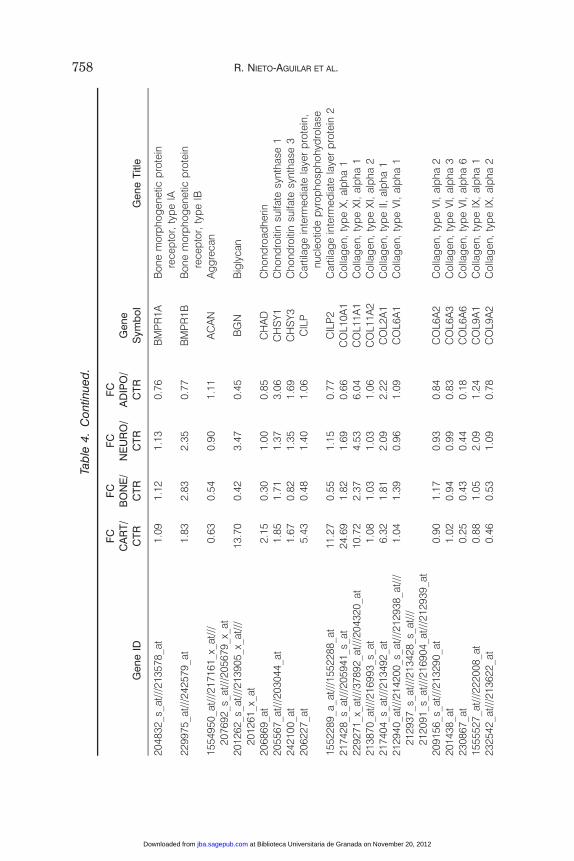
In this work, we have selected 95 genes with relevant functions relatedto each transdifferentiated cell type (osteocyte-like, adipocyte-like,chondrocyte-like, and neuron-like cells). To identify genes that becameupregulated after the induction process, we first calculated the averageexpression of control noninduced ASCs and of the different types oftransdifferentiated 2D-ACC. Then, the fold-change relative expression ofnoninduced controls versus transdifferentiated 2D-ACC was calculatedfor each cell type. At this step, all genes with a relative fold-change of atleast 1.2 (i.e., genes whose average expression became upregulated by atleast 20% in comparison with the control ASCs) were selected.
RESULTS
Induction to the Osteogenic Lineage
As shown in Figure 2 and Table 1, histological analysis of 2D-ACCinduced to osteoblast-type cells revealed that the production of a calcifiedextracellular matrix was absent for the first 24 h of induction (0% of areastained per field) for both alizarin red S staining and alkaline phosphataseimmunofluorescence. Then, when the cells were cultured in the osteo-inductive medium for 10 days, an initial generation of calcified extra-cellular matrix was detected in some scattered areas of the culture (5.24%of cells stained per field), with alkaline phosphatase being expressed by6.26% of the cells. Finally, cells incubated in the osteoinductive mediumfor 20 days showed large areas of calcification scattered all along the cellculture corresponding to 47.29% of stained cells per field and 40.6% of thecells showing alkaline phosphatase expression.
Then, the histological analysis of the 3D tissue constructs analyzed inthis study revealed that some of the cells in the 3D-FAH became trans-differentiated into osteoblast-like cells after 10 and 20 days of induction(7.60% positive cells by alizarin red S staining and 93.8 by alkalinephosphatase after 10 days of induction and 74.56% and 100%,respectively after 20 days of induction) (Figure 2 and Table 2). Asshown in Table 3, the percentage of positive cells was significantly higherin the 3D-FAH model than in 2D-ACC for alkaline phosphatase after 10days and for alkaline phosphatase and alizarin red S staining after 20 days( p¼ 0.0000).
Induction to the Chondrogenic Lineage
First, our results revealed that induction of 2D-ACC to thechondrogenic lineage during 24 h was not able to generate any
750 R. NIETO-AGUILAR ET AL.
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

Figure 2. 2D-ACC and 3D-FAH transdifferentiation to the osteogenic lineage. (A)–(C)Alizarin red staining of 2D-ACC cultured for 24 h, 10 days and 20 days, respectively.
(G)–(I): Alkaline phosphatase immunofluorescence in 2D-ACC induced for 24 h, 10 days
and 20 days, respectively. Orange signals correspond to positive alkaline phosphatase
expression. (D)–(F) Alizarin red staining of 3D-FAH cultured for 24 h, 10 days and 20 days,respectively. (J)–(L): Alkaline phosphatase immunofluorescence in 3D-FAH induced for
24 h, 10 days and 20 days, respectively.
Table 1. Percentage of positive transdifferentiated ASCs on the 2D-ACCmodel when induced to the osteogenic, chondrogenic, adipogenic and
neurogenic lineages as determined by histological staining for alizarin red S,alcian blue, oil red O, and hematoxylin and eosin, and by
immunofluorescence for alkaline phosphatase, type II collagen,leptin-OB and nestin after 24 h, 10 days, and 20 days of induction.
Osteogenicinduction
Chondrogenicinduction
Adipogenicinduction
Neurogenicinduction
2D-ACCAlizarinred S
Alkalinephosphatase
Alcianblue
Type IIcollagen
Oilred O Leptin-OB H&E Nestin
24 h 0.00 0.00 0.00 0.00 0.00 0.00 31.00 43.4210 days 5.24 6.26 84.08 92.34 0.00 90.50 44.00 57.3420 days 47.29 40.6 94.94 93.00 33.33 98.10 44.00 85.71
Pluripotential Differentiation Capability of ASCs 751
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

detectable levels of cell transdifferentiation as determined by alcian bluestaining and type II collagen expression (0% of cells stained per field)(Figure 3 and Table 1). However, 2D-ACC cultured for 10 days inchondrogenic medium resulted in a high percentage of cells transdiffer-entiated to chondrocyte-like cells (84.08% of alcian blue positive cellsand 92.34% of cells showing collagen II expression). This percentage waseven higher after 20 days of induction (94.94% by alcian blue and 93.00%by immunofluorescence).
Similarly, the analysis of 3D-FAH induced to the chondrogenic lineageshowed that the process of cell transdifferentiation was not effectiveafter 24 h, with 0% of cells transdifferentiated (Figure 3 and Table 2).Nevertheless, when these bioengineered tissues were induced during 10
Table 2. Percentage of ASCs for the 3D-FAH model transdifferentiated positivelyto the osteogenic, chondrogenic, adipogenic, and neurogenic lineages as
determined by histological staining for alizarin red S, alcian blue, oil red O andhematoxylin and eosin, and by immunofluorescence for alkaline phosphatase, type
II collagen, leptin-OB and nestin after different induction times.
Osteogenicinduction
Chondrogenicinduction
Adipogenicinduction
Neurogenicinduction
3D-FAHAlizarinred S
Alkalinephosphatase
Alcianblue
Type IIcollagen
Oilred O leptin-OB H&E Nestin
24 h 0.00 0.00 0.00 0.00 0.00 0.00 50.00 78.8010 days 7.60 93.80 14.80 80.00 0.00 87.00 55.60 85.3020 days 74.56 100.00 98.70* 86.70 92.30 87.90 60.00 95.74
Table 3. Differentiation analysis between 2D-ACC and 3D-FAH models induced tothe osteogenic, chondrogenic, adipogenic, and neurogenic lineages as deter-
mined by alizarin red S, alcian blue, oil red O and hematoxylin and eosin (H&E) andby immunofluorescence for alkaline phosphatase, type II collagen, leptin-OB and
nestin after 24 h, 10 days, and 20 days of induction process. Significantdifferences at p50.05 are considered as statistically significant.
2D-ACC
Osteogenicinduction
Chondrogenicinduction
Adipogenicinduction
NeurogenicInduction
vs 3D-FAH(FET)
Alizarinred S
Alkalinephosphatase
Alcianblue
Type IIcollagen
Oilred O Leptin-OB H&E Nestin
24 h 1.0000 1.0000 1.0000 1.0000 1.0000 1.0000 0.0093 0.000010 days 0.5679 0.0000 0.0000 0.0236 1.0000 0.5149 0.1196 0.000020 days 0.0000 0.0000 0.2116 0.2380 0.0000 0.0101 0.0335 0.0238
752 R. NIETO-AGUILAR ET AL.
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

days, 14.8% of the cells became positive by alcian blue staining, with80% of cells expressing collagen type II by immunofluorescence.The statistical analysis demonstrated that these percentages weresignificantly lower than those of the 2D-ACC model ( p¼ 0.0000 for thealcian blue and p¼ 0.0236 for the immunofluorescence) (Table 3).Finally, tissues induced to the chondrogenic lineage during 20 daysshowed 98.7% of the tissue area strongly stained by alcian blue and86.7% of the cells expressing collagen type II.
Induction to the Adipogenic Lineage
Histological analysis of 2D-ACC induced for 24 h or 10 days to theadipogenic lineage did not show the presence of any intracellular lipid
Figure 3. 2D-ACC and 3D-FAH chondrogenic differentiation: (A)–(C) Alcian blue
staining of ASCs induced after 24 h, 10 days and 20 days, respectively. (G)–(I) Type IIcollagen immunofluorescence in 2D-ACC chondroinduced for 24 h, 10 and 20 days,
correspondingly. (D)–(F) Alcian blue staining of 3D-FAH cultured for 24 h, 10 days and 20
days, respectively. (J)–(L) Type II collagen immunofluorescence in 3D-FAH induced for
24 h, 10 days and 20 days, respectively.
Pluripotential Differentiation Capability of ASCs 753
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

bodies by oil red O staining, with 0% of the cells expressing leptin-OB byimmunofluorescence after 24 h of induction. However, 90.5% of the cellsincubated for 10 days in the adipogenic medium showed detectableleptin-OB expression by immunofluorescence (Figure 4 and Table 1).Finally, one third of the cells induced for 20 days became stained by oilred O histochemistry, whereas 98.1% of cells expressed leptin-OB.
Similarly, our analysis of 3D-FAH showed that the adipogenic trans-differentiation process was not efficient at 24 h, and that 0% of cellsbecame stained by oil red O after 10 days in culture. In addition, 87% ofcells induced for 10 days expressed high amounts of leptin-OB. When thebioengineered 3D-FAH tissue constructs were induced during 20 days, thepercentage of oil red O-positive cells was significantly higher than that ofthe 2D-ACC (92.3% in 3D-FAH and 33.33% in 2D-ACC; p¼ 0.0000),
Figure 4. 2D-ACC and 3D-FAH adipogenic induction: (A)–(C) Oil red O staining of
induced ASCs to the adipogenic lineage for 24 h, 10 days and 20 days, respectively. (G)–(I)
Leptin-OB immunofluorescence in ASCs cultured for 24 h, 10 and 20 days, correspond-ingly. (D)–(F) Oil red O staining of 3D-FAH cultured for 24 h, 10 days and 20 days,
respectively. (J)–(L) Leptin OB immunofluorescence in 3D-FAH induced for 24 h, 10 days
and 20 days, respectively.
754 R. NIETO-AGUILAR ET AL.
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

although the proportion of cells that expressed leptin-OB was signifi-cantly lower in comparison to the 2D-ACC (87.9% in 3D-FAH and 98.1%in 2D-ACC; p¼ 0.0101) (Table 3).
Induction to the Neurogenic Lineage
H&E staining of 2D-ACC induced to the neurogenic lineage revealedthat the transdifferentiation process initiated after 24 h of induction,with 31% of the cells developing a cytoplasmic cell prolongation thatresembled a rudimentary axon-like process and 43.42% showing nestinexpression by immunofluorescence (Figure 5 and Table 1). When thecells were incubated in the neurogenic medium for 10 and 20 days, 44%of the cells showed large axon-like prolongations as well as some small
Figure 5. 2D-ACC and 3D-FAH induction to the neurogenic lineage: (A)–(C) 2D-ACC
induced in neurogenic medium for 24 h, 10 days and 20 days, respectively; and stained with
H&E. (G)–(I) Nestin immunofluorescence in neuroinduced ASCs for 24 h, 10 and 20 dayscorrespondingly. (D)–(F) H&E staining of 3D-FAH cultured for 24 h, 10 days and 20 days,
respectively. (J)–(L) Nestin immunofluorescence in 3D-FAH induced for 24 h, 10 days and
20 days, respectively.
Pluripotential Differentiation Capability of ASCs 755
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

dentritiform cellular extensions, with 57.34% of the cells being positivefor nestin at day 10 and 85.71% of cells at day 20.
The analysis of the induced bioengineered 3D-FAH tissues revealedthat 50% of the cells in the tissues displayed significant morphologicalmodification compatible with the process of neuron-like transdiffer-entiation after 24 h of induction, with 78.8% of the cells expressingnestin protein by immunofluorescence (Figure 5 and Table 2). Then,induction during 10 days resulted in 55.6% of neuron-like cells and85.3% of cells with nestin expression, whereas the incubation of thetissues in the inductive medium for 20 days was able to generate 60% ofneuron-like cells and 95.74% of cells expressing nestin protein. All thesenestin expression percentages were significantly higher in 3D-FAH thanin 2D-ACC, with the morphological changes being higher only after20 days of induction (Table 3).
Gene Expression Analysis of Transdifferentiated 2D-ACC
Gene expression analysis of 2D-ACC induced to the osteogenic lineagefor 20 days showed significant upregulation of several genes related toosteogenic differentiation and bone function (more than 20% expressionincrease in comparison with controls), including ALPL, BGLAP, BMP15,BMP3, BMP6, BMP7, BMP8B and BMPR1B (Table 4). In addition,BMP8A and B, BMPER and BMPR1A showed a gene expressionincrement that was lower than 20%. Moreover, the expression of somegenes decreased after osteogenic induction, including BMP1, BMP10,BMP2, BMP4, BMP5, and BMP8A. The gene expression analysis of 2D-ACC induced to the chondrogenic lineage after 20 days of inductionshowed upregulation of the genes BGN, CHAD, CHSY1, CHSY3, CILP,CILP2, COL10A1, COL11A1, COL2A1, CSGALNACT2, HAS1, andHAS2 (Table 4), whereas COL11A2, COL6A1, and COL6A3 showed agene expression increment that was lower than 20% and some genes(ACAN, COL6A2, COL6A6, COL9A1, COL9A2, COL9A3, and HAS3)decreased after chondrogenic induction. Similarly, 2D-ACC incubated inadipogenic medium showed clear upregulation (more than 20% overcontrols) of the genes ACACB, ADFP, ADIPOQ, ADIPOR1, ADIPOR2,CFD, DGAT2, FAB1, FAB4, FAB7, FADS1, FADS2, FADS3, GPAM,GPD1, LBP, LEP, LEPROT, LEPROLT1, LPIN1, LPIN3, LPL, PLIN,and PNPLA3, with two genes downregulated after the induction processLPIN2 and RETN (Table 4). Finally, after induction of 2D-ACC to theneurogenic lineage, the microarray gene expression analysis revealedthat several genes related to neurogenic development and differentiationbecame upregulated by 20%, including DDN, HNT, NAV1, NAV2, NAV3,
756 R. NIETO-AGUILAR ET AL.
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

Tab
le4
.G
en
eex
pre
ssio
nan
aly
sis
of
co
ntr
olA
SC
san
dA
SC
str
an
sdiffe
ren
tiate
dto
the
ost
eo
ge
nic
,ad
ipo
ge
nic
,c
ho
nd
rog
en
ic,
an
dn
eu
rog
en
iclin
eag
es
as
de
term
ine
db
ym
icro
arr
ay.
Ge
ne
ID:
Aff
yme
trix
refe
ren
ce
of
the
spe
cifi
cg
en
eo
rp
rob
e-s
et.
FC
CA
RT
/CT
R:
fold
-ch
an
ge
exp
ress
ion
of
ce
llsin
du
ce
dto
the
ch
on
dro
ge
nic
line
ag
evs
co
ntr
olA
SC
s.F
CB
ON
E/C
TR
:fo
ld-c
han
ge
exp
ress
ion
ofA
SC
sin
du
ce
din
too
ste
oc
yte
-lik
ec
ells
vsc
on
tro
lAS
Cs.
FC
NE
UR
O/C
TR
:fo
ld-c
han
ge
exp
ress
ion
ofc
ells
ind
uc
ed
toth
en
eu
rog
en
iclin
eag
evs
co
ntr
olA
SC
s.F
CA
DIP
O/C
TR
:fo
ld-c
han
ge
exp
ress
ion
ofc
ells
ind
uc
ed
toth
ead
ipo
ge
nic
line
ag
evs
co
ntr
ol
AS
Cs.
Ge
ne
ssh
ow
ing
co
nsi
de
rab
leove
rexp
ress
ion
(mo
reth
an
20
%in
cre
ase
)afte
rth
ein
du
ctio
nto
the
diffe
ren
tc
ell
line
ag
es
we
rese
lec
ted
.
Ge
ne
ID
FC
CA
RT
/C
TR
FC
BO
NE
/C
TR
FC
NE
UR
O/
CT
R
FC
AD
IPO
/C
TR
Ge
ne
Sym
bo
lG
en
eT
itle
1557924_s
_at/
//215783_s
_at
0.7
11.5
51.3
31.3
8A
LPL
Alk
alin
ep
ho
sph
ata
se,
liver/
bo
ne/k
idn
ey
206956_a
t0.9
61.2
90.8
01.0
6B
GLA
PB
on
eg
am
ma-c
arb
oxy
glu
tam
ate
(gla
)p
rote
in1570383_a
t///
202701_a
t///
1569002_
x_at/
//205574_x
_at/
//206725_x
_at/
//207595_s
_at/
//1569001_a
t
1.0
00.6
20.8
80.7
9B
MP
1B
on
em
orp
ho
gen
etic
pro
tein
1
208292_a
t1.0
10.5
91.5
50.8
1B
MP
10
Bo
ne
mo
rph
og
en
etic
pro
tein
10
221332_a
t2.3
71.8
91.7
41.5
2B
MP
15
Bo
ne
mo
rph
og
en
etic
pro
tein
15
205290_s
_at/
//205289_a
t0.5
00.2
60.6
30.2
2B
MP
2B
on
em
orp
ho
gen
etic
pro
tein
2208244_a
t4.9
01.9
00.6
09.9
0B
MP
3B
on
em
orp
ho
gen
etic
pro
tein
3211518_s
_at
0.0
70.0
50.0
80.0
6B
MP
4B
on
em
orp
ho
gen
etic
pro
tein
4205431_s
_at/
//205430_a
t0.3
80.8
30.7
10.5
9B
MP
5B
on
em
orp
ho
gen
etic
pro
tein
5241141_a
t///
206176_a
t///
215042_a
t1.1
82.5
01.3
41.2
6B
MP
6B
on
em
orp
ho
gen
etic
pro
tein
6211260_a
t///
209591_s
_at
1.4
22.2
61.5
11.8
7B
MP
7B
on
em
orp
ho
gen
etic
pro
tein
7207866_a
t///
220203_a
t///
220204_s
_at
1.4
40.7
70.8
21.1
0B
MP
8A
Bo
ne
mo
rph
og
en
etic
pro
tein
8a
221615_a
t0.8
81.0
10.9
01.0
5B
MP
8A
///
BM
P8B
Bo
ne
mo
rph
og
en
etic
pro
tein
8a
///
bo
ne
mo
rph
og
en
etic
pro
tein
8b
235275_a
t///
207865_s
_at
0.8
22.5
50.6
80.7
6B
MP
8B
Bo
ne
mo
rph
og
en
etic
pro
tein
8b
241986_a
t0.1
11.1
30.4
11.0
8B
MP
ER
BM
Pb
ind
ing
en
do
thelia
lre
gu
lad
or
(co
ntin
ue
d)
Pluripotential Differentiation Capability of ASCs 757
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from
Tab
le4
.C
on
tinu
ed
.
Ge
ne
ID
FC
CA
RT
/C
TR
FC
BO
NE
/C
TR
FC
NE
UR
O/
CT
R
FC
AD
IPO
/C
TR
Ge
ne
Sym
bo
lG
en
eT
itle
204832_s
_at/
//213578_a
t1.0
91.1
21.1
30.7
6B
MP
R1A
Bo
ne
mo
rph
og
en
etic
pro
tein
rece
pto
r,ty
pe
IA229975_a
t///
242579_a
t1.8
32.8
32.3
50.7
7B
MP
R1B
Bo
ne
mo
rph
og
en
etic
pro
tein
rece
pto
r,ty
pe
IB1554950_a
t///
217161_x
_at/
//207692_s
_at/
//205679_x
_at
0.6
30.5
40.9
01.1
1A
CA
NA
gg
reca
n
201262_s
_at/
//213905_x
_at/
//201261_x
_at
13.7
00.4
23.4
70.4
5B
GN
Big
lyca
n
206869_a
t2.1
50.3
01.0
00.8
5C
HA
DC
ho
nd
road
herin
205567_a
t///
203044_a
t1.8
51.7
11.3
73.0
6C
HS
Y1
Ch
on
dro
itin
sulfa
tesy
nth
ase
1242100_a
t1.6
70.8
21.3
51.6
9C
HS
Y3
Ch
on
dro
itin
sulfa
tesy
nth
ase
3206227_a
t5.4
30.4
81.4
01.0
6C
ILP
Cart
ilag
ein
term
ed
iate
laye
rp
rote
in,
nu
cleo
tide
pyr
op
ho
sph
oh
ydro
lase
1552289_a
_at/
//1552288_a
t11.2
70.5
51.1
50.7
7C
ILP
2C
art
ilag
ein
term
ed
iate
laye
rp
rote
in2
217428_s
_at/
//205941_s
_at
24.6
91.8
21.6
90.6
6C
OL1
0A
1C
olla
gen
,ty
pe
X,
alp
ha
1229271_x
_at/
//37892_a
t///
204320_a
t10.7
22.3
74.5
36.0
4C
OL1
1A
1C
olla
gen
,ty
pe
XI,
alp
ha
1213870_a
t///
216993_s
_at
1.0
81.0
31.0
31.0
6C
OL1
1A
2C
olla
gen
,ty
pe
XI,
alp
ha
2217404_s
_at/
//213492_a
t6.3
21.8
12.0
92.2
2C
OL2
A1
Co
llag
en
,ty
pe
II,alp
ha
1212940_a
t///
214200_s
_at/
//212938_a
t///
212937_s
_at/
//213428_s
_at/
//212091_s
_at/
//216904_a
t///
212939_a
t
1.0
41.3
90.9
61.0
9C
OL6
A1
Co
llag
en
,ty
pe
VI,
alp
ha
1
209156_s
_at/
//213290_a
t0.9
01.1
70.9
30.8
4C
OL6
A2
Co
llag
en
,ty
pe
VI,
alp
ha
2201438_a
t1.0
20.9
40.9
90.8
3C
OL6
A3
Co
llag
en
,ty
pe
VI,
alp
ha
3230867_a
t0.2
50.4
30.4
40.1
8C
OL6
A6
Co
llag
en
,ty
pe
VI,
alp
ha
61555527_a
t///
222008_a
t0.8
81.0
52.0
91.2
4C
OL9
A1
Co
llag
en
,ty
pe
IX,
alp
ha
1232542_a
t///
213622_a
t0.4
60.5
31.0
90.7
8C
OL9
A2
Co
llag
en
,ty
pe
IX,
alp
ha
2
758 R. NIETO-AGUILAR ET AL.
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

204724_s
_at
0.8
50.3
50.3
00.4
5C
OL9
A3
Co
llag
en
,ty
pe
IX,
alp
ha
3222235_s
_at/
//218871_x
_at/
//239077_a
t1.5
00.8
51.2
40.8
0C
SG
ALN
AC
T2
Ch
on
dro
itin
sulfa
teN
-ace
tylg
ala
cto
sam
inyl
tran
sfera
se2
207316_a
t8.6
30.7
01.7
35.1
5H
AS
1H
yalu
ron
an
syn
thase
1230372_a
t///
206432_a
t23.6
04.1
91.6
59.2
8H
AS
2H
yalu
ron
an
syn
thase
2223541_a
t///
228179_a
t///
1552980_a
t0.9
00.9
00.7
00.3
8H
AS
3H
yalu
ron
an
syn
thase
3214788_x
_at
0.7
31.5
31.7
31.3
3D
DN
Den
drin
227566_a
t///
222020_s
_at/
//241934_a
t10.7
40.5
52.8
81.3
4H
NT
Neu
rotr
imin
233567_a
t///
233870_a
t///
224774_s
_at/
//227584_a
t///
224771_a
t///
224770_
s_at/
//224773_a
t///
224772_a
t
2.8
71.6
61.6
91.2
8N
AV
1N
eu
ron
navi
gato
r1
1567357_a
t///
222599_s
_at/
//1556606_
at/
//1567358_a
t///
218330_s
_at/
//222598_s
_at
0.9
31.6
81.2
21.7
1N
AV
2N
eu
ron
navi
gato
r2
1552658_a
_at/
//1562234_a
_at/
//204823_a
t///
216632_a
t///
216466_a
t1.5
51.0
41.5
21.8
1N
AV
3N
eu
ron
navi
gato
r3
214952_a
t///
229799_s
_at/
//231532_a
t///
212843_a
t///
217359_s
_at/
//227394_a
t///
209968_s
_at
0.8
40.7
71.0
10.8
0N
CA
M1
Neu
ral
cell
ad
hesi
on
mo
lecu
le1
205669_a
t///
232390_a
t0.3
00.9
50.8
31.1
2N
CA
M2
Neu
ral
cell
ad
hesi
on
mo
lecu
le2
205143_a
t0.7
82.5
62.6
15.0
6N
CA
NN
eu
roca
n204412_s
_at/
//33767_a
t1.2
73.0
41.5
42.7
2N
EF
HN
eu
rofil
am
en
t,h
eavy
po
lyp
ep
tide
221805_a
t///
221801_x
_at/
//221916_a
t///
2.0
64.6
32.8
78.5
2N
EF
LN
eu
rofil
am
en
t,lig
ht
po
lyp
ep
tide
205113_a
t///
223902_a
t0.8
20.9
90.8
41.3
8N
EF
MN
eu
rofil
am
en
t,m
ed
ium
po
lyp
ep
tide
1553194_a
t///
229461_x
_at/
//243357_a
t///
239548_a
t3.1
96.3
92.7
28.0
7N
EG
R1
Neu
ron
al
gro
wth
reg
ula
tor
1
1556057_s
_at/
//206282_a
t0.6
22.6
42.8
41.0
2N
EU
RO
D1
Neu
rog
en
icd
iffere
ntia
tion
1210271_a
t///
1552953_a
_at
6.2
11.4
42.1
02.1
7N
EU
RO
D2
Neu
rog
en
icd
iffere
ntia
tion
2221318_a
t1.3
40.3
11.1
40.8
0N
EU
RO
D4
Neu
rog
en
icd
iffere
ntia
tion
4220045_a
t1.5
00.5
01.2
12.4
3N
EU
RO
D6
Neu
rog
en
icd
iffere
ntia
tion
6208497_x
_at
2.8
87.1
25.3
58.9
4N
EU
RO
G1
Neu
rog
en
in1
(co
ntin
ue
d)
Pluripotential Differentiation Capability of ASCs 759
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

Tab
le4
.C
on
tinu
ed
.
Ge
ne
ID
FC
CA
RT
/C
TR
FC
BO
NE
/C
TR
FC
NE
UR
O/
CT
R
FC
AD
IPO
/C
TR
Ge
ne
Sym
bo
lG
en
eT
itle
215632_a
t1.1
71.1
72.1
72.6
7N
EU
RO
G2
Neu
rog
en
in2
207965_a
t1.3
70.2
40.3
10.3
1N
EU
RO
G3
Neu
rog
en
in3
206814_a
t0.6
10.1
50.7
60.4
9N
GF
Nerv
eg
row
thfa
cto
r(b
eta
po
lyp
ep
tide)
205858_a
t3.6
041.6
04.4
0108.4
0N
GF
RN
erv
eg
row
thfa
cto
rre
cep
tor
(TN
FR
sup
erf
am
ily,
mem
ber
16)
217963_s
_at
0.8
61.2
30.9
91.1
1N
GF
RA
P1
Nerv
eg
row
thfa
cto
rre
cep
tor
(TN
FR
SF
16)
ass
oci
ate
dp
rote
in1
204105_s
_at/
//216959_x
_at
2.0
25.4
43.1
213.6
9N
RC
AM
Neu
ron
al
cell
ad
hesi
on
mo
lecu
le240532_a
t0.7
06.9
00.8
01.1
0S
LC32A
1S
olu
teca
rrie
rfa
mily
32
(GA
BA
vesi
cula
rtr
an
spo
rter)
,m
em
ber
11554724_a
t9.2
01.4
01.2
01.8
0S
LC6A
11
So
lute
carr
ier
fam
ily6
(neu
rotr
an
smitt
er
tran
spo
rter,
GA
BA
),m
em
ber
11
237058_x
_at
1.0
50.6
70.8
20.7
8S
LC6A
13
So
lute
carr
ier
fam
ily6
(neu
rotr
an
smitt
er
tran
spo
rter,
GA
BA
),m
em
ber
13
217213_a
t///
215715_a
t///
217214_s
_at/
//210353_s
_at/
//2.1
80.6
91.4
82.4
3S
LC6A
2S
olu
teca
rrie
rfa
mily
6(n
eu
rotr
an
smitt
er
tran
spo
rter,
no
rad
ren
alin
),m
em
ber
2207519_a
t2.7
12.0
62.4
60.9
4S
LC6A
4S
olu
teca
rrie
rfa
mily
6(n
eu
rotr
an
smitt
er
tran
spo
rter,
sero
ton
in),
mem
ber
4210854_x
_at/
//202219_a
t///
213843_
x_at/
//2.1
21.2
70.9
13.0
8S
LC6A
8S
olu
teca
rrie
rfa
mily
6(n
eu
rotr
an
smitt
er
tran
spo
rter,
creatin
e),
mem
ber
8207043_s
_at
1.0
80.2
61.2
50.4
9S
LC6A
9S
olu
teca
rrie
rfa
mily
6(n
eu
rotr
an
smitt
er
tran
spo
rter,
gly
cin
e),
mem
ber
9214584_x
_at/
//221928_a
t///
49452_a
t///
1552616_a
_at/
//1552615_a
t///
43427_a
t0.8
11.8
91.7
951.5
0A
CA
CB
Ace
tyl-C
oen
zym
eA
carb
oxy
lase
beta
215895_x
_at/
//209122_a
t0.4
20.7
30.9
51.6
7A
DF
PA
dip
ose
diff
ere
ntia
tion
-rela
ted
pro
tein
///A
dip
op
hili
n
760 R. NIETO-AGUILAR ET AL.
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

207175_a
t0.3
09.0
26.3
6556.2
1A
DIP
OQ
Ad
ipo
nect
in,
C1Q
an
dco
llag
en
do
main
con
tain
ing
217748_a
t1.2
81.1
20.8
81.3
3A
DIP
OR
1A
dip
on
ect
inre
cep
tor
1201346_a
t1.3
51.0
30.6
41.7
6A
DIP
OR
2A
dip
on
ect
inre
cep
tor
2205382_s
_at
0.1
01.5
70.9
33.5
4C
FD
Co
mp
lem
en
tfa
cto
rD
(ad
ipsi
n)
224327_s
_at/
//226064_s
_at
0.1
81.3
21.3
152.0
7D
GA
T2
Dia
cylg
lyce
rol
O-a
cyltr
an
sfera
seh
om
olo
g2
(mo
use
)205892_s
_at/
//231693_a
t0.7
81.3
71.8
71.2
8F
AB
P1
Fatty
aci
db
ind
ing
pro
tein
1,
liver,
mR
NA
(cD
NA
clo
ne
IMA
GE
:4712653)/
//fa
tty
aci
db
ind
ing
pro
tein
1,
liver
235978_a
t///
203980_a
t0.9
716.0
58.2
2924.0
5F
AB
P4
Fatty
aci
db
ind
ing
pro
tein
4,
ad
ipo
cyte
205029_s
_at/
//205030_a
t///
216192_a
t1.7
21.5
52.6
910.8
4F
AB
P7
Fatty
aci
db
ind
ing
pro
tein
7,
bra
in208964_s
_at/
//208963_x
_at/
//208962_s
_at
0.7
51.5
91.2
95.7
2F
AD
S1
Fatty
aci
dd
esa
tura
se1
202218_s
_at/
//243953_a
t1.8
02.2
61.3
43.4
4F
AD
S2
Fatty
aci
dd
esa
tura
se2
204257_a
t///
216080_s
_at
1.2
91.5
11.2
23.4
3F
AD
S3
Fatty
aci
dd
esa
tura
se3
225420_a
t///
225424_a
t3.2
31.6
62.0
344.3
2G
PA
MG
lyce
rol-3-p
ho
sph
ate
acy
ltran
sfera
se,
mito
cho
nd
rial
204997_a
t///
213706_a
t1.0
07.4
52.2
0997.9
3G
PD
1G
lyce
rol-3-p
ho
sph
ate
deh
ydro
gen
ase
1(s
olu
ble
)214461_a
t0.1
63.6
90.2
592.4
1LB
PLi
po
po
lysa
cch
arid
eb
ind
ing
pro
tein
207092_a
t13.2
070.2
06.1
019.8
0LE
PLe
ptin
227095_a
t///
202378_s
_at
0.8
21.2
41.3
01.4
4LE
PR
OT
Lep
tinre
cep
tor
ove
rlap
pin
gtr
an
scrip
t202595_s
_at
1.9
01.1
51.3
41.4
9LE
PR
OTL1
Lep
tinre
cep
tor
ove
rlap
pin
gtra
nscr
ipt-
like
1212276_a
t///
212274_a
t///
212272_a
t0.3
60.9
30.9
82.8
2LP
IN1
Lip
in1
202460_s
_at/
//202459_s
_at
0.8
81.0
90.8
10.9
5LP
IN2
Lip
in2
232966_a
t1.2
51.3
51.1
51.7
5LP
IN3
Lip
in3
203549_s
_at/
//203548_s
_at
0.2
19.5
24.0
8438.9
5LP
LLi
po
pro
tein
lipase
205913_a
t0.8
224.2
45.8
21879.9
4P
LIN
Peril
ipin
220675_s
_at/
//233030_a
t0.4
57.8
21.7
459.9
5P
NP
LA3
Pata
tin-lik
ep
ho
sph
olip
ase
do
main
con
tain
ing
3220570_a
t0.4
00.3
10.1
90.1
3R
ETN
Resi
stin
Pluripotential Differentiation Capability of ASCs 761
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

NCAN, NEFH, NEFL, NEGR1, NEUROD1, NEUROD2, NEUROD6,NEUROG1, NEUROG2, NGFR, NRCAM, SLC6A2, SLC6A4, andSLC6A9. In contrast, NCAM1, NEUROD4, and SLC6A11 showed agene expression increment that was lower than 20% and the genesNCAM2, NEFM, NEUROG3, NGF, NGFRAP1, SLC32A1, SLC6A13,and SLC6A8 decreased their expression after induction towards theneurogenic lineage (Table 4).
DISCUSSION
Generation of artificial tissues using adult stem cells and biocompa-tible biomaterials is one of the main objectives of current biomedicalresearch. Although these bioengineered tissues could be potentiallyuseful for the clinical substitution of damaged tissues, harvesting ofnative cells with proliferation and differentiation capabilities is notalways possible from these damaged tissues. For that reason, the searchof alternative cell sources for use as substitutes of the native cells is oneof the current challenges of the field.
In this context, ASCs are one of the most promising sources of humanadult mesenchymal stem cells [16,23] with high proliferative anddifferentiation capabilities [16,17]. One of the main properties of thesecells is their accessibility and the possibility of obtaining different types ofcells from a single pluripotential cell source. Although the potentiality ofASCs cultured on 2D systems was previously established [24,25], verylittle is so far known about the phenotype and differentiation potentialityof cells in 3D culture systems [26,27], in which cells resemble the naturalstructure of native tissues. For that reason, in this work we havecompared the behavior of ASCs cultured on 2D and 3D conditions usingfibrin-agarose biomaterials previously described by our research group[8,12–14]. Ideally, a good biomaterial should be suitable to act as a cellcarrier [28–37], guiding cell differentiation and proliferation [28].However, the influence of fibrin-agarose biomaterials on the differentia-tion capabilities of ASCs has not been elucidated to the date.
In this regard, our results demonstrated that 2D-ACC can beefficiently transdifferentitated to several tissue types, which wasconfirmed not only by histological analysis, but also by histochemistry,immunofluorescence, and high-throughput mRNA expression as deter-mined by microarray. In addition, our analyses confirmed the cellularplasticity of ASCs when subjected to the osteogenic, chondrogenic,adipogenic, and neurogenic induction media on 3D culture conditions(3D-FAH) as well. Strikingly the transdifferentiation potential of ASCssubjected to 3D cultures was comparable or even higher than that of 2D
762 R. NIETO-AGUILAR ET AL.
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

cultures, except for the 10 days chondrogenic induction. These resultsimply that the fibrin-agarose 3D culture model could support theefficient transdifferentiation capability of ASCs, suggesting that thegeneration of 3D human tissue substitutes using ASCs is a feasibletechnique in the laboratory.
When ASCs induced to the osteogenic lineage were analyzed, weobserved that a high percentage of the cells acquired the osteogenicphenotype after 20 days of induction, especially in the 3D culturesystem. In addition to the histological analysis, the gene expressionanalysis of 2D-ACC revealed that a high number of genes with a role inbone development and function became overexpressed at day 20 ofinduction. These results imply that transdifferentiated osteocyte-likecells not only resembled the morphology of bone cells, but they alsobehave like bone cells from the genetic point of view, supporting thepotential clinical use of these cells. In contrast, other relevantosteogenic-related genes did not show considerable levels of upregula-tion, likely due to the fact that the cells have been induced to theosteogenic lineage for only 20 days or to a lack of specific in situ bonesignals that would develop in an in vivo environment. Further in vivoexperiments could give new information of genes expression of inducedand transdifferentiated ASCs.
On the other hand, when we evaluated ASCs transdifferentiation to thechondrogenic lineage, we found that both the 2D-ACC and the 3D-FAHwere able to synthesize high amounts of mucopolysaccharides andcollagen type II at day 10 of induction, suggesting that the chondrogenicphenotype could be acquired before the osteogenic phenotype. Also,microarray gene expression of 2D-ACC confirmed that several genesrelated to the cartilage lineage were overexpressed upon induction,including BGN, CHAD, CHSY1, CHSY3, CILP, CILP2, COL10A1,COLL11A1, CSGALNACT2, HAS1, HAS2, and COL2A1. This latterhas been reported in previous studies as an important gene expressed inchondroinduced stem cells [24,25,38]. Quite the opposite, some genesencoding for some collagen isoforms and the gene HAS3 showeddownregulation after chondrogenic induction. This reduced gene expres-sion could be due to the fact that an in vivo environment or a higherex vivo induction period could be necessary for the proper upregulation ofcertain types of genes. All these data support the idea that ASCs inducedto the chondrogenic lineage have a very similar behavior to the nativecartilage and may have a potential usefulness in regenerative medicine.
Similarly, ASCs induced to adipocyte-like cells displayed high levels ofmRNA gene expression for most of the genes related to the adiposelineage. Indeed, 92% of the genes related to adipose tissue that were
Pluripotential Differentiation Capability of ASCs 763
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

analyzed in this work, became overexpressed after induction. Mostlikely, this is due to the fact that ASCs reside in adipose tissue and evenafter being isolated and expanded in culture, they could maintain a highamount of growing factors or intra/extracellular signals which couldreact once the adipogenic induction medium was applied ex vivo to thesecells. Also, our microarray analysis revealed that this gene over-expression was very high for several genes such as ADIPOQ, FABP4,GPD1, LPL, and PLIN, which are essential for adipocyte differentiationand function [39–42]. Nevertheless, cell staining with oil red O showedthat 2D-ACC and 3D-FAH did not show any cytoplasmatic droplets withtriglycerides content until the 20 day of adipoinduction, although asignificant leptin-OB expression was found from day 10. All these resultssuggest that ASCs have a high potential to differentiate into adipocyte-like cells ex vivo, which could be potentially useful in medicine.
In respect to the neurogenic induction, 2D-ACC overexpressed 63% ofthe related genes to the neural lineage after 20 days of neurogenicinduction, including among others, NEUROD6, NEUROG1, NEUROG2,and the nerve growth factor receptor NGFR. Interestingly, induced cellson both the 2D and the 3D culture models showed importantmorphological changes, with the formation of axon-like cell prolonga-tions, and rudimentary dendritic extensions at the cell membrane afteronly 24 h of induction. These findings coincide with the upregulation ofgenes that are specific of neuron development, including two types ofneurofilaments (the light and the heavy polypeptides), the neural celladhesion molecule NRCAM and the gene encoding for dendrin, with animportant role in the development of neuronal dendrites [43]. Accordingto the nestin protein expression determined by immunofluorescence,both the 2D-ACC and the 3D-FAH resulted in most of the cells (485%)expressing nestin after 20 days of induction. The potential clinical use ofautologous neuron-like cells developed on both models from adult stemcells is unlimited.
One of the main advantages of the methods described here is theirsimplicity, the rapidness of the transdifferentiation induction and thefact that a single cell type has been used for the generation of severaladult tissue types. Thus, by using specific conditioning culture media,ASCs can be efficiently differentiated to adipocyte-like cells in 2D-ACCand 3D-FAH models after 20 days in culture, transdifferentiated toneurons and chondrocytes-like cells after 10 and 20 days, respectivelyand into osteocytes-like cells after 20 days. Another advantage of thismethodology is that it does not imply any genetic manipulation of thecells. Therefore, the possibility of inducing a severe genomic alterationwith oncogenic potential is very low.
764 R. NIETO-AGUILAR ET AL.
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

In summary, in this work we have demonstrated that highly accessiblestem cells obtained from the adipose tissue (ASCs) are an efficientsource of cells for use in tissue engineering with high differentiationpotentiality on both 2D and 3D culture systems. The use of highlybiocompatible fibrin-agarose scaffolds results in comparable or evenhigher efficiencies of cell transdifferentiation ex vivo.
ACKNOWLEDGEMENTS
This work was supported by grants P06-CTS-2191 and SAS PI0135/2007 of Junta de Andalucia. This study was approved by the localResearch and Ethics committees of the University of Granada (number2006/2191).
REFERENCES
1. Holcomb, J.D. and Gentile, R.D. Aesthetic Facial Surgery of MalePatients: Demographics and Market Trends, Facial Plast. Surg., 2005: 21:223–231.
2. Housman, T.S., Hancox, J.G., Mir, M.R. et al. What Specialties Perform theMost Common Outpatient Cosmetic Procedures in the United States?Dermatol. Surg., 2008: 34: 1–7.
3. Chiu, R.C. Msc Immune Tolerance in Cellular Cardiomyoplasty, Semin.Thorac. Cardiovasc. Surg., 2008: 20: 115–118.
4. Baillie, D.R., Stawicki, S.P., Eustance, N., Warsaw, D. and Desai, D. Use ofHuman and Porcine Dermal-derived Bioprostheses in Complex AbdominalWall Reconstructions: A Literature Review and Case Report, Ostomy WoundManage., 2007: 53: 30–37.
5. Kim, I.K., Bedi, D.S., Denecke, C., Ge, X. and Tullius, S.G. Impact of Innateand Adaptive Immunity on Rejection and Tolerance, Transplantation, 2008:86: 889–894.
6. Atala, A. Advances in Tissue and Organ Replacement, Curr. Stem Cell Res.Ther., 2008: 3: 21–31.
7. Priya, S.G., Jungvid, H. and Kumar, A. Skin Tissue Engineering for TissueRepair and Regeneration, Tissue Eng. Part B Rev., 2008: 14: 105–118.
8. Alaminos, M., Del Carmen Sanchez-Quevedo, M., Munoz-Avila, J.I. et al.Construction of a Complete Rabbit Cornea Substitute Using a Fibrin-agarose Scaffold, Invest. Ophthalmol. Vis. Sci., 2006: 47: 3311–3317.
9. Alaminos, M., Sanchez-Quevedo, M.C., Munoz-Avila, J.I. et al. Evaluation ofthe Viability of Cultured Corneal Endothelial Cells by Quantitative ElectronProbe X-ray Microanalysis, J. Cell. Physiol., 2007: 211: 692–698.
10. Stevens, B., Yang, Y., Mohandas, A., Stucker, B. and Nguyen, K.T. A Reviewof Materials, Fabrication Methods, and Strategies Used to Enhance BoneRegeneration in Engineered Bone Tissues, J. Biomed. Mater. Res. B Appl.Biomater., 2008: 85: 573–582.
Pluripotential Differentiation Capability of ASCs 765
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

11. Fuchs, S., Ghanaati, S., Orth, C. et al. Contribution of Outgrowth EndothelialCells from Human Peripheral Blood on In Vivo Vascularization of BoneTissue Engineered Constructs Based on Starch Polycaprolactone Scaffolds,Biomaterials, 2009: 30: 526–534.
12. Alaminos, M., Garzon, I., Sanchez-Quevedo, M.C. et al. Time-course Study ofHistological and Genetic Patterns of Differentiation in Human EngineeredOral Mucosa, J. Tissue Eng. Regen. Med., 2007: 1: 350–359.
13. Sanchez-Quevedo, M.C., Alaminos, M., Capitan, L.M. et al. Histological andHistochemical Evaluation of Human Oral Mucosa Constructs Developed byTissue Engineering, Histol. Histopathol., 2007: 22: 631–640.
14. Garzon, I., Sanchez-Quevedo, M.C., Moreu, G. et al. In Vitro and In VivoCytokeratin Patterns of Expression in Bioengineered Human PeriodontalMucosa, J. Periodontal Res., 2009: 44(5): 588–597.
15. Kinikoglu, B., Auxenfans, C., Pierrillas, P. et al. Reconstruction of a Full-Thickness Collagen-based Human Oral Mucosal Equivalent, Biomaterials,2009: 30: 6418–6425.
16. De Ugarte, D.A., Alfonso, Z., Zuk, P.A. et al. Differential Expression of StemCell Mobilization-associated Molecules on Multi-lineage Cells from AdiposeTissue and Bone Marrow, Immunol. Lett., 2003: 89: 267–270.
17. Schaffler, A. and Buchler, C. Concise Review: Adipose Tissue-derivedStromal Cells – Basic and Clinical Implications for Novel Cell-basedTherapies, Stem Cells, 2007: 25: 818–827.
18. Weinzierl, K., Hemprich, A. and Frerich, B. Bone Engineering with AdiposeTissue Derived Stromal Cells, J. Craniomaxillofac. Surg., 2006: 34:466–471.
19. Xu, Y., Malladi, P., Wagner, D.R. and Longaker, M.T. Adipose-derivedMesenchymal Cells as a Potential Cell Source for Skeletal Regeneration,Curr. Opin. Mol. Ther., 2005: 7: 300–305.
20. Wei, Y., Sun, X., Wang, W. and Hu, Y. Adipose-derived Stem Cells andChondrogenesis, Cytotherapy, 2007: 9: 712–716.
21. Lin, C.S., Xin, Z.C., Deng, C.H., Ning, H., Lin, G. and Lue, T.F. RecentAdvances in Andrology-related Stem Cell Research, Asian J. Androl., 2008:10: 171–175.
22. Yoshimura, K., Sato, K., Aoi, N. et al. Cell-assisted Lipotransfer for FacialLipoatrophy: Efficacy of Clinical Use of Adipose-derived Stem Cells,Dermatol Surg., 2008: 34(9): 1178–85.
23. Zuk, P.A, Zhu, M., Ashjian, P. et al. Human Adipose Tissue is a Source ofMultipotent Stem Cells, Mol. Biol. Cell, 2002: 13: 4279–4295.
24. Guilak, F., Lott, K.E., Awad, H.A. et al. Clonal Analysis of theDifferentiation Potential of Human Adipose-derived Adult Stem Cells,J. Cell. Physiol., 2006: 206: 229–237.
25. Matsumoto, T., Kano, K., Kondo, D. et al. Mature Adipocyte-derivedDedifferentiated Fat Cells Exhibit Multilineage Potential, J. Cell. Physiol.,2008: 215: 210–222.
26. Duggal, S., Fronsdal, K.B., Szoke, K., Shahdadfar, A., Melvik, J.E. andBrinchmann, J.E. Phenotype and Gene Expression of Human MesenchymalStem Cells in Alginate Scaffolds, Tissue Eng. Part A, 2009: 15: 1763–1773.
766 R. NIETO-AGUILAR ET AL.
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

27. Mauney, J.R., Nguyen, T., Gillen, K., Kirker-Head, C., Gimble, J.M. andKaplan, D.L. Engineering Adipose-like Tissue In Vitro and In Vivo UtilizingHuman Bone Marrow and Adipose-derived Mesenchymal Stem Cells withSilk Fibroin 3D Scaffolds, Biomaterials, 2007: 28: 5280–5290.
28. Hutmacher, D.W., Goh, J.C. and Teoh, S.H. An Introduction toBiodegradable Materials for Tissue Engineering Applications, Ann. Acad.Med. Singapore, 2001: 30: 183–191.
29. Bumgardner, J.D., Wiser, R., Gerard, P.D. et al. Chitosan: Potential Use as aBioactive Coating for Orthopaedic and Craniofacial/Dental Implants,J. Biomater. Sci. Polym. Ed., 2003: 14: 423–438.
30. Saadeh, P.B., Khosla, R.K., Mehrara, B.J. et al. Repair of a Critical SizeDefect in the Rat Mandible Using Allogenic Type I Collagen, J. Craniofac.Surg., 2001: 12: 573–579.
31. Seol, Y.J., Lee, J.Y., Park, Y.J. et al. Chitosan Sponges as TissueEngineering Scaffolds for Bone Formation, Biotechnol. Lett., 2004: 26:1037–1041.
32. Solchaga, L.A., Gao, J., Dennis, J.E. et al. Treatment of OsteochondralDefects with Autologous Bone Marrow in a Hyaluronan-based DeliveryVehicle, Tissue Eng., 2002: 8: 333–347.
33. Trubiani, O., Orsini, G., Zini, N. et al. Regenerative Potential of HumanPeriodontal Ligament Derived Stem Cells on Three-dimensional Biomaterials:A Morphological Report, J. Biomed. Mater. Res. A, 2008: 87: 986–993.
34. Behravesh, E., Yasko, A.W., Engel, P.S. and Mikos, A.G. SyntheticBiodegradable Polymers for Orthopaedic Applications, Clin. Orthop. Relat.Res., 1999: 367(S): 118–129.
35. Ishaug, S.L., Crane, G.M., Miller, M.J., Yasko, A.W., Yaszemski, M.J. andMikos, A.G. Bone Formation by Three-dimensional Stromal OsteoblastCulture in Biodegradable Polymer Scaffolds, J. Biomed. Mater. Res., 1997:36: 17–28.
36. Neuss, S., Apel, C., Buttler, P. et al. Assessment of Stem Cell/BiomaterialCombinations for Stem Cell-based Tissue Engineering, Biomaterials, 2008:29: 302–313.
37. Munirah, S., Kim, S.H., Ruszymah, B.H. and Khang, G. The Use of Fibrinand Poly(lactic-co-glycolic acid) Hybrid Scaffold for Articular CartilageTissue Engineering: An In Vivo Analysis, Eur. Cell. Mater., 2008: 15: 41–52.
38. Mehlhorn, A.T., Zwingmann, J., Finkenzeller, G. et al. Chondrogenesis ofAdipose-derived Adult Stem Cells in a Poly-lactide-co-glycolide Scaffold,Tissue Eng. Part A, 2009: 15: 1159–1167.
39. Suga, H., Eto, H., Inoue, K. et al. Cellular and Molecular Features of LipomaTissue: Comparison with Normal Adipose Tissue, Br. J. Dermatol., 2009:161: 819–825.
40. De Rosa, A., De Francesco, F., Tirino, V. et al. A New Method for theCryopreserving ASCs: An Attractive and Suitable Large-scale and Long-termCell Banking Technology, Tissue Eng. Part C Methods, 2009: 15: 659–667.
41. Noer, A., Boquest, A.C. and Collas, P. Dynamics of Adipogenic PromoterDNA Methylation During Clonal Culture of Human Adipose Stem Cells toSenescence, BMC Cell. Biol., 2007: 8: 18–29.
Pluripotential Differentiation Capability of ASCs 767
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from

42. Qi, L., Zhang, C., Greenberg, A. and Hu, F.B. Common Variations inPerilipin Gene, Central Obesity, and Risk of Type 2 Diabetes in US Women,Obesity (Silver Spring), 2008: 16: 1061–1065.
43. Neuner-Jehle, M., Denizot, J.P., Borbely, A.A. and Mallet, J.Characterization and Sleep Deprivation-induced Expression Modulation ofDendrin, a Novel Dendritic Protein in Rat Brain Neurons, J. Neurosci. Res.,1996: 46: 138–151.
768 R. NIETO-AGUILAR ET AL.
at Biblioteca Universitaria de Granada on November 20, 2012jba.sagepub.comDownloaded from














![Journal of Biomaterials Applications ‘Green’ biocompatible ... Biomater Appl-20… · PVA/ chitosan/nano-ZnO composite nanofibrous membranes Antibacterial and antifungal [16]](https://static.fdocuments.us/doc/165x107/605be37fd9239d416832e8c2/journal-of-biomaterials-applications-agreena-biocompatible-biomater-appl-20.jpg)



