
Histologiabasicadegartner1eraedicion 150127211651-conversion-gate01
353
-
Upload
betty-martinez-rodriguez -
Category
Documents
-
view
239 -
download
1
Transcript of Histologiabasicadegartner1eraedicion 150127211651-conversion-gate01


HistologıabasicaLESLIE P. GARTNER, PhDProfessor of Anatomy (Retirada)
Department of Biomedical Sciences
Baltimore College of Dental Surgery
Dental School
University of Maryland
Baltimore, Maryland
JAMES L. HIATT, PhDProfessor Emeritus
Department of Biomedical Sciences
Baltimore College of Dental Surgery
Dental School
University of Maryland
Baltimore, Maryland

Edicion en espanol de la obra original en ingl�es
Concise Histology
Copyright � MMXI by Saunders, an imprint of Elsevier Inc. All rights reserved.
Revision cientıfica
M.a Jesus Fern�andez Acenero
Doctora en Medicina. Especialista en Anatomıa Patologica
Hospital Fundacion Jim�enez DıazProfesora Asociada de Anatomıa Patologica
Universidad Autonoma de Madrid
� 2011 Elsevier Espana, S.L.Travessera de Gr�acia, 17-21 – 08021 Barcelona, Espana
Fotocopiar es un delito (Art. 270 C.P.)
Para que existan libros es necesario el trabajo de un importante colectivo (autores, traductores, dibujantes,correctores, impresores, editores...). El principal beneficiario de ese esfuerzo es el lector que aprovecha su
contenido.
Quien fotocopia un libro, en las circunstancias previstas por la ley, delinque y contribuye a la «no» existencia
de nuevas ediciones. Adem�as, a corto plazo, encarece el precio de las ya existentes.Este libro est�a legalmente protegido por los derechos de propiedad intelectual. Cualquier uso fuera de los
lımites establecidos por la legislacion vigente, sin el consentimiento del editor, es ilegal. Esto se aplica en
particular a la reproduccion, fotocopia, traduccion, grabacion o cualquier otro sistema de recuperacion y
almacenaje de informacion.
ISBN edicion original: 978-0-7020-3114-4
ISBN edicion espanola: 978-84-8086-868-6
Traduccion y produccion editorial:GEACONSULTORIA EDITORIAL, S.L.
Advertencia
La medicina es un �area en constante evolucion. Aunque deben seguirse unas precauciones de seguridadest�andar, a medida que aumenten nuestros conocimientos gracias a la investigacion b�asica y clınica habr�a
que introducir cambios en los tratamientos y en los f�armacos. En consecuencia, se recomienda a los lectores
que analicen los ultimos datos aportados por los fabricantes sobre cada f�armaco para comprobar las dosis
recomendadas, la vıa y duracion de la administracion y las contraindicaciones. Es responsabilidad inelu-dible del m�edico determinar las dosis y el tratamiento m�as indicados para cada paciente, en funcion de su
experiencia y del conocimiento de cada caso concreto. Ni los editores ni los directores asumen respon-
sabilidad alguna por los danos que pudieran generarse a personas o propiedades como consecuencia del
contenido de esta obra.El editor

A mi esposa, Roseann;
mi hija, Jennifer;
y mi madre, Mary
LPG
A mis nietos,
Nathan David,
James Mallary,
Hanna Elisabeth,
Alexandra Renate,
Eric James,
y Elise Victoria
JLH



Prefacio
Una vez m�as, nos complace presentar una nueva obrade histologıa, basada en la tercera edicion de nuestrotratado Color Textbook of Histology, un libro de textoque disfruta de reconocimiento no solamente en elidioma en el que fue redactado inicialmente, sinotambi�en en otras lenguas.
A lo largo de las tres ultimas d�ecadas, la histologıaha dejado de ocuparse de la mera descripcion de laanatomıa microscopica para transformarse en unadisciplina compleja que conjuga la anatomıa funcio-nal con la biologıa molecular y celular. Esta obra pre-senta un diseno peculiar, ya que cada p�agina par con-tiene un texto que se ilustra en la p�agina imparcorrespondientemediante bellısimas im�agenes a todocolor procedentes de la tercera edicion de Color Text-book of Histology. Por consiguiente, cada pareja dep�aginas enfrentadas pretende actuar como una uni-dad independiente de aprendizaje. Casi todas las uni-dades de aprendizaje se acompanan de consideracionesclınicas de inter�es para el tema abordado con el fin deponer de manifiesto la relevancia de la informacionallı presentada para un profesional de las ciencias de lasalud. Tanto los estudiantes como el profesoradopercibir�an la ausencia de im�agenes de microscopiaoptica y electronica en esta nueva obra, ya que se haprescindido de ellas de manera deliberada en el textoimpreso para incluirlas en la p�agina web Student Con-sult vinculada con el mismo. Pretendemos reducir eltamano del libro y, por tanto, facilitar las cosas alos estudiantes actuales, los cuales han de adquirir
diversos conceptos que hace 10 anos se presentabana lo largo de 16 semanas en pr�acticamente la mitad deese tiempo. Adem�as de las im�agenes impresas en lasp�aginas impares de este libro de texto, la web de Stu-dent Consult alberga 150 im�agenes de microscopiaoptica y electronica divididas por capıtulos que seacompanan de preguntas y respuestas de examen parafacilitar la evaluacion por parte del estudiante de sucapacidad de reconocimiento de organos/tejidos/c�elulas relevantes y del conocimientos de sus caracte-rısticas funcionales. Asimismo, en la p�agina web figu-ran casos clınicos con preguntas que ponen de relievela importancia de la histologıa en las ciencias de lasalud al tiempo que preparan al estudiante para elcomponente histologico de sus ex�amenes. Tanto enel libro de texto como en el material complementariode la p�agina web se ha procurado destacar los concep-tos clave que sustentan nuestra presentacion de lahistologıa, a saber, la existencia de una estrecha rela-cion entre la estructura y la funcion.
Hemos tratado de ofrecer un panorama completo yexacto de esta materia, si bien somos conscientes dehaber cometido errores y omisiones en una tarea de talmagnitud. En consecuencia, seguimos confiando enlas sugerencias, consejos y crıticas de los lectores paramejorar las proximas ediciones de esta obra.
Leslie P. Gartner
James L. Hiatt
v


Agradecimientos
La histologıa es una asignatura visual, por lo que suestudio ha de basarse necesariamente en im�agenes degran calidad. Agradecemos enormemente a ToddSmith su minuciosidad a la hora de revisar y crearnuevas ilustraciones. Asimismo, deseamos expresarnuestra gratitud a un gran numero de colegas de todoel mundo y sus editores por su generosidad al permi-tirnos utilizar su material gr�afico.
Por ultimo, agradecemos su colaboracion al equipoencargado de este proyecto en Elsevier: Kate Dimock,Barbara Cicalese, Lou Forgione y Carol Emery, asıcomo a Linnea Hermanson por su trabajo incansableen la produccion de esta obra.
vii


�Indice de capıtulos
1 Introduccion a la histologıa . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
2 Citoplasma . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8
3 Nucleo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26
4 Matriz extracelular. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40
5 Epitelio y gl�andulas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48
6 Tejido conjuntivo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 62
7 Cartılago y hueso . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 74
8 Musculo. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 94
9 Tejido nervioso . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 108
10 Sangre y hematopoyesis . . . . . . . . . . . . . . . . . . . . . . . . . . 132
11 Aparato circulatorio. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 152
12 Sistema linf�atico (inmunitario) . . . . . . . . . . . . . . . . . . . . . . . 168
13 Sistema endocrino . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 188
14 Sistema tegumentario . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 204
15 Aparato respiratorio . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 218
16 Aparato digestivo: cavidad bucal . . . . . . . . . . . . . . . . . . . . 230
17 Aparato digestivo: tubo digestivo . . . . . . . . . . . . . . . . . . . . 238
18 Aparato digestivo: gl�andulas . . . . . . . . . . . . . . . . . . . . . . . 250
19 Aparato urinario. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 260
20 Aparato genital femenino . . . . . . . . . . . . . . . . . . . . . . . . . . 272
21 Aparato genital masculino . . . . . . . . . . . . . . . . . . . . . . . . . 286
22 Organos de los sentidos . . . . . . . . . . . . . . . . . . . . . . . . . . . 304
�Indice alfab�etico . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .325
ix

Histologıa basica

1 INTRODUCCIONA LA HISTOLOGIA
La histologıa es el �area de conocimiento que estudia lostejidos de animales y plantas, si bien la obra Histologıab�asica se ocupa exclusivamente de los tejidos de losmamıferos y, en particular, de los humanos. Adem�asde la estructura de dichos tejidos, este texto librodescribe las de las c�elulas, los organosy los sistemas org�anicos, por lo que lamateria aquı presentadadeberıa deno-minarse anatomıa microscopica. Sabe-mos que el cuerpo se compone de:
. C�elulas
. Matriz extracelular (MEC), en la quese encuentran inmersas las c�elulas
. Lıquido extracelular, que atraviesala MEC para transportar nutrientes,oxıgeno y mol�eculas desenalizacion a las c�elulas y paraeliminar productos de desecho,dioxido de carbono, otrasmol�eculas de senalizacion, hormonas y f�armacosdel ambiente extracelular.. El lıquido extracelular proviene del plasmasanguıneo y se extravasa hacia la MEC en la caraarterial de los lechos capilares; la mayor parte delmismo regresa al plasma a trav�es de la cara venosade dichos lechos.
. La fraccion restante de este lıquido pasa a losvasos del sistema linf�atico, cuya presion esmenor, para regresar al torrente circulatorio en launion de la vena yugular interna y las venassubclavias derecha e izquierda.
En los textos de histologıa modernos no se tratade manera aislada la morfologıa microscopica delorganismo, sino que tambi�en se analiza su funciona-miento. La materia que es objeto de esta obra serelaciona, asimismo, con la biologıa celular, lafisiologıa, la biologıamolecular, la bioquımica, la ana-tomıa macroscopica, la embriologıa e, incluso, conalgunos aspectos de la medicina clınica en el apartado«Consideraciones clınicas». Esperamos que el estudiode la histologıa haga percibir al lector la importanciade la relacion entre estructura y funcion. Sin em-bargo, antes de que esto fuera posible, fue precisodesarrollar diversas t�ecnicas de visualizacion de c�elulasy tejidos muertos que conservan, en gran medida, suaspecto en condiciones in vivo.
Microscopia optica
PREPARACION DE LAS MUESTRAS TISULARES
Un pequeno bloque de tejido, obtenido de un sujetoanestesiado o que ha muerto recientemente:
1. Se fija, por lo general con formoltamponado neutro tratado para per-mitir el r�apido entrecruzamiento conlas proteınas tisulares, con el fin defijarlas en la localizacion que ocupa-ban en el tejido vivo.
2. Tras la fijacion, la muestra sedeshidrata en una serie gradual debanos de alcohol.
3. La muestra se sumerge en xileno, elcual hace que el tejido se tornetransparente.
4. La muestra debe introducirse enparafina lıquida, que impregnar�a el
tejido, para hacer posible la visualizacion de losdelgados cortes tisulares en el microscopio. Lamuestra tisular se introduce en un recipiente pequenopara enfriarse y formar un bloque de parafina.
5. El bloque se corta en secciones delgadas de 5 a10mm por medio de un microtomo dotado de unacuchilla afilada que separa finas l�aminas tisularesdel mismo.
6. Los cortes se transfieren a portaobjetos recubiertosde una sustancia adhesiva, se retira la parafinamediante un bano con xileno y se rehidrata lamuestra con una serie gradual de banos en alcohol(por orden inverso a la secuencia dedeshidratacion).
7. Las secciones ası hidratadas se tinenmediante distintos colorantes hidrosolubles(tabla 1.1); la tincion de hematoxilina-eosina (H-E)es una de las m�as utilizadas en las preparacioneshistologicas habituales. La hematoxilina confiere unacoloracion azulada a los componentes �acidos dec�elulas y tejidos, mientras que la eosina tine de colorrosado los componentes b�asicos.
Los microscopios opticos actuales constan de una seriede lentes ordenadas para lograr el m�aximo aumento altiempo que se mantiene el poder de resolucion. Esteinstrumento posee m�as de una lente, por lo que recibeel nombre de microscopio compuesto (fig. 1.1).
� 2011 Elsevier Espana, S.L. Reservados todos los derechos
TERMINOS CLAVE. Microscopia optica. Inmunocitoquımica. Autorradiografıa. Microscopiaconfocal. Microscopia
electronica detransmision
. Microscopiaelectronicadebarrido
2

Una l�ampara de gran intensidad emite luz, la cualse enfoca en la muestra desde abajo a trav�es de uncondensador. La luz que atraviesa la muestra es reco-gida por una de las lentes del objetivo acopladas altambor giratorio, el cual permite modificar elaumento de bajo a intermedio y a alto, y una lentede inmersion, que aumentan la imagen 4, 10, 20, 40 y100 veces en los microscopios convencionales. Lostres primeros aumentos corresponden a lentes secas,mientras que la de inmersion requiere aceite de inmer-sion, el cual actua como interfaz entre el cristal de la
preparacion y la lente del objetivo. La luz recogida porel objetivo es captada por el ocular, el cual la amplificageneralmente 10 veces para obtener un aumentofinal de 40, 100, 200, 400 y 1.000 de la imagen quevisualizar�a la retina.
INTERPRETACION DE LOS CORTES MICROSCOPICOS
Los cortes histologicos son planos bidimension-ales extraıdos de una estructura tridimensional. Ini-cialmente, el estudiante presenta dificultades para
3
1
INTRODUCCIONALA
HISTO
LOGIA
Tabla 1.1 TINCIONES Y REACCIONES HISTOLOGICAS FRECUENTES
Reactivo Resultado
Hematoxilina Azul: nucleo; regiones �acidas del citoplasma; matriz de cartılagoEosina Rosa: regiones b�asicas del citoplasma; fibras de col�ageno
Tricromico de Masson Azul oscuro: nucleo
Rojo: musculo, queratina, citoplasma
Azul claro: mucinogeno, col�agenoColorante de orceına para fibras el�asticas Marron: fibras el�asticas
Colorante de Weigert para fibras el�asticas Azul: fibras el�asticas
Tincion de plata Negro: fibras reticulares
Hematoxilina f�errica Negro: estriaciones musculares, nucleo, eritrocitosAcido peryodico de Schiff Magenta: glucogeno y mol�eculas ricas en hidratos de carbono
Colorantes de Wright y Giemsa* Rosa: eritrocitos, gr�anulos eosinofilos
Azul: citoplasma de monocitos, eritrocitos y linfocitos
*Utilizada en la tincion diferencial de gr�anulos de las celulas hem�aticas.
Figura 1.1 Comparacion de los microscopios optico, electronico de transmision y electronico de barrido. (Tomado de Gartner LP, Hiatt JL:
Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p. 4.)�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

relacionar la imagen observada en el microscopio conel tejido u organo de procedencia de la muestra. Lasim�agenes de un tubo enroscado cortado a distintos�angulos (fig. 1.2)muestran como se puede reconstruirla morfologıa tridimensional a partir de una serie deim�agenes bidimensionales.
METODOS AVANZADOS DE VISUALIZACION
Se han desarrollado varias t�ecnicas que permitenestudiar el funcionamiento de las c�elulas, los tejidosy los organos por medio del microscopio. Las moda-lidades m�as utilizadas son la histoquımica (y lacitoquımica), la inmunohistoquımica y la autorra-diografıa.
. Los m�etodos histoquımicos y citoquımicos sebasan en reacciones quımicas, procesos enzim�aticosy procesos fisicoquımicos que confieren coloracional tejido, al tiempo que permiten identificar lalocalizacion de diversas macromol�eculas intra- yextracelulares.. Uno de los m�etodos histoquımicos m�asutilizados es la tincion con �acido peryodico deSchiff (PAS), a trav�es de la cual se confiere
coloracion viol�acea-rojiza al glucogeno y lasmol�eculas ricas en hidratos de carbono.La ausencia de color rojo indica la desapariciondel glucogeno en un punto dado comoconsecuencia del tratamiento de cortesconsecutivos con la enzima amilasa, la cualhidroliza el glucogeno.
. Otras t�ecnicas histoquımicas y citoquımicaspermiten localizar enzimas; sin embargo, no sevisualiza la propia enzima sino la presencia delproducto de la reaccion que precipita en formade deposito coloreado en el lugar de lareaccion.
. Las t�ecnicas inmunocitoquımicas identifican demanera m�as precisa la localizacion de unamacromol�ecula determinada que los m�etodoshistoquımicos o citoquımicos.. Noobstante, se trata de una t�ecnicam�as compleja,ya que precisa de anticuerpos frente a la mol�eculade inter�es en el m�etodo directo, o bien
. Depende de la preparacion de un anticuerpofrente a un anticuerpo primario en su m�etodoindirecto (fig. 1.3) y elmarcado del aquel con unamol�ecula fluorescente, como rodamina ofluoresceına. La sensibilidad y la precision del
4
1
INTRODUCCIONALA
HISTO
LOGIA
Figura 1.2 Diagrama bidimensional de un tubo tridimensional cortado en distintos planos. (Tomado de Gartner LP, Hiatt JL: ColorTextbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 4.)

m�etodo indirecto son m�as altas, puesto que unnumero mayor de mol�eculas de anticuerpomarcadas se une al anticuerpo primario que en elm�etodo directo. Por otra parte, los anticuerposprimarios suelen ser m�as costosos y encontrarsedisponibles en cantidades limitadas.
. Asimismo, la inmunocitoquımica puedeutilizarse en microscopia electronica mediante launion del metal pesado ferritina en lugar delmarcador fluorescente.
. En la autorradiografıa se emplea un isotoporadioactivo (normalmente, tritio, 3H) que seincorpora a la mol�ecula de inter�es.. Para estudiar la sıntesis de una proteına dada,se introducen amino�acidos tritiados en elsistema y se recogen muestras en perıodosdefinidos.
. Los cortes se procesan del modo habitual, si bienlas preparaciones se recubren de una capa deemulsion fotogr�afica en lugar de un cubreobjetosy se mantienen en la oscuridad durante variassemanas.
. Se revela y se fija la emulsion de manera similar auna placa fotogr�afica y se dispone uncubreobjetos sobre el corte.
. Al observar la muestra en el microscopio, sevisualizan gr�anulos de plata sobre las zonas de lamuestra a las que se incorporaron las mol�eculasmarcadas con el isotopo.
. Se ha adaptado la t�ecnica de autorradiografıa a lamicroscopia electronica.
Microscopia confocal
En la microscopia confocal se dirige un haz de l�asersobre una muestra impregnada en colorantes fluores-centes; el haz incidente que atraviesa un espejo
dicroico excita dichas mol�eculas, que emiten fluores-cencia (fig. 1.4).
. El rayo l�aser atraviesa una pequena aberturacontrolada inform�aticamente, de modo que lamuestra emite fluorescencia amedida que es barridapor el haz.
. La luz fluorescente emitida por la muestra escapturada conforme atraviesa la abertura en sentidocontrario al de la luz l�aser.
. Un detector fotomultiplicador captura la luzemitida; el sistema inform�atico recoge cada uno delos pıxeles ası obtenidos para elaborar una imagende la muestra.
. En cada toma se observa unicamente un planomuy delgado de la muestra, por lo que es precisoefectuar numerosos barridos a distintos nivelescon el fin de crear una imagen tridimensional dela misma.
Microscopia electronica
En la microscopia electronica se utiliza un haz de elec-trones en lugar de fotones como fuente de luz, el cual seamplıa y enfoca por medio de electroimanes (fig. 1.5).
. La resolucion del microscopio depende de lalongitud de onda de la luz emitida y la longitud deonda de un haz de electrones es mucho menor quela de la luz visible; la resolucion del haz deelectrones es, aproximadamente, 1.000 veces mayorque la de la luz visible. El poder de resolucion de unmicroscopio optico compuesto es de unos 200nm,mientras que el de un microscopio electronico detransmision es de 0,2 nm, lo que supone unaamplificacion de unas 150.000 veces y permitevisualizar mol�eculas individuales, como lamiosina.
5
1
INTRODUCCIONALA
HISTO
LOGIA
Figura 1.3 M�etodos directos e indirectos de inmunocitoquımica. Izquierda. Se marca un anticuerpo frente a un antıgeno con un
colorante fluorescente y se visualiza con un microscopio de fluorescencia. La region que emite luz fluorescente indica la localizacion
del anticuerpo marcado. Derecha. Se preparan anticuerpos marcados frente a un anticuerpo que reacciona frente a un antıgeno dado.
En el microscopio de fluorescencia, la senal fluorescente representa el lugar de reaccion del antıgeno con el anticuerpo. (Tomado deGartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 5.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

6
1
INTRODUCCIONALA
HISTO
LOGIA
Figura 1.4 Esquema de unmicroscopio confocal en el que se observan la abertura que atraviesa el haz de l�aser para explorar lamuestra y
la trayectoria de la luz fluorescente que es emitida posteriormente por lamuestra hasta alcanzar el detector fotomultiplicador. (Tomado
de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 8.)
Figura 1.5 Comparacion de los microscopios optico, electronico de transmision y electronico de barrido. (Tomado de Gartner LP, Hiatt JL:Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p. 4.)

. Existen dos modalidades de microscopiaelectronica: la microscopia electronica detransmision (MET) y la microscopia electronicade barrido (MEB).. Como indica su propio nombre, en la MET(v. fig. 1.3, derecha) los electrones han deatravesar un corte muy delgado tenido conmetales pesados (p. ej., fosfato de plomoo acetatode uranilo) y chocan contra una placafluorescente, que los absorbe y emite puntoslumınicos cuya intensidad depende de la energıacin�etica del electron incidente. Esta energıadisminuye conforme interacciona el electron conla muestra, de modo que cuanto mayor sea lacantidad de metales pesados absorbida por unaregion dada de la muestra, m�as acusada ser�a ladisminucion de la energıa cin�etica del electron. Laimagen ası formada se compone de puntos de luzde distintas intensidades que comprenden del grisclaro al gris oscuro. La imagen puede registrarse al
sustituir la placa fluorescente por una pelıculafotogr�afica sensible a electrones, que se revelar�adel modo habitual para obtener una fotografıa enblanco y negro.
. Los electrones no han de atravesar lamuestra en laMEB (v. fig. 1.5), sino que son dirigidos hacia lasuperficie de la misma para generar una imagentridimensional de la misma. Para ello, la muestradebe revestirse de una capa de un metal pesado,como oro o paladio. A medida que el haz deelectrones recorre la superficie de la muestra,algunos electrones se reflejan en dicha capa(electrones retrodispersos) y otros provocanla expulsion de electrones del metal pesado(electrones secundarios). Tanto los electronesdispersos como los secundarios son capturadospor detectores de electrones y generan una imagentridimensional que se muestra en un monitor.La imagen digitalizada se puede guardar en unarchivo e imprimir como una fotografıa.
7
1
INTRODUCCIONALA
HISTO
LOGIA
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

2 CITOPLASMALos organismos complejos se componen de c�elulas ymaterial extracelular. Existen m�as de 200 tipos dife-rentes de c�elulas, cadaunade las cualesrealiza una funcion diferente, que seorganizan junto a la matriz celularpara formar cuatro tipos b�asicos detejido: epitelio, tejido conjuntivo,musculo y tejido nervioso. Los tejidosforman organos y los organos se com-binan en sistemas org�anicos.
Por lo general, la c�elula es unaestructura rodeada por una mem-brana y que contiene el protoplasma,el cual se subdivide en dos comparti-mentos: el citoplasma y el cario-plasma (fig. 2.1).
. El carioplasma corresponde al nucleoy se rodea de la envoltura nuclear.
. Este capıtulo se ocupa de la membrana celular y elcitoplasma de una c�elula tıpica.. La sustancia m�as abundante del citoplasma es elcitosol, una suspension lıquida en la que seencuentran disueltas o suspendidas diversasmol�eculas inorg�anicas y org�anicas,macromol�eculas, pigmentos, cristales yorg�anulos.
. El citosol se rodea de una membrana celularformada por una bicapa lipıdica semipermeable(plasmalema, membrana plasm�atica) conproteınas embebidas.
Membrana celular (plasmalema,membrana plasm�atica)
Lamembrana celular presenta un grosor de 7 a 8nmyse compone de una bicapa lipıdica formada por fos-folıpidos anfip�aticos, colesterol y proteınas embebi-das o unidas (fig. 2.2). En el microscopio electronico,el plasmalema consta de dos capas densas:. Una hojuela interna (citopl�asmica). Una hojuela externa, que delimita una capaintermedia hidrofoba transparente
Esta estructura trilaminar recibe el nombre de unidaddemembrana, ya que no solo forma parte de la mem-brana celular, sino tambi�en de los org�anulos conmembrana presentes en la c�elula. Las proteınas suelenrepresentar el 50% del peso de una membrana tıpica.Los fosfolıpidos se disponen de tal modo que:
. Las cabezas hidrofilas se orientan hacia la periferia yforman las superficies intra- y extracelulares.
. Las cadenas hidrofobas de los �acidos grasos de lasdos l�aminas fosfolipıdicas enfrentadas (hojuelas
interna y externa) se proyectanhacia la region central de lamembrana y forman la capaintermedia transparente.
El colesterol aparece, habitual-mente, entre las colas de �acidos grasosde las mol�eculas de fosfolıpidos.Cuando lamembrana celular se sometea criofractura, la separacion tiene lugarpreferentemente a lo largo de la capahidrofoba transparente, lo que deja aldescubierto la superficie interna deambas hojuelas (fig. 2.3).
. La superficie de la hojuela interna (m�as cercana alcitoplasma) se denomina cara P.
. La superficie de la hojuela externa (m�as proxima alespacio extracelular) recibe el nombre de cara E.
Las proteınas asociadas a la membrana celular seclasifican en dos grupos: integrales y perif�ericas. Lasproteınas integrales son:
. Proteınas transmembrana, que atraviesan lamembrana para alcanzar el citoplasma y el espacioextracelular.
. Proteınas perif�ericas, que no se encuentran en elinterior de la membrana, sino que se unen a susuperficie citopl�asmica o extracelular. Durante elprocesode criofractura, unnumeromayor de proteınasse mantienen unidas a la cara P que a la cara E.
. La superficie extracelular de la membrana celular,que puede recubrirse de un glucoc�aliz (cubiertacelular), se compone de hidratos de carbono queestablecen enlaces con proteınas integrales ofosfolıpidos para formar glucoproteınas oglucolıpidos, respectivamente.
Las proteınas integrales y perif�ericas poseen uncierto grado de movimiento en la bicapa fosfolipıdicabidimensional, la cual remeda un mosaico sometidocontinuamente a cambios. La capacidad de desplaza-miento de estas proteınas es limitada y el modelo demembrana que recibıa el nombre de modelo del mosaicofluido se conoce en la actualidad como modelo delmosaico fluido modificado. Algunas regiones de lamembrana presentan engrosamientos formados porla concentracion de glucoesfingolıpidos y colesterolalrededor de ciertas proteınas de membrana. Estaszonas especializadas, llamadas balsas lipıdicas, inter-vienen en la senalizacion celular.
� 2011 Elsevier Espana, S.L. Reservados todos los derechos
TERMINOS CLAVE
. C�elula
. Canales ionicos
. Proteınastransportadoras
. Org�anulos
. Sıntesis de proteınas
. Tr�afico de membrana
. Citoesqueleto
. Inclusiones
8

9
2
CITO
PLASMA
Figura 2.1 C�elula tipo y sus org�anulos. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders,
2007, p 14.)
Figura 2.2 Modelo del mosaico fluido de la membrana celular. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.
Philadelphia, Saunders, 2007, p 16.)
Figura 2.3 Caras E y P de la membrana plasm�atica. (Tomado de Gartner
LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders,
2007, p 16.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

10
2
CITO
PLASMA
PROTEINAS TRANSPORTADORAS DE MEMBRANA
La membrana plasm�atica es permeable a las mol�ecu-las apolares, como el oxıgeno, y mol�eculas polares sincarga, como el agua y el glicerol, las cuales puedenatravesarla por difusion simple a favor de un gradientede concentracion. Sin embargo, los iones y lasmol�eculas polares de pequeno tamano han deser transportadas por ciertas proteınas integralesmultipaso, denominadas proteınas transportadorasde membrana, las cuales se ocupan de transferir estasmol�eculas a trav�es del plasmalema.
. El transporte a trav�es de la membrana plasm�aticarecibe el nombre de transporte pasivo cuando norequiere gasto de energıa.
. La transferencia de mol�eculas con gasto energ�eticose conoce como transporte activo (fig. 2.4).
Las proteınas de transporte de membrana seclasifican como proteınas de canal y proteınas trans-portadoras.
. Las proteınas de canal tan solo est�an implicadas enel transporte pasivo, ya que carecen de la capacidadde utilizar energıa para transportar mol�eculas encontra de un gradiente de concentracion.. Estas proteınas se pliegan de tal modo que creancanales ionicoshidrofilos a trav�es de lamembranaplasm�atica para desempenar su funcion.
. La mayorıa de estos canales controla el paso demol�eculas hacia el interior de los mismos pormedio de unas barreras denominadascompuertas que impiden su entrada o su salida.La apertura de estos canales con compuertadepende de varios mecanismos de control.
. Los canales con compuerta de voltaje, como loscanales deNa+ de las fibras nerviosas, se abren comoconsecuencia de la despolarizacion de la membrana(v. capıtulo 9).
. Los canales con compuerta de ligando se abrencomo consecuencia de la union de una mol�ecula desenalizacion (ligando) al canal ionico. Algunoscanales con compuerta de ligando responden a laestimulacion por neurotransmisores, por lo que seconocen como canales con compuerta deneurotransmisores (como los presentes en elmusculo esquel�etico).
. Otros canales responden a nucleotidos, como elmonofosfato de adenosina cıclico (AMPc) oel monofosfato de guanosina cıclico (GMPc), y sedenominan canales de compuerta de nucleotidos(p. ej., los que aparecen en los bastones de las retinas).
. La apertura de los canales de compuerta mec�anicaobedece al contacto fısico, como sucede en ladeflexion de los estereocilios de las c�elulas pilosasdel oıdo interno.
. Los canales ionicos con compuerta de proteınaG, como los receptores de acetilcolina de las
c�elulas del musculo cardıaco, precisan de laactivacion de una proteına G para la apertura de lacompuerta.. Los canales sin compuerta se encuentran abiertospermanentemente. Los m�as frecuentes son loscanales de escape de K+, los cuales se ocupan delmantenimiento de los potenciales de reposo delas c�elulas nerviosas. Las acuaporinas, unoscanales encargados del transporte de H2O,tambi�en pertenecen a este grupo.
. Las proteınas transportadoras son proteınasmultipaso; estas proteınas no solamente actuancomo canales pasivos que permiten el paso a favorde un gradiente de concentracion, sino que tambi�entransportan mol�eculas en contra de dicho gradientemediante mecanismos basados en el trifosfato deadenosina (ATP). Tambi�en se diferencian de loscanales ionicos por la presencia de sitios de unionpara iones o mol�eculas especıficas en su superficieinterna. Pueden transportar una mol�ecula o ion enuna direccion (uniporte), o bien acoplar dosmol�eculas diferentes en el:. Mismo sentido (simporte), o bien. Sentido contrario (antiporte)
La proteına transportadora m�as abundante esla bomba de Na+-K+, la cual utiliza a la ATPasa deNa+-K+ para expulsar tres iones de sodio en contrade gradiente de concentracion al espacio extracelulare introducir dos iones de potasio en el citoplasma.Algunas proteınas transportadoras emplean la con-centracion diferencial deNa+ en el citoplasma y el espa-cio extracelular para impulsar el movimiento de ciertosionesomol�eculaspequenasencontradegradiente. Esteproceso, realizado por proteınas transportadoras aco-pladas, recibe el nombre de transporte activo secunda-rio; la glucosa y enNa+ suelen transportarse demaneraconjunta mediante este mecanismo.
SENALIZACION CELULAR
Las c�elulas se comunican entre sı a trav�es de pequenasmol�eculas (mol�eculas de senalizacion, ligandos) queson secretadas y se unen a receptores presentes en otrasc�elulas. La c�elula que libera la mol�ecula de senalizacionse denomina c�elula senalizadora y la c�elula portadoradel receptor se conoce como c�elula diana.
Con frecuencia, las funciones de estas c�elulas puedeninvertirse, ya que, amenudo, la comunicacion es bidire-ccional. Cuando el ligando es una mol�ecula polar, losreceptores suelen localizarse en la superficie celular.Los receptores intracelulares o intranucleares correspon-den a mol�eculas hidrofobas apolares (p. ej., una hor-mona esteroidea)obienun receptor situado en la super-ficie de la c�elula transduce la senal a trav�es de laactivaciondeun sistema intracelularde segundosmen-sajeros (p. ej., receptores ligados a proteına G).

CONSIDERACIONES CL�INICAS
Una proteına transportadora elimina el
amino�acido cisteına del tubulo proximal de lanefrona. Algunas personas que portan dos copiasde la misma mutacion, cada una de las cualesproviene de uno de sus progenitores, que originaproteınas transportadoras anomalas, padecen untrastorno conocido como cistinuria. Laconcentracion del amino�acido en la orina de estossujetos es lo suficientemente alta para formarpiedras de cisteına. La cistinuria se manifiestaentre los 10 y los 30 anos de edad y provoca unanefrolitiasis recurrente. El diagnostico se sustentaen la deteccion de cristales de cisteına en la orinaen el estudio microscopico y las concentraciones
elevadas de este amino�acido en el an�alisis deorina. Se trata de una enfermedad que puederesultar muy dolorosa, si bien la ingesta de lıquidoabundante logra diluir la orina en un gradosuficiente para evitar la formacion de piedras enla mayorıa de los afectados.
11
2
CITO
PLASMA
Figura 2.4 Tipos de transporte. A. Transporte pasivo que no requiere gasto de energıa. B. El transporte activo es un mecanismo que
requiere gasto de energıa. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 18.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

12
2
CITO
PLASMA
Receptores ligados a proteına G y segundosmensajeros en la c�elula
Los receptores ligados a proteına G (proteınas deunion a nucleotidos de guanina) son proteınas trans-membrana cuyos dominios extracelulares poseen sitiosde union para mol�eculas de senalizacion especıficas(ligandos) y cuyos dominios intracelulares se asociana una proteına G en la hojuela interna de lamembranaplasm�atica. Cuando lamol�ecula de senalizacion se uneal sitio extracitopl�asmico del receptor, su dominiointracitopl�asmico sufre un cambio conformacionalque provoca la activacion de la proteına G (fig. 2.5).Existen varios tipos de proteınas G: estimuladoras (Gs),inhibidoras (Gi), sensibles e insensibles a la toxinapertussis (G0 y GBq) y transducina (Gt).
. Las proteınas Gs son trim�ericas, dado que secomponen de tres subunidades: a, b, y g . Suelenencontrarse en estado inactivo, en el cual portan unamol�ecula de difosfato de guanosina (GDP) unidoa su dominio citopl�asmico.
. Cuando la proteına Gs se activa, sustituye el GDPpor una mol�ecula de trifosfato de guanosina (GTP);la subunidad a se disocia de las otras dos parainteraccionar con la enzima adenilato ciclasa, lacual se activa y cataliza la transformacion del ATPcitopl�asmico en AMPc.
. Al desacoplarse el ligando del receptor unido a laproteına G, el GTP la subunidad a pierde un grupofosfato y se separa de la adenilato ciclasa paraasociarse, de nuevo, a las subunidades b y g .
. El AMPc, uno de losmensajeros celulares secundarios,activa la cinasa A, la cual pone en marcha unarespuesta especıfica por parte de la c�elula.
. En otras c�elulas, el AMPc pasa al nucleo y activala proteına de union a CRE, que se une a ciertasregiones reguladoras de los genes conocidas comoCRE (elementos de respuesta a AMPc), lo queposibilita la transcripcion de un gen determinadoque est�a vinculado con la respuesta especıfica dela c�elula en cuestion.
Maquinaria celular implicada en la sıntesisde proteınas
La sıntesis de proteınas es una funcion destacada de lamayorıa de las c�elulas, que son utilizadas por ellasmismas o bien exportadas a otras zonas del orga-nismo. La sıntesis de proteınas engloba:
. Un componente intranuclear, la transcripcion, esdecir, la sıntesis de unamol�ecula deARNmensajero(ARNm), y
. La traduccion, el componente citopl�asmico, queconsiste en el ensamblaje de amino�acidos en unasecuencia correcta,basadaenelmoldedenucleotidosde ARNm especıfico para una proteına determinada
El componente citopl�asmico de la maquinariaimplicada en la sıntesis proteica unicamente emplearibosomas cuando el destino de la proteına es el cito-plasma, mientras que utiliza ribosomas y el retıculoendopl�asmico rugoso (RER) (fig. 2.6) en el caso de lasque se empaquetan para almacenarse en la c�elula obien secretarse al espacio extracelular.
. Los ribosomas son unas partıculas pequenas(12 nm�25nm) formadas por una subunidadmayor y una subunidad menor. Las subunidadesse fabrican en el nucleo y se componen de ARNribosomico (ARNr) y proteınas. La subunidadmenor posee sitios de union para el ARNm,adem�as de otros tres sitios: uno al que se une elpeptidil-ARN de transferencia (ARNt) (sitio P),otro al que se une el aminoacil-ARNt (sitio A) y untercer sitio de liberacion (sitio E), en el que elARNt vacıo se separa del ribosoma. La subunidadmayor se une a la menor y posee un ARNrespecial con actividad enzim�atica llamadoribozima, que cataliza la formacion de enlacespeptıdicos que permiten que los amino�acidos seunan entre sı.
. Existen dos tipos de retıculo endopl�asmico (RE):retıculo endopl�asmico liso (REL) y RER. A pesar deno estar implicado en la sıntesis de proteınas, en esteapartado se trata la estructura del primero de elloscon el fin de ofrecer una visionm�as completa de esteorg�anulo.. El REL est�a formado por una serie de tubulos yvesıculas aplanadas cuya luz podrıa estarconectada con la del RER. Entre las funciones delREL figuran la sıntesis de lıpidos y esteroides, elmetabolismo del glucogeno y la detoxificacion deproductos toxicos, y el almacenamiento de calcioen las c�elulas musculares.
. El RER interviene en la sıntesis de proteınas quese empaquetar�an para almacenarse en la c�elula obien ser secretadas al espacio extracelular. Est�aconstituido por un sistema de cisternas aplanadasinterconectadas a cuya cara citopl�asmica seadhieren ribosomas y polirribosomas con unaelevada actividad de traduccion de ARNm ysıntesis de proteınas. El RER posee el receptor dela partıcula de reconocimiento de la senal(proteına de anclaje), riboforinas I y II ytranslocadores, unas proteınas que anclan losribosomas al RER y funcionan como un poro atrav�es del cual las proteınas en proceso deformacion ingresan en la cara interna (luminal)de la membrana de este org�anulo. Esta caraalberga la enzima peptidasa de senales ydolicol fosfato, el cual participa en laN-glucosilacion. Las cisternas del RER secomunican con la cisterna perinuclear de laenvoltura nuclear.

13
2
CITO
PLASMA
Figura 2.6 C�elula tipo y sus org�anulos. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders,
2007, p 14.)
Figura 2.5 Receptor ligado a proteına G. PPi, pirofosfato inorg�anico. (Tomado deGartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.Philadelphia, Saunders, 2007, p 21.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Sıntesis de proteınas
El proceso de sıntesis de proteınas se pone en marchacomo consecuencia de la union de una mol�ecula deARNm que se une a un ribosoma en el citoplasma yconcluye en ese mismo compartimento en el casode las proteınas citosolicas. El ARNm correspondientea las proteınas que se empaquetan contiene un p�eptidosenal que, al ser traducido, crea una senal de transportedel complejo ribosoma-ARNm hacia el RER.
SINTESIS DE PROTEINAS CITOSOLICAS
A continuacion se describe el proceso de sıntesis de lasproteınas que no deben empaquetarse (fig. 2.7):
. El ARNm sale del nucleo a trav�es de un complejo deporos nucleares (v. capıtulo 3), pasa al citoplasma yse asocia a una unidad ribosomica menor, cuyositio P se encuentra ocupado por unARNt iniciadorunido al amino�acido metionina. El anticodon delARNt corresponde al codon del ARNm, lo quepermite la alineacion correcta de los componentesdel sistema. Una subunidad ribosomica mayor seacopla al complejo ası formado y la traduccion sepone enmarcha al recorrer el ribosoma un codon dela cadena de ARNm en sentido 5’ a 3’.
. Un ARNt unido a un amino�acido (aminoacil-ARNt)y portador de un anticodon correcto se ancla al sitioA de la unidad ribosomica menor y se forma unenlace peptıdico entre dicho amino�acido y lametionina localizada en el sitio P. La metionina sedesprende del ARNt fijado al sitio P, demodo que elARNt que ocupa el sitio A porta un dip�eptido(metionina y el amino�acido reci�en anadido). ElARNt desaminado se desplaza al sitio E y el ARNtunido a los dos amino�acidos pasa al sitio P. Porultimo, el ribosoma recorre la distancia de un codonen la cadena del ARNm en sentido 5’ a 3’.
. Otro aminoacil-ARNt con el anticodon correctose une al sitio A. Interacciona con el dip�eptidodel ARNt localizado en el sitio P, que se une a estenuevo ARNt, el cual porta tres amino�acidos. El ARNtdel sitio E se separa del complejo y el ARNt vacıo sedesplaza al sitio E desocupado. El ARNt unido altrip�eptido pasa del sitio P al sitio A y el complejoribosomico recorre un unico codon en sentido5’ a 3’. Un nuevo aminoacil-ARNt con el codonadecuado se fija al sitio A, vacıo de nuevo.
. El proceso continua con la union de nuevassubunidades menores al extremo 5’ del ARNm, demodo que varios ribosomas traducen de manerasimult�anea lamismamol�ecula de ARNm. La cadenade ARNm leıda por varios ribosomassimult�aneamente recibe el nombre depolirribosoma o polisoma.
. El proceso de union de nuevos aminoacil-ARNt serepite hasta que el complejo llega a un codon determinacion, el cual indica que se ha anadido elultimo amino�acido al polip�eptido en formacion. Selibera, ası, el ultimoARNt desaminado del sitio E sinque otros aminoacil-ARNt se unan al sitio A, y lassubunidades ribosomicas mayor y menor sedisocian del ARNm.
SINTESIS DE PROTEINAS QUE SE EMPAQUETARAN
La sıntesis de proteınas no citosolicas (fig. 2.8) seinicia en el citoplasma de manera similar a la descritaen el apartado anterior.
. La cadena peptıdica que se va sintetizando es elp�eptido senal, el cual es reconocido por lapartıcula de reconocimiento de la senal (PRS),una mol�ecula formada por proteına y ARN que selocaliza en el citoplasma. La sıntesis proteica sedetiene como consecuencia de la union de la PRSal p�eptido senal y el complejo formado por elribosoma, el ARNm y la PRS migra hacia el RER.
. La PRS se ancla al receptor de la PRS (proteına deanclaje), situado en la membrana del RER, y elribosoma se fija a las proteınas translocadoras–proteınas integrales– de dicha membrana. La PRSse libera debido a estas interacciones; la traduccionprosigue y la base de las proteınas translocadoras seabre para crear un poro en la cisterna del RER. Laproteına en formacion pasa a la luz del RER a trav�esde dicho poro.
. La enzima peptidasa de senales escinde el p�eptidosenal y algunas de las proteınas en formacion sufrenun proceso de N-glucosilacion por dolicol fosfatopresente en la cara luminal de la membrana del RER.En este proceso intervienen las proteınas riboforina Iy riboforina II, exclusivas del RER y localizadas en sumembrana. La traduccion concluye cuando lamaquinaria alcanza el codon de terminacion.
. La proteına reci�en sintetizada se transloca a lascisternas del RER, en las que sufrir�a diversasmodificaciones con el fin de quedar plegadacorrectamente en presencia de chaperonas.
. Las proteınas modificadas se encapsulan envesıculas de transferencia que abandonan el RER ymigran hacia el aparato de Golgi, donde ser�ansometidas a otras modificaciones y se realizar�a elempaquetamiento final.
. Las proteınas con plegamiento incorrecto regresanal RE a trav�es de una proteına translocadorasemejante a la que permitio su paso a este org�anulodurante el proceso de sıntesis. Estas proteınaserroneas sufren reacciones de ubiquitinacion y sondestruidas por los proteasomas en el citoplasma.
14
2
CITO
PLASMA

15
2
CITO
PLASMA
CONSIDERACIONES CL�INICAS
La secuencia de amino�acidos de una proteınadetermina su estructura primaria. Por lo general, lasalteraciones leves de dicha estructura no tienenconsecuencias funcionales; en algunos casos; sinembargo, las mutaciones puntuales –es decir, la
sustitucion de un unico amino�acido por otro–transforman radicalmente el funcionamiento de laproteına afectada. Un ejemplo de ello es la presencia de
este tipo de mutaciones perjudiciales en la mol�eculade la hemoglobina, en la que un residuo de glutamina
que suele ocupar la sexta posicion de la cadena b essustituido por una valina, lo que origina un trastornoconocido como anemia falciforme. Cuando la tension
de oxıgeno es baja, como sucede despu�es de practicaruna actividad fısica vigorosa, los eritrocitos se deformandebido a la presencia de la cadena b modificada paraadoptar una morfologıa falciforme que reduce en granmedida su capacidad de transporte de oxıgeno. Estoseritrocitos anomalosmuestran una acusada tendencia ala fragmentacion al perder su flexibilidad normal.
Figura 2.7 La sıntesis de proteınas citosolicas tiene lugar en el citoplasma. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology,
3rd ed. Philadelphia, Saunders, 2007, p 26.)
Figura 2.8 La sıntesis de proteınas que se empaquetar�an tiene lugar en la superficie del RER. (Tomado de Gartner LP, Hiatt JL: Color
Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 27.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

16
2
CITO
PLASMA
Aparato de Golgi
El aparato de Golgi (complejo de Golgi) consta deun conjunto de s�aculos orientados y una serie devesıculas convexas aplanadas y apiladas unidas a lamembrana, cada una de las cuales remeda un pan depita sin cortar con una luz central, la cisterna (fig.2.9). La c�elula puede poseer uno o m�as complejosde Golgi, cada uno de los cuales presenta:
. Cara convexa de entrada cercana al nucleo,conocida como la red del Golgi cis (RGC)
. Cara cis, a trav�es de la cual las proteınasreci�en sintetizadas en el RER llegan al aparato deGolgi
. Cara concava de salida, orientada hacia lamembrana celular, llamada cara trans
. Una om�as caras intermedias que se disponen entrelas caras cis y trans
. Un complejo de vesıculas y s�aculos, que reciben elnombre de complejo tubulovesicular (CTV,anteriormente RECIG) y se localizan entre la cara detransicion del RER y la red del Golgi cis
. Otro grupo de vesıculas, asociadas a la cara trans,bautizado como red del Golgi trans (RGT)
Entre las funciones del aparato de Golgi figuranla sıntesis de hidratos de carbono y el transporte y lasecrecion de proteınas
Tr�afico de proteınas
Las vesıculas que transportan mol�eculas (p. ej., pro-teınas o hidratos de carbono) desde un org�anulo hastaotro o bien entre distintas regiones de un mismoorg�anulo reciben el nombre de vesıculas de trans-porte,mientras que el material transportado se deno-mina cargo. Las vesıculas de transporte est�an dotadasde una envoltura proteica (vesıculas recubiertas) ensu cara citosolica que permite que se desprendan yadhieran al org�anulo diana. Se han identificado trestipos principales de proteınas de envoltura (adem�asde algunos subtipos) utilizadas por las c�elulas para eltr�afico de proteınas:
. Coatomero I (COP I)
. Coatomero II (COP II)
. Clatrina
Las cubiertas formadas por estas proteınasaseguran la inclusion de las mol�eculas correctasen la vesıcula, ası como la creacion de una vesıculamembranosa de tamano y morfologıa adecuadas.Cada tipo de cubierta se vincula con un tipo especıficode transporte (fig. 2.10). La envoltura de la vesıcularecubierta se desprende al alcanzar la membrana delorg�anulo diana y se fusiona con ella. El reconoci-miento de la vesıcula y esta membrana depende delas llamadas proteınas SNARE (receptor del factor de
la proteına de fusion soluble sensible a N-etilmalei-mida) y un grupo de GTPasas especializadas en elreconocimiento de dianas, conocidas como Rab. Lasproteınas SNARE unicamente permiten la union de lavesıcula correcta con su org�anulo diana. La proteınaRab interviene en el anclaje inicial de la vesıcula. Lamembrana celular presenta ciertas regiones en las queabunda las proteınas SNARE, conocidas como poro-somas, a las que se fijan las vesıculas para vaciarse enel espacio extracelular.
Las proteınas abandonan el RE de transicion, unaregion del RER exenta de ribosomas, en pequenasvesıculas de transporte cuya membrana provienedel RER y est�a recubierta de COP II (v. fig. 2.10). Estasvesıculas recubiertas de COP II se dirigen al complejotubulovesicular, se desprenden de la envoltura deCOP II y se fusionan con este. La c�elula posee unmecanismo de control del contenido de estas vesıcu-las, de modo que la deteccion de alguna mol�eculapropia del RE supone su devolucion a este org�anuloen vesıculas recubiertas de COP I (transporte retro-grado); las mol�eculas cargo restantes se transportan alaparato de Golgi en vesıculas recubiertas de COP I(transporte anterogrado). Las proteınas atraviesanlas distintas caras del aparato de Golgi, posiblementeen vesıculas recubiertas de COP I, en cada una de lascuales sufrir�an diversas modificaciones, y se dirigen ala RGT para su empaquetamiento final. Las proteınasmodificadas se almacenan en vesıculas recubiertas declatrina o COP II, en las que son transportadas hacia:
. La membrana celular, en la que se insertan comoproteınas de membrana o bien se fusionan con lamembrana para secretar su contenido al espacioextracelular (exocitosis continua)
. Las proximidades del plasmalema, dondepermanecen temporalmente como vesıculas dealmacenamiento (secrecion) a la espera de laliberacion final de su contenido en el espacioextracelular (exocitosis discontinua)
. Los endosomas tardıos, los cuales se convierten enlisosomas
El proceso de exocitosis discontinua se relacionacon la envoltura de clatrina y sigue la ruta secretoraregulada de las proteınas, mientras que la exocitosiscontinua cuenta con vesıculas recubiertas de COP II ysigue la vıa secretora constitutiva de las proteınas.
Adem�as de las proteınas que conforman suenvoltura, estas vesıculas de transporte poseen ungran numero de marcadores de membrana quehacen posible su anclaje a los microtubulos para sutransporte mediante motores moleculares a lo largode estos hasta su destino final. Asimismo, las vesıculaspresentanmarcadores que actuan como etiquetas pos-tales, ya que las utilizan para anclarse a los org�anulosdiana.

17
2
CITO
PLASMA
Figura 2.10 Tr�afico de proteınas en el aparato de Golgi y vesıculas asociadas. (Tomado deGartner LP, Hiatt JL: Color Textbook of Histology,
3rd ed. Philadelphia, Saunders, 2007, p 30.)
Figura 2.9 Retıculo endopl�asmico rugoso y aparato de Golgi. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.
Philadelphia, Saunders, 2007, p 28.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Tr�afico de membrana
ENDOCITOSIS: FAGOSOMAS Y VESICULAS PINOCITICAS
Se denomina endocitosis al proceso de transportede material desde el espacio extracelular hacia elcitoplasma.
. Las mol�eculas de mayor tamano se fagocitanen una vesıcula llamada fagosoma.
. Las mol�eculas m�as pequenas (ligandos) sepinocitan en una vesıcula pinocıtica.. Lapinocitosis es unproceso sometido a un estrictocontrol en el que ciertas proteınas receptoras decargo localizadas en la membrana celularreconocen las mol�eculas a englobar alinteraccionar con el ligando en el espacioextracelular y con clatrina en el citoplasma.
. La capacidad de reconocimiento y union a lasmol�eculas de clatrina induce la formacion de unavesıcula de pinocitosis, la cual puede contenercientos de mol�eculas de ligando.
. De igual modo, las c�elulas pueden transportarcompuestos desde el citoplasma hacia el espaciointercelular a trav�es de un proceso denominadoexocitosis.
. Durante la endocitosis se desprenden fragmentosde la membrana plasm�atica que pasan a formarparte de las vesıculas procedentes de la mismay se incorporan las membranas de las vesıculasderivadas de la RGT en el transcurso de laexocitosis. Este reciclaje continuo de lasmembranas recibe el nombre de tr�aficode membrana (fig. 2.11).
ENDOSOMAS (COMPARTIMENTO ENDOSOMICO)
Las vesıculas de pinocitosis se desprenden de la envol-tura de clatrina y se fusionan con:
. Endosomas tempranos, unas vesıculasmembranosas situadas en la periferia celularcuya membrana posee bombas de H+
dirigidas por ATP que reducen el pH de su luzhasta un valor de 6.
. En algunos endosomas tempranos, los endosomasde reciclaje, el ligando y el receptor se disocian, demodo que este regresa a la membrana celular yaquel pasa al citoplasma o bien se transfiere a
. Endosomas tardıos, otros compartimentosdotados de membrana m�as alejados de la periferia.Las bombas deH+ situadas en lamembrana de estosendosomas provocan una reduccion de la luz de lasvesıculas, en las que prosigue el proceso de digestionde sus contenidos y los compuestos parcialmentedegradados se transportan hasta los lisosomas parasu destruccion final.
LISOSOMAS (ENDOLISOSOMAS)
Los lisosomas son unas pequenas vesıculas membra-nosas que contienen un gran numero de enzimashidrolıticas activas a un pH interno bajo, de 5, el cualse mantiene a trav�es de bombas de H+ presentes en sumembrana. En los lisosomas se digieren diversos com-puestos, de modo que sus productos de degradacionpasan al citoplasma y los derivados toxicos permane-cen en el interior de estas vesıculas, que se transfor-man en cuerpos residuales.
PEROXISOMAS
Los peroxisomas presentan una morfologıa seme-jante a la de los lisosomas, aunque albergan nume-rosas enzimas oxidativas sintetizadas en ribosomascitosolicos que posteriormente son transportadas aestos org�anulos mediante senales especıficas paraperoxisomas que se unen a ciertos receptores situa-dos en la membrana del peroxisoma.
. La enzima m�as abundante en este org�anulo es lacatalasa, la cual cataliza la escision del H2O2 enagua y oxıgeno. Por otra parte, el peroxisomainterviene en la biosıntesis de los lıpidos, enespecial del colesterol; el catabolismo lipıdicoa trav�es de la b-oxidacion de los �acidos grasos decadena larga; y la formacion de �acidos biliares enlos hepatocitos.
. En el sistema nervioso central, los rinones, lostestıculos y el corazon, los peroxisomas portanenzimas que participan en la sıntesis deplasmalogeno, un fosfolıpido de membrana queconfiere proteccion a la c�elula frente al oxıgenomolecular.
PROTEASOMAS
Los proteasomas son unos pequenos org�anulos enforma de tonel que intervienen en:
. La degradacion de las proteınas plegadasincorrectamente, danadas, desnaturalizadas oportadoras de alguna anomalıa.
. La escision de proteınas antig�enicas en fragmentosde menor tamano conocidos como epıtopos(v. capıtulo 12).
La proteolisis mediada por los proteasomas est�asometida a un estricto control celular a trav�es de laadicion de numerosas copias de ubiquinona, unareaccion que requiere energıa, a la proteına indicadapara generar una proteına poliubiquinada. Lasmol�eculas de ubiquitina y los productos de su degra-dacion pasan al citoplasma con gasto de energıa.
18
2
CITO
PLASMA

CONSIDERACIONES CL�INICAS
El sındrome de Zellweger es una enfermedad
cong�enita incurable de consecuencias mortales
que afecta a reci�en nacidos; la muerte sobrevienea lo largo del ano siguiente por insuficiencia
hep�atica, insuficiencia respiratoria o ambas. Laenfermedad se vincula con la incapacidad delos peroxisomas de incorporar enzimasperoxisomicas debido a la ausencia de receptoresde senales peroxisomicas en la membrana deestas vesıculas. Como consecuencia de ello, losperoxisomas no llevan a cabo la b-oxidacion de
los �acidos grasos de cadena larga para sintetizarplasmalogenos.
19
2
CITO
PLASMA
Figura 2.11 Endocitosis, endosomas y lisosomas. CURL, compartimento para el desacoplamiento del receptor y el ligando. (TomadodeGartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 33.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

20
2
CITO
PLASMA
Mitocondrias
Las mitocondrias son unos org�anulos de grantamano; algunas presentan una longitud de 7mm yun di�ametro de 1mm. La vida media de una mitocon-dria se situa en unos 10 dıas, despu�es de los cualesaumenta su longitud y sufre un proceso de biparti-cion. La mitocondria consta de:
. Una membrana externa lisa
. Una membrana interna replegada en estructurasaplanadas o tubulares, llamadas crestas, queincrementan notablemente el �area superficial de lamembrana interna.
La funcion principal de lamitocondria es la sıntesisde ATP mediante un proceso conocido como fosfo-rilacion oxidativa. Las membranas mitocondrialesdelimitan dos espacios (fig. 2.12B):
. Espacio intermembranoso, el cual se localiza entrelas membranas externa‘ e interna, y
. Espacio (intersticial) de la matriz, definido por lamembrana interna (fig. 2.12A), el cual alberga lamatriz, un lıquido viscoso que contiene numerosasproteınas, ribosomas, ARN, ADN circular (quesolamente codifica 13 proteınas mitocondriales)y gr�anulos densos de fosfolipoproteınas,denominados gr�anulos de la matriz, que podrıanunirse a iones de calcio y de magnesio.
Las membranas interna y externa aparecen conec-tadas en algunas zonas, en las que diversas proteınasreguladoras y transportadoras facilitan el movimientode distintasmol�eculas hacia los distintos compartimen-tosmitocondriales. Lasmacromol�eculas cuyodestino esalguna de las dos membranas mitocondriales o lamatriz se transportan a trav�es de las regiones membra-nosas que no se comunican entre sı, dotadas de recep-tores capaces de reconocer dichas macromol�eculas.
. La membrana mitocondrial externa es lisa yrelativamente permeable a iones de pequenotamano; las abundantes porinas presentes en ellahacen posible el movimiento de H2O a trav�es dela misma. La composicion del espaciointermembranoso se asemeja, en gran medida, a ladel citoplasma.
. La membrana interna replegada presenta un grannumero de cardiolipinas, unos fosfolıpidos queposeen cuatro cadenas de �acidos grasos, en lugar dedos, lo que reduce notablemente su permeabilidadfrente a protones y electrones. En esta membranaabunda, asimismo, el complejo enzim�aticosintetasa de ATP, implicada en la produccion deATP a partir de ADP y fosfato inorg�anico.. La sintetasa de ATP consta de dos porciones, F0y F1; la porcion F0 se encuentra enterrada, en sumayor parte, en la membrana interna, mientrasque la porcion F1 (tambi�en conocida como
cabeza) se halla suspendida en la matriz, seconecta con la porcion F0 a trav�es del tallo detransicion y se estabilizamediante otras proteınas(v. fig. 2.12B).
. Cada porcion F0 presenta tres sitios defosforilacion de ADP en ATP. La porcion F1 poseeunamanga externa fija y unamanga internamovilformada por 10 a 14 subunidades. Igualmente, eltallo cuenta con una manga interna movil quecomunica con la porcion F0 y una manga externafija.
. El conjunto de las mangas moviles del tallo y laporcion F1 recibe el nombre de rotor. Las mangasexternas fijas est�an conectadas con la porcion F0 ylos tres componentes conforman el llamadoestator.
Lamatriz contiene las enzimas que procesan el piru-vato obtenido en la glucolisis y los �acidos grasosderivados de lıpidos que se transportan a este compar-timento para convertirlos en acetil-coenzima A (CoA),cuyo grupo acetilo interviene en el ciclo del �acido cıtricopara reducir mol�eculas de dinucleotido de nicotina-mida adenina (NAD+) oxidadas a NADH y dinucleo-tido de flavina adenina (FAD) en FADH2. Estasmol�ecu-las reducidas actuan como aceptores de electrones dealta energıa procedentes del ciclo del �acido cıtrico y lostransfieren a una serie de proteınas integrales de lamembrana interna que configuran la cadena de trans-porte de electrones (fig. 2.12C). Los electrones pasan alo largo de esta cadena y su energıa se emplea paratransportarH+ (es decir, protones) desde lamatriz haciael espacio intermembranoso. A medida que la concen-tracion de H+ en este espacio aumenta con relacion a lamatriz, el gradiente de concentracion (fuerzamotriz deprotones) ası creado impulsa el retorno de los protoneshacia este compartimento a trav�es de la unica vıa posi-ble para ello, la sintetasa de ATP.
El movimiento de protones hacia el rotor de lasintetasa de ATP hace que gire en contacto con elestator, de modo que se genera energıa queaprovechar�an los tres sitios de la porcion F0 paracatalizar la fosforilacion de ADP en ATP, un com-puesto rico en energıa. Algunas de las mol�eculas deATP ası formadas ser�an utilizadas por la mitocon-dria, si bien la mayor parte de ellas se transportar�aal citoplasma para satisfacer las necesidades energ�eti-cas de la c�elula.
Lagrasapardaabundaespecialmente en losanimalesque hibernan. Lasmitocondrias de sus lipocitos poseentermogeninas en lugar de sintetasa de ATP. Aunqueestas enzimas pueden desviar a los protones del espaciointermembranoso hacia la matriz, las reacciones oxida-tivas que tienen lugarenestas c�elulasnoest�anacopladasa la fosforilacion y la fuerzamotriz de protones producecalor en lugar de ATP. El calor se utiliza para despertar alanimal del estado de hibernacion.

CONSIDERACIONES CL�INICAS
Las miopatıas mitocondriales son trastornoshereditarios por vıa materna, ya que lasmitocondrias provienen del ovulo. Estasinfrecuentes miopatıas afectan por igual a ambossexos. Su pronostico depende de los gruposmusculares afectados. Pueden manifestarse condebilidad muscular y cansancio tras el ejercicio,
aunque las variantes m�as graves pueden tenerconsecuencias mortales. Por lo general, eltrastorno semanifiesta hacia el final de la segunda
d�ecada de vida. Algunasmiopatıas frecuentes sonel sındrome de Kearns-Sayre, la epilepsiamioclonica y la encefalomiopatıa mitocondrial.No existe ningun tratamiento conocido para estegrupo de enfermedades.
21
2
CITO
PLASMA
Figura 2.12 A. Representacion tridimensional de una mito-
condria con crestas aplanadas. B. Diagrama de las crestas enparrilla a gran aumento. C. Diagrama de la cadena de trans-
porte de electrones y la sintetasa de ATP en la membrana
mitocondrial interna. (Tomado de Gartner LP, Hiatt JL: Color
Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007,p 39.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Inclusiones y citoesqueleto
INCLUSIONES
Las inclusiones son elementos celulares inertesque flotan libremente en el citoplasma y no est�an limi-tados pormembranas. Las inclusionesm�as abundantesson de glucogeno, lıpidos, pigmentos y cristales.
. Habitualmente, el glucogeno se almacena en elcitoplasma en rosetas de partıculas b, las cualesaparecen proximas a elementos del RER. Estaspartıculas constituyen depositos de energıa que sesometer�an a la glucogenolisis para obtener glucosa,la cual se transformar�a en mol�eculas de piruvato,que pasar�an al ciclo del �acido cıtrico.
. Los lıpidos se almacenan en forma de triglic�eridos,cuya degradacion origina �acidos grasos que secatabolizan en el ciclo del �acido cıtrico para formarpiruvato. Los lıpidos constituyen una forma m�aseficaz de almacenamiento de energıa que elglucogeno, ya que el ATP generado por 1 g delıpidos duplica la cantidad obtenida a partir de 1 gde aquel.
. Por lo general, los pigmentos carecen de actividadmetabolica, si bien pueden desempenar unafuncion protectora, como la melanina presente enla piel, la cual confiere proteccion a las c�elulasepid�ermicas frente a lesiones en el ADN de suscromosomas al absorber la radiacion ultravioleta.La melanina tambi�en participa en la funcion visualde la retina. Otro pigmento, la lipofucsina, podrıaformarse por fusion de varios cuerpos residuales, lasvesıculas limitadas por membrana que contienenlos residuos no degradables de la actividadlisosomica.
. Las c�elulas de mamıfero no suelen contenercristales, aunque las c�elulas de Sertoli de lostestıculos presentan, a menudo, cristales deCharcot-Bottscher, cuya funcion, de haberla,no se ha definido aun.
CITOESQUELETO
El citoesqueleto, la red tridimensional que sostienea la c�elula, est�a formado por microtubulos, filamen-tos delgados y filamentos intermedios. Adem�as delmantenimiento de la morfologıa celular, esta redinterviene en la adhesion intercelular, el movimientode la c�elula a lo largo de elementos del tejido conjun-tivo, y facilita la exocitosis, la endocitosis y el tr�afico demembrana en el citoplasma. El citoesqueleto participaen la creacion de compartimentos celulares en los quese concentran ciertos sistemas enzim�aticos con el finde potenciar la consecucion de ciertas reacciones bio-quımicas.
. Losmicrotubulos son estructuras cilındricas largas,flexibles y huecas formadas por heterodımeros detubulina a y b (fig. 2.13A). Los dımeros de tubulinase disponen para formar unos elementos linealesdependientes de GTP conocidos comoprotofilamentos; la asociacion de13 protofilamentos da lugar a un microtubulocilındrico de 25nm de di�ametro con una luz de15nm de di�ametro. Cada microtubulo poseeun extremo positivo a partir del cual crece y unextremo negativo que tiende a acortarse a no serque se encuentre incluido en unas estructurasanulares formadas por tubulina g. La estabilizaciondel extremo positivo se logra a trav�es de unacaperuza de proteınas especıficas asociadas amicrotubulos (MAP), las cuales impiden supolimerizacion. Los microtubulos est�anpolarizados. Pueden crecer (en un procesodenominado rescate) o bien acortarse (en unproceso denominado cat�astrofe); esta actividadcıclica se conoce como inestabilidad din�amica.. Otras MAP actuan comomotores moleculares, lasproteınas cinesina y dineına, ya que permiten eltransporte de material celular a lo largo dedistancias considerables sobre los microtubulos,que representarıan autopistas celulares, hacia elextremo positivo (cinesina) o el negativo(dineına).
. Algunas MAP intervienen como elementosespaciadores de microtubulos cercanos; porejemplo, MAP2 aleja a un microtubulo de otro,mientras que otras, como tau, facilitan suacercamiento.
. Generalmente, los extremos negativos dela mayorıa de los microtubulos proceden de lamisma region celular, el centrosoma o centroorganizador de microtubulos (COMT) de lac�elula. Los microtubulos mantienen lamorfologıa celular, facilitan el transporteintracelular, forman el huso mitotico y el husomeiotico, constituyen el eje citoesquel�etico decilios y flagelos, y conforman los centrıolos ylos corpusculos basales.
. Los centrıolos son dos pequenas estructurascilındricas, cada una de las cuales est�a constituidapor nueve tripletes de microtubulos y con unadisposicion perpendicular respecto al otrocentrıolo (fig. 2.13D). Cada integrante del par decentrıolos se replica a lo largo de la fase S del ciclocelular. Los centrıolos conforman el centrosoma yactuan como centros de organizacion del aparatodel huso en el transcurso de la division celular.Asimismo, constituyen los corpusculos basalesque dirigen la organizacion de los cilios y losflagelos.
22
2
CITO
PLASMA

23
2
CITO
PLASMA
Figura 2.13 Esquemas tridimensionales de los distintos compo-
nentes del citoesqueleto. A.Microtubulo. B. Filamento delgado.
C. Filamento intermedio.D. Centrıolo. (Tomado de Gartner LP,Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders,
2007, p 43.)
CONSIDERACIONES CL�INICAS
TRASTORNOS RELACIONADOSCON LOS DEPOSITOS DE GLUCOGENOAlgunas personas presentan trastornosrelacionados con depositos de glucogeno al ser
incapaces de degradar esta mol�ecula, lo que da
lugar a su acumulacion excesiva en las c�elulas.Se distinguen tres variantes: 1) afectacion
hep�atica; 2) afectacion miop�atica, y
3) miscel�anea. La enfermedad se debe a laausencia o el funcionamiento incorrecto dealguna de las enzimas implicadas en ladegradacion.
TRASTORNOS RELACIONADOSCON LA MELANINALos sujetos que carecen de la capacidad desintetizar melanina, por lo general debido a unamutacion en el gen que codifica la enzima
tirosinasa,muestran una coloracion cut�aneamuy
p�alida y ojos rojos. Estas personas presentanalbinismo. Los individuos que producen unacantidad de melanina mayor de lo normal
presentan una tez m�as oscura y placas
escamiformes de color oscuro. Se dice que est�anafectados por un trastorno conocido comoictiosis laminar. Otros sujetos carecen de
melanocitos, las c�elulas responsables de lasıntesis de melanina. Presentan vitıligo.
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

. Los filamentos delgados (microfilamentos) est�anformados por monomeros de actina G que seorganizan en dos cadenas enrolladashelicoidalmente de actina F que configuran unfilamento polarizado de 7nm de di�ametro enun proceso que requiere ATP (fig. 2.14B). Losmonomeros y los filamentos de actina representan,aproximadamente, el 15% del contenido proteicode la c�elula, por lo que constituye una de lasproteınas intracelulares m�as abundantes. Demanera similar a los microtubulos, losmicrofilamentos poseen un extremo positivo(romo debido a la presencia del sitio de union demiosina) y un extremo negativo (afilado debido ala ausencia de dicho sitio). El ritmo depolimerizacion del filamento es m�as r�apido en elextremo positivo.. Cuando un filamento delgado alcanza la longitudindicada, sus dos extremos se recubren conproteınas de coronamiento, como gelsolina, lascuales estabilizan ambos extremos al impedir supolimerizacion o despolimerizacion. La proteınagelsolina interviene, adem�as, en la division de unmicrofilamento en dos y la adicion de unacaperuza a los extremos ası creados.
. La accion de la proteına cofilina, que induce ladespolimerizacion mediante la eliminacion demonomeros de actina G en el extremo negativo,provoca el acortamiento de los filamentosdelgados. Su polimerizacion depende de lapresencia de un gran numero de monomeros deactina G, que son secuestrados por la proteınatimosina en el citoplasma y transferidos desdeesta al extremo positivo del microfilamento por laproteına profilina.
. Este complejo proteico controla la ramificacion delos filamentos delgados, ya que regula la unionde monomeros de actina G a los microfilamentosexistentes. Laprofilina cataliza lapolimerizaciondela ramificacion a partir de ese sitio. Las asociacionesque crean los microfilamentos al interaccionarentre sı se dividen en haces contr�actiles, redessimilaresagel yhacesparalelos.Asimismo, laactinainterviene en el establecimiento y elmantenimiento de los contactos focales de la c�elulacon la matriz extracelular.
. Los haces contr�actiles se vinculan con variostipos demiosina, de lamiosina I a lamiosina IX, yest�an implicados en los procesos de contraccion,
la contraccion muscular y el movimientointracelular de mol�eculas.
. Las redes similares a gel se relacionan con laproteına filamina y conforman matrices muyviscosas, como las que se encuentran presentesen la corteza celular.
. Los haces paralelos son filamentos delgadosasociados a las proteınas villina y fimbrina.Conservan la disposicion paralela de losmicrofilamentos, como sucede en el haz internode las microvellosidades y las microespıculas y enla red terminal.
. Los filamentos intermedios son unoscomponentes citoesquel�eticos filiformes con undi�ametro comprendido entre 8 y 10nm quemantienen la estructura celular, anclan el nucleoen su posicion, unen a las proteınas integrales demembrana al citoesqueleto y reaccionan ante losmovimientos de la matriz extracelular. Estosfilamentos (fig. 2.14C) se componen de tetr�amerosformados por proteınas cilındricas, ocho de lascuales se organizan en h�elices densas deprotofilamentos. La asociacion de dos de estosorigina protofibrillas, que se organizan en gruposde cuatro para formar un filamento intermedio.Se han identificado alrededor de 40 tipos defilamentos intermedios con arreglo a sucomposicion proteica y su distribucion celular.Las clases m�as comunes son los filamentos dequeratina, los filamentos de desmina, losfilamentos de vimentina, los filamentos deproteına acıdica fibrilar glial, los neurofilamentosy los filamentos de la l�amina nuclear. Lasproteınas de union a los filamentos intermediosse anclan a los filamentos intermedios, a los queunen entre sı para facilitar la formacion de la redtridimensional del citoesqueleto. Las proteınasmejor conocidas de este sistema son filagrina,sinemina, plectina y las plaquinas.. Las filagrinas unen filamentos de queratina enhaces.
. La sinemina se une a la desmina,mientras que laplectina lo hace a la vimentina, para crear una redtridimensional en el compartimentocitopl�asmico.
. Las plaquinas anclan los filamentos de queratinaa los hemidesmosomas de las c�elulas epiteliales ylos neurofilamentos a los filamentos delgados enlas neuronas de los ganglios dorsales.
24
2
CITO
PLASMA

25
2
CITO
PLASMA
Figura 2.14 Esquemas tridimensionales de los distintos componentes del citoesqueleto. A. Microtubulo. B. Filamento delgado.
C. Filamento intermedio. D. Centrıolo. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders,
2007, p 43.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

3 NUCLEOEl nucleo, el org�anulo celular de mayor tamano, nosolamente contiene el ADN de la c�elula, sino quetambi�en alberga la maquinaria responsable de la sın-tesis del ADN y el ARN. El nucleoengloba tres compartimentos princi-pales: la cromatina, el material gen�e-tico de la c�elula; el nucl�eolo, en elque tiene lugar la sıntesis del ARNribosomico (ARNr) y el ensamblajede las subunidades ribosomicas; y elnucleoplasma, una matriz formadapor diversas macromol�eculas y partı-culas nucleares. El nucleo se rodea dela envoltura nuclear, integrada pordos membranas. A pesar de que lamorfologıa, la localizacion y eltamano del nucleo difieren en distin-tos tipos celulares, en la mayorıa delas c�elulas adopta una posicion cen-tral y es esf�erico.
Envoltura nuclear
La envoltura nuclear est�a formada por las membra-nas nucleares internas y externas, que se separanpor la cisterna perinuclear (10 a 30nm de grosor) ypresenta unas perforaciones conocidas como porosnucleares, unas regiones en las que ambas membra-nas se fusionan y a trav�es de las cuales se inter-cambian sustancias entre el nucleo y el citoplasma(fig. 3.1).
. La membrana nuclear interna, cuyo grosor esde unos 6 nm, est�a en contacto con la l�aminanuclear, una malla de filamentos intermediosformada por las l�aminas A, B y C y localizadaen la periferia del nucleo. Adem�as de organizary mantener a la cromatina perinuclear y lamembrana nuclear interna, estas l�aminasintervienen en el ensamblaje de la envolturanuclear tras la division celular. Las proteınastransmembrana de la membrana nuclear interna,que suelen asociarse a proteınas de la matriz,actuan como sitios de contacto para mol�eculasde ARN y cromosomas nucleares.
. Lamembrana nuclear externa, de 6nm de grosor ya la que se adosan ribosomas, se continua con elretıculo endopl�asmico rugoso; su superficiecitopl�asmica est�a rodeada por una malla devimentina (filamentos intermedios).
POROS NUCLEARES Y COMPLEJOS DEL PORO NUCLEAR
Los poros nucleares aparecen en regiones de la envol-tura en las que se fusionan las membranas interna y
externa para permitir la comunicaciondel nucleo con el citoplasma. En la peri-feria del poro nuclear aparecen gluco-proteınas que intervienen en la forma-cion del complejo del poro nuclear. Loscomplejos se comunican entre sı a trav�esde la l�amina nuclear para permitir elpaso de mol�eculas a trav�es de los poros.
. El complejo del poro nuclear constade tres estructuras anulares denaturaleza proteica, con simetrıaoctam�erica, conectadas entre sı porrayos en disposicion vertical y queatraviesan ambas membranas(di�ametro, 100 a 125nm).
. Los tres anillos superpuestos recibenel nombre de anillo citopl�asmico,anillo luminal y anillo nuclear.
Adem�as, existe una canastilla nuclear en la caranuclear de cada complejo del poro (fig. 3.2).
. En el reborde de la porcion citopl�asmica del poronuclear se halla el anillo citopl�asmico, que constade ocho subunidades, cada una de las cuales poseeun filamento citopl�asmico formado por unaproteına de union a Ran (proteına de union a GTP)que facilita el transporte de mol�eculas desde elcitoplasma hacia el nucleo.
. Otro grupo de ocho proteınas transmembrana que seproyectan hacia la luz del poro y la cisternaperinuclear conforman el anillo luminal (anillomedio), cuya luz central podrıa corresponder a uncanal con compuerta que limitarıa la difusion pasiva.Otras proteınas asociadas al complejo intervienen enla regulacion del transporte a trav�es del poro nuclear.
. En ocasiones puede visualizarse una estructuraovalada, el transportador, en la luz central. Esposible que este elemento corresponda amaterial queest�a siendo transportado hacia el nucleo o fuera de �el.
. El anillo nuclear (anillo nucleopl�asmico) selocaliza en el borde de la cara nuclear del complejodel poro, integrado tambi�en por ocho subunidades.Este anillo interno participa en la exportacion demol�eculas de ARN hacia el citoplasma.
. Suspendida del anillo nuclear est�a la canastillanuclear, una estructura filamentosa flexible en formadecesta, yunanillodistal,unidoa laporciondistal dela canastilla, aparecen suspendidos del anillo nuclear.
� 2011 Elsevier Espana, S.L. Reservados todos los derechos
TERMINOS CLAVE. Complejo del poro
nuclear. Cromosomas. Acido
desoxirribonucleico(ADN)
. Acido ribonucleico(ARN)
. Ciclo celular
. Mitosis
. Meiosis
. Apoptosis
26

27
3
NUCLEO
Figura 3.2 Complejo del poro nuclear. (Tomado de
Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.Philadelphia, Saunders, 2007, p 54.)
Figura 3.1 Diagrama de un nucleo tıpico. (Tomado de Gartner
LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia,
Saunders, 2007, p 52.)
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

Funcionamiento del poro nuclear
El tamano de la luz del complejo del poro nuclear sereduce como consecuencia de la presencia de las pro-teınas que lo integran, de tal modo que las mol�eculasmayores de 11 nm no pueden atravesarlo en ningunsentido salvo en un proceso de transporte mediadopor receptores, dependiente de energıa.
. Unos receptores, las importinas y las exportinas,del complejo del poro nuclear han de reconocer lassecuencias senal que portan los compuestos atransportar; la regulacion del transporte depende denucleoproteınas asociadas a Ran y al complejo delporo nuclear.
. Las importinas presentan senales de localizacionnuclear.
. Las exportinas poseen senales de exportacionnuclear.
Un ejemplo del funcionamiento de las importinases el transporte de las proteınas que integran los ribo-somas hacia el citoplasma, mientras que la transferen-cia de ciertas macromol�eculas, como el ARN, haciadicho compartimento ilustra la funcion de las expor-tinas (fig. 3.3).
Cromatina
El material gen�etico (ADN) de la c�elula se encuentraen el nucleo y forma parte de los cromosomas, unasestructuras tan compactadas a lo largo de la mitosisque pueden visualizarse en la microscopia optica,pero durante las restantes etapas del ciclo celular sedesenrollan hasta convertirse en unas hebras laxas decromatina.
. La mayor parte de la cromatina nuclear est�aparcialmente descondensada, corresponde a �areasinactivas desde el punto de vista de la transcripcion yse localiza en la periferia nuclear; recibe el nombrede heterocromatina.
. La cromatina activa, conocida como eucromatina,est�a descondensada y representa una hebra de ADNde 2nm de grosor enrollada alrededor de unascuentas, los nucleosomas, cuya trascripcion dar�alugar al ARN.. Cada nucleosoma es un complejo octam�ericoformado por proteınas, las histonas (H2A, H2B,H3 y H4), alrededor del cual se enrollan dosvueltas de la cadena del ADN, que equivalen aunos 150 pares de nucleotidos.
. ElADN espaciador comprende unos 200 pares debases que separan nucleosomas contiguos. Estasestructuras mantienen la hebra del ADN yparticipan en la regulacion de la replicacion, lareparacion y la transcripcion del ADN.
. La cromatina se enrolla helicoidalmente paraformar una fibra de 30 nm que contiene seisnucleosomas por vuelta de h�elice y seestabilizan por la interaccion con la histona H1
(fig. 3.4).
CROMOSOMAS
Las fibras de cromatina se condensan antes delcomienzo de la mitosis o la meiosis para formar loscromosomas, que alcanzan el m�aximo grado deempaquetamiento durante la metafase (v. fig. 3.4).
. Cada especie posee un numero especıfico decromosomas, denominado genoma o constituciongen�etica total.
. El genoma humano consta de 46 cromosomas:23 pares de cromosomas homologos, cada uno delpar de cada progenitor.. La especie humana posee 22 pares decromosomas som�aticos (autosomas) y un unicopar de cromosomas sexuales.
. El par de cromosomas sexuales femeninos secompone de dos cromosomas X (XX), mientrasque el parmasculino lo hace por un cromosoma Xy un cromosoma Y (XY).
28
3
NUCLEO

29
3
NUCLEO
Figura 3.4 Empaquetamiento de la cromatina para dar lugar a los cromosomas. (Tomado de Gartner LP, Hiatt JL: Color Textbook of
Histology, 3rd ed. Philadelphia, Saunders, 2007, p 55.)
Figura 3.3 Funcion de Ran en la importacion nuclear. GAP, proteına activadora de GTPasa; GDP, difosfato de guanosina; SLN, senalesde localizacion nuclear. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 54.)
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

ACIDO DESOXIRRIBONUCLEICO Y ACIDO RIBONUCLEICO
La mol�ecula de ADN se compone de dos tipos debases nitrogenadas, purinas (adenina y guanina) ypirimidinas (citosina y timidina), que se unen a dosesqueletos de desoxirribofuranosas, lo que da lugar auna secuencia lineal de nucleotidos. Entre las basescomplementarias enfrentadas se establecen enlaces dehidrogeno que conectan entre sı a las dos hebras y danlugar a la doble h�elice.
La mol�ecula de ARN es semejante a la de ADN, sibien se compone de una cadena sencilla en lugar deuna hebra doble y las bases nitrogenadas est�an unidasa esqueletos de ribofuranosa (no obstante algunosvirus poseen ARN bicatenario). Asimismo, el ARNpresenta uracilo en lugar de timidina. La sıntesis deARN se conoce como transcripcion, en la que una delas hebras del ADN actua como molde para la forma-cion de una hebra monocatenaria complementaria deARN. Se distinguen tres tipos diferentes de ARN, quese transcriben de manera similar por una polimerasade ARN especıfica para cada uno de ellos.
. El ARN mensajero (ARNm), sintetizado por lapolimerasa II, contiene informacion gen�eticatranscrita a partir del ADN para la sıntesis de unasecuencia de amino�acidos, que tendr�a lugar en elcitoplasma. La mol�ecula de ARNm representa unacopia exacta de una secuencia determinada de lamol�ecula de ADN que corresponde a un gen.
. El ARN de transferencia (ARNt), sintetizado porla polimerasa III, transporta amino�acidos activadoshasta el complejo ribosoma-ARNm encargado dela sıntesis proteica (v. apartado sobre la sıntesis deproteınas en el capıtulo 2).
. El ARN ribosomico (ARNr), sintetizado por lapolimerasa I, se fabrica en el nucl�eolo y se asocia aproteınas para formar las subunidades ribosomicas.
Transcripcion
La union de ciertos cofactores a la polimerasa II facilitael desenrollamiento de dos vueltas de la doble h�elice
del ADN con el fin de exponer los nucleotidos presen-tes en esa region.
. La polimerasa II emplea una de las hebras del ADNcomo molde para sintetizar la mol�eculacomplementaria de ARNm.
. La doble h�elice continua separ�andose conformeavanza la transcripcion, en la que la misma hebradel ADN actua como molde para formar unamol�ecula complementaria de ARN.
. La hebra de ARN crece a medida que se anadennuevos nucleotidos hasta liberarse delmoldedeADN,que recuperar�a su conformacion anterior (fig. 3.5).
Lamol�ecula transcrita de ARN (tr�anscrito primario)separada del molde de ADN se denomina ARN men-sajero precursor (ARNm-pre) y contiene elementoscodificantes (exones) y no codificantes (intrones).
. Los intrones no codificantes deben eliminarse paracrear una secuencia continua de exones.
. Las mol�eculas de ARNm-pre han de formarcomplejos con partıculas de ribonucleoproteınasnucleares heterog�eneas (RNPnh) para poner enmarcha el proceso de corte y empalme y reducir lalongitud del ARNm-pre. En este proceso intervienenotros mecanismos de procesamiento.
. En el proceso participan cinco ribonucleoproteınaspequenas nucleares (RNPpn) y muchos otrosfactores de corte y empalme diferentes de lasRNPpn que conforman los espliceosomas, los cualesproducen ribonucleoproteına mensajera (RNPm).
. Tras concluir el proceso de corte y empalme yeliminarse las proteınas nucleares, la mol�ecula deARNm atraviesa el complejo del poro nuclear parapasar al citoplasma.
A pesar de que las secuencias del ARN intronicoque se eliminan de la mol�ecula inmadura de ARNconstituyen un porcentaje mayor del ARN nuclearque los fragmentos de ARN exonicos, se cree que nodesempenan funcion alguna. Los indicios m�as recien-tes sugieren que podrıan llevar a cabo funciones deregulacion junto a proteınas reguladoras.
30
3
NUCLEO

31
3
NUCLEO
Figura 3.5 Transcripcion del ADNenARNm. (Modificado deAlberts B, BrayD, Lewis J, et al: Molecular Biology of the Cell, 3rd ed. New York,
Garland Publishing, 1994.)
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

NUCLEOPLASMA
El nucleoplasma se compone de gr�anulos de intercro-matina y pericromatina, RNPpn y la matriz nuclear(fig. 3.6).
. Gr�anulos de intercromatina (20 a 25nm dedi�ametro), que se diseminan entre la cromatina,contienen RNP y varias enzimas, como trifosfatasade adenosina (ATPasa), trifosfatasa de guanosina(GTPasa), b-glicerofosfatasa y pirofosfatasa dedinucleotidos de nicotinamida-adenina (NAD).Se ignora cu�al es su funcion.
. Gr�anulos de pericromatina (30 a 50nm dedi�ametro), rodeados de un halo de composiciondesconocida y un grosor de 25nm, se localizan en laperiferia de la heterocromatina y contienenpartıculas similares a RNPnh. Unos complejosformados por secuencias cortas de ARN y proteınas,denominados RNPpn, manipulan y transportan lasRNPnh, que est�an implicadas en el procesamientode los ARNm-pre.
Matriz nuclear
Desde el punto de vista estructural, la matriz nuclearcontiene elementos fibrilares, nucl�eolos residuales,restos de redes de RNP y el complejo integrado porla l�amina nuclear y los poros nucleares. Reciente-mente se ha identificado un retıculo nucleopl�asmicoen el seno de estamatriz que parece continuarse con elretıculo endopl�asmico del citoplasma y almacenarıacalcio para su uso por el nucleo. Asimismo, se handescubierto receptores e inositol 1,4,5-trifosfato, queregulan la senalizacion mediada por el calcio nuclear,en especial de las senales relacionadas con el trans-porte de proteınas y la transcripcion de ciertos genes.
Lamatriz nuclear puede dividirse en compartimen-tos que interaccionan entre sı para permitir la regula-
cion de la expresion de genes especıficos en momen-tos determinados, la transcripcion y el procesamientodemol�eculas de ARNt y ARNm, y la union de distintasmol�eculas de senalizacion y virus.
NUCLEOLO
El nucl�eolo, una estructura nuclear formada por ARNmuy basofilo y en la que abundan las proteınas, tansolo es visible a lo largo de la interfase. Aunque lasc�elulas contienen normalmente un unico nucl�eolo, enalgunos tipos celulares aparecen dos o tres nucl�eolos;el nucl�eolo se expande durante la sıntesis de ARNm yse asocia a una porcion de los cromosomas, la croma-tina asociada al nucl�eolo, que se transcribe para ge-nerar ARNm o ARNr. Pueden distinguirse cuatroregiones en el nucl�eolo:
. Centro fibrilar de tincion p�alida, que se caracterizapor la presencia de los extremos de los cromosomas13, 14, 15, 21, y 22 (en el ser humano), los cualesportan los genes correspondientes al ARNr.
. Parte fibrosa, en la que se transcribe el ARNnucleolar.
. Parte granulosa, la region nucleolar en la que seensamblan las subunidades ribosomicas.
. Matriz nucleolar, una red de fibras que mantienenla organizacion del nucl�eolo.
El nucl�eolo est�a implicado en el ensamblaje y laorganizacion de las subunidades ribosomicas nomitocondriales (fig. 3.7), la regulacion de determi-nados procesos del ciclo celular mediante el secues-tro o la inactivacion de ciclasas dependientes deciclinas, el ensamblaje de las RNP, la regulacion dela exportacion de compuestos desde el nucleo y,posiblemente, la regulacion del proceso de envejeci-miento.
32
3
NUCLEO

33
3
NUCLEO
Figura 3.6 Nucleo. (Tomado de Gartner LP, Hiatt JL: Color
Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 52.)
Figura 3.7 Formacion de los ribosomas. (Modificado de Alberts B, Bray D, Lewis J, et al: Molecular Biology of the Cell, 3rd ed. New York,
Garland Publishing, 1994.)�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

Ciclo celular
El ciclo celular, una secuencia de acontecimientoscelulares que preparan a la c�elula para su division,consta de la interfase, a lo largo de la cual se incre-menta su tamano y se duplica el material gen�etico, y lamitosis, un proceso que da lugar a dos c�elulas hijasid�enticas. Por lo general, se considera que el ciclocelular se inicia al concluir la division celular paradar comienzo a la interfase (fig. 3.8).
. La mitosis puede suspenderse en algunas c�elulasmuy diferenciadas (p. ej., c�elulas musculares yneuronas), que permanecerıan en la fase dereposo G0.
. Otras c�elulas, como los linfocitos perif�ericos, pasana la fase G0 de forma temporal hasta la reanudaciondel ciclo celular en un momento posterior.
Algunos acontecimientos, como fuerzas mec�anicas,la isquemia o la muerte de c�elulas de un linaje celulardeterminado, pueden inducir la liberacion de factoresde crecimiento por parte de c�elulas de senalizacion queestimulan la expresion de protooncogenes, los cualesponen en marcha las vıas proliferativas de la c�elula. Seactiva, ası, una cascadade cinasasdeproteınas citopl�as-micasque inducenuna seriede factoresde transcripcionnucleares, los cuales regulan la expresion de protoonco-genes vinculados con la division celular. Muchas neo-plasias malignas se deben a la existencia de mutacionesen protooncogenes que permiten la proliferacion des-controlada de la c�elula portadora de la mutacion.
Al unirse a cinasas dependientes de ciclinas(CDK), un grupo de proteınas llamadas ciclinas nosolamente las activan, sino que las encauzan hacia lasproteınas diana y, de este modo, rigen el ingreso y elavance de la c�elula en el ciclo celular. Existen tres tiposprincipales de puntos de control, en los cuales puedenactuar los mecanismos de control para impedir laentrada de la c�elula en el ciclo celular o interrumpirlo.En cada punto de control, la c�elula puede optar poringresar o finalizar el ciclo celular, detenerlo tempo-ralmente o bien abandonarlo. Estos puntos de controlson los siguientes:
. Punto de inicio/restriccion en la fase gap 1, el cualpermite la duplicacion de los cromosomas y el pasoa la fase gap 2;
. Punto de control G2/M, el cual pone en marchala condensacion de los cromosomas y otrosprocesos necesarios para el comienzo de lamitosis; y
. Punto de control de metafase/anafase, el cualposibilita la separacion de las crom�atidas hermanas,la finalizacion de la fase M y la citocinesis.
A continuacion, se enumeran las cuatro clases deciclinas y las CDK con las que interaccionan:
. Ciclinas G1. La ciclina D se une a CDK4 y CDK6 enuna etapa temprana de la fase G1.
. Ciclinas G1/S. La ciclina E se sintetiza a finales de lafase G1 y se une a la CDK2. Estos tres complejos,junto a otros mediadores, permiten el inicio de lafase S y su progresion.
. Ciclinas S. La ciclina A se une a CDK2 y CDK1 paraformar complejos que permiten a la c�elula salir de lafase S y pasar a la fase G2 e inducir la sıntesis dela ciclina B.
. Ciclinas M. La ciclina B forma un complejo con laciclina CDK1, el cual regula la salida de la fase G2 yel ingreso en la fase M.
Las ciclinas se degradan cuando han cumplido sumision para impedir su intervencion en otras etapas.
INTERFASE
La interfase se divide en tres fases: la fase G1(gap), a lolargo de la cual la c�elula se prepara para la sıntesis deADN; la fase S (fase de sıntesis), en la que tiene lugarla replicacion del ADN; y la fase G2, la etapa de pre-paracion para la mitosis (v. fig. 3.8).
. Fase G1. Al concluir la mitosis, las nuevas c�elulashijas entran en la fase G1 el ciclo celular, que sedistingue por la sıntesis de las proteınas reguladorasimplicadas en la replicacion del ADN, elrestablecimiento de los nucl�eolos y el volumencelular normal en la c�elula hija, y el comienzode la duplicacion de los centrıolos.
. Fase S. La fase S corresponde a una etapa de sıntesisen la que se duplica el material genomico.El complemento de ADN habitual de la c�elula seduplica de un valor de (2n) a otro de (4n) conanterioridad a la division mitotica.
. Fase G2. Se denomina ası al perıodo comprendidoentre el final de la sıntesis de ADN y el comienzo dela mitosis; a lo largo de este lapso se sintetizan ARN,tubulina y otras proteınas necesarias para la divisioncelular. Asimismo, se registra un aumento de lasreservas de trifosfato de adenosina (ATP) y severifica y se corrige cualquier error presente el ADNde nueva sıntesis.
34
3
NUCLEO

CONSIDERACIONES CL�INICAS
La quimioterapia antitumoral se ha beneficiado delos adelantos logrados en la comprension del ciclo
celular y la mitosis. Algunos f�armacos se utilizanenmomentos especıficos con el objeto de detenerla proliferacion celular mediante la alteracion deciertas etapas del ciclo celular. La vincristinaaltera el husomitotico, lo que provoca la detenciondel ciclo en la fase de mitosis. La administracionde colchicina, un alcaloide de origen vegetal,persigue este mismo fin; se utiliza en estudioscromosomicos personalizados y en lacaracterizacion del cariotipo. Elmetotrexato, queinhibe la sıntesis de las purinas, y el5-fluorouracilo, que inhibe la sıntesis depirimidinas, actuan a lo largo de la fase S del ciclocelular, impiden la division celular y forman parte
de algunas pautas quimioter�apicas.
35
3
NUCLEO
Figura 3.8 El ciclo celular en una c�elula de division activa. Las
c�elulas que no se dividen, como las neuronas, salen del ciclo
para pasar a la fase G0 (fase de reposo). Otras c�elulas, como loslinfocitos, pueden volver al ciclo celular. (Tomado de Gartner LP,
Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders,
2007, p 61.)
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

MITOSIS
La mitosis es el perıodo del ciclo celular que sucede ala fase G2 y supone la formacion de dos c�elulas hijasid�enticas, aunque de menor tamano, a partir de unac�elula madre. Inicialmente se produce la division delmaterial nuclear o cariocinesis, hacia el final de lacual tiene lugar la citocinesis o division del cito-plasma. Aunque la mitosis constituye un proceso con-tinuo, pueden diferenciarse cinco etapas: profase,prometafase, metafase, anafase y telofase (fig. 3.9).
. En la primera fase de lamitosis, laprofase, se observala condensacion paulatina de los cromosomas, ladesaparicion del nucl�eolo, el comienzo de ladesorganizacion del nucleo, y la division delcentrosoma en dos mitades que migran hacia polosopuestos de la c�elula. Cada una de estas mitadesposee un centrıolo y un centro organizador de losmicrotubulos (COMT). A medida que loscromosomas (formados por dos crom�atidashermanas unidas a nivel del centromero pormol�eculas de cohesina, una proteına de union a lacromatina) continuan condens�andose, se crea otroCOMT, el cinetocoro, y se pone en marcha laformacion del aparato del huso mitotico. El husodirige lamigracion de las crom�atidas hermanas haciapolos opuestos del nucleo.
. A lo largo de la prometafase se produce ladesaparicion de la envoltura nuclear comoconsecuencia de la fosforilacion de las l�aminasnucleares. Prosigue la condensacion de loscromosomas, que se orientan de manera aleatoriaen el citoplasma. Los microtubulos unidos a loscinetocoros, llamados microtubulos del husomitotico, y los microtubulos polares, que sedisponen entre ambos centrosomas, definen laestructura del huso mitotico. Los primerosmantienen a los cromosomas en una orientacioncorrecta, mientras que estos ultimos parecenconservar la distancia entre ambos centrosomas.
. En el transcurso de la metafase, los cromosomas,en su m�aximo grado de condensacion, se disponenen el plano ecuatorial (placa metaf�asica) del husomitotico, de tal modo que las crom�atidas
hermanas est�en orientadas en paralelo al ecuadorcelular.
. La desaparicion de las proteınas de cohesion queunen entre sı a las crom�atidas hermanas a nivel delcentromero inaugura la etapa de anafase, durante lacual comienzan a separarse las crom�atidashermanas (cromosomas). Se cree que loscromosomas desempenarıan un papel pasivo en lamigracion hacia polos opuestos de la c�elula, la cualse debe a la despolimerizacion de los microtubulosdel huso mitotico en asociacion con la dineına. Enla anafase tardıa aparece un surco de division queindica la preparacion de lamembrana plasm�atica decara a la citocinesis.
. En la telofase los cromosomas han alcanzado lospolos opuestos de la c�elula y la envoltura nuclear seorganiza de nuevo debido a la desfosforilacion delas l�aminas nucleares. Se observa ladescondensacion de los cromosomas y el desplieguede las regiones de organizacion nucleolar de cincopares de cromosomas.. A pesar de que la citocinesis (division delcitoplasma en dos mitades para formar dosc�elulas hija) se inicia a lo largo de la anafase,llega a t�ermino durante la telofase.
. A medida que el surco de division estrangula elperımetro celular, la c�elula se asemeja a unamancuerna en la que ambas esferas se encuentranproximas entre sı.
. Finalmente, los citoplasmas de ambas c�elulashijas aparecen conectados por el cuerpo medio,una estructura formada por los microtubulospolares rodeados de un delgado halocitopl�asmico.
. En cada una de las c�elulas hijas, un anillocontr�actil compuesto por actina y miosina seocupa del proceso de constriccion, que concluyecon la despolimerizacion de los microtubulos delcuerpo medio.
. El huso mitotico se despolimeriza tras laseparacion total de ambas c�elulas hijas y finalizala citocinesis.
. Las dos c�elulas hijas diploides (2n) son id�enticasentre sı.
36
3
NUCLEO

37
3
NUCLEO
Figura 3.9 Etapas de la mitosis en una c�elula con una dotacion diploide de seis cromosomas. (Tomado de Gartner LP, Hiatt JL: Color
Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 64.)
CONSIDERACIONES CL�INICAS
El estudio del cariotipo puede revelar la existencia deuna aneuploidıa (un numero anomalo decromosomas). El sındrome de Down, en el que elafectado porta una copia adicional del cromosoma 21(trisomıa del cromosoma 21), es un ejemplo de estetipo de trastornos. Los sujetos con sındrome deDown presentan manos cortas y anchas, retrasomental, malformaciones cardıacas y muchas otras
anomalıas cong�enitas. Una aneuploidıa de loscromosomas sexuales es el sındrome deKlinefelter. Esta entidad afecta a los hombres, sibien poseen una copia adicional del cromosoma X(XXY). Presentan un fenotipo masculino, pero nodesarrollan los caracteres sexuales secundarios y
suelen ser est�eriles. La existencia de un numero decromosomas menor del normal se define comomonosomıa, un ejemplo de la cual es el sındromede Turner (XO). Afecta a mujeres con retraso
mental, inmadurez ov�arica y mamaria, y tamanouterino pequeno.
Los oncogenes representan formas mutadas deunos genes normales denominadosprotooncogenes, los cuales codifican proteınas
que controlan la division celular. Losprotooncogenes utilizan cuatro mecanismos decontrol del crecimiento celular y pueden codificarfactores de crecimiento, receptores de factores de
crecimiento, mol�eculas de transduccion desenales, y factores de transcripcion nuclear. La
aparicion de un oncog�en puede deberse a una
infeccion vırica o a errores gen�eticos aleatorios.Los oncogenes ejercen una influencia dominantesobre los alelos normales y provocan la division yla proliferacion celulares descontroladas. Algunostipos tumorales debidos a la expresion de
oncogenes son el c�ancer de vesıcula y laleucemia mielocıtica aguda. El linfoma deBurkitt se relaciona con la translocacion de un
protooncog�en presente habitualmente en elcromosoma 8 al cromosoma 14, lo que provocasu separacion del elemento encargado deregularlo. Este linfoma es una enfermedad
end�emica en algunas regiones africanas, en lasque afecta a ninos y adultos jovenes; semanifiesta en el maxilar y la mandıbula. El linfomade Burkitt es susceptible de tratamiento conquimioterapia.
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

MEIOSIS
Lameiosis es un tipo especial de division celular en laque una c�elula diploide (2n) produce cuatro c�elulasgerminales haploides (1n). En la mujer, una de estascuatro c�elulas haploides es un ovulo y las otras tresson cuerpos polares que terminar�an por desaparecer.En el hombre, las cuatro c�elulas haploides son esper-matozoides. Lameiosis –que consta de dos divisionessucesivas, la primera division meiotica y la segundadivision meiotica– da lugar a una reduccion del com-plemento gen�etico de las c�elulas germinales, promuevela recombinacion gen�etica a trav�es de la reorganizacionde los genes, e introduce variabilidad gen�etica.
Primera division meiotica (division reduccional)
El ADN de las c�elulas germinales se duplica a (4n) enla fase S, aunque el numero de cromosomas continuasiendo de (2n), con anterioridad al comienzo de lameiosis (fig. 3.10).
. La profase I se subdivide en las siguientes etapas:. Leptoteno. Comienza la condensacion de loscromosomas.
. Cigoteno. Los cromosomas homologos se unenestrechamente entre sı y originan unas estructurasdenominadas complejos sinapton�emicos.
. Paquiteno. Prosigue la condensacion de loscromosomas homologos; se forman puntos desobrecruzamiento (quiasmas) entre crom�atidasno hermanas que permiten el intercambioaleatorio de material gen�etico.
. Diploteno. El grado de condensacion de loscromosomas homologos continua aumentando;comienzan a separarse los pares de cromosomashomologos.
. Diacinesis.Cuando los cromosomas alcanzan ungrado m�aximo de condensacion, los complejossinapton�emicos se desorganizan, el nucl�eolo y laenvoltura nuclear desaparecen, y los cromosomasse localizan en el compartimento citopl�asmico.Se pone en marcha la formacion de un husointegrado por microtubulos.
. Metafase I. Los cromosomas homologos se disponende manera aleatoria en el plano ecuatorial, lo quepermite la redistribucion de los cromosomasmaternos y paternos. Los microtubulos cinetocoricosse unen a los cinetocoros.
. Anafase I. Los cromosomas homologos, que aunconstan de dos crom�atidas hermanas, migran haciapolos opuestos.
. Telofase I. La telofase I de la meiosis es similar a latelofase mitotica. Los cromosomas alcanzan lospolos opuestos, los nucleos reaparecen y la c�elulaoriginal se divide en dos c�elulas hijas, cada una delas cuales posee una dotacion cromosomica (1n)
(23 cromosomas integrados por 2 crom�atidas,por lo que el complemento de ADN es (2n) en lacitocinesis (v. fig. 3.10). Al final de esta etapacomienza la meiosis II en cada una de las c�elulashijas ası formadas.
Segunda division meiotica (division ecuatorial)
La division ecuatorial no se precede de una fase S. Lasegunda division meiotica equivale a una mitosis y sedivide enprofase II,metafase II, anafase II y telofase II.Los cromosomas, que aun se componen de crom�atidashermanas, se disponen sobre el plano ecuatorial y lascrom�atidas hermanas se separan hacia polos opuestospor accion de los microtubulos cinetocoricos. Al alcan-zar los polos, cada c�elula hija formada en la primeradivision meiotica da lugar a dos nuevas c�elulas hijas enla citocinesis, de modo que se obtienen cuatro c�elulashaploides con un complemento gen�etico exclusivo(fig. 3.11).
Apoptosis
Cuando una c�elula deja de recibir nutrientes o sufreun traumatismo inesperado, se ve sometida a un pro-ceso de necrosis, el cual pone en marcha una res-puesta inflamatoria. La mayorıa de las c�elulas utilizaun proceso gen�etico de autodestruccion conocidocomo apoptosis, la modalidad m�as conocida de lamuerte celular programada. En algunas c�elulas, cier-tas condiciones ambientales, como el hacinamiento,inducen la apoptosis; otras sufren este proceso debidoa su edad; y ciertas c�elulas, como las transformadaspor un virus, lo hacen por accion de determinadasc�elulas pertenecientes al sistema inmunitario. La apop-tosis provoca diversos cambios morfologicos en lac�elula: reduccion del volumen celular, desorganiza-cion de la red citoesquel�etica, desaparicion de laenvoltura nuclear y fragmentacion de la cromatinanuclear. Posteriormente, los restos celulares se englo-ban en vesıculas apoptosicas que ser�an fagocitadaspor los macrofagos. El proceso de apoptosis est�a re-gulado por las caspasas, unas enzimas proteolıticasque reconocen ciertos residuos de aspartato en lasproteınas diana. Cada c�elula presenta la forma inac-tiva de estas enzimas, las procaspasas, algunas de lascuales son activadas para formar caspasas iniciado-ras, que inducen una cascada que dar�a lugar a procas-pasas ejecutoras y proteınas diana en la c�elula paradesencadenar los acontecimientos celulares enumera-dos anteriormente. Algunas senales extracelulares acti-van a receptores apoptosicos localizados en la mem-brana celular, los cuales inducen a las caspasas queintervienen en la vıa extrınseca de la apoptosis. Laliberacion de citocromo c al citoplasma por partede las mitocondrias pone en marcha la vıa intrınseca
38
3
NUCLEO

39
3
NUCLEO
de la apoptosis. Esta mol�ecula se une a la proteınaadaptadora activadora de procaspasas apoptosicas(Apaf1), que se combina con otras Apaf para crear unapoptosoma similar a una rueda, el cual induce unacascada de caspasas que provoca la muerte c�elula pro-gramada. La vıa extrınseca es incapaz de inducir latotalidad de la cascada de las caspasas por sı misma,por lo que ha de activar a la vıa intrınseca para poneren marcha la cascada completa.
CONSIDERACIONES CL�INICAS
Durante la primera division meiotica, a lo largo de lacual se separan los cromosomas homologos paramigrar hacia polos opuestos (anafase I), puedeproducirse la no disyuncion de un par de
homologos, demodo que una c�elula hija albergarıaambos cromosomas homologos y, por tanto,24 cromosomas, mientras que la otra carecerıa deesa copia y poseerıa 22 cromosomas. En lafecundacion normal, un cigoto presentarıa47 cromosomas (trisomıa), mientras que el otroestarıa dotado de 45 (monosomıa). El sındrome deDown es un ejemplo de una trisomıa del cromosoma
21. La no disyuncion se produce m�as a menudo enlos cromosomas 8, 9, 13, 18 y 21, y originatrastornos diferentes en cada caso. Esta anomalıa
es m�as frecuente en mujeres mayores de 35 anos.
Figura 3.10 Etapas de la meiosis en una c�elula con una dotacion diploide (2n) de 4 cromosomas. Primera division meiotica. (Tomadode Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 66.)
Figura 3.11 Segundadivisionmeiotica. (TomadodeGartnerLP,Hiatt JL:ColorTextbookofHistology,3rded.Philadelphia, Saunders,2007, p67.)
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

4 MATRIZ EXTRACELULARLas c�elulas que comparten caracterısticas estructuralesy funcionales se organizan en tejidos que llevan a cabotareas especializadas. En los mamıferos se distinguencuatro tejidos, a saber, el tejido epite-lial, el tejido conjuntivo, el tejidomuscular y el tejido nervioso. Los teji-dos no se componen exclusivamentede c�elulas, sino que tambi�en contie-nen material inerte, la matriz extrace-lular (MEC), formada por sustanciafundamental y fibras (fig. 4.1). Lac�elula produce la MEC, que secretaal espacio extracelular. Anterior-mente se creıa que tan solo prestabasustento fısico, aunque hoy en dıa sesabe que desempena muchos otrospapeles, como:
. Influencia en el desarrollo, la migracion, la mitosis,la morfologıa y la funcion de la c�elula.
. Migracion de las c�elulas en su seno.
El lıquido que se extravasa de los vasos sanguıneos,conocido como lıquido extracelular, transportanutrientes, oxıgeno y mol�eculas de senalizacion a lasc�elulas que integran el organismo y retira productosde desecho, oxıgeno y otros productos celulares altorrente circulatorio. Algunas c�elulas pueden abando-nar los vasos sanguıneos y desplazarse en el seno de laMEC con el fin de eliminar compuestos toxicos, antı-genos, microorganismos, restos de c�elulas muertas yotras sustancias no deseadas de dicha matriz.
Sustancia fundamental
La sustancia fundamental es un gel compuesto porglucosaminoglucanos (GAG), proteoglucanos y glu-coproteınas.
. Los GAG son polisac�aridos de cadena larga noramificada y carga negativa formados por larepeticion de unidades de disac�aridos, el �acidouronico (�acido iduronico o �acido glucuronico)y un aminoazucar (N-acetilglucosamina oN-acetilgalactosamina) (tabla 4.1).. La carga negativa atrae iones Na+, que se unen amol�eculas de agua procedentes del lıquidoextracelular, por lo que los GAG est�an muyhidratados. Estas macromol�eculas suelenasociarse a nucleos proteicos, por lo quepresentan un grado elevado de empaquetamientoque no solo les confiere resistencia a la
compresion, sino que tambi�en las hacedeslizantes.
. Exceptuando al �acido hialuronico, un GAG degran peso molecular formado porm�as de 10.000 disac�aridos, losrestantes GAG est�an sulfatados.
. Los GAG m�as frecuentes(queratano sulfato, condroitina-4-sulfato, condroitina-6-sulfato,heparano sulfato, heparina ydermatano sulfato) constan deunos 300 disac�aridos, se sintetizanen el aparato de Golgi y establecenenlaces covalentes con un nucleoproteico lineal.
. La enzima hialuronano sintetasafabrica el �acido hialuronico en la
superficie citopl�asmica de la membrana celular,que se transporta al espacio extracelular paraincorporarse a la MEC. La mol�ecula de �acidohialuronico puede alcanzar una longitud de20mm.
. Los proteoglucanos de peso molecular muy altoque se componen de un nucleo proteico al que seunen GAG sulfatados mediante enlaces covalentes(v. fig. 4.1). La mol�ecula de proteoglucano remedaun cepillo para limpiar botellas, en el que la porcionproteica corresponderıa al alambre y los GAGrepresentarıan las cerdas. El tamano y lacomposicion de estas macromol�eculas sonvariables:. La decorina presenta un peso molecular de unos50 kDa y posee una unicamol�ecula de GAGunidaa la fraccion proteica, mientras que
. El agrecano, integrado por 200 GAG, tiene unpeso molecular de 3 millones de Da.
. Los proteoglucanos presentan un volumenimportante, ya que todas las mol�eculas de GAGest�an hidratadas.. Muchos proteoglucanos, como el agrecano, seasocian al �acido hialuronico para dar lugar a unasenormes macromol�eculas, alguna de las cualespueden alcanzar un peso molecular de varioscientos de millones de daltons y poseen undominio responsable del estado de gel de lasustancia fundamental.
. Los proteoglucanos desempenan diversasfunciones, como la resistencia a la compresion, launion a mol�eculas de senalizacion, la facilitaciondel movimiento celular, el bloqueo de ladiseminacion de infecciones y la colaboracion enla sıntesis del col�ageno. Los proteoglucanos
� 2011 Elsevier Espana, S.L. Reservados todos los derechos
TERMINOS CLAVE. Sustancia
fundamental. Col�ageno. Sıntesis de col�ageno. Fibras el�asticas. Membrana basal. L�amina basal. Integrinas
40

41
4
MATRIZEXTR
ACELU
LAR
Figura 4.1 Diagrama de la MEC. A. Imagen de bajo aumento en la que se muestra el bandeado de las fibras de col�ageno y losproteoglucanos unidos a ellas.B.GAGunidas al nucleo proteico y las proteınas que los enlazan con lasmol�eculas de �acido hialuronico,
lo que da lugar a grandes macromol�eculas que pueden presentar un peso molecular de varios cientos de millones de daltons.
(Adaptado de Fawcett DW: Bloom and Fawcett’s A Textbook of Histology, 11th ed. Philadelphia, Saunders, 1986.)
Tabla 4.1 TIPOS DE GLUCOSAMINOGLUCANOS (GAG)
GAG Masa (Da) Disac�aridos repetidosLocalizacionen el organismo
Acido
hialuronico
107-108 Acido D-glucuronico-b-1,3-N-acetil-D-glucosamina La mayor parte del tejido
conjuntivo, lıquido
sinovial, cartılago, dermis
Queratanosulfato I y II
10.000-30.000 Galactosa-b-1,4-N-acetil-D-glucosamina-6-SO4 Cornea (queratano sulfato I),cartılago (queratano
sulfato II)
Heparano
sulfato
15.000-20.000 Acido D-glucuronico-b-1,3-N-acetilgalactosamina
Acido L-iduronico 2 o -SO4-b-1,3-N-acetil-D-galactosamina
Vasos sanguıneos, pulmon,
l�amina basal
Heparina (90%)
(10%)
15.000-20.000 Acido L-iduronico-b-1,4-sulfo-D-glucosamina-6-SO4
Acido D-glucuronico-b-1,4-N-acetilglucosamina-6-SO4
Gr�anulos de mastocitos,
hıgado, pulmon, piel
Condroitina-4-
sulfato
10.000-30.000 Acido D-glucuronico-b-1,3-N-acetilgalactosamina-6-
SO4
Cartılago, hueso, cornea, vasos
sanguıneos
Condroitina-6-sulfato
10.000-30.000 Acido D-glucuronico-b-1,3-N-acetilgalactosamina-6-SO4
Cartılago, gelatina deWharton, vasos sanguıneos
Dermatano
sulfato
10.000-30.000 Acido L-iduronico-a-1,3-N-acetilglucosamina-4-SO4 V�alvulas cardıacas, piel, vaso
sanguıneos
Adaptado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 71.�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

asociados a la membrana celular potencian laadhesion de la c�elula a la MEC y pueden actuarcomo mol�eculas receptoras.
. Las glucoproteınas (glucoproteınas de adhesioncelular) son unas proteınas de gran tamano queportan residuos de azucares. Presentan variosdominios de union especıficos para ciertasintegrinas y mol�eculas de la MEC, lo que haceposible la adhesion mutua de las c�elulas y loselementos que integran esta matriz. Lasglucoproteınas mejor conocidas son:. Fibronectina, un gran dımero con forma de V y unpeso cercano a 440.000Da que fabrican algunasc�elulas del tejido conjuntivo, como losfibroblastos. Posee sitios de union especıficos paraun gran numero de integrinas, por lo que potenciala adhesion de las c�elulas a laMEC. La fibronectinaplasm�atica, una forma soluble de fibronectinapresente en el torrente circulatorio, participa en lacoagulacion, la fagocitosis y la cicatrizacion.
. La laminina, una glucoproteına de gran tamanode origen epitelial (950.000Da) se compone detres cadenas polipeptıdicas. Suele localizarse en lacara epitelial de la l�amina basal y presenta sitios deunion para algunos componentes de estaestructura, ası como para integrinas.
. La entactina (nidogeno) se une a la laminina y alcol�ageno de tipo IV; esta glucoproteına potenciala adhesion entre la laminina y la l�amina basal.
. La tenascina es una glucoproteına de elevado pesomolecular (1.700 kDa) que se compone de seispolip�eptidos y remedauna arana con seis patas quesalen de una masa central; posee sitios de unionpara fibronectina y sindecano, un proteoglucanode membrana. Generalmente aparece en el tejidoconjuntivo embrionario, en el que define ciertasrutas para lamigracion de las c�elulas embrionarias.
. La osteopontina se encuentra en el hueso, en elque est�a implicada en la calcificacion, y se une aintegrinas de los osteoclastos.
. La condronectina y la osteonectina sonsemejantes a la fibronectina, aunque aparecenen el cartılago y el hueso, respectivamente.Presentan sitios de union para c�elulas de ambostejidos, ası como para mol�eculas presentes en laMEC de cada uno de estos tejidos.
Fibras
Tradicionalmente, se ha considerado que las fibras dela MEC eran las fibras de col�ageno, las fibras reticu-lares y las fibras el�asticas, aunque en la actualidad sesabe que las fibras reticulares corresponden, en reali-dad, a fibras de col�ageno de tipo II.
El col�ageno, que representa alrededor del 25%de las proteınas del organismo, es una mol�ecula in-
el�astica y confiere resistencia a la tension al tejidoconjuntivo no calcificado. Se distinguen, aproximada-mente, 25 tipos de col�ageno con arreglo a la secuenciaaminoacıdica, que se asignan a tres tipos en funcion alamodalidad de polimerizacion:formador de fibrillas,asociado a fibrillas y formador de redes. Algunos auto-res han descrito proteınas similares a col�ageno.
. Col�agenos formadores de fibrillas (los m�asabundantes de los cuales son los tipos I, II, III, V yXI), organizados en mol�eculas filiformes que seasocian para formar estructuras flexibles similares aun cable cuya resistencia a la tension supera a la delacero inoxidable. Se denominan, tambi�en, fibrasblancas por su coloracion blanca.
. Col�agenos asociados a fibrillas (tipos IX y XII),localizados sobre las fibrillas de col�ageno, quefacilitan su interaccion con otras fibrillas decol�ageno y con elementos de la MEC.
. Col�agenos formadores de redes (tipos IV y VII),que no presentan una estructura filiforme, sino quecrean una malla similar al fieltro que representa elcomponente principal de la l�amina densa de lal�amina basal (tipo IV) y las fibrillas de anclaje(tipo VII), las cuales colaboran en la union de lal�amina basal a la l�amina reticular del tejido conjuntivo.
. Proteınas similares a col�ageno, entre las quefiguran las de tipo XVII (asociadas a loshemidesmosomas) y las de tipo XVIII (situadasen la l�amina basal de los vasos sanguıneos).
ESTRUCTURA DEL COLAGENO FORMADOR DE FIBRILLAS
En muestras no tenidas, las fibras de col�ageno sonincoloras y presentan un tamano muy largo, si biensu di�ametro apenas alcanza los 10mm, en la micros-copia optica. En el microscopio electronico, se obser-va un bandeado transversal distintivo a intervalos de67nm junto a estrıas longitudinales, lo que indica quese componen de fibras m�as delgadas de un di�ametrocomprendido entre 10 y 300nm. Estas finas fibrillasest�an formadas por tropocol�ageno:
. Muchas subunidades de tropocol�ageno se alineande extremo a final y unas junto a otras.
. Una mol�ecula de tropocol�ageno, de 280nm delongitud y 1,5 nm de di�ametro, se compone detres cadenas alfa enroscadas entre sı (fig. 4.2).
. La cadena alfa contiene unos 1.000 amino�acidos;cada tercer amino�acido de esta secuencia es unresiduo de glicina.
. Las cadenas alfa presentan un gran numero deresiduos de hidroxiprolina, que mantienen unidasa dichas cadenas; hidroxilisina, que une lasmol�eculas de tropocol�ageno entre sı, y prolina, lacual suele aparecer a continuacion de glicina en lasecuencia aminoacıdica.
42
4
MATRIZEXTR
ACELU
LAR

43
4
MATRIZEXTR
ACELU
LAR
Figura 4.2 Componentes de las fibras de col�ageno de tipo I. La organizacion de las regiones de intersticio y superpuestas de las mol�eculas
cercanas de tropocol�ageno origina el tıpico bandeado de 67nm en las microfotografıas electronicas. (Tomado de Gartner LP, Hiatt JL:
Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 74.)
CONSIDERACIONES CL�INICAS
La colitis colagenosa afecta principalmente amujeres de mediana edad y ancianas quemanifiestan una diarrea acuosa con una capa
relativamente gruesa de col�ageno acelular hasta elepitelio que reviste el intestino grueso. Desde elpunto de vista histologico, el epitelio de estaspacientes se encuentra infiltrado por linfocitos yneutrofilos. Se desconoce la etiologıa de estaentidad, aunque se ha propuesto que podrıa tener unorigen autoinmunitario. El tratamiento habitual deesta enfermedad consiste en la administracion de
f�armacos antidiarreicos o antiinflamatorios, oambos. Asimismo, la antibioterapia puede dar lugara unamejorıa en caso de sospecha de una infeccion.
Lahepatitis alcoholica se acompana, amenudo, de
la acumulacion de col�ageno a lo largo del fragmentode la vena central correspondiente al lobulo
hep�atico. De no interrumpirse el consumo excesivode alcohol, la enfermedad puede avanzar a un
estadio m�as grave, la esclerosis hialina central, enla cual la v�enula eferente y los sinusoides que larodean se envuelven de tejido conjuntivocolagenoso, lo que reduce la irrigacion y provocahipertension. Los sujetos afectados por estetrastorno presentan fiebre, dolor en el cuadranteabdominal superior derecho e ictericia; la
enfermedad puede provocar insuficiencia hep�aticay muerte en el 20 al 25% de los casos.
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Sıntesis de col�ageno
La sıntesis de col�ageno tiene lugar en el retıculoendopl�asmico rugoso (RER) en forma de cadenas deprecol�ageno, codificadas por mol�eculas individualesde ARNm (fig. 4.3).
. Los extremos amino y carboxilo de estospolip�eptidos reci�en sintetizados poseen secuenciasadicionales de polip�eptidos.
. Adem�as de la escision de los p�eptidos senal, en lascisternas del RER se produce la hidroxilacion dealgunos residuos de prolina y lisina por parte de lasenzimas hidroxilasa de peptidil-prolina ehidroxilasa de peptidil-lisina, respectivamente.
. Otras modificaciones post-traduccion son laglucosilacion selectiva de algunos residuosde lisina.
. Las tres mol�eculas de precol�ageno modificadas sealinean por medio de los prop�eptidos para adoptaruna configuracion helicoidal compacta, aunque losprop�eptidos no se enroscan entre sı.
. El conjunto formado por las tres mol�eculas deprecol�ageno recibe el nombre de procol�ageno, unamol�ecula que se asemeja a una cuerda corta conextremos deshilachados. Las mol�eculas deprocol�ageno no se adhieren unas a otras, lo quepodrıa deberse a los prop�eptidos, sino que salen delRER hacia el aparato de Golgi, donde se asocian aoligosac�aridos.
. El procol�ageno modificado se empaqueta envesıculas recubiertas de proteına coatomero(COP) I, que abandonan el aparato de Golgi transy se exportan al espacio extracelular por mediode la vıa constitutiva.
. Los prop�eptidos se separaran del procol�ageno poraccion de las enzimas de membrana procol�agenopeptidasa amedida que dichasmol�eculas se liberanal espacio extracelular, como consecuencia de locual se forman mol�eculas de tropocol�ageno(v. fig. 4.3).
. La ausencia de estos polip�eptidos permite elautoensamblaje espont�aneo de las mol�eculas detropocol�ageno para originar col�ageno de tipo I.La formacion de esta variante depende de lapresencia de col�ageno de tipo XI, el cualconstituye el eje del col�ageno de tipo I.Por otra parte, los col�agenos de tipos III y V seentremezclan en la matriz de las fibrillas decol�ageno de tipo I. La c�elula encargada deproducir las fibras de col�ageno determina laalineacion de las mol�eculas de tropocol�ageno y lamorfologıa de la fibra de col�ageno a sintetizar.
. Los col�agenos formadores de redes (tipos IV y VII)conservan las propiedades de las mol�eculas deprocol�ageno que los integran; no se organizan enfibras de col�ageno, sino que forman dımeros quecrean una malla similar al fieltro.
. En algunos ganglios linf�aticos, el bazo, la m�edulaosea y el timo, las c�elulas reticulares especializadasfabrican fibras reticulares (col�ageno de tipo III), alas que rodean para aislarlas de su entorno. En casitodas las restantes zonas del organismo, losfibroblastos o las c�elulas del musculo liso (en vasossanguıneos) y las c�elulas de Schwann (en nerviosperif�ericos) se encargan de la sıntesis de estasmol�eculas.
FIBRAS ELASTICAS
A diferencia de las fibras de col�ageno, que no sonel�asticas, las fibras el�asticas pueden estirarse hasta el150%de su longitud en reposo y recuperar su longitudinicial cuando se libera la tension.
. Las fibras el�asticas, tambi�en llamadas fibrasamarillas por su color en muestras frescas, selocalizan en elementos de tejido conjuntivo nocalcificado (sintetizados por los fibroblastos), losvasos sanguıneos (fabricados por c�elulas demusculo liso) y cartılago el�astico (sintetizados porcondrocitos).
. Estas fibras pueden ser filamentos muy delgados obien organizarse en haces m�as gruesos. Rara vez seobservan en cortes desecados tenidos conhematoxilina-eosina (H-E), pero puedenvisualizarse por medio de colorantes especiales. Lasfibras el�asticas constan de un nucleo amorfo deelastina rodeado de microfibrillas (fig. 4.4).. La elastina es una proteına rica en glicina(72 kDa) en la que tambi�en abundan los residuosde alanina, lisina, prolina y valina, con la notableausencia de hidroxilisina.
. Cuatro mol�eculas de lisina pertenecientes acadenas distintas de esta mol�ecula establecenenlaces covalentes muy deformables, llamadosenlaces cruzados de desmosina. Estos enlacescruzados confieren gran elasticidad a las fibras deelastina.
. Las microfibrillas que rodean al nucleo dedesmina est�an formadas por fibrilina, unaglucoproteına de 350 kDa.
. En la sıntesisde las fibrasel�asticas, la c�elulaproduceen primer lugar las microfibrillas y posteriormentese deposita el componente de elastina amorfa en laregion delimitada por aquellas.
44
4
MATRIZEXTR
ACELU
LAR

45
4
MATRIZEXTR
ACELU
LAR
CONSIDERACIONES CL�INICAS
La elastosis solar es un trastorno cut�aneo debido ala exposicion excesiva al sol y a rayos ultravioletas encabinas de bronceado. La piel danada por el sol
aparece m�as arrugada de lo normal, parecedescolgarse y tiene un aspecto y tacto similar alcuero. Esta entidad es consecuencia de los danosocasionados a la dermis, la cual presenta un menor
contenido en col�ageno y un mayor contenido en
fibras el�asticas. Es posible que las fibras el�asticaspierdan un cierto grado de elasticidad a raız de ladesorganizacion de las fibrillas que las integran. Laenfermedad puede evolucionar hacia una neoplasiamaligna.
El escorbuto es una enfermedad que aparece comoconsecuencia de la carencia de vitamina C, una
mol�ecula necesaria para la hidroxilacion de los
residuos de prolina en el preprocol�ageno. Laescasez de hidroxiprolina dificulta la organizacion
normal de las mol�eculas de tropocol�ageno; los
tejidos con un recambio intenso de col�agenoprovocan la descimentacion de los dientes y elsangrado gingival. La entidad se corrige mediante elconsumo de alimentos ricos en vitamina C.
Figura 4.3 Sıntesis y ensamblaje de las mol�eculas de col�ageno detipo I. Los tipos III, V y XI no se incluyen en esta representacion.
(Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology,
3rd ed. Philadelphia, Saunders, 2007, p 77.)
Figura 4.4 Fibra el�astica. El nucleo amorfo de elastina se rodeade microfibrillas. (Tomado de Gartner LP, Hiatt JL: Color
Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 80.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Membrana basal
En el microscopio optico, una delgada capa acelular,denominada membrana basal, separa el tejido con-juntivo del epitelio. En las microfotografıas electroni-cas se distinguen dos componentes en la membranabasal:
. L�amina basal, sintetizada por c�elulas epiteliales y deescaso espesor.
. L�amina reticular, procedente del tejido conjuntivo yde mayor grosor (fig. 4.5).
La l�amina basal se compone de dos capas en lasmicrofotografıas electronicas:
. Una region transparente de 50nm de grosor,conocida como la l�amina lucida, en contacto conlas membranas de las c�elulas basales del epitelio.
. Una malla densa, denominada l�amina densa, queocupa la region comprendida entre la l�amina luciday la l�amina reticular.
Algunos investigadores han senalado, no obstante,que la l�amina lucida podrıa representar un artefactogenerado por el proceso de fijacion de la muestra, demodo que la l�amina basal se compondrıa unicamentede la l�amina densa. Adem�as de separar el epitelio deltejido conjuntivo, las l�aminas basales, denominadasl�aminas externas, pueden rodear a las c�elulas deSchwann, las c�elulas de la musculatura lisa y esquel-�etica, y los adipocitos. El glom�erulo renal posee unal�amina densa engrosada.
LAMINA BASAL Y LAMINA RETICULAR
La l�amina lucida (exista o no como entidad morfolo-gica independiente) de la l�amina basal (v. fig. 4.5)alberga las porciones extracelulares de las proteınasintegrales de membrana integrina y distroglucano,las cuales son receptoras de lamininas. Por otra parte,dos glucoproteınas estructurales, laminina y entac-tina, configuran una fina capa en la superficie dela l�amina densa, la malla de aspecto compacto,de la l�amina basal, cuyo componente principal esel col�ageno de tipo IV (v. fig. 4.5). Las superficies dela l�amina lucida y la l�amina reticular aparecen recu-biertas de un proteoglucano rico enheparano sulfato,elperlecano. Por otra parte, la fibronectina abunda enla cara de la l�amina densa que est�a en contacto con lal�amina reticular.
La laminina se une a integrinas y distroglucanos delas c�elulas epiteliales y al heparano sulfato y el
col�ageno de tipo IV de la l�amina densa, de modoque el epitelio se ancla fijamente a la l�amina basal.La l�amina densa se adhiere firmemente a la l�aminareticular subyacente a trav�es demol�eculas de fibronec-tina, col�ageno de tipo VII (fibras de anclaje) y micro-fibrillas (fibrilina), lo que no solamente refuerza launion del epitelio a la l�amina basal, sino tambi�en ala l�amina reticular.
Entre las funciones de la l�amina basal figuran:
. Refuerzo de la adhesion epitelial
. Filtro molecular debido a la presencia de fibras decol�ageno de tipo IV y la carga negativa de lasmol�eculas de heparano sulfato
. Potenciacion de la actividad mitotica de las c�elulas
. Union a mol�eculas de senalizacion
. Colaboracion en la reorganizacion de proteınasintegrales de la membrana celular
. Asistencia en la reepitelizacion de heridas y laregeneracion de las uniones neuromusculares
Los fibroblastos se encargan de sintetizar la l�aminareticular (v. fig. 4.5), integrada por fibras de col�agenode tipos I y III. El grosor de esta capa es variable ydepende de las fuerzas abrasivas que actuen sobre elepitelio que lo recubre; se extiende hasta el epitelio enlas palmas de las manos y las plantas de los pies ymuestra un espesor reducido bajo el epitelio delpar�enquima pulmonar. Las fibras de col�ageno de lal�amina reticular proceden de las mol�eculas decol�ageno del tejido conjuntivo y se continuan conellas para asegurar este tejido a la l�amina reticularadem�as de la l�amina basal a la l�amina reticular, detal manera que se mantenga una asociacion estrechaentre el epitelio y el tejido conjuntivo.
Integrinas y distroglucanos
Las integrinas son proteınas demembrana cuyo grupoextracelular se une a ciertos ligandos presentes en laMEC en presencia de cationes divalentes y cuyos extre-mos carboxilo intracelulares se asocian a las talinas ya-actininas citoesquel�eticas. Las integrinas puedentransducir senales extracelulares para desencadenaracontecimientos intracelulares vinculados con la divi-sion celular, la expresion g�enica o ambas.
Los distroglucanos son proteınas transmembranaformadas por heterodımeros cuyo dominio extracelu-lar se une a un sitio determinado de la laminina,mientras que el dominio intracelular lo hace a la dis-trofina, una proteına de union a actina que se asociaal citoesqueleto.
46
4
MATRIZEXTR
ACELU
LAR

CONSIDERACIONES CL�INICAS
El sındrome de Goodpasture es un trastornoautoinmunitario que afecta a los rinones y lospulmones. La enfermedad limitada a los primerosrecibe el nombre de glomerulonefritis poranticuerpos frente a la membrana basal de los
glom�erulos. En ambos casos, el organismodesarrolla una respuesta inmunitaria frente al
col�ageno de tipo IV de la l�amina basal. El sındromede Goodpasture suele debutar con posterioridad auna infeccion respiratoria y la afectacion pulmonarsuele relacionarse con el tabaquismo. Los afectadossuelen ser hombres jovenes, aunque la enfermedadse manifiesta en sujetos de ambos sexos y cualquiergrupo etario. Este trastorno puede evolucionar congran rapidez hacia insuficiencia renal que requiere untrasplante renal. En las etapas iniciales de laenfermedad, el tratamiento englobacorticoesteroides, compuestos citotoxicos y
plasmaf�eresis. A menudo, este sındrome tieneconsecuencias mortales y la tasa de supervivencia
tan solo alcanza el 50% a los 2 anos del comienzo dela enfermedad incluso cuando se instaura untratamiento agresivo.
El sındrome de Alport (nefritis hereditaria) es una
enfermedad gen�etica producida por unamutacion en
el gen COL4A5 que codifica el col�ageno de tipo IV;
los sujetos afectados no pueden fabricar una l�amina
basal normal. La l�amina basal glomerular de estospacientes presenta un grosor excesivo y parecedividirse en dos capas interconectadas como siestuviera formada por ampollas. La prevalencia y lagravedad de esta entidad sonmayores en el hombre,aunque afecta a ambos sexos. La enfermedadevoluciona hacia insuficiencia renal terminal hacia la
quinta d�ecada de vida en casi todos los hombres yalrededor del 20% de las mujeres. Por otra parte, almenos el 50% de los pacientes de ambos sexospresenta hipoacusia progresiva y lesiones en elcristalino.
CONSIDERACIONES CL�INICAS
El tratamiento de algunos casos de artrosis
consiste en inyecciones repetidas de �acidohialuronico, un componente del lıquido sinovial,directamente en la articulacion con el fin delubricarla y aliviar la sintomatologıa durante unperıodo prolongado.
47
4
MATRIZEXTR
ACELU
LAR
Figura 4.5 La membrana basal consta de dos elementos, la l�amina basal y la l�amina reticular. (Adaptado de Fawcett DW: Bloom andFawcett’s a Textbook of Histology, 12th ed. New York, Chapman & Hall, 1994.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

5 EPITELIO Y GLANDULASEl cuerpo humano se compone de m�as de 200 tiposdiferentes de c�elulas, que se organizan en cuatro tiposb�asicos de tejido: epitelio, tejido conjuntivo, musculoy tejido nervioso. Los tejidos se combi-nanpara formarunasunidades funciona-les denominadas organos, que se agrupanen sistemas org�anicos.
Tejido epitelial
El tejido epitelial aparece en forma del�aminas de c�elulas contiguas, querecubren o revisten la superficie cor-poral, o bien en forma de gl�andulas,estructuras secretoras derivadas dec�elulas epiteliales. La mayor parte delos epitelios proviene del ectodermo yel endodermo, aunque algunos proce-den del mesodermo.
. El ectodermo origina la epidermis de la piel, elrevestimiento de las cavidades bucal y nasal, lacornea, las gl�andulas sudorıparas y seb�aceas y lasgl�andulas mamarias.
. El endodermo da lugar al revestimiento de losaparatos digestivo y respiratorio, ası como a lasgl�andulas del primero.
. Delmesodermo proceden los tubulos urinıferos delrinon, el revestimiento de los aparatos reproductor ycirculatorio, y el revestimiento de las cavidadescorporales.
El epitelio, un tejido avascular que se dispone enl�aminas, recibe nutrientes de la irrigacion vascular deltejido conjuntivo adyacente. Est�a formado por c�elulasde empaquetamiento muy compacto, unidas por
complejos de union, entre las que apenas queda espa-cio extracelular y cimentadas por un reducido volu-men de matriz extracelular. La l�amina basal, de ori-
gen epitelial, separa ambos tejidos. Elepitelio interviene en las siguientesfunciones:
. Proteccion de los tejidos querecubre o reviste,
. Transporte transcelular demol�eculas a trav�es de las distintascapas que lo integran,
. Secrecion de diversas mol�eculaspor parte de las gl�andulas,
. Absorcion (p. ej., tubo digestivo ytubulos renales),
. Control del movimiento de iones ymol�eculas mediantepermeabilidad selectiva, y
. Deteccion de sensaciones (p. ej.,gusto, vista, oıdo).
CLASIFICACION DE LAS MEMBRANAS EPITELIALES
La clasificacion del tejido epitelial se basa en elnumero de capas celulares dispuestas entre la l�aminabasal y la superficie libre y en la morfologıa celular. Elepitelio simple se compone de una monocapa dec�elulas epiteliales, mientras que el epitelio estratifi-cado consta de dos o m�as capas. Las c�elulas epitelialesadosadas a la superficie libre pueden presentar unamorfologıa escamosa (aplanada), cubica o cilındrica,lo que conforma distintos tipos de epitelio (fig. 5.1 ytabla 5.1). Existen otros dos tipos de tejido epitelial: elepitelio cilındrico seudoestratificado y el epitelio detransicion.
� 2011 Elsevier Espana, S.L. Reservados todos los derechos
TERMINOS CLAVE
. Epitelio
. Epitelio simple
. Epitelio estratificado
. Microvellosidades
. Complejo de union
. Gl�andulas exocrinasunicelulares
. Gl�andulas exocrinasmulticelulares
. Gl�andulas endocrinas
48

49
5
EPITELIOYGLANDULAS
Figura 5.1 Tipos de epitelio. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 87.)
Tabla 5.1 CLASIFICACION DE LOS EPITELIOS
TipoMorfologıa delas c�elulas de superficie
Ejemplos Funciones
Simple
Escamoso simple Aplanada Revestimiento: alv�eolos, asa de Henle,
capa parietal de la c�apsula de Bowman,
oıdo interno y medio, vasos
sanguıneos y linf�aticos, cavidades
pleural y peritoneal
Membrana limitante, transporte de
lıquidos, intercambio gaseoso,
lubricacion, reduccion de friccion
(ayuda al movimiento de
vısceras), membrana de
revestimiento
Cubico simple Cubica Conductos demuchas gl�andulas, cubre el
ovario, forma los tumulos renales
Secrecion, absorcion, proteccion
Cilındrico simple Cilındrica Revestimiento: oviductos, conductillos
eferentes de testıculos, utero,
bronquıolos, gran parte del tubo
digestivo, vesıcula biliar y conductos
grandes de algunas gl�andulas
Transporte, absorcion, secrecion,
proteccion
Seudoestratificado Todas las c�elulas descansan en
la l�amina basal, pero sin
alcanzar la superficie
epitelial; las c�elulas de
superficie son cilındricas
Revestimiento: lamayor parte de la tr�aquea,
bronquıolos primarios, epidıdimo y
conducto deferente, trompa auditiva,
parte de la cavidad timp�anica, cavidad
nasal, saco lacrimal, uretra masculina,
conductos excretores grandes
Secrecion, absorcion, lubricacion,
proteccion, transporte
Estratificado
Escamoso
estratificado (no
queratinizado)
Aplanada (con nucleos) Revestimiento: cavidad bucal, epiglotis,
esofago, pliegues vocales, vagina
Proteccion, secrecion
Escamoso
estratificado
(queratinizado)
Aplanada (sin nucleos) Epidermis de la piel Proteccion
Cubico estratificado Cubica Revestimiento: conductos de gl�andulas
sudorıparas
Absorcion, secrecion
Cilındrico
estratificado
Cilındrico Conjuntiva ocular, algunos conductos
excretores grandes, porciones de la
uretra masculina
Secrecion, absorcion, proteccion
De transicion Cupular (relajado), aplanada
(distendido)
Revestimiento: vıas urinarias desde los
c�alices renales hasta la uretra
Proteccion, distensibilidad
Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 86.�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

POLARIDAD Y ESPECIALIZACIONES DE LA SUPERFICIE
CELULAR
Por lo general, las c�elulas epiteliales presentan regio-nes especıficas –dominios– que les confieren unapolaridad definida. Estos dominios aparecen en laregion apical o basolateral de la c�elula. Las unionesherm�eticas, estructuras especializadas de membrana,rodean a la region apical de la c�elula, separan ambosdominios y le confieren polaridad. En cada dominioexisten modificaciones especiales.
Dominio apical
En el dominio apical, la region de la c�elula epitelial encontacto con la superficie libre, abundan los canalesionicos, las proteınas transportadoras, la bomba deH+-ATPasa, las acuaporinas, las glucoproteınas y lasenzimas hidrolıticas. Adem�as, a trav�es de esta regionse secretan diversos compuestos hacia el espacio extra-celular. Algunas modificaciones del dominio apical,como las microvellosidades y el glucoc�aliz asociado,los cilios, los estereocilios y los flagelos, participan enun gran numero de funciones celulares.
. Las microvellosidades (fig. 5.2) son proyeccionesdigitiformes limitadas por membrana de 1 a 2mmde longitud que se localizan en las regiones apicalesde las c�elulas que integran los epitelios cubicosimple y cilındrico simple. En el microscopiooptico, corresponden a los extremos estriados y encepillo de estos tejidos; cuando presentanempaquetamiento compacto, pueden incrementarel �area superficial de la c�elula hasta 20 veces.. El eje de cada microvellosidad consta de 20 a30 filamentos de actina unidos entre sı por villinay fimbrina; los filamentos localizados en laperiferia del haz se adhieren a la membranaplasm�atica por medio de las proteınascalmodulina ymiosina I.
. Los extremos positivos de los filamentos de actinase extienden hasta la punta de la microvellosidad,que aparece incluida en una sustancia amorfa.
. Los extremos citopl�asmicos del haz de actina seunen a la red terminal y est�an formados porfilamentos intermedios, espectrina, actina y otroselementos citoesquel�eticos.
. La cara extracelular de la membrana de lasmicrovellosidades se recubre de un glucoc�alizcuya composicion depende de la localizacion y lafuncion de la c�elula.
. Lasmicrovellosidades rıgidas, inmoviles y de granlongitud, exclusivas del epidıdimo y las c�elulas
pilosas sensoriales de la coclea (oıdo interno)reciben en nombre de estereocilios. Estasestructuras incrementan el �area de superficie conel fin de facilitar la absorcion en el epidıdimo,mientras que en el oıdo colaboran con las c�elulaspilosas en la produccion de senales.
. Los cilios (fig. 5.3) son estructuras digitiformeslargas (con una longitud comprendida entre 7 y10mm y un di�ametro de 0,2mm) que se proyectandesde el dominio apical de la c�elula. Se trata de unasestructuras muy conservadas que est�an presentes enlos organismos unicelulares, las plantas y todos losanimales. Los cilios son estructuras contr�actiles queposibilitan el desplazamiento de los organismosunicelulares en el agua; en los organismossuperiores, en los que algunas capas epiteliales,como el revestimiento de las vıas respiratorias,pueden poseer 2.000millones de cilios/cm2, actuande manera coordinada para impulsar lıquido a lolargo de capas de c�elulas epiteliales.. El eje central de los cilios, llamado axonema, esuna estructura longitudinal muy organizadaintegrada por nueve dobletes que rodean a dosmicrotubulos solitarios, dineına y proteınasel�asticas asociadas.
. Cada doblete se compone de un microtubulocompleto (subunidad A), formado por13 protofilamentos, y un microtubuloincompleto (subunidad B), el cual posee10 protofilamentos completos y comparte tresprotofilamentos con elmicrofilamento completo.
. La subunidad A de cada doblete presenta brazosde dineına que se disponen a lo largo de lamismaa intervalos de 24nm, de manera similar a laspatas de un miri�apodo (ciempi�es). Los extremoslibres de estos brazos portan sitios de uniondependientes de trifosfato de adenosina (ATP)para la subunidad B.
. La disposicion de las proteınas el�asticas asociadasal axonema es la siguiente: los dos microtubulossolitarios se rodean de una vaina central, hacia laque se proyecta unaproyeccion radialprocedentede cada subunidad A.
. Asimismo, la subunidad A de un doblete seconecta con la subunidad B de los dobletesadyacentes mediante un puente de nexina.
. La estructura tridimensional de la vaina centralcorresponde a un cilindro que rodea a losmicrotubulos solitarios; cada puente de nexina ycada brazo radial constituyen un cuadril�atero dematerial el�astico.
50
5
EPITELIOYGLANDULAS
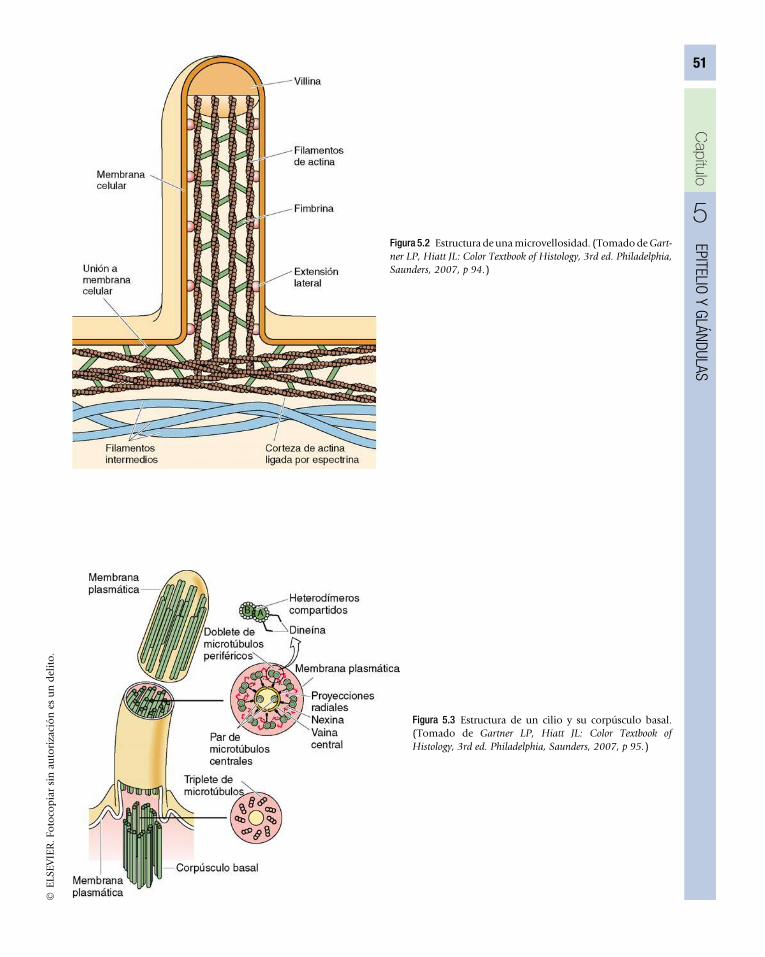
51
5
EPITELIOYGLANDULAS
Figura 5.2 Estructura de unamicrovellosidad. (TomadodeGart-
ner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia,Saunders, 2007, p 94.)
Figura 5.3 Estructura de un cilio y su corpusculo basal.(Tomado de Gartner LP, Hiatt JL: Color Textbook of
Histology, 3rd ed. Philadelphia, Saunders, 2007, p 95.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Movimiento ciliar, cuerpo basal y flagelos
Los brazos de dineına se unen y separan de la sub-unidad B del doblete adyacente ascendiendo haciael extremo del cilio durante su movimiento. Lanexina y las proyecciones radiales tienden a limitarla accion de ascenso, por lo que el cilio ha dedoblarse.
. Las proteınas el�asticas se distienden comoconsecuencia del doblamiento del cilio(dependiente de ATP); sin embargo, cuando seinterrumpe el movimiento ascendente de los brazosde dineına, las proteınas el�asticas recuperan sulongitud normal y el cilio adopta de nuevo suposicion erguida (en un proceso que no requiereATP).
. La r�apida alternancia de ambos procesos permite eldesplazamiento de compuestos sobre la superficiedel epitelio.
La estructura descrita se interrumpe en la base delcilio, en la que el axonema se une al cuerpo basal(fig. 5.4), una estructura formada por nueve triple-tes de microtubulos (subunidades A, B y C) quecarece de microtubulos solitarios en su porcion central.El cuerpo basal es similar a un centrıolo y se ensamblaa partir de los organizadores de los procentrıolos.
. Las subunidades A y B del cilio se continuan con lassubunidades A y B del cuerpo basal.
. La subunidad C del cuerpo basal no forma parte dela estructura del cilio.
Algunas c�elulas, como los fibroblastos, las neuronasy ciertas c�elulas epiteliales, como las de los tubulosrenales, pueden presentar un cilio inmovil solitariocuyo axonema est�a exento de brazos de dineına. Estetipode cilio recibe elnombrede cilioprimario ypodrıaactuar como organo sensorial o receptor de senales.
Los flagelos, restringidos a los espermatozoides en elser humano, son cilios modificados dotados de un axo-nema y un complejo robusto de proteınas el�asticas queimpulsan a estas c�elulas en el seno del aparato reproduc-tor femenino. Los flagelos se abordan en el capıtulo 21.
Dominio basolateral
El dominio basolateral de las c�elulas epitelialesengloba sus membranas lateral y basal. Presenta
un gran numero de complejos de union, receptoresde senales, canales ionicos y ATPasas de Na+/K+;esta region participa, adem�as, en la secrecionconstitutiva.
Especializaciones de la membrana lateral
Las barras terminales, que pueden visualizarse en lamicroscopia optica, son �areas de posible union de lasc�elulas epiteliales; se sabe que forman una estructuracontinua alrededor de la c�elula. Las barras terminalesocupan regiones restringidas cercanas al �apice de lac�elula. Las im�agenes de microscopia electronicaponen de relieve que las barras terminales correspon-den a complejos de union que potencian la adhe-sion de c�elulas contiguas (fig. 5.5). La barra terminalse compone de tres tipos de uniones celulares: lazonula oclusiva, que ocupa la posicion m�as apical,inmediatamente por debajo de la cual aparece lazonula adherente, dos zonas de contacto continuasen forma de cinturon que rodean el perımetro de lac�elula, y las m�aculas adherentes (desmosomas),que corresponden a uniones puntuales, no conti-nuas, que jalonan la superficie celular. La c�elulaposee otras uniones celulares que no se localizanen las barras terminales, por lo que no pertenecenal complejo de union. Entre ellas figuran las unionesgap, los desmosomas, los hemidesmosomas y lasadhesiones de matriz celular ligadas por actina. Lasuniones existentes entre c�elulas epiteliales puedendividirse en tres grandes grupos con arreglo a la fun-cion que desempenan:
. Uniones oclusivas (zonulas oclusivas), que actuancomo barrera impermeable o con permeabilidadselectiva que impide el paso de compuestos a trav�esde la membrana de dos c�elulas epitelialesadyacentes (vıa paracelular).
. Uniones de anclaje (zonulas adherentes, m�aculasadherentes, hemidesmosomas, adhesiones dematriz celular ligadas por actina), a trav�es de lascuales las c�elulas epiteliales se unen entre sı, a lamembrana basal o ambas.
. Uniones comunicantes (uniones gap), quepermiten el movimiento transcitopl�asmico de ionesy mol�eculas pequenas, acoplando su carga y sumetabolismo.
52
5
EPITELIOYGLANDULAS

53
5
EPITELIOYGLANDULAS
Figura 5.5 Complejos de union. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 97.)
Figura 5.4 Estructura de un cilio y su corpusculo basal. (Tomado de
Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia,
Saunders, 2007, p 95.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Las zonulas oclusivas (uniones estrechas), elcomponente m�as apical del complejo de union, secrean como consecuencia de la fusion de las hojuelasexternas de las membranas de c�elulas adyacentes(fig. 5.6). La zona de fusion circunda por completoa la c�elula epitelial; en las im�agenes de microscopiaelectronica, se observa la presencia de hebras defusion, unas estructuras lineales formadas por pro-teınas transmembrana, en la cara P y surcos linealesen la cara E.
. Segun el grado de integridad de la union estrecha,pueden existir varias modalidades de fusion, comoun sistema divergente de hebras de fusion, de modoque la zonula oclusiva serıa m�as permeable alposeer un numerom�as bajo de dichas hebras, o bienuna estructura menos permeable, integrada porabundantes hebras de fusion.
. Estas proteınas transmembrana se localizan en lasmembranas de ambas c�elulas y establecen contactode manera independiente del calcio en el espacioextracelular hasta ocluirlo. Se han identificado tresclases de proteınas transmembrana en las unionesestrechas:. Las claudinas constituyen el grupo m�asimportante; se trata de proteınas que bloquean elespacio extracelular en las zonas de contacto dedos c�elulas.
. Las tricelulinas sustituyen a las claudinas enaquellas regiones en las que establecen contactotres c�elulas.
. Las ocludinas conforman la tercera clase deproteınas; se desconoce cu�al es su funcion.
. Los otros dos elementos que integran el complejode union, las zonulas adherentes y las m�aculasadherentes, refuerzan las uniones estrechas.. Tres proteınas citopl�asmicas de andamiaje–proteınas de union estrecha (zonulas oclusivas)ZO1, ZO2 y ZO3– verifican que la alineacion delas claudinas, las ocludinas y las tricelulinas de lasc�elulas adyacentes sea la correcta, si bien no se hadefinido aun cu�al serıa su mecanismo de accion.
. Otro complejo proteico, el complejo afadina-nectina, se une a la proteına ZO1; se cree quepodrıa interaccionar con la mol�ecula homologade la c�elula adyacente para potenciar la adhesionde unas claudinas con otras.
Las uniones estrechas limitan o evitan el movi-miento paracelular de material a trav�es del epitelio,adem�as de impedir la migracion de proteınas integra-les entre los dominios apical y basolateral de la mem-brana celular.
Al igual que las zonulas oclusivas, las zonulasadherentes son zonas de contacto con forma de cintu-ron que rodean a la c�elula (v. fig. 5.6). En estas uniones
de adhesion intervienen proteınas ligadoras trans-membrana dependientes de calcio, las cadherinas,que mantienen unidas a c�elulas adyacentes. El domi-nio sensible al calcio de las cadherinas se localiza en lacara extracelular de lamembrana y se compone de unaestructura flexible similar a una bisagra.
. En presencia de Ca++, la region similar a una bisagrase mantiene fija; al extenderse, establece contactocon el dominio extendido de una cadherina de lac�elula adyacente y se une al mismo, aunque ladistancia entre ambasmembranas no puede superarlos 15 a 20nm. Los dominios intracelulares de lascadherinas se asocian ahaces de filamentos de actinaque se disponen en paralelo a la membrana celular.
. Los enlaces con los filamentos de actina seestablecen a trav�es de cateninas, vinculina, ya-actinina. De este modo, las cadherinastransmembrana acoplan el citoesqueleto de unac�elula con el de la c�elula adyacente. Al igual quesucede en la zonula oclusiva, la union de adhesionse refuerza por medio de un complejo afadina-nectina.
. Por otra parte, las uniones adherentes puedenadoptar una estructura similar a un lazo, como en elendotelio de los capilares, que no recorre latotalidad del perımetro de la c�elula; este tipo deuniones recibe el nombre de fascias adherentes.
Otras uniones celulares similares a redes, denomi-nadas desmosomas (400�250� 10nm, aproxima-damente), parecen presentar una distribucion aleato-ria en la membrana basolateral de las c�elulas delepitelio simple y las membranas de c�elulas adyacentesdel epitelio escamoso estratificado, como el de la epi-dermis (v. fig. 5.6). Cada mitad del desmosoma seacopla a las superficies intracelulares de las membra-nas de las c�elulas epiteliales adyacentes.
. Las desmoplaquinas y las pacoglobinas actuancomo proteınas de union en cada placa.
. Se cree que los filamentos de citoqueratina(filamentos intermedios) reducen las fuerzas decizallamiento en la c�elula, ya que atraviesan la placay giran sobre sı mismos para ingresar, de nuevo, enel citoplasma.
. El espacio intercelular comprendido entre dosplacas de desmosomas (con un anchuraaproximada de 30nm) contiene proteınasligadoras transmembrana filamentosasdependientes de Ca++ pertenecientes a la familia delas cadherinas, desmogleına y desmocolina.
. En presencia de Ca++, se establece un enlace entre lasproteınas ligadoras de cada c�elula. Este enlace serompe en ausencia de este cation, de modo que lasdos mitades del desmosoma no puedenmantenerseunidas firmemente y las c�elulas se separan.
54
5
EPITELIOYGLANDULAS

CONSIDERACIONES CL�INICAS
El p�enfigo vulgar es una enfermedad autoinmuneque afecta a la piel caracterizada por laproduccion de anticuerpos frente a proteınasdesmosomicas. La union de los anticuerpos aestas proteınas incide en la adhesion celular. Estaalteracion provoca la formacion de ampollas en la
epidermis con p�erdida de lıquidos corporales.En ausencia de tratamiento, este trastorno tieneconsecuencias mortales. Se emplean
corticoesteroides sist�emicos y f�armacosinmunosupresores como tratamiento de estaentidad.
55
5
EPITELIOYGLANDULAS
Figura 5.6 Complejos de union. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 97.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

UNIONES GAP
Los complejos de union m�as abundantes, las unionesgap, aparecen en la mayorıa de los tejidos epiteliales,las neuronas y las c�elulas de musculo liso y cardıaco.Las uniones gap son zonas de comunicacion interce-lular, ya que permiten el paso demol�eculas pequena atrav�es de un espacio intercelular estrecho (2 a 4nm).Estas uniones equilibran a las c�elulas a nivel de carga ycomposicion quımica, y facilitan la comunicacionintercelular tanto en los tejidos adultos como en losembrionarios.
. En la membrana celular, la asociacion de seisconexinas, proteınas transmembrana formadorasde canales, da lugar a unos canales acuososdenominados conexones (fig. 5.7).
. En una union gap pueden aparecer de unos pocos avarios miles de conexones.
. Los conexones de un lado de la union gap que est�anconectados con los del lado opuesto se unen entre sıpara formar un canal hidrofilo de comunicacion,con un di�ametro de 1,5 a 2 nm, a trav�es del cualpueden pasarmol�eculas de tamanomenor de 1 kDade una c�elula a otra contigua.
. No se conoce adecuadamente el mecanismo detransporte de mol�eculas a trav�es de las uniones gap,si bien se sabe que el aumento de lasconcentraciones citopl�asmicas de Ca++ o ladisminucion del pH citopl�asmico clausura estasuniones, mientras que el aumento de dicho pH o ladisminucion de dichas concentraciones da lugar asu apertura.
Especializaciones de la superficie basal
La l�amina basal, los repliegues de lamembrana celulary los hemidesmosomas representan las tres especiali-zaciones principales de las superficies basales de lasc�elulas epiteliales (v. fig. 5.7). Los hemidesmosomasse localizan en la superficie basal de la c�elula y parti-cipan en el anclaje de la membrana basal a la l�aminabasal subyacente.
. La l�amina basal, un producto del epitelio quesepara el epitelio del tejido conjuntivo situado
por debajo del mismo, se describio en elcapıtulo 4.
. Los repliegues de lamembrana basal, en particularlos implicados en el transporte ionico, incrementanel �area de superficie de la membrana plasm�atica ycrean compartimentos en la porcion basal delcitoplasma en los que se concentran lasmitocondrias. La presencia de estos org�anulos juntoa los pliegues de la membrana citopl�asmica confiereun aspecto estriado a la c�elula en la microscopiaoptica.
. Los hemidesmosomas parecen ser la mitad de undesmosoma y se localizan en la membranaplasm�atica basal. Intervienen en la union de lamembrana plasm�atica basal a la l�amina basal, loque facilita el anclaje de la c�elula al tejidoconjuntivo subyacente.. Los hemidesmosomas se localizan en la caracitopl�asmica de lamembrana plasm�atica y poseenplacas de union formadas por desmoplaquinas,plectina y otras proteınas secundarias, en las quese embeben los extremos terminales de losfilamentos intermedios de queratina(tonofilamentos).
. Las proteınas ligadoras transmembrana,pertenecientes a la familia de las integrinas, sonun grupo de receptores de la matriz extracelularque ingresan en la placa en su cara citopl�asmicay atraviesan la membrana celular; su dominioextracelular se une a mol�eculas de laminina ycol�ageno de tipo IV presentes en la l�aminabasal.
Recambio de las c�elulas epiteliales
La tasa de recambio de las c�elulas epiteliales es ele-vada; el recambio es m�as intenso en ciertos organos,como en el revestimiento del tubo digestivo, ym�as lento en otras zonas del organismo, como laepidermis de la piel. No obstante, la tasa de recam-bio de cada organo suele ser constante. La destruc-cion de un numero alto de c�elulas debido a unainfeccion o una lesion induce un aumento de lamitosis encaminado al restablecimiento de la pobla-cion celular normal.
56
5
EPITELIOYGLANDULAS

57
5
EPITELIOYGLANDULAS
Figura 5.7 Complejos de union. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 97.)
CONSIDERACIONES CL�INICAS
La hipoacusia no sindromica y la eritroqueratodermia
variable, una afeccion cut�anea, se deben a mutacionesen genes que codifican las conexinas. Las mutaciones
en estos genes se vinculan, adem�as, con anomalıas en la
migracion de las c�elulas de la cresta neural que dan lugar a
anomalıas cong�enitas de los vasos pulmonares del
corazon.
A pesar de que cada poblacion de c�elulas epitelialespresenta caracterısticas distintivas relacionadas con
diversos factores, como su localizacion y su entorno, todas
ellas guardan relacion con su funcion. En ciertos trastornos,
las c�elulas epiteliales pueden transformarse en otro tipo de
c�elulas epiteliales mediante un proceso conocido como
metaplasia. El epitelio respiratorio (epitelio cilındricoseudoestratificado ciliado) de un fumador habitual puede
sufrir un procesodemetaplasia escamosa, el cual da lugara la transformacion de este tejido en un epitelio escamoso
estratificado capaz de realizar un numero menor de
funciones. Esta transformacion puede invertirse comoconsecuencia de la desaparicion del factor ambiental
desencadenante.
Los tumores de c�elulas epiteliales pueden ser benignos o
malignos. Los tumores malignos de origen epitelial reciben
el nombre de carcinomas, mientras que los procedentesde tejido glandular se denominan adenocarcinomas. Enlos ninos menores de 10 anos, es poco probable que los
tumores provengan del epitelio, mientras que los
adenocarcinomas prevalecen en los adultos. Alrededor del90% de los casos de neoplasias malignas en sujetos
mayores de 45 anos son de origen epitelial.
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Gl�andulas
A lo largo del desarrollo de algunas regiones del orga-nismo, las c�elulas epiteliales invaden el tejido conjun-tivo subyacente para dar lugar al par�enquima (unida-des secretoras y conductos) de las gl�andulas y serodean de una l�amina basal secretada por ellas mis-mas. El tejido conjuntivo circundante, conocidocomo estroma, sustenta al par�enquima glandular alaportar su irrigacion vascular, inervacion y ciertos ele-mentos estructurales, como c�apsulas, que engloban ala gl�andula en su totalidad, y tabiques, los cuales sub-dividen a la c�elula en lobulos y lobulillos. Cada una delas c�elulas que integra la unidad secretora de lagl�andula fabrica productos de secrecion, que alma-cena en unos compartimentos intracelulares llamadosgr�anulos secretores para su posterior liberacion. Elcontenido de estos gr�anulos es variado y depende de lagl�andula encargada de su sıntesis:
. Una hormona, como la insulina, producida por losislotes de Langerhans;
. Una enzima, como la amilasa salival de la gl�andulaparotida, o la secrecion rica en bicarbonato de lasgl�andulas de Brunner del duodeno; o bien
. Una l�agrima, una secrecion acuosa fabricada por lagl�andula lacrimal.
Sehandefinidodos tipos principales de gl�andulas enfuncion del mecanismo de secrecion de sus productos:
. Gl�andulas exocrinas, las cuales poseen conductos atrav�es de los que se liberan las mol�eculas quesintetizan.
. Gl�andulas endocrinas, que carecen de conductos,por lo que secretan sus productos directamente altorrente circulatorio o los vasos linf�aticos.
A menudo, las c�elulas se comunican entre sı pormedio de citocinas, unas mol�eculas de senalizacionque actuan sobre unas c�elulas especıficas denomina-das c�elulas diana. Las c�elulas que liberan citocinasreciben el nombre de c�elulas de senalizacion; lasmol�eculas por ellas secretadas se unen a receptoresque inducen funciones especıficas en las c�elulas diana(v. capıtulo 2). Los efectos de las citocinas puedenasignarse a una de las tres categorıas siguientes, basa-das en la distancia que separa a la c�elula de senaliza-cion de la c�elula diana:
. Autocrino. La c�elula de senalizacion y la c�eluladiana son la misma c�elula –la c�elula se estimula a sımisma.
. Paracrino. La c�elula diana y la c�elula desenalizacion se encuentran proximas entre sı, demodo que la citocina alcanza a aquella mediantedifusion.
. Endocrino. La distancia entre la c�elula desenalizacion y la c�elula diana es lo bastanteamplia para que la citocina se transporte hasta sudestino a trav�es del torrente circulatorio o losvasos linf�aticos.
GLANDULAS EXOCRINAS
Las gl�andulas exocrinas pueden clasificarse con arre-glo al numero de c�elulas que las integran:
. Unicelulares: la gl�andula se compone de una unicac�elula (p. ej., c�elulas caliciformes).
. Multicelulares: la gl�andula consta de m�as de unac�elula (p. ej., gl�andula submandibular).
Se puede utilizar una clasificacion basada en el tipode secrecion sintetizada por la gl�andula:
. Serosa: acuosa (p. ej., gl�andula parotida)
. Mucosa: viscosa (p. ej., gl�andulas salivales menoresdel paladar)
. Mixta: serosa y mucosa (p. ej., gl�andula sublingual)
Existen otros sistemas de clasificacion que sesustentan en el mecanismo de liberacion de los pro-ductos de secrecion por parte de las gl�andulas(fig. 5.8):
. Merocrino: tan solo se libera el producto desecrecion (como en la gl�andula parotida).
. Apocrino: el producto se acompana de unapequena porcion del citoplasma de la c�elula (comopodrıa ser el caso de la gl�andula mamaria durante elperıodo de lactancia).
. Holocrino: la c�elula muere y se convierte en elproducto secretado (como sucede en las gl�andulasseb�aceas).
Gl�andulas exocrinas unicelulares
La c�elula caliciforme, localizada en el revestimientoepitelial de los intestinos delgado y grueso, y la por-cion conductora de las vıas respiratorias, constituye elejemplo m�as destacado de una gl�andula exocrina uni-celular (fig. 5.9). La base estrecha de la c�elula calici-forme, llamada tallo, est�a en contacto con la l�aminabasal. La teca, la porcion apical de la c�elula que seexpande debido a la presencia de un gran numero degr�anulos de secrecion con mucinogeno, se adosa a laluz del intestino o la porcion conductora de las vıasrespiratorias. El mucinogeno, que se libera a raız de laexposicion a compuestos nocivos o a mol�eculas neu-rotransmisoras procedentes del sistema nervioso para-simp�atico, se hidrata para dar lugar a una sustanciaviscosa resbaladiza llamada mucina, que recibe elnombre demucosidad al combinarse con otros com-puestos presentes en la luz.
58
5
EPITELIOYGLANDULAS

CONSIDERACIONES CL�INICAS
El sındrome de Sj€ogren es una enfermedadautoinmune inflamatoria cronica que afecta a las
gl�andulas salivales y lagrimales, que no puedenliberar los productos de secrecion, de modo que elpaciente presenta xerostomıa y xeroftalmıa. Laenfermedad puede aparecer por sı sola o bien
asociarse a otros trastornos de base, como artritisreumatoide, lupus y esclerodermia; se asocia,
tambi�en, a linfomas. El sındrome de Sj€ogren es
nueve veces m�as frecuente en mujeres que enhombres. No existe ningun tratamiento frente a estaentidad en la actualidad.
59
5
EPITELIOYGLANDULAS
Figura 5.8 Modos de secrecion glandular. A. Holocrina. B. Merocrina. C. Apocrina. (Tomado de Gartner LP, Hiatt JL: Color Textbook of
Histology, 3rd ed. Philadelphia, Saunders, 2007, p 105.)
Figura 5.9 Esquema de la ultraestructura de una c�elula calici-
forme. (Tomado de Lentz TL: Cell Fine Structure: An Atlas ofDrawings of Whole-Cell Structure. Philadelphia, Saunders, 1971.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Gl�andulas exocrinas multicelulares
Las c�elulas secretoras forman asociaciones para actuarconjuntamente como organos secretores denomina-dos gl�andulas exocrinas multicelulares. Algunas deestas gl�andulas poseen una estructura sencilla (p. ej.,mucosa g�astrica y utero),mientras que otras presentanuna estructura compleja integrada por varios tipos deunidades secretoras con elementos ramificados (p. ej.,gl�andula submandibular). La clasificacion de las gl�an-dulasmulticelulares se fundamenta en lamorfologıa yla organizacion de las unidades secretoras y sus con-ductos, segun lo cual pueden ser:
. Sencillas, carentes de conductos ramificados.
. Compuestas, dotadas de conductos ramificados.
Lamorfologıa de las unidades secretoras de los con-ductos compuestos permite clasificarlas en acinares(alveolares), tubulares o tubuloalveolares (fig. 5.10).
El tejido conjuntivo colagenoso forma c�apsulasalrededor de las gl�andulas multicelulares de grantamano, ası como unos tabiques denominados septosque confieren apoyo estructural a la gl�andula al sub-dividirla en lobulos y lobulillos (fig. 5.11). Los ner-vios, los vasos sanguıneos y los conductos ingresan yabandonan la gl�andula a trav�es de orificios presentesen los tabiques.
Las c�elulasmioepiteliales –c�elulas de origen epite-lial que pueden contraerse– aparecen en las gl�andulassalivales mayores y las gl�andulas sudorıparas, en lasque comparten la l�amina basal con los �acinos glandu-lares. Tanto los �acinos como los conductos depequeno calibre se rodean de hebras fibrilares de cito-plasma procedentes de dichas c�elulas. La contraccionde las c�elulas mioepiteliales comprime los �acinos y lospequenos conductos, que se ven obligados a liberarsus productos de secrecion.
GLANDULAS ENDOCRINAS
Las gl�andulas endocrinas son las gl�andulas suprarrena-les (adrenales), la gl�andula tiroides, la hipofisis, lagl�andula paratiroides, los ovarios, los testıculos, la pla-centa y la gl�andula pineal. Todas ellas est�an exentas deconductos, por lo que secretan sus productos (hormo-nas) al torrente circulatorioo los vasos linf�aticospara sudistribucion a los organos diana. Algunas de estasgl�andulas (como los islotes de Langerhans del p�ancreasy las c�elulas intersticiales de Leydig de los testıculos) secomponen de grupos de c�elulas incluidas en el estromaconjuntivo de los organos que las contienen.
Las hormonas secretadas por las gl�andulas endocri-nas pueden ser proteınas, p�eptidos, esteroides, amino-�acidos modificados y glucoproteınas (v. capıtulo 13).Las c�elulas secretoras endocrinas se organizanformando cordones o folıculos. Los primeros, m�as fre-
cuentes, suelen anastomosarse alrededor de los capila-res o los sinusoides sanguıneos. La liberacion de lashormonas que contienen en el interior de la c�elulaobedece a la estimulacion neural o la recepcion deuna mol�ecula de senalizacion. Las gl�andulas endocri-nas con morfologıa filiforme son las gl�andulas parati-roidea y suprarrenales y la adenohipofisis. En lasgl�andulas endocrinas con organizacion folicular, lasc�elulas foliculares (c�elulas secretoras) se disponen alre-dedor de una depresion o cavidad; no almacenan losproductos que sintetizan, por lo que los liberan a dichacavidad, en la que se almacenan. Al recibir una senaladecuada, las c�elulas foliculares reabsorben las hormo-nas de la cavidad y la secretan a capilares sanguıneospresentes en el tejido conjuntivo asociado (p. ej.,gl�andula tiroidea).
Otras gl�andulas son mixtas, es decir, poseen tantounidades secretoras exocrinas como endocrinas. Este esel caso del p�ancreas, los ovarios y los testıculos. La por-cion exocrina libera la hormona a un conducto, mien-tras que la endocrina lo hace al torrente circulatorio.
Sistema neuroendocrino difuso
Las c�elulas endocrinas tambi�en aparecen entre las c�e-lulas epiteliales que revisten el tubo digestivo. Estasc�elulas endocrinas constituyen el sistemaneuroendo-crino difuso (SNED) y sintetizan algunas hormonasparacrinas y endocrinas. El t�ermino SNED ha susti-tuido a los de c�elulas argentafines, c�elulas argirofilasy c�elulas APUD (v. capıtulo 17).
CONSIDERACIONES CL�INICAS
Los tumores carcinoides provienen de c�elulasSNED, principalmente del tubo digestivo. Con
anterioridad al ano 2000, este t�ermino se aplicabaa las variantes benignas y malignas derivadas de
dichas c�elulas. A partir de entonces, los tumoresbenignos procedentes de SNED se denominan
tumores neuroendocrinos; las c�elulas quemigran a otras regiones del organismo dan lugar aunas lesiones denominadas carcinoides. Seutilizan los siguientes t�erminos para describirestos tumores: tumores neuroendocrinos(carcinomas), bien diferenciados (menos
agresivos) y poco diferenciados (m�as agresivos).
Sin embargo, muchos m�edicos se inclinan, aun,
por el t�ermino carcinoide para los tumoresbenignos bien diferenciados. A medida que sedesarrollan y diseminan, estos tumores yneoplasias procedentes de SNED secretansustancias similares a hormonas que originanrubefaccion, estornudos, diarrea y taquicardia,sıntomas que reciben, en su conjunto, el nombrede sındrome carcinoide. Por otra parte, estostumores y neoplasias pueden manifestarse consıntomas en cualquier region del organismo.
60
5
EPITELIOYGLANDULAS

61
5
EPITELIOYGLANDULAS
Figura 5.10 Clasificacion de las gl�andulas exocrinas multicelulares. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.Philadelphia, Saunders, 2007, p 107.)
Figura 5.11 Gl�andula salival: organizacion, uni-
dades secretoras y sistema de conductos.
(Tomado de Gartner LP, Hiatt JL: Color Textbookof Histology, 3rd ed. Philadelphia, Saunders, 2007,
p 108.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

6 TEJIDO CONJUNTIVOEl tejido conjuntivo, uno de los cuatro tejidos b�asicosdel organismo, procede mayoritariamente del meso-dermo y conecta entre sı a los otros tres tejidos b�asicosy los distintos tipos de tejido conjun-tivo. A lo largo del desarrolloembrionario, las c�elulas mesenqui-matosas pluripotenciales del tejidoconjuntivo embrionario primitivo,el mes�enquima, migran por todo elorganismo para diferenciarse enc�elulas maduras de tejido conjuntivoespecializado, como el tejido cartila-ginoso, el tejido oseo y la sangre.Asimismo, las c�elulas mesenquima-tosas dan lugar a las c�elulas del tejidoconjuntivo no especializado, eltejido conjuntivo propiamentedicho, como fibroblastos, adipocitosy mastocitos.
Los distintos tipos de tejido con-juntivo llevan a cabo funciones diversas y de granalcance:
. El cartılago, el hueso, los tendones, los ligamentos ylas c�apsulas de los organos ofrecen soporteestructural.
. La sangre, la linfa y el tejido conjuntivopropiamente dicho actuan como medio deintercambio de nutrientes, residuos y mol�eculas desenalizacion para muchas c�elulas del organismo.
. Algunas c�elulas que migran en el torrentecirculatorio pueden extravasarse hacia el tejidoconjuntivo propiamente dicho para desempenarfunciones de defensa y proteccion del organismofrente a sustancias o microorganismosposiblemente nocivos.
. Los adipocitos almacenan lıpidos y se agrupan en eltejido adiposo para formar depositos locales degrasa.
El tejido conjuntivo propiamente dicho se com-pone de matriz extracelular y c�elulas, algunas de lascuales intervienen en la sıntesis de la matriz en la quetanto ellas como otras c�elulas se encuentran embebi-das. La predominancia de c�elulas omatriz extracelularpara formar su elemento principal en cada tipo detejido conjuntivo se relaciona con su funcion. De estemodo, las fibras son m�as importantes que las c�elulas,los fibroblastos, en los tendones y los ligamentos,mientras que los filamentos desempenan una funcionm�as relevante que las fibras en el tejido conjuntivolaxo. En otros casos, como en la respuesta inmuni-
taria, la sustancia fundamental juega un papel m�asdestacado que las c�elulas y las fibras, ya que la funciondefensiva del organismo depende de las caracterısticas
de dicha sustancia.La matriz extracelular, el compo-
nente inerte del tejido conjuntivo,est�a formada por sustancia funda-mental y fibras; aunque se describeen el capıtulo 4, en este apartado se re-pasar�an sus caracterısticas m�as sobresa-lientes. La sustancia fundamental secompone de:
. Glucosaminoglucanos, sulfatados(p. ej., queratano sulfato, heparina,condroitina sulfato, dermatanosulfato y heparano sulfato)o no sulfatados (p. ej., �acidohialuronico).
. Proteoglucanos, los cuales formanmacromol�eculas de agrecano al establecer enlacescovalentes con mol�eculas de �acido hialuronico,lo que hace posible el estado de gel de la matrizextracelular.
. Algunas glucoproteınas de adhesion, como lafibronectina, que se distribuye en la matrizextracelular, y la laminina, de distribucion amplia ylocalizada en la l�amina basal. Otras proteınas, comola condronectina, se encuentran en el cartılago,mientras que la osteonectina aparece en el hueso.
Las fibras, inertes, se agrupan en dos grandesclases:
. Fibras de col�ageno, subdivididas en 25 tiposdiferentes segun la secuencia aminoacıdica de lastres cadenas alfa, si bien unicamente seis resultan deinter�es para los lectores de esta obra (tabla 6.1). Lamayorıa de las fibras de col�ageno poseen una granfuerza de tension. Los amino�acidos m�as frecuentesen el col�ageno son la glicina, la prolina, lahidroxiprolina y la hidroxilisina.
. Fibras de elastina, formadas por elastina ymicrofibrillas. La proteına amorfa elastina,compuesta principalmente por glicina y prolina,les confiere elasticidad (p. ej., las fibras el�asticaspueden estirarse hasta el 150% de su longitud),mientras que su estabilidad depende de lasmicrofibrillas. De igual modo, en la elastina abundala lisina, un amino�acido implicado en la formacionde enlaces de desmosina caracterizados por suelasticidad y deformabilidad.
TERMINOS CLAVE. Matriz extracelular. C�elulas del tejido
conjuntivo. Almacenamiento de
lıpidos enadipocitos
. Respuestainflamatoria
. Tipos de tejidoconjuntivo
� 2011 Elsevier Espana, S.L. Reservados todos los derechos62

CONSIDERACIONES CL�INICAS
El sındrome de Ehlers-Danlos engloba un grupo de
enfermedades gen�eticas infrecuentes en el serhumano debidas a anomalıas en la sıntesis de
col�ageno. Los sıntomas muestran una notablevariabilidad en funcion del tipo de sındrome. En cadacaso, la sintomatologıa se debe, en ultima instancia,a la sıntesis defectuosa o reducida de fibras de
col�ageno; entre los sıntomasm�as comunes figuran lainestabilidad articular, de modo que lasarticulaciones se dislocan con facilidad y presentanhipermovilidad como consecuencia de ladistensibilidad excesiva de los ligamentos formados
por fibras anomalas de col�ageno. Algunas variantesafectan a la piel, mientras que otras lo hacen a lasparedes de los vasos sanguıneos. La gravedadde las modalidades de este trastorno incurablecomprende de leve a potencialmente mortal.
El sındrome de Marfan es un trastorno autosomicodominante caracterizado por la debilidad del tejido
el�astico debido a una mutacion en el gen quecodifica la fibrilina. Este trastorno afecta a las fibras
el�asticas de los sistemas cardiovascular, ocular y
esquel�etico. Las personas aquejadas de estesındrome son muy altas y poseen brazos, piernas,pies y dedos de pies y manos muy largos. Lasalteraciones cardiovasculares pueden provocar lamuerte y engloban problemas vasculares ydilatacion de la aorta ascendente. Lasmanifestaciones oculares incluyen miopıa ydesprendimiento del cristalino. Las alteraciones
esquel�eticas comprenden debilidad anomala delperiostio como consecuencia de defectos en las
fibras el�asticas, que no pueden ejercer una fuerzade aposicion en el desarrollo oseo.
63
6
TEJIDOCONJUNTIVO
Tabla 6.1 TIPOS PRINCIPALES Y CARACTERISTICAS DEL COLAGENO
Tipo molecular Formula molecularC�elulas encargadasde la sıntesis Funcion
Localizacionen organismo
I (formadoras de
fibrillas); son las
m�as frecuentes
[a(I)]2 a2(I) Fibroblastos,
osteoblastos,
odontoblastos,
cementoblastos
Resistencia a la tension Dermis, tendones,
ligamentos,
c�apsulas de
organos, hueso,dentina, cemento
II (formadoras de
fibrillas)
[a1(II)]3 Condroblastos Resistencia a la presion Cartılago hialino,
cartılago el�astico
III (formadoras defibrillas); tambi�en
llamadas fibras
reticulares; elevadogrado de
glucosilacion
[a1(III)]3 Fibroblastos, c�elulasreticulares, c�elulas
del musculo liso,
hepatocitos
Marco estructural delbazo, hıgado,
ganglios linf�aticos,
musculo liso, tejidoadiposo
Sistema linf�atico,bazo, hıgado,
sistema
cardiovascular,pulmon, piel
IV (formadoras de
redes); carecen debandeado de
67 nm, las cadenas
alfa conservan los
prop�eptidos
[a1(IV)]2a2(IV) C�elulas epiteliales,
c�elulas musculares,c�elulas de Schwann
Malla de la l�amina
densa de la l�aminabasal que ofrece
sost�en y filtracion
L�amina basal
V (formadoras de
fibrillas)
[a1(V)]2a2(V) Fibroblastos, c�elulas
mesenquimatosas
Asociadas al col�ageno
de tipo I, tambi�en en
la sustanciafundamental
placentaria
Dermis, tendones,
ligamentos,
c�apsulas deorganos, hueso,
cemento, placenta
VII (formadoras de
redes); formandımeros que se
organizan en
fibrillas de anclaje
[a1(VII)]3 C�elulas epid�ermicas Formacion de fibrillas
de anclaje quefavorecen la union
de la l�amina densa
con la l�amina
reticular subyacente
Union de epidermis y
dermis
Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 76.
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Componente celular del tejido conjuntivo
Las c�elulas del tejido conjuntivo propiamente dichose dividen en dos grupos: fijas (residentes), aquellasque nomigran; y transitorias, aquellas que utilizan eltorrente circulatorio y los vasos linf�aticos para accederal tejido conjuntivo propiamente dicho, donde mue-ren o bien se desplazan hacia otras zonas del orga-nismo (tabla 6.2).
CELULAS FIJAS DEL TEJIDO CONJUNTIVO
Fibroblastos
Los fibroblastos (figs. 6.1 y 6.2), las c�elulasm�as abun-dantes en el tejido conjuntivo, provienen de c�elulasmesenquimatosas y se ocupan de fabricar la matrizextracelular. Pueden ser activos o inactivos; los miofi-broblastos constituyen una subpoblacion de los fibro-blastos.
. Los fibroblastos activos se disponen en paralelo aleje largo de los haces de col�ageno; se trata de c�elulasfusiformes alongadas con un citoplasma de tincionp�alida y un gran nucleo ovalado de color oscuro. Elaparato de Golgi y el retıculo endopl�asmico rugoso(RER) de los fibroblastos que est�an sintetizando lamatriz se encuentran bien desarrollados. Lamiosinase distribuye en el citoplasma,mientras que la actinay la a-actinina aparecen preferentemente en laperiferia celular.
. Los fibroblastos inactivos presentan un tamanomenor, poseen un citoplasma acidofilo y un nucleom�as denso de tincion muy oscura. El RER y elaparato de Golgi no son tan prominentes en estasc�elulas, en las que abundan los ribosomas.
. Los fibroblastos pueden diferenciarse enmiofibroblastos en zonas de cicatrizacion.Presentan rasgos similares a los de los fibroblastos ylas c�elulas del musculo liso, aunque a diferencia deestas ultimas carecen de l�amina externa. Losmiofibroblastos participan en la contraccion de lasheridas; en el ligamento periodontal aparecen comoc�elulas residentes y podrıan favorecer la erupciondental.
Pericitos
Los pericitos, derivados de las c�elulas mesenquima-tosas, rodean parcialmente las c�elulas endotelialesde los capilares sanguıneos y las v�enulas pequenas(v. fig. 6.2). Est�an dotados de una l�amina basal, quepuede fusionarse con la de las c�elulas endotelialescercanas. Los pericitos comparten algunas caracterıs-ticas de las c�elulas del musculo liso y las c�elulas endote-liales; pueden diferenciarse en fibroblastos, c�elulasendoteliales o c�elulas del musculo liso vascular al serinducidas por una agresion ambiental.
Adipocitos
Las c�elulas adiposas (adipocitos) son c�elulas amito-ticas encargadas de sintetizar y almacenar triglic�eridos(v. fig. 6.2). Se distinguen dos tipos de adipocitos:uniloculares y multiloculares.
. Los adipocitos uniloculares, unas c�elulasredondeadas de gran tamano (di�ametro � 120mm)rellenas de una sola gotıcula de lıpido, predominanen el tejido adiposo blanco. En la microscopiaelectronica, presentan un delgado halocitopl�asmico con abundantes ribosomas, unaparato de Golgi poco desarrollado, escasasmitocondrias, RER y numerosas vesıculaspinocıticas en las proximidades de la caracitopl�asmica de la membrana celular.
. Los adipocitos multiloculares poseen unamorfologıa poligonal, un tamano menor que losadipocitos del tejido adiposo blanco, y numerosasgotıculas con lıpidos dispersas en su citoplasma. Lasmicrofotografıas electronicas ponen de manifiesto lapresencia de abundantesmitocondrias, que confierena la c�elula su coloracion oscura –de donde provieneel nombre de adipocitos del tejido adiposo pardo, elprincipal componente celular de este tejido.
CONSIDERACIONES CL�INICAS
A pesar de que los fibroblastos se clasifican como
c�elulas fijas, est�an dotados de una cierta
capacidad de movimiento. Estas c�elulas puedensufrir divisiones en ciertas condiciones, comosucede en el transcurso de la cicatrizacion deheridas. Por otra parte, la distension de lostendones debido a su utilizacion excesiva estimulala diferenciacion de los fibroblastos encondrocitos, los cuales se rodean de matrizcartilaginosa sintetizada por ellos y transformanel tendon en fibrocartılago. Asimismo, losfibroblastos pueden diferenciarse en adipocitos;en condiciones patologicas, incluso llegan adiferenciarse en osteoblastos.
64
6
TEJIDOCONJUNTIVO
Tabla 6.2 CELULAS FIJAS Y TRANSITORIAS
C�elulas fijas C�elulas transitorias
Fibroblastos C�elulas plasm�aticas
Adipocitos LinfocitosPericitos Neutrofilos
Mastocitos Eosinofilos
Macrofagos BasofilosMonocitos
Macrofagos

65
6
TEJIDOCONJUNTIVO
Figura 6.2 Tipos de c�elulas y fibras del tejido conjuntivo laxo. (Tomado deGartner LP,
Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 113.)
Figura 6.1 Origen de las c�elulas del tejido
conjuntivo. (Tomado de Gartner LP, HiattJL: Color Textbook of Histology, 3rd ed.
Philadelphia, Saunders, 2007, p 112.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Almacenamiento y liberacion de lıpidos por los adipocitos
Durante la digestion, la lipasa pancre�atica catabolizalos lıpidos presentes en la luz del intestino delgado en�acidos grasos y glicerol, los cuales son absorbidospor las c�elulas de absorcion de superficie del revesti-miento epitelial. En el citoplasma de estas c�elulas, los�acidos grasos y el glicerol pasan al retıculo endopl�as-mico liso, en el cual se esterifican de nuevo y se trans-portan al aparato de Golgi, en el que se rodean deproteınas. Estos triglic�eridos recubiertos de proteınas,los quilomicrones, se liberan hacia la l�amina propiadel intestino delgado y pasan a unos canales linf�aticosllamados lacteales para alcanzar, m�as tarde, el torren-te circulatorio.
Los capilares que irrigan el tejido adiposo poseenuna enzima, la lipoproteına lipasa, que fabrican losadipocitos y se localiza en la superficie luminal delas c�elulas endoteliales (fig. 6.3). Esta enzima cata-boliza los quilomicrones y otros lıpidos s�ericos,como las lipoproteınas de densidad muy baja(VLDL), que convierte en glicerol y �acidos grasos.Estos ultimos abandonan los capilares, atraviesanla membrana plasm�atica de los adipocitos y sereconstituyen en triglic�eridos en el citoplasma delos adipocitos, en los que se almacenan en formade gotıculas lipıdicas, una vıa eficaz y de bajo peso dealmacenamiento de energıa. La union de noradre-nalina y adrenalina a sus respectivos receptores enla membrana de los adipocitos provoca la activaciondel sistema de la adenilato ciclasa, la cual producemonofosfato de adenosina cıclico (AMP) para indu-cir la degradacion de los triglic�eridos de la gotıcula delıpidos por accion de una lipasa sensible a hormo-nas presente en el citoplasma. Los �acidos grasos y elglicerol salen del adipocito para pasar a los capilares(v. fig. 6.3).
Mastocitos
Los mastocitos (di�ametro, 20 a 30mm) provienen dec�elulas progenitoras de m�edula osea, que migran altejido conjuntivo para madurar, subsistir y, rara vez,dividirse. Estas c�elulas ovoides dotadas de un nucleocentral poseen gr�anulos limitados por membrana(fig. 6.4. y tabla 6.3) que confieren la propiedad demetacromasia. Los mastocitos almacenan algunasmol�eculas farmacologicas, llamadas mediadores pri-marios o preformados, en estos gr�anulos y sintetizanotras, los mediadores secundarios, con arreglo a lasnecesidades celulares.
. Los mediadores primarios son histamina yheparina (en los mastocitos del tejido conjuntivo)o bien histamina y condroitina sulfato (en losmastocitos de la mucosa de las vıas respiratorias yel tubo digestivo), proteasas neutras (triptasa,
quimasa y carboxipeptidasas), arilsulfatasa,b-glucuronidasa, cininogenasa, peroxidasa,superoxido dismutasa, factor quimiot�acticode eosinofilos y factor quimiot�actico deneutrofilos.
. Los mediadores secundarios, sintetizados a partirde precursores del �acido araquidonico de lamembrana son los leucotrienos (C4, D4 y E4), lostromboxanos (tromboxano A2 y tromboxano B2)y las prostaglandinas (prostaglandina D2).
. Entre los mediadores secundarios que noprovienen de precursores del �acido araquidonicofiguran el factor activador de plaquetas, lasbradicininas, las interleucinas (IL-4, IL-5 e IL-6) y elfactor de necrosis tumoral a. (En la tabla 6.3 seofrece una relacion de los principales mediadoresprimarios y secundarios liberados por losmastocitos.)
Activacion y desgranulacion de los mastocitos
Lasmembranas plasm�aticas de losmastocitos cuentancon receptores de Fc de superficie de gran afinidad(FceRI) especıficos para mol�eculas de IgE que seproyectan hacia el espacio extracelular. Estas c�elulaspueden liberarmediadores quımicos que inducen unarespuesta localizada denominada reaccion de hiper-sensibilidad inmediata o, en casos extremos, unareaccion de consecuencias mortales denominadareaccion anafil�actica. Algunos f�armacos, venenos deciertos insectos, granos de polen y otros antıgenospueden desencadenar estas respuestas del modo quese describe a continuacion (v. fig. 6.4):
1. Los mastocitos se sensibilizan al unirse aanticuerpos IgE frente a un antıgeno determinado atrav�es de sus receptores FceRI, aunque noresponden al antıgeno en esta primera exposicion.
2. Al entrar en contacto con el tejido conjuntivo porsegunda vez, el antıgeno se une a las mol�eculas deIgE presentes en la superficie de los mastocitos, loque provoca su entrecruzamiento y aproxima a losreceptores.De estemodo, se estimula la activacion dela adenilato ciclasa y la fosfolipasa A2 por parte defactores de acoplamiento a receptores.
3. La adenilato ciclasa sintetiza AMP cıclico, cuyasconcentraciones citosolicas aumentan. Se induce,ası, la liberacion de iones de Ca++ desde loscompartimentos celulares en los que se almacenan,lo que desencadena la exocitosis de mediadorespreformados mediante un proceso dedesgranulacion.
4. La fosfolipasa A2 induce la sıntesis de �acidoaraquidonico, el cual se transforma enmediadores secundarios que pasan de inmediatoal espacio extracelular.
66
6
TEJIDOCONJUNTIVO
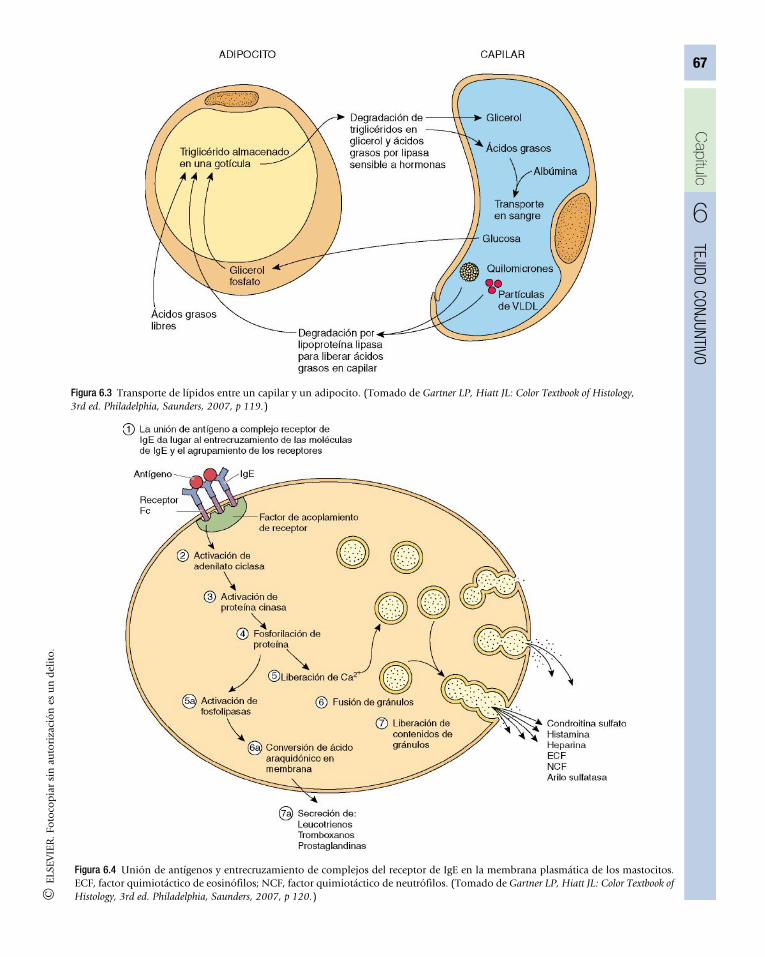
67
6
TEJIDOCONJUNTIVO
Figura 6.3 Transporte de lıpidos entre un capilar y un adipocito. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology,
3rd ed. Philadelphia, Saunders, 2007, p 119.)
Figura 6.4 Union de antıgenos y entrecruzamiento de complejos del receptor de IgE en la membrana plasm�atica de los mastocitos.
ECF, factor quimiot�actico de eosinofilos; NCF, factor quimiot�actico de neutrofilos. (Tomado de Gartner LP, Hiatt JL: Color Textbook of
Histology, 3rd ed. Philadelphia, Saunders, 2007, p 120.)�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Mastocitos y respuesta inflamatoria
La liberacion de mediadores (primarios y secunda-rios) por parte de los mastocitos (v. tabla 6.3) comorespuesta a la acumulacion de antıgenos en sus IgE desuperficie desencadena la secuencia de acontecimien-tos que se describe a continuacion:
1. La histamina ejerce una accion vasodilatadora eincrementa la permeabilidad vascular; asimismo,actua como vasoconstrictor y, adem�as de reducir eldi�ametro de la luz de los bronquıolos, estimula laproduccion de mucosidad.
2. El tejido conjuntivo recibe mol�eculas delcomplemento debido a la extravasacion de plasmadesde los vasos sanguıneos; las proteasas neutrasmetabolizan el complemento en macromol�eculasque intervienen en el proceso inflamatorio.
3. Los factores quimiot�acticos de neutrofilos yeosinofilos reclutan neutrofilos y eosinofiloshacia el foco de la inflamacion; los primerosdestruyen microorganismos, mientras que lossegundos fagocitan complejos antıgeno-anticuerpoy destruyen par�asitos.
4. De igual modo, las bradicininas incrementan lapermeabilidad vascular y producen sensacion dedolor en el �area de inflamacion.
5. Los leucotrienos C4, D4 y E4 desempenan unasfunciones semejantes a las de la histamina, si bienejercen una accion mucho m�as potente; no est�animplicados en la produccion de mucosidad.
6. La prostaglandina D2 origina la contraccion de lamusculatura lisa de los bronquıolos y potencia laproduccion de mucosidad.
7. El factor activador de plaquetas recluta neutrofilosy eosinofilos hasta el �area de inflamacion,incrementa la permeabilidad vascular y actua comobroncoconstrictor.
8. El tromboxano A2, el cual se inactiva con granrapidez al convertirse en tromboxano B2 ejerce unaaccion vasoconstrictora e induce la agregacionplaquetaria.
Macrofagos
Losmacrofagos, unas c�elulas de morfologıa irregular yun di�ametro comprendido entre 10 y 30mm, son fago-citos y pertenecen al sistema fagocıtico mononuclear,un grupo de c�elulas derivadas de precursores hemato-poy�eticos comunes. Los macrofagos se desplazan a lolargo del torrente circulatorio en forma demonocitos ymaduran para diferenciarse en macrofagos al pasar al
tejido conjuntivo. Algunosmacrofagos se asientan en el�area del organismo a la que acceden al abandonarlos vasos sanguıneos y reciben el nombre de macrofa-gos residentes (fijos) (p. ej., las c�elulas de Kupffer, lasc�elulas de Langerhans, las c�elulas de polvo, las c�elulasde la microglia), mientras que los macrofagos transi-torios (libres) son aquellos que llevan a cabo sus fun-ciones y despu�es mueren en el �area en el que hanactuado o bien migran hacia otras regiones.
Algunos macrofagos que han de eliminar partıculasde gran tamano se fusionan entre sı con el fin dedesempenar esta tarea, como los osteoclastos y lasc�elulas gigantes de cuerpo extrano. Las membranascelulares de los macrofagos son lisas; sin embargo,cuando se desplazan de forma activa o fagocitan com-puestos exogenos o residuos celulares, presentanrepliegues. Algunos macrofagos han de ser activadospor mol�eculas de senalizacion secretadas por linfoci-tos implicados en la respuesta inmunitaria para reali-zar sus funciones (v. capıtulo 12). El citoplasma de losmacrofagos maduros contiene numerosas vacuolas,un aparato de Golgi bien desarrollado, abundanteslisosomas, un gran numero de microtubulos y unRER prominente. Presentan un nucleo denso de mor-fologıa arrinonada. Aparte de la fagocitosis de micro-organismos invasores y residuos celulares y extracelu-lares, los macrofagos sintetizan y liberan mol�eculas desenalizacion, como el factor de necrosis tumoral-a eIL-1, adem�as de actuar como c�elulas presentadorasde antıgenos que transportan fragmentos antig�enicosen receptores de membrana hasta los linfocitos, loscuales ponen en marcha la respuesta inmunitaria.
CELULAS TRANSITORIAS DEL TEJIDO CONJUNTIVO
C�elulas plasm�aticas
Las c�elulas plasm�aticas, que derivan de un subgrupode linfocitos (linfocitos B) activados por contacto conun antıgeno, son c�elulas ovaladas de gran tamano(aproximadamente, 20mmde di�ametro) con un nucleoexc�entrico denso cuya heterocromatina muestra unaconfiguracion tıpica de esfera de reloj o rueda de carro(fig. 6.5). El citoplasma de estas c�elulas posee un apa-rato de Golgi y un RER bien desarrollados, ya que parti-cipan activamente en la sıntesis de anticuerpos frente aantıgenos. La vida media de estas c�elulas comprende de2 a 3 semanas. Se distribuyen en los tejidos conjuntivosdel organismo, aunque sonm�as abundantes en �areas deinflamacion cronica y zonas susceptibles a la invasionpor antıgenos o microorganismos, como la l�amina pro-pia del tubo digestivo y las vıas respiratorias.
68
6
TEJIDOCONJUNTIVO

69
6
TEJIDOCONJUNTIVO
Figura 6.5 Esquema de una c�elula plasm�atica basado en una microfotografıa. (Tomado de Lentz TL: Cell Fine Structure: An Atlas ofDrawings of Whole-Cell Structure. Philadelphia, Saunders, 1971.)
Tabla 6.3 PRINCIPALES MEDIADORES PRIMARIOS Y SECUNDARIOS LIBERADOS POR LOS MASTOCITOS
SustanciaTipo demediador Origen Accion
Histamina Primario Gr�anulo Aumenta la permeabilidad vascular, vasodilatacion,
contraccion de musculatura lisa bronquiolar, produccion
de mucosidad
Heparina Primario Gr�anulo Anticoagulante que se une a la histamina para inactivarlaCondroitina sulfato Primario Gr�anulo Se une a la histamina y la inactiva
Arilo sulfatasa Primario Gr�anulo Inactiva el leucotrieno C4 para restringir la respuesta
inflamatoria
Proteasas neutras Primario Gr�anulo Escinden las proteınas para activar el complemento(en especial, C3a); potencian la respuesta inflamatoria
Factor quimiot�actico
de eosinofilos
Primario Gr�anulo Recluta eosinofilos hacia el �area inflamada
Factor quimiot�actico
de neutrofilos
Primario Gr�anulo Recluta eosinofilos hacia el �area inflamada
Leucotrienos C4, D4
y E4
Secundario Lıpidos de membrana Vasodilatadora; aumentan la permeabilidad vascular;
origina la contraccion de musculatura lisa bronquiolarProstaglandina D2 Secundario Lıpidos de membrana Origina la contraccion de musculatura lisa bronquiolar;
aumenta la secrecion de mucosidad; vasoconstriccion
Tromboxano A2 Secundario Lıpidos de membrana Origina la agregacion plaquetaria, vasoconstriccion
Bradicininas Secundario Formadas por accion deenzimas incluidas en
gr�anulos
Aumentan la permeabilidad vascular y originan sensacionde dolor
Factor activador deplaquetas
Secundario Activado por lafosfolipasa A2
Atrae los neutrofilos y eosinofilos; aumenta lapermeabilidad vascular y la contraccion de musculatura
lisa bronquiolar
Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 121.
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Leucocitos
Los leucocitos, o globulos blancos, circulan en el to-rrente circulatorio y migran al tejido conjuntivo al serreclutados por citocinas o bien reconocerlas a trav�es desus receptores de localizacion (v. fig. 6.4). En los capıtu-los 10 y 12 se describe con mayor detalle a estas c�elulas.
. Los monocitos se han abordado en un apartadoanterior relativo a los macrofagos.
. Los neutrofilos responden al factor quimiot�acticode neutrofilos liberado por los mastocitos y actuanfagocitando y digiriendo bacterias en las reaccionesde inflamacion aguda. Estas c�elulas mueren y pasana formar parte del pus tras su desgranulacion paradestruir a las bacterias.
. Los eosinofilos acuden al �area de inflamacion poraccion del factor quimiot�actico de eosinofilossintetizado por los mastocitos y actuan en lainflamacion aguda. Destruyen par�asitos y fagocitancomplejos antıgeno-anticuerpo.
. Los basofilos son unas c�elulas similares a losmastocitos con funciones similares a estos.
. Los linfocitos abundan especialmente en las �areasde inflamacion cronica.
Clasificacion del tejido conjuntivo
Se distinguen tres tipos de tejido conjuntivo. El tejidoconjuntivo embrionario est�a restringido a las fases dedesarrollo embrionario y fetal, si bien algunos autores
estiman que podrıa incluirse en la categorıa de tejidoconjuntivo propiamente dicho, el cual aparece entodo el organismo. El tejido conjuntivo especiali-zado engloba el cartılago, el hueso y la sangre. En latabla 6.4 se sintetizan las categorıas y subcategorıas deltejido conjuntivo.
TEJIDO CONJUNTIVO EMBRIONARIO
Se conocen dos tipos de tejido conjuntivo embrionario:
. Tejido conjuntivo mesenquimatoso, abunda en elembrion y el feto; se compone de una sustanciafundamental gelatinosa rica en �acido hialuronicoy fibras reticulares (fibras de col�ageno de tipo II)en la que se embeben las c�elulasmesenquimatosas.Estas c�elulas son c�elulas pluripotenciales conproyecciones relativamente largas en variasdirecciones que se alejan del cuerpo celular. Cadac�elula mesenquimatosa posee un nucleo ovoideunico de tincion p�alida con un nucl�eolo biendefinido que se rodea de finas fibras de cromatina.Las c�elulas mesenquimatosas tan solo est�anpresentes en algunas regiones del organismo adulto.
. Tejido conjuntivo mucoso, aparece por debajo dela epidermis del embrion y en el cordon umbilical,se compone de una sustancia fundamental rica en�acido hialuronico que alberga a fibroblastos y fibrasdelgadas de col�ageno de tipos I y III. En el cordonumbilical, este tejido recibe el nombre de gelatinade Wharton.
70
6
TEJIDOCONJUNTIVO
CONSIDERACIONES CL�INICAS
Las personas aquejadas de rinitis polınicapresentan edema localizado y tumefaccion de lamucosa nasal, lo que impide la respiracion y originala sensacion de taponamiento. Estos sıntomas sedeben a la liberacion de histamina por parte de losmastocitos de la mucosa nasal, lo cual incrementa lapermeabilidad de los vasos sanguıneos de pequenocalibre y produce edema localizado. La disnea sueleacompanarse de asma como consecuencia de laliberacion de leucotrienos en los pulmones, queprovoca broncoespasmo.
Normalmente, la desgranulacion de los mastocitoses una reaccion localizada que produce unarespuesta inflamatoria leve tıpica. Sin embargo, los
sujetos hiperal�ergicos presentan un riesgo de
anafilaxia sist�emica tras la reexposicion a un
al�ergeno (p. ej., picadura de abeja). Esta nuevaexposicion, que da lugar a una reaccion de
hipersensibilidad inmediata grave y sist�emica, recibe
el nombre de choque anafil�actico. Los sıntomas
aparecen inmediatamente despu�es o pasados unos
minutos y la muerte puede sobrevenir en el plazo deunas horas en ausencia de tratamiento. La
sintomatologıa engloba una disminucion r�apida de latension arterial y disnea. Se recomienda a estos
individuos que porten una pulsera m�edica, ya que
alerta al profesional m�edico de la necesidad deadministrar un tratamiento farmacologico sindemora.
Por lo general, el lıquido extracelular presente en lostejidos regresa a los capilares directamente o bien es
recogido por los vasos linf�aticos, desde los cualesretorna al torrente circulatorio. Este lıquido seacumula en el tejido conjuntivo laxo durante larespuesta inflamatoria, lo que impide su retorno a losvasos sanguıneos. Esta alteracion origina un edema(tumefaccion extensa), que puede deberse a laliberacion de cantidades excesivas de histamina y
leucotrienos C4 y D4, unas mol�eculas sintetizadaspor losmastocitos que incrementan la permeabilidadcapilar. De igual modo, el edema puede deberse a
obstrucciones en las venas o los vasos linf�aticos.

CONSIDERACIONES CL�INICAS
Se distinguen dos tipos de obesidad en el adulto:
. Obesidad hipertrofica, debida a un desequilibrioentre el aporte calorico y el gasto de energıa queda lugar a la acumulacion y el almacenamiento delıpidos en adipocitos uniloculares que llegan acuadriplicar su tamano.
. Obesidad hipercelular (hiperpl�asica), debida aun exceso de poblacion de adipocitos. Losadipocitos maduros no sufren mitosis, pero susprecursores sı lo hacen durante un breve perıododel desarrollo posnatal. Se dispone de datos que
indican que la sobrealimentacion del reci�ennacido, incluso durante varias semanas, provoca
la proliferacion de las c�elulas precursoras de losadipocitos, lo que da lugar a un aumento anomalo
de sus poblaciones y obesidad hiperpl�asica quepodrıa iniciarse en la ninez. Los lactantes consobrepeso tienen una probabilidad dos o tresveces mayor de padecer obesidad en la vidaadulta que los ninos con peso normal.
Los tumores localizados en el tejido adiposo puedenser benignos o malignos. Los lipomas son tumoresbenignos de los adipocitos, mientras que losliposarcomas son lesionesmalignas de los adipocitoso sus precursores. Los liposarcomas representan unaentidad frecuente, de modo que en EE. UU. sediagnostican alrededor de 2.000 nuevos casos cadaano. Se conocen tres tipos de liposarcoma:
. Liposarcoma bien diferenciado odesdiferenciado (aproximadamente, el 50% de
los casos; representa el tipom�as frecuente), el cualse desarrolla en forma de unmasa indolora de grantamano localizada en la cavidad abdominal o unaextremidad. El tratamiento de eleccion esquirurgico y la tasa de recidiva en la regionabdominal se situa entre el 70 y el 80%. La variante
«desdiferenciada» es la m�as agresiva, si bien no seconsidera un sarcoma de grado alto.
. Liposarcoma de c�elulas mixoides o redondas(alrededor del 40%).
. Liposarcoma pleomorfo (el 10%; es el tipomenos frecuente), el cual afecta a una extremidad,es agresivo y puede diseminarse a otras regiones,como el pulmon y las partes blandas.
Dos grupos de hormonas rigen la acumulacion delıpidos: las hormonas responsables del control delpeso a corto plazo, grelina y el p�eptido YY, y lasimplicadas en el control del peso a largo plazo,leptina e insulina.
. Las c�elulas P/D1 del epitelio g�astrico y el
p�ancreas sintetizan la grelina, la cual induce una
sensacion de apetito, mientras que las c�elulas Ldel revestimiento epitelial del ıleon y el colon
fabrican el p�eptido YY, que induce una sensacionde saciedad. La falta de sueno da lugar a unaumento de las concentraciones de grelina einduce sensacion de apetito.
. La leptina, sintetizada por los adipocitos del
tejido adiposo blanco, las c�elulas ov�aricas y los
miocitos, se une a receptores de c�elulaspertenecientes al «centro del apetito» del
hipot�alamo e induce sensacion de saciedad.Algunos sujetos presentan resistencia a laleptina y pueden padecer obesidad morbida a
pesar de unas concentraciones s�ericas elevadasde esta hormona. El tratamiento de estospacientes con leptina humana recombinante seha asociado a una elevada eficacia. La insulinaincrementa la cantidad de lıpidos almacenadosen los adipocitos uniloculares del tejido adiposoblanco al inducir la conversion de glucosa en
triglic�eridos en estas c�elulas.
71
6
TEJIDOCONJUNTIVO
Tabla 6.4 CLASIFICACION DEL TEJIDO CONJUNTIVO
A. Tejido conjuntivo embrionario
1. Tejido conjuntivo mesenquimatoso2. Tejido conjuntivo mucoso
B. Tejido conjuntivo propiamente dicho
1. Tejido conjuntivo laxo (areolar)2. Tejido conjuntivo denso
a. Tejido conjuntivo irregular denso
b. Tejido conjuntivo regular denso
(1) Colagenoso(2) El�astico
3. Tejido reticular
4. Tejido adiposo
C. Tejido conjuntivo especializado1. Cartılago
2. Hueso
3. Sangre
Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology,
3rd ed. Philadelphia, Saunders, 2007, p 126.
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

TEJIDO CONJUNTIVO PROPIAMENTE DICHO
El tejido conjuntivo propiamente dicho puede divi-dirse en cuatro tipos principales: tejido conjuntivolaxo, tejido conjuntivo denso, tejido reticular y tejidoadiposo, cada uno de los cuales presenta rasgos yfunciones distintivas.
Tejido conjuntivo laxo (areolar)
La matriz extracelular del tejido conjuntivo laxo (areo-lar) est�a formada por una red irregular de fibras decol�ageno de tipos I y III en la que aparecen algunasfibras el�asticas delgadas y largas, todas las cuales seembeben en una sustancia fundamental gelatinosa.Las c�elulas que integran el tejido conjuntivo propia-mente dicho tambi�en se hallan en el tejido conjuntivolaxo sano y reciben sus nutrientes del lıquido extrace-lular que se extravasa de las abundantes arteriolas y lacara arterial de los lechos capilares, se percola a trav�es dela sustancia fundamental y regresa a la cara venosade los lechos capilares y la tupida red de v�enulas y vasoslinf�aticos. El tejido conjuntivo laxo est�a presentepor debajo de la piel y envuelve a los haces neurovas-culares.
Tejido reticular
El tejido reticular, que contiene principalmente fibrasde col�ageno de tipo III, conforma elmarco reticular dealgunos organos, como el hıgado, el bazo, la m�edulaosea, el musculo liso, el tejido adiposo y los ganglioslinf�aticos. Los fibroblastos suelen encargarse de lasıntesis de las fibras reticulares, aunque son las c�elulasdelmusculo liso las que fabrican las fibras de col�agenode tipo III en ese tejido.
Tejido conjuntivo denso
En el tejido conjuntivo denso, las fibras son muchom�as abundantes y las c�elulas m�as escasas que en ellaxo. El tejido conjuntivo denso se divide en regular eirregular con arreglo a la precision de la disposicion delas fibras que lo integran:
. Los haces de fibras de col�ageno de tipo I del tejidoconjuntivo colagenoso denso irregular se disponende manera aparentemente aleatoria, lo que confiere aeste tejido gran flexibilidad y elasticidad, adem�as deresistencia a las fuerzas de tension. El tejidoconjuntivo colagenoso denso irregular forma ladermis de la piel, las c�apsulas de un gran numero deorganos y las vainas de tejido conjuntivo de losnervios y los vasos sanguıneos de gran calibre.
. El tejido conjuntivo denso regular se subdivide endos grupos, colagenoso y el�astico, en funcion
de cu�ales sean las fibras predominantes en elmismo.. Tejido conjuntivo colagenoso regular denso, elcual constituye los tendones, las aponeurosis y losligamentos. Est�a formado mayoritariamente porhaces gruesos de fibras de col�ageno de tipo Iorientados en paralelo y empaquetados de talmanera que apenas contienen sustanciafundamental y fibroblastos comprimidos entreellos.
. Tejido conjuntivo el�astico regular denso,formado por haces de fibras el�asticas muyempaquetados dispuestos en paralelo entre loscuales aparecen fibroblastos. Este tejido formal�aminas perforadas, como en la membranafenestrada de la aorta, o bien haces cortos ygruesos, como en el ligamento nucal de lacolumna vertebral.
Tejido adiposo
Se distinguen dos clases de tejido adiposo con arregloal tipo de adipocitos presentes en el mismo: adipo-citos uniloculares del tejido adiposo blanco oadipocitos multiloculares del tejido adiposo pardo.El tejido adiposo pardo se forma a lo largo del desa-rrollo prenatal, mientras que el blanco lo hace en elperıodo posnatal.
. El tejido adiposo pardo (multilocular) presentauna organizacion m�as lobular que el tejido adiposoblanco y, a diferencia de este, las fibras nerviosasinervan tanto a los vasos sanguıneos como a losadipocitos multiloculares. Se cree que este tejidounicamente est�a presente en embriones y neonatosen el ser humano; despu�es del nacimiento, lasgotitas de grasa se unen hasta formar una sola gota.Los adipocitos multiloculares aparecen de nuevo enalgunas personas de edad avanzada afectadas porenfermedades de desgaste.
. El tejido adiposo blanco (unilocular) se componede adipocitos uniloculares (fig. 6.6) agrupados enlobulos divididos por tabiques incompletos detejido conjuntivo a trav�es de los cuales recibe suinervacion e irrigacion. La irrigacion vascular formaamplios lechos capilares en el interior de loslobulos, de modo que cada adipocito establece unaestrecha relacion con los capilares cercanos. Adem�asde localizarse en el tejido conjuntivo subcut�aneo, elepiplon, los mesenterios y las nalgas, existe unaacumulacion de tejido adiposo blanco que dependedel sexo. En la mujer, aparece en las mamas, lascaderas y los muslos, mientras que en el varondestaca en el cuello, los hombros y las caderas.
72
6
TEJIDOCONJUNTIVO

CONSIDERACIONES CL�INICAS
Los adipocitos multiloculares son muy abundantesen los animales que hibernan. Al recibir la senaladecuada (liberacion de noradrenalina por partede las fibras nerviosas que sinaptan con ellos),
estas c�elulas generan calor para despertar alanimal del estado de hibernacion. La capacidad deproduccion de calor de los adipocitos del tejidoadiposo se sustenta en la actividad de unaproteına transmembrana mitocondrial, llamadaproteına de desacoplamiento 1 (UPC-1) otermogenina,exclusiva de este tipo c�elulas.
En lugar de permitir el flujo de protones a trav�es del
complejo enzim�atico de la sintetasa de trifosfatode adenosina (ATP), dirige el flujo retrogrado deprotones y desacopla la oxidacion de lafosforilacion. De este modo, el flujo protonico nogenera ATP, sino que se disipa y produce calor.La sangre de la profusa red vascular del tejidoadiposo pardo se distribuye por todo elorganismo, como consecuencia de lo cualaumenta su temperatura corporal y despierta desu estado de hibernacion.
73
6
TEJIDOCONJUNTIVO
Figura 6.6 Origen de las c�elulas del tejido conjuntivo. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.
Philadelphia, Saunders, 2007, p 112.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

7 CARTILAGO Y HUESOEl cartılago y el hueso son los dos tipos de tejido con-juntivo especializado que se tratan en este capıtulo. Elcartılago es una estructura lisa y firme con una matrizflexible, mientras que el hueso est�aformado por una matriz calcificadaque lo hace inflexible. Las c�elulas delcartılago y el hueso quedan embebi-das en la matriz a medida que la vansecretando. El cartılago y el hueso serelacionan ıntimamente por su capa-cidad de resistencia al estr�es y elsoporte de los distintos elementosque integran el organismo; de igualmodo, el cartılago hialino se fabricaen primer lugar durante el desarrolloembrionario para formar la estructurasobre la que se sintetizar�an los huesoslargos. El cartılago se reabsorbe me-diante unproceso conocido como for-macion endocondral de hueso conforme se sintetizael hueso. La formacion de la mayor parte de los huesosdel esqueleto se sustenta en unproceso conocido comoformacion intermembranosa de hueso, en el cual seconstruye tejido oseo en el interior de una vaina mem-branosa en ausencia de un molde cartilaginoso.
Cartılago
Las c�elulas del cartılago, denominadas condroblastos ycondrocitos, secretan una matriz extracelular de glu-cosaminoglucano y proteoglucanos que se refuerzacon fibras colagenosas y el�asticas. A lo largo del procesode secrecion, los condroblastos quedan incluidos en lamatriz que fabrican y se convierten en condrocitos queocupan pequenas cavidades denominadas lagunas. Elcartılago carece de irrigacion, por lo que sus nutrientesdifunden a trav�es de la matriz desde los vasos sanguı-neos del tejido conjuntivoque lo rodea, conocido comopericondrio. Gracias a su flexibilidad y resistencia a lacompresion, el cartılago es capaz de:
. Absorber choques.
. Recubrir las superficies de la mayorıa de lasarticulaciones oseas; su superficie lisa elimina lasfuerzas de friccion asociadas a su movimiento.
Se distinguen tres tipos de cartılago en funcion deltipo de fibras presente en sumatriz (fig. 7.1 y tabla 7.1):
. El cartılago el�astico se asemeja al cartılago hialino,si bien contiene fibras el�asticas gruesas en su matrizque le confieren una coloracion amarillenta opaca yun mayor grado de flexibilidad. Existen otras tres
diferencias leves entre ambos tejidos: loscondrocitos del cartılago el�astico son m�as
numerosos y grandes que los delhialino, y la matriz de aquel esmenos abundante que la de este.El cartılago el�astico aparece en lospabellones auditivos, la laringe, laepiglotis y las trompas auditivasexterna e interna.
. El fibrocartılago no se rodea de unpericondrio; presenta una estructurasemejante a la de los tendones, ya queest�a formado por gruesos hacesparalelos de fibras de col�ageno detipo I inmersas en una matriz escasa.La matriz de este tipo de cartılago secompone fundamentalmente dedermatano sulfato y condroitina
sulfato y embebe a los condrocitos pequenosalojados entre dichoshaces. Amenudo, la formacionde fibrocartılago obedece a la aplicacion de fuerzasde tension excesivas en los tendones; los fibroblastosse diferencian en condrocitos que sintetizan matriz,demodo que el tendon se convierte en fibrocartılagopara resistir las potentes fuerzas de tension a lasque est�a sometido. El fibrocartılago aparece en losdiscos articulares, los discos intervertebrales, lasınfisis pubica y en las �areas de anclaje de tendonesy ligamentos al hueso.
. El cartılago hialino se halla en todo el organismo,comoen las articulacionesde las costillas y el esternon,la estructura de las vıas respiratorias en el aparatorespiratorio, y la estructura de la laringe y gran parte dela nariz. Este tejido recubre, adem�as, las superficies delas articulaciones oseas. Las placas epifisarias de losextremos de los huesos largos se componen de estetipo de cartılago. El cartılago hialino que se fabrica a lolargo del desarrollo embrionario constituye laestructura sobre la cual se construir�a el hueso en laformacion endocondral de hueso.
CONSIDERACIONES CL�INICAS
A pesar de que el fibrocartılago es un tejido muyresistente, en ocasiones las fuerzas que soporta lacolumna vertebral pueden ser de tal magnitud queprovocan la herniacion o la rotura de algun discointervertebral. Estas alteraciones suelen ser muydolorosas como consecuencia de la presionejercida por el disco herniado sobre los nerviosraquıdeos. La herniacion y la rotura de los discos
se producen m�as a menudo en las carasposteriores de la region lumbar.
TERMINOS CLAVE. Cartılago hialino. Cartılago el�astico. Matriz osea. C�elulas oseas. Sistemas laminares. Formacion osea. Remodelacion osea. Reparacion osea
74 � 2011 Elsevier Espana, S.L. Reservados todos los derechos
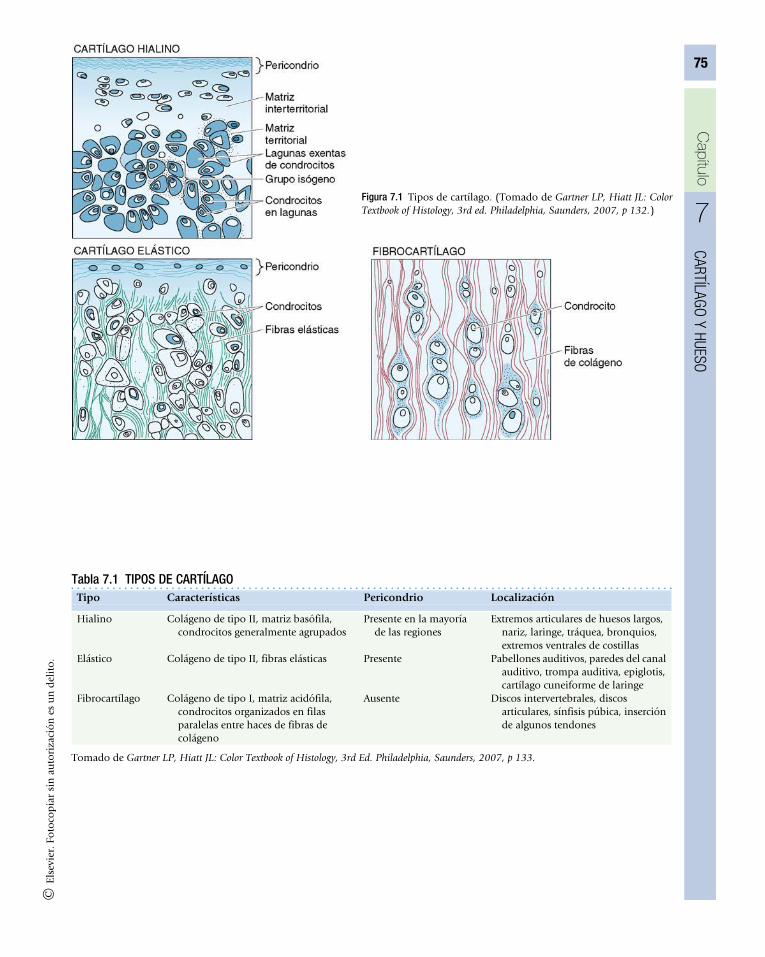
75
7
CARTILA
GOYHUESO
Figura 7.1 Tipos de cartılago. (Tomado de Gartner LP, Hiatt JL: ColorTextbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 132.)
Tabla 7.1 TIPOS DE CARTILAGO
Tipo Caracterısticas Pericondrio Localizacion
Hialino Col�ageno de tipo II, matriz basofila,condrocitos generalmente agrupados
Presente en la mayorıade las regiones
Extremos articulares de huesos largos,nariz, laringe, tr�aquea, bronquios,
extremos ventrales de costillas
El�astico Col�ageno de tipo II, fibras el�asticas Presente Pabellones auditivos, paredes del canal
auditivo, trompa auditiva, epiglotis,cartılago cuneiforme de laringe
Fibrocartılago Col�ageno de tipo I, matriz acidofila,
condrocitos organizados en filas
paralelas entre haces de fibras decol�ageno
Ausente Discos intervertebrales, discos
articulares, sınfisis pubica, insercion
de algunos tendones
Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd Ed. Philadelphia, Saunders, 2007, p 133.
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

76
7
CARTILA
GOYHUESO
HISTOGENIA Y CRECIMIENTO DEL CART�ILAGO HIALINO
Las c�elulas mesenquimatosas cercanas a las �areas enlas que se formar�a cartılago se agrupan en centros decondrificacion, expresan el gen Sox9 y se diferencianen condroblastos. Estas c�elulas reci�en formadas secre-tan la matriz cartilaginosa alrededor de los condro-blastos, cada uno de los cuales queda atrapado en unespacio denominado laguna (fig. 7.2). Estas c�elulas,que aun mantienen la capacidad de division, recibenel nombre de condrocitos.
. Durante el proceso de crecimiento intersticial decartılago, la division del condrocito origina dos ocuatro c�elulas, llamadas grupos isogenos, en elseno de las lagunas. Conforme secretan matriz lasc�elulas que integran cada uno de estos grupos, lalaguna se divide en dos o cuatro compartimentos,los condrocitos se separan unos de otros y el tamanodel cartılago va aumentando.
. Otras c�elulas mesenquimatosas que rodean alcartılago en formacion se diferencian enfibroblastos, los cuales dan lugar a un tejidoconjuntivo vascular denso, el pericondrio, queconsta de dos capas:. La capa fibrosa externa, fabricada con tejidofibroso, alberga a los fibroblastos y posee unairrigacion profusa.
. La capa celular interna se compone de c�elulascondrogenas en mitosis activa que se diferencianen condroblastos.
. Los condroblastos de la capa interna delpericondrio sintetizan una matriz en la periferia delcartılago mediante crecimiento aposicional. Esteproceso representa el principal m�etodo decrecimiento del tejido cartilaginoso, con excepcionde aquellas regiones carentes de este tejido (comolas articulaciones articulares y las placas epifisariasde los huesos largos).
Las c�elulas condrogenasno solamente se diferencianen condroblastos, sino que pueden dar lugar a unosprecursores oseos conocidos como c�elulas osteoprogeni-toras. Varias hormonas y vitaminas influyen en el creci-miento y el desarrollo del cartılago hialino (tabla 7.2).
MATRIZ DEL CARTILAGO HIALINO
El col�ageno de tipo II constituye uno de los compo-nentesm�as importantes de lamatriz (el 40%de su pesoseco, acompanado de algunas fibras de col�ageno detipos IX, X y XI), mientras que la proporcion restantecorresponde a glucoproteınas (condroitina-4-sulfato,condroitina-6-sulfato y heparano sulfato), proteoglu-canos (condronectina) y lıquido extracelular. Se distin-guen dos regiones especializadas en la matriz:
. Matriz territorial, que rodea a cada una de laslagunas.
. Matriz interterritorial, que constituye la matriz quese encuentra entre las otras matrices. En la matrizterritorial apenas aparecen fibras de col�ageno y sonabundantes los condroitina sulfatos, mientras quela interterritorial es rica en fibras de col�ageno ypresenta una cantidad menor de proteoglucanos.
. La c�apsula pericelular, una estrecha banda dematriz territorial, remeda la l�amina basal y seencuentra en contacto directo con los condrocitos,a los que podrıa proteger de danos fısicos.
La matriz del cartılago posee un gran numero deagrecanos, unas mol�eculas de proteoglucanos de ele-vado peso molecular cuyos centros proteicos estable-cen enlaces covalentes con glucosaminoglucanos.Muchas de ellas se unen a mol�eculas de �acido hialu-ronico, lo que da lugar a enormes complejos de agre-cano de carga negativa y una longitud de 3 a 4mmqueatraen iones de Na+ debido a dicha carga. Esta capa deiones de carga positiva atrae a las mol�eculas de agua,que forman una cubierta hidratada sobre los comple-jos de agrecano a la que cartılago hialino debe suelevado contenido hıdrico (aproximadamente, el80%) y su capacidad de resistencia a la compresion.Adem�as de formar enlaces electrost�aticos con lamatriz, las fibras de col�ageno de tipo II incluidasen ella resisten a las fuerzas de tension. La glucopro-teınas de adhesion condronectina posee sitios deunion a las c�elulas del cartılago, las fibras de tipo IIy los elementos que integran los complejos de agre-cano, de modo que favorece la adhesion de los dis-tintos componentes celulares y extracelulares de estetejido. La textura lisa de la superficie del cartılagohialino y su capacidad de resistencia a las fuerzas decompresion y tension lo convierten en un materialideal para recubrir las superficies articulares de loshuesos largos.
CONSIDERACIONES CL�INICAS
Los condrocitos del cartılago hialino que sehipertrofian y mueren crean una matriz calcificadaque provoca la degeneracion del cartılago. Estasecuencia de acontecimientos de degradacion delcartılago hialino sucede normalmente en laformacion endocondral de hueso; sin embargo,estas etapas forman parte del patron normal deenvejecimiento que da lugar a artralgias agudas ycronicas y movilidad reducida. La regeneraciondel cartılago se da principalmente en los ninos,siendo muy limitada en adultos mayores; cuando
est�a activa, las c�elulas condrogenasmigran desdeel pericondrio hacia la lesion. La lesion se rellenade cartılago de nueva formacion cuando susdimensiones no son grandes; de lo contrario, seforma tejido colagenoso denso para ocupar elespacio danado.

77
7
CARTILA
GOYHUESO
Tabla 7.2 EFECTOS DE HORMONAS Y VITAMINAS SOBRE EL CARTILAGO HIALINO
Sustancia Efectos
Hormonas
Tiroxina, testosterona y somatotropina
(a trav�es de factores de crecimiento insulinoides)
Estimulan el crecimiento del cartılago
y la formacion de la matriz
Cortisona, hidrocortisona, y estradiol Inhiben el crecimiento del cartılago
y la formacion de la matrizVitaminas
Hipovitaminosis A Reduce la anchura de las placas epifisarias
Hipervitaminosis A Acelera la osificacion de las placas epifisarias
Hipovitaminosis C Inhibe la sıntesis de la matriz y deforma la estructura de lasplacas epifisarias; produce escorbuto
Ausencia de vitamina D, que origina hipoabsorcion
de calcio y fosforo
Proliferacion normal de condrocitos, pero la calcificacion
de la matriz es deficiente; origina raquitismo
Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 135.
Figura 7.2 Diagrama de lamatriz extracelular. Arriba, imagen de bajo aumento en la que se observa el bandeado de las fibras de col�ageno
con proteoglucanos adheridos. Abajo, glucosaminoglucanos unidos al nucleo proteico y proteınas de union que los anclan al �acidohialuronico para crear unas enormesmacromol�eculas con un pesomolecular de cientos demillones de daltons. (Tomado deGartner LP,
Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 72.)
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

Hueso
El hueso, el tercer tejido m�as duro, est�a sometido auna continua remodelacionmediante resorcion comorespuesta a la presion y osteogenia como respuesta a latension.
. El esqueleto oseo no se limita a sostener alorganismo, sino que tambi�en configura unaarmadura defensiva que protege a los organosvitales, como el cerebro y la m�edula espinal.
. Los musculos esquel�eticos se unen a los huesos atrav�es de las articulaciones y hacen posible elmovimiento relativo de las distintas partes delorganismo y el desplazamiento del mismo.
. Pr�acticamente la totalidad del calcio se almacena enel esqueleto oseo, el cual puede liberarse delesqueleto para mantener unas concentracioneshem�aticas equilibradas.
. Asimismo, en la cavidadmedulardel hueso se aloja lam�edula osea, la cual se ocupa de la hematopoyesis.
La superficie externa del hueso se recubre de tejidoconjuntivo blando, el periostio, el cual consta de doscapas:
. Una capa fibrosa externa formada por tejidoconjuntivo colagenoso denso irregular.
. Una capa celular interna con capacidadosteog�enica, que alberga c�elulas osteoprogenitoras(c�elulas osteog�enicas), osteoblastos y, a veces,osteoclastos.
Las cavidades medulares est�an tapizadas de endos-tio, una delgada capa celular formada por c�elulasosteoprogenitoras, osteoblastos, osteoclastos en oca-siones, y elementos aislados de tejido conjuntivo.
La matriz osea se compone de elementos org�a-nicos e inorg�anicos:
. El calcio y el fosforo representan la mayorıa de loscomponentes inorg�anicos (alrededor del 65% delpeso seco). Casi todo el calcio y el fosforo aparecenen forma de cristales de hidroxiapatita[Ca10(PO4)6(OH)2] que se distribuyen en losespacios intersticiales que separan las fibras decol�ageno de tipo I y se alinean a lo largo de lasmismas. Los cristales atraen agua, que crea unacubierta de hidratacion que facilita el intercambioionico con el lıquido extracelular.
. El col�ageno de tipo I, el componente principal de lafraccion org�anica, constituye entre el 80 y el 90% dela misma. La mayor parte de los restantescomponentes org�anicos se hallan en forma decomplejos de agrecano; aparecen, asimismo,osteocalcina, osteopontina, sialoproteınas oseasy glucoproteınas adhesivas. Estas ultimas
favorecen la adhesion de las proteınas de la matrizosea a los cristales de hidroxiapatita y las integrinaspresentes en las membranas plasm�aticas de lasc�elulas oseas.
CELULAS DEL HUESO
El hueso se compone de cuatro clases de c�elulas, lastres primeras (c�elulas osteoprogenitoras, osteoblastosy osteocitos) pertenecen al mismo linaje celular,mientras que la cuarta (osteoclastos) proviene de pre-cursores monocıticos (fig. 7.3).
. Las c�elulas osteoprogenitoras se encuentran en lacapa celular interna del periostio; revisten loscanales de Havers y las cavidades medulares.Estas c�elulas proliferan y expanden la poblacionde c�elulas osteoprogenitoras y osteoblastos cuandola tension de oxıgeno es elevada o bien c�elulascondrogenas cuando es baja.
. La formacion de osteoblastos depende de lapresencia de proteınas morfogenas de hueso yfactor transformador de crecimiento b. Lososteoblastos forman una capa de c�elulas quesecretan los elementos org�anicos de la matriz osea,ası como las mol�eculas de senalizacion, losreceptores para la activacion del ligando del factornuclear kappa b (RANKL) y el factor estimulador decolonias de macrofagos (M-CSF). Conformefabrican la matriz, los osteoblastos emiten delgadasproyecciones que entran en contacto con lasproyecciones de otros osteoblastos cercanos y creanuniones gap con estos. Los osteoblastos quedanatrapados en la matriz que se acumula alrededor deellos; se denominan osteocitos y el espacio queocupan en dichamatriz recibe el nombre de laguna.Adem�as de sintetizar matriz osea y diferenciarse enosteocitos, los osteoblastos intervienen en lacalcificacion de la matriz. Al terminar la formaciondel hueso, su superficie externa conserva una capade osteoblastos inactivos que han dejado desintetizar matriz y presentan una morfologıaaplanada; estas c�elulas se llaman c�elulas derevestimiento del hueso. Una delgada capa dematriz no calcificada separa a estas c�elulas de lososteocitos, llamada osteoide, separa a estas c�elulasy a los osteocitos de la matriz calcificada. De sernecesario, las c�elulas que tapizan el hueso puedenactivarse de nuevo para sintetizar matriz osea. Lasmembranas plasm�aticas de los osteoblastos est�andotadas de integrinas y receptores de la hormonaparatiroidea (PTH); la primera facilita la adhesionde los osteoblastos a los componentes de la matrizosea, mientras que la ultima desencadena lasecrecion de RANKL y factor estimulador deosteoclastos por parte de los osteoblastos trasunirse a dicha hormona. La proteına RANKL
78
7
CARTILA
GOYHUESO

favorece la diferenciacion de los preosteoclastosen osteoclastos, mientras que el factorestimulador de osteoclastos induce la actividadde resorcion osea en esta poblacion celular.No obstante, los osteoblastos han de eliminarpreviamente el osteoide de la superficie del huesopara permitir el acceso de los osteoclastos alhueso calcificado.
. Los osteocitos se encuentran en las lagunas delhueso y sus delgadas proyecciones osteocıticas se
extienden a trav�es de los canalıculos, unos delgadoscanales presentes en la matriz calcificada, y entranen contacto con osteocitos adyacentes; formanuniones gap con las proyecciones de otrososteocitos y osteoblastos con el fin de facilitar lacomunicacion de las c�elulas que integran el tejidooseo. El lıquido extracelular rellena los espaciosperiosteocıticos y los canalıculos y aportanutrientes y mol�eculas de senalizacion a las c�elulasdel hueso, adem�as de eliminar productos de
79
7
CARTILA
GOYHUESO
Figura 7.3 Origen de las c�elulas del tejido conjuntivo (v. osteoblastos, osteocitos y osteoclastos). (Tomado deGartner LP, Hiatt JL: ColorTextbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 112.)
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

desecho y mol�eculas de senalizacion secretadas porestas c�elulas. Estos espacios contienen m�as de 1 l delıquido extracelular al que los osteocitos puedensecretar hasta 20 g de calcio en un breve espacio detiempo.
. Los osteoclastos, procedentes del sistemafagocıtico mononuclear, son c�elulas de grantamano (di�ametro, 150mm) multinucleadas(� 50 nucleos) encargadas de la resorcion oseay que presentan numerosos receptores demembrana, como el receptor del factorestimulador de osteoclastos 1, el receptor decalcitonina y RANK. Los osteoblastos estimuladospor la PHT activan la resorcion osea por lososteoclastos, que es inhibida por la union de lacalcitonina a receptores de calcitonina presentesen su membrana plasm�atica. El M-CSF, secretadopor los osteoblastos, se une a receptores de M-CSFpresentes en las c�elulas precursoras de osteoclastos,como consecuencia de lo cual estimulan suproliferacion y la expresion de receptores de RANKen susmembranas celulares. Demanera simult�anea,los osteoblastos expresan receptores para RANK,RANKL, a trav�es de los cuales se unen losprecursores de los osteoclastos a los osteoblastos.. La interaccion de RANK con RANKL induce latrimerizacion de aquella en la superficie de lasc�elulas precursoras de osteoclastos, lo que activa alas mol�eculas adaptadoras para poner en marchala transcripcion.
. Los factores nucleares sintetizados convierten alprecursor mononucleado de osteoclastos en unosteoclasto multinucleado inactivo que se separadel osteoblasto.
. Asimismo, los osteoblastos fabricanosteoprotegerina (OPG), un ligandocaracterizado por una gran afinidad a RANKL,que impide su union a RANK y la asociacion delprecursor de osteoclastos con un osteoblasto,lo que evita la formacion de osteoclastos.
. En presencia de PTH, los osteoblastos producenun numeromayor demol�eculas de RANKL que deOPG, de tal modo que favorecen laosteoclastogenia (desarrollo de osteoclastos).
. Los osteoclastos inactivos expresan integrinas avb3,a trav�es de las cuales pueden adherirse a la superficiedel hueso.
. Tras la eliminacion del osteoide de la superficieosea por parte de los osteoblastos, estas c�elulasmigran y son sustituidas por osteoclastosinactivos que se convierten en c�elulas activas alunirse a dicha superficie. Los osteoclastos activosse alojan en unas depresiones someras localizadasen la superficie osea, las lagunas de Howship.
CONSIDERACIONES CL�INICAS
La fosfatasa alcalina es una enzima muyabundante en las membranas celulares de lososteoblastos. Cuando se secreta hueso, estas
c�elulas secretan concentraciones elevadas defosfatasa alcalina que provocan un aumento de
sus concentraciones hem�aticas. La valoracion de
las concentraciones hem�aticas de esta enzimapermite controlar la formacion de hueso.
Los osteoclastos activos en la resorcion osea pre-sentan cuatro regiones reconocibles (fig. 7.4):
. La zona basal contiene la mayorıa de los org�anulosdel osteoclasto, con excepcion de las mitocondrias,que se tienden a concentrarse en el borde en cepillo.
. El borde en cepillo se encuentra en la zona decontacto del osteoclasto con el hueso en la que tienelugar la resorcion osea. El osteoclasto presenta unasproyecciones citopl�asmicas digitiformes dotadas deunamembrana plasm�atica engrosada que confierenproteccion a la c�elula en el transcurso de la resorcional delimitar un compartimento subosteocl�astico.
. La zona clara,un �area exenta de org�anulos situada enla periferia del borde en cepillo, expresa integrinasavb3. El dominio extracelular de estas proteınas seasocia a la osteopontina de la superficie osea paracrear una zona de sellado que aısla el microentornodel compartimento subosteocl�astico. La porcionintracelular de las integrinas se une a filamentos deactina para dar lugar a un anillo de actina.
. La zona vesicular, localizada entre la zona basal y elborde en cepillo, presenta un gran numero devesıculas de exocitosis y endocitosis. En las primerasse transporta la catepsina K, la cual degrada fibrasde col�ageno y otras proteınas de la matriz osea,hacia el compartimento subosteocl�astico, mientrasque estas ultimas translocan los residuos generadospor la degradacion osea al interior del osteoclasto.
MECANISMO DE RESORCION OSEA
El componente inorg�anico se desprende de la matrizosea debido al pH �acido del entorno y las sales mine-rales disueltas pasan al citoplasma del osteoclasto, enel que se transportan en vesıculas de exocitosis hacialos capilares locales cercanos a la zona basal. Lososteoclastos secretan catepsina K en el compartimentosubosteocl�astico para degradar los componentesorg�anicos de dicha matriz. Los materiales degradadosparcialmente son captados en vesıculas de endocitosispor los osteoclastos, que los someten a nuevas reac-ciones de degradacion antes de liberarse en la regionbasal (v. fig. 7.4).
80
7
CARTILA
GOYHUESO

CONSIDERACIONES CL�INICAS
La osteopetrosis se debe a una anomalıa
gen�etica caracterizada por el desarrollo deosteoclastos incapaces de llevar a cabo laresorcion osea al no poder crear un borde encepillo. Los pacientes con osteopetrosis poseenhuesos muy densos y, con frecuencia, anemiacomo consecuencia de la reduccion del volumende la cavidad medular. De igual modo, puedenmanifestar ceguera, hipoacusia y anomalıas delos nervios craneales debido al estrechamiento
de los agujeros a trav�es de los cuales salen
del cr�aneo.
81
7
CARTILA
GOYHUESOFigura 7.4 Funcion del osteoclasto. RER, retıculo endopl�asmico rugoso. (Tomado de Gartner LP, Hiatt JL, Strum JM: Cell Biology and
Histology [Board Review Series]. Philadelphia, Lippincott Williams & Wilkins, 1998, p. 100.)
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

ASPECTO MACROSCOPICO DEL HUESO
El hueso puede clasificarse con arreglo a sumorfologıaexterna:
. Huesos largos, formados por un eje delgado, ladi�afisis, y dos cabezas, las epıfisis
. Huesos cortos, de anchura similar a su longitud
. Huesos planos, compuestos por dos placas planasde hueso compacto entre las que se encuentra unacapa de hueso esponjoso
. Huesos irregulares, sin una morfologıa definida
. Huesos sesamoideos, formados en el interior de lostendones
Segun su densidad, el hueso puede ser denso, comoes el caso del hueso compacto, o espongiforme,como el hueso esponjoso. Este ultimo se rodeasiempre de hueso compacto. La cavidad medular delos huesos largos, tapizados por una capa delgada dehueso esponjoso, alberga la m�edula osea roja en losindividuos jovenes, que se convierte en la m�edulaosea amarilla en el adulto como consecuencia de laacumulacion de lıpidos en esta cavidad con el paso deltiempo. La m�edula osea fabrica las c�elulas sanguıneas,mientras que la amarilla no est�a implicada en la hema-topoyesis, aunque conserva esta capacidad. El huesoesponjoso posee espacios medulares tapizados porosteoblastos en reposo; el tejido oseo que se formaen el perımetro de estos espacios consta de unasl�aminas oseas irregulares m�as pequenas y alargadas,las espıculas y la trab�eculas.
Los extremos articulares de las epıfisis se compo-nen de una delgada capa de hueso compacto querecubre al hueso esponjoso y sobre la que se disponeuna capa de cartılago hialino. En los sujetos en creci-miento, la epıfisis y la di�afisis se separan por una placaepifisaria de cartılago hialino. Lamet�afisis es un �areaen abanico del eje que se dispone entre la di�afisis ydicha placa.
La superficie externa de la di�afisis y las superficiesque no se articulan de las epıfisis se recubren de unperiostio formado por dos capas que se inserta en elhueso por medio de fibras de col�ageno, las fibras deSharpey (fig. 7.5).
. La capa fibrosa externa del periostio est�a formadapor tejido conjuntivo fibroso irregular denso cuyoselementos vasculares irrigan la region externa delhueso compacto.
. La capa celular interna posee c�elulasosteoprogenitoras y osteoblastos.
Los huesos de la boveda craneal (cubierta delcr�aneo) constan de las tablas externa e interna dehueso compacto que rodean una masa de hueso
esponjoso, llamada dıploe, situada entre ellas. El peri-ostio que recubre la tabla externa recibe el nombre depericr�aneo, mientras que el que reviste la tablainterna de la boveda craneal se conoce como dura-madre, la capa m�as externa de las meninges que recu-bre y protege al cerebro. Adem�as, esta capa actua comoperiostio de la tabla interna.
TIPOS DE HUESO SEGUN LA MORFOLOGIA
MICROSCOPICA
La morfologıa microscopica del hueso permite distin-guir dos tipos: hueso primario y hueso secundario.
. El hueso primario (hueso inmaduro oentrelazado) es el primero que se forma a lo largodel desarrollo y durante el proceso de reparacionosea. Se distingue por la predominancia delcomponente celular, el menor grado decalcificacion y la distribucion aleatoria de las fibrasde col�ageno. Es sustituido por hueso secundario,con excepcion de los alv�eolos dentales y lasinserciones tendinosas.
. El hueso secundario (huesomaduro o laminar) esun tejido muy organizado en l�aminas oseasconc�entricas (grosor, 3 a 7mm); su mayor grado decalcificacion y la disposicion ordenada de las fibrasde col�ageno le confieren una dureza mayor que ladel hueso primario. Los osteocitos alojados en laslagunas se distribuyen a intervalos regulares entrelas laminillas o, de manera menos frecuente, en elinterior de las mismas (v. fig. 7.5). Estas c�elulas secomunican a trav�es de sus proyeccionesosteocıticas que forman uniones gap entre sı enunos angostos canales denominados canalıculos.
Sistemas laminares del hueso compacto
El hueso compacto se compone de unas capas muydelgadas de hueso conocidas como l�aminas que sedisponen en cuatro sistemas laminares –l�aminas cir-culares externas, l�aminas circulares internas, l�aminasintersticiales y osteonas (sistemas de canales deHavers)– que pueden distinguirse f�acilmente en loshuesos largos (v. fig. 7.5).
. La capa calcificadam�as externa de la di�afisis, situadainmediatamente por debajo del periostio, es elsistema laminar circular externo, en el que seinsertan las fibras de Sharpey.
. Las l�aminas oseas que rodean a la cavidadmedular conforman el sistema laminar circularinterno. El hueso esponjoso que reviste estesistema proyecta trab�eculas y espıculas hacia dichacavidad.
82
7
CARTILA
GOYHUESO

83
7
CARTILA
GOYHUESO
Figura 7.5 Diagrama de la estructura del hueso en el que aparece hueso cortical compacto, osteonas, l�aminas, canales de Volkmann,
canales de Havers, lagunas, canalıculos y hueso esponjoso. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.Philadelphia, Saunders, 2007, p 144.)
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

. Los sistemas de canales de Havers (osteonas),con un di�ametro comprendido entre 20 y 100mm,representan el principal sistema laminar del huesocompacto. Est�an formados por l�aminas similares aobleas de hueso calcificado que originan cilindrosconc�entricos con un canal de Havers central querecibe inervacion e irrigacion y est�a tapizado porc�elulas osteoprogenitoras y osteoblastos (fig. 7.6).Las osteonas reflejan las ramificaciones ybifurcaciones de la irrigacion vascular. Est�anlimitados por una lınea de cementacion, formadapor una sustancia fundamental calcificada quecontiene escasas fibras de col�ageno.. Las fibras de col�ageno presentan una estructurahelicoidal muy organizada; en un cortetransversal, las fibras son paralelas entre sı dentrode cada l�amina, pero est�an orientadas enperpendicular a las de las l�aminas adyacentes. Lavariacion de la inclinacion de la h�elice da lugar aeste patron, que reduce la probabilidad defractura osea.
. Los canales de Havers est�an conectados a los de laosteona adyacente pormedio de canales oblicuos,los canales de Volkmann, que hacen posible lairrigacion de otros canales de Havers (v. fig. 7.6).
. El proceso de formacion de las osteonas es elsiguiente: en primer lugar se desarrolla la l�aminam�as externa, m�as cercana a la lınea decementacion; las l�aminas siguientes se adosan a lasya formadas; la l�amina m�as interna, en contactocon el canal de Havers, es la ultima en sintetizarse.Dado que el mantenimiento de los osteocitosdepende de los ineficientes canalıculos, el grosorde cada osteona no suele superar las 20 l�aminas.
. El hueso est�a sometido a una remodelacioncontinua a medida que los osteoclastos reabsorbenosteonas y son sustituidos por osteoblastos. Losrestos de las osteonas que han sufrido este procesopermanecen en forma de fragmentos laminares conaspecto de arco, las l�aminas intersticiales, rodeadosde osteonas no reabsorbidas.
HISTOGENIA DEL HUESO
El hueso se desarrolla a lo largo de la etapa embrionariamediante formacion intermembranosa de hueso obien formacion endocondral de hueso. A pesar deque ambos procesos difieren notablemente, histologica-mente sus productos finales son id�enticos. De maneraindependiente de la vıa de formacion, el primer huesoque se forma es el primario; posteriormente se reab-sorbe y sustituye por hueso secundario, un tejido
maduro sometido continuamente a reabsorcion yremodelacion como respuesta a las fuerzas ambientalesa las que se enfrentar�a a lo largo de la vida (fig. 7.7).
. La mayorıa de los huesos planos se desarrollamediante el proceso de formacionintramembranosa de hueso.. El proceso comienza en un entorno muyvascularizado de tejido mesenquimatoso en elque las c�elulas mesenquimatosas est�an encontacto entre sı.
. Estas c�elulas expresan los reguladores maestrososteog�enicos, factores de transcripcion Cbfa1/Runx2 y el factor de transcripcion con dedo decinc osterix, y se diferencian en osteoblastos, lasc�elulas encargadas de secretar la matriz osea.
. En ausencia del factor osterix, las c�elulasmesenquimatosas se convierten enpreosteoblastos que no pueden diferenciarse enosteoblastos funcionales capaces de secretar lamatriz.
. La osteogenia se pone en marcha a medida que lamatriz inicial establece complejos trabeculares encuyas superficies se alojan los osteoblastos. Esta�area representa un centro de osificacion primariaen el que se forma hueso primario.
. La calcificacion se inicia tras la secrecion delosteoide, de modo que los osteoblastos quedanatrapados en las lagunas. Estas y la matriz que lasrodea reciben el nombre de osteocitos. La matrizse calcifica y se crean canalıculos alrededor de lasproyecciones de los osteocitos.
. Las trab�eculas crecen y proliferan para formarredes alrededor de los elementos vasculares, quese convierten en m�edula osea.
. Los huesos planos de mayor tamano, como losque integran el cr�aneo, precisan de otros centrosde osificacion. Estos centros se fusionan paraoriginar un unico hueso conforme avanza laformacion del hueso, salvo en el caso de lasfontanelas del cr�aneo neonatal, en el que loscentros de osificacion de los huesos frontal yparietal no se fusionan hasta despu�es delnacimiento con la sustitucion de los puntos blandospor hueso.
. Las regiones del tejido conjuntivomesenquimatoso que no intervienen en laosteogenia originan el periostio y el endostio.
Formacion endocondral de hueso
La mayorıa de los huesos del organismo, excep-tuando a los huesos planos, se desarrollan a trav�es
84
7
CARTILA
GOYHUESO

85
7
CARTILA
GOYHUESO
Figura 7.6 Diagrama de la estructura del hueso en el que
aparece hueso cortical compacto, osteonas, l�aminas,
canales de Volkmann, canales de Havers, lagunas, cana-
lıculos y hueso esponjoso. (Tomado de Gartner LP,Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia,
Saunders, 2007, p 144.)
Figura 7.7 Formacion intramembranosa dehueso. (Tomado de Gartner LP, Hiatt JL: Color
Textbook of Histology, 3rd ed. Philadelphia, Saun-
ders, 2007, p 146.)
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

de la vıa de formacion endocondral de hueso, unproceso dividido en varias etapas que se representagr�aficamente en la figura 7.8 y se sintetiza en latabla 7.3.
. Un molde de cartılago hialino actua comoesqueleto para la formacion del hueso.
. A medida que el hueso en formacion vaadquiriendo estabilidad para sostener al organismo,el molde cartilaginoso se reabsorbe y sustituye portejido oseo.
. La primera zona del cartılago que se sustituyecorresponde a la di�afisis, el centro primario deosificacion, y posteriormente se forma hueso en lasepıfisis, los centros secundarios de osificacion.
El proceso de formacion endocondral de huesoconsta de una secuencia din�amica de acontecimientosrelacionados que se pone en marcha durante el desa-rrollo fetal y continua hasta la vida adulta, en la que seprolonga en funcion de las necesidades de reparacionosea. Estos mecanismos actuan en el adulto, ya que el
86
7
CARTILA
GOYHUESO
Tabla 7.3 ACONTECIMIENTOS EN LA FORMACION ENDOCONDRAL DE HUESO
Acontecimiento Descripcion
Formacion del molde de cartılago hialino Se forma un molde de cartılago hialino en miniatura en lasregiones del embrion en las que se desarrollar�a el hueso.
Algunos condrocitos maduran, se hipertrofian y mueren.
La matriz de cartılago se calcifica
Centro primario de osificacion
Se vasculariza el pericondrio de la membrana media de la
di�afisis
La vascularizacion del pericondrio lo transforma en periostio.
Las c�elulas condrogenas se convierten en c�elulas
osteoprogenitoras
Los osteoblastos secretan matriz y forman un collar oseoperiostico
Se forma un collar oseo periostico de hueso primario(formacion intramembranosa de hueso)
Los condrocitos del nucleo de la di�afisis se hipertrofian,
mueren y degeneran
La presencia de periostio y hueso impide la difusion de
nutrientes a los condrocitos. Su muerte crea lagunas queabren unos espacios amplios en los tabiques del cartılago
Los osteoclastos taladran el collar oseo periostico y crean
orificios a trav�es de los cuales entrar�a la yema osteog�enica
Los orificios hacen posible la invasion del molde de cartılago, ya
calcificado, por parte de capilares y c�elulas osteoprogenitoras,
que comienzan a sintetizar la matriz oseaFormacion del complejo cartılago calcificado/hueso calcificado La matriz osea depositada en los tabiques del cartılago
calcificado da lugar a este complejo. Desde el punto de vista
histologico, el cartılago calcificado se tine de color azul,
mientras que el hueso calcificado lo hace de rojoLos osteoclastos reabsorben el complejo cartılago calcificado/
hueso calcificado
La cavidad medular se amplıa como consecuencia de la
destruccion del complejo cartılago calcificado/hueso
calcificadoEl collar oseo subperiostico se engrosa y comienza a crecer hacia
las epıfisis
Con el paso del tiempo, este proceso termina por sustituir el
cartılago diafisario por hueso
Centro secundario de osificacion
La osificacion comienza en las epıfisis El inicio es id�entico al del centro primario, si bien no existecollar oseo. Los osteoblastos depositan matriz en el esqueleto
de cartılago calcificado
Crecimiento de hueso en la placa epifisaria Se conserva la superficie articular cartilaginosa del hueso. La
placa epifisaria se mantiene (el crecimiento tiene lugar en suextremo epifisario). Se anade tejido oseo en el extremo
diafisario de la misma
La epıfisis y la di�afisis se conectan Al concluir el crecimiento del hueso, se interrumpe la
proliferacion del cartılago de la placa epifisaria. El desarrollooseo prosigue hasta conectar la di�afisis con la epıfisis

87
7
CARTILA
GOYHUESO
Figura 7.8 Formacion de hueso endocondral. El color azul representa el molde de cartılago sobre el que crece el hueso, por el cual es
sustituido. A. Molde de cartılago hialino. B. El cartılago del eje (di�afisis) es invadido por elementos vasculares. C. Se forma un collar
subperiostico de hueso.D. El collar oseo impide la difusion de nutrientes hasta las c�elulas del cartılago, que mueren y forman lagunascomunicadas. E.Complejo de hueso calcificado/cartılago calcificado en los extremos epifisarios del hueso en formacion. F. Elongacion
de la placa epifisaria en el extremo en el que el hueso sustituye al cartılago. (Tomado deGartner LP, Hiatt JL: Color Textbook of Histology,
3rd ed. Philadelphia, Saunders, 2007, p 147.)
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

hueso es un tejido din�amico sometido a una constanteremodelacion para adaptarse a las presiones ambien-tales.
Crecimiento longitudinal del hueso
El crecimiento longitudinal del hueso se debe a laproliferacion de los condrocitos de la placa epifisaria.La superficie epifisaria de la placa est�a formada porcartılago, mientras que la cara diafisaria se componede hueso. La placa epifisaria se divide en cincoregiones que comienzan en el lado epifisario (fig. 7.9):
. Zona de reserva del cartılago. Los condrocitos enmitosis activa se distribuyen de forma aleatoria.
. Zona de proliferacion. Los condrocitos secretan laproteına Indian hedgehog, que impide lahipertrofia de estas c�elulas e induce la secrecion dela proteına relacionada con la PTH (PR-PTH), lacual favorece la division celular en los condrocitosde la zona de proliferacion. Estos condrocitos enmitosis activa forman lıneas paralelas a la direccionde elongacion del hueso.
. Zona de maduracion e hipertrofia. Loscondrocitos en fase de maduracion acumulanglucogeno y expresan los factores de trascripcionCbfa1/Runx2, que les permiten hipertrofiarse.Asimismo, estos condrocitos liberan fibras decol�ageno de tipo X y el factor de crecimiento delendotelio vascular, el cual favorece lavascularizacion.
. Zona de calcificacion. Las c�elulas hipertrofiadasatraen macrofagos, los cuales destruyen lostabiques calcificados de las lagunas hipertrofiadasadyacentes; los condrocitos sufren procesos deapoptosis y mueren.
. Zona de osificacion. Las c�elulas osteoprogenitoraspasan a la zona de osificacion y se diferencian enosteoblastos, los cuales sintetizan matriz osea queposteriormente se calcificar�a sobre la superficie delcartılago calcificado. El complejo cartılagocalcificado/hueso calcificado se reabsorbe ysustituye por hueso.
El crecimiento longitudinal del hueso continuamientras las zonas de proliferacion y la tasa de reab-sorcion en la zona de osificacion se encuentren equi-libradas. Hacia los 20 anos de edad, la tasa de reabsor-cion en la zona de osificacion supera a la tasa demitosis en la zona de proliferacion, de modo que seagota la reserva de cartılago. Al concluir la reabsorciondel ultimo complejo cartılago calcificado/hueso calci-ficado, la placa epifisaria no separa la epıfisis de ladi�afisis, las cavidades medulares de ambas regiones
se conectan y se interrumpe el crecimiento longitudi-nal del hueso.
Crecimiento transversal del hueso
El crecimiento longitudinal del hueso se lleva a cabopor medio de la proliferacion intersticial del cartılagode la placa epifisaria, mientras que el crecimientotransversal se sustenta en el crecimiento aposicionalpor debajo del periostio. Los osteoblastos procedentesde c�elulas osteoprogenitoras del periostio secretanmatriz osea en la superficie del hueso, un procesoque recibe el nombre de formacion intramembra-nosa subperiostica de hueso y continua a lo largodel desarrollo y el crecimiento del hueso.
Los procesos de resorcion y deposito del huesodeben mantenerse equilibrados a lo largo de la vida.La formacion de hueso en la superficie externa de ladi�afisis debe compensarse con la actividad de resor-cion ejercida por los osteoclastos en la cara internapara elongar la cavidad medular.
CALCIFICACION DEL HUESO
El proceso de calcificacion no se conoce por completo,si bien se ha determinado que los proteoglucanos, laosteonectina y la sialoproteına osea estimulan lacalcificacion. La nocion prevalente en la actualidadsobre la calcificacion implica la liberacion de vesıcu-las de matriz envueltas en una membrana (con undi�ametro comprendido entre 100 y 200nm) por partede los osteoblastos.
. Las vesıculas de matriz contienen concentracionesaltas de iones Ca++ y PO4
3-, monofosfato cıclico deadenosina, ATPasa, pirofosfatasa, proteınas deunion al calcio y fosfoserina.
. En las membranas de estas vesıculas aparecenbombas de calcio que transportan estos iones haciasu luz; el aumento de las concentraciones de Ca++
induce la formacion de cristales de hidroxiapatitac�alcica, que crecen, perforan la membrana de lavesıcula y provocan la liberacion de sus contenidos.
. Los cristales libres de hidroxiapatita c�alcica actuancomo focos de cristalizacion en el seno de la matriz.
. Las enzimas de estas vesıculas liberan iones fosfatoque se combinan con iones de calcio para calcificarla matriz que rodea a los focos de cristalizacion.
. Se reabsorbe el agua de la matriz y se depositancristales de hidroxiapatita en el interior de lasregiones intersticiales de las fibras de col�ageno.
. Los distintos focos de cristalizacion crecen hastafusionarse entre sı y calcificar la matriz en sutotalidad.
88
7
CARTILA
GOYHUESO

CONSIDERACIONES CL�INICAS
Los ninos con carencia de somatropina padecenenanismo, mientras que los que presentanconcentraciones excesivamente altas de estahormona durante el perıodo de crecimientopadecen gigantismo hipofisario. Los adultosque sintetizan cantidades demasiado altas desomatotropina se caracterizan por el aumento deldeposito oseo en ausencia de una resorcionnormal. Este trastorno, conocido comoacromegalia, provoca el engrosamiento de loshuesos, en especial de los que conforman elrostro, lo que altera la morfologıa normal de laspartes blandas que los recubren.
89
7
CARTILA
GOYHUESO
Figura 7.9 Zonas de la placa epifisaria.
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

REMODELACION OSEA
En el adulto, el tejido oseo est�a sometido a una con-tinua remodelacion y los sistemas de Havers se susti-tuyen constantemente con el fin de adaptarse a lascambiantes fuerzas ambientales a las que est�a some-tido conforme se reabsorben �areas del hueso en unaregion y se anade tejido oseo en otras.
. La morfologıa general del hueso continuareflejando la de los moldes de cartılagoembrionario a pesar de presentar un tamanovarias veces mayor que estos. Esta caracterıstica sedebe a la capacidad de remodelacion superficial,ya que el deposito y la resorcion actuan de maneracoordinada en las superficies del periostio y elendostio.
. Las c�elulas del hueso compacto responden a losfactores sist�emicos calcitonina y PTH,mientras quelas del hueso esponjoso lo hacen a factoressintetizados por la m�edula osea, como el factorestimulador de colonias 1, el factor de necrosistumoral, la interleucina 1, la osteoprotegerina(OPG, una mol�ecula homologa de RANK), elligando de osteoprotegerina (OPGL, unamol�ecula homologa de RANKL) y el factortransformador del crecimiento b.
. La sustitucion de los sistemas de Havers provoca lamuerte de los osteocitos y los osteoclastos se ocupande la reabsorcion de una fraccion de los antiguossistemas, lo que da lugar a la aparicion de cavidadesde resorcion.
. El tamano de estas cavidades aumenta amedida quecontinua la actividad de los osteoclastos y soninvadidas por vasos sanguıneos.
. La sıntesis de l�aminas sucesivas que rodean los vasossanguıneos y crean un nuevo sistema de Haverssupone el comienzo de la formacion de hueso. Lasecuencia de resorcion seguida de sustitucion recibeel nombre de acoplamiento; los residuos de lasosteonas reabsorbidas constituyen el sistemalaminar intersticial.
REPARACION OSEA
Los danos graves ocasionados al hueso pueden pro-vocar el desplazamiento de los extremos del huesofracturado o la separacion de fragmentos oseos delhueso danado o ambos. Adem�as, la seccion de losvasos sanguıneos en la zona de fractura da lugar auna hemorragia local con formacion de un co�aguloque rellena el �area danada (fig. 7.10).
. La irrigacion de la zona se interrumpe de maneraretrograda hasta un punto de anastomosis que
permita el establecimiento de una nueva vıa deirrigacion.
. La irrigacion de muchos sistemas de Haversdesaparece, lo que origina la muerte de lososteocitos, que dejan lagunas vacıas y amplıan el�area danada.
. La desaparicion de la irrigacion afecta en menormedida al periostio y la m�edula osea, ya quedisfrutan de una profusa vascularizacionprocedente de distintas �areas.
. A lo largo de los 2 dıas siguientes a la lesion,pequenos capilares y fibroblastos invaden elco�agulo que ocupa la zona danada y se forma tejidode granulacion.
. La capa osteog�enica del periostio, el endostio, y lasc�elulas indiferenciadas de la m�edula osea proliferany dan lugar a c�elulas osteoprogenitoras, las cuales sediferencian en osteoblastos.
. Los nuevos osteoblastos secretan matriz osea, quecementa el hueso muerto en el �area danada paracrear hueso sano, un proceso que comienza con laformacion de un collar oseo –el callo externo–.
. De manera simult�anea, c�elulas pluripotencialesprocedentes de la m�edula osea y c�elulasosteoprogenitoras derivadas del endostio invaden elco�agulo de la cavidad medular para crear el callointerno, formado por trab�eculas oseas, a lo largo dela semana siguiente a la lesion.
La tasa de proliferacion de las c�elulas osteoproge-nitoras en el callo externo es mayor que la de vascula-rizacion, por lo que algunas de las c�elulas osteopro-genitoras alejadas de los lechos capilares est�anexpuestas a una tension de oxıgeno m�as baja. Estasc�elulas se diferencian en c�elulas condrogenas quedan lugar a condroblastos y secretan matriz de cartı-lago en la superficie del collar oseo. Las c�elulas osteo-progenitoras irrigadas por capilares continuan proli-ferando para ampliar su poblacion. El callo externoconsta de tres capas:
. Capa de collar oseo cementada al hueso
. Capa intermedia de cartılago
. Capa superficial con c�elulas osteog�enicas
La matriz de cartılago adyacente al hueso del collarse calcifica y se sustituye por hueso primariomedianteformacion endocondral de hueso. En ultima instan-cia, los fragmentos oseos terminan por unirse a trav�esde hueso esponjoso. Por ultimo, el �area danada seremodela a trav�es de la sustitucion del hueso primariopor hueso secundario y la resorcion del callo. La zonaafectada recupera finalmente su morfologıa y su capa-cidad de resistencia iniciales. La reparacion satisfac-toria de las lesiones oseas depende de la formacionintramembranosa y endocondral de hueso.
90
7
CARTILA
GOYHUESO

91
7
CARTILA
GOYHUESO
Figura 7.10 Secuencia de acontecimientos en la reparacion de una fractura osea. (Tomado de Gartner LP, Hiatt JL: Color Textbook of
Histology, 3rd ed. Philadelphia, Saunders, 2007, p 153.)
CONSIDERACIONES CL�INICAS
Cuando el hueso sufre danos de tal magnitud quehacen necesaria la eliminacion de algunosfragmentos, es posible que la distancia que separalos segmentos ası creados sea excesivamentegrande para permitir la formacion de un callo oseopara crear una union osea. En este caso, se obtienehueso viable para su implantacion a partir de unbanco de huesos en el que se almacenan fragmentosde hueso congelados con el fin de mantener su
capacidad osteog�enica. Existen tres tipos de injertososeos. Los autoinjertos son injertos oseosprocedentes del receptor del trasplante y son los que
tienen m�as �exito. Los homoinjertos son injertosque provienen de otro sujeto de la misma especie.Estos trasplantes entranan un riesgo de rechazoinmunitario del injerto. Los heteroinjertos sonfragmentos oseos obtenidos de una especiediferente y se asocian a los resultados menossatisfactorios.
Los androgenos y los estrogenos sintetizados porel aparato reproductormasculino y femenino influyenen la maduracion del esqueleto al incidir en el cierre
de las placas epifisarias. El desarrollo esquel�etico seinterrumpe en sujetos con una maduracion sexualtemprana, ya que este proceso estimula el cierreprematuro de dichas placas. En los individuos con
una maduracion sexual diferida se produce elfenomeno contrario, ya que su crecimiento oseo
prosigue durante un perıodo m�as prolongado al
continuar activas las placas epifisarias despu�es delespacio temporal normal.
La osteoporosis se vincula con una disminucion dela masa osea y afecta a unos 10 millones deestadounidenses, en particular a las mujeres
posmenop�ausicas y las mayores de 40 anos. La
disminucion de la sıntesis estrog�enica en estosgrupos da lugar a una reduccion de la poblacion deosteoblastos implicados en la produccion de matrizosea. Asimismo, la actividad de los osteclastossupera la actividad de deposito de hueso, lo queprovoca un descenso de la masa osea. La gravedadde la reduccion puede ser de tal magnitud queincremente la fragilidad de los huesos del sujetoafectado. Lasmujeres osteoporoticas se han tratadocon hormonoterapia sustitutiva de estrogenos. Noobstante, se ha descubierto que este tratamientoincrementa el riesgo de afecciones cardıacas,
c�ancer de mama, accidente cerebrovascular y
formacion de co�agulos. Por consiguiente, lahormonoterapia se ha sustituido por un nuevo grupo
de f�armacos (bisfosfonatos) que reducen laincidencia de las fracturas osteoporoticas.
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

MANTENIMIENTO DE LAS CONCENTRACIONES
HEMATICAS DE CALCIO
Las concentraciones hem�aticas de calcio est�an some-tidas a un estrecho control y se mantienen en unosvalores de 9 a 11mg/dl. El hueso constituye la reservade este mineral en el organismo, ya que alrededor del99% del calcio corporal se almacena en forma decristales de hidroxiapatita; la fraccion restante (1%)se localiza en las osteonas reci�en formadas, de los quepuedemovilizarse con rapidez para su utilizacion. Losiones presentes en el hueso y el torrente circulatorioest�an sometidos a un intercambio permanente.
INFLUENCIA HORMONAL
La disminucion de las concentraciones hem�aticas decalcio induce la secrecion de PTH por las c�elulas para-tiroides, como consecuencia de lo cual los osteoblas-tos comienzan a secretar el factor estimulador deosteoclastos y OPGL y se detiene la osteogenia. Seactivan los osteoclastos inactivos y se induce el desa-rrollo de nuevos osteoclastos, que ponen enmarcha laresorcion osea y provocan, en ultima instancia, laliberacion de iones de calcio almacenados en el huesoque se dirigen al torrente circulatorio.
Las c�elulas parafoliculares (c�elulas C) de la gl�andulatiroidea vigilan las concentraciones plasm�aticas deiones de calcio. El aumento de estas concentracionesdesencadena la secrecion de calcitoninapor esta pobla-cion celular, una hormona que sensibiliza receptorespresentes en la membrana de los osteoclastos paradetener la resorcion osea. De manera simult�anea, seinduce la sıntesis de osteoides por los osteoblastos yse recluta y deposita calcio en el hueso en formacion. Ellobulo anterior de la hipofisis produce somatotropina,la cual controla el desarrollo oseo a trav�es de factoresde crecimiento insulinoides, conocidos anterior-mente como somatomedinas, que estimulan el creci-miento de la placa epifisaria. Otras mol�eculas queinfluyen en el metabolismo oseo son:
. Interleucina 1, sintetizada por los osteoblastos, activala proliferacion de precursores de los osteoclastos yestimula de manera indirecta a los osteoclastos.
. Interleucina 6, procedente de c�elulas oseas, inducela formacion de nuevos osteoclastos.
. OPG, que inhibe la diferenciacion de lososteoclastos.
. Factor de necrosis tumoral, fabricado por losmacrofagos activados, desempena una funcionsimilar a la interleucina-1.
. Interferon g, formado por los linfocitos T, impide laformacion de osteoclastos.
. Factor estimulador de colonias 1, derivado de lasc�elulas del estroma, induce la formacion deosteoclastos.
. Factor transformador del crecimiento b, liberadopor lamatriz osea en el transcurso de la degradaciondel hueso, induce la osteogenia e inhibe laformacion de osteoclastos.
En el desarrollo del esqueleto tambi�en influyen lasvitaminas, adem�as de las hormonas citadas (tabla 7.4).
ARTICULACIONES
El hueso puede articularse con otro hueso en unaarticulacion movil o bien puede acercarse a otro enuna articulacion inmovil. La clasificacion de las articu-laciones se sustenta en la ausencia de movimiento(sinartrosis) o el movimiento libre (diartrosis) delos dos huesos de la articulacion (fig. 7.11).
Las articulaciones de sinartrosis se dividen en tressubgrupos:
. Sinostosis: el movimiento es reducido oinexistente; el hueso es tejido que une la articulacion(p. ej., huesos parietal derecho e izquierdo delcr�aneo adulto).
. Sincondrosis: el rango de movimiento es limitado;el tejido que une la articulacion se compone decartılago hialino (p. ej., articulacion esternocostal).
. Sindesmosis: el rango de movimiento es escaso;el tejido que une la articulacion est�a formadopor tejido conjuntivo denso (p. ej., articulaciontibiofibular inferior unida por el ligamentointeroseo).
Las articulaciones de diartrosis son las m�asabundantes en las extremidades (v. fig. 7.10). Lassuperficies que se articulan de los huesos de estasarticulaciones presentan una cubierta permanentede cartılago hialino (cartılago articular). Los liga-mentos unidos ambos huesos suelen mantener encontacto entre ellos. Existe una c�apsula articular querodea y sella la articulacion. La capa fibrosa externade esta c�apsula consta de tejido conjuntivo fibroso,que se continua con el periostio de ambos huesos dela articulacion. La capa interna, la capa sinovialcelular (membrana sinovial), recubre todas lassuperficies articulares exceptuando a las superficiesque se articulan. La capa sinovial est�a formada pordos tipos celulares:
. Las c�elulas de tipo A son macrofagos que fagocitanresiduos presentes en la cavidad articular.
. Las c�elulas de tipo B secretan lıquido sinovial.
El lıquido sinovial aporta nutrientes y oxıgeno a loscondrocitos del cartılago articular. Presenta leucoci-tos, abundante �acido hialuronico y lubricina, unaglucoproteına que se combina con el plasma extrava-sado para lubricar la articulacion.
92
7
CARTILA
GOYHUESO

93
7
CARTILA
GOYHUESO
CONSIDERACIONES CL�INICAS
La artrosis es una enfermedad degenerativa queafecta a las articulaciones sinoviales y que serelaciona con el desgaste por uso del cartılagoarticular de los condilos de uno o ambas piezasoseas de la articulacion. El cartılago hialino comienzaa degenerar hasta erosionarse y los huesoscorticales de los condilos contactan entre sı alarticularse, lo que origina un dolor lo suficientementeintenso para limitar el rango de movimiento de laarticulacion y debilitar al individuo.
La artritis reumatoide es un trastorno de lasarticulaciones sinoviales originado por ladestruccion de la membrana sinovial. La membranasinovial sufre un proceso de engrosamiento e
infiltracion por c�elulas plasm�aticas y linfocitos.El cartılago articular termina por destruirse y sesustituye por tejido conjuntivo fibrovascular, lo queprovoca un dolor intenso al mover la articulacion.
El raquitismo es una enfermedad de lactantes yninos que se debe a la carencia de la vitamina D.En ausencia de esta vitamina, la mucosa intestinalno puede absorber el calcio incluso cuando elaporte de este mineral en la alimentacion esadecuado. La carencia de calcio altera la
osificacion en los cartılagos epifisarios y la
orientacion de las c�elulas metafisarias, lo que dalugar a una calcificacion insuficiente de la matrizosea. El raquitismo se caracteriza por ladeformacion y la debilidad de los huesos del nino,en especial de los de las piernas, ya que nopueden soportar el peso corporal.
La osteomalacia es una afeccion producida por ladeficiencia prolongada de vitamina D en el adulto(raquitismo del adulto). La carencia de esta vitaminadurante un perıodo prolongado impide lacalcificacion normal del hueso en formacion duranteel proceso de remodelacion. Esta entidad puedereagudizarse durante el embarazo, ya que el fetorequiere calcio y la unica fuente para obtenerlo es lamadre.
El escorbuto es una enfermedad causada por lacarencia de vitamina C. La ingesta inadecuada de
esta vitamina altera la sıntesis de col�ageno, lo quese traduce en la formacion inadecuada de matrizosea y el desarrollo oseo anomalo. Asimismo, estetrastorno da lugar a un retraso de la cicatrizacional ser inadecuadas las concentraciones de
col�ageno.
Figura 7.11 Anatomıa de una articulacion de diartrosis. (Tomado
de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Phila-
delphia, Saunders, 2007, p 156.)
Tabla 7.4 VITAMINAS Y SUS EFECTOSEN EL DESARROLLO DEL ESQUELETO
Vitamina Efectos en el desarrollo esquel�etico
Carencia devitamina A
Inhibe la formacion osea normal, yaque altera la coordinacion de las
actividades de osteoblastos y
osteoclastos. La resorcion y la
remodelacion deficientes de laboveda craneal, necesarias para
acomodar el enc�efalo, ocasionan
lesiones graves al sistema nerviosocentral
Hipervitaminosis
A
Erosion de las columnas de cartılago sin
aumentar la poblacion celular de la
zona de proliferacion. Las placasepifisarias pueden obstruirse, lo que
interrumpirıa el crecimiento en una
etapa prematura
Carencia devitamina C
Afectacion del tejido mesenquimatoso,ya que el tejido conjuntivo no puede
producir ni mantener la matriz
extracelular. La sıntesis deficiente decol�ageno ymatriz osea origina retraso
del crecimiento y cicatrizacion
diferida. Escorbuto
Carencia devitamina D
Alteracion de la osificacion del cartılagoepifisario. Las c�elulas se desorganizan
en las met�afisis, lo que provoca una
calcificacion deficiente de los huesos,
que se deforman al soportar el peso.En ninos: raquitismo. En adultos:
osteomalacia
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

8 MUSCULOLos animales puedenmoverse y desplazar la sangre y otrosmateriales a lo largo de la luz de las estructuras tubularesque forman parte de su organismo gracias a la presencia dec�elulas musculares elongadas especiali-zadasdotadasde la capacidadde contrac-cion. Estas c�elulas musculares se dividenen dos grupos: estriadas, las cuales pre-sentan bandas alternas claras y oscuras; ylisas, carentes de estas bandas. Se distin-guen dos tipos de musculo estriado:
. Esquel�etico, encargado de losmovimientos voluntarios, y
. Cardıaco, implicado en el bombeode la sangre (fig. 8.1).
Para describir a estas c�elulasespecializadas se aplica una terminologıaespecial. Su membrana plasm�atica recibe el nombre desarcolema, su retıculo endopl�asmico liso se conoce comoretıculo sarcopl�asmico y sus mitocondrias se denominansarcosomas en algunas ocasiones. Su longitud supera concreces su anchura, por lo que suelen llamarse fibras mus-culares. Los tres tipos de c�elulas musculares derivan delmesodermo.
Musculo esquel�etico
Las c�elulas del musculo esquel�etico se componen decientos de mioblastos que se alinean y fusionan paraformarmiotubos. Cadamiotubo se encarga de la sıntesisde sus propios elementos contr�actiles, los miofilamen-tos, que se organizan para formarmiofibrillas, ası comocomponentes citopl�asmicos y org�anulos. Las c�elulas delmusculo esquel�etico:
. Pueden tener una longitud de varios centımetros y undi�ametro comprendido entre 10 y 100mm, y
. Se organizan de tal modo que se encuentran enparalelo y las bandas oscuras y claras de c�elulasadyacentes.
Los espacios extracelulares est�an ocupados por capi-lares continuos.
La fuerza del musculo esquel�etico es proporcional alnumero y el di�ametro de las fibras musculares que inte-gran un musculo dado.
. Las fibras blancas (p. ej., pechuga de pollo) secontraen con rapidez, pero se fatigan con facilidad.
. Las fibras rojas (p.ej., carne roja) se contraen demanera m�as lenta y no se fatigan con facilidad.
. Las caracterısticas de las fibras intermedias seencuentran a mitad de camino entre las de las fibrasblancas y rojas.
Las fibras blancas presentan una vascularizacionmenos profusa, un numero m�as bajo de mitocondrias,un numero inferior de enzimas oxidativas y una cantidad
menor de mioglobina, la proteına quetransporta oxıgeno, aunque su grosor esmayor y su retıculo sarcopl�asmico est�am�as desarrollado. La inervacion de lafibra determina el tipo al que pertenece(roja o blanca); el traslado experimen-tal de una c�elula muscular de un tipo defibra a otro tipo induce su adaptacion ala nueva inervacion.
Adem�as de aprovechar la energıagenerada por la contraccion del mus-culo, los elementosdel tejido conjuntivodel musculo esquel�etico conducen loselementos neurovasculares hacia cada
c�elulamuscular y delimitan lamasamuscular enunidadesmenores, denominadas fascıculos. Cada fascıculo serodea de su perimisio (v. fig. 8.1), que est�a formado porun gran numero de fibras de musculo esquel�etico, cadauna de las cuales posee un revestimiento de tejido con-juntivo menos denso, el endomisio, cuyas fibras reticu-lares se entremezclan con las de las c�elulas adyacentes.El tejido conjuntivo que rodea al musculo, el epimisio(v. fig. 8.1), se continua con los tendones y las aponeu-rosis del mismo y establecen una estrecha relacion con lasfibras reticulares del endomisio que se entremezclan conlos extremos de la c�elula muscular; esta conexion recibe ladenominacion de unionmiotendinosa.
MICROSCOPIA OPTICA DEL MUSCULO ESQUELETICO
A lo largo de la superficie de la fibra de musculoesquel�etico se encuentran unas pequenas c�elulas rege-nerativas, llamadas c�elulas sat�elite, que comparten lal�amina externa con aquella. Asimismo, en el endomisioaparecen algunos fibroblastos. El citoplasma de lasfibras de musculo esquel�etico contiene un gran numerode miofibrillas cilındricas.
. Las miofibrillas se disponen de manera precisa, demodo que las bandas claras y oscuras coincidan con lasde las c�elulas vecinas; estas bandas se encuentranalineadas en toda la fibra muscular.
. Un disco Z (lınea) atraviesa las bandas I.
. Las bandas A, de color oscuro, se dividen en dos por labanda H, atravesada por una delgada lınea M.
. La unidad contr�actil del musculo esquel�etico, elsarcomero, comprende la distancia entre dos discos Zsucesivos.
. El sarcomero se acorta a lo largo de la contraccionmuscular, los discos Z se aproximan, la banda Hdesaparece y las bandas I se estrechan, mientras que labanda A permanece inalterada.
� 2011 Elsevier Espana, S.L. Reservados todos los derechos
TERMINOS CLAVE
. Musculo esquel�etico
. Miofibrillas
. Sarcomero
. Miofilamentos
. Unionneuromuscular
. Musculo cardıaco
. Musculo liso
94

95
8
MUSCULO
CONSIDERACIONES CL�INICAS
La miositis temporal es una inflamacion de leve a
intensa de la musculatura esquel�etica debida a unalesion accidental, una infeccion, el ejercicio intenso,
una infeccion vırica o ciertos f�armacos con receta
m�edica. Cursa con mialgia, debilidad muscular,dolor a la palpacion en la region que recubre almusculo afectado, rubefaccion y reduccion oalteracion de la funcion muscular. Como su nombreindica, no representa un trastorno grave; se trata de
una entidad de car�acter temporal que remite por sısola tras la desaparicion del factor desencadenante.
Lamiositis puede constituir una afeccion muy graveque engloba a un gran numero de miopatıasinflamatorias: dermatomiositis, miositis porcuerpos de inclusion, miositis juvenil, y polimiositis.
Todas ellas son idiom�aticas, aunque pueden ser deorigen autoinmunitario. Los sıntomas generales deestos trastornos engloban mialgia y debilidadmuscular; malestar general; movilidad reducida (en
especial, al subir escaleras y levantarse despu�es deuna caıda); y dificultad frecuente para deglutir(disfagia).
Figura 8.1 Diagrama de los tres tipos de
musculo. A. Esquel�etico. B. Liso.C. Cardıaco. (Tomado de Gartner LP,
Hiatt JL: Color Textbook of Histology, 3rd ed.
Philadelphia, Saunders, 2007, p 159.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

96
8
MUSCULO
MICROSCOPIA ELECTRONICA DEL MUSCULO
ESQUELETICO
El sarcolema se asemeja, en gran medida, a las membra-nas de otros tipos celulares, si bien presenta numerosasinvaginaciones tubulares y profundas en el musculoesquel�etico.
. Los tubulos T (fig. 8.2) se extienden hacia elcitoplasma y se entremezclan en el interior de la fibramuscular en la zona de union de las bandas I y A. Dostubulos T de cada sarcomero conducen ondas dedespolarizacion hacia el interior de la fibra muscular.
. Dos cisternas terminales, regiones dilatadas delretıculo sarcopl�asmico que almacenan calcio,flanquean a cada tubulo T en las regiones de union I-A(formando una trıada) alrededor de cada miofibrilla.
. Los canales de liberacion de calcio con compuerta devoltaje (receptores de rianodina) de las cisternasterminales mantienen una estrecha relacion con losreceptores sensibles a dihidropiridina (RSDH)sensibles al voltaje de los tubulos T (que conformanun complejo llamado pie de union). Los RSDHinducen la apertura de los canales de liberacion decalcio conforme avanza la onda de despolarizacionhacia el interior de la c�elula muscular, de modo queeste ion pasa de las cisternas terminales hacia elsarcoplasma (fig. 8.3; v. fig. 8.2).
Las bandas A e I de los miofilamentos adyacentes seencuentran alineadas entre sı.
. Esta relacion se mantiene gracias a la accion de ladesmina, que envuelve a los discos Z de lasmiofibrillasadyacentes, asegur�andolas entre sı y a los discos Z conayuda de la plectina.
. La proteına de choque t�ermico ab-cristalina confiereproteccion a la desmina frente a las fuerzas a las queest�a sometida.
. La proteına de union a actina distrofina fija la desminaa los cost�ameros del sarcolema.
. Entre los haces demiofilamentos y a nivel profundo enel sarcoplasma aparecen mitocondrias tubularesalargadas. La mioglobina abunda en el sarcoplasma.
ESTRUCTURA DE LAS MIOFIBRILLAS
Las bandas claras y oscuras que se observan en la micros-copia optica se deben a la presencia de miofilamentosinterdigitados paralelos:
. Miofilamentos delgados (1mm de longitud, 7 nm dedi�ametro, formados principalmente por actina) y
. Miofilamentos gruesos (1,5mmde longitud, 15 nmdedi�ametro, compuestos fundamentalmente pormiosina II).
Los filamentos delgados parten de ambos lados deldisco Z en sentidos opuestos hacia el centro de dos sar-comeros sucesivos. Los dos discos Z de un mismo sarco-mero poseen filamentos delgados que se proyectan hacia
el centro del mismo y hacia el centro de los situados a suderecha e izquierda.
Los filamentos gruesos y delgados no se extienden a lolargo del sarcomero en una c�elula del musculoesquel�etico relajada, y el �area de un disco Z determinado,formado solamente por filamentos delgados, representala banda I observada en la microscopia optica.
. La banda I se divide en dos mitades, cada una de lascuales pertenece a un sarcomero.
. El �area de un sarcomero relajado dado formado por unfilamento grueso en su longitud total corresponde a labanda A. El centro de la banda A de un sarcomerorelajado est�a exento de filamentos delgados, lo querepresenta la banda H, una region que contienenumerosas mol�eculas de creatina cinasa, la enzimaque cataliza la transferencia de grupos fosfato de altaenergıa del fosfato de creatina para sintetizartrifosfato de adenosina (ATP).
. Enel centrode labandaHse localiza la lıneaM, formadamayoritariamente por proteına C ymiomesina, dosmacromol�eculas que conectan los filamentos gruesosentre sı y participan en elmantenimiento de su posicioncorrecta con el fin de permitir la conexion de losfilamentos gruesos con los delgados.
Cuandoelmusculosecontrae, los filamentosdelgadossedeslizan m�as all�a de los gruesos y acercan los discos Z entresı, de modo que el sarcomero se acorta aproximadamente0,4mm.Una sola c�elula demusculo esquel�etico puede estarformada por una secuencia de 100.000 sarcomeros, por loque ladisminucionde su longituden0,4mmpuede suponerun acortamiento de 4cm del musculo contraıdo. La organi-zacion estructural de los filamentos gruesos debe mante-nerse para permitir su interaccion con los delgados.
En el musculo esquel�etico de mamıferos, cada fila-mento grueso se rodea de seis filamentos delgados aintervalos de 60�. En un corte transversal, estos filamen-tos configuran un hex�agono cuyo centro ocupa un fila-mento grueso (fig. 8.4). El mantenimiento de la disposi-cion relativa de los componentes del sarcomero corre acargo de cinco proteınas:
. Dos mol�eculas de titina, unas proteınas el�asticas degran tamano que parten de cada disco Z de un mismosarcomero hasta la lınea M, aseguran a los filamentosgruesos en su posicion correcta.
. Las a-actininas anclan los filamentos delgados aldisco Z.
. Dosmol�eculas de nebulina se extienden desde el discoZ hacia el extremo de cada filamento delgado paraasegurar la posicion de los filamentos delgados y elmantenimiento de su longitud adecuada.
. Adem�as, Cap Z y tropomodulina, unas mol�eculas queevitan la adicion o la delecion de actina G en elfilamento delgado, intervienen en el mantenimientode su longitud correcta. La primera actua en el extremopositivo irregular (en el disco Z), mientras que lasegunda lo hace en el extremo negativo afilado delfilamento delgado (v. fig. 8.4).

97
8
MUSCULO
Figura 8.2 Organizacion de los sarcomeros y las miofibrillas de una c�elula de musculo esquel�etico. (Tomado de Gartner LP, Hiatt JL:
Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 161.)
Figura 8.3 Organizacion de
trıadas y sarcomeros de lasfibras del musculo esquel�etico.
(Tomado de Gartner LP,
Hiatt JL: Color Textbook of
Histology,3rd ed. Philadelphia, Saunders,
2007,
p 162.)
Figura 8.4 A-D. Miofilamentos de un sarcomero. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia,
Saunders, 2007, p 164.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Filamentos gruesos
Cada filamento grueso se compone aproximadamente de300mol�eculas demiosina II, con un di�ametro compren-dido entre 2 y 3nm y una longitud de 150 nm. La mio-sina II consta de:
. Dos cadenas pesadas
. Dos pares de cadenas ligeras, cada uno de los cualesintegra una cadena ligera esencial y una cadena ligerareguladora (fig. 8.5); la cadena reguladora essusceptible de fosforilacion por parte de la cinasa de lacadena ligera de la miosina (MLCK)
Cada una de las dos cadenas pesadas id�enticas seasemeja a un palo de golf, de modo que las cadenaspolipeptıdicas (asas) de ambas cadenas forman unah�elice al enroscarse entre sı. La enzima tripsina puedeescindir la cadena pesada para formar:
. Una mol�ecula de meromiosina ligera similar a unbaston
. Una mol�ecula de meromiosina pesada formadapor dos cabezas globulares y un corto tallo; lasprimeras se componen de dos cadenas polipeptıdicasque se envuelven entre sı; la papaına escinde estamol�ecula en dos regiones globulares (S1) y un tallocorto (S2)
Cada porcion S1 posee tres sitios de union, a saber,sitio de union a ATP, sitio de union a miosina de cadenaligera y sitio de union a actina F. La disposicion de lasmol�eculas de miosina es tal que suceden longitudinal-mente en un filamento grueso de modo que su centro esliso y sus extremos son irregulares debido a la presenciade las porciones S1. Lasmol�eculas demiosina poseen dosregiones flexibles, una en la zona de union de las por-ciones S1 y S2 y otra en la union de las meromiosinaspesada y ligera, que permiten a la miosina II entrar encontacto con el filamento delgado para acercarlo hacia elcentro del sarcomero.
Filamentos delgados
Los filamentos delgados, formados por actina F, tropo-miosina y troponina, presentan un extremo positivoirregular unido al disco Z y un extremo negativo afiladorecubierto por unamol�ecula de tropomodulina (fig. 8.6).
. La actina F consta de dos cadenas de polımerosde actina G, semejantes a dos hileras de perlasenroscadas entre sı. En los dos surcos someros asıformados se alojanmol�eculas de tropomiosina linealesde 40nm de longitud dispuestas de cabeza a cola.
. Las mol�eculas de tropomiosina ocultan el sitio activode cada mol�ecula de actina G, de modo que impidenque entre en contacto con la subunidad S1 de lamol�ecula de miosina II.
. Una mol�ecula trim�erica de troponina aparece unida acada mol�ecula de tropomiosina. Se compone de
troponina C (TnC), la cual se une a iones libres decalcio; troponina T (TnT), que se une a la mol�eculade troponina de tropomiosina; y troponina I (TnI),que inhibe la interaccion de la subunidad S1 con laactina G.
. Si hay iones libres de calcio, se unen a lasubunidad TnC para inducir un cambioconformacional en la troponina que empuja a latropomiosina a una porcion m�as profunda del surcodel filamento de actina F, de modo que se descubre elsitio activo y se produce una union temporal con lasubunidad S1.
CONTRACCION MUSCULAR
La contraccion muscular suele suceder a un impulsonervioso y obedece la ley de todo o nada en cadafibra nerviosa, segun la cual la c�elula se contrae o no. Elgrado de acortamiento depende del numero de sarcome-ros de una miofibrilla dada y la potencia de la contra-ccion de todo un musculo se relaciona con el numero dec�elulas musculares que se contraen. Los miofilamentosno se contraen, sino que los filamentos delgados se des-plazan sobre los gruesos de acuerdo con la teorıa delfilamento deslizante de Huxley del siguiente modo:
. Los tubulos T conducen el impulso generado en launion mioneural hasta las cisternas terminales. Loscanales de liberacion de calcio con compuerta devoltaje de las cisternas se abren y los iones Ca++
liberados al sarcoplasma se unen a TnC, lo que provocaun cambio conformacional y empuja a latropomiosina hacia una porcion m�as profunda delsurco, lo que deja al descubierto el sitio activo de laactina G.
. La hidrolisis de ATP del fragmento S1 de la miosina IIda como resultado difosfato de adenosina (ADP)y fosfato inorg�anico (Pi), los cuales permanecenunidos al grupo S1. La cabeza de la miosina sedesplaza y el complejo se une al sitio de union amiosina de actina G (v. fig. 8.6).
. El Pi se separa del complejo, demodo que se refuerza elenlace formado entre la miosina y la actina y seproduce un nuevo cambio conformacional en elgrupo S1 para liberar el ADP; se modifica laconformacion de la cabeza de la miosina, que arrastraal filamento delgado hacia el centro del sarcomero.Este movimiento recibe el nombre de golpe de fuerzade la contraccion muscular.
. El grupo S1 acepta una nueva mol�ecula de ATP, con loque se rompe el enlace formado entre la actina y lamiosina (v. fig. 8.6).
. Los ciclos de union y disociacion han de repetirse entre200 y 300 veces para que la contraccion muscular seacompleta y cada ciclo requiere la hidrolisis de unamol�ecula de ATP.
98
8
MUSCULO

99
8
MUSCULO
CONSIDERACIONES CL�INICAS
Las mutaciones de algunas proteınas estructurales
responsables de mantener la integridad de la organizacion
de las miofibrillas del musculo esquel�etico pueden tener
unos efectos notabilısimos. La alteracion de la estructura
primaria del filamento intermedio desmina o la proteına de
choque t�ermico cristalina aB dificulta el mantenimiento de
la posicion normal de las miofibrillas en el espacio
tridimensional, de modo que se destruyen al ser sometidas
a fuerzas intensas de contraccion.
El rıgor mortis es un estado que aparece con
posterioridad a la muerte. El ATP del grupo S1 (cabeza de
la miosina) de la miosina II se hidroliza en ADP y Pi durante
la contraccion muscular en un sujeto vivo, y ambosproductos permanecen unidos a la cabeza de la miosina.
El cambio conformacional de la miosina II permite que
su cabeza se acerque al sitio de union a miosina de la
actina G del filamento delgado. El Pi se libera a raız de
este contacto y se establece un enlace m�as firme entre la
miosina y la actina; a continuacion, el ADP se disocia de
la cabeza de la miosina, lo que origina un golpe de fuerza.
Otra mol�ecula de ATP se une a la cabeza de la miosina y
se rompe el enlace entre el grupo S1 de la miosina II y laactina G del filamento delgado. En una persona muerta, el
ATP no se regenera, de modo que al cabo de cierto
tiempo terminan por agotarse las reservas de esta
mol�ecula; el retıculo sarcopl�asmico es incapaz de
secuestrar iones de calcio y la contraccion muscular se
mantiene hasta que no quede ninguna mol�ecula de ATPpara separar el grupo S1 de la miosina del filamento
delgado, y sobreviene una contraccion muscular
mantenida (es decir, rigidez muscular). Este estado recibe
el nombre de rıgor mortis. En funcion de la temperaturaambiental, las enzimas lisosomicas abandonan los
lisosomas algun tiempo despu�es y degradan las
mol�eculas de actina y miosina, de modo que desaparece
el rıgor mortis. A finales de la primavera en las regiones
templadas, el rıgor mortis comienza entre 3 y 8 h despu�esde la muerte y se mantiene a lo largo de un perıodo
comprendido entre 16 y 24h; los musculos se relajan unas
36h despu�es del fallecimiento.
Figura 8.5 A-D.Miofilamentos gruesos y delgados de un sarcomero. (Tomado deGartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.
Philadelphia, Saunders, 2007, p 164.)
Figura 8.6 Papel del ATP en la contraccion
muscular. (Modificado de Alberts B, Bray D,
Lewis J, et al: Molecular Biology of the Cell. New York,Garland Publishing, 1994.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

RELAJACION MUSCULAR
El proceso de contraccion muscular requiere la presenciade iones libres de calcio en el sarcoplasma. Al interrum-pirse el estımulo nervioso, los tubulos T dejan de con-ducir la onda de despolarizacion hacia el interior de lac�elula muscular, se cierran los canales de liberacion decalcio con compuerta de voltaje de las cisternas termi-nales.
. Las bombas de calcio transportan el calcio presente enel sarcoplasma, que regresa al retıculo sarcopl�asmico,en el que ser�a secuestrado por la calsecuestrina.
. La disminucion de las concentraciones de calcio inducela liberacion de este ion de las subunidades TnC, querecuperan su conformacion relajada; la mol�ecula detropomiosina regresa a su posicion anterior paraocultar el sitio activo de la mol�ecula de actina G,a la que la miosina no puede ya unirse.
INERVACION DEL MUSCULO ESQUELETICO
El musculo esquel�etico recibe fibras nerviosas motoras,que inducen la contraccion muscular; fibras nerviosassensoriales, que inervan los haces musculares y los orga-nos tendinosos deGolgi que protegen almusculo frente alesiones, y fibras autonomicas, que controlan la irriga-cion del musculo. En funcion del grado de coordinacionfina de un musculo dado, puede contar con:
. Inervacion rica, como es el caso de los musculosoculares, en las que una unica neuronamotora tan solopuede controlar cinco c�elulas musculares, o bien
. Inervacion cruda, como sucede en los musculos de laespalda, en los que una neurona motora aislada puedecontrolar varios cientos de c�elulas musculares.
La neurona motora y todas las c�elulas musculares con-troladas por ella reciben el nombre de unidad motora.Todas las fibras musculares de una unidad motora dadapueden contraerse de forma simult�anea o bien no hacerloen absoluto.
Transmision del impulso en la union neuromuscular
Los axones mielinizados de las neuronas motoras ainervan las c�elulas del musculo esquel�etico. Estos axonesemplean elementos del tejido conjuntivo del musculo amedida que se ramifican para inervar cada c�elula delmusculo esquel�etico de la unidad motora a la que perte-necen. La ramificacion del axon pierde su vaina de mie-lina al alcanzar la c�elula muscular correspondiente, aun-que continua rode�andose de las c�elulas de Schwann yforma un terminal axonal expandido (membrana pre-sin�aptica) sobre la placa terminal motora (membranapostsin�aptica), una region modificada del sarcolema. Lacombinacion de la placa terminal motora, la hendidurasin�aptica (primaria) (espacio comprendido entre las
membranas presin�aptica y postsin�aptica) y el terminalaxonal recibe el nombre de union neuromuscular(fig. 8.7).
La membrana postsin�aptica presenta numerososrepliegues que delimitan unos espacios denominadoshendiduras sin�apticas secundarias (pliegues de launion). Los pliegues y las hendiduras sin�apticas secun-darias est�an recubiertos por una l�amina externa. Lasc�elulas de Schwann recubren el terminal axonal, quealberga mitocondrias, el retıculo sarcopl�asmico y varioscientos demiles de vesıculas sin�apticas que contienen elneurotransmisor acetilcolina, proteoglucanos, ATP yotras mol�eculas. La membrana presin�aptica posee barrasdensas proximas a canales de calcio reguladas por vol-taje. La transmision del estımulo se produce del siguientemodo:
. El estımulo, conducido a lo largo del axon, alcanza lamembrana presin�aptica, cuya despolarizacion suscitala apertura de los canales de calcio con compuerta devoltaje y la entrada de calcio al terminal axonal.
. Alrededor de 120 vesıculas sin�apticas por cada impulsose fusionan con los sitios activos de la membranapresin�aptica en las barras densas, como consecuenciade lo cual se libera una quanta de acetilcolina(aproximadamente, 20.000 mol�eculas),proteoglucanos y ATP a la hendidura sin�apticaprimaria (fig. 8.8).
. Los receptores de acetilcolina de la membrana(muscular) postsin�aptica se unen a la acetilcolinaliberada, con lo que se abren los canales de sodio concompuerta de ligando de esta membrana y la entradade sodio despolariza el sarcolema y los tubulos T.La onda de despolarizacion alcanza las cisternasterminales y se libera calcio en la union I-A para poneren marcha la contraccion muscular.
. En menos de 500 ms, la enzima acetilcolinesterasa,situada en la l�amina externa de las hendidurassin�apticas primaria y secundaria, degrada mol�eculasde acetilcolina en colina y acetato; el potencial dereposo de la membrana postsin�aptica se restablece,de modo que se impide la liberacion de otrasmol�eculas de acetilcolina que inducirıa nuevascontracciones.
. El gradiente de concentracion de sodio impulsa elsimporte de sodio-colina para devolver las mol�eculasde colina al terminal axonal, en el que el acetatoactivado procedente de las mitocondrias se combinacon la colina con intervencion de la enzima colinaO-acetiltransferasa. La acetilcolina se transporta alinterior de vesıculas sin�apticas mediante un gradienteprotonico impulsado por proteınas transportadorasantiporte.
. El �area superficial de la membrana presin�aptica semantiene gracias al mecanismo de tr�afico demembranas.
100
8
MUSCULO

CONSIDERACIONES CL�INICAS
Clostridium tetani es una bacteria esporuladora quehabita en el suelo; en condiciones de anaerobiosis,sintetiza una toxina que bloquea la glicina,un neurotransmisor inhibidor producido poralgunas neuronas del sistema nervioso central.Generalmente, la infeccion se contrae cuando la
bacteria se introduce en una herida abierta a trav�esde restos de suelo o contaminantes. Al proliferar, losmicroorganismos fabrican la toxina, la cual llega a la
m�edula espinal e inhibe la liberacion deglicina, lo cualprovoca una contraccion muscular espasmodicaconocida como tetania. La enfermedad debuta conrigidez de los musculos al masticar y los sıntomasiniciales pueden observarse entre 2 y 50 dıas
despu�es de la infeccion. La rigidez inicial puede darpaso a la incapacidad de abrir la boca, lo que recibe
el nombre comun de trismo. Otros sıntomas son la
rigidezdeotrosgruposmusculares; en los casosm�asgraves, los musculos del cuello, el abdomen y laespalda pueden sufrir espasmos de gran intensidadque llegan a arquear el torax y el abdomen haciadelante y empujan la cabeza y las extremidades
inferiores hacia atr�as, unaposicion tıpica de la tetaniaconocida como opistotonos. La tasa de mortalidadglobal anual es de unas 50.000 personas. La tetaniapuede prevenirse mediante la administracion de unavacuna frente a la infeccion seguida de una dosis derecuerdo cada 10 anos. El tratamiento englobaantibioterapia combinada con inmunoglobulinas
antitet�anicas para inactivar la toxina. El paciente
puede requerir analg�esicos, sedantes, relajantes
musculares y ventilacion mec�anica para respirar.
101
8
MUSCULO
Figura 8.8 Acontecimientos que tienen lugar en una sinapsis de
una neurona motora y una c�elula de musculo esquel�etico.AcCoA, acetil coenzima A; ACh, acetilcolina; AChE,
acetilcolinesterasa; PG, prostaglandina; RE, retıculo
endopl�asmico. (Modificado de Katzung BG: Basic and ClinicalPharmacology, 4th ed. East Norwalk, CT, Appleton & Lange,
1989.)
Figura 8.7 Union neuromuscular. (Tomado de Gartner LP,Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia,
Saunders, 2007, p 171.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

SISTEMA SENSORIAL DEL MUSCULO ESQUELETICO
El organismo controla la actividadmuscular con el fin deevitar danos a los musculos o los tendones.
. Los husos musculares vigilan la alteracion de lalongitud del musculo y el ındice de alteracion.
. El organo tendinoso de Golgi controla las fuerzas detension y la velocidaddedesarrollo de lasmismas enuntendon, a medida que se acorta el musculo.
La informacion adquirida por ambos organos senso-riales llega a la m�edula espinal encargada de procesarla.Asimismo, la informacion se transmite al cerebelo parasu procesamiento inconsciente y a la corteza cerebral, enla que puede alcanzar niveles conscientes para que elsujeto perciba la posicion de sus musculos.
Husos musculares
Los husos musculares (fig. 8.9) son receptores senso-riales encapsulados dispersos entre las fibras del musculoesquel�etico que inducen la contraccion autom�atica de losmusculos distendidos, una respuesta propioceptoradenominada reflejo de estiramiento. Estos haces encap-sulados est�an formados por un pequeno numero defibras de musculo esquel�etico modificadas, las llamadasfibras intrafusales, que se localizan en espacio periaxialrelleno de lıquido; se disponen en paralelo al eje longi-tudinal del musculo. Aunque las c�elulas del musculoesquel�etico que rodean el huso muscular no son impor-tantes, reciben el nombre de fibras musculares extrafu-sales.
Se distinguen dos tipos de fibras intrafusales: lasfibras de bolsa nuclear y las fibras de cadena nuclear.Las primeras son m�as anchas y escasas que las segundas.Los nucleos de ambos tipos se hallan en el centro de lac�elula y sus elementos contr�actiles lo hacen en los polos.
. Los nucleos de las fibras de bolsa nuclear se agrupan enla region expandida en el centro de las c�elulas.
. Los nucleos de las fibras de cadena nuclear se alineanen una hilera, pero no se agregan, en el centro de estasc�elulas.
Las fibras de bolsa nuclear se dividen en dos clases:din�amicas y est�aticas. A pesar de que la inervacion de lasfibras intrafusales parece ser compleja, en realidad esbastante sencilla, ya que reciben dos tipos de fibras sen-soriales, las cuales inervan las regiones nucleares, y dostipos de fibras motoras, que se ocupan de inervar lasregiones contr�actiles.
. Las regiones nucleares de las fibras de cadena nuclear yambos tipos de fibras de bolsa nuclear de un husomuscular son inervadas por ramificaciones de una solafibra nerviosamielinizada grande de grupo Ia (tambi�enllamada terminaciones sensoriales Ia o din�amicas), lacual se envuelve en espiral alrededor de estas regiones.
. Las regiones nucleares de todas las fibras de cadenanuclear y unicamente las fibras de bolsa nuclearest�aticas de un huso muscular son inervadas porramificaciones de una sola fibra nerviosa sensorial degrupo II (tambi�en conocidas como terminacionesnerviosas sensoriales II o est�aticas) que envuelvenestas zonas de la c�elula (v. fig. 8.9).
. La inervacion motora de las regiones polares(contr�actiles) de todas las fibras de cadena nuclear ysolamente las fibras de bolsa nuclear est�aticas procedede los axones de neuronas motoras g est�aticas,mientras que las regiones polares de las fibras de bolsanuclear din�amica la reciben de los axones de neuronasmotoras g din�amicas.
. La inervacion de todas las fibras extrafusales procedede los axones mielinizados de neuronas motoras g(v. fig. 8.5A y B).
La distension de un musculo esquel�etico estira elhuso muscular y estimula las fibras sensoriales de losgrupos Ia (din�amicas) y II (est�aticas). A menudo, estasfibras se activan conforme se estira el musculo. Adem�as,las fibras del grupo Ia responden a cambios en la tasa dedistension muscular; el huso muscular no se limita aaportar informacion acerca de la velocidad de distensiondel musculo, sino que tambi�en lo hace sobre la disten-sion inesperada del mismo. Las neuronas motoras ginducen la contraccion de ambas regiones polares delas fibras intrafusales, provocando su distension y sen-sibilizacion a modificaciones mınimas del estado deestiramiento del musculo.
Organos tendinosos de Golgi
A diferencia de los husos musculares, los organos tendi-nosos de Golgi controlan las fuerzas de tension (y lavelocidad a la que se desarrollan) a las que est�an some-tidos los tendones como consecuencia del acortamiento(contraccion) de la musculatura esquel�etica. Se localizanen la union del musculo con el tendon, presentan unalongitud de alrededor de 1mmyun di�ametro de 0,1mm,y se disponen en paralelo al eje longitudinal delmusculo.Est�an formados por fibras de col�ageno ondulado encuyos espacios intersticiales aparecen ramificaciones nomielinizadas de axones de tipo Ib. Las fibras onduladasse estiran y comprimen las terminaciones nerviosas libresa medida que el musculo se contrae y ejerce fuerzas detension sobre el tendon. La velocidad de la producciondel impulso en estas fibras nerviosas depende de lasfuerzas de tension a las que est�a sometido el tendon.Cuando la fuerza se acerca a unos valores por encimade los cuales podrıa ocasionar danos al tendon, el mus-culo o el hueso, el organo tendinoso de Golgi interrumpela contraccion delmusculo. Los husosmusculares vigilanel estiramiento y los organos de Golgi se ocupande la contraccion del mismo musculo con el objeto decoordinar el control espinal de los reflejos del musculoesquel�etico.
102
8
MUSCULO

103
8
MUSCULO
CONSIDERACIONES CL�INICAS
MIASTENIA GRAVE
La miastenia grave es una enfermedad
autoinmunitaria que alcanza su m�axima prevalenciaen mujeres de edades comprendidas entre 20 y40 anos, si bien puede afectar a sujetos de ambossexos y de cualquier grupo etario. Alrededor del10% de estos pacientes porta algun tumor tımico;los anticuerpos pueden atravesar la barreraplacentaria, por lo que los neonatos presentanmiastenia grave temporal que remite antes deconcluir el segundo mes de vida en el 10 al 12% delas embarazadas. Los individuos afectados por estaentidad sintetizan anticuerpos frente a sus propiosreceptores de acetilcolina, lo que reduce lacapacidad normal de contraccion de lamusculatura.
A pesar de que las c�elulas musculares internalizan ysustituyen los receptores bloqueados, laenfermedad supera los mecanismos de reparacionmuscular. Esta enfermedad afecta especialmente ala musculatura facial, en particular a los musculosextrınsecos del ojo. Asimismo, altera los musculosde la garganta y el resto del organismo, lo que dalugar a dificultades del habla y la deglucion y adebilidadmuscular generalizada de lamayorıa de losgrupos musculares del organismo. El grado de
debilidad oscila de leve a intenso. En su forma m�asgrave, el trastorno recibe el nombre de crisis
miast�enica y llega a afectar a los musculosrespiratorios con consecuencias mortales. Amenudo, la administracion de los inmunosupresores
y los f�armacos que potencian la sıntesis deacetilcolina permite controlar esta enfermedad.
ARCO REFLEJO SENCILLO
Los husos musculares, como los que median elreflejo rotuliano, son dos receptores de reflejosneuronales que reaccionan iniciando la contraccionmusculo progenitor ante su estiramiento. Elsiguiente ejemplo ilustra la importancia que revisteeste reflejo: una persona que se encuentra enbipedestacion recibe una patada en la fosa poplıteaderecha (en la cara posterior de la rodilla derecha)propinada por un sujeto que se acerco a ella desde
atr�as. Esta accion suscita la flexion de dicha rodilla;la rodilla se desplaza hacia delante y la izquierda
comienza a ceder. El musculo cu�adriceps mayor(cuatro musculos localizados en la cara anterior delmuslo) de la pierna derecha se estira; las fibrassensoriales de los husos musculares se activan y laonda de despolarizacion se transmite hasta la
m�edula espinal. En la sinapsis se liberanneurotransmisores que estimulan las neuronas
motoras a del asta ventral de la m�edula espinalque inervan las fibras musculares extrafusales
del cu�adriceps derecho y provocan sucontraccion. Conforme se contrae este musculo,la pierna derecha se estira y evita la caıda delindividuo atacado. Este sistema se activa altropezar, de modo que el arco reflejo impidela caıda.
Figura 8.9 A. Diagrama de los componentes del huso
muscular. B. Representacion desplegada de distintos tipos
de fibras de un huso muscular y su inervacion.(A.Modificado de Krstic RV: Die Gewebe des Menschen und der
Saugertiere. Berlin, Springer-Verlag, 1978. B. Modificado de
Hulliger M: The mammalian muscle spindle and its centralcontrol. Rev Physiol Biochem Pharmacol 101:1–110, 1984.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Musculo cardıaco
El musculo cardıaco tambi�en se compone de musculoestriado, si bien presenta diversas diferencias con el mus-culo esquel�etico. El musculo cardıaco no est�a sometido aun control voluntario; se localiza en el corazon y las por-ciones proximales de los grandes vasos que irrigan esteorgano. El miocardio, que constituye la masa musculardel corazon, est�a formado por l�aminas, o capas super-puestas, de c�elulas de musculo cardıaco. Las l�aminas seseparan por medio de elementos delgados de tejido con-juntivo, que transportan vasos sanguıneos y nervios. Unared capilar densa irriga a cada c�elula de musculo cardıaco.
. Las c�elulas de musculo cardıaco son c�elulascilındricas ramificadas y cortas de 80mm de longitud y15mm de di�ametro dotadas de un nucleo centralsolitario (o, algunas veces, dos nucleos). Presentandepositos de glucogeno y triglic�eridos en ambos polos.
. Alrededor del 50% del sarcoplasma est�a ocupado pormitocondrias, que se disponen en paralelo al ejelongitudinal de la c�elula intercaladas entre susmiofibrillas. Una proteına transportadora de oxıgeno,la mioglobina, es muy abundante.
A diferencia de las fibras del musculo esquel�etico, lasdel musculo cardıaco poseen la capacidad de contraersede forma espont�anea y una ritmicidad inherente. Lasc�elulas modificadas de musculo cardıaco (nodulo sino-auricular, nodulo auriculoventricular, haz de His y fibrasde Purkinje), las cuales se tratan en el capıtulo 11, actuancomo elementos neurales que regulan y controlan laactividad cardıaca.
. Las c�elulas musculares del ventrıculo son mayores quelas presentes en la aurıcula.
. Las c�elulas musculares de la aurıcula presentangr�anulos auriculares rellenos de factor natriur�eticoauricular y factor natriur�etico cerebral, unasmol�eculas con efecto diur�etico que inhiben lasecrecion de aldosterona por la corteza adrenal y derenina por las c�elulas yuxtaglomerulares del rinon, locual atenua la capacidad de conservacion de sodio yagua por parte de este organo y da lugar a unadisminucion de la tension arterial.
Las c�elulas del musculo cardıaco presentan extremosaflautados que se aproximan entre sı al alinearse y danlugar a uniones interdigitantes, conocidos como discosintercalados (fig. 8.10). Cada uno de estos discos posee:
. Una porcion lateral rica en uniones gap.
. Una porcion transversal en las que abundan losdesmosomas y las fascias adherentes.
. Miofilamentos delgados unidos a la fascia adherente,que actua como un disco Z, a trav�es de a-actinina yvinculina.
. Uniones gap que facilitan el paso de informacion entrec�elulas contiguas, de manera que el proceso decontraccion puede darse de manera concertada paraque los ventrıculos se doblen sobre sı mismos yexpulsen la sangre de forma eficiente hacia la aorta y eltronco pulmonar.
Al igual que las fibras del musculo esquel�etico, lasc�elulas del musculo cardıaco presentan un bandeadocaracterıstico formado por bandas A e I y la organizacionde los sarcomeros de los dos tipos demusculo estriado esid�entica entre sı. La teorıa del filamento deslizante deHuxley tambi�en se aplica al musculo cardıaco, si bienexisten algunas diferencias relevantes:
. Los tubulos T poseen un di�ametro mayor (algo m�asdel doble del de las c�elulas del musculo esquel�etico) yest�an cubiertos por una l�amina externa con carganegativa que almacena iones de calcio a trav�es deenlaces de baja energıa. Los tubulos T no se localizanen la zona de union de las bandas A e I, sino que lohacen en el disco Z del sarcomero.
. El retıculo sarcopl�asmico de las c�elulas del musculocardıaco es menos abundante, por lo que no puedesecuestrar suficientes iones de Ca++ para poner enmarcha la contraccion.
. Por otra parte, no aparecen cisternas dilatadas delretıculo sarcopl�asmico a cada lado del el tubulo T paraformar trıadas, sino que una sola terminal de retıculosarcopl�asmico se adosa a un tubulo T de cada disco Zen una configuracion de dıadas.
. El sarcolema de las c�elulas del musculo cardıaco poseecanales de sodio r�apidos (canales de sodio de lasc�elulas del musculo esquel�etico) y canales de sodiolentos (canales de calcio-sodio) que se mantienenabiertos varias d�ecimas de segundo. Durante ladespolarizacion, los canales lentos de sodio de lostubulos se abren, de modo que los iones de calcio y desodio pasan a una region citosolica proxima al retıculosarcopl�asmico.
. Los iones de Ca++ provocan la apertura de los canalesde liberacion de calcio del retıculo sarcopl�asmico,como consecuencia de lo cual se incrementan enmayor medida las concentraciones sarcopl�asmicas deeste ion. La contraccion muscular comienza demanerasemejante a la del musculo esquel�etico. La duracion esmayor que en este ultimo, ya que los iones de K+ nopueden salir con rapidez de las c�elulas del musculocardıaco, lo que retrasa la repolarizacion delsarcolema.
104
8
MUSCULO

CONSIDERACIONES CL�INICAS
INFARTO DE MIOCARDIO
El t�ermino infarto de miocardio se refiere a losdanos ocasionados al musculo cardıaco por la
ausencia de irrigacion en el �area afectada. Lainterrupcion del flujo sanguıneo puede deberse a laoclusion total de una arteria coronariaateroesclerotica o la migracion de un trombo quese alojaba en un vaso sanguıneo de calibresuficientemente pequeno para ser obstruido por elmismo. El dano es reversible cuando se reanuda lairrigacion en el plazo de 20 minutos, a partir de loscuales se convierte en permanente y mueren las
c�elulas del musculo cardıaco que no reciben
perfusion. Las c�elulas muertas liberan troponina Iespecıfica de cardiocitos (TnI especıfica decardiocitos), un marcador del infarto de
miocardio, entre 3 y 10 h despu�es de la lesion, quese permanece en el torrente durante de 2 a 3
semanas. Otros marcadores s�ericos menosespecıficos son la creatina cinasa y la isozimaMB de la creatina cinasa.
MARCAPASOS ARTIFICIALES
Los marcapasos artificiales se implantan enpersonas afectadas por arritmia no susceptible detratamiento farmacologico. Estos marcapasos sondispositivos electronicos que se insertaninmediatamente por debajo de la piel de la clavıculay se conectan con un electrodo introducido por elsistema venoso hasta la aurıcula y el ventrıculoderechos. En funcion del tipo de marcapasosutilizado, puede controlar de manera continua lafrecuencia del latido cardıaco o bien actuar bajodemanda para regular dicha frecuencia cuando elmarcapasos biologico no funciona del modocorrecto. Otros marcadores pueden adaptar lafrecuencia cardıaca en funcion del nivel deactividad del individuo. Las baterıas de losmarcapasos duran unos 15 anos. La implantacionde estos dispositivos puede realizarse en el marcoambulatorio; la intervencion requiere entre 1 y 2 h, oincluso menos.
105
8
MUSCULO
Figura 8.10 A. Representacion tridimensional del musculo cardıaco a nivel del disco intercalado. B. Representacion bidimensional
de un disco intercalado y sus porciones transversal y lateral. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.
Philadelphia, Saunders, 2007, p 178.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Musculo liso
El musculo liso carece tanto de estriaciones como detubulos T. Se encuentra en las paredes de las vısceras;no est�a sometido a un control voluntario, sino que loregulan diversos factores locales, hormonas y el sistemanervioso autonomo; se divide en:
. Musculo liso multiunitario, en el que cada c�elulaposee su propia inervacion, o bien
. Musculo liso unitario (de unidades unicas ovasculares), en el que solamente algunas c�elulasdisfrutan de inervacion propia; la informacion seconduce a otras c�elulas del musculo liso a trav�es deuniones gap.
Adem�as de intervenir en la contraccion, las c�elulas delmusculo liso sintetizan diversasmacromol�eculas que for-man parte de la matriz extracelular.
MICROSCOPIA OPTICA DE LAS FIBRAS
DE MUSCULO LISO
Las fibras del musculo liso son c�elulas fusiformes elon-gadas cortas con un promedio de longitud de 200mm omenos y un promedio de di�ametro de 5 a 6mm. El nucleoovalado unico se localiza en el centro de la c�elula alrealizar un corte longitudinal, si bien adopta una posi-cion ac�entrica en un corte transversal. La c�elula gira sobresı misma durante la contraccion, de tal modo que elnucleo remeda un sacacorchos (fig. 8.11). Las c�elulasde musculo liso se rodean de una l�amina externa queest�a ausente en las uniones gap. Las fibras reticulares seincluyen en la sustancia de dicha l�amina y aprovechan lasfuerzas de contraccion.
Algunas tinciones especiales, como la de hematoxi-lina-hierro, permite visualizar unas delgadas estrıas lon-gitudinales que representan grupos de miofilamentos.Por otra parte, los cuerpos densos que actuan comodiscos Z aparecen en el citoplasma y a lo largo de la caracitopl�asmica del sarcolema. Generalmente, las c�elulas delmusculo liso suelen formar una hoja en la que cada c�elulaest�a sometida a un grado alto de empaquetamiento y susextremos afilados se ajustan casi exactamente a las regio-nes m�as anchas de las c�elulas adyacentes.
MICROSCOPIA ELECTRONICA DEL MUSCULO LISO
El sarcoplasma de las c�elulas del musculo liso de cadapolo del nucleo alberga mitocondrias, el retıculo sar-copl�asmico, el aparato de Golgi y depositos de gluco-geno. Aparecen, adem�as, miofilamentos, aunque no seasocian en una configuracion paracristalina como sucedeen el musculo estriado.
. Los filamentos delgados son semejantes a losdel musculo estriado; no obstante, se asocian a laproteına caldesmona, que enmascara el sitio activode la actina G, en lugar de la troponina.
. Los filamentos gruesos est�an formados porsubunidades de miosina II, aunque en lugar dealinearse de cabeza a cola, las cabezas de miosinasobresalen a lo largo de la longitud del filamento. Estaorganizacion hace posible unas contracciones deduracion m�as prolongada que en el musculo estriado,que no se rigen por la ley de todo o nada. Puedecontraerse solamente una porcion de la c�elula demusculo liso.
. Los filamentos intermedios vimentina y desmina en elmusculo liso unitario y desmina en el musculo lisomultiunitario aprovechan las fuerzas de contraccionproducidas por los miofilamentos.
. Los filamentos intermedios y delgados se anclan en loscuerpos densos, unas estructuras integradas poractinina a y otras mol�eculas vinculadas con los discosZ que aparecen adosados al sarcolema y dispersos en elcitoplasma. Los cuerpos densos originan un complejointerconectado que se encarga de hacer girar a lasc�elulas de musculo liso sobre sı mismas en eltranscurso de la contraccion.
. El sarcolema de las c�elulas de musculo liso posee unsistema bien desarrollado de balsas lipıdicas ricas encolesterol y esfingolıpidos. Estas balsas se asocian conproteınas caveolina para crear cav�eolas, unasvesıculas de reducido tamano que actuan comotubulos T primitivos e inducen la liberacion de calciohacia el sarcoplasma por parte del retıculosarcopl�asmico.
CONTROL DE LA CONTRACCION DEL MUSCULO LISO
La contraccion del musculo liso depende de las con-centraciones sarcopl�asmicas de calcio y de la disposi-cion de las mol�eculas de miosina II, cuya porcion demeromiosina ligera se acerca al sitio de union a actinadel dominio S1 y lo oculta. Los iones de calcio pasandel retıculo sarcopl�asmico al sarcoplasma a trav�es de lascav�eolas y atraviesan el sarcolema a trav�es de canales decalcio. La calmodulina se une a cuatro iones de Ca++, yel complejo ası formado activa la fosforilacion de lacadena ligera de la miosina reguladora por accion deMLCK, lo que permite la separacion de la meromiosinaligera y el grupo S1; la mol�ecula de miosina II se estira(v. fig. 8.11B) y se une a otras mol�eculas de miosina IIpara crear un filamento grueso. Asimismo, el calcio seune a la caldesmona para descubrir el sitio de union delfilamento delgado, ambos filamentos se deslizan unosobre otro, y tiene lugar la contraccion muscular. Lahidrolisis del ATP es una reaccion lenta en el musculoliso, por lo que la contraccion tiene una duracion pro-longada y requiere una cantidad menor de energıa. Lacalmodulina se inactiva como consecuencia de la dis-minucion de las concentraciones sarcopl�asmicas de cal-cio, al igual que la MLCK. La enzima miosina fosfatasaretira grupos fosfato de la cadena ligera de la miosina,la miosina II se pliega sobre sı misma y el miofilamentogrueso se despolimeriza.
106
8
MUSCULO

CONSIDERACIONES CL�INICAS
LEIOMIOMA
El leiomioma es un tumor benigno de las c�elulas demusculo liso que suele localizarse en los vasossanguıneoso lapareddel tubodigestivo, enparticularen el esofago y el intestino delgado, donde forma
nodulos pequenos de c�elulas entrelazadas demusculo liso. Los leiomiomas gastrointestinalessuelen afectar a adultos de edades comprendidasentre los 30 y los 60 anos y no se consideran motivode preocupacion, a no ser que produzcan dolor o sehipertrofienhastadificultar ladeglucion, obstruir la luzdel tubo digestivo u originar estrangulacion intestinal.Suelen tratarse mediante electrocauterizacion o, encaso necesario, escision quirurgica.
LEIOMIOSARCOMA
El leiomiosarcoma es un tumor maligno infrecuente
de las c�elulas de musculo liso que aparece en lasparedes de los vasos sanguıneos. Suelen presentarun tamano mayor que los leiomiomas, si bien su
dureza es inferior, y pueden presentar �areas
necroticas y hemorr�agicas. Las c�elulas de musculoliso se dividen activamente y dan lugar a fascıculosabundantes. En la mayorıa de los casos, el tumor se
disemina; las met�astasis pueden aparecer entre 10 y
15 anos despu�es de la reseccion del tumor primario;el pronostico a largo plazo de los pacientesafectados es desfavorable.
107
8
MUSCULO
Figura 8.11 A.C�elula de musculo liso relajada y contraıda. B. Activacion de la mol�ecula demiosina del musculo liso. P, fosfato unido ala cadena ligera de lamiosina. (A. Tomado deGartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 182.
B. Modificado de Alberts B, Bray D, Lewis J, et al: Molecular Biology of the Cell. New York, Garland Publishing, 1994.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

9 TEJIDO NERVIOSOEl tejido nervioso, formado por cientos de miles demillones de neuronas que forman una red compleja eintricada de conexiones con otras neu-ronas y con un gran numero de c�elulasno nerviosas, actua como el centro decomunicaciones y la base de datos delorganismo. Este centro de comunica-cion se sustenta en la presencia dereceptores que reciben informacionprocedente del exterior y del interiordel organismo y transmiten estosdatos a los centros encargados de suprocesamiento, que procesan losdatos nuevos y los comparan con lainformacion almacenada en la basede datos, y generan respuestas que seenvıan a los organos efectores quellevar�an a cabo la accion especificada.Unas c�elulas no neurales, las c�elulas de la neuroglia,soportan fısicamente a las neuronas y las mantienenmetabolicamente.
La organizacion anatomica del sistema nervioso esla siguiente:
. El sistema nervioso central (SNC) se compone delenc�efalo y la m�edula espinal.
. El sistema nervioso perif�erico (SNP), formadopor 12 pares de nervios craneales y 31 pares denervios espinales y sus respectivos ganglios,colabora con el sistema nervioso en el desempenode sus numerosas funciones.
Desde el punto de vista funcional, el SNP puededividirse en componentes sensoriales (aferentes),que perciben el estımulo y lo transmiten a centrossuperiores para su procesamiento, y componentesmotores (eferentes), los cuales parten del enc�efalo ola m�edula espinal y conducen el estımulo al organoefector (p. ej., musculo esquel�etico, musculo cardıaco,musculo liso, gl�andula). El componente motor delsistema nervioso se subdivide, a su vez, en:
. Sistema nervioso som�atico, que solamentetransmite impulsos motores a las c�elulas delmusculo esquel�etico del organismo a trav�es de unaunica neurona.
. Sistema nervioso autonomo, que conduceimpulsos motores al musculo cardıaco, el musculo
liso y las gl�andulas a trav�es de unsistema integrado por dos neuronas, en elque un ganglio se interpone entre laneurona preganglionar que procededel SNC y la neurona posganglionarque parte de dicho ganglio. Otrasneuronas, las c�elulas neurogliales,sustentan a las neuronas encargadasde la transmision del impulso(fig. 9.1).
Desarrollo delsistema nervioso
Las citocinas sintetizadas por la noto-corda estimulan el ectodermo supraya-
cente, el cual se diferencia para formar el neuroepi-telio que m�as tarde se engrosar�a para dar lugar ala placa neural. El plegamiento inicial de los m�arge-nes de esta placa origina el surco neural; losm�argenesterminan por fusionarse entre sı para formar un tuboneural cilındrico. El enc�efalo se desarrolla en elextremo rostral del tubo neural, mientras que lam�edula espinal se forma en el extremo caudal. Otrasestructuras del sistema nervioso, como las c�elulas dela neuroglia, las neuronas, el plexo coroideo y elep�endimo, proceden igualmente del tubo neural.Antes de su fusion, una delgada lınea de c�elulas(c�elulas de la cresta neural) procedentes de losm�argenes de la placa neural se aleja de �esta para ori-ginar los siguientes elementos:
. Ganglios sensoriales, ganglios autonomos yneuronas que parten de ellos
. La mayor parte del mes�enquima y sus derivados encabeza y region anterior del cuello
. Odontoblastos
. Melanocitos
. C�elulas cromafines de la m�edula suprarrenal
. C�elulas de la aracnoides y la piamadre
. C�elulas sat�elite de los ganglios perif�ericos
. C�elulas de Schwann
� 2011 Elsevier Espana, S.L. Reservados todos los derechos
TERMINOS CLAVE. Neuronas. Neuroglia. Impulsos nerviosos. Sinapsis. Neurotransmisores. Sistema nervioso
som�atico. Sistema nervioso
autonomo. Meninges
108

CONSIDERACIONES CL�INICAS
El desarrollo del sistema nervioso tiene lugar en unaetapa temprana y es un proceso de grancomplejidad, por lo que pueden producirse
numerosas anomalıas y malformaciones cong�enitasa lo largo de la embriogenia. La espina bıfida es unaanomalıa debida a la fusion incompleta del tubo
neural que permite la salida de lam�edula espinal y las
meninges a trav�es de la zona no fusionada. Laespina bıfida anterior se debe al cierre incompleto
de las v�ertebras. Las vısceras tor�acicas yabdominales pueden presentar malformaciones en
los casos m�as graves.
La fusion deficiente del neuroporo anterior origina
una boveda craneal con un enc�efalo poco
desarrollado, una anomalıa cong�enita que recibe elnombre de anencefalia y tiene consecuenciasmortales.
La migracion anomala de las c�elulas corticalespuede alterar el funcionamiento de lasinterneuronas, lo que da lugar a la epilepsia.
La enfermedad de Hirschsprung o megacolon
cong�enito proviene de la ausencia de migracion de
las c�elulas de la cresta neural hacia la pared del colondistal en formacion. La ausencia del plexo deAuerbach del sistema nervioso ent�erico, encargadode la inervacion del colon distal, lo que originahipertrofia colonica.
109
9
TEJIDONERVIO
SO
Figura 9.1 Comparacion de reflejos som�aticos y viscerales. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.Philadelphia, Saunders, 2007, p 207.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

110
9
TEJIDONERVIO
SO
C�elulas del sistema nervioso
El sistemanervioso se compone de dos grupos celularesdiferenciados. Las neuronas son c�elulas nerviosas fun-cionales; en este grupo se incluyen las c�elulas m�aspequenas (5mm) y m�as grandes (150mm) del orga-nismo; se encargan de transmitir informacion al SNCy conducirla desde este. Las c�elulas de la neurogliaaportan sustento fısico y metabolico a las neuronas.
ESTRUCTURA Y FUNCION DE LAS NEURONAS
Una neurona tıpica est�a formada por un cuerpocelular (pericarion o soma) que consta de un nucleorodeado de citoplasma y dos tipos de proyecciones,varias dendritas y un axon solitario (fig. 9.2).
. Los cuerpos celulares presentan tamanos ymorfologıas variadas, si bien la mayorıa de ellostiende a mostrar una forma poligonal en el SNC yuna morfologıa esf�erica en los ganglios sensoriales.El cuerpo celular alberga al nucleo y diversosorg�anulos, entre los que destacan el retıculoendopl�asmico rugoso (RER) (corpusculo de Nisslen la microscopia optica), un gran aparato de Golgiperinuclear, mitocondrias abundantes, y un sistemabien desarrollado de microtubulos,microfilamentos y neurofilamentos. Losmicrotubulos poseen proteına asociada amicrotubulos 2 (MAP-2). De igual modo, contieneinclusiones, como lipofucsina, una mol�ecularelacionada con la edad que podrıa corresponder alos restos no degradables de la actividad lisosomica;melanina, un pigmento de color marron oscuroque podrıa representar los residuos de la sıntesis dealgunos neurotransmisores (p. ej., noradrenalina ydopamina); gr�anulos secretores, que podrıancontener neurotransmisores; y gotıculas lipıdicas.
. Las dendritas, proyecciones celulares encargadas dela recepcion de estımulos tanto externos comointernos, suelen ramificarse y pueden arborizarsepara recibir estımulos de diverso origen de manerasimult�anea, que transmiten en forma de impulsohacia el cuerpo celular. Una misma neurona sueleposeer varias dendritas, cada una de las cualescontiene org�anulos, aunque carece del aparato deGolgi, en sus regiones proximales. Las proyeccionesse ensanchan en las proximidades del soma y seestrechan conforme se alejan de �el. Losneurofilamentos de las dendritas suelen estar encontacto con microtubulos portadores de proteınasMAP-2. A medida que se ramifican, las dendritasestablecenun grannumero de sinapsis y las dendritasde algunasneuronas formanpequenosmontıculos, oespinas, en su superficie con el fin deaumentar el�areasuperficial disponible para la creacion de sinapsis.
. El cuerpo celular de una neurona posee solamenteun axon que se proyecta desde una region
especializada del cuerpo celular denominada conoaxonal. El axon puede recorrer distancias grandescon el fin de aportar inervacion motora a musculosy gl�andulas. El di�ametro axonal es variable ydetermina la velocidad de conduccion (es decir, lavelocidad se incrementa al hacerlo el di�ametro delaxon). No obstante, el di�ametro es especıfico decada tipo de neurona. El axon solitario puederamificarse en �angulos rectos, formando axonescolaterales, y puede arborizarse al aproximarse a sudiana. Los axones finalizan en terminales axonales(bulbos terminales, pie terminal, botonesterminales) en los que establecen unionessin�apticas (sinapsis) con otras c�elulas.. El cono axonal es una region especializada delcuerpo celular del que parten las dendritas. Elcitoplasma del interior de esta region carece deRER, aparato de Golgi, ribosomas y corpusculosde Nissl, si bien contiene abundantesmicrotubulos y neurofilamentos que podrıanregular el di�ametro axonal.
. Al abandonar el cuerpo celular, el segmentoinicial del axon est�a exento de mielina y sedenomina zona de estimulacion m�axima (spiketrigger), en la que se combinan los impulsosexcitadores e inhibitorios y se evaluan para decidiracerca de su transmision.
. El axoplasma (citoplasma axonal) no presentaRER ni polirribosomas, por lo que el cuerpocelular se ocupa de su sustento. Esta region sıcontiene retıculo endopl�asmico liso (REL),abundantes mitocondrias alargadas,microtubulos conMAP-3 y neurofilamentos en suextremo distal.
. La oligodendroglia del SNC y las c�elulas deSchwann en el SNP forman una vaina demielina (de color blanco) alrededor de algunosaxones. El SNC se divide en sustancia blanca, enla que la mayorıa de los axones est�anmielinizados, y sustancia gris, en la que no loest�an.
. Las sustancias contenidas en el interior delaxoplasma y el cuerpo celular se transportanmediante un proceso conocido como transporteaxonal, que puede darse en dos sentidos:. Transporte anterogrado, encargado de trasladarmaterial como org�anulos, vesıculas, actina,miosina, clatrina y enzimas implicadas en lasıntesis de neurotransmisores en el terminal axonalhacia el pie terminal. El axon utiliza a la proteınamotriz cinesina para llevar a cabo este proceso.
. Transporte retrogrado de material, comomonomeros y dımeros de tubulina, subunidadesde neurofilamentos, enzimas, virus y mol�eculas adegradar, hacia el soma. El axon emplea laproteına motriz dineına para trasladar estosmateriales.

CONSIDERACIONES CL�INICAS
Algunos virus, como el virus herpes simple y el virusde la rabia, aprovechan el transporte axonalretrogrado para diseminarse de una neurona a otradentro de una cadena. De manera similar sediseminan algunas toxinas, como la producida por
Clostridium tetani, que ocasiona t�etanos, desde laperiferia hacia el SNC.
La mayorıa de los tumores intracraneales son deorigen neuroglial; de manera infrecuente provienende neuronas pertenecientes al SNC.
Entre los tumores neurogliales figuran losoligodendrogliomas benignos y losastrocitomas malignos mortales. Otros tumoresintracraneales procedentes del tejido conjuntivodel sistema nervioso son el fibroma benigno y elsarcoma maligno. El neuroblastoma, un tumorextremadamente maligno que afecta en su mayorıaa lactantes y ninos de corta edad, es un tumor del
SNP localizado en el seno de la gl�andulasuprarrenal.
111
9
TEJIDONERVIO
SO
Figura 9.2 Esquema de la ultraestructura del cuerpo celular de una neurona. (Tomado de Gartner LP, Hiatt JL: Color Textbook
of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 190.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

CLASIFICACION DE LAS NEURONAS
Las neuronas pueden dividirse en tres grupos conarreglo a su morfologıa y la organizacion de susproyecciones (fig. 9.3):
. Las neuronas unipolares (neuronasseudounipolares) se localizan en el ganglio de laraız dorsal y algunos ganglios de los nervioscraneales. Poseen solamente una proyeccion; noobstante, esa unica proyeccion se bifurca en unarama perif�erica que continua hasta la c�elula a la queinerva y una rama central que accede al SNC. Larama perif�erica da lugar a terminaciones receptorassemejantes a una dendrita y su funcion es receptora.El impulso pasa al proceso central sin atravesar elcuerpo som�atico.
. Las neuronas bipolares se encuentran en elepitelio olfativo y los ganglios del nerviovestibulococlear. Poseen dos proyecciones:una dendrita y un axon.
. Las neuronas multipolares son ubicuas,generalmente se trata de neuronas motoras, y selocalizan en lam�edula espinal y la corteza cerebral ycerebelosa.
Se distinguen tres clases de neuronas segun sufuncion:
. Las neuronas sensoriales (aferentes) sonestimuladas por sus receptores dendrıticos en laperiferia para responder a los estımulos ambientalesexternos, y en el interior del organismo pararesponder a los estımulos ambientales internos ytransmitir la informacion al SNC, donde ser�aprocesada.
. Las neuronas motoras (eferentes) parten del SNCy transmiten sus impulsos a otras neuronas,musculos y gl�andulas.
. Las interneuronas, exclusivas del SNC, actuancomo intermediarias entre las neuronas sensorialesy las neuronas motoras; establecen e integran lasactividades de los distintos circuitos neuronales.
CELULAS NEUROGLIALES
Las c�elulas neurogliales (fig. 9.4) son, al menos,10 veces m�as abundantes que las neuronas; a pesarde carecer de la capacidad de transmision de losimpulsos nerviosos, desempenan la funcion clave desostener y proteger a las neuronas al envolver su soma,dendritas y axones. A diferencia de las neuronas, lasc�elulas neurogliales pueden sufrir divisiones celulares.Las c�elulas neurogliales que forman parte del SNC sonlos oligodendrocitos, la microglia, los astrocitos y las
c�elulas ependimarias; las c�elulas de Schwann y las c�e-lulas de la neuroglia intervienen en el SNP.
. Los oligodendrocitos se dividen en dos grupos:. Oligodendrocitos interfasciculares, quesintetizan mielina para aislar los axonespertenecientes al SNC. Uno de estosoligodendrocitos puede envolver varios axonesen mielina.
. Oligodendrocitos sat�elite, que rodean el somade las neuronas grandes, a las que podrıanproteger de contactos indeseados.
. Las c�elulas de la microglia son unas c�elulaspequenas que proceden de la m�edula osea y actuancomo macrofagos, ya que se incluyen en el sistemafagocıtico mononuclear. Residen en el SNC, en elque fagocitan residuos y c�elulas danadas y confierenproteccion frente a la infeccion por virus ymicroorganismos y la invasion tumoral. Asimismo,actuan como c�elulas presentadoras de antıgenos ysecretan citocinas.
. Se han identificado dos tipos de astrocitos:astrocitos protopl�asmicos, localizados en lasustancia gris del SNC, y astrocitos fibrosos, tıpicosde la sustancia blanca. Sin embargo, se ha propuestoque solamente existirıa un tipo de astrocito, cuyapresencia en dos ambientes diferentes darıa lugar ala expresion de caracterısticas dispares. Ambos tiposposeen filamentos intermedios, los cuales sedistinguen por la proteına acıdica fibrilar glial,exclusiva de estas c�elulas. Los astrocitos retiranmol�eculas acumuladas, como iones yneurotransmisores, ası como sus restos metabolicosen el �area en la que se encuentran. Otras funcionesde los astrocitos serıan la reparacion de danos en elSNC, en el que crean tejido cicatricial formadosolamente por c�elulas; la liberacion de glucosa paranutrir a las neuronas de la corteza cerebral y laparticipacion en el desarrollo de la barrerahematoencef�alica (BHE) junto a las c�elulasendoteliales de los vasos sanguıneos.. Los astrocitos protopl�asmicos poseen pedicelos(pies vasculares) que est�an en contacto con losvasos sanguıneos. Otros astrocitos de estesubgrupo situados junto a la piamadre o lam�edula espinal presentan pedıculos que seconectan entre sı para crear una delgada capaadosada a la piamadre, la membrana piaglial.
. Los astrocitos fibrosos est�an dotados deproyecciones prolongadas que se asocian a losvasos sanguıneos y la piamadre sin llegar a entraren contacto con ellos debido a la presencia de lal�amina basal.
112
9
TEJIDONERVIO
SO

113
9
TEJIDONERVIO
SO
Figura 9.3 Tipos de neuronas. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 189.)
Figura 9.4 Tipos de c�elulas neurogliales. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders,
2007, p 193.)�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

. Las c�elulas ependimarias son c�elulas cubicas querevisten los ventrıculos cerebrales y el canal centralde la m�edula espinal. Intervienen en la formaciondel plexo coroideo, la estructura encargada desintetizar el lıquido cefalorraquıdeo (LCR).Algunas c�elulas ependimarias poseen cilios quepotencian la circulacion del LCR, mientras queotras, los tanicitos, participan en la transferencia deLCR a las c�elulas neurosecretoras del hipot�alamo.
. Las c�elulas de Schwann proceden de c�elulas de lacresta neural y, a pesar de considerarse c�elulasneurogliales, est�an restringidas al SNP (fig. 9.5). Demanera similar a los oligodendrocitos, las c�elulas deSchwann forman una vaina mielinizada odesmielinizada alrededor de los axones que los aısladel exterior; no obstante, a diferencia de aquellas,una sola c�elula de Schwann puede mielinizar unsolo axon, aunque puede envolver varios axones nomielinizados. La vaina de mielina corresponde alplasmalema de la c�elula de Schwann que se enroscasobre el axon hasta 50 veces. Las c�elulas de Schwannse alinean unas junto a otras y cada una de ellasrecubren una seccion corta del axon con sumembrana plasm�atica. La region del axon rodeadapor una c�elula de Schwann recibe el nombre desegmento internodular. La region comprendidaentre dos segmentos internodulares no est�amielinizada y se conoce como nodulo de Ranvier.Cada c�elula de Schwann posee su propia l�aminabasal, por lo que la porcion del axon localizada endicho nodo est�a cubierta por interdigitaciones de lasproyecciones de las c�elulas de Schwann y por lal�amina basal de estas c�elulas; por lo tanto, el axonno est�a expuesto de forma directa a su entorno. Losoligodendrocitos no forman proyecciones en losnodulos de Ranvier, sino que esa region es ocupadapor las proyecciones de un astrocito (fig. 9.6).. Los axones de muchas neuronas est�anmielinizados en el adulto, si bien no todos poseenuna cubierta demielina a lo largo del desarrollo.La mielinizacion de los nervios sensoriales noconcluye hasta varios meses despu�es delnacimiento. En el SNC, los axones de algunas delas fibras carecen de mielina durante los primerosanos de vida.
. Lamielinizacion es un proceso complejo que nose conoce bien aun. La membrana de la c�elula deSchwann (u oligodendrocitos en el SNC) seenrolla alrededor del axon y el citoplasma seexprime hacia el interior del soma. La cara internadel plasmalema se aproxima a la cara interna del
plasmalema y la cara externa se acerca a la caraexterna, una relacion estrecha que se repite encada vuelta de la espiral.
. En la microscopia electronica, aparece en lamembrana enrollada una lınea densa de coloroscuro, la lınea densa mayor, que indica la zonade contacto entre las dos caras citopl�asmicas de lamembrana plasm�atica de la c�elula de Schwann. Laregion en la que se adosan las superficies externasdel plasmalema corresponde a una lınea demenor grosor llamada lınea intraperıodo. Lalınea densa mayor y la lınea intraperıodo sealternan entre sı. En las im�agenes de granresolucion se observa la existencia de espaciosangostos dentro de la lınea intraperıodo, losintersticios intraperıodo; se trata de un espacioextracelular muy pequeno que permite lacomunicacion entre el axon y el ambiente externoa la vaina de mielina. Por supuesto, tan solo losiones de pequeno tamano pueden atravesardichos intersticios.
. Algunas regiones de la vaina demielina conservancitoplasma residual, que puede visualizarse enforma de hendiduras conocidas como incisionesde Schmidt-Lanterman.
. La membrana de la c�elula de Schwann que forma lavaina de mielina contiene numerosasglucoproteınas y esfingomielina, ası como dosproteınas clave: proteına cero de mielina (MPZ) yproteına b�asica de mielina (MBP). Adem�as decolaborar en la sıntesis de mielina, la MPZ ayuda aestabilizar la vaina de mielina. Se cree que MBPtambi�en ayuda a mantener la estabilidad de dichavaina.MPZno aparece en lamielina del SNC, ya quela proteına proteolipıdica (PLP) asume susfunciones.
. Las caras externas de las membranas celulares(lıneas intraperıodo) se mantienen unidasmediante uniones estrechas que no solamentecontienen las proteınas tıpicas, claudinas y lasproteınas de la zonula oclusiva, sino que tambi�enpresentan conexina 32 (Cx32).
. La region de la vaina demielina en la que el extremode la espiral demielina se encuentram�as alejado delaxolema (membrana axonal) se denominamesaxon externo.
. La region de la vaina demielina en la que el extremode la espiral se halla m�as cercano al axolema sellama mesaxon interno.. El intersticio intraperıodo comprende delmesaxon externo al mesaxon interno.
114
9
TEJIDONERVIO
SO

115
9
TEJIDONERVIO
SO
CONSIDERACIONES CL�INICAS
La esclerosis multiple, una alteracion de lamielinizacion del SNC, es una entidad frecuente. Afectaa sujetos de edades comprendidas entre 15 y 45 anos
de edad y es 1,5 veces m�as prevalente en la mujer. Sedesmielinizan el cerebelo, la sustancia blanca
encef�alica, m�edula espinal y nervios craneales yraquıdeos. Se producen episodios de inflamacionmultifocal acompanados de edema y desmielinizacionde los axones del SNC. Cada uno de estos episodiospuede ocasionar un deterioro intenso, neoplasiamaligna o ambos en los nervios afectados; en funcionde las regiones afectadas, la muerte puede sobreveniren el plazo de unos meses. Estas crisis se siguen deperıodos de remision que pueden prolongarse varios
meses o d�ecadas. Cada episodio merma la vitalidad delpaciente. Se cree que la esclerosis multiple representauna enfermedad autoinmunitaria debida a la presenciade un microorganismo. El tratamiento de eleccion
consiste en f�armacos inmunosupresores combinadoscon corticoesteroides y antiinflamatorios.
La radioterapia en el enc�efalo o la m�edula espinalprovoca la desmielinizacion de los nervios que seencuentran en el campo de radiacion. Por otra parte, las
mol�eculas citotoxicas utilizadas en la quimioterapiainducen desmielinizacion de los axones del sistemanervioso, lo que da lugar a alteraciones neurologicas.
El sındrome de Guillain-Barr�e es un trastornoinmunitario derivado de una infeccion respiratoria ogastrointestinal reciente. Produce inflamacion y
desmielinizacion de los nervios perif�ericos, lo que setraduce en debilidad muscular en las extremidades. Laenfermedad se inicia en un plazo breve y alcanza su
m�axima intensidad despu�es de varias semanas. Esconveniente elaborar un diagnostico precoz e instaurarun tratamiento con inmunoglobulinas y fisioterapia.
Figura 9.5 Esquema de la estructura fina de una fibra nerviosa
mielinizada y su c�elula de Schwann. (Tomado de Gartner LP,
Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders,2007, p 192.)
Figura 9.6 Diagrama de la estructura de la mielina en unnodulo de Ranvier de axones del SNC y SNP (recuadro).
(Tomado de Gartner LP, Hiatt JL: Color Textbook of
Histology, 3rd ed. Philadelphia, Saunders, 2007, p 197.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Generacion y conduccionde los impulsos nerviosos
Las membranas de las c�elulas est�an polarizadas el�ec-tricamente de modo que la cara interna es menospositiva que la externa debido a la diferencia entrelas concentraciones ionicas; concretamente, la con-centracion de iones Na+ y Cl- es mayor fuera de lac�elula que en su interior, mientras que la concentra-cion de iones K+ es m�as alta dentro de la c�elula quefuera de ella. Este rasgo de las membranas es m�asacusado en las c�elulas de mamıfero, en las que elpotencial de reposo de las neuronas grandes es de–90mV, aunque es menos negativo en las neuronasm�as pequenas y las fibras musculares (figs. 9.7 y 9.8).Las neuronas se comunican modulando el potencialde membrana mediante su despolarizacion y repo-larizacion,demodoque una onda de despolarizacionse transmite a lo largo de las proyecciones de la neu-rona hasta llegar a otra neurona, c�elula muscular oc�elula de una gl�andula a trav�es de una union especia-lizada conocida como sinapsis. La membrana plas-m�atica del axon posee, al menos, estos tres canalesionicos y una bomba de Na+-K+:
. Canales de escape de K+, que permiten la salida delos iones K+ de la c�elula siguiendo un gradiente deconcentracion de potasio que origina un aumentode las cargas positivas a lo largo de la cara externa dela membrana celular. El canal de escape de K+
establece el potencial de membrana de reposo,con la intervencion minoritaria de las bombasde Na+-K+.
. Bombas deNa+-K+ de lamembrana plasm�atica, queextraen tres iones Na+ por cada dos iones K+ quepasan al citoplasma.
. Canales de Na+con compuerta de voltaje, cuyaapertura permite la entrada de iones Na+ al citosol.Estos canales se abren como consecuencia de la
despolarizacion de la membrana, si bien se trata deuna configuracion inestable que da lugar a lainactivacion del canal (es decir, su cierre hasta lasiguiente repolarizacion de membrana al potencialde reposo). Esta capacidad se atribuye a la existenciade dos compuertas, una de las cuales se encuentra enla superficie extracitopl�asmica, la compuerta deactivacion, y otra lo hace en la superficiecitopl�asmica, la compuerta de inactivacion.Aunque la compuerta de activacion permaneceabierta debido al cambio de voltaje, la compuerta deinactivacion se cierra para impedir el paso de losiones Na+ a trav�es del canal; este estado recibe elnombre de perıodo refractario. Los canales de Na+
con compuerta de voltaje pueden encontrarse:. Cerrados (compuerta de activacion cerrada,compuerta de inactivacion abierta).
. Abiertos (compuerta de activacion abierta,compuerta de inactivacion abierta).
. Inactivados (perıodo refractario: compuerta deactivacion abierta, compuerta de inactivacioncerrada).
. Canales de K+con compuerta de voltaje, que seabren –demaneramuy lenta– cuando lamembranase despolariza, lo que posibilita la salida de iones K+
de la neurona. Estos canales se cierran comoconsecuencia de la repolarizacion de la membrana.
Por lo general, la neurona recibe en estımulo en lazona de estimulacion m�axima del axon. Se modifica,con ello, el potencial demembrana en esa �area, y tienelugar la siguiente secuencia de acontecimientos:
1. Los canales de Na+ con compuerta de voltaje seabren en la zona de estimulacion m�axima, losiones Na+ pasan al axon y las elevadasconcentraciones de iones Na+ en la cara interna dela membrana invierten el potencial de membrana,que se despolariza.
116
9
TEJIDONERVIO
SO
Continua en la pag. 118

117
9
TEJIDONERVIO
SO
Figura 9.8 Esquema de la propagacion de un potencial de accion en un axon no mielinizado (A) y mielinizado (B). (Tomado
de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 200.)
Figura 9.7 Esquema del establecimiento del potencial de reposo en una neurona tıpica. Los canales de escape de K+ son m�as
numerosos que los de Na+ y Cl-; en consecuencia, el numero de iones K+ que salen de la c�elula es mayor que el de iones Na+ y Cl-
que entran en la misma. Los iones positivos son m�as abundantes en el espacio extracitopl�asmico que en el citosol, de modo que se
crea una diferencia de potencial en la membrana. No se representan los canales ionicos y las bombas de iones que no intervienendirectamente en la creacion del potencial de reposo. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia,
Saunders, 2007, p 199.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

2. Los canales de Na+con compuerta de voltaje quese habıan abierto se inactivan durante unos 2 ms.Los canales de K+con compuerta de voltaje seabren, los iones K+ abandonan el axon en la zonade estimulacionm�axima, y la region de esta zona serepolariza e, incluso, hiperpolariza durante unafraccion de milisegundo (fig. 9.9).
3. Muchos de los iones Na+ que pasaron al axon en laetapa 1 fluyen en ambas direcciones y podrıanprovocar la despolarizacion de las regionesadyacentes del axon. Esta onda de despolarizacionse diseminarıa en ambos sentidos hacia y desde elsoma; no obstante, los canales de Na+concompuerta de voltaje hacia el soma se encuentranen el perıodo refractario y permanecen cerrados.La propagacion del impulso (onda dedespolarizacion) no puede avanzar hacia el soma(propagacion retrograda), pero sı alejarse de �elhacia las terminales axonales.
4. Las variaciones del voltaje de membrana descritasse denominan potencial de accion; se trata de unproceso de todo o nada que puede repetirse hasta1.000 veces por segundo.
SINAPSIS
Las sinapsis, uniones especializadas a trav�es de lascuales se establece la comunicacion entre unas c�elulasnerviosas con otras o con c�elulas efectoras (comoc�elulas musculares o gl�andulas), pueden dividirse endos tipos: el�ectricas y quımicas. Las primeras corres-ponden a uniones gap, aunque apenas aparecen enmamıferos salvo en algunas regiones del SNC. Lassegundas requieren la liberacion de una sustancianeurotransmisora a un espacio intercelular adaptadollamado hendidura sin�aptica, la cual se localiza entreel plasmalema del extremo terminal de un axon(membrana presin�aptica) y una region especializadade la membrana plasm�atica (membrana postsin�ap-tica) de otra neurona, c�elula muscular o c�elula de unagl�andula. En la tabla 9.1 se enumeran distintos tiposde sinapsis, que se representan en la figura 9.10. Elneurotransmisor secretado al espacio presin�aptico seune a receptores de lamembrana postsin�aptica, lo queinduce la apertura de los canales ionicos asociados areceptores que, a su vez, suscita el movimiento deiones a trav�es de la luz del canal. Si el trasiego de ionesorigina una:
. Despolarizacion de suficiente magnitud de lamembrana postsin�aptica para iniciar un potencialde accion, el estımulo se denomina potencialpostsin�aptico excitador, o bien
. Hiperpolarizacion de la membrana postsin�aptica,lo que no desencadena un potencial de accion; el
estımulo recibe el nombre de potencialpostsin�aptico inhibitorio.
El terminal presin�aptico, el pie terminal, alberga lasvesıculasdelREL,ası comounasvesıculaspequenas conneurotransmisores denominadas vesıculas sin�apticas,de40 a60mmdedi�ametro. Estas vesıculas se agregan enla periferia de la membrana presin�aptica y en las pro-ximidadesdeunas regiones conocidas como sitios acti-vos, ya que en ellas tiene lugar la fusion de las vesıculascon la membrana sin�aptica, con la consiguiente libera-cion de sus contenidos en la hendidura sin�aptica. Lasvesıculas sin�apticas localizadas en el sitio activo est�anpreparadas para liberar su contenido, mientras que lassituadas alrededor del mismo se reservan mediante:
. Las proteınas transmembrana de la vesıculasinapsina I y sinapsina II, que inmovilizan a lasvesıculas al anclarlas a filamentos de actina.
. Fosforilacion de ambas proteınas, que liberan a lasvesıculas sin�apticas de dichos filamentos y permitensu traslado al sitio activo.
La fusion de las vesıculas sin�apticas del sitio activocon la membrana presin�aptica se ve facilitada por:
. Entrada de iones Ca++ al boton terminal a trav�es decanales de Ca++ abiertos como consecuencia de lallegada del potencial de accion al plasmalema delboton terminal.
. Presencia de iones Ca++ en el citoplasma que hacenposible la interaccion mutua de las proteınastransmembrana de la vesıcula sin�aptica y lamembrana presin�aptica rab3A, sinaptotagmina,sinaptobrevina, sintaxina, SNAP-25 (proteına deunion para la proteına de fusion sensible aN-etilmaleimida soluble-25), y sinaptofisina, conel fin de completar el proceso de fusion y permitir laliberacion de mol�eculas neurotransmisoras a lahendidura sin�aptica.
. La membrana de la vesıcula que se incorporo a lamembrana presin�aptica se recupera mediante unproceso de endocitosismediado por la envoltura declatrina, en el que colaboran proteınas integralescomo la proteına de la envoltura de las vesıculasAP-2 y la sinaptotagmina. El fragmento demembrana recuperado se transporta de nuevo alREL, en el que ser�a reciclado.
La membrana postsin�aptica, separada de la pre-sin�aptica por el intersticio sin�aptico, presenta un grosormayor que la membrana de la c�elula postsin�aptica yalberga receptores para los neurotransmisores libera-dos en el sitio activo del pie terminal de la neuronapresin�aptica. El grosor de la membrana postsin�apticasuele reflejar su respuesta al neurotransmisor secretado.
118
9
TEJIDONERVIO
SO

CONSIDERACIONES CL�INICAS
La bacteria Clostridium botulinum secreta la toxinabotulınica, una neurotoxina con efecto mortal enconcentraciones muy bajas (DL50 en administracionintravenosa de alrededor de 1ng/kg). Aunque latoxina es sensible al calor y se desnaturaliza a 60 �C,las esporas bacterianas continuan siendo viables ypueden germinar en condiciones de anaerobiosis.Los microorganismos vegetativos fabrican la toxina;pueden subsistir en alimentos sometidos a unamanipulacion incorrecta o en latas danadas. Latoxina es una proteasa que escinde especıficamenteuna de las proteınas de fusion (SNAP-25, sintaxina o
sinaptobrevina) en las uniones mioneuronales. Lapresencia de fragmentos de estas proteınas impide
la fusion de las vesıculas sin�apticas de la membrana
presin�aptica y bloquea la liberacion de acetilcolina,
lo que ocasiona par�alisis fl�acida de la musculatura
afectada. En general, la muerte se debe a la par�alisisde los musculos respiratorios; no obstante, la toxinaactua durante varios dıas y, de ser reconocida atiempo, se puede evitar la muerte mediante laventilacion artificial y la administracion deantitoxinas botulınicas.
119
9
TEJIDONERVIO
SO
Tabla 9.1 TIPOS DE SINAPSIS ENTRE DOS NEURONAS
Tipo de sinapsisRegiones neuronalesimplicadas
Axodendrıtica Entre axon y dendrita
Axosom�atica Entre axon y soma
Axoaxonica Entre dos axonesDendrodendrıtica Entre dos dendritas
Figura 9.9 Esquema de la propagacion
de un potencial de accion en un axon no
mielinizado (A) y mielinizado (B).(Tomado de Gartner LP, Hiatt JL: Color
Textbook of Histology, 3rd ed. Philadelphia,
Saunders, 2007, p 200.)
Figura 9.10 Esquema de los tiposde sinapsis. (Tomado de Gartner LP,
Hiatt JL: Color Textbook of Histology, 3rd ed.
Philadelphia, Saunders, 2007, p 201.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Neurotransmisores (mol�eculas de senalizacion)
La interaccion de los neurotransmisores con losreceptores situados en las c�elulas diana pone enmarcha una respuesta especıfica por parte de estas.Los receptores pueden ser:
. De accion r�apida (el proceso requiere � 1 ms),acoplados a los canales ionicos; las mol�eculas desenalizacion que los activan se conocen comoneurotransmisores.
. De accion lenta (el proceso puede prolongarsedurante varios minutos), acoplados a proteınas G;las mol�eculas de senalizacion (que activan unsistema de segundosmensajeros) reciben el nombrede neuromoduladores o neurohormonas.
Se conocen m�as de 100neurotransmisores/neuro-hormonas, que pueden asignarse a tres grupos dife-rentes (tabla 9.2):
. Transmisores de pequeno tamano molecular(acetilcolina, amino�acidos, aminas biog�enicas)
. Neurop�eptidos (p�eptidos opioides, p�eptidosgastrointestinales, hormonas liberadorashipotal�amicas, hormonas almacenadas en laneurohipofisis)
. Gases (oxido nıtrico y monoxido de carbono)
Los neurotransmisores pueden inducir distintasrespuestas en condiciones diferentes y la configura-cion del receptor postsin�aptico puede determinar suefecto en la c�elula postsin�aptica. Habitualmente, lacomunicacion interneuronal en las sinapsis precisade la intervencion de varios neurotransmisores (trans-mision de volumen), en particular en las c�elulas delenc�efalo, en las que los neurotransmisores se encuen-tran en el lıquido intercelular y provocan la activacionde grupos de c�elulas portadoras de los receptores co-rrespondientes en lugar de activar una sola c�elula. Latransmision de volumen es un proceso de accionlenta que podrıa relacionarse con el estado de vigilia,la funcion autonoma, la sensibilidad al dolor y elestado de �animo. Por el contrario, la comunicacionsin�aptica es un proceso de accion r�apida.
Nervios perif�ericos
Los nervios perif�ericos se componen de fibras nervio-sas sensoriales y motoras recubiertas por revesti-
mientos que permiten su visualizacion a simple vista.Los haces ası formados, llamados fascıculos, mues-tran una coloracion blanquecina debido a la pre-sencia de mielina en muchas de las fibras que losintegran.
REVESTIMIENTOS DE TEJIDO CONJUNTIVO
Los nervios del fascıculo se rodean de tres capas detejido conjuntivo (fig. 9.11):
. Epineuro, la envoltura m�as externa, recubre porcompleto al nervio y se continua con la duramadredel SNC. Es m�as grueso en el origen del nervio quesale del SNC y se adelgaza conforme se ramifica elnervio hasta desaparecer. Est�a formado por tejidoconjuntivo colagenoso denso irregular que seentremezcla con fibras gruesas de elastina.La organizacion de las fibras de col�ageno de lavaina pretende impedir su distension.
. Perineuro, la capa media de los revestimientosde tejido conjuntivo, rodea a cada fascıculonervioso. Se compone de una delgada l�aminade tejido conjuntivo irregular denso en el queaparecen escasas fibras de col�ageno y fibrasel�asticas. La superficie interna del perineuro serecubre de capas de c�elulas epitelioides y unal�amina basal que separa el compartimentoneuronal del tejido conjuntivo.
. Endoneuro, la capa m�as interna, envuelve acada fibra nerviosa. Est�a en contacto con la l�aminabasal de las c�elulas de Schwann, de modo que seaısla del perineuro y dichas c�elulas. En lasproximidades del extremo de la fibra, secompone solamente de algunas fibras de col�agenode tipo III.
CLASIFICACION FUNCIONAL DE LOS NERVIOS
Los nervios se componen de fibras sensoriales omotoras o de ambas. Los primeros, llamados fibrasnerviosas aferentes, conducen senales nerviosasdesde los receptores sensoriales hasta el SNC parasu procesamiento. Las ultimas, las fibras nerviosaseferentes, parten del SNC y transmiten impulsosmotores hacia los organos efectores. Los nervios mix-tos son los m�as frecuentes y est�an formados por fibrasnerviosas aferentes (fibras sensoriales) y eferentes(fibras motoras).
120
9
TEJIDONERVIO
SO

CONSIDERACIONES CL�INICAS
La corea de Huntington es una enfermedad
gen�etica que comienza con dolor articular, seguidode sacudidas de las articulaciones de lasextremidades. Estas manifestaciones evolucionandando lugar a espasmos de las articulaciones, condeformaciones acompanadas de demencia ydisfuncionmotora. La enfermedad hace su aparicion
entre la tercera y la cuarta d�ecada de vida. Se
atribuye a la desaparicion de las c�elulas encargadas
de sintetizar el �acido g-aminobutırico (GABA), unneurotransmisor inhibidor. La demencia se ha
vinculado con la p�erdida de las c�elulas que secretanacetilcolina.
La enfermedad de Parkinson, la segunda
enfermedad neurodegenerativa m�as frecuente, secaracteriza por el temblor de reposo, labradicinesia, la rigidez y la facies similar a una
m�ascara. Esta entidad se debe a la desaparicion de
las neuronas dopamin�ergicas de la sustancia negra,lo cual se traduce en la ausencia de dopamina en el
enc�efalo. Se han propuesto y estudiado distintostratamientos, aunque la mayorıa de ellos solamenteaporta un alivio temporal sin evitar la muerte de las
neuronas dopamin�ergicas. Se est�a investigando el
trasplante de c�elulas modificadas mediante
ingenierıa gen�etica para secretar dopamina ycapaces de restablecer conexiones con ciertas
c�elulas del enc�efalo que requieren dopamina.Un tratamiento utilizado en la actualidad, laestimulacion cerebral profunda (un tratamientobasado en un dispositivo similar a un marcapasos),supone la implantacion de electrodos en el
t�alamo y el globo p�alido con la finalidad dereducir la rigidez y el temblor y potenciar elequilibrio.
121
9
TEJIDONERVIO
SOTabla 9.2 NEUROTRANSMISORES FRECUENTES Y FUNCIONES INDUCIDAS POR SU RECEPTOR
Neurotransmisor Grupo de compuestos Funcion
Acetilcolina Mol�ecula transmisora pequena; no proviene deamino�acidos
Uniones mioneuronales, todas las sinapsisparasimp�aticas y las sinapsis simp�aticas
preganglionares
Noradrenalina Mol�ecula transmisora pequena; amina biog�enica;
catecolamina
Sinapsis simp�aticas posganglionares (salvo en
gl�andulas sudorıparas ecrinas)Acido glut�amico Mol�ecula transmisora pequena; amino�acido Sensorial presin�aptico y corteza; neurotransmisor
excitador m�as frecuente en el SNC
GABA Mol�ecula transmisora pequena; amino�acido Neurotransmisor inhibidor m�as frecuente en el
SNCDopamina Mol�ecula transmisora pequena; amina biog�enica;
catecolamina
Ganglios basales del SNC; inhibidor o excitador
segun el receptor
Serotonina Mol�ecula transmisora pequena; amina biog�enica Inhibicion del dolor; control del estado de �animo;sueno
Glicina Mol�ecula transmisora pequena; amino�acido Tallo encef�alico y m�edula espinal; inhibidor
Endorfinas Neurop�eptido; p�eptido opioide Analg�esica; ¿inhibicion de la transmision del dolor?
Encefalinas Neurop�eptido; p�eptido opioide Analg�esica; ¿inhibicion de la transmision del dolor?
Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 204.
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

VELOCIDAD DE CONDUCCION
La velocidad de conduccion nerviosa es directa-mente proporcional al grado de mielinizacion(tabla 9.3). Los iones solamente pueden atravesar lamembrana plasm�atica en los nodulos de Ranvier enlos nervios mielinizados, ya que la vaina de mielinapresente en los internodulos impide el intercambioionico en la membrana y los canales de Na+ con com-puerta de voltaje se concentran en dichos nodos. Lospotenciales de accion viajan de un nodo a otro en unproceso denominado conduccion saltatoria. Lasfibras no mielinizadas, que tan solo est�an rodeadaspor una capa de plasmalema de las c�elulas deSchwann, no pueden impedir la salida de iones Na+
a favor de gradiente y los canales de Na+ con com-puerta de voltaje se distribuyen en la membrana plas-m�atica axonal. El proceso de conduccion, conocidocomo conduccion continua, esm�as lento y se asocia aun gasto mayor de energıa.
Sistemas nerviosos motor som�aticoy autonomo
La musculatura esquel�etica recibe su inervacionmotora som�atica a trav�es de neuronas eferentes indi-viduales cuyos cuerpos celulares pertenecen al SNC. Elmusculo liso, el musculo cardıaco y las gl�andulas reci-ben inervacion motora autonoma por medio de unacadena bineuronal en la que el soma de la primeraneurona se halla en el SNC y el de la segunda lo haceen un ganglio autonomo del SNP.
. El sistema nervioso motor som�atico (fig. 9.12) secompone denerviosmotores raquıdeos que partendel asa ventral de la m�edula espinal y nerviosmotores craneales que inervan musculosesquel�eticos a partir de nucleos motores de ciertosnervios craneales. Al salir del SNC, las neuronasmotoras raquıdeas viajan en los nervios raquıdeoshasta el musculo, en el que hacen sinapsis con laplaca terminal motora.
. El sistema nervioso autonomo (v. fig. 9.12) esun sistema motor involuntario que inerva almusculo liso, el musculo cardıaco y las gl�andulas.A diferencia del anterior, el sistema nerviosoautonomo utiliza dos neuronas para alcanzar a losorganos efectores. La primera neurona motora de lacadena, la neurona preganglionar, sale del SNC ysu axon se prolonga hasta un ganglio autonomoque no pertenece al dicho sistema y en el queestablece sinapsis con cuerpos celularesmultipolares de neuronas posganglionareslocalizadas en dicho ganglio. Los axones de lasneuronas posganglionares abandonan el ganglioy viajan hasta el organo efector (musculo liso,musculo cardıaco, gl�andula). Las sinapsisposganglionares de los organos efectores sonm�as generalizadas que las del sistema nerviososom�atico, puesto que el neurotransmisor sedistribuye en un �area m�as amplia y su efecto esm�as extenso. Por otra parte, los musculos que seactivan para contraerse pueden transmitir laestimulacion a otros musculos adyacentes a trav�esde las uniones gap.
122
9
TEJIDONERVIO
SO
Tabla 9.3 CLASIFICACION DE LAS FIBRAS NERVIOSAS PERIFERICAS
Grupo de fibras Di�ametro (mm)Velocidad deconduccion (m/s) Funcion
Fibras de tipo A:
mielinizacion
intensa
1-20 15-120 Fibras som�aticas eferentes de gran velocidad; fibras que
registran dolor agudo, temperatura, contacto, presion y
propiocepcion
Fibras de tipo B:mielinizacion
menos
intensa
1-3 3-15 Fibras de velocidad moderada; fibras viscerales aferentes;fibras autonomas preganglionares
Fibras de tipo C:ausencia
de mielinizacion
0,5-1,5 0,5-2 Fibras de velocidad baja; fibras autonomas posganglionares;dolor cronico
Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 206.

123
9
TEJIDONERVIO
SO
Figura 9.11 Estructura de un haz nervioso. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia,
Saunders, 2007, p 205.)
Figura 9.12 Comparacion de reflejos som�aticos y viscerales. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology,
3rd ed. Philadelphia, Saunders, 2007, p 207.)�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

La inervacion autonoma puede subdividirse en dosgrupos diferentes desde el punto de vista funcional: elsistema nervioso simp�atico y el sistema nervioso para-simp�atico. En t�erminos generales, se puede afirmarque el sistema nervioso simp�atico desempena unafuncion vasoconstrictora, mientras que el parasimp�a-tico lleva a cabo funciones secretoras y motoras.
. El sistema nervioso simp�atico prepara alorganismo para la respuesta de lucha o huida alralentizar la actividad de las vısceras; dilatar laspupilas; aumentar la presion arterial, la frecuenciacardıaca y la frecuencia respiratoria; e intensificarla irrigacion de la musculatura esquel�etica(fig. 9.13).
. El sistema nervioso parasimp�atico prepara alorganismo para el descanso y la digestion al potenciarla actividad visceral; contraer las pupilas; reducir lapresion arterial, la frecuencia cardıaca y la frecuenciarespiratoria; y reducir la irrigacion de los musculosesquel�eticos (v. fig. 9.13).
El neurotransmisor que actua entre las neuronaspreganglionar y posganglionar de los sistemas ner-vioso simp�atico y parasimp�atico es la acetilcolina,que tambi�en actua entre la neurona posganglionar yel organo efector en el sistema nervioso parasimp�a-tico. El neurotransmisor presente entre la neurona pos-ganglionar y el organo efector en el sistema nerviososimp�atico es la noradrenalina.
Ganglios
Los ganglios son acumulaciones de cuerpos celularesde neuronas que se localizan fuera del SNC y desem-penan la misma funcion general (v. fig. 9.13). Se dis-tinguen dos grupos de ganglios:
. Ganglios sensoriales, asociados a todos los nerviossensoriales que parten de la m�edula espinal y losnervios craneales V, VII, IX y X. Los gangliossensoriales vinculados a lam�edula espinal se llamanganglios de la raız dorsal, mientras que los
asociados a los nervios craneales reciben nombresespeciales para cada uno de estos nervios.Los ganglios sensoriales contienen neuronasunipolares. El endoneuro del axon se continuacon el tejido conjuntivo que rodea al ganglio.Los receptores especializados que envuelven losextremos distales de los nervios perif�ericos poseen lacapacidad de transduccion de varios estımulos y degenerar un potencial de accion, que pasadirectamente al enc�efalo o lam�edula espinal para suprocesamiento.
. Ganglios autonomos, cuya funcion es motora. Loscuerpos celulares de las neuronas parasimp�aticaspreganglionares se encuentran en el enc�efalo y lam�edula espinal sacra, mientras que los de lasneuronas simp�aticas se hallan en ciertossegmentos de la m�edula espinal tor�acica y lumbar.Los axones de las neuronas motoraspreganglionares conectan con sus ganglios en lassinapsis con los cuerpos celulares de las neuronasmotoras posganglionares. Los axones de lasneuronas posganglionares pueden unirse de nuevoal nervio perif�erico del que partieron en su recorridohacia los organos efectores. Muchas fibrasparasimp�aticas posganglionares localizadas en lacabeza se unen a ramificaciones del nerviotrig�emino (PCV) al abandonar el ganglio para viajarhasta los organos efectores. Las neuronasparasimp�aticas posganglionares que proceden delplexo de Meissner o de Auerbach, situado en lasparedes del aparato gastrointestinal, hacen sinapsisen los organos efectores cercanos.
Los ganglios simp�aticos se restringen a los gangliosde la cadena simp�atica paralela a la columna vertebralo bien a los ganglios colaterales localizados a lo largode la aorta abdominal. Los ganglios parasimp�aticosasociados a los nervios craneales se encuentran en lacabeza (con excepcion de los pertenecientes al PC X),mientras que los asociados a los nervios sacros lohacen en el organo al que inervan.
124
9
TEJIDONERVIO
SO

125
9
TEJIDONERVIO
SO
Figura 9.13 Sistema nervioso autonomo. Izquierda. Division simp�atica. Derecha. Division parasimp�atica. (Tomado de Gartner LP,
Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 209.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Sistema nervioso central
El enc�efalo y la m�edula espinal se componen de:
. Sustancia blanca, la cual contiene principalmentefibras nerviosas mielinizadas acompanadasde fibras no mielinizadas y c�elulas de la neuroglia.La abundante mielina que recubre los axonesle confiere su coloracion blanquecina.
. Sustancia gris, formada por acumulaciones decuerpos celulares de neuronas y sus dendritas juntoa axones nomielinizados. El color gris�aceo refleja laausencia de mielina.
La red enmaranada formada por los axones, lasdendritas y las proyecciones de las c�elulas de la neu-roglia conforman el neuropilo. Los agregados locali-zados de cuerpos neuronales en la sustancia blancareciben el nombre de nucleos. La sustancia gris selocaliza en la periferia del enc�efalo, mientras que lablanca se halla en niveles m�as profundos; en lam�edula espinal, la sustancia gris se encuentra pordebajo de la blanca. En un corte transversal de lam�edula espinal, la sustancia gris forma una H y sucentro representa el conducto central, un angostocanal revestido por c�elulas ependimarias que contie-nen LCR. Las proyecciones centrales de las neuronassensoriales llegan a los cuerpos celulares de las inter-neuronas de las astas dorsales, las caras superioresde las barras verticales de la H. Los axones de lasinterneuronas terminan en los cuerpos celulares delas neuronas motoras en la parte inferior de las barrasverticales de laH, las astas ventrales. Los axones de lasneuronas motoras atraviesan las raıces centrales parasalir de la m�edula espinal.
MENINGES
Las meninges representan los tres tejidos conjuntivosque recubren el cerebro y la m�edula espinal: la capaexterna o duramadre, la capa intermedia o aracnoi-des y la capa interna o piamadre (fig. 9.14).
1. La duramadre, la capa m�as externa de lasmeninges, difiere en el enc�efalo y la m�edulaespinal. La duramadre craneal est�a formada portejido conjuntivo denso, el cual consta de doscomponentes distintos:. Una capa periostica adosada ıntimamente alcr�aneo, que actua tambi�en como periostio de lacara interna de la boveda craneal. Presenta unavascularizacion rica y contiene c�elulasosteoprogenitoras, fibroblastos y haces decol�ageno de tipo I.
. La capa interna de la duramadre, la capamenıngea, posee fibroblastos de tincion oscuracon proyecciones largas, fibras delgadas de
col�ageno organizadas en l�aminas y pequenasarterias. La region m�as interna de la capamenıngea, la capa celular limıtrofe, se componede una delgada capa de fibroblastos envueltospor una matriz extracelular desorganizada quecarece de fibras de col�ageno y se extiende hacia lacapa menıngea.
La duramadre raquıdea no se dispone en capas. Adiferencia de la capa periostica, no se adhiere al canalvertebral, sino que forma un tubo continuo que rodeaa la m�edula espinal desde el agujero occipital hasta elsegundo segmento craneal. A lo largo de este con-ducto, los nervios raquıdeos atraviesan la duramadreraquıdea y el espacio comprendido entre el canal ver-tebral oseo y esta membrana, llamado espacio epidu-ral, contiene grasa epidural y un plexo venoso.
2. La aracnoides avascular se compone de dos capas:. Una l�amina aplanada que se encuentra adosada ala duramadre.
. Una segunda capa integrada por fibroblastosmodificados de organizacion laxa (c�elulastrabeculares aracnoideas) junto a algunas fibrasde col�ageno y fibras el�asticas de las trab�eculas encontacto con la piamadre.
. El espacio comprendido entre la l�aminaaplanada adyacente a la duramadre y lastrab�eculas en contacto con la piamadre se conocecomo espacio subaracnoideo.
. Los vasos sanguıneos atraviesan la aracnoidesconforme avanzan desde la duramadre hacia lapiamadre, aunque est�an aislados de la aracnoidesy el espacio subaracnoideo por una l�amina defibroblastos procedentes de aquella. El espaciosubaracnoideo es un compartimento real ocupadopor LCR, mientras que el espacio subdural,comprendido entre la duramadre y la capalaminar de la aracnoides en contacto con esta,representa un espacio potencial. Unas regionesespecializadas de la aracnoides, las vellosidadesaracnoideas, se prolongan hacia los senosvenosos de la duramadre y transportan el LCRdesde el espacio subaracnoideo a dichos senos.
3. La capa interna de las meninges, la piamadre,contiene fibroblastos aplanados, mastocitos,macrofagos y linfocitos; se encuentra en contactoestrecho con el enc�efalo y la m�edula espinal. Sinembargo, una delgada membrana formada porproyecciones neurogliales, que se adhieren a lasfinas fibras reticulares y el�asticas de la piamadrepara formar una barrera fısica en la periferia delSNC, separa esta capa del par�enquima encef�alico.La profusa red vascular que irriga la piamadre serecubre de una capa de c�elulas de la piamadre,que son sustituidas por c�elulas de la neurogliaconforme van atravesando el tejido nervioso.
126
9
TEJIDONERVIO
SO

CONSIDERACIONES CL�INICAS
Los tumores menıngeos, llamados meningiomas,suelen desarrollarse con lentitud y ser benignos, sibien pueden manifestarse con sıntomas clınicosgraves, como compresion cerebral o aumento de lapresion intracraneal.
La meningitis, una inflamacion de las meninges,puede deberse a la infeccion por bacterias o virusque han logrado acceder al LCR. La meningitis deetiologıa vırica no suele entranar un riesgo alto,mientras que la de origen bacteriano es muy
contagiosa adem�as de considerarse unaenfermedad muy grave. El acceso del patogeno al
organismo puede darse a trav�es de la nariz, el oıdoo la garganta, desde donde puede transmitirse pormedio de las secreciones respiratorias al toser obesarse. La meningitis debuta con fiebre, rigidez
nucal, n�auseas y vomitos. El diagnostico de esta
entidad se sustenta en los resultados del an�alisis delLCR obtenido mediante puncion lumbar; sutratamiento consiste en antibioterapia. Actualmentese dispone de vacunas que confieren proteccionfrente a algunas de las bacterias frecuentes queproducen meningitis.
La barrera hematoencef�alica (BHE) posee unanotable capacidad de discriminacion a la hora de
permitir el paso al SNC de mol�eculas procedentes
del torrente circulatorio, de modo que evita la
entrada de la mayorıa de los f�armacos, muchosantibioticos, toxinas y algunos neurotransmisores,como dopamina, en el tejido neural. La perfusion deuna solucion hipertonica de manitol alteratemporalmente las uniones estrechas de la BHE enun grado suficiente para hacer posible el paso de
algunos f�armacos. Otro m�etodo utilizado para
superar la BHE es la union del f�armaco de inter�es aanticuerpos frente a receptores de transferrinalocalizados en las c�elulas endoteliales de loscapilares, lo que facilita su transporte hacia el SNC.
Algunas enfermedades o trastornos que afectan alSNC, como los accidentes cerebrovasculares, lostumores y las infecciones, alteran laBHEal reducir sufuncionamiento y permitir el transporte de sustanciastoxicas y metabolitos indeseados al tejido neural.
El espacio subaracnoideo es un espacio realocupado por LCR, mientras que el espaciosubdural, localizado entre la duramadre y una capalaminar de la aracnoides adosada a esta, solamenterepresenta un espacio potencial. Puede convertirseen un espacio real cuando la sangre de unahemorragia separa las dos capas; este trastorno seconoce como hemorragia subdural.
127
9
TEJIDONERVIO
SO
Figura 9.14 El cr�aneo y las capas de las meninges que recubren el enc�efalo. (Tomado de Gartner LP, Hiatt JL: Color Textbook of
Histology, 3rd ed. Philadelphia, Saunders, 2007, p 212.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

BARRERA HEMATOENCEFALICA
Las c�elulas endoteliales de los capilares continuos deltejido neural establecen uniones estrechas entre sıpara crear la barrera hematoencef�alica (BHE), la cualrestringe el paso de las sustancias transportadas en eltorrente circulatorio hacia el territorio del SNC.
. Ciertas mol�eculas, como el O2, el CO2, el agua y loslıpidos de pequeno tamano, pueden atravesarlibremente la BHE.
. La mayorıa de las restantes sustancias, como laglucosa, los nucleosidos y los amino�acidos, han deser translocadas por proteınas transportadoras ycanales ionicos especıficos para ellas.
. Otros compuestos se transfieren por medio demecanismos de transporte mediado porreceptores.
Los astrocitos, cuyas proyecciones generan piesterminales que rodean por completo la l�amina basalde los capilares pertenecientes al SNC, refuerzan laBHE. Los botones terminales crean una envolturacilındrica, la glıa perivascular limitante. Los astroci-tos participan, igualmente, en el transporte de meta-bolitos de los capilares a las neuronas y en la captacionde iones K+ y neurotransmisor de los espacios extra-celulares en los que se encuentran inmersas las neu-ronas y sus proyecciones.
PLEXO COROIDEO
El plexo coroideo, que se compone de pliegues muyvascularizados de piamadre rodeados de c�elulas epen-dimarias cubicas, se proyecta hacia los ventrıculoscerebrales y fabrica alrededor del 50% del volumendel LCR. Se desconoce en qu�e lugar del enc�efalo seproduce el volumen restante. El LCR rellena los ven-trıculos cerebrales, el conducto central de la m�edulaespinal y los espacios subaracnoideos.
Lıquido cefalorraquıdeo
El LCR es un lıquido transparente en el que escaseanlas proteınas y abundan los electrolitos y que contieneuna reducida poblacion de linfocitos y otras c�elulas(tabla 9.4). Este lıquido se sintetiza de manera cons-tante a una velocidad de 0,2 a 0,6ml/min y es trans-ferido hacia los senos venosos durales por las vellosi-dades aracnoideas a una velocidad similar, de modoque su formacion y resorcion funcionan como unabomba que impulsa su circulacion a trav�es de losventrıculos cerebrales, el conducto central de lam�edula espinal y los espacios subaracnoideos. Estelıquido sustenta la actividad metabolica del SNC yconfiere proteccion frente a fuerzas repentinas quepodrıan afectar al enc�efalo y la m�edula espinal.
CORTEZA CEREBRAL
El enc�efalo se divide en dos hemisferios cuya capam�asexterna se compone de sustancia gris, la corteza cere-bral, y recubre una gruesa capa de sustancia blancasituada en niveles m�as profundos del mismo. La cor-teza cerebral se pliega en �areas elevadas denominadascircunvoluciones, las cuales se separanmediante unasdepresiones conocidas como surcos.
. El enc�efalo lleva a cabo multitud de funciones,como la memoria, el aprendizaje, la integracion dela informacion sensorial, el an�alisis de datos, elcomienzo de la respuesta motora y elprocesamiento del pensamiento.
. Lacorteza cerebralenglobaseis capashorizontales; lasneuronas de cada una de ellas presentan unos rasgosmorfologicos especıficos de cada capa (tabla 9.5).
. La capa externa se dispone inmediatamente pordebajo de la piamadre, mientras que la sexta capaest�a en contacto con la sustancia blanca de la cortezacerebral.
. Todas las capas contienen neuronas y c�elulas de laneuroglia especıficas.
128
9
TEJIDONERVIO
SO

CONSIDERACIONES CL�INICAS
El LCR se sintetiza continuamente, por lo que elbloqueo de los ventrıculos o el funcionamientodeficiente de las vellosidades aracnoideas ocasionauna hipertrofia ventricular denominada hidrocefalia.Esta entidad tiene consecuencias graves, ya que lahipertrofia de los ventrıculos incrementa la presionintracraneal. Las fontanelas y las suturas oseas del
cr�aneo no se encuentran fusionadas aun en losneonatos, de modo que esta enfermedad ocasiona
hipertrofia del cr�aneo, deterioro mental,funcionamiento incorrecto de los musculos y muerteen ausencia de tratamiento.
La enfermedad de Alzheimer, el trastornoneurodegenerativo m�as prevalente, afecta a unoscinco millones de sujetos en EE. UU. y originademencia progresiva y terminal. El diagnostico sueleelaborarse en sujetosmayores de 65 anos, si bien lossıntomas pueden haber comenzado mucho tiempo
antes. El sıntoma temprano m�as frecuente es la
p�erdida de memoria, al que suceden la confusion, lairritabilidad, la agresividad y los cambios del estado
de �animo. Las manifestaciones de la enfermedadavanzada engloban alteraciones del habla, la
p�erdida de memoria a largo plazo, la disminucion dela sensibilidad, el aislamiento del entorno, ladesaparicion de las funciones corporales y, porultimo, la muerte. No se conoce adecuadamente laetiologıa de esta enfermedad, aunque se ha descritola desaparicion de neuronas y sinapsis,principalmente en la corteza cerebral, seguida dela atrofia macroscopica de los lobulos cerebrales.Las autopsias de sujetos aquejados de laenfermedad de Alzheimer han revelado la presenciade placas amiloides y ovillos neurofibrilares en el
enc�efalo cuya expansion afecta a un numero cadavez mayor de neuronas, que dejan de funcionar connormalidad.
129
9
TEJIDONERVIO
SOTabla 9.4 COMPARACION DEL SUERO Y EL LIQUIDOCEFALORRAQUIDEO (LCR)
Componente Suero LCR
Leucocitos(c�elulas/ml)
0 0-5
Proteına (g/l) 60-80 Insignificante
Glucosa (mmol/l) 4-5,5 2,1-4
Na+ (mmol/l) 135–150 135-150K+(mmol/l) 4–5,1 2,8-3,2
Cl- (mmol/l) 100–105 115-130
Ca++ (mmol/l) 2,1–2,5 1–1,4Mg++ (mmol/l) 0,7–1 0,8–1,3
pH 7,4 7,3
Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology,
3rd ed. Philadelphia, Saunders, 2007, p 215.
Tabla 9.5 CAPAS DE LA CORTEZA CEREBRAL
Capa Denominacion Tipos neuronales*
1 Molecular C�elulas horizontales2 Granular externa C�elulas granulares muy
empaquetadas
3 Piramidal externa C�elulas piramidales grandes
4 Granular interna C�elulas piramidales, c�elulasgranulares pequenas;
densidad celular alta en una
delgada capa
5 Piramidal interna C�elulas piramidales grandes;densidad celular baja
6 Multiforme C�elulas de Martinotti
*En todas las capas se alojan celulas de la neuroglia.
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

130
9
TEJIDONERVIO
SO
CORTEZA CEREBELOSA
El cerebelo se compone de dos hemisferios laterales yuna porcion central que los conecta llamada vermis.La capa m�as externa del cerebelo, la corteza cerebe-losa, est�a formada por sustancia gris que recubre lasustancia blanca. Esta corteza se encarga de mantenerel equilibrio durante todas las fases de la postura cor-poral y coordina la actividad de los musculos volun-tarios y la conservacion del tono muscular. La cortezacerebelosa se divide en tres capas:
. Capa molecular, integrada fundamentalmente pordendritas de c�elulas de Purkinje y axones nomielinizados de la capa granular, ası como algunasc�elulas estrelladas y c�elulas en canasta.
. Capa de c�elulas de Purkinje, que contiene c�elulasde Purkinje grandes (exclusivas del cerebelo) cuyasdendritas arborizadas se localizan en la capamolecular, mientras que sus axones mielinizados seextienden hacia la sustancia blanca.
. Capa granular, formada por agregados de nucleosde c�elulas granulares pequenas y glom�erulos (islascerebelosas) que representan las sinapsis de losaxones que ingresan en el cerebelo y las c�elulasgranulares residentes en el mismo.
Las c�elulas de Purkinje tan solo emiten impulsosinhibitorios y procesan e integran simult�aneamenteinformacion procedente de cientos de miles de sinap-sis excitadoras e inhibitorias antes de generar unarespuesta. El unico neurotransmisor que secretan estasc�elulas es GABA, y constituyen las unicas c�elulas cere-belosas cuyas proyecciones y respuestas viajan a regio-nes fuera del cerebelo.
Regeneracion nerviosa
Es posible que algunas neuronas puedan proliferar, porlo general se estima que lamayorıa de las neuronas y losnervios del SNC danados por un traumatismo no pue-den regenerarse debido a su incapacidad de prolifera-cion. Los danos ocasionados a las neuronas y susproyecciones en el seno del SNC son permanentes.Sin embargo, los nervios perif�ericos danados puedenreparar las lesiones a trav�es de una serie de sucesos quereciben el nombre de reaccion de axon (fig. 9.15).
REACCION DE AXON
La reaccion frente al dano engloba cambios en tresregiones especıficas de la neurona: cambios locales,cambios anterogrados y cambios retrogrados. Algu-nas de las reacciones frente al dano se suceden conrapidez, mientras que lamayorıa de las modificacionesde reparacion, regeneracion y restablecimiento funcio-nal no concluir�an antes de varias semanas o meses.
. En el lugar del dano se produce una reaccion local.Si el axon hubiera sido seccionado, los extremos
cortados se alejan y son recubiertos por el axolema,lo que impide la perdida de axoplasma. Losmacrofagos, con la ayuda de las c�elulas de Schwann,comienzan a fagocitar el tejido lesionado.
. La reaccion anterograda consiste en ladegeneracion del segmento axonal comprendidoentre el lugar danado y el boton terminal.. A lo largo de los 7 dıas siguientes al dano, el pieterminal se hipertrofia y comienza a degenerar,como consecuencia de lo cual deja de estar encontacto con la membrana postsin�aptica. Lasc�elulas de Schwann proliferan y migran hacia laantigua zona de sinapsis, en la que fagocitan losrestos del boton terminal.
. La region del axon y su vaina de mielina,localizada entre el lugar de la lesion y la hendidurasin�aptica anterior, se fragmentan en unproceso dedegeneracion walleriana. Las c�elulas de Schwanndejan de sintetizar mielina y comienzan afagocitar los restos del axon distal y su vaina demielina, aunque se conserva la l�amina basal delendoneuro.
. La proliferacion de la poblacion de las c�elulas deSchwann prosigue y las nuevas c�elulas forman untubo de Schwann envuelto por la l�amina basalderivada del endoneuro.
. La reaccion retrograda y la regeneracion afectana la region neuronal localizada entre el lugarde la lesion y el soma en el interior delSNC.. La neurona danada sufre un proceso decromatolisis: los corpusculos de Nissldifunden, el nucleo se desplaza, el somase hipertrofia y la neurona fabricamacromol�eculas que se emplean para laregeneracion del axon lesionado.
. El extremo proximal del axon comienza aproyectar numerosos brotes axonales, uno de loscuales alcanza la luz del tubode Schwann a trav�esdel endoneuro; los restantes brotes axonalesdegeneran y son fagocitados por macrofagos yc�elulas de Schwann.
. El brote axonal se prolonga a lo largo del tubo deSchwann guiado por factores inductoressecretados por las c�elulas de Schwann, losfibroblastos y los macrofagos; a un ritmo de 3 a4mm/dıa, el brote axonal alcanza la membranapostsin�aptica y restablece la sinapsis.
La regeneracion podrıa interrumpirse cuando laobstruccion del tubo de Schwann debido a la presen-cia de tejido cicatricial impida el avance del broteaxonal a lo largo de su luz. En este caso, la c�elulapostsin�aptica sufrirıa un proceso de atrofia conocidocomodegeneracion transneuronal, lo que indica quela neurona ejerce una influencia trofica sobre lac�elula con la que establece la sinapsis.

CONSIDERACIONES CL�INICAS
Hasta hace poco tiempo, se creıa que el SNC nopodıa regenerarse por diversos motivos, como lapresencia de unos macrofagos llamadosmicroglia
que fagocitaban las c�elulas danadas con granrapidez y la ocupacion del espacio despejado por
una masa de c�elulas gliales para crear una cicatriz
glial. El SNC posee c�elulas progenitorasneuronales capaces de proliferar al ser inducidascon el fin de dar lugar a nuevas neuronas que
asumirıan las funciones de las c�elulas danadas porla lesion. Los resultados de algunos estudios
recientes centrados en las c�elulas progenitoras, laplasticidad neuronal, el factor de crecimientonervioso, lo inhibidores de la proliferacion nerviosay las neurotrofinas nos permiten albergaresperanzas sobre la reparacion y la inversion de laslesiones medulares.
131
9
TEJIDONERVIO
SO
Figura 9.15 Esquema de la regeneracion
nerviosa. A. Neurona normal. B-D.Aspecto a las 2 semanas. B. A las 3 semanas.
C. A los 3 meses. D. Despu�es de la lesion.
E. Aspecto varios meses despu�es de la
lesion en un caso de regeneracion neuronalinsatisfactoria. (Tomado de Gartner LP,
Hiatt JL: Color Textbook of Histology, 3rd ed.
Philadelphia, Saunders, 2007, p 217.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

10 SANGREY HEMATOPOYESIS
Una personal normal posee unos 5 l de sangre, unlıquido rojo y viscoso cuyo pH es de 7,4. El corazonbombea sangre a trav�es de los vasos del sistema cir-culatorio para distribuir nutrientes,mol�eculas de senalizacion, electro-litos, y oxıgeno a las c�elulas delorganismo, ası como retirar de estassustancias de desecho y dioxido decarbono que ser�an eliminados porlos organos encargados de estatarea. Por otra parte, en el torrentecirculatorio viajan c�elulas especialesy elementos formes, los cualesdesempenan sus funciones en elmismo torrente o bien abandonanel sistema circulatorio al llegar a sudestino y migran hacia el comparti-mento de tejido conjuntivo en elque actuar�an. De igual modo, lasangre interviene en la regulaciondel equilibrio osmotico y acidob�asico y de la tempe-ratura corporal. La sangre es un lıquido, por lo queest�a dotada de un mecanismo protector de coagu-lacion, dirigido por las plaquetas, que reduce lap�erdida de sangre en caso de danos a los vasos san-guıneos.
Al centrifugar 100ml de sangre en un tubo deensayo heparinizado, se separa en una fraccion quecontiene c�elulas y elementos formes y una fraccionlıquida, el plasma.
. Los 44ml que ocupan el fondo del tubocorresponden a eritrocitos (globulos rojos)empaquetados.
. Una capa leucocitaria de 1ml, formada porleucocitos (globulos blancos), se dispone porencima de la anterior.
. Por encima de los leucocitos se situan 55ml deplasma.
El 44% que representan los eritrocitos constituye elhematocrito, el volumen eritrocıtico total.
Las c�elulas sanguıneas y las plaquetas tienenuna vida limitada, por lo que deben reponersediariamente para mantener unas poblaciones cons-tantes de cada tipo celular en el torrente circula-
torio. El proceso de renovacion continua se conocecomo hematopoyesis.
Sangre
PLASMA
El plasma constituye el 55% delvolumen sanguıneo:
. El 90% corresponde a agua
. El 9% son proteınas
. El 1% se compone de electrolitos,nutrientes y gases disueltos.
En la tabla 10.1 se muestra lacomposicion proteica del plasma.El componente lıquido de la san-gre, con escasas proteınas, sale delas v�enulas de pequeno calibre ylos lechos capilares para extrava-
sarse al compartimento de tejido conjuntivo, en elque se denomina lıquido extracelular (lıquido deltejido intersticial). La proteına albumina crea unapresion coloidosmotica en el seno del sistema cir-culatorio gracias a la cual el lıquido permanecedentro de sus confines y se limita el volumen del lıqui-do extracelular.
Al coagularse la sangre, una parte de las proteınas ylos factores presentes en el plasma desaparecen duranteel proceso de coagulacion. El lıquido de color pajizo asıformado, el suero, es un derivado del plasma en el queabundan las proteınas y est�a ausente el fibrinogeno.
ELEMENTOS FORMES
Los elementos formes de la sangre son los eritrocitos,los leucocitos y unos fragmentos celulares llamadosplaquetas (fig. 10.1). En el estudiomicroscopico de lasc�elulas sanguıneas se utilizan t�ecnicas y tincionesespeciales:
. Por lo general, se extiende una gota de sangre om�edula osea en un portaobjetos de vidrio y se dejasecar al aire.
. El portaobjetos se sumerge enmetanol absoluto y seseca al aire.
� 2011. Elsevier Espana, S.L. Reservados todos los derechos
TERMINOS CLAVE
. Plasma
. Eritrocitos
. Agranulocitos
. Granulocitos
. C�elulas madre
. C�elulas progenitoras
. C�elulas precursoras
. Factores decrecimientohematopoy�eticos
132

. La preparacion se tine mediante la modificacion deWright o Giemsa de la tincion de Romanovsky,que incluıa inicialmente una mezcla de eosina yazul de metileno.
. La preparacion se lava con agua, se deja secar al airenuevamente y se recubre, o no, con un cubreobjetospara ser observada.
En este texto, las descripciones de las c�elulassanguıneas (salvo los reticulocitos, que se abor-dar�an en el apartado relativo a la eritropoyesis) sebasan en la coloracion obtenida mediante estas tin-ciones.
133
10
SANGREYHEM
ATOPO
YESIS
Tabla 10.1 PROTEINAS PRESENTES EN EL PLASMA
Proteına Tamano Origen Funcion
Albumina 60.000–69.000 Da Hıgado Mantenimiento de presion
coloidosmotica y
transporte de algunosmetabolitos insolubles
Globulinas
a-globulinas y b-globulinas 80.000–1� 106 Da Hıgado Transporte de iones
met�alicos, lıpidos unidos aproteınas y vitaminas
liposolubles
g-globulinas C�elulas plasm�aticas Anticuerpos del sistema
inmunitarioProteınas de coagulacion
(p. ej., protrombina,
fibrinogeno, globulinaaceleradora)
Variado Hıgado Formacion de filamentos de
fibrina
Proteınas del complemento
C1-C9
Variado Hıgado Destruccion de
microorganismos y
comienzo de la respuestainflamatoria
Lipoproteınas plasm�aticas
Quilomicrones 100–500mm C�elulas epiteliales del
intestino
Transporte de triglic�eridos al
hıgadoLipoproteına de muy baja
densidad
25-70 nm Hıgado Transporte de triglic�eridos
del hıgado a las c�elulas del
organismoLipoproteına de baja
densidad
3�106 Da Hıgado Transporte de colesterol del
hıgado a las c�elulas del
organismo
Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 221.
Figura 10.1 Elementos formes de la sangre circulante. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia,
Saunders, 2007, p 220.)
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

Eritrocitos
Los eritrocitos se asemejan a discos biconcavos; repre-sentan las c�elulas sanguıneas m�as numerosas y peque-nas (di�ametro, 7,2mm) (v. fig. 10.1). El numero deeritrocitos por unidad de volumen difiere en ambossexos: las mujeres portan 4,5�106 eritrocitos pormm3, mientras que esta cifra es de 5� 106 en loshombres. Este numero aumenta en personas deambos sexos que residen en lugares a gran altitud.A diferencia de otras c�elulas del organismo, los eritro-citos carecen de org�anulos o nucleo, que se expulsarona lo largo de su desarrollo antes de incorporarse altorrente circulatorio. La morfologıa de los eritrocitosles proporciona un �area de superficie grande con rela-cion a su volumen, lo que facilita el desempeno de sufuncion: el transporte e intercambio de O2 por CO2
y viceversa. La presencia de numerosas mol�eculasde hemoglobina en los eritrocitos facilita la realiza-cion de dichas tareas; estas c�elulas poseen, asimismo,la enzima anhidrasa carbonica. Preferentemente,estas c�elulas:
. Ceden O2 y recogen CO2 en regiones con unatension baja deO2 y tension alta de CO2 –los tejidosque integran el organismo.
. Captan O2 y liberan CO2 en las regiones ricas en O2
y pobres en CO2 –los pulmones.
Membrana celular de los eritrocitos
La composicion de la membrana celular de los eri-trocitos (fig. 10.2) es similar a la de otros tipos celu-lares, con un 40% de fosfolıpidos en una bicapa, un10% de hidratos de carbono y un 50% de proteınas,que corresponden, en esencia, a:
. Glucoforina A, una de las dos proteınastransmembrana que forma parte del complejo deunion de proteınas de union a espectrina
. Canales ionicos
. Proteınas transportadoras, como la proteınabanda 3 transportadora de iones, que intercambiaCl� y HCO3
� entre ambas caras de la membranay posibilita la cesion de CO2 en los pulmones
. Proteına perif�erica banda 4.1, la cual une laglucoforina A a la actina y la tropomiosina
La membrana plasm�atica se sostiene en una red detetr�ameros de espectrina, unas proteınas que creanun esqueleto hexagonal por debajo de aquella, con lacolaboracion de anquirina, la cual une espectrina a laproteına banda 3. El complejo de union de proteınasintegrado por la proteına banda 4.1, actina, aducina,tropomiosina y glucoforina proporciona un apoyoadicional al entramado de espectrina. Adem�as de con-ferir un grado alto de flexibilidad al eritrocito, el entra-
mado proteico confiere una extraordinaria estabilidady capacidad de resistencia a las fuerzas de cizalla-miento. La duracion de la vida de estas c�elulas es deunos 120 dıas, durante los cuales atraviesan estrechoscapilares en innumerables ocasiones, en las que sedistorsionan y son sometidos a potentes fuerzas decizallamiento, para retomar su morfologıa normal alsalir de estos conductos.
La superficie extracelular de la membrana plas-m�atica del eritrocito presenta polisac�aridos heredita-rios de naturaleza antig�enica que deben tenerseen cuenta en las transfusiones sanguıneas. Los dosantıgenos principales son los antıgenos A y B, quedan lugar a cuatro grupos sanguıneos (tabla 10.2).Asimismo, el 85% de la poblacion estadounidenseporta uno de los tres antıgenos Rh m�as importantes(C, D y E), por lo que se dice que son Rh-positivos,mientras que el 15% restante se considera Rh-negativo(v. «Consideraciones clınicas»).
Transporte de dioxido de carbono y oxıgeno
Los eritrocitos transportan CO2 y O2 a trav�es de dosmecanismos distintos. La mayor parte del CO2
se transporta en forma de iones HCO3- (formado
por accion de la anhidrasa carbonica a partir deH2O y CO2, que se combinan para dar lugar aH2CO3, el cual se disocia de inmediato en H+ yHCO3
-). En los pulmones, caracterizados por una ten-sion baja de CO2, los iones HCO3
- salen del cito-plasma de los eritrocitos y los iones Cl- pasan al inte-rior de los mismos por medio de proteınas banda 3,las cuales est�an implicadas en el intercambio ionico(este proceso de intercambio recibe el nombre dedesviacion de cloruros).
La proteına hemoglobina, una mol�ecula tetra-m�erica de gran tamano, se encarga del transporte deoxıgeno. Cada una de las cuatro cadenas polipeptıdi-cas de la hemoglobina establece un enlace covalentefuerte con un grupo hemo que contiene hierro. En latabla 10.3 se describen los principales tipos de hemo-globina.
. El grupo globina de la hemoglobina transportauna pequena cantidad de CO2 y recibe el nombrede carbaminohemoglobina; libera CO2 en �areasde baja tension de CO2 (pulmones).
. El O2, que es captado en los pulmones, en los queabunda, se une a la porcion hemo para formaroxihemoglobina, que se transporta hasta regionescon una baja tension de O2, a las que ceder�a este�atomo.
. Una mol�ecula de 2,3-difosfoglicerato ocupa ellugar correspondiente al O2 y la hemoglobina seconvierte en desoxihemoglobina.
134
10
SANGREYHEM
ATOPO
YESIS

CONSIDERACIONES CL�INICAS
La eritroblastosis fetal, un trastorno que puedetener consecuencias mortales para el feto, se debe ala destruccion de los eritrocitos fetales por parte delsistema inmunitario materno. A finales de lagestacion y en el transcurso del parto, la sangre fetalpuede pasar al torrente circulatorio materno. Si lamadre presenta un Rh negativo y el feto posee Rhpositivo, la primera desarrollarıa anticuerpos frente alantıgeno Rh. En un primer momento, sintetizaanticuerpos IgM, cuyo tamano es excesivamentegrande para atravesar la barrera placentaria, por loque no afectan al feto. En los embarazos posteriorespueden aparecer complicaciones cuando el fetopresente Rh positivo, ya que el sistema inmunitariomaterno habrıa cambiado los isotipos y producirıaIgG frente a dicho antıgeno en lugar de IgM. Eltamano de los anticuerpos IgG es menor, por lo quepueden atravesar la barrera placentaria para unirse alos antıgenos Rh de los eritrocitos fetales y provocarsu hemolisis, lo que destruye al feto. Para evitar este
trastorno, la madre recibe aglutininas frente al factor
Rh despu�es del nacimiento del primer hijo con Rhpositivo con el fin de ocultar los epıtopos presentes
en las c�elulas sanguıneas fetales y evitar el desarrollode una respuesta inmunitaria completa frente a dichoantıgeno.
La morfologıa del eritrocito presenta una ıntimarelacion con su funcion: las mutaciones que alteranla forma normal de los eritrocitos pueden dar lugar adistintas variantes de anemia. La alteracion de lacadena polipeptıdica de espectrina puede reducir sucapacidad de union a la proteına banda 4.1, demodo
que las mol�eculas de espectrina no podrıan sostenerde manera adecuada el plasmalema de dicha
c�elulas. Esta entidad recibe el nombre deesferocitosis hereditaria. Los eritrocitos del sujetoafectado son incapaces de transportar una cantidad
suficiente de oxıgeno, son fr�agiles y tienden a serdestruidos en el bazo, lo que origina anemia.
135
10
SANGREYHEM
ATOPO
YESIS
Tabla 10.3 TIPOS PRINCIPALES DE HEMOGLOBINA
Cadenaspolipeptıdicas
Tipo dehemoglobina Designacion
aagg Hemoglobina fetal HbFaabb Hemoglobina
adulta (m�as
frecuente, 96%)
HbA1
aadd Hemoglobina
adulta
(infrecuente, 2%)
HbA2
Figura 10.2 Membrana plasm�atica de un eritrocito y proteınas asociadas a ella. (Tomado de Gartner LP, Hiatt JL: Color Textbook ofHistology, 3rd ed. Philadelphia, Saunders, 2007, p 224.)
Tabla 10.2 SISTEMA DE GRUPOS SANGUINEOS AB0
Gruposanguıneo
Antıgenospresentes Varios
A Antıgeno AB Antıgeno B
AB Antıgenos A y B Aceptor universal
0 Ausencia de
antıgenos A y B
Donante
universal
Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology,
3rd ed. Philadelphia, Saunders, 2007, p 224.
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

Leucocitos
El numero de leucocitos es de 6.500 a 10.000 pormm3 de sangre. No desempenan funcion alguna enel seno del torrente circulatorio, ya que lo utilizanexclusivamente para acceder a su destino. Al llegar allugar de inter�es, los leucocitos abandonan los capi-lares o las v�enulas mediante diap�edesis (migracionentre las c�elulas endoteliales) para pasar al comparti-mento de tejido conjuntivo en el que realizar�an susfunciones. Se han establecido dos grandes grupos deleucocitos (tabla 10.4):
. Agranulocitos, leucocitos carentes de gr�anulosespecıficos:. Linfocitos. Monocitos
. Granulocitos, leucocitos con gr�anulos especıficos:. Neutrofilos. Eosinofilos. Basofilos
Linfocitos
Los linfocitos, c�elulas redondeadas con un nucleoac�entrico que ocupa gran parte del citoplasma, pre-sentan un tamano ligeramente mayor que los eritro-citos y constituyen del 20 al 25% de la poblacion deleucocitos (v. tabla 10.4). Los linfocitos son agranu-locitos, lo que supone la ausencia de gr�anulos espe-cıficos, si bien poseen algunos gr�anulos azurofilos quese han identificado como lisosomas en los estudios demicroscopia electronica. Se distinguen tres tipos delinfocitos:
. Linfocitos B, representan el 15% dela poblacion linfocıtica y se ocupan de larespuesta inmunitaria humoral. Estas c�elulasadquieren la competencia inmunitaria en unaregion desconocida de la m�edula osea. Setransforman en c�elulas plasm�aticas quesintetizan anticuerpos al recibir estimulacionantig�enica.
. Linfocitos T, constituyen el 80% de los linfocitos yse encargan de la respuesta inmunitaria celular. Seconvierten en c�elulas competentes desde el punto devista inmunitario en el timo.
. C�elulas nulas, conforman el 5% de la poblacionlinfocıtica, y se dividen en dos clases: c�elulas madrey linfocitos citotoxicos naturales (NK).. Las c�elulas madre circulantes puedendiferenciarse para originar todos los tipos dec�elulas sanguıneas y las plaquetas.
. Los linfocitos NK son c�elulas citotoxicas que nointeraccionan con el timo ni con otros linfocitos Tpara desarrollar sus funciones.
En el capıtulo 12 se abordan las funciones de loslinfocitos B, los linfocitos T y las c�elulas NK. Lasc�elulas madre se describen en un apartado posteriorde este capıtulo.
Monocitos
Los monocitos, unas c�elulas redondeadas con unnucleo arrinonado, son las c�elulas sanguıneas demayor tamano en el torrente circulatorio (v. tabla10.4). En la microscopia electronica, se observan liso-somas abundantes y un aparato de Golgi poco desa-rrollado en la proximidad de las indentaciones nuclea-res. Los monocitos permanecen en el torrentecirculatorio unos pocos dıas despu�es de salir de lam�edula osea y pronto abandonan la circulacion, pasanal compartimento de tejido conjuntivo, se diferencianen macrofagos, que forman parte del sistema fagocı-tico mononuclear. Los macrofagos:
. Fagocitan principalmente c�elulas muertas oalteradas y patogenos invasores, ya sea antıgenosinertes o microorganismos vivos
. Secretan mol�eculas de senalizacion que inducenrespuestas inflamatorias y la proliferacion de c�elulasimplicadas en la respuesta inmunitaria
. Pueden fusionarse entre sı para convertirse enc�elulas gigantes de cuerpo extrano capaces defagocitar mol�eculas de mayor tamano cuando eltamano de una partıcula sea excesivamente grandepara un macrofago aislado
. Se diferencian en c�elulas presentadoras deantıgenos que fagocitan antıgenos, los degradan enunidades antig�enicas menores llamadas epıtopos,las asocian a los antıgenos del complejo principalde histocompatibilidad II (MHC II, tambi�enllamados antıgenos leucocitarios humanos [HLA]de clase II), que portan en sus membranas, ypresentan estos fragmentos proteicos a c�elulasinmunocompetentes.
CONSIDERACIONES CL�INICAS
La inflamacion es la respuesta del organismofrente a estımulos perjudiciales, como agresionesfısicas o quımicas o la invasion por patogenos.La respuesta vascular inicial se conoce comoinflamacion aguda; se elimina, ası, a los agentesnocivos y se pone en marcha el proceso decicatrizacion. La inflamacion prolongada se llamainflamacion cronica y supone el reclutamiento de
monocitos, linfocitos, c�elulas plasm�aticas yfibroblastos que tratan de atenuar las causas de lareaccion inflamatoria.
136
10
SANGREYHEM
ATOPO
YESIS

137
10
SANGREYHEM
ATOPO
YESISTabla10.4
LEUCOCITOS:RASGOS,CATEGORIASYFUNCIONES
GRANULOCITOS
AGRANULOCITOS
Rasgos
Neu
trofilos
Eosinofilos
Basofilos
Linfocitos
Monocitos
N.�/m
m3
3.500-7.000
150-400
50-100
1.500-2.500
200-800
%leucocitos
60-70
2-4
<1
20-25
3-8
Di� ametro
(mm)
Cortetran
sversal
8-9
9-11
7-8
7-8
10-12
Frotis
9-12
10-14
8-10
8-10
12-15
Nucleo
Tresacuatro
lobulos
Doslobulos(form
ade
salchicha)
Form
adeS
Red
ondeado
Arrinonad
o
Gr� anulosespecıficos
0,1mm
,colorrosa
claro*
1-1,5mm
,colorrosa
oscuro
*0,5mm
,colorazul-negro
*Ausentes
Ausentes
Contenidosdegr� an
ulos
especıficos
ColagenasatipoIV,
fosfolipasaA2,
lactoferrina,
lisozima,
fago
citina,
fosfatasa
alcalina,
proteınade
unionavitaminaB12
Arilo
sulfatasa,
histaminasa,
b-glucuronidasa,
fosfatasa
� acida,
fosfolipasa,
proteınab� asicaprincipal,
proteınacationica
eosinofila,neu
rotoxina,
ribonucleasa,catepsina,
peroxidasa
Histamina,
hep
arina,
factor
quim
iot� acticode
eosinofilos,peroxidasa,
proteasas
neu
tras,
condroitinasulfato
Ausentes
Ausentes
Marcadoresdesuperficie
Recep
toresFc,receptordel
factoractivadorde
plaquetas,receptorde
leucotrienoB4,mol� ecula
dead
hesionaleucocitos1
Recep
toresIgE,receptordel
factorquim
iot� acticode
eosinofilos
Recep
toresdeIgE
LinfocitosT:receptoresde
linfocitosT,mol� eculas
CD,receptoresdeIL
LinfocitosB:
inmunoglobulinas
de
superficie
HLAdeclaseII,receptores
Fc
Perıododevida
<1seman
a<2seman
as1–2an
os(enraton)
Demeses
avariosan
os
Unosdıasen
lasangre,
variosmeses
entejido
conjuntivo
Funcion
Fago
citosisydestruccionde
bacterias
Fago
citosisdecomplejo
antıgeno-anticuerpo;
destrucciondepar� asitos
Similar
amastocitosal
actuar
comomed
iadores
enrespuestas
inflam
atorias
LinfocitosT:respuesta
inmunitaria
celular
LinfocitosB:respuesta
inmunitaria
humoral
Sediferen
cian
en
macrofago
s:fago
citosis,
presentacionan
tig� enica
*Tinciones
detipoRoman
ovsky(o
modificaciones
delasmismas).
CD,grupodediferen
ciacion;HLA,an
tıgenoleucocitariohuman
o;IL,interleu
cina.
Tomad
odeGartner
LP,HiattJL:Color
Textbookof
Histology,3rd
ed.Philadelphia,Saunders,2007,p226.

Neutrofilos
Los neutrofilos (leucocitos polimorfonucleares,polis) representan entre el 60 y el 70% de la pobla-cion leucocitaria (v. tabla 10.4). El nucleo de unneutrofilo inmaduro tan solo posee de dos a treslobulos; conforme avanza el proceso de maduracioncelular se incrementa el numero de lobulos, que seconectan al cuerpo principal del nucleo (o entre sı)por delgados hilos de cromatina. En la mujer, losnucleos de los neutrofilos suelen poseer un pequenoap�endice o «palillo de tambor» que alberga al cor-pusculo de Barr, el segundo cromosoma X queparece encontrarse inactivo. La membrana celularde los neutrofilos porta el receptor de Fc y el recep-tor del componente C3b del complemento, asıcomo L-selectina e integrinas que facilitan la adhe-sion al revestimiento endotelial como paso previopara la diap�edesis. El citoplasma de estas c�elulascontiene tres tipos de gr�anulos (v. tabla 10.4):
. Gr�anulos especıficos, de pequeno tamano,formados por mol�eculas utilizadas por la c�elulapara destruir microorganismos
. Gr�anulos azurofilos (lisosomas), compuestos porenzimas hidrolıticas y oxidativas
. Gr�anulos terciarios, rellenos de gelatinasa,catepsinas y algunas glucoproteınas.
Las mol�eculas quimiot�acticas que secretan algunasc�elulas, como los mastocitos, atraen a los neutrofiloshacia focos de inflamacion aguda, en los que atacan alas bacterias invasoras. Para delimitar el lugar de salidaen el vaso sanguıneo, las c�elulas endoteliales (activa-das por interleucina 1 (IL-1) y el factor de necrosistumoral (TNF) sintetizados por c�elulas del tejido con-juntivo) expresan mol�eculas de adhesion intercelu-lar 1 y 2 (ICAM-1 e ICAM-2) a las que se unir�an lasintegrinas de los neutrofilos, los cuales interrumpir�ansu viaje en el torrente circulatorio y abandonar�an elvaso sanguıneo. Para entrar en el tejido conjuntivo yfagocitar las bacterias patogenas:
. Los gr�anulos terciarios liberan la enzima gelatinasa,que digiere la l�amina basal con el fin de facilitar ladiap�edesis
. Los neutrofilos secretan enzimas de los gr�anulosespecıficos, las cuales destruyen las bacterias queinvadieron el tejido conjuntivo.
. Los neutrofilos fagocitan bacterias mediante losreceptores de membrana Fc y C3b (fig. 10.3A y B).
. Los gr�anulos azurofilos liberan su contenidoenzim�atico en los fagosomas cargados de bacterias(fig. 10.3C).
. Los gr�anulos especıficos secretan su cargaenzim�atica en los fagosomas con bacterias, lo quedestruye a los microorganismos mediante
degradacion enzim�atica y formacion desuperoxidos por accion de la oxidasa dedinucleotidos de nicotinamida-adenina-fosfato(NADPH) en el O2. Se pone en marcha, de estemodo, una explosion respiratoria que da lugara �acido hipocloroso y peroxido de hidrogeno(fig. 10.3D).
Los neutrofilos mueren tras haber destruido lospatogenos invasores y forman el pus junto a las bac-terias muertas y el lıquido extracelular.
Eosinofilos
Los eosinofilos, que engloban del 2 al 4% de losleucocitos, son unas c�elulas redondeadas grandesdotadas de un nucleo bilobulado. En su citoplasmaabundan los gr�anulos especıficos eosinofilos degran tamano y gr�anulos azurofilos similares a loslisosomas de los neutrofilos (v. tabla 10.4). La mem-brana celular porta IgG, IgE y receptores del comple-mento, ası como receptores para el factor quimiot�ac-tico de eosinofilos, histamina y leucotrienos. Loseosinofilos se encargan de destruir par�asitos, adem�asde fagocitar y degradar complejos antıgeno-anti-cuerpo. Los gr�anulos especıficos de los eosinofilosposeen dos regiones:
. La region externa contiene diversas enzimashidrolıticas y catepsinas, peroxidasa ehistaminasa, la cual limita la respuestainflamatoria (v. tabla 10.4).
. La region interna alberga la proteına b�asica mayor(MBP), la proteına cationica de eosinofilos (ECP)y la neurotoxina derivada de neutrofilos. La MBP yla ECP crean poros en las envolturas de los par�asitosa trav�es de los que entran el peroxido de hidrogeno ylos aniones superoxido generados por las enzimasfrente a estos microorganismos.
Basofilos
Los basofilos representan solamente un 1% de lapoblacion leucocıtica (v. tabla 10.4). Su membranaplasm�atica contiene receptores de gran afinidad porIgE; poseen un nucleo en forma de S y un citoplasmarico en gr�anulos especıficos oscuros con histamina,heparina, factor quimiot�actico de eosinofilos, factorquimiot�actico de neutrofilos, peroxidasa, proteasasneutras y condroitina sulfato. Adem�as, albergan gr�anu-los azurofilos semejantes a los de los eosinofilos.
Aunque es probable que carezcan de relacioncon los mastocitos, los basofilos desempenan unasfunciones muy parecidas, ya que ambos ponenen marcha respuestas inflamatorias. La funcionde los mastocitos se aborda con mayor detalle enel capıtulo 6.
138
10
SANGREYHEM
ATOPO
YESIS

139
10
SANGREYHEM
ATOPO
YESIS
Figura 10.3 A-D. Proceso de fagocitosis bacteriana basado en los receptores C3b y Fc y su destruccion por los neutrofilos.
H2O2, peroxido de hidrogeno; HOCl, �acido hipocloroso; MPO, mieloperoxidasa; O2-, anion superoxido; PLA2, fosfolipasa A2.
(Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 228.)
CONSIDERACIONES CL�INICAS
Lasmutaciones en genes que codifican ligandos quese unen a selectinas impiden la marginacion y laralentizacion de los leucocitos, como los neutrofilos,al entrar en contacto con las superficies luminales de
las c�elulas endoteliales. Por lo general, estosligandos entran en contacto con selectinas
presentes en la superficie de estas c�elulas; loscontactos son reversibles, por lo que los leucocitosno se detienen y tan solo se ralentizan. Estaenfermedad se denomina deficiencia de adhesionleucocitaria tipo I. Los macrofagos del tejidoconjuntivo liberan IL-1 y TNF-a, que inducen laexpresion de ICAM-1 e ICAM-2 por parte de las
c�elulas endoteliales localizadas en el foco deinflamacion. Dos de las integrinas leucocitarias, elantıgeno asociado a la funcion de los leucocitos 1(LFA-1) y macrofagos 1 (Mac-1) se adhieren aICAM-1 y -2, como consecuencia de lo cual se
detienen los leucocitos. Las c�elulas inflamatorias deltejido conjuntivo fabrican IL-8, que estimula la
diap�edesis en los leucocitos y su salida del vaso
sanguıneo para migrar en el seno del tejidoconjuntivo. Los sujetos portadores demutaciones enlos genes que codifican las integrinas leucocitariaspadecen un trastorno conocido como deficienciade adhesion leucocitaria tipo I y algunos de susleucocitos no intervienen en la respuestainflamatoria.
Las mutaciones que afectan al gen que codifica laenzima oxidasa de NADPH alteran la capacidad de
algunas c�elulas, como los neutrofilos, de someterseal estallido respiratorio ante la invasion tisular porbacterias patogenas. Los neutrofilos acceden al focode infeccion y fagocitan las bacterias invasoras, sibien no pueden destruirlas con la suficiente rapidez yeficacia para lograr reducir sus poblaciones a unosniveles seguros. Este trastorno afecta
fundamentalmente a la poblacion pedi�atrica y recibeel nombre de deficiencia hereditaria de la oxidasade NADPH, una entidad que se asocia a infeccionesbacterianas frecuentes.
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

Plaquetas
Las plaquetas, fragmentos celulares cuyo di�ametrooscila de 2 a 4mm (fig. 10.4; v. tabla 10.4), se formana partir de los megacariocitos de la m�edula osea eintervienen en la coagulacion sanguınea y la protec-cion de vasos danados. Las plaquetas circulan duranteunas 2 semanas antes de ser destruidas. Presentan unamorfologıa discoide con un halo m�as claro, el hialo-mero, y una region central oscura, el granulomero. Enla microscopia electronica, se puede observar unanillo integrado por 10 a 15 microtubulos paralelosy flanqueado pormonomeros de actina ymiosina querodean el perımetro celular. El hialomero posee dossistemas de canales citopl�asmicos (v. fig. 10.4):
. Un canal que desemboca en la superficie, llamadosistema tubular de apertura en superficie, queconstituye una extension de la superficie externa dela plaqueta y multiplica entre siete y ocho veces su�area de superficie
. Un canal que no desemboca en la superficie, elsistema tubular denso
En la microfotografıa electronica, el granulomerocontiene peroxisomas, mitocondrias, gr�anulos de glu-cogeno, sistemas enzim�aticos que intervienen en elmetabolismo plaquetario y en la produccion de trifos-fato de adenosina (ATP), y tres tipos de gr�anulos cuyoscontenidos se enumeran en la tabla 10.5:
. Gr�anulos alfa
. Gr�anulos delta
. Gr�anulos gamma (lisosomas)
Las plaquetas limitan la salida de sangre de losvasos danados por medio de la formacion de unco�agulo que tapona la abertura en la pared del vaso.Cuando se altera el revestimiento endotelial yentran en contacto con el col�ageno, las plaquetasse activan y se fijan al �area danada y entre sı ensendos procesos conocidos como adhesion plaque-taria y agregacion plaquetaria, respectivamente. Laformacion del co�agulo depende de la interaccion dedistintos factores derivados del plasma, las plaque-tas y el tejido danado. La capa endotelial no danadalibera oxido nıtrico y prostaciclinas para evitar laagregacion plaquetaria, ası como mol�ecula hepa-rinoide y trombomodulina para impedir la coagu-lacion. El proceso de coagulacion sanguınearequiere un abanico de factores que actuan en unacascada en la que la formacion o activacion de unode ellos induce la formacion o activacion de otro.En esta obra no se describe la secuencia completa de
acontecimientos, que son tratados en muchos textosde histologıa. Las principales etapas de la coagula-cion sanguınea son las siguientes:
. El endotelio danado deja de sintetizar factores queimpiden la agregacion plaquetaria y la coagulaciony comienza a secretar tromboplastina tisular,factor de von Willebrand y endotelina, la cualestimula la mitosis de las c�elulas endoteliales yejerce una accion vasoconstrictora.
. El factor de von Willebrand activa a las plaquetas,que se adhieren al col�ageno y la laminina quesobresalen hacia la luz del vaso sanguıneolesionado, se adhieren a otras plaquetas y liberan loscontenidos de sus gr�anulos.
. El difosfato de adenosina (ADP) y latromboplastina secretadas por las plaquetasincrementan su afinidad por otras plaquetas yprovocan la desgranulacion de las plaquetas reci�enadheridas.
. El plasmalema de las plaquetas activadas producetromboxano A2, derivado del �acido araquidonico,con acciones vasoconstrictora y activadora deplaquetas.
. Las plaquetas se agregan y expresan el factorplaquetario 3 en su membrana plasm�atica, el cualfavorece la precipitacion de los factores decoagulacion.
. En presencia de estos factores, la protrombina seconvierte en trombina, una reaccion catalizada porla tromboplastina derivada de las plaquetas y eltejido danado.
. La trombina cataliza la conversion de fibrinogenoen fibrina en una reaccion que requiere la presenciade calcio.
. El factor XIII entrecruza losmonomeros de fibrina yda lugar a un retıculo de co�agulo que se transformaen un trombo (co�agulo sanguıneo) conforme vaincorporando nuevas c�elulas sanguıneas yplaquetas.
. Alrededor de 1h despu�es, los monomeros de actinay miosina liberados por las plaquetas se agreganpara formar miofilamentos, que reducen el tamanodel co�agulo al contraerse y acercan los extremosseccionados del vaso, lo que reduce en mayormedida la p�erdida de sangre a partir del vasodanado.
. Tras la reparacion del vaso, el activador delplasminogeno convierteplasminogeno enplasmina,una proteasa que disuelve el co�agulo de fibrina demanera concertada con enzimas hidrolıticasprocedentes de gr�anulos lambda de las plaquetas.
140
10
SANGREYHEM
ATOPO
YESIS

141
10
SANGREYHEM
ATOPO
YESIS
Figura 10.4 Anatomıa de una plaqueta. (Tomado deGartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007,p 233.)
Tabla 10.5 TUBULOS Y GRANULOS DE LAS PLAQUETAS
Estructura (tamano) Localizacion Contenidos Funcion
Sistema de tubulos de
apertura en superficie
Hialomero Agiliza la captacion y liberacion de
mol�eculas por las plaquetas activadasSistema tubular denso Hialomero Es probable que secuestre iones de calcio
para evitar la «viscosidad» de las
plaquetas
Gr�anulos alfa (300–500 nm) Granulomero Fibrinogeno, factor de crecimientoderivado de plaquetas,
tromboplastina plaquetaria,
trombospondina, factores decoagulacion
Contienen factores que facilitan lareparacion vascular, la agregacion
plaquetaria y la coagulacion
sanguınea
Gr�anulos delta (cuerpos
densos) (250–300 nm)
Granulomero Calcio, ADP, ATP, serotonina,
histamina, pirofosfatasa
Contienen factores que potencian la
agregacion y la adhesion plaquetarias
y la vasoconstriccionGr�anulos gamma (lisosomas)
(200–250 nm)
Granulomero Enzimas hidrolıticas Contienen enzimas que facilitan la
resorcion del co�agulo
Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 236.
CONSIDERACIONES CL�INICAS
La vitamina K, un cofactor en la produccion deprotrombina y ciertos factores de coagulacion, sueleencontrarse a concentraciones bajas en losneonatos, por lo que habitualmente se administrauna inyeccion de esta vitamina para prevenir su
muerte debido a la enfermedad hemorr�agica delneonato. Los lactantes alimentados mediantelactancia natural y que no han recibido tal inyeccion
presentan un riesgo especialmente alto al presentarunas concentraciones excesivamente bajas de estefactor. Los adultos afectados por hipoabsorcion de
lıpidos tambi�en pueden padecer una deficiencia devitamina K, la cual se manifiesta con hemorragiaprofusa y formacion frecuente de hematomas. Laadministracion de un complemento de vitamina Ksuele aminorar los sıntomas.
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

M�edula osea
Lam�edula osea (fig. 10.5), un tejido conjuntivo muyvascularizado y gelatinoso que contiene numerosasc�elulas, ocupa las cavidades medulares de los huesosgrandes y lo espacios intratrabeculares de los hue-sos esponjosos y conforma un entorno propicio a lahematopoyesis, la formacion de c�elulas sanguıneas yplaquetas. En lactantes y sujetos jovenes, la m�edulaosea es activa en su totalidad y recibe el nombre dem�edula roja, ya que la mayorıa de las c�elulas que sedesarrollan en ella son eritrocitos. Hacia los 20 anosde edad, gran parte de la m�edula presente en las di�afi-sis de los huesos largos acumula lıpidos en cantidadestan grandes que se interrumpe la actividad hemato-poy�etica y se conoce comom�edula amarilla. La irriga-cion medular proviene de:
. Arterias que ingresan en la cavidadmedular a trav�esde canales de nutrientes.
. Un sistema de sinusoides grandes que desembocanen la vena longitudinal central, la cual drena en ungran numero de venas que abandonan la cavidadmedular a trav�es de los canales de nutrientes.
A diferencia de la mayorıa de las venas, las venasmedulares presentan un calibre menor que sus homo-logas arteriales y la presion hidrost�atica medular bastapara mantener la permeabilidad de los sinusoides. Elcomponente vascular de la m�edula engloba los vasos
sanguıneos y los sinusoides, y sus espacios intersti-ciales aparecen ocupados por grupos de c�elulas hema-topoy�eticas (islas de c�elulas hematopoy�eticas) queconfiguran el compartimento hematopoy�etico. Lossinusoides se rodean de:
. Una l�amina basal
. Una delgada capa de fibras reticulares
. C�elulas reticulares adventicias que est�an encontacto con dicha l�amina y recubren la mayorıa delas superficies de los sinusoides
Las extensiones citopl�asmicas de estas ultimas sealejan de los sinusoides y establecen contactos conlas proyecciones citopl�asmicas de otras c�elulas reti-culares adventicias, de tal modo que definen unosespacios en los que se alojan las islas hemato-poy�eticas (cordones hematopoy�eticos). Estos agre-gados de c�elulas hematopoy�eticas est�an presentes envarias etapas de su desarrollo, aunque suelen estarformados por un linaje celular determinado. Junto alas distintas c�elulas en maduracion, existen macro-fagos que destruyen los nucleos expulsados, fagoci-tan restos citopl�asmicos y proporcionan hierro a lasc�elulas de la serie eritrocıtica. Las c�elulas reticularesadventicias controlan el volumen de m�edula oseadisponible para la hematopoyesis; conforme se acu-mula lıquido la m�edula osea, su tamano aumenta yreduce el volumen del compartimento hematopo-y�etico.
142
10
SANGREYHEM
ATOPO
YESIS

143
10
SANGREYHEM
ATOPO
YESIS
Figura 10.5 Diagrama de lam�edula osea de un conejo a la que se ha inyectado carmın de litio y tinta china. (Modificado de Fawcett DW:
Bloom and Fawcett A Textbook of Histology, 12th ed. New York, Chapman & Hall, 1994, p 238.)
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

HEMATOPOYESIS
La hematopoyesis se divide en dos etapas: prenatal yposnatal. La hematopoyesis prenatal comienza alre-dedor del 14 dıa del desarrollo y consta de cuatrofases:
. La fase mesobl�astica se inicia con la formacion delos islotes sanguıneos; las c�elulas del borde de estosislotes se diferencian en c�elulas endoteliales, queforman los vasos sanguıneos, mientras que el restode la poblacion da lugar a eritroblastos queoriginar�an eritrocitos.
. La fase hep�atica sucede a la mesobl�astica hacia elfinal de la quinta semana despu�es de la concepcion;los eritrocitos son, aun, c�elulas nucleadas; eldesarrollo de los leucocitos comienza hacia laoctava semana del embarazo.
. La fase espl�enica se pone en marcha a lo largo delcuarto mes del desarrollo, y el bazo y el hıgadoconservan la actividad hematopoy�etica hasta elmomento del parto.
. La fase mieloide (fase de m�edula osea) se iniciahacia el sexto mes y adquiere una importancia cadavez mayor conforme se aproxima el momento delparto; despu�es del nacimiento, la actividad
hematopoy�etica se restringe a la m�edula osea,aunque el hıgado y el bazo pueden reanudar estaactividad en caso necesario.
La etapa posnatal de la hematopoyesis comienzaen el momento del parto y continua a lo largo de lavida del sujeto para producir innumerables c�elulassanguıneas. Lam�edula osea fabrica y sustituye 1.000mi-llones de c�elulas sanguıneas cada dıa por medio delproceso de hematopoyesis. Las c�elulas madre, querepresentan las c�elulas hematopoy�eticas con un gradom�as bajo de diferenciacion, se someten a divisionescelulares para originar c�elulas m�as diferenciadas, lasc�elulas progenitoras, que se dividen para formarc�elulas precursoras (tabla 10.6). No existe ningunadiferencia histologica entre las c�elulas madre y lasc�elulas progenitoras. Por el contrario, las c�elulas pre-cursoras pertenecen a linajes celulares especıficos, delque reciben su denominacion. Algunas c�elulas precur-soras conservan la capacidad de proliferacion, mien-tras que otras se comportan como c�elulas posmitoti-cas conforme maduran para transformarse en c�elulassanguıneas circulantes. El proceso hematopoy�eticoest�a sometido a una estrecha vigilancia y un estrictocontrol por parte de citocinas y factores de creci-miento.
144
10
SANGREYHEM
ATOPO
YESIS

145
10
SANGREYHEM
ATOPO
YESISTabla10.6
CELULASHEM
ATO
POYETICAS
BFU
,unidad
form
adora
debrotes(E,eritrocito);CFU
,unidad
form
adora
decolonias(E,eritrocito);CMHP,celulasmad
rehem
atopoyeticaspluripotenciales;G,gran
ulocito;
GEMM,gran
ulocito,eritrocito,monocito,megacariocito;GM,gran
ulocito-m
onocito;Linf,linfocito;LinfB,linfocito
B;LinfT,linfocito
T;M,monocito;Meg,megacarioblasto;neu
tro,
neu
trofilo.
Modificad
odeGartner
LP,HiattJL,Strum
J:Histology.Baltimore,William
s&Wilkins,1988.

C�elulas madre, c�elulas progenitoras y c�elulasprecursoras
Las c�elulas madre, llamadas c�elulas madre hemato-poy�eticas pluripotenciales (CMHP), de las que pro-vienen todas las c�elulas sanguıneas y las plaquetas, seencuentran en la m�edula osea, constituyen aproxima-damente el 0,1% de la poblacion medular de c�elulasnucleadas y se asemejan a los linfocitos. Las CMHPrara vez siguen el ciclo celular, pueden sufrir episodiossubitos de actividad mitotica para ampliar su pobla-cion. Las c�elulasmadre no se diferencian entre sı desdeel punto de vista morfologico, presentan marcadoresde membrana diferentes por los que son reconocidos.Las CMHP dan lugar a dos grupos de c�elulas madre(v. tabla 10.6), llamadas c�elulas madre hematopo-y�eticas multipotenciales (CMHM):
. Las c�elulas unidades formadores de colonias degranulocitos, eritrocitos, monocitos ymegacariocitos (CFU-GEMM) se encargan de laformacion de c�elulas progenitoras que dar�an lugar alos linajes mieloides.. BFU-E (unidad formadora de brote-eritrocitos),que originan CFU-E y, a continuacion,eritrocitos.
. CFU-Meg, que dan lugar a losmegacariocitos quese diferenciar�an en plaquetas.
. CFU-Eosinofilos, que se transformar�an eneosinofilos.
. CFU-Basofilos, que se convertir�an en basofilos.
. CFU-GM, que formar�an CFU-G y CFU-M, lascuales originar�an neutrofilos y monocitos.
. Las unidades formadoras de colonias delinfocitos (CFU-Linf) se ocupan de la formacionde las c�elulas progenitoras de los linajeslinfoides, CFU-LinfT (linfocitos T) y CFU-LinfB(linfocitos B).
. A diferencia de las c�elulas madre y las c�elulasprogenitoras, las c�elulas precursoras sonincapaces de regenerarse (es decir, no puedenproducir nuevas c�elulas precursoras) y carecen derasgos histologicos claros que permitan suidentificacion como predecesoras de alguna c�elulasanguınea circulante (fig. 10.6, v. tabla 10.6).Las primeras c�elulas precursoras de cada linajecelular son:. Proeritroblastos, que dan lugar a los eritrocitos. Megacarioblastos, que originan las plaquetas. Mieloblastos, una excepcion a la regla anterior, yaque pueden reconocerse como los precursores deneutrofilos, eosinofilos y basofilos
. Promonocitos, que se diferencian en monocitos
. Linfocitos T vırgenes
. Linfocitos B vırgenes
146
10
SANGREYHEM
ATOPO
YESIS

147
10
SANGREYHEM
ATOPO
YESIS
Figura 10.6 C�elulas precursoras de las series eritrocıtica y granulocıtica. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology,
3rd ed. Philadelphia, Saunders, 2007, p 240.)
CONSIDERACIONES CL�INICAS
La mielofibrosis es un trastorno debido a la sıntesis
excesiva de tejido conjuntivo fibroso por los fibroblastos
de la m�edula osea en lugar de producir delgadas fibras
de col�ageno para reforzar los vasos sanguıneos, los
sinusoides, y los islotes sanguıneos
de la m�edula osea. Conforme se acumula el tejido
fibroso, la m�edula osea se rellena de material fibroso
de tal modo que disminuye el volumen disponible
anteriormente para la hematopoyesis y se reduce la
eritropoyesis, con lo que el paciente se torna an�emico.
La formacion de leucocitos puede aumentar o disminuir,
mientras que el desarrollo de plaquetas se restringe.
Esta infrecuente enfermedad, que afecta a uno de cada
50.000 sujetos en EE. UU., suele afectar
a personas de edades comprendidas entre
50 y 70 anos.
A menudo, no se puede identificar el origen de la
enfermedad (mielofibrosis idiop�atica) o bien acompana a
otras mielopatıas o infecciones de m�edula osea. Muchos
pacientes aquejados de esta entidad presentan
exposicion a radiacion ionizante o benceno. En su estadio
inicial, la mielofibrosis es una entidad asintom�atica y no se
manifiesta hasta que la anemia provoca un descenso de
los niveles de energıa, disminucion del peso corporal,
debilidad ymalestar general. La disminucion concomitante
de las poblaciones de leucocito y plaquetas incrementa la
susceptibilidad a la infeccion, el desarrollo de petequias y
la formacion de hematomas. La m�edula osea no puede
realizar la hematopoyesis normal, por lo que el bazo y el
hıgado se hipertrofian al asumir esta funcion, lo que origina
dolor abdominal. El diagnostico solo se confirmamediante
la obtencion de una biopsia medular. Los sujetos
afectados por esta entidad pueden vivir 10 anos o m�as, si
bien en algunos casos la progresion es muchom�as r�apida
(mielofibrosis aguda o maligna). No se dispone de ningun
tratamiento curativo frente a esta enfermedad, aunque el
trasplante de m�edula puede reducir en cierta medida su
sintomatologıa.
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

Factores de crecimiento hematopoy�eticos(factores estimuladores de colonias)
Algunas c�elulas del organismo sintetizan un grannumero de glucoproteınas que estimulan la hemato-poyesis. Estos factores de crecimiento hemato-poy�eticos (factores estimuladores de colonias)actuan sobre sus c�elulas diana como hormonas endo-crinas, hormonas paracrinas o bien a trav�es del con-tacto intercelular. Cada uno de ellos estimula la pro-liferacion, diferenciacion, o ambas, de una c�elulamadre, c�elula progenitora o c�elula precursora especı-fica, de modo que esa poblacion celular mantenga unnivel normal en la sangre circulante (tabla 10.7).
. Factor de Steel (factor de c�elulas madre), factorestimulador de colonias de monocitos, IL-3 e IL-7estimulan la mitosis en las CMHP, CFU-GEMM yCFU-Linf con el fin de mantener sus densidadespoblacionales.
. El factor estimulador de colonias degranulocitos, el factor estimulador de coloniasde monocitos, la IL-2, la IL-5, la IL-6, la IL-11,
la IL-12, la proteına inhibidora de macrofagos-ay la eritropoyetina inducen la diferenciacion delas CMHP, CFU-GEMM y CFU-Linf en c�elulasprogenitoras (v. tabla 10.7).
Por otra parte, los factores estimuladores de colo-nias estimulan la formacion de neutrofilos, eosinofi-los, basofilos y monocitos a partir de c�elulas precur-soras unipotenciales; la eritropoyetina induce el desa-rrollo de los eritrocitos; y la trombopoyetina incita laformacion de las plaquetas.
El factor de Steel, sintetizado por las c�elulas estro-males de la m�edula osea, se expresa en la membranaplasm�atica de esta poblacion celular. La activacion delas CMHP, CFU-GEMM y CFU-Linf requiere de suinteraccion con el factor de Steel presente en dichamembrana. La hematopoyesis tan solo puede tenerlugar en las �areas en las que las c�elulas estromalesexpresan este factor en su plasmalema. En ausenciade contacto con los factores de crecimiento hemato-poy�eticos, las c�elulas hematopoy�eticas sufren apopto-sis, mueren y son fagocitadas por los macrofagos.
148
10
SANGREYHEM
ATOPO
YESIS

149
10
SANGREYHEM
ATOPO
YESISTabla 10.7 FACTORES DE CRECIMIENTO HEMATOPOYETICOS
Factores Accion principal Origen
Factor de c�elulas madre Favorece la hematopoyesis C�elulas estromales de m�edula osea
CSF-GM Induce la mitosis y la diferenciacion de
las CFU-GM; facilita la actividad granulocıtica
Linfocitos T; c�elulas endoteliales
CSF-G Induce la mitosis y la diferenciacion de las CFU-G;
facilita la actividad neutrofila
Macrofagos, c�elulas endoteliales
CSF-M Induce la mitosis y la diferenciacion de las CFU-M Macrofagos, c�elulas endotelialesIL-1 Junto a IL-3 e IL-6, favorece la proliferacion de
CMHP, CFU-GEMM y CFU-Linf; inhibe a los
precursores eritroides
Monocitos; macrofagos; c�elulas endoteliales
IL-2 Estimula la mitosis de linfocitos T activados ylinfocitos B; induce la diferenciacion de
linfocitos NK
Linfocitos T activados
IL-3 Junto a IL-1 e IL-6, favorece la proliferacion de
CMHP, CFU-GEMM, CFU-Linf y todos losprecursores unipotenciales (salvo los de
linfocitos B y T)
Linfocitos T y B activados
IL-4 Estimula la activacion de linfocitos T y B, ası comoel desarrollo de mastocitos y basofilos
Linfocitos T activados
IL-5 Favorece la mitosis de las CFU-Eo y activa a los
eosinofilos
Linfocitos T
IL-6 Junto a IL-1 e IL-3, favorece la proliferacion deCMHP, CFU-GEMM, CFU-Linf; igualmente,
estimula la diferenciacion de LTC y linfocitos B
Monocitos y fibroblastos
IL-7 Favorece la diferenciacion de CFU-Linf; potencia la
diferenciacion de los linfocitos NK
C�elulas estromales
IL-8 Induce la migracion y la desgranulacion de
neutrofilos
Leucocitos, c�elulas endoteliales, y c�elulas de
musculo liso
IL-9 Induce la activacion y la proliferacion demastocitos; modula la sıntesis de IgE; favorece la
proliferacion de linfocitos T cooperadores
Linfocitos T cooperadores
IL-10 Inhibe la produccion de citocinas por los
macrofagos, linfocitos T y linfocitos NK; facilitala diferenciacion de LTC y la proliferacion de
linfocitos B y mastocitos
Macrofagos y linfocitos T
IL-12 Estimula a linfocitos NK; potencia la funcion de
LTC y linfocitos NK
Macrofagos
Interferones g Activa a linfocitos B y monocitos; potencia la
diferenciacion de LTC; incrementa la expresion
de HLA de clase II
Linfocitos T y linfocitos NK
Eritropoyetina Diferenciacion de CFU-E; mitosis de BFU-E C�elulas endoteliales de red capilar peritubularrenal; hepatocitos
Trombopoyetina Proliferacion y diferenciacion de CFU-Meg y
megacarioblastos
Desconocido
BFU, unidades formadoras de brotes (E, eritrocito); CFU, unidad formadora de colonias (Eo, eosinofilo; G, granulocito;
GEMM, granulocito, eritrocito, monocito, megacariocito; GM, granulocito-monocito; Linf, linfocito); CMHP, c�elula madrehematopoy�etica pluripotencial; CSF, factor estimulador de colonias (G, granulocito; GM, granulocito-monocito; M, monocito);
IL, interleucina; LTC, linfocito T citotoxico; NK, citotoxico natural.Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 242.
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

Eritropoyesis, granulocitopoyesis, monocitopoyesisy linfopoyesis
La formacion de los eritrocitos, conocida como eritro-poyesis, depende de dos clases de c�elulas progenitorasparaproducir2,5�1011 eritrocitosdiarios. Si elnumerode eritrocitos esmenorde la cantidadnormal, las c�elulasendoteliales de los capilares peritubulares renales y loshepatocitos secretan eritropoyetina. De manera con-junta con el factor de Steel, IL-3, IL-9 y el factor estimu-lador de colonias de granulocitos-macrofagos, este fac-tor estimula la diferenciacion de las CFU-GEMM en ungran numero de BFU-E, las cuales se dividen para origi-narunapoblacion aunm�as amplia deCFU-E. La sıntesisde eritropoyetina en las c�elulas renales y hep�aticas seinterrumpe tras la formacion de estas ultimas, demodoque las bajas concentraciones de este factor estimulan eldesarrollo de proeritroblastos a partir de las CFU-E. Enla tabla 10.8 se enumeran las c�elulas pertenecientes a laserie eritrobl�astica y sus caracterısticas.
La formacion de los granulocitos, la granulocito-poyesis, se inicia a partir de las CFU-GM, que dan lugara otras dos clases de c�elulas progenitoras: CFU-M,responsables del desarrollo de los monocitos, y CFU-G,encargadas del de los neutrofilos. Los eosinofilos y losbasofilos provienen de las CFU-Eo y CFU-Ba. Los fa-ctores IL-1, IL-6 y TNF-a suscitan la liberacion de losfactores de crecimiento factor estimulador de las colo-nias de granulocitos, factor estimulador de las colo-nias de granulocitos-monocitos, e IL-5, los cuales esti-mulan el desarrollo de los neutrofilos, los eosinofilosy los basofilos. La primera c�elula reconocible desde el
punto de vista morfologico del linaje granulocıtico eselmieloblasto y la segunda es el promielocito. Tantolos mieloblastos como los promielocitos carecen degr�anulos especıficos y los tres tipos de granulocitosprovienen de estos precursores. La siguiente c�elula deeste linaje posee gr�anulos especıficos y caracterısticasque hacen posible su identificacion como mielocitosneutrofilos, eosinofilos o basofilos. Las c�elulas de laserie neutrofila se muestran en la tabla 10.9.
La c�elula progenitora de la monocitopoyesis es laCFU-GM bipotencial, la cual se diferencia en la CFU-M unipotencial, de los que proceden los promonoci-tos. Estas c�elulas originanmonocitos, los cuales pasanal torrente circulatorio.
Las plaquetas provienen de la CFU-Meg, que sediferencia enmegacarioblastos; estas c�elulas se hiper-trofian a trav�es de endomitosis sucesivas, en el trans-curso de las cuales la c�elula sufre unamitosis que no seacompana de citocinesis y da lugar a unas c�elulas degran tamano, los megacariocitos. Estas enormesc�elulas residen en las proximidades de los sinusoides,hacia cuya luz extienden su citoplasma. Las proyec-ciones citopl�asmicas se fragmentan a lo largo de loscanales de demarcacion y liberan proplaquetas enestos vasos. Las proplaquetas se convierten en plaque-tas antes de pasar al torrente circulatorio.
La linfopoyesis se pone en marcha con la c�elulamadre CFU-Linf, que origina las c�elulas progenitorasCFU-LinfT y CFU-LinfB. Estas c�elulas se diferencian enlinfocitos T vırgenes (CFU-LinfT) y linfocitos B vır-genes (CFU-LinfB).
150
10
SANGREYHEM
ATOPO
YESIS
Tabla 10.8 CELULAS DE LA SERIE ERITROPOYETICA
C�elulaTamano(mm) Nucleo* y mitosis Nucl�eolos Citoplasma*
Microfotografıaelectronica
Proeritroblasto 14-19 Redondeado, colorburdeos; fina red de
cromatina; mitosis
3-5 Gris-azul, agregados enperiferia
RER poco desarrollado;numerosos polisomas,
escasas mitocondrias;
ferritina
Eritroblastobasofilo
12-17 Igual que el anterior,aunque red de
cromatinam�as gruesa;
mitosis
¿1-2? Semejante al anterior,aunque coloracion
rosada poco intensa
Semejante al anterior;presencia de
hemoglobina
Eritroblastopolicromatofilo
12-15 Redondeada, tincionintensa; red de
cromatina muy
gruesa; mitosis
Ninguno Rosa amarillento sobreun trasfondo azulado
Similar al anterior;mayor cantidad de
hemoglobina
Eritroblastoortocromatofilo
8-12 Pequeno, redondo,denso; exc�entrico o en
proceso de expulsion;
ausencia de mitosis
Ninguno Rosa en un trasfondoazulado poco intenso
Escasas mitocondrias ypolisomas, abundante
hemoglobina
Reticulocito 7-8 Ninguno Ninguno Semejante a eritrocitos
maduros, pero en
tincion con azul de
cresilo muestra unretıculo azulado
Grupos de ribosomas;
c�elula rellena de
hemoglobina
Eritrocito 7,5 Ninguno Ninguno Citoplasma rosado Unicamente
hemoglobina
*Los colores aparecen al utilizar tinciones de tipo Romanovsky.RER, retıculo endopl�asmico rugoso.Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 246.

151
10
SANGREYHEM
ATOPO
YESIS
Tabla10.9
CELULASDELA
SER
IENEU
TROFILA
C� elula
Taman
o(mm)
Nucleo
*ymitosis
Nucl� eo
los
Citoplasm
a*
Gr� anulos
Microfotografıas
electronicas
Mieloblasto
12-14
Red
ondeado,azul
rojizo
;finaredde
cromatina;
mitosis
2-3
Agregad
osazuladosen
trasfondoazulclaro;
vesıculas
citopl� asm
icas
enperiferia
celular
Ningu
no
RER,Golgipoco
desarrollad
o,
much
as
mitocondrias
ypolisomas
Promielocito
16-24
Deredondeadoa
ovalado,azulrojizo
;
reddecromatina
gruesa;
mitosis
1-2
Citoplasm
aazulado;
ausenciadevesıculas
citopl� asm
icas
enperiferia
Gr� anulosazurofilos
RER,Golgiextenso,
much
as
mitocondrias,
abundan
tes
lisosomas
Mielocito
neu
trofilo
10-12
Aplanad
o,ac� en
trico;
reddecromatina
gruesa;
mitosis
0-1
Citoplasm
aazulclaro
Gr� anulosazurofilosy
especıficos
RER,Golgi
desarrollad
o,
numerosas
mitocondrias,
lisosomas
ygr� an
ulosespecıficos
Metam
ielocito
neu
trofilo
10-12
Conform
aarrinonad
a,
den
so;redde
cromatinagruesa;
ausenciademitosis
Ningu
no
Citoplasm
aazulclaro
Gr� anulosazurofilosy
especıficos
Numeroreducidode
org� an
ulos,au
nque
gr� an
ulossemejan
tes
alan
terior
Ban
daneu
trofila
(estab
le;juvenil)
9-12
Form
adeherradura;red
decromatinamuy
gruesa;
ausenciade
mitosis
Ningu
no
Citoplasm
aazulclaro
Gr� anulosazurofilosy
especıficos
Id� en
tico
alan
terior
Neu
trofilo
9-12
Multilobulado;redde
cromatinamuy
gruesa;
ausenciade
mitosis
Ningu
no
Rosa
azuladoclaro
Gr� anulosazurofilosy
especıficos
Id� en
tico
alan
terior
*Loscoloresaparecenalu
tilizartincionesdetipoRomanovsky
(omodificacionesdelasmismas).RER,retıculo
endopl� asm
icorugoso
.Tomad
odeGartner
LP,HiattJL:Color
Textbookof
Histology,3rd
ed.Philadelphia,Saunders,2007,p248.

11 APARATOCIRCULATORIO
Aparato cardiovascular
El aparato cardiovascular est�a formado por un cora-zon con cuatro c�amaras que se dividen en c�amarasauriculares derecha e izquierda (derecepcion de sangre) y c�amaras ventri-culares derecha e izquierda (de expul-sion de sangre). El hemicardio dere-cho, el cual contiene la aurıculaderecha y el ventrıculo derecho, con-forma el circuito pulmonar que trans-porta sangre hasta los pulmones parasu oxigenacion y la liberacion del di-oxido de carbono. La sangre oxigenadaregresa al hemicardio izquierdo (cir-cuito sist�emico) del corazon y se bom-bea desde el ventrıculo izquierdo paradistribuirse a los tejidos corporales.
Los vasos que forman parte del apa-rato cardiovascular son los siguientes
. Arterias, que salen del corazon y conducen la sangrehacia otras regiones del organismo; su calibredisminuye conforme se ramifican al alejarse delcorazon.
. Venas, a trav�es de las cuales la sangre regresa alcorazon; los vasos de menor calibre son los m�asalejados de este organo, mientras que los de mayorcalibre se encuentran cerca del mismo.
. Capilares, los vasos m�as pequenos y con paredesm�as delgadas, conectan los sistemas arterial yvenoso; posibilitan el intercambio de mol�eculasentre las c�elulas y el torrente circulatorio. Loscapilares reciben sangre procedente de arterias demenor calibre, las arteriolas (y lasmetaarteriolas),que ceden a las venas m�as pequenas, las v�enulas.
TUNICAS DE LOS VASOS
Las paredes de las arterias y las venas se componen detres capas: tunica ıntima, tunica media y tunicaadventicia (fig. 11.1).
. La capa m�as interna de las tunicas, la tunica ıntima,consta de un epitelio escamoso simple y unendotelio que tapiza la luz del vaso.
. Lamembrana basal se dispone entre el endotelio yel tejido conjuntivo subendotelial.
. En las arterias musculares, el tejido conjuntivosubendotelial alberga algunas c�elulas de
musculo liso.. El tejido conjuntivo subendotelial serodea de una l�amina el�asticainterna, una membrana el�asticaperforada formada esencialmentepor elastina.
. En los cortes transversales de vasosde pequeno calibre, como loscapilares, bastan una o dos c�elulasendoteliales para envolver la luz delvaso, mientras que se requierendocenas de estas c�elulas en los demayor tamano.
. Las c�elulas endoteliales crean unasuperficie lisa, exenta de friccionesy secretan numerosas mol�eculas,
como lamina; endotelina; col�ageno de tipo II, IV yV; oxido nıtrico (NO) y factor de von Willebrand(vWF).
. En su cara luminal, las membranas de las c�elulasendoteliales presentan la enzima conversora deangiotensina y otras enzimas que inactivan a unamplio abanico de sustancias de transportehem�atico, como bradicininas, trombina,prostaglandinas y serotonina.
. La lipasa de lipoproteınas se une a la superficieluminal de las membranas de las c�elulasendoteliales para escindir estas proteınas.
. La capa m�as gruesa, en especial en las arterias, esla tunica media, formada por varias capas dec�elulas de musculo liso en una disposicionhelicoidal. La matriz extracelular de la tunicamedia consta de fibras el�asticas integradas porc�elulas de musculo liso, fibras de col�ageno de lostipos I y III, y sustancia fundamental. En la capaexterna de la tunica media, al menos en lasgrandes arterias musculares, se alojan fibrasel�asticas delgadas, que conforman la l�aminael�astica externa. Los capilares poseen unasc�elulas contr�actiles denominadas pericitos quesustituyen a la tunica media.
� 2011 Elsevier Espana, S.L. Reservados todos los derechos
TERMINOS CLAVE. Tunicas de los vasos. Arterias. Arteriolas. Regulacion de la
presion arterial. Capilares. Venas. Corazon. Vasos linf�aticos
152

CONSIDERACIONES CL�INICAS
Las c�elulas endoteliales fabrican una proteınaespecıfica, el factor de von Willebrand (vWF),un factor de coagulacion, aunque solamente sealmacena en los cuerpos de Weibel-Palade de lasarterias. El vWF favorece la coagulacion sanguınea alunirse a las plaquetas en el transcurso del procesode coagulacion. La enfermedad de vonWillebrand esuna coagulopatıa hereditaria que altera lacoagulacion sanguınea. Suele deberse a unadeficiencia del vWF o a alguna anomalıa en el mismo.
Un aneurisma es una deformacion en globo de lapared de una arteria (o, de manera menos frecuente,
una vena) causada por el debilitamiento de la pareddel vaso. Generalmente, los aneurismas se vinculancon el envejecimiento, como sucede en laateroesclerosis, o bien pueden deberse a otrostrastornos, como el sındrome deMarfan, el sındromede Ehlers-Danlos y la sıfilis. Los aneurismas pueden
afectar a diversas arterias, si bien son m�asfrecuentes en la aorta abdominal. El diagnosticoprecoz hace posible la reparacion de la anomalıa;pero la rotura de un aneurisma no detectado
ocasiona una p�erdida masiva de sangre queconduce a la muerte del paciente.
153
11
APARATO
CIRCULATORIO
Figura 11.1 Arteria tıpica. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 252.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

. El revestimiento m�as externo, la tunica adventicia,est�a formado por tejido conjuntivo fibroel�asticoque fija los vasos sanguıneos a las estructurasadyacentes (fig. 11.2).. Los nutrientes y el oxıgeno presentes en el torrentecirculatorio no pueden atravesar la pared de losvasos sanguıneos de gran calibre; los vasavasorum, arterias pequenas, penetran la tunicaadventicia, se ramifican en la pared del vaso yaportan nutrientes y oxıgeno a las c�elulaslocalizadas en la tunica adventicia y la tunicamedia. Los vasa vasorum est�an m�as desarrolladosen las venas que en las arterias.
. La inervacion de los vasos sanguıneos puedeatravesar la tunica adventicia; los nerviosvasomotores secretan el neurotransmisornoradrenalina, que se difunde hacia las c�elulas demusculo liso de la tunica media. Se trata de fibrasvasomotoras simp�aticas que estimulan lacontraccion de las c�elulas de musculo liso y laonda de contraccion viaja a trav�es de las unionesgap entre c�elulas vecinas de musculo liso paradesencadenar la vasoconstriccion.
ARTERIAS
Las arterias (tabla 11.1) son unos vasos sanguıneosmusculares de gran calibre cuyo di�ametro disminuyepaulatinamente conforme se alejan del corazon parasuministrar sangre a los lechos capilares. A pesar deque las definiciones no son claras, se distinguen tresclases de arterias segun su di�ametro, grosor de la paredy otros rasgos histologicos:
. Arterias el�asticas (de conduccion), demayor calibre.
. Arteriolas, cuyo calibre es el m�as pequeno.
. Arterias musculares (de distribucion), de calibrecomprendido entre los de las anteriores.
Estructuras sensoriales especializadas de las arterias
Las arterias musculares est�an dotadas de unos organossensoriales especializados, el seno carotıdeo y elcuerpo carotıdeo, mientras que el cayado aorticoposee una estructura sensorial equivalente, el cuerpoaortico.
. El seno carotıdeo, localizado en la tunica adventiciade la arteria carotida interna, recibe su inervaciondel nervio craneal IX (nervio glosofarıngeo) y actuacomo barorreceptor al vigilar la presion arterial. La
informacion procedente del seno se conduce haciael centro vasomotor, que genera una respuestaencaminada a la conservacion de unos valoresnormales de presion arterial.
. El cuerpo carotıdeo, un organo quimiorreceptor depequeno tamano que disfruta de una excelenteirrigacion por los lechos capilares, se encuentra en labifurcacion de la arteria carotida primitiva y recibesu inervacion de los nervios craneales IX y X (nerviosglosofarıngeo y vago). Responde a las variaciones delas concentraciones sanguıneas de CO2, O2 y H
+. Enlas im�agenes de microscopia electronica se puedenidentificar dos tipos celulares:. El citoplasma de las c�elulas glomicas (c�elulasde tipo I) contiene gr�anulos de catecolaminas yemite proyecciones que interaccionan con lasc�elulas endoteliales de los capilares y las c�elulasglomicas vecinas.
. Las extensiones de las c�elulas de la vaina(c�elulas de tipo II) envuelven las proyeccionesde las primeras y sustituyen la envoltura dec�elulas de Schwann de las fibras nerviosasdesnudas que atraviesan los grupos de c�elulasglomicas.
. Los cuerpos aorticos, localizados en el cayadoaortico, remedan a los cuerpos carotıdeos desdeel punto de vista morfologico y funcional.
Regulacion de la presion arterial
Las vıas neural y bioquımica controlan la presionarterial.
. El centro vasomotor encef�alico se encargade mantener una presion arterial adecuada de90-119/60-79mmHg a trav�es de la vıa neural, loque logra al conservar un tono constante de lasc�elulas de musculo liso de la tunica media de losvasos sanguıneos.. La disminucion de la presion arterial favorece lacontraccion muscular mediante la liberacion delneurotransmisor noradrenalina por parte delsistema nervioso simp�atico.
. El sistema nervioso parasimp�atico reduce eltono muscular al secretar el neurotransmisoracetilcolina en caso de aumento excesivo de lapresion arterial; este neurotransmisor inducela liberacion de NO por parte de las c�elulasendoteliales. Las c�elulas de musculo liso de latunica media se relajan al recibir este mediador.
154
11
APARATO
CIRCULATORIO

155
11
APARATO
CIRCULATORIO
Figura 11.2 Arteria tıpica. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 252.)
Tabla 11.1 CARACTER�ISTICAS DE LOS DISTINTOS TIPOS DE ARTERIAS
Arteria Tunica ıntima Tunica media Tunica adventicia
El�astica (conduccion)
(p. ej., aorta, tronco
pulmonar y arterias)
Endotelio con cuerpos de
Weibel-Palade, l�amina
basal, capa subendotelial,l�amina el�astica interna
incompleta
40-70 membranas el�asticas
fenestradas, c�elulas de
musculo liso mezcladas conmembranas el�asticas,
l�amina el�astica externa
delgada, vasa vasorum en
superficie externa
Delgada capa de tejido
conjuntivo fibroel�astico,
vasa vasorum, vasoslinf�aticos, fibras nerviosas
Arteria muscular
(distribucion)
(p. ej., arterias carotidas,
arteria femoral)
Endotelio con cuerpos de
Weibel-Palade, l�amina
basal, capa subendotelial,
l�amina el�astica internagruesa
� 40 capas de c�elulas de
musculo liso, l�amina
el�astica externa gruesa
Capa delgada de tejido
conjuntivo fibroel�astico,
vasa vasorum poco
desarrollados, vasoslinf�aticos, fibras nerviosas
Arteriola Endotelio con cuerpos de
Weibel-Palade, l�aminabasal, capa subendotelial
poco desarrollada, algunas
fibras el�asticas en lugar de
una l�amina el�astica definida
1-2 capas de c�elulas
de musculo liso
Tejido conjuntivo laxo, fibras
nerviosas
Metaarteriola Endotelio, l�amina basal C�elulas de musculo liso que
forman el esfınter precapilar
Tejido conjuntivo laxo
disperso
Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 254.
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

. Los rinones y la hipofisis rigen la vıa bioquımica.. Los rinones liberan la enzima renina al torrentecirculatorio. Esta enzima escinde elangiotensinogeno circulante en angiotensina I,que se transforma en angiotensina II, unamol�ecula con una potente accion constrictora delmusculo liso de la tunica media, por accion de laenzima conversora de angiotensina de lamembrana de la luz del endotelio capilar.
. La hipofisis secreta un potente vasoconstrictor,la vasopresina (hormona antidiur�etica).
Asimismo, las membranas el�asticas de lasarterias musculares de gran calibre y, especialmente,en las arterias el�asticas, intervienen en la regulacionde la presion arterial.
. Al contraerse los ventrıculos del corazon, bombeanun gran volumen de sangre hacia la aorta y lasarterias pulmonares, en cuyas paredes abundan lasfibras el�asticas y las membranas el�asticas(membranas fenestradas). La pared de los vasos seexpande, la membrana el�astica se distiende yrecupera de manera lenta su tamano normal, demodo que se evitan las variaciones r�apidas develocidad del flujo sanguıneo y la presion arterial.
CAPILARES
Los capilares (fig. 11.3) son los vasos sanguıneos demenor calibre y paredesm�as delgadas. Se componen deun epitelio escamoso simple dispuesto en una estruc-tura tubular de menos de 50mm de longitud y 8 a10mmde di�ametro. Las zonas de contacto de las c�elulasendoteliales consigo mismas u otras c�elulas endote-liales en la estructura tubular dan lugar a una pequenasolapa, el pliegue marginal, que proyecta hacia la luz.De igual modo, las c�elulas endoteliales forman fasciasoclusivas (uniones estrechas). Los capilares crean unared anastomosada, llamada lecho capilar, que se inter-pone entre las arteriolas y las v�enulas.
. Las c�elulas endoteliales capilares son muy delgadas;su grosor es menor de 0,2mm y sus nucleos formanextensiones que se proyectan hacia la luz del vaso.
. En el citoplasma aparece un reducido numero de losorg�anulos habituales y filamentos intermediosformados por desmina, vimentina o ambas.
. Una caracterıstica distintiva de los capilares es laabundancia de vesıculas pinocıticas asociadas a sumembrana plasm�atica.
. Los capilares forman una l�amina basal que revistesu superficie abluminal.
. Los pericitos, unas c�elulas contr�actiles asociadas alos capilares y las v�enulas pequenas, comparten lal�amina basal de los capilares, forman uniones gapcon las c�elulas endoteliales y pueden colaborar en laregulacion del flujo sanguıneo. Asimismo, puedenactuar como c�elulas regenerativas que colaboranen la reparacion de vasos danados.
Se diferencian tres tipos de capilares en las micro-fotografıas electronicas:
. Capilares continuos, que se localizan en el tejidoconjuntivo, el musculo y el tejido nervioso, ycapilares continuos modificados, que se restringenal enc�efalo. Los capilares continuos presentanmuchas vesıculas pinocıticas y las unionesintercelulares son fascias oclusivas, de modo que eltransporte de amino�acidos, glucosa, nucleosidos ypurinas depende de la intervencion de proteınastransportadoras. A pesar de que las c�elulasendoteliales regulan la barrera hematoencef�alica,los astrocitos tambi�en parecen intervenir en esteproceso.
. Capilares fenestrados, propios de las gl�andulasendocrinas, el p�ancreas y el intestino, poseenfenestraciones (poros, 60 a 80nm de di�ametro)en sus paredes que se recubren de un diafragma.Estos complejos diafragma/poro se disponen aintervalos de 50 nm, si bien pueden organizarseen agregados.
. Capilares sinusoides, presentes en la m�edulaosea, el bazo, el hıgado, los ganglios linf�aticos yalgunas gl�andulas endocrinas, son canales amorfos(sinusoides) tapizados por c�elulas endotelialesdotadas de un gran numero de fenestracionesgrandes carentes de diafragmas. En algunoscasos, la l�amina basal de la pared endotelialpuede presentar discontinuidades, lo quefacilita en gran medida el intercambio demol�eculas entre el torrente circulatorio y lostejidos.
156
11
APARATO
CIRCULATORIO

CONSIDERACIONES CL�INICAS
CAMBIO VASCULARLas arterias de mayor calibre continuan creciendohasta los 25 anos de edad mediante la adicion
continuada de l�amina el�astica a sus paredes. A partirde la edad madura, las paredes de las arterias
musculares se engrosan con col�ageno yproteoglicano, lo que ocasiona una reduccion de suflexibilidad. Los vasos coronarios son los primerosen mostrar cambios relacionados con elenvejecimiento, en particular a nivel de la tunicaıntima. Estas alteraciones son semejantes a lasobservadas en la arterioesclerosis.
ARTERIOESCLEROSISA menudo, la arterioesclerosis se vincula conhipertension y diabetes. Esta enfermedad sedistingue por la acumulacion de una sustanciahialina en las paredes de la tunica media de lasarterias pequenas y las arteriolas (especialmente,
en los rinones). La calcificacion de dichas paredesprovoca rigidez en los vasos afectados.
ATEROESCLEROSISLa ateroesclerosis es la causa m�as frecuente demorbilidad en las afecciones vasculares; secaracteriza por la formacion de depositos deplacas lipıdicas amarillentas sin componentecelular (ateromas) en las paredes de la ıntima, lo
cual da lugar a una reduccion del di�ametro de la luzde las arterias coronarias (p. ej., arterias carotidas)y las arterias grandes del cerebro. La acumulacion
de este material estrecha el di�ametro de la luz ydificulta el flujo sanguıneo hacia la region afectadaalrededor de los 25 anos de edad. El dolor quesiente el paciente puede preceder a un infarto demiocardio y un accidente cerebrovascular cuandola restriccion del flujo sanguıneo se da en los vasoscoronarios.
157
11
APARATO
CIRCULATORIO
Figura 11.3 A-C. Tres tipos de capilares. (Tomado deGartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007,
p 262.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Regulacion del flujo sanguıneo en un lecho capilar
La regulacion del flujo sanguıneo en un lecho capilarse lleva a cabo a trav�es de las anastomosis arteriove-nosas (AAV) y los canales centrales (fig. 11.4).
. Las AAV derivan los lechos capilares, en los queexiste una conexion directa entre los extremosarteriales y venosos. El vaso conector posee tresregiones: un extremo arterial, un extremo venoso yun segmento intermedio. El segmento intermediopresenta:. Una tunica media engrosada y c�elulasmodificadas de musculo liso en la capasubendotelial, y
. Una inervacion adren�ergica y colin�ergica biendesarrollada que se encuentra bajo el controldirecto del centro termorregulador encef�alico.
. La apertura o el cierre de estas derivaciones AAVcontrola la irrigacion.. Al cerrarse la derivacion, la sangre fluye connormalidad a trav�es del lecho capilar.
. Al abrirse la derivacion, la sangre fluye a trav�esde esta y deriva el lecho capilar.
Las AAV se distribuyen en todo el organismo,si bien son m�as abundantes en la piel, en la queintervienen en la regulacion t�ermica.
. Los canales centrales est�an formados por unametaarteriola y su continuacion, el canal dedesag€ue.. Las metaarteriolas, que provienen de lasarteriolas, presentan esfınteres precapilares quepermiten el flujo de sangre en el lecho capilar alabrirse.
. La sangre procedente del lecho capilar pasa a loscanales de desag€ue; estos conductos carecen deesfınteres, por lo que siempre pueden recibirsangre, que se distribuye a las v�enulas pequenas.
Histofisiologıa de los capilares
El estudio fisiologico de la permeabilidad de los capi-lares puso de manifiesto la existencia de dos tipos deporos en sus paredes (fig. 11.5): poros pequenos, quepodrıan corresponder a pequenas interrupcionesentre uniones de c�elulas epiteliales (9 a 11nm dedi�ametro), y poros grandes, que representarıan fenes-traciones y vesıculas de transporte (50 a 70nm de di�a-metro).
. Las mol�eculas pequenas pueden difundir a trav�esde la c�elula endotelial o de uniones intercelulares.
. Las mol�eculas de mayor tamano pasan del espacioextracelular hacia la luz (o viceversa) a trav�es devesıculas pinocıticas, un proceso llamadotranscitosis.
. Otras sustancias, como las que se encuentran en elaparato de Golgi de las c�elulas endoteliales, setransportan a la superficie luminal de la membranaplasm�atica en vesıculas recubiertas de clatrina, enlas que su contenido es sustituido por otro diferente,que se transporta hasta la cara abluminal de lamembrana celular para secretarse a la matrizextracelular.
. Los leucocitos abandonan la luz del vaso mediantediap�edesis: atraviesan la c�elula endotelial o lasuniones entre estas para pasar al espacioextracelular. A menudo, la presencia de receptoresde mol�eculas de adhesion en la superficie luminalde las c�elulas endoteliales, que son reconocidos porlas mol�eculas de adhesion presentes en lasmembranas leucocitarias, favorece la diap�edesis.
Los mediadores histamina y bradicininaincrementan la permeabilidad capilar, lo quefacilita la extravasacion de un volumen excesivode lıquido intraluminal e incrementa el volumendel lıquido extracelular. El aumento notable delvolumen de este lıquido recibe el nombre deedema.El endotelio capilar sintetiza, asimismo:
. Macromol�eculas destinadas al espacioextravascular, como laminina, fibronectina ycol�ageno (tipos II, IV y V).
. Sustancias que intervienen en el mecanismo decoagulacion, la regulacion del tono de lamusculatura lisa de la tunicamedia, y la diap�edesisde los neutrofilos.
. Algunas mol�eculas mediadoras, como elvasodilatador prostaciclina, tambi�en inhiben laagregacion plaquetaria.
. Enzimas que degradan e inactivan la noradrenalina,las prostaglandinas, la serotonina, la trombina y labradicinina.
. Enzimas, como la lipoproteına lipasa, queescinden lipoproteınas y triglic�eridos en glicerol y�acidos grasos, los cuales se almacenan en losadipocitos, y la enzima conversora deangiotensina, que convierte la angiotensina I,un vasoconstrictor d�ebil, en angiotensina II, unvasoconstrictor potente.
158
11
APARATO
CIRCULATORIO

159
11
APARATO
CIRCULATORIO
Figura 11.5 A-C.M�etodos de transporte en el endotelio capilar.(Adaptado de Simionescu N, Simionescu M: In Ussing H,
Bindslev N, Sten-Knudsen O [eds]: Water Transport Across Epithe-
lia. Copenhagen, Munksgaard, 1981.)
Figura 11.4 Control de la irrigacion a trav�es de un lechocapilar. El canal central, formado por lametaarteriola de
la superficie arterial y el canal de desag€ue del lado
venoso, puede derivar el lecho capilar mediante la
clausura de los esfınteres precapilares. (Tomado deGartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.
Philadelphia, Saunders, 2007, p 264.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

VENAS
La sangre de los lechos capilares pasa a las v�enulas,desde las que se conduce hacia venas de calibre cadavez mayor hasta desembocar en las aurıculas del cora-zon. Las venas son unos vasos caracterizados por unabaja presion, por lo que su numero supera al de lasarterias y su di�ametro luminal es mayor, de modo quealbergan alrededor del 70% del volumen sanguıneototal.
. Las venas y las arterias suelen encontrarse proximas,si bien las paredes de las venas aparecen aplanadasdebido a su menor espesor, elasticidad y contenidoen tejido muscular.
. Al igual que las arterias, las venas poseen las trestunicas, si bien la zona de contacto de la tunicamedia y la tunica ıntima est�a poco definida; latunica media presenta un espesor m�as reducido,mientras que la tunica adventicia se caracterizapor su mayor grosor.
. Las venas se distribuyen en tres grupos: v�enulas,venas medianas y pequenas, y venas grandes(tabla 11.2).
Las venas de calibre mediano, en especial, las delas extremidades inferiores, est�an dotadas dev�alvulas que garantizan el flujo unidireccionalde la sangre con el fin de impedir la inversion de lairrigacion. Las v�alvulas venosas:
. Se componen de dos hojuelas procedentes de latunica ıntima que se proyectan hacia la luz.
. Su estructura es poco firme, aunque se refuerza confibras el�asticas y de col�ageno derivadas de la tunicaıntima.
. Se comprimen con la superficie luminal de la pareddel vaso a medida que la sangre fluye hacia elcorazon.
. Se doblan hacia la luz hasta obstruirla, de manerasimilar a dos palmas de las manos en forma de copapara contener agua, e impiden el flujo retrogrado dela sangre.
160
11
APARATO
CIRCULATORIO

CONSIDERACIONES CL�INICAS
Las venas varicosas son venas superficialeshipertrofiadas tortuosas. Su aparicion suelerelacionarse con el proceso de envejecimientoconforme degeneran las paredes de los vasos,disminuye el tono de la musculatura del vaso
afectado o se tornan incompetentes las v�alvulasvenosas. De igual modo, las venas varicosaspueden localizarse en el extremo del esofago
(varices esof�agicas) y el extremo terminal delconducto anal (hemorroides).
161
11
APARATO
CIRCULATORIO
Tabla 11.2 CARACTER�ISTICAS DE LAS VENAS
Tipo Tunica ıntima Tunica media Tunica adventicia
Venas grandes Endotelio, l�amina basal, v�alvulasen algunas, tejido conjuntivo
subendotelial
Tejido conjuntivo, c�elulasde musculo liso
C�elulas de musculo lisodispuestas en haces
longitudinales, c�elulas de
musculo liso cerca de la zonade union al corazon, capas de
col�ageno con fibroblastos
Venas medianas
y pequenas
Endotelio, l�amina basal, v�alvulas
en algunas, tejido conjuntivosubendotelial
Fibras reticulares y el�asticas,
algunas c�elulas de musculo liso
Capas de col�ageno con
fibroblastos
V�enulas Endotelio, l�amina basal (pericitos,
v�enulas poscapilares)
Tejido conjuntivo disperso y
algunas fibras de musculo liso
Cierta cantidad de col�ageno
y algunos fibroblastos
Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 265.
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

CORAZON
El corazon (fig. 11.6), un vaso sanguıneo muymodificado, presenta tres capas: endocardio (corres-pondiente a la tunica ıntima); miocardio (correspon-diente a la tunica media), formado por musculocardıaco, y epicardio (correspondiente a la tunicaadventicia).
. El endocardio reviste la luz del corazon; se continuacon la tunica ıntima de los vasos sanguıneos, por loque consta de un epitelio escamoso simple querecubre una capa de tejido conjuntivo fibroel�asticocon algunos fibroblastos. En una capa m�asprofunda de tejido conjuntivo denso aparecennumerosas fibras el�asticas mezcladas con c�elulas demusculo liso. La capa m�as profunda, la capasubendoc�ardica, que separa el endocardio delmiocardio, se compone de tejido conjuntivo laxoacompanado de vasos sanguıneos, fibras nerviosas yfibras de Purkinje.
. Elmiocardio, la capa intermedia y m�as gruesa de lapared del corazon, contiene c�elulas de musculocardıaco organizadas en espirales que rodean cadauna de las cuatro c�amaras del organo que nos ocupa.Las c�elulas delmusculo cardıaco desempenan variasfunciones:. Union del miocardio al esqueleto fibroso delcorazon.
. Sıntesis y secrecion de hormonas, como elp�eptido natriur�etico auricular, la cardionatrinay la cardiodilatina, que participan en elmantenimiento del equilibrio hidroelectrolıtico yla reduccion de la presion arterial.
. Produccion y conduccion de impulsos.
. La generacion y la conduccion de impulsos corre acargo de:. Un grupo de c�elulas cardıacas especializadas cuyoconjunto conforma el nodulo sinoauricular (SA)de la pared auricular derecha en la zona de unioncon la vena cava superior. Estas c�elulas nodales sedespolarizan de manera espont�anea y generanimpulsos que inician el latido cardıaco a un ritmode unos 70 latidos/min.
. Estos impulsos viajan a trav�es de las c�amarasauriculares y las vıas del noduloauriculoventricular (AV) situado en la paredseptal inmediatamente por encima de la v�alvulatricuspide.
. Las c�elulas cardıacas modificadas localizadas enel nodulo AV reciben los impulsos procedentesdel nodulo SA y transmiten las senales a trav�es delhaz AV (haz de His) hacia el v�ertice de las paredesventriculares y las ramas de los haces AV,denominados fibras de Purkinje, unas c�elulascardıacasmodificadas grandes, para transmitir losimpulsos hacia las c�elulas del musculo cardıaco.
. Estas c�elulas especializadas del musculo cardıacooriginan el latido cardıaco, si bien es el sistemanervioso autonomo el que se ocupa de controlar lafrecuencia cardıaca y el volumen sistolico:. Las fibras simp�aticas incrementan la frecuenciacardıaca.
. Las fibras parasimp�aticas reducen dichafrecuencia.
162
11
APARATO
CIRCULATORIO

CONSIDERACIONES CL�INICAS
La cardiopatıa reum�atica afecta a sujetos
aquejados de fiebre reum�atica durante su infancia.
La fiebre reum�atica da lugar a la creacion de
cicatrices en las v�alvulas debido a la formacion detejido fibrotico durante el proceso de resolucion,lo que reduce su flexibilidad e impide el cierre(incompetencia) o la apertura (estenosis) normales
de estas v�alvulas. La enfermedad afecta m�as a
menudo a la v�alvula AV bicuspide, seguida de las
v�alvulas aorticas.
Las infecciones de la cavidad peric�ardica recibenel nombre de pericarditis; esta cavidad puederellenarse de lıquido y adhesiones entre la capaserosa del pericardio y el endocardio en los casos
m�as graves, en los que puede impedir que el corazonlata de manera normal.
El fenomeno de Raynaud representa una entidadque cursa con decoloracion de los dedos de las
manos o los pies, o ambos, tras la exposicion a
variaciones t�ermicas (calor o frıo) o bien sucesos queinciden en la estabilidad emocional del sujeto.La decoloracion se atribuye a espasmos anomalosen los vasos sanguıneos y la disminucion de lairrigacion. Los dedos afectados adquieren un colorazulado como consecuencia de la deficienciaprolongada de oxıgeno. Los vasos sanguıneosterminan por abrirse, lo que da lugar a un fenomenode «sofoco», que confiere una coloracion rojiza a losdedos. Esta secuencia de modificacion de la
coloracion en tres etapas se asocia, m�as a menudo,a la exposicion al frıo y es tıpica del fenomeno deRaynaud. Esta enfermedad suele afectar a mujeres,en particular a lo largo de la segunda, tercera o
cuarta d�ecadas de vida. Las pacientes puedenpresentar el fenomeno de Raynaud aislado o
acompanando a otras enfermedades reum�aticas.Se desconoce su etiologia.
163
11
APARATO
CIRCULATORIO
Figura 11.6 Diagrama de la estructura del corazon en la
que se representan la localizacion de los nodulos SA yVA, las fibras de Purkinje y el haz de His. (Tomado de
Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.
Philadelphia, Saunders, 2007, p 267.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

. El epicardio, la capa m�as externa del corazon(pericardio visceral), se compone de mesotelio,un epitelio escamoso simple que recubre la capasubepic�ardica de tejido conjuntivo cargado conlıpidos con sus vasos coronarios, nervios y ganglios.El pericardio parietal, formado por una capaserosa interna y una capa fibrosa externa, envuelveal corazon y se prolonga por el pericardio visceral delos grandes vasos que entran y salen de este organo.La cavidad peric�ardica, localizada entre lospericardios visceral y parietal, contiene un lıquidoseroso que reduce la friccion existente entre las dossuperficies peric�ardicas durante el latido cardıaco(fig. 11.7).
El corazon es la bomba que impulsa lacirculacion de la sangre por todo el organismo, paralo cual est�a dotada de cuatro c�amaras: dosaurıculas, que reciben sangre del sistema venoso,y dos ventrıculos, que impulsan la sangredesde el corazon hacia el resto del organismo.Estas cuatro c�amaras configuran dos circuitos:
el circuito pulmonar y el circuito sist�emico(v. fig. 11.7).
. La sangre procedente de los tejidos desemboca en laaurıcula derecha, atraviesa la v�alvula AV derecha(v�alvula tricuspide) y entra en el ventrıculoderecho.
. La sangre abandona el ventrıculo derecho a trav�es dela v�alvula semilunar para entrar en el troncopulmonar, desde el que la sangre desoxigenada sedirige a los pulmones, en los que se oxigenar�a.
. La sangre oxigenada que regresa de los pulmonesentra en la aurıcula izquierda, atraviesa la v�alvulaAV izquierda (v�alvula bicuspide, tambi�enllamada v�alvula mitral) y rellena el ventrıculoizquierdo.
. La sangre sale del ventrıculo izquierdo a trav�es deotra v�alvula semilunar e ingresa en la aorta, que ladistribuye a los tejidos del organismo.
Las v�alvulas impiden el flujo retrogrado de lasangre hacia la c�amara de la que partio.
164
11
APARATO
CIRCULATORIO

CONSIDERACIONES CL�INICAS
La cardiopatıa coronaria afecta a unos14 millones de individuos en EE. UU. Se debe a laacumulacion de calcio y tejido cicatricial en lasarterias coronarias que irrigan el miocardio. Con elpaso del tiempo, los depositos de calcio y placaoriginan ateroesclerosis, la cual reduce la luz de laarteria coronaria afectada y, por tanto, la irrigaciondel musculo cardıaco. Este trastorno ocasiona
dolor tor�acico y angina (dolor referidodescendente en el brazo izquierdo). Elestrechamiento de la arteria hasta obstruirlapuede causar infarto de miocardio o paradacardiorrespiratoria. El tratamiento de eleccion enpacientes con obstruccion arterial parcial sueleser la angioplastia.
165
11
APARATO
CIRCULATORIO
Figura 11.7 Diagrama de la estructura del corazon en la
que se representan la localizacion de los nodulos SA yVA, las fibras de Purkinje y el haz de His. (Tomado de
Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.
Philadelphia, Saunders, 2007, p 267.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Aparato vascular linf�atico
La linfa, el lıquido extracelular que bana los espaciosdel tejido intersticial del organismo, es recogido porcapilares linf�aticos ciegos (fig. 11.8) de los comparti-mentos de tejido conjuntivo, desde los que pasa avasos de calibre creciente hasta regresar a trav�es dedos conductos linf�aticos a las venas de la base delcuello que pertenecen al aparato cardiovascular. Losvasos que forman parte del sistema linf�atico se distri-buyen en todo el organismo, salvo el sistema nerviosocentral, la orbita, el cartılago y el hueso, el oıdointerno y la epidermis. El aparato vascular linf�aticoes un sistema abierto; la linfa no circula ni es impul-sada por bomba alguna. Los ganglios linf�aticos seinterponen a lo largo del recorrido de los vasoslinf�aticos y filtran la linfa.
. Los vasos linf�aticos aferentes conducen la linfahacia los ganglios linf�aticos, que poseen numerososcanales tapizados de endotelio y abundantesmacrofagos que eliminan la materia particulada dela linfa.
. Los linfocitos se incorporan a la linfa al salir delganglio linf�atico en el que ha sido procesada; la linfaregresa al vaso linf�atico a trav�es de los vasoslinf�aticos eferentes.
CAPILARES Y VASOS LINFATICOS
Los capilares linf�aticos ciegos constan de un epitelioescamoso simple muy atenuado, una l�amina basalincompleta; suelen existir espacios intercelulares entrelas c�elulas endoteliales adyacentes debido a la ausen-cia de uniones estrechas. Los filamentos linf�aticos deanclaje (5 a 10nmde di�ametro), que se insertan en lasmembranas plasm�aticas abluminales, mantienen lapermeabilidad de estos delicados vasos.
La linfa de los capilares linf�aticos drena en vasoslinf�aticos de calibre mediano y pequeno de estructurasimilar a la de las venas pequenas, si bien presentanuna luz mayor y paredes m�as delgadas. Los vasoslinf�aticos de mayor calibre poseen una fina capa defibras el�asticas y musculo liso recubierta por unal�amina de fibras el�asticas que se mezclan con el tejidoconjuntivo circundante. Los dos vasos linf�aticos m�asgrandes, el conducto linf�atico derecho y el conductotor�acico, desembocan en el sistema venoso del cuelloy presentan una composicion semejante a la de lasgrandes venas, con tres tunicas definidas y vasosnutrientes similares a los vasa vasorum de las arteriasy venas.
166
11
APARATO
CIRCULATORIO

CONSIDERACIONES CL�INICAS
El t�ermino linfedema se aplica a una acumulacionanomala de lıquido intersticial que producetumefaccion, por lo general en las extremidadessuperiores o inferiores. Se debe a la desaparicion,
alteracion, lesion o reseccion de vasos linf�aticos o
ganglios linf�aticos. El linfedema primario constituyeuna entidad infrecuente ocasionada por la ausencia
cong�enita de ciertos vasos linf�aticos o poranomalıas de los mismos. El linfedema secundarioaparece como consecuencia de la obstruccion o la
interrupcion de alguna vıa linf�atica. Puede deberse,
tambi�en, a una infeccion, neoplasia maligna,intervencion quirurgica, formacion de tejidocicatricial, traumatismo, trombosis venosa profunda,exposicion a radiacion o algun otro tratamiento
antineopl�asico.
Las c�elulas de tumores malignos acceden al
sistema linf�atico desde el tumor primario. A lo largo
de su recorrido en las vıas linf�aticas, estas c�elulas
pueden entrar en un ganglio linf�atico que limitarıa su
diseminacion. Sin embargo, las c�elulas tumoralespueden proliferar en el seno del ganglio, del que
saldrıan para formar una met�astasis en unalocalizacion secundaria. En la cirugıa de reseccion
de un ganglio invadido por c�elulas tumorales, se
extirpan tambi�en otros ganglios linf�aticos
hipertrofiados pertenecientes a lamisma vıa linf�atica,
ası como los vasos linf�aticos asociados, con lafinalidad de evitar la diseminacion secundaria de las
c�elulas tumorales y, por ende, la formacion de
met�astasis.
167
11
APARATO
CIRCULATORIO
Figura 11.8 Diagrama de la ultraestructura de un capilar linf�atico. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.
Philadelphia, Saunders, 2007, p 270.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

12 SISTEMA LINFATICO(INMUNITARIO)
El sistema linf�atico protege al organismo frente a lainvasion por agentes exogenos, como macro-mol�eculas y microorganismos, ası como c�elulas infec-tadas por virus. Este sistema se com-pone de diversas poblaciones dec�elulas no encapsuladas, que recibenel nombre de sistema linf�atico difuso,junto a la linfa, los ganglios, las amıg-dalas, el timo y el bazo.
Aspectos generales delsistema linf�atico
El organismo posee tres niveles defensi-vos: el epitelio, que lo aısla del medio-ambiente; la epidermis, y las mucosas.Estas tres lıneas de defensa representanbarreras fısicas que suelen impedir el acceso de lospatogenos a los compartimentos est�eriles del orga-nismo. Estas barreras, relativamente delgadas, pue-den verse alteradas como consecuencia de un trau-matismo y algunos microorganismos patogenos soncapaces de atravesarlas incluso cuando se encuentranintactas. Los sistemas inmunitarios innato (inespecı-fico) y adaptativo (adquirido) representan otras doslıneas defensivas. En la mayorıa de los casos, estossistemas logran proteger al organismo ante el fracasode las anteriores.
SISTEMA INMUNITARIO INNATO
El sistema inmunitario innato (natural), el m�as pri-mitivo y antiguo desde el punto de vista evolutivo, perotambi�en el m�as r�apido, est�a formado por el comple-mento, diversos p�eptidos antimicrobianos, citocinas,macrofagos, neutrofilos, linfocitos citotoxicos natura-les (NK) y receptores de tipo Toll. Se trata de un sistemainespecıfico carente de memoria inmunitaria frente almicroorganismo o la mol�ecula que desencadeno suaccion. En la tabla 12.1 se enumeran los acronimosque se utilizar�an a lo largo de este capıtulo.
. El complemento, un conjunto de mol�eculascirculantes, precipita a trav�es de una secuenciaespecıfica y da lugar a un complejo de ataque demembrana que actua sobre las membranas de lospatogenos que han logrado acceder al torrente
circulatorio. Los neutrofilos y losmacrofagos portanreceptores de C3b, cuya interaccion con elcomponente C3b unido a la superficie de dichos
microorganismos induce su fagocitosispor estas c�elulas.. Los p�eptidos antimicrobianos,como lisozima y defensina,destruyen los microorganismos altiempo que reclutan linfocitos T yc�elulas dendrıticas.
. Se distinguen varios grupos demol�eculas de senalizacion, conocidascomo citocinas, segun su origen yfunciones:. Las mol�eculas sintetizadas por loslinfocitos son las interleucinas.
. Las quimiocinas ejercen unaaccion quimioatrayente.
. Las mol�eculas inductoras de la proliferacion y ladiferenciacion son los factores estimuladores decolonias (CSF).
. Las citocinas frente a virus se llaman interferones.
. Los macrofagos son fagocitos que reconocen lasporciones Fc de los anticuerpos, el componenteC3b del complemento, y residuos glucıdicospresentes en la membrana del microorganismoinvasor. Estas c�elulas interaccionan con loslinfocitos T y B presentadoras de antıgenos.De igual modo, inducen la proliferacionde las CFU-GM y CFU-G.
. Los linfocitos NK intervienen en la citotoxicidadcelular dependiente de anticuerpos, por lo que sonsimilares a los linfocitos T citotoxicos (LTC). Adiferencia de �estos, no obstante, los linfocitos NKno han de madurar en el timo para diferenciarse enc�elulas citotoxicas. Estos linfocitos poseenreceptores activadores de linfocitos citotoxicosy receptores inhibidores de linfocitoscitotoxicos. Los primeros destruyen las c�elulasunidas a la porcion variable de los anticuerpos IgGal reconocer su porcion Fc, a no ser que presentenmol�eculas tipo I del complejo principalde histocompatibilidad en su superficie celular.
. Los receptores de tipo Toll, unas proteınasintegrales presentes en la membrana plasm�atica delas c�elulas pertenecientes al sistema inmunitario
� 2011. Elsevier Espana, S.L. Reservados todos los derechos
TERMINOS CLAVE. Sistema inmunitario
innato. Sistema inmunitario
adaptativo. Inmunoglobulinas. Linfocitos T. Linfocitos B. Mol�eculas MHC
168

innato, se disponen en parejas para funcionar.Algunos de ellos son proteınas transmembrana,mientras que otros se asocian exclusivamente a lasuperficie citopl�asmica de la membrana plasm�atica.Casi todos los receptores de tipo Toll inducen la vıadel factor nuclear kB para poner en marcha unacascada intracelular que culminarıa con laliberacion de citocinas especıficas. De igual modo,estos receptores pueden activar una respuestainflamatoria y suscitar una respuesta celular en laque participan los linfocitos T y B del sistemainmunitario adquirido. En la tabla 12.2 se ofreceuna relacion de las posibles funciones de losreceptores de tipo Toll.
SISTEMA INMUNITARIO ADAPTATIVO
El sistema inmunitario adaptativo (adquirido) esespecıfico y est�a formado por los linfocitos T y B ylas c�elulas presentadoras de antıgenos (CPA), aunque
utiliza tambi�en algunos componentes del sistemainmunitario innato para proteger al organismo.Adem�as de secretar citocinas para comunicarse entresı, estas c�elulas interaccionan unas con otras e inducenrespuestas especıficas en otras poblaciones celulares atrav�es del reconocimiento de ciertas mol�eculas unidasa la membrana plasm�atica con el fin de actuar sobreunas sustancias exogenas conocidas como antıgenos.Por definicion:
. Todos los antıgenos interaccionan con losanticuerpos de manera independiente de sucapacidad de induccion de una respuestainmunitaria.
. Un inmunogeno es una mol�ecula exogena concapacidad de poner en marcha una respuestainmunitaria.
Las c�elulas que pertenecen al sistema inmunitarioadaptativo liberan citocinas, las cuales reclutandiversas c�elulas del sistema inmunitario innato que
169
12
SISTEMALIN
FATIC
O(INMUNITA
RIO)
Tabla 12.1 ACRONIMOS Y ABREVIATURAS
Acronimo/Abreviatura Definicion
BALT Tejido linf�atico asociado a los bronquiosC3b Complemento 3b
CAM Complejo de ataque a membrana
CCDA Citotoxicidad celular dependiente de anticuerpos
CD Mol�ecula del grupo de diferenciacion (seguido de un numero ar�abico)C�elula M C�elula de micropliegue
CLIP Proteına invariable asociada a clase II
CPA C�elula presentadora de antıgenos
CSF Factor estimulador de coloniasCSF-G Factor estimulador de colonias de granulocitos
CSF-GM Factor estimulador de colonias de granulocitos-macrofagos
Fab Fragmento de union a antıgenos de un anticuerpoFc Fragmento cristalizado (fragmento constante de un anticuerpo)
GALT Tejido linf�atico asociado al intestino
IFN-g Interferon gIgs Inmunoglobulinas de superficieIL Interleucina (seguida de un numero ar�abico)
Linfocito B Linfocito derivado de la bolsa (linfocito derivado de la m�edula osea)
Linfocito NK Linfocito citotoxico natural
Linfocito T Linfocito derivado del timoLinfocito T reg Linfocito T regulador
Linfocito TH Linfocito T cooperador (seguido de un numero ar�abico)
LTC Linfocito T citotoxicoLTMC Linfocito T central de memoria
LTME Linfocito T efector de memoria
MALT Tejido linf�atico asociado a mucosas
MHC I y II Mol�eculas del complejo principal de histocompatibilidad I y IIPTra Proteına transportadora (1 y 2)
TCR Receptor de linfocitos T
TLR Receptor de tipo Toll
TNF-a Factor de necrosis tumoral-aTSH Hormona estimuladora del tiroides
VEA V�enula del endotelio alto
Vesıcula MIIC Compartimento con abundantes mol�eculas de MHC II
VLPA Vaina linf�atica periarterial
Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 274.
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

colaborar�an en la respuesta frente a los antıgenosexogenos. El sistema inmunitario adaptativo se carac-teriza por cuatro rasgos principales: la especifici-dad, la diversidad, la memoria y la capacidad dediferenciar las mol�eculas propias de las exogenas.Las reacciones inmunitarias desplegadas por el sis-tema inmunitario adaptativo pueden ser de dostipos:
. La respuesta inmunitaria humoral se sustenta enlas inmunoglobulinas (anticuerpos) sintetizadospor los linfocitos B diferenciados, llamados c�elulasplasm�aticas. Los anticuerpos se unen a losantıgenos para inactivarlos o senalarlos para suposterior destruccion por parte de los macrofagos.
. En la respuesta inmunitaria celular, los LTC, unasubpoblacion especıfica de linfocitos T, se activapara interaccionar con una c�elula exogena o alteradapor un virus y provocar su apoptosis.
Las c�elulas del sistema inmunitario adaptativo seforman en lam�edula osea, en la que los linfocitos B sediferencian en c�elulas inmunocompetentes. La adqui-sicion de la inmunocompetencia por parte de loslinfocitos T requiere su salida de la m�edula oseay su migracion a la corteza tımica. Los linfocitosB y T inmunocompetentes abandonan los organoslinf�aticos primarios (m�edula osea y timo) y se dirigenal tejido linf�atico difuso, los ganglios linf�aticos y elbazo (cuyo conjunto recibe el nombre de organoslinf�aticos secundarios), en los que tratan de detectarantıgenos e interaccionan con ellos.
SELECCION CLONAL Y EXPANSION
A lo largo del desarrollo embrionario se desarrollan,aproximadamente, 1015 linfocitos que reconocer�any eliminaran todos los antıgenos y patogenos con los
que el individuo pueda entrar en contacto durantesu vida. Cada uno de estos linfocitos es capaz dereconocer un antıgeno exogeno determinado; al pro-liferar, cada linfocito origina una subpoblacion dec�elulas id�enticas que recibe el nombre de clon. Las c�e-lulas que pertenecen a un mismo clon presentan losmismos anticuerpos de membrana (inmunoglobu-linas de superficie [Igs]) o el mismo receptor delinfocitos T (TCR). Durante la embriogenia, losclones que portan inmunoglobulinas o los recep-tores que reconocen macromol�eculas propias soneliminados (delecion clonal) o inactivados paraimpedir la induccion de una respuesta inmunitaria(anergia clonal), lo que evita la aparicion de reac-ciones autoinmunitarias.
. El primer contacto con un antıgeno determinadosuscita una respuesta lenta y d�ebil por parte delsistema inmunitario adaptativo, la respuestainmunitaria primaria, dado que los linfocitos T y Bno han estado expuestos al antıgeno se consideranc�elulas vırgenes (naıve).
. Con posterioridad a este contacto, las c�elulasvırgenes proliferan y forman una subpoblacion dec�elulas efectoras (c�elulas plasm�aticas en el caso dela respuesta humoral y LTC, linfocitos Tcooperadores [TH] TH1, TH2, TH17 y linfocitos Treguladores CD [linfocitos T reg] en la respuestainmunitaria), c�elulas que reaccionan y eliminan alantıgeno, y c�elulas de memoria que se asemejan alas c�elulas vırgenes. Las c�elulas efectoras vivenmucho tiempo (anos), responden con mayorrapidez e intensidad ante una nueva exposicion almismo antıgeno (respuesta inmunitariasecundaria, respuesta anamn�esica) y puedenampliar de manera considerable el tamano de supoblacion (expansion clonal).
170
12
SISTEMALIN
FTICO(INMUNITA
RIO)
Tabla 12.2 RECEPTORES DE TIPO TOLL Y POSIBLES FUNCIONES
Dominios Pareja de receptores Funcion
Intracelular y extracelular
(en membrana celular)
TLR1-TLR2 Union a lipoproteınas bacterianas; union a ciertas proteınas
de par�asitosTLR2-TLR6 Union a �acido lipoteicoico de pared de bacterias grampositivas
y a cimosan
TLR4-TLR4 Union a LPS de bacterias gramnegativas
TLR5-¿?* Union a flagelina de flagelos bacterianosTLR11-¿?* Reconocimiento de Toxoplasma gondii
Solamente intracelular TLR3-¿?* Union a ARN vırico bicatenario
TLR7-¿?* Union a ARN vırico monocatenario
TLR8-¿?* Union a ARN vırico monocatenarioTLR9-¿?* Union a ADN bacteriano y vırico
TLR10-¿?* Desconocida
TLR12-¿?* Desconocida
*Se desconoce, aun, cu�al es el otro miembro de la pareja.LPS, lipopolisac�arido.Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 275.

Inmunoglobulinas (anticuerpos)
Las c�elulas plasm�aticas sintetizan gran cantidad deuna familia especial de glucoproteınas, los anticuer-pos (inmunoglobulinas), que tambi�en son fabrica-dos en menores cantidades por los linfocitos B (quelos expresan en sus membranas plasm�aticas en formade Igs, receptores de linfocitos B). Un anticuerpotıpico (IgG), que se asemeja a la letra Y, consta decuatro cadenas polipeptıdicas (fig. 12.1).
. Dos grandes cadenas pesadas id�enticas, unidasentre sı por puentes disulfuro, que conformanel tronco y los brazos de la Y (en la que el brazo y eltronco se unen pormedio de una region de bisagra).
. Dos cadenas ligeras cortas id�enticas queconstituyen los brazos de la estructura en Y y semantienen unidas a la cadena pesadacorrespondiente mediante enlaces disulfuro.
La escision enzim�atica de un antibiotico por partede la papaına se produce en la region de bisagra y dalugar a un fragmento Fc, el tronco, cuya secuenciaaminoacıdica es constante, y dos fragmentos Fab(de union a antıgenos), cada uno de los cuales est�aformado por una cadena ligera y una porcion de unacadena pesada cuyos extremos distales se distinguenpor la capacidad de unirse exclusivamente a un epıtopodeterminado. Se han definido cinco clases diferentesde inmunoglobulinas en funcion de sus caracterısticasdistintivas (tabla 12.3).
CELULAS DE LOS SISTEMAS INMUNITARIOS INNATO
Y ADAPTATIVO
Los sistemas inmunitarios innato y adaptativo se com-ponen de las siguientes c�elulas: linfocitos B, linfocitos T,macrofagos y las CPA, derivadas de ellos, y linfoci-tos NK.
Linfocitos B
Los linfocitos B se desarrollan en lam�edula osea, en laque adquieren su inmunocompetencia. Sintetizananticuerpos IgM e IgD, cuyos extremos Fc se insertanen la membrana (Igs) de tal modo que la porcion Fcqueda expuesta al entorno extracelular. La porcionFc se ancla a la membrana plasm�atica a trav�es de dosproteınas transmembrana, Igb e Iga, que transduceninformacion al compartimento intracelular comoconsecuencia de la interaccion de la Igs con un epı-topo y desencadenan una secuencia de senalizacionque dar�a lugar a:
. Activacion de los linfocitos B, que se ocupan de larespuesta inmunitaria humoral.
. Proliferacion de los linfocitos B activados paraoriginar c�elulas plasm�aticas y linfocitos B dememoria.. Las c�elulas de memoria se encargan de laexpansion clonal.
. Las c�elulas plasm�aticas fabrican inicialmenteIgM y, m�as tarde, sintetizan otros isotipos(tabla 12.4).
Algunos polisac�aridos, como los peptidoglucanospresentes en las membranas bacterianas, son antıge-nos independientes del timo, ya que pueden suscitaruna respuesta inmunitaria humoral en ausencia delinfocitos T intermediarios. Unicamente se sintetizananticuerpos IgM y no se forman linfocitos B de me-moria.
Linfocitos T
A pesar de que el desarrollo de los linfocitos T tienelugar en la m�edula osea, estas c�elulas han de migrar ala corteza tımica para expresar ciertas mol�eculas ensu membrana plasm�atica y convertirse en c�elulas
171
12
SISTEMALIN
FATIC
O(INMUNITA
RIO)
Figura 12.1 Esquema de una inmunoglobulina tıpica. (Tomado deGartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia,
Saunders, 2007, p 278.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

172
12
SISTEMALIN
FTICO(INMUNITA
RIO)
Tabla 12.3 ISOTIPOS DE INMUNOGLOBULINAS
Clase y n.� deunidades* Citocinas† Union a c�elulas Rasgos biologicos
IgA 1 o 2 TGF-b Temporal a c�elulas
epiteliales durante
el proceso de
secrecion
Presente en l�agrimas, saliva, luz intestinal y cavidad nasal
en forma de dımeros; las unidades que integran el
dımero se unen mediante la proteına J fabricada por las
c�elulas plasm�aticas y se protegen de la degradacionenzim�atica con un componente secretor sintetizado por
las c�elulas epiteliales; destruye antıgenos y
microorganismos en la luz del tubo digestivo, la cavidad
nasal, la vagina y el saco conjuntival; se secreta en la lechey confiere proteccion al neonatomediante la inmunidad
pasiva; aparece en forma de monomeros en la sangre;
colabora con los eosinofilos en el reconocimiento y ladestruccion de par�asitos
IgD 1 Membrana
plasm�atica de
linfocitos B
Inmunoglobulina de superficie; colabora con los
linfocitos B en el reconocimiento de antıgenos
especıficos; interviene en la activacion de linfocitos B trasla exposicion antig�enica, lo que da lugar a su
diferenciacion en c�elulas plasm�aticas
IgE 1 IL-4, IL-5 Mastocitos y
basofilos
Anticuerpo reagınico; la IgE favorece la desgranulacion de
basofilos y mastocitos como consecuencia delentrecruzamiento de varios anticuerpos de membrana
con antıgenos, lo que provoca la liberacion de diversos
mediadores, como heparina, histamina, factoresquimiot�acticos de neutrofilos y eosinofilos y
leucotrienos; suscita reacciones de hipersensibilidad
inmediata; colabora con los eosinofilos en el
reconocimiento y la destruccion de par�asitosIgG 1 IFN-g , IL-4, IL-6 Macrofagos y
neutrofilos
Atraviesa la placenta y confiere proteccion pasiva al feto;
se secreta en la leche y confiere inmunidad pasiva al
neonato; fija la cascada del complemento; actua como
opsonina, es decir, recubre a los microorganismos parapotenciar su fagocitosis por los macrofagos y los
neutrofilos, c�elulas dotadas de receptores Fc para la
region Fc de estos anticuerpos; interviene en la
citotoxicidad celular dependiente de anticuerpos atrav�es de la activacion de linfocitos NK; se sintetiza en
grandes cantidades en la respuesta inmunitaria
secundariaIgM 1 o 5 Linfocitos B
(en forma
monom�erica)
La estructura pentam�erica se mantiene por medio de
enlaces con las proteınas J que unen las regiones Fc
de cada unidad; activa la cascada del sistema del
complemento; constituye el primer isotipo que sesintetiza en la respuesta inmunitaria primaria
*Una unidad es una unica inmunoglobulina formada por dos cadenas pesadas y dos cadenas ligeras; la IgA aparece en forma demonomero y de dımero.yCitocinas implicadas en el cambio de isotipo.Fc, fragmento cristalizable; IFN, interferon; IL, interleucina; NK, citotoxico natural; TGF, factor transformador del crecimiento.
Tabla 12.4 CAMBIO DE ISOTIPO DE IgM
Cambio a
Citocinasintetizada porlinfocito TH Microorganismo Funcion
IgE IL-4, IL-5 Helmintos par�asitos Union a mastocitos
IgG IL-6, IFN-g Bacterias y virus Opsonizacion de bacterias, fijacion del complemento,
induccion de destruccion de c�elulas transformadaspor virus por linfocitos NK (CCDA)
IgA TGF-b Bacterias y virus Secrecion en superficie de mucosas
CCDA, citotoxicidad celular dependiente de anticuerpos; IFN, interferon; IL, interleucina; NK, citotoxico natural; TGF, factor
transformador del crecimiento.Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 281.

inmunocompetentes (v. apartado «Timo»). A diferen-cia de los linfocitos B, los linfocitos T:
. Presentan TCR en lugar de Igs.. Los TCR son similares a los anticuerpos, ya que suregion constante se entierra en la membranacelular y su region variable se extiende hacia elespacio extracelular y se une a los epıtopos.
. No son capaces de reconocer antıgenos que no seanpresentados por las CPA.
. Expresan grupos de proteınas de diferenciacion(mol�eculas CD) en su membrana plasm�atica(tabla 12.5).. Se han identificado unas 200 mol�eculas CDdistintas. El complejo TCR, formado por TCR,CD3 y CD4 o CD8, reconoce a los epıtopospresentados por las CPA y se une a ellas.
. Tan solo actuan en el espacio inmediato que lasrodea.
. Ignoran los antıgenos de naturaleza no proteica.
. Unicamente reconocen epıtopos asociados a una delas dos clases de mol�eculas MHC de las CPA. Estasmol�eculas est�an determinadas gen�eticamente y sondistintas en cada individuo; caracterizan «lo que espropio».. Las MHC I se localizan en las membranasplasm�aticas de las c�elulas nucleadas.
. Las MHC II (y MHC I) se encuentran en lasmembranas celulares de las CPA.
La activacion de los linfocitos T no depende sola-mente del reconocimiento del epıtopo, sino tambi�en
de lamol�eculaMHC. El sistema inmunitario no puededesplegar una respuesta inmunitaria cuando el linfo-cito T no reconoce dicha mol�ecula; por consiguiente,se dice que estos linfocitos est�an restringidospor MHC. Los linfocitos T se dividen en tres clasesamplias:
. Linfocitos T vırgenes
. Linfocitos T de memoria
. Linfocitos T efectores
Los linfocitos T vırgenes son c�elulas inmunocom-petentes que portan mol�eculas CD45RA en su plas-malema, aunque aun no se han visto expuestas a nin-gun antıgeno. La exposicion antig�enica de estas c�elulasinduce su proliferacion para originar linfocitos T dememoria y efectores.
Los linfocitos T de memoria poseen mol�eculasCD45R0 en su membrana plasm�atica y se distribuyenen dos grupos: linfocitos T de memoria centrales(LTMC), que presentan mol�eculas de CR7+ en sumembrana y linfocitos T de memoria efectores (lin-focitos CR7-, LTME), carentes de esta mol�ecula en susuperficie celular. La memoria inmunitaria del orga-nismo se sustenta en estas c�elulas. Los LTMC se hallanen la paracorteza de los ganglios linf�aticos, en la que seunen a las CPA para inducir la secrecion de IL-12 porestas. Este mediador citocınico induce la proliferacionde los LTMC y la formacion de LTME. Los LTME asıformados migran hacia el foco de inflamacion, sediferencian en linfocitos T efectores y responden ala exposicion a un antıgeno.
173
12
SISTEMALIN
FATIC
O(INMUNITA
RIO)
Tabla 12.5 CIERTOS MARCADORES DE SUPERFICIE IMPLICADOS EN REACCIONES INMUNITARIAS
Proteına Superficie celularLigando yc�elula diana Funcion
CD3 Todos los linfocitos
T
Ninguno Transduccion de union de complejo MHC-epıtopo en senal
intracelular; activacion de linfocito T
CD4 Linfocitos T
cooperadores
MHC II en CPA Correceptor de union de TCR a complejo MHC II-epıtopo,
activacion de linfocitos T cooperadoresCD8 Linfocitos T
citotoxicos y
linfocitos T
supresores
MHC I en la
mayorıa de las
c�elulas nucleadas
Correceptor de union de TCR a complejo MHC I-epıtopo,
activacion de linfocitos T citotoxicos
CD28 Linfocitos T
cooperadores
7 en CPA Colabora en activacion de linfocitos T cooperadores
CD40 Linfocitos B Receptor de CD40presente en
linfocitos T
cooperadores
activados
Union de CD40 a receptor de CD40 induce la activacion delinfocitos B por parte de linfocitos T cooperadores, lo que
provoca su diferenciacion en linfocitos B de memoria y
c�elulas plasm�aticas
CD, molecula de grupo de diferenciacion; CPA, celula presentadora de antıgenos; MHC, complejo principal de histocompatibilidad;
TCR, receptor de linfocitos T.Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 281.
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

CONSIDERACIONES CL�INICAS
El anticuerpo que sintetizan inicialmente loslinfocitos B es IgM hasta que los linfocitos THinducen un cambio de isotipo a IgG. En los sujetoscon deficiencia de ligandos de CD40 no se realizael cambio de isotipos, por lo que presentan unasconcentraciones excesivamente altas de IgM, untrastorno llamado sındrome de hiper-IgM queorigina infecciones cronicas debidas a estainmunodeficiencia humoral.
Todas las c�elulas nucleadas poseen mol�eculasMHC I, que han de ser reconocidas por los LTCpara inducir una respuesta inmunitaria. Muchas
c�elulas tumorales y c�elulas infectadas por virus
interrumpen la sıntesis de mol�eculas MHC I con elfin de evitar su reconocimiento y ulteriordestruccion por esta poblacion de linfocitos.
Linfocitos T efectores
Los linfocitos T efectores de memoria originan linfo-citos T efectores, que engloban tres grupos diferentesde c�elulas inmunocompetentes dotadas de la capaci-
dad de desplegar una respuesta inmunitaria, a saber,linfocitos TH, LTC y linfocitos T citotoxicos, y linfo-citos T reg.
Todos los linfocitos TH portanmol�eculas CD4 en elplasmalema y poseen la capacidad de colaborar tantocon c�elulas pertenecientes al sistema inmunitario innatocomo al adaptativo. Asimismo, los linfocitos TH indu-cen la destruccion de c�elulas exogenas e infectadas porvirus por parte de los LTC y la diferenciacion de loslinfocitos B en c�elulas plasm�aticas que fabrican anti-cuerpos. Se distinguen cuatro clases de linfocitos TH(unaquintaclase sehaasignadoalgrupode los linfocitosT reg), todas las cuales secretan citocinas (tabla 12.6):
. Linfocitos TH0, precursores de las otras tres clases delinfocitos TH; sintetizan un gran numero decitocinas diferentes.
. Linfocitos TH1:. Respuestas directas frente a patogenosintracitopl�asmicos.
. Comienzo de respuestas inmunitarias celulares.
. Secrecion de IL-2, un mediador que induce lamitosis en los linfocitos T CD4 y CD8 y lacitotoxicidad en los LTC.
174
12
SISTEMALIN
FTICO(INMUNITA
RIO) Tabla 12.6 ORIGEN Y FUNCIONES DESTACADAS DE CIERTAS CITOCINAS
Citocina Origen celular C�elula diana Funcion
IL-1a e IL-1b Macrofagos y c�elulas epiteliales Linfocitos T y macrofagos Activacion de linfocitos T y macrofagosIL-2 Linfocitos TH1 Linfocitos T y B activados Favorece la proliferacion de linfocitos T
y B activados
IL-4 Linfocitos TH2 Linfocitos B Favorece la proliferacion de linfocitos B
y su diferenciacion en c�elulasplasm�aticas; favorece el paso de
produccion de IgM a IgG e IgE
IL-5 Linfocitos TH2 Linfocitos B Favorece la proliferacion de linfocitos B
y su diferenciacion en c�elulasplasm�aticas; favorece el paso de
produccion de IgM a IgE
IL-6 C�elulas presentadoras deantıgenos y linfocitos TH2
Linfocitos T; linfocitos Bactivados
Activacion de linfocitos T; favorece lamaduracion de linfocitos B en c�elulas
plasm�aticas que sintetizan IgG
IL-10 Linfocitos TH2 Linfocitos TH1 Inhibe el desarrollo de linfocitos TH1;
inhibe la secrecion de citocinas porestas c�elulas
IL-12 Linfocitos B y macrofagos Linfocitos NK y linfocitos T Activacion de linfocitos NK e induccion
de formacion de c�elulas similares a
linfocitos TH1TNF-a Macrofagos
Linfocitos TH1
Macrofagos
Macrofagos hiperactivos
Autoactivacion de macrofagos para
secretar IL-12
Estimula a los macrofagos hiperactivospara producir radicales de oxıgeno,
favoreciendo la destruccion bacteriana
IFN-a C�elulas infectadas por un virus Linfocitos NK y macrofagos Activacion de macrofagos y linfocitos NK
IFN-b C�elulas infectadas por un virus Linfocitos NK y macrofagos Activacion de macrofagos y linfocitos NKIFN-g Linfocitos TH1 Macrofagos y linfocitos T Favorece la destruccion celular por
linfocitos T citotoxicos y fagocitosis
por macrofagos
IFN, interferon; IL, interleucina; NK, citotoxico natural; TH, T cooperador; TNF, factor de necrosis tumoral.Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 284.

. Liberacion de IFN-g , el cual induce la destruccionde microorganismos fagocitados por losmacrofagos y activa la subpoblacion de linfocitosNK. Los macrofagos secretan IL-12, una citocinaque favorece la expansion de los linfocitos TH1 yrestringe la ampliacion de la poblacion delinfocitos TH2.
. Sıntesis del factor de necrosis tumoral-b, el cualfavorece la inflamacion aguda por accion de losneutrofilos.
. Los linfocitos TH2 intervienen en la puesta enmarcha de respuestas humorales frente a par�asitos einfecciones en la mucosa; estas c�elulas sintetizan:. IL-4, que activa la produccion de IgE por parte delos linfocitos B en respuestas al�ergicas y, junto aIL-10, impide el desarrollo de linfocitos TH1.
. IL-5, la cual favorece el desarrollo de loseosinofilos.
. IL-6, que activa la formacion de linfocitos T y B enel asma y el lupus sist�emico eritematoso.
. IL-9, que amplifica las respuestas de losmastocitos y la proliferacion de los linfocitos TH2.
. IL-13, que induce la formacion de linfocitos B ydemora el desarrollo de linfocitos TH1.
. Los linfocitos TH17 sintetizan IL-17 y amplifican larespuesta mediada por los neutrofilos al potenciarsu reclutamiento; asimismo, pueden diferenciarse apartir de los linfocitos T vırgenes en presencia deIL-6 y el factor transformador del crecimiento b.
. A diferencia de los linfocitos TH, el plasmalema delos LTC posee mol�eculas CD8. Los TCR de estoslinfocitos se unen a epıtopos presentes en lamembrana plasm�atica de c�elulas exogenas,infectadas por un virus o tumorales; adem�as:. Insertan perforinas en el plasmalema de la c�eluladiana para formar poros en la misma.
. Secretan granzimas que pasan al citoplasma de lac�elula diana a trav�es de los poros ası formados einducen la apoptosis en la misma.
. Su el plasmalema contiene CD95L (ligandode apoptosis), que se une a CD95 (receptor deapoptosis) en la membrana de la c�elula dianapara activarlo, lo que induce una cascadaapoptosica en la misma.
. Los linfocitos T reg tambi�en poseenmol�eculas CD4en el plasmalema e intervienen en la inhibicion dela respuesta inmunitaria. Se conocen dos categorıasde linfocitos T reg que pueden actuar de maneracoordinada para bloquear la respuesta inmunitaria:. Linfocitos T reg naturales, los cuales detienen larespuesta inmunitaria de manera independientede antıgenos a trav�es de la union a CPA.
. Linfocitos T reg inducibles (anteriormenteconocidos como linfocitos TH3), que secretanIL-10 y FTG-b con el fin de impedir la formacionde linfocitos TH1.
. Adiferencia de otros linfocitos T, los linfocitos TNKresponden ante antıgenos lipıdicos presentados porCPA portadoras de mol�eculas CD1 en su superficie.Los linfocitos T NK son similares a las c�elulas NK, yaque pueden ser activados directamente, aunquepara ello han de convertirse en c�elulasinmunocompetentes en la corteza tımica. Estasc�elulas secretan IL-4, IL-10 e IFN-g .
CONSIDERACIONES CL�INICAS
En algunos sujetos, el sistema inmunitariopresenta una alteracion conocida comoenfermedaddeGraves, en la cual no se reconoce
como propios a los receptores de las c�elulas delos folıculos tiroideos para la hormonaestimuladora del tiroides. Por el contrario, estosreceptores se interpretan como antıgenos. Los
trastornos en los que las mol�eculas propias se
consideran erroneamente mol�eculas exogenas seconocen como enfermedadesautoinmunitarias. Los anticuerpos se unen a losreceptores de la TSH, lo que ocasiona una sıntesisexcesiva de esta hormona. Los pacientesaquejados de la enfermedad de Graves presentanhipertrofia tiroidea y exoftalmia (globos ocularessobresalientes).
Mol�eculas del complejo principalde histocompatibilidad
Las mol�eculas MHC, localizadas en la superficie de lasCPA, como las c�elulas infectadas o alteradas por unvirus, retienen unos pequenos p�eptidos derivados delos antıgenos, llamados epıtopos, para su presenta-cion a los linfocitos T. Las mol�eculas MHC de cadapersona son diferentes de las de otros sujetos; loslinfocitos T pueden reconocer las mol�eculas propias.Se distinguen dos grupos de mol�eculas MHC:
. Las MHC I se encargan de la presentacionde epıtopos (cuya secuencia contiene de 8 a12 amino�acidos) escindidos de proteınassintetizadas por la c�elula (proteınas endogenas);todas las c�elulas nucleadas, como las CPA, fabricanestas mol�eculas MHC I.
. Las MHC II se ocupan de presentar epıtopos(integrados por 13 a 25 amino�acidos) procedentesde proteınas fagocitadas (proteınas exogenas); susıntesis corresponde unicamente a las CPA.
Carga de epıtopos en las mol�eculas del complejo principalde histocompatibilidad de clase I
La degradacion de las proteınas endogenas por losproteasomas genera epıtopos de 8 a 12 amino�acidos
175
12
SISTEMALIN
FATIC
O(INMUNITA
RIO)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

de longitud. Las proteınas transportadoras TAP1 yTAP2 transportan a los epıtopos al retıculo endopl�as-mico rugoso, donde se unen a MHC I; el complejo asıformado se transfiere hacia el aparato de Golgi, en elcual se empaquetar�a para ser transportado. El com-plejo MHC I-epıtopo se traslada a la membrana plas-m�atica para su presentacion a los LTC, los cuales deci-dir�an si deben ser destruidas. Se induce la apoptosisde las c�elulas portadoras de antıgenos virales, mien-tras que las que expresan antıgenos propios no sonatacadas.
Carga de epıtopos en las mol�eculas del complejo principalde histocompatibilidad de clase II
. Las proteınas exogenas fagocitadas por losmacrofagos y las CPA se escinden en fragmentoscada vez m�as pequenos en los endosomastempranos y tardıos (secuencia de 13 a25 amino�acidos).
. Almismo tiempo, estas c�elulas sintetizanmol�eculasMHC II en el retıculo endopl�asmico rugoso, en cuyaluz se unen de manera temporal a la proteınainvariable relacionada con la clase II (CLIP).
. El complejo MHC II-CLIP pasa al aparato de Golgipara empaquetarse en vesıculas MIIC(compartimento enriquecido en MHC II), quetambi�en reciben epıtopos procesados en losendosomas tardıos.
. En el interior de la vesıcula MIIC, los epıtopossustituyen a las proteınas CLIP; el complejoMHC II-epıtopo se traslada a la membranaplasm�atica para insertarse en ella.
. Las CPA y los macrofagos presentan el complejoMHC-epıtopo a los linfocitos TH, los cualesdeterminan la conveniencia de desplegar unarespuesta inmunitaria.
C�elulas presentadoras de antıgenos
Se conocen dos tipos de CPA:
. Miembros del sistema fagocıtico mononuclear,como macrofagos y c�elulas dendrıticas
. Linfocitos B y c�elulas reticulares del epitelio tımico
Las CPA fagocitan y procesan antıgenos, cargan losepıtopos ası formados enmol�eculasMHC II e insertanel complejo en su membrana plasm�atica para presen-tarlo a los linfocitos T. Las CPA secretan citocinas quemodulan la respuesta inmunitaria, como IL-1, IL-6,IL-12 y TNF-a, ası como otras mol�eculas de senaliza-cion que participan en procesos no inmunitarios.
Interaccion entre las c�elulas linf�aticas
Las c�elulas linf�aticas interaccionan entre sı y analizanlas mol�eculas de superficie de sus homologas. Si no
logran identificar dichas mol�eculas en una c�elula pre-sentadora, el linfocito incapaz de reconocerlas se des-truye mediante apoptosis. En caso contrario, el linfo-cito que reconoce las mol�eculas expuestas en lamembrana se activa, es decir, prolifera y se diferencia.La activacion requiere:
. El reconocimiento del epıtopo.
. El reconocimiento de una senal coestimuladora(libre o de membrana).
Respuesta inmunitaria humoral mediadapor linfocitos TH2
Los linfocitos B internalizan y escinden todos los antı-genos dependientes del timo que forman complejoscon Igs, los asocian a las MHC II e insertan dichocomplejo en su membrana plasm�atica para su presen-tacion a un linfocito TH2 (fig. 12.2).
. Paso 1: El linfocito TH2 reconoce el epıtopo a trav�esde su TCR y el MHC II por medio de su mol�eculaCD4.
. Paso 2: El receptor de CD40 del linfocito TH2 y lamol�ecula CD28 se unen a la mol�eculas CD40 yCD80 del linfocito B, lo que induce la formacion delinfocitos B de memoria y c�elulas plasm�aticas.
CONSIDERACIONES CL�INICAS
El sındrome de inmunodeficiencia adquirida(SIDA) se debe a la infeccion por el virus de lainmunodeficiencia humana (VIH), el cual se une a
las mol�eculas CD4 de los linfocitos TH. El nucleovırico pasa al citoplasma de estos linfocitos tras launion a CD4, lo que tiene efectos debilitadores en
estas c�elulas. El patogeno vırico se multiplica einfecta a otros linfocitos TH, cuya poblaciondisminuye. El paciente no puede desplegar unarespuesta inmunitaria y sucumbe ante infeccionesoportunistas.
Destruccion mediada por linfocitos TH1 de c�elulastransformadas por virus
La capacidad de destruccion de una c�elula transfor-mada por un virus por parte de un LTC depende de lasatisfaccion de las siguientes condiciones:
1. Recepcion por este linfocito de mol�eculas desenalizacion procedentes de un linfocito TH1activado.
2. Union del LTC a lamisma CPA que est�a activandoal linfocito TH1 (fig. 12.3).
176
12
SISTEMALIN
FTICO(INMUNITA
RIO)

177
12
SISTEMALIN
FATIC
O(INMUNITA
RIO)
Figura 12.2 Activacion de linfocitos B por linfocitos TH2 para generar linfocitos B de memoria y c�elulas plasm�aticas que sintetizananticuerpos. No se requieren una respuesta humoral frente a antıgenos independientes del timo ni la interaccion con linfocitos TH2.
(Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 285.)
Figura 12.3 Activacion de los LTC por parte de los linfocitos TH1. El linfocito TH1 y el LTC deben asociarse a lamismaCPA. (Tomado deGartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 286.)�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

. La activacion del linfocito TH1 tiene lugar cuando:. Paso 1: Los TCR y mol�eculas CD4 del linfocitoTH1 han de unirse al complejo epıtopo-MHC II enla CPA, lo que induce la expresion de unamol�ecula de B7 en su plasmalema.
. Paso 2: La mol�ecula CD28 del linfocito TH1 debeunirse a la mol�ecula B7 de la CPA para que elprimero secrete IL-2, IFN-g y TNF.
. La activacion del LTC se produce cuando secumplen estos requisitos:. Paso 1: La mol�ecula CD8 del LTC y el TCR han dereconocer el complejo epıtopo-MHC II de laCPA, y la mol�ecula CD28 del LTC debe unirse a lamol�ecula B7 de la CPA.
. Paso 2: El linfocito TH1 libera IL-2, que se unir�a alreceptor de IL-2 presente en el LTC. El IFN-ginduce la proliferacion del LTC activado.
El LTC activado se une al complejo epıtopo-MHC IIa trav�es de las mol�eculas TCR y CD8 de las c�elulastransformadas, a las que destruye mediante:
. La insercion de perforinas en su membranaplasm�atica, que dan lugar a poros de gran tamano atrav�es de los cuales salen los componentescitosolicos.
. La insercion de perforinas en el plasmalema de lasc�elulas transformadas y la entrada de granzimas alcitoplasma, lo que induce la muerte celular.
. Otra posibilidad es la union del ligando Fas(mol�ecula CD95L, ligando de apoptosis) del LTCa la proteına Fas de la c�elula transformada (CD95,receptor de apoptosis), lo cual suscita la apoptosis.
Los linfocitos TH1 colaboran con los macrofagosen la destruccion de las bacterias fagocitadas
La destruccion de las bacterias fagocitadas por losmacrofagos precisa de su activacion por parte de loslinfocitos TH1. Para ello, estos linfocitos han de acti-varse en primer lugar; a continuacion, el linfocito TH1activado ordena al macrofago que destruya la bacteriaen el seno de sus fagosomas (fig. 12.4). La activaciondel linfocito TH1 se da en dos pasos:
. Paso 1: Las mol�eculas TCR y CD4 del linfocito TH1deben reconocer el complejo epıtopo-MHC II delmacrofago.
. Paso 2: El linfocito TH1 se activa a sı mismomediante la expresion de receptores para IL-2 y lasecrecion de esta citocina, la cual se une a aquellospara inducir la mitosis de estos linfocitos.
Los linfocitos TH1 activados ası formados se unena los macrofagos cuyos fagosomas albergan a las bac-terias.
. Paso 1: Las mol�eculas TCR y CD4 del linfocito TH1han de reconocer el complejo epıtopo-MHC II delmacrofago y el linfocito secreta IFN-g .
. Paso 2: El IFN-g activa al macrofago, el cual liberaTNF-a, que se une a esta c�elula; estos dosmediadores ponen en marcha el proceso dedestruccion de las bacterias fagocitadas mediantela formacion de radicales de oxıgeno.
178
12
SISTEMALIN
FTICO(INMUNITA
RIO)
Figura 12.4 Activacion demacrofagos por parte de los linfocitos TH1. (Tomado deGartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.Philadelphia, Saunders, 2007, p 287.)

Organos linf�aticos
Los organos linf�aticos se dividen en dos clases:
. Organos linf�aticos primarios (centrales) (hıgadofetal, m�edula osea posnatal, y timo), en los que loslinfocitos se convierten en c�elulasinmunocompetentes
. Organos linf�aticos secundarios (perif�ericos)(ganglios linf�aticos, bazo, m�edula osea posnatal ytejido linf�atico asociado a mucosas [MALT]), en losque las c�elulas inmunocompetentes interaccionanentre sı y con antıgenos con el fin de poner enmarcha una respuesta inmunitaria frente apatogenos y antıgenos.
TIMO
El timo, un pequeno organo de origen endod�ermicolocalizado en el mediastino superior, se divide en doslobulos mediante una c�apsula de tejido conjuntivo;los linfocitos T se diferencian en c�elulas inmunocom-petentes en este organo. Comienza a involucionar(degenerar) hacia la pubertad y es infiltrado porc�elulas adiposas, si bien continua funcionando en laedad adulta. Cada lobulo del timo se subdivide enlobulillos incompletos, de modo que cada uno deellos posee una corteza individual y comparte lam�edula con otros lobulillos (fig. 12.5).
La corteza tımicaalberganumerosos linfocitos cuyosnucleos grandes y citoplasma escaso confieren unaspecto basofilo oscuro a las im�agenes histologicas.
179
12
SISTEMALIN
FATIC
O(INMUNITA
RIO)
Figura 12.5 Esquema del timo en el que se representa su histologıa e irrigacion. (Tomado de Gartner LP, Hiatt JL: Color Textbook of
Histology, 3rd ed. Philadelphia, Saunders, 2007, p 288.)�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Los precursores de linfocitos T inmunoincompetentesde la m�edula osea pasan a la corteza del timo para pro-liferar y convertirse en linfocitos T inmunocompetentes.Para ello, estas c�elulas han de interaccionar condistintasc�elulas epiteliales reticulares de la corteza con el fin deexpresar algunos marcadores de superficie y eliminarotros.
. Los precursores de los linfocitos T procedentes dela m�edula osea migran a la union corticomedulardel timo y se dirigen a la corteza externa, en la quereciben el nombre de timocitos.
. Los receptores Notch-1 del plasmalema de estasc�elulas reciben mol�eculas de senalizacion secretadapor las c�elulas epiteliales reticulares de la corteza, loque supone su compromiso con el linaje de loslinfocitos T.
. Los timocitos comienzan a expresar algunosmarcadores de linfocitos T (CD2, pero nocomplejo CD3-TCR y tampoco CD4 ni CD8),por lo que se conocen como timocitos doblenegativos.
. Todos los timocitos doble negativos se desplazanhacia el interior de la corteza (en la periferiamedular) y expresan otras proteınas en susuperficie, que posteriormente ser�an eliminadas.
. Estos timocitos doble negativos expresanreceptores de pre-linfocitos T (pre-TCR) queinducen su proliferacion.
. Estos timocitos reci�en formados expresanmol�eculasCD4 y CD8, lo que los convierte en timocitos doblepositivos.
. Los genes que codifican la region variable del TCRde los timocitos dobles positivos se reorganizan y seexpresan un numero reducido de unidades delcomplejo CD3-TCR en su superficie.
. Las c�elulas reticulares del epitelio corticalverifican la capacidad de reconocimiento decomplejos MHC propias-epıtopos propios delos timocitos doble positivos con escasoscomplejos CD3-TCR.. La mayorıa de los timocitos doble positivos(alrededor del 90%) no reconocen estoscomplejos y sufren procesos de apoptosis; susrestos celulares son fagocitados por losmacrofagos corticales.
. Algunos linfocitos doble positivos (10%)reconocen dichos complejos y pasan a una fase de
maduracion, expresan abundantes TCR einterrumpen la expresion de mol�eculas CD4 yCD8.
. Los linfocitos T que expresanmol�eculas CD4 o CD8se denominan timocitos positivos sencillos, yabandonan la corteza para ingresar en lam�edula deltimo.
. Los timocitos positivos sencillos interaccionan conlas c�elulas epiteliales reticulares de la m�edula y lasc�elulas dendrıticas, que comprueban su capacidadde reconocimiento de epıtopos propios a los que noestuvieron expuestos en la corteza.. Los timocitos positivos sencillos que pondrıanen marcha una respuesta frente a autoantıgenossufren un proceso de apoptosis y son eliminadospor los macrofagos medulares (delecion clonal).
. Los timocitos positivos sencillos que noreconocen antıgenos propios abandonan el timoy migran hacia los organos linf�aticos secundariosen forma de linfocitos T vırgenes.
C�elulas epiteliales reticulares
Existen seis tipos de c�elulas epiteliales reticulares,tres de los cuales se localizan en la corteza y otrostantos lo hacen en la m�edula:
. Las c�elulas de tipo I aıslan la corteza de la c�apsula detejido conjuntivo y las trab�eculas, y envuelven losvasos sanguıneos corticales.
. Las c�elulas de tipo II se hallan en la zonamedia de lacorteza y rodean islotes de timocitos; presentanantıgenos propios y mol�eculas MHC I y II a lostimocitos.
. Las c�elulas de tipo III se localizan en la zona deunion corticomedular, presentan antıgenos propiosy mol�eculas MHC I y II a los timocitos.
. Las c�elulas de tipo IV se encuentran en la zona deunion corticomedularde lam�edula; colaborancon lasde tipo III en la separacion de la corteza y la m�edula.
. Las c�elulas de tipo V construyen el marco estructuralde la m�edula.
. Las c�elulas de tipo VI forman corpusculos tımicos(de Hassall), sintetizan linfopoyetina estromaltımica que favorece la delecion clonal, y colaboranen la induccion de la apoptosis en linfocitos Tpositivos sencillos.
180
12
SISTEMALIN
FTICO(INMUNITA
RIO)

CONSIDERACIONES CL�INICAS
Los vasos sanguıneos que irrigan el timo llegan a la
m�edula y formanun lechocapilar en la zonadeunion
de lacortezay lam�edula.Lasramificacionesdeestoscapilarespasana lacortezayse rodeande inmediato
deunavainadec�elulasepiteliales reticularesde tipo Ique semantienen unidas por medio de fascias
oclusivas. Estas c�elulas configuran la barrerahematotımica en la corteza de este organo, la cual
impide el pasodemacromol�eculas transportadas enel torrente circulatorio a la corteza para alterar el
desarrollode los linfocitosT.Lasc�elulasepitelialesde
los capilares corticales y las c�elulas epiteliales
reticulares de tipo I est�an dotadas de una l�aminabasal propia que refuerza dicha barrera. Losmacrofagos vigilan el espacio comprendido entre lavaina epitelial y el endotelio y destruyen las
macromol�eculasque logranaccederal timodesdeeltorrente circulatorio. Las venas de la corteza del timo
son tributarias de la red venosa de la m�edula.
Algunos sujetos con ausencia cong�enita del timo,un trastorno conocido como sındrome de DiGeorge,no pueden fabricar linfocitos T, por lo que son inca-paces de desplegar una respuesta inmunitaria celular.
Casi todos estos pacientes son inmunodeficientes, yaque el inicio de la mayorıa de las respuestas inmuni-tarias humorales precisa de la intervencion de los lin-focitos TH. Los sujetos afectados pueden sobrevivirsiempre y cuando no contraigan ninguna infeccion;no obstante, la mayor parte de ellos muere debido ainfecciones o bien a tetania calcio (hipocalcemiagrave) debida a la ausencia cong�enita concomitantede las gl�andulas paratiroides.
GANGLIOS LINFATICOS
Los ganglios linf�aticos son estructuras ovaladas depequeno tamano (di�ametro � 3 cm) con una super-ficie convexa y otra concava (hilio) envueltas por unac�apsula conjuntiva (fig. 12.6) que suele rodearse detejido adiposo. Por debajo de la c�apsula, el par�en-quima del ganglio se subdivide en:
. Una corteza externa, la cual alberga linfocitos B queconstituyen los nodulos linf�aticos primarios ysecundarios.
. Una paracortezamedia, que contiene linfocitos TH.
. Una m�edula profunda, en la que las c�elulaspredominantes son los linfocitos,las c�elulas plasm�aticas y los macrofagos.
181
12
SISTEMALIN
FATIC
O(INMUNITA
RIO)
Figura 12.6 Diagrama de un ganglio linf�atico tıpico. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia,
Saunders, 2007, p 291.)�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

182
12
SISTEMALIN
FTICO(INMUNITA
RIO)
La c�apsula de la superficie convexa proyectatrab�eculas hacia la corteza que la subdividen en com-partimentos incompletos; conforme se extienden haciala paracorteza y lam�edula, las trab�eculas se tornanm�astortuosas y menos definidas (v. fig. 12.6). Los ganglioslinf�aticos contienen linfocitos T, linfocitos B, c�elulasdendrıticas, macrofagos y CPA; se ocupan de depurarla linfa y poner en marcha reacciones inmunitariasfrente a antıgenos exogenos. La linfa ingresa en el gan-glio linf�atico a trav�es de los vasos linf�aticos aferentes,
que perforan la superficie convexa y poseen v�alvulasque impiden el flujo retrogrado de la linfa. La linfaatraviesa el par�enquima del ganglio, del que sale atrav�es de los vasos linf�aticos eferentes, tambi�en dota-dos de v�alvulas que evitar el regreso de la linfa a niveldel hilio. Las arterias ingresan en el ganglio linf�aticoen el hilio, desde el que parten las venas; estos vasossanguıneos atraviesan el par�enquima ganglionar atrav�es de las trab�eculas. En la paracorteza, las venasforman v�enulas del endotelio alto (VEA).
CONSIDERACIONES CL�INICAS
En los sujetos sanos, los ganglios linf�aticos sonexcesivamente blandos y no pueden palparse. Sinembargo, en presencia de una infeccion regional, los
linfocitos del ganglio que drena el �area afectadaproliferan, de modo que el ganglio se hipertrofia, setorna duro, doloroso y palpable sin dificultad. Una
cadena de ganglios linf�aticos conectados entre sı
por vasos linf�aticos drena cada region del
organismo. Esta secuencia de ganglios linf�aticossuele permitir la diseminacion a distancia de
infecciones o lamet�astasis de c�elulas tumorales. Losmacrofagos fagocitan alrededor del 99% de lamateria particulada inutil para eliminarla conforme
percola la linfa a trav�es de los sinusoides del ganglio
linf�atico.
Las CPA que han detectado algun antıgeno migran
hacia el ganglio linf�atico m�as proximo a ellas,presentan el complejoMHC-epıtopo a los linfocitos Tcooperadores y desencadenan una respuesta
inmunitaria. Estas c�elulas reciben el nombre de
c�elulas dendrıticas migratorias cuando se hallan
en el seno del ganglio linf�atico.
Las c�elulas dendrıticas foliculares captan los
antıgenos que son transportados al ganglio linf�atico
a trav�es de los vasos linf�aticos aferentes parapresentarlos a los linfocitos residentes.
El reconocimiento del antıgeno supone la activacionde los linfocitos B en la zona de contacto de laparacorteza y la corteza, que migran hacia el nodulo
linf�atico primario y sufren divisiones celulares r�apidaspara formar el centro germinal que transforma el
nodulo linf�atico primario en un nodulo secundario.Las c�elulas dendrıticas foliculares inducen laapoptosis en los linfocitos B que expresan Igsincorrectas, mientras que aquellas con Igs normalespueden diferenciarse en linfocitos B de memoria y
c�elulas plasm�aticas. Las c�elulas reci�en diferenciadasmigran hacia la m�edula del ganglio y formancordones medulares. Alrededor del 90% de las
c�elulas plasm�aticas utilizan los vasos linf�aticos
eferentes para salir del ganglio linf�atico y migrar hacia
la m�edula osea, en la que sintetizar�an y secretar�ananticuerpos hasta su muerte. El 10% restantepermanece en el cordon medular y fabricaanticuerpos hasta el final de su vida. De igual modo, lamayorıa de los linfocitos B de memoria abandona el
ganglio linf�atico original para colonizar otros organos
linf�aticos secundarios, en los que formar�an clonesreducidos a la espera de una nueva exposicion alantıgeno correspondiente. Una pequena poblacion de
linfocitos B de memoria subsiste en el ganglio linf�aticoinicial, en el que crea un clon formado por un reducido
numero de c�elulas.

La c�apsula de tejido conjuntivo define la fronterasuperior de los compartimentos incompletos de la cor-teza del ganglio linf�atico, mientras que las trab�eculasemitidas por dicha c�apsula constituyen sus lımites late-rales (v. fig. 12.6). Amedidaque atraviesan la c�apsula, losvasos linf�aticos aferentes vacıan la linfa en el seno sub-capsular,desde los que sedirige a los senosparatrabecu-lares,paralelos a las trab�eculas, quedesembocan enunossenosmedularesmuy tortuosos quedrenan en los vasoslinf�aticos eferentes. Los senos linf�aticos est�an tapizadospor c�elulas endoteliales escamosas simples y su luz con-tieneuna redde c�elulas reticulares estrelladasque ralen-tizan el flujo linf�atico, adem�as de sustentar a losmacro-fagos que fagocitan antıgenos particulados.
Los compartimentos corticales contienen unosnodulos linf�aticos primarios o secundarios de morfo-logıa esf�erica y coloracion oscura.
. La formacion de losnodulos secundarios (v. fig. 12.6)obedece a la estimulacion antig�enica; en estasestructuras se desarrollan los linfocitos B(centroblastos) que no expresan, aun, Igs. Laproliferacion inicial de estas c�elulas tiene lugar en lazonam�as oscura, ym�as tarde se desplaza haciala zona central clara (el llamado centro germinal); loscentroblastosdesplazanaloslinfocitosBenreposo,quese alejan para formar un densomanto (corona) querecubreel centrogerminalhaciael senosubcapsular.Enel folıculo secundario aparecen, adem�as:. C�elulas dendrıticas migratorias, como lasc�elulas de Langerhans de la piel, unas c�elulasderivadas de la m�edula osea que se distribuyen entodo el organismo; migran al ganglio linf�aticom�as proximo al detectar antıgenos exogenos conel fin de iniciar una respuesta inmunitaria.
. C�elulas dendrıticas foliculares, que no provienende lam�edula osea y residen en el ganglio linf�atico; seencargan de la presentacion de antıgenos a loscentrocitos,quecorrespondena linfocitosB jovenesque expresan Igs. Las c�elulas dendrıticas folicularesinducenlaapoptosisdelos linfocitosBportadoresdeIgs erroneos y permiten la diferenciacion de otroslinfocitos B en linfocitos B dememoria y c�elulasplasm�aticas,quemigranhacia lam�edula y salen delganglio linf�atico.
. C�elulas reticulares, las cuales sintetizan fibras decol�ageno de tipo III (fibras reticulares) queconforman la estructura arquitectonica de losganglios linf�aticos.
. Macrofagos, que eliminan a las c�elulasapoptosicas.
. Losnodulos primarios (v. fig. 12.6) son nodulos enreposo, ya que carecen de centros germinales ymanto; lamigracion hacia el nodulo primario de loslinfocitos B activados por los linfocitos Tcooperadores en la periferia de la corteza y la
paracorteza supone su transformacion en unnodulo secundario.
La paracorteza (v. fig. 12.6) es una region gan-glionar en la que abundan los linfocitos T. Los linfo-citos B y T acceden al ganglio linf�atico a trav�es de lasVEA. Los primeros migran a la corteza, mientras queestos ultimos permanecen en la paracorteza.
La m�edula (v. fig. 12.6) contiene sinusoides medu-lares, trab�eculas y cordonesmedulares,unas estructurasformadas por fibras reticulares, c�elulas reticulares ymacrofagos, ası como linfocitos B y c�elulas plasm�aticasformadas en los folıculos linf�aticos secundarios.
BAZO
El bazo posee una c�apsula de tejido conjuntivo cola-genoso irregular denso recubierta por el peritoneo, unepitelio escamoso simple. Constituye el organo lin-f�atico de mayor tamano; presenta una superficie con-vexa y otra concava, el hilio, a partir del cual la c�apsulaemite trab�eculas de tejido conjuntivo que transportana los vasos sanguıneos y nervios hacia el par�enquima deeste organo. La estructura fısica del bazo se sustenta enuna red tridimensional de fibras de col�ageno de tipoIII con c�elulas reticulares asociadas que aparece ado-sada a la c�apsula y las trab�eculas. A diferencia de losganglios linf�aticos, el bazo no consta de una corteza,una paracorteza y una m�edula, sino que se divide en lapulpa blanca, la zona marginal y la pulpa roja(dotada de abundantes sinusoides tortuosos) que seentremezclan (fig. 12.7) para desempenar las fun-ciones de las que se ocupa este organo:
. Filtrado de sangre y destruccion de eritrocitossenescentes
. Formacion de linfocitos T y B e inicio de respuestasinmunitarias
. Hematopoyesis fet al y, en caso necesario, enadultos.
Irrigacion del bazo
La arteria de gran calibre que irriga el bazo, la arteriaespl�enica, se ramifica varias veces antes de atravesar elhilio para recorrer el par�enquima de este organo(fig. 12.8; v. fig. 12.7).
. Los vasos viajan a trav�es de las trab�eculas en formade arterias trabeculares, las cuales emiten un grannumero de ramificaciones de calibre decrecienteconforme disminuye el tamano de aquellas.
. Las arterias abandonan la trab�ecula correspondienteal alcanzar un di�ametro � 200mm; su tunicaadventicia se despliega para rodearse de una vainade linfocitos T, la vaina linf�atica periarterial(VLPA). La arteria que ocupa el centro de esta vainarecibe el nombre de arteria central.
183
12
SISTEMALIN
FATIC
O(INMUNITA
RIO)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

. Conforme disminuye su calibre, la arteria centralpierde su VLPA y origina una serie de arteriolasrectas cortas paralelas que ingresan en la pulpa roja,la arterias peniciliares, cada una de las cuales constade tres secciones:. Arteriola de la pulpa. Arteriola envainada por una capa de macrofagos(vaina de Schweigger-Seidel)
. Capilar arterial terminal, que desembocadirectamente en un sinusoide (circulacioncerrada), en el par�enquima de la pulpa rojaen las proximidades de un sinusoide (circulacionabierta) o bien, como suponen muchosinvestigadores, en circulaciones abiertas ycerradas.
. Las venas de la pulpa (v. fig. 12.8) reciben sangre delos sinusoides y desembocan en venas de mayorcalibre, asociadas a las arterias de tamanocorrespondiente en las trab�eculas, que conducen alas venas grandes del hilio, en las que se fusionanpara formar la vena espl�enica.
Pulpa blanca, zona marginal y pulpa roja
El bazo se compone de pulpa blanca, zona marginal ypulpa roja.
. La pulpa blanca corresponde a la vaina delinfocitos T, los VLPA, que rodea a la arteriacentral. A menudo, el VLPA alberga un nodulolinf�atico, formado por linfocitos B, de tal modo que
184
12
SISTEMALIN
FTICO(INMUNITA
RIO)
Figura 12.7 Diagrama del bazo. (Tomado deGartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 294.)

los linfocitos T se disponen alrededor de unaacumulacion esf�erica de linfocitos B. Los nodulosque se encuentran inmersos en una respuestainmunitaria contienen, adem�as, un centrogerminal. En el bazo, al igual que en los ganglioslinf�aticos, los linfocitos B y T aparecen enlocalizaciones especıficas (v. figs. 12.7 y 12.8).
. La zona marginal, una region con un grosorcercano a 1/10mm, representa el �areade separacion de ambas pulpas (v. fig. 12.7).Aparecen c�elulas dendrıticas interdigitantes(CPA), macrofagos, c�elulas plasm�aticas,y linfocitos T y B. En esta zona son abundantesunos sinusoides pequenos, los sinusoidesmarginales. Los capilares procedentes de la arteriacentral recorren durante una distancia corta la pulparoja, salen de ella y desembocan en los senosmarginales.
. La pulpa roja (v. figs. 12.7 y 12.8) constade espacios vasculares, los sinusoides, queaparecen rodeados del estroma de la pulpa roja,los cordones espl�enicos, integrados por una redde fibras reticulares rodeadas de una capa c�elulasreticulares estrelladas que evitan el contactode las fibras de col�ageno con la sangre extravasadaen sus intersticios y el inicio de la cascadade coagulacion. Las c�elulas endoteliales de lossinusoides son especiales, ya que muestranuna morfologıa fusiforme y sus ejes longitudinalesse disponen en paralelo al eje mayor de lossinusoides. El endotelio es relativamentepermeable por la presencia de espacios ampliosentre c�elulas adyacentes, que utilizan las c�elulassanguıneas para pasar con facilidad de la luzhacia los cordones espl�enicos. Las escasasfibras reticulares filiformes, envueltas por unmaterial similar a una l�amina basal discontinua,rodean el revestimiento endotelial de lossinusoides.
Las funciones del bazo presentan una estrecha rela-cion con la anatomıa de la red vascular encargada desu irrigacion.
. La primera region de contacto de la sangre con elpar�enquima espl�enico corresponde a lossinusoidesmarginales, en los que lasCPA tratan dedetectar antıgenos y los macrofagos atacan a losmicroorganismos presentes en el torrentecirculatorio. Los linfocitos T y B migran hacia laVLPA y los nodulos linf�aticos desde los vasossanguıneos.
. En la zona marginal tiene lugar la presentacion delcomplejo MHC-epıtopo a los linfocitos T por partede las c�elulas dendrıticas interdigitantes. Elreconocimiento de antıgenos dependientes deltimo por los linfocitos B pone en marcha larespuesta inmunitaria; estas c�elulas se diferencianen c�elulas plasm�aticas, la mayorıa de las cuales sedesplaza hasta la m�edula osea y fabrica anticuerpos.
. El material que no es destruido en la zonamarginal,como plaquetas y eritrocitos senescentes, pasa a lossinusoides de la pulpa roja, en la que ser�aeliminado por los macrofagos. Los eritrocitossenescentes pierden los residuos de �acido si�alico yportan residuos de galactosa en sus membranasplasm�aticas, lo que induce su reconocimiento.
TEJIDO LINFATICO ASOCIADO A LA MUCOSA
Las mucosas de los aparatos respiratorio, gastrointes-tinal y urinario contienen agregados no encapsuladosde nodulos linf�aticos e infiltraciones linfocıticas lla-mados MALT, como el tejido linf�atico asociado alintestino (GALT), el tejido linf�atico asociado a losbronquios (BALT) y las amıgdalas.
. Los folıculos linf�aticos presentes en la mucosa deltubo digestivo, que reciben en nombre de GALT,est�an formados por linfocitos B que se asocian a
185
12
SISTEMALIN
FATIC
O(INMUNITA
RIO)
Figura 12.8 Diagrama de la circulacion cerrada y
abierta del bazo.
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

linfocitos T perif�ericos. El GALT m�as conocido sonlas placas de Peyer, localizadas en la mucosa delıleon (fig. 12.9A). Las arteriolas que irrigan estasplacas desembocan en venas, algunas de las cualesson VEA que permiten la salida de linfocitos ymacrofagos desde su luz (fig. 2.9B-D). Las c�elulasM(c�elulas de micropliegues), asociadas a las placasde Peyer, engullen antıgenos presentes en la luzintestinal y los transfieren a las CPA de dichas placaspara su procesamiento.
. El BALT presenta una morfologıa y una funcionsimilares a las del GALT, si bien los folıculos sehallan en la mucosa del �arbol respiratorio.
Amıgdalas
Las amıgdalas, unos agregados de nodulos linf�aticosdotados de c�apsulas incompletas (amıgdalas palatinas,farıngeas, linguales y un gran numero de amıgalas depequeno tamano) se encuentran en la entrada de la fa-ringe bucal, a la que confieren proteccion frente a an-tıgenos inhalados. Los linfocitos se activan como con-secuencia de la exposicion antig�enica y proliferan, loque conduce a la hipertrofia de la amıgdala afectada.
. Las amıgdalas palatinas bilaterales, localizadasentre los pliegues palatogloso y palatofarıngeo, se
recubren de un epitelio escamoso estratificado ypresentan alrededor de una docena de criptasprofundas que pueden albergar alimentos y otrosresiduos, ası como microorganismos y c�elulasepiteliales descamadas. En el par�enquima de estasamıgdalas abundan los nodulos linf�aticos, algunosde los cuales contienen centros germinales. La carainterna de las amıgdalas posee una c�apsula fibrosadensa.
. La amıgdala farıngea unilateral se encuentra en lafaringe nasal; presenta una estructura semejante alas amıgdalas palatinas, aunque se rodea de unepitelio respiratorio y posee repliegues superficiales,llamados plegamientos, en lugar de criptas y unac�apsula m�as delgada. La amıgdala farıngeainflamada recibe el nombre de adenoide.
. Las amıgdalas linguales, presentes en la superficiedorsal del tercio posterior de la lengua, se recubrende un epitelio escamoso estratificado que tapizalas abundantes criptas, en cuya base desembocanlas gl�andulas salivales menores mucosasposteriores. La superficie profunda de estasamıgdalas presenta una fina c�apsula. El par�enquimade las amıgdalas linguales est�a formado pornodulos linf�aticos, muchos de los cuales poseencentros germinales.
186
12
SISTEMALIN
FTICO(INMUNITA
RIO)

187
12
SISTEMALIN
FATIC
O(INMUNITA
RIO)
Figura 12.9 Microfotografıas de transmision electronica. A. Vaso ALPA (L) del �area interfolicular en la que abundan los linfocitos;
se observa un canal intraendotelial que contiene linfocitos (flecha) en la pared endotelial (3.000�). B-D.Cortes ultrafinos en los que semuestran las distintas etapas que atraviesa un canal endotelial formado por una (1) y dos (2) c�elulas endoteliales (9.000�). l, linfocito.
(Tomado de Azzali G, Arcari MA: Ultrastructural and three-dimensional aspects of the lymphatic vessels of the absorbing peripheral lymphatic
apparatus in Peyer’s patches of the rabbit. Anat Rec 258:76, 2000.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

13 SISTEMA ENDOCRINOEl sistema nervioso autonomo y el sistema endocrinose ocupan del mantenimiento de la homeostasis y elcontrol del metabolismo de ciertosorganos y sistemas org�anicos. El pri-mero actua con rapidez mediante laliberacion de mol�eculas neurotrans-misoras en el medio que circunda alsistema org�anico de inter�es, mientrasque el segundo lo hace de maneram�as lenta y a distancia a trav�es dela secrecion de hormonas, unasmol�eculas mensajeras que utilizan eltorrente circulatorio para alcanzarel organo diana. Sin embargo, estosdos sistemas separados funcionanjuntos para dirigir las actividadesmetabolicas del cuerpo. El sistemaendocrino se compone de:
. Gl�andulas dotadas de una red vascular profusa: lagl�andula hipofisis, la gl�andula tiroides, la gl�andulaparatiroides, las gl�andulas suprarrenales y el cuerpopineal.
. Agregados de c�elulas endocrinas, como los islotesde Langerhans en el p�ancreas.
. C�elulas endocrinas solitarias dispersas en elrevestimiento epitelial del tubo digestivo y elaparato respiratorio (c�elulas del sistemaneuroendocrino difuso).
Hormonas
Las hormonas se dividen en tres grupos en funcion desu naturaleza quımica:
. Proteınas y polip�eptidos, como insulina yhormona luteinizante (LH), son mol�eculashidrofilas que se unen a receptores de la superficieextracelular de la membrana plasm�atica.
. Derivados de amino�acidos, como tiroxinay noradrenalina, son compuestos hidrofobos que seunen a receptores extracelulares del plasmalema.
. Derivados de esteroides y �acidos grasos, comoestrogenos y androgenos, sonmol�eculas hidrofobasque se unen a receptores citosolicos.
La union de una hormona a su receptor (ya sea areceptores de la superficie extracelular o a recep-tores intracelulares) pone en marcha una cadena de
transduccion de senales que representa la reaccioncelular frente a la hormona. La transduccion de
senales iniciada por la union a unreceptor de la superficie extracelularorigina la activacion de:
. Una proteına cinasa, la cual activaa ciertas proteınas reguladoras,como la ciclasa de adenilato,que sintetizan un segundomensajero, el monofosfatocıclico de adenosina. Otrossistemas utilizan mensajerosdiferentes, como el monofosfatocıclico de guanina, derivados defosfatidilinositol, iones de calcioe iones de sodio.
. Proteınas G, que activan un sistemade segundos mensajeros.
. Receptores con actividad catalıtica, los cualesactivan diversas proteına cinasas para poneren marcha una cascada de fosforilacion.
La transduccion de senales desencadenada por launion a un receptor intracelular comienza a raız dela entrada del complejo hormona-receptor en elnucleo, en el que se une a una region del ADN pro-xima a un promotor para inducir la transcripcion deARN mensajero (ARNm) que se traducir�a, en ultimainstancia, en la proteına deseada.
Cuando la cantidad de hormona liberada es insu-ficiente para poner en marcha la transduccion desenales, la c�elula diana utiliza un mecanismode retroalimentacion positiva con el fin de garanti-zar la secrecion de la cantidad necesaria de este pro-ducto. No obstante, la activacion de una c�elula dianapuede dar lugar a una respuesta inhibitoria adem�asde la respuesta deseada, en la cual se sintetiza unamol�ecula de senalizacion que induce unmecanismode retroalimentacion que desactiva a la gl�andula/c�elula endocrina para detener la liberacion de lahormona.
Hipofisis
Lahipofisis, una gl�andula que fabrica numerosas hor-monas, se encuentra suspendida del hipot�alamo y selocaliza en la silla turca del esfenoides (fig. 13.1). Esta
� 2011 Elsevier Espana, S.L. Reservados todos los derechos
TERMINOS CLAVE. Hormonas. Hipofisis. Vıa
hipotalamohipofisaria. Gl�andula tiroides. Gl�andulas
paratiroides. Corteza suprarrenal. M�edula suprarrenal. Gl�andula pineal
188

gl�andula de pequeno tamano, similar al de un gui-sante, proviene de dos fuentes diferentes:
. La neurohipofisis procede de la evaginacion deldienc�efalo.
. La adenohipofisis representa una evaginacion de lacavidad bucal (bolsa de Rathke).
La hipofisis y su red vascular reciben fibras nerviosasy neurotransmisores de origen hipotal�amico, respecti-vamente, que regulan la liberacion de las hormonasproducidas por esta gl�andula o almacenadas en ella. En
la hipofisis se distinguen la adenohipofisis (hipofisisanterior) y la neurohipofisis (hipofisis posterior),cada una de las cuales se subdivide en otros comparti-mentos (tabla 13.1). Las c�elulas residuales de la bolsa
189
13
SISTEMAENDOCRINO
Figura 13.1 Hipofisis y organos diana. ACTH, hormona adrenocorticotropa; ADH, hormona antidiur�etica; FSH, hormona foliculo-
estimulante; TSH, tirotropina. (Tomado deGartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 305.)
Tabla 13.1 COMPONENTES DE LA HIPOFISIS
Adenohipofisis(hipofisis anterior)
Neurohipofisis(hipofisis posterior)
Parte distal (parte anterior) Eminencia media
Parte intermedia Infundıbulo
Parte tuberosa Parte nerviosa
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

de Rathke se insertan entre la adenohipofisis y la neu-rohipofisis en vesıculas rellenas de material coloidal. Elinfundıbulo se rodea de una envoltura de c�elulas endo-crinas que recibe el nombre de parte tuberosa. La irri-gacion de la hipofisis proviene de las arterias supe-riores e inferiores de la hipofisis, las cuales parten dela arteria carotida interna.
Las dos arterias superiores de la hipofisis irrigan elinfundıbulo y la parte tuberosa; se ramifican para darlugar al plexo capilar primario (formado por capi-lares fenestrados) de la eminencia media. Las arteriasinferiores de la hipofisis irrigan fundamentalmente ala hipofisis posterior. La vena porta de la hipofisisdrena el lecho capilar primario y desemboca en ellecho capilar secundario (que tambi�en contiene capi-lares fenestrados), el cual irriga la adenohipofisis.
Los axones de las neuronas que inervan elhipot�alamo terminan en el �area del lecho capilar pri-mario, en el que liberan abundantes hormonas neu-rosecretoras hipotal�amicas (hormonas liberadoraso inhibidoras), que alcanzan dicho lecho. Las venasporta de la hipofisis transportan las hormonas neuro-secretoras al lecho capilar secundario. El hipot�alamoregula la actividad de la adenohipofisis a trav�es de laliberacion de ciertas hormonas (factores) que se enu-meran en la tabla 13.2.
ADENOHIPOFISIS (HIPOFISIS ANTERIOR)
La adenohipofisis, la cual se desarrolla a partir de labolsa de Rathke, posee tres regiones: la parte distal,la parte intermedia y la parte tuberosa (fig. 13.2).
. La c�apsula de la parte distal emite fibras reticulareshacia la sustancia de los cordones que sustentan alas c�elulas parenquimatosas y los capilaressinusoides del lecho capilar secundario. Estasc�elulas parenquimatosas son de dos tipos: 1) c�elulascuyos gr�anulos secretores captan coloranteshistologicos, llamadas cromofilas, y 2) c�elulascuyos gr�anulos secretores no adquieren estoscolorantes, y reciben el nombre de cromofobas. Losgr�anulos de algunas cromofilas presentan una
mayor afinidad por colorantes �acidos (acidofilas),mientras que los de otras lo hacen por los b�asicos(basofilas).. Las c�elulas acidofilas,m�as abundantes, se dividenen: somatotropas, que sintetizansomatotropina, la hormona del crecimiento, ymamotropas, que secretan prolactina, lahormona que favorece el desarrollo de lasgl�andulas mamarias en la mujer gestante y lalactancia para alimentar al neonato.
. Las c�elulas basofilas se encuentran en la periferiade la parte distal. Se distinguen tres subtipos:1) corticotropas, que sintetizan hormonaadrenocorticotropa (ACTH) y hormonalipotropa; 2) tirotropas, que fabricantirotropina, y 3) gonadotropas, que secretanhormona foliculoestimulante (FSH) y hormonaluteinizante (LH).
. Las c�elulas cromofobas poseen un citoplasmaescaso, un pequeno numero de gr�anulossecretores y no captan colorantes histologicos. Secree que podrıan representar c�elulas cromofilasque han secretado sus gr�anulos secretores, aunquetambi�en se ha propuesto que podrıa tratarse dec�elulas madre. Las c�elulas m�as destacadas de laparte distal son las c�elulas foliculoestrelladas, defuncion desconocida.
. La parte intermedia (zona intermedia), localizadaentre la parte anterior y la parte nerviosa, contienequistes rellenos de material coloidal derivados de labolsa de Rathke y agregados de basofilos quesintetizan proopiomelanocortina. La escision deesta prohormona a lugar a las hormonasestimuladoras de melanocitos a (a-MSH),endorfina b, corticotropina y lipotropina. Adiferencia de los animales inferiores, la a-MSHinduce la liberacion de prolactina en el ser humanoy recibe el nombre de factor liberador deprolactina.
. La parte tuberosa envuelve parcialmente el tallohipofisario. No parece sintetizar ninguna hormona,si bien algunas de sus c�elulas contienen FSH y LH.
190
13
SISTEMAENDOCRINO

191
13
SISTEMAENDOCRINO
Figura 13.2 Hipofisis y organos diana. ACTH, hormona adrenocorticotropa; ADH, hormona antidiur�etica; FSH, hormona foliculo-
estimulante; TSH, tirotropina. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 305.)
Tabla 13.2 HORMONAS LIBERADORAS DEL HIPOTALAMO Y SUS FUNCIONES
Hormona liberadora Funcion
Tiroliberina Liberacion de tirotropinaCorticoliberina Liberacion de adrenocorticotropina
Somatoliberina Liberacion de somatotropina (hormona del crecimiento)
Luliberina Liberacion de LH y FSH
Hormona liberadora de prolactina Liberacion de prolactinaHormona inhibidora de prolactina Inhibe la secrecion de prolactina
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

192
13
SISTEMAENDOCRINO
NEUROHIPOFISIS
La neurohipofisis (o hipofisis posterior) se desarrollaa partir del hipot�alamo y se divide en tres regiones(figs. 13.3 y 13.4): eminencia media, infundıbulo yparte nerviosa. Se considera que la neurohipofisisconstituye una prolongacion del hipot�alamo. La vıahipotalamohipofisaria est�a formada por los axonesno mielinizados de c�elulas neurosecretoras localiza-das en los dos nucleos hipotal�amicos:
. Supraoptico
. Paraventricular
Las c�elulas neurosecretoras de estos nucleos sin-tetizan hormona antidiur�etica (ADH, vasopresina)
y oxitocina, ası como la proteına transportadoraneurofisina, a la que se asocian las hormonas(fig. 13.5).
Parte nerviosa
La vıa hipotalamohipofisaria concluye en la parte ner-viosa; los pituicitos, unas c�elulas similares a las de laglıa tıpicas de esta region hipofisaria, sostienen susaxones. Las hormonas ADH y oxitocina se almacenanen estado activo en unas distensiones axonales co-nocidas como cuerpos de Herring; se liberan, bajodemanda, en la proximidad del lecho capilar fenes-trado creado por las dos arterias inferiores de la hipo-fisis (tabla 13.3; v. tabla 13.2).
CONSIDERACIONES CL�INICAS
Los adenomas hipofisarios son unos tumoresfrecuentes de la adenohipofisis. La hipofisis seencuentra en la fosa hipofisaria del esfenoides, por loque su crecimiento e hipertrofia inciden en la sıntesishormonal en la parte distal. En ausencia detratamiento, estos tumores llegan a erosionar elhueso y otros tejidos neurales.
La diabetes insıpida podrıa presentar una relacion
con alteraciones en el hipot�alamo, la parte nerviosa,o ambas, que reduzcan la produccion deADH, lo queocasionarıa una disfuncion renal debida a laimposibilidad de concentrar la orina. Los sujetosafectados pueden ingerir cantidades enormes delıquidos y secretar hasta 20 l de orina al dıa (poliuria).

193
13
SISTEMAENDOCRINO
Figura 13.4 Hipofisis y organos diana. ACTH, hormona adrenocorticotropa; ADH, hormona antidiur�etica; FSH, hormona foliculo-
estimulante; TSH, tirotropina. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 305.)
Figura 13.3 Hipofisis y su irrigacion. (Tomado de
Gartner LP, Hiatt JL: Color Textbook of Histology,
3rd ed. Philadelphia, Saunders, 2007, p 306.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

194
13
SISTEMAENDOCRINO
Tabla 13.3 EFECTOS FISIOLOGICOS DE LAS HORMONAS HIPOFISARIAS
Hormona Liberacion/inhibicion Funcion
Parte distal
Somatotropina (hormona
del crecimiento)
Liberacion: SRH/inhibicion:
somatostatina
Efecto generalizado de aumento de tasas metabolicas en la
mayorıa de las c�elulas; estimulacion de la secrecion de
somatomedinas (factores de crecimiento insulinoides I
y II) por hepatocitos, que inducen la proliferacion delcartılago y favorecen el crecimiento de los huesos largos
Prolactina Liberacion: PRH/inhibicion: PIF Favorece el desarrollo de gl�andulas mamarias durante el
embarazo; estimula produccion de leche tras el parto
(la succion estimula la secrecion de prolactina)Hormona adrenocorticotropa
(ACTH, corticotropina)
Liberacion: CRH Estimula la sıntesis y la liberacion de hormona (cortisol y
corticosterona) por corteza suprarrenal
FSH Liberacion: LHRH/inhibicion:inhibina (en el hombre)
Estimula el desarrollo de folıculos ov�aricos secundarios y lasecrecion de estrogenos; estimula la sıntesis de proteına
de union a androgenos por c�elulas de Sertoli de tubulos
seminıferos
LH Liberacion: LHRH Colabora con la FSH en induccion de la ovulacion,formacion del cuerpo luteo y secrecion de progesterona y
estrogenos; interviene en la retroalimentacion negativa
para inhibir la secrecion de LHRH por el hipot�alamo en
la mujerHormona estimuladora de
c�elulas intersticiales (ICSH)
en el hombre
Estimula las c�elulas de Leydig para que secreten y liberen
testosterona, lo que constituye un circuito de
retroalimentacion negativa para inhibir la secrecion deLHRH por el hipot�alamo en hombres
TSH (tirotropina) Liberacion: TRH/inhibicion:
suprime retroalimentacion
negativa a trav�es del SNC
Estimula la sıntesis y la liberacion de hormona tiroidea,
la cual incrementa la tasa metabolica
Parte nerviosa
Oxitocina Estimula la contraccion del musculo liso uterino durante
el orgasmo; estimula la contraccion del musculo
liso uterino durante el parto (la estimulacion del cuellouterino envıa una senal al hipot�alamo para secretar
cantidades mayores de oxitocina); la succion remite una
senal al hipot�alamo, que incrementa la sıntesis de
oxitocina, lo que provoca la contraccion de las c�elulasmioepiteliales de las gl�andulas mamarias y, por tanto, la
salida de la leche
Vasopresina (hormonaantidiur�etica [ADH])
Conserva el contenido hıdrico al potenciar la resorcionrenal de agua; podrıa estar controlada por la presion
osmotica; provoca la contraccion del musculo liso de las
arterias, lo que incrementa la presion arterial; puede
restablecer la presion arterial normal tras unahemorragia intensa
PIF, factor inhibidor de la liberacion de prolactina; SNC, sistema nervioso central.Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 307.

CONSIDERACIONES CL�INICAS
El t�ermino bocio no toxico se refiere a una hipertrofia
de la gl�andula tiroides que carece de relacion con unaproduccion excesiva de hormona tiroidea o un tumor
maligno. La hipertrofia de esta gl�andula puedeobedecer a diversas causas. La carencia de yodo enla dieta origina bocio, si bien se trata de una etiologıainfrecuente debido a la presencia de este mineral en
los alimentos. Una causa m�as comun de bocio es elaumento de la tirotropina (TSH) debido a la sıntesis
hormonal deficiente en la gl�andula tiroides. En estospacientes, la TSH estimula la hipertrofia de la
gl�andula a lo largo de varios anos. La mayor parte delas alteraciones de tamano pequeno a moderado sonsusceptibles de tratamiento con la hormona tiroideaen comprimidos. Este tratamiento provoca unadisminucion de la produccion hipofisaria de TSH, loque suele traducirse en la estabilizacion del tamano
de esta gl�andula. A menudo, el tratamiento no lograreducir el tamano de la alteracion, aunque impide suhipertrofia. Los sujetos que no responden a este
tratamiento habr�an de someterse a una reseccionquirurgica.
195
13
SISTEMAENDOCRINO
Figura 13.5 Hipofisis y organos diana. ACTH, hormona adrenocorticotropa; ADH, hormona antidiur�etica; FSH, hormona foliculo-
estimulante; TSH, tirotropina. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 305.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Gl�andula tiroides
La gl�andula tiroides es una gl�andula bilobulada quese localiza en el cuello en posicion anteroinferiorrespecto a la laringe (fig. 13.6). Los lobulos derechoe izquierdo se unen en la lınea media a trav�es delistmo. En ocasiones aparece un lobulo piramidal,que representa un remanente embrionario del con-ducto tirogloso, a partir del cual se forma la gl�andulaen la region posterior de la lengua del embrion. Lagl�andula tiroides se rodea de una delgada c�apsula; ensu porcion posterior se halla la gl�andula paratiroides.La c�apsula se extiende para formar tabiques en elpar�enquima glandular que la subdividen en lobulosy conducen los vasos sanguıneos, los nervios y losvasos linf�aticos; el par�enquima de la gl�andula se orga-niza en folıculos cistiformes (di�ametro� 1mm) cuyaluz contiene material coloidal rodeado de c�elulas foli-culares cubicas simples y algunas c�elulas parafolicu-lares.
Cada folıculo posee una envoltura de l�amina basalfabricada por las c�elulas foliculares (fig. 13.7).
. La union de TSH, sintetizada por la adenohipofisis,a receptores de TSH localizados en las membranasbasales y la presencia de yodo, que pasa a la c�elula atrav�es de bombas de yodo situadas en la membranabasal de las c�elulas foliculares, inducen la sıntesis delas hormonas tetrayodotironina (tiroxina T4) ytriyodotironina (T3) por parte de estas c�elulas.. La oxidacion del yodo en la zona de contacto delas c�elulas foliculares y el material coloidal poraccion de la enzimaperoxidasa tiroideaprecede ala yodacion de las hormonas.
. Los residuos de tirosina ligados a la glucoproteınasecretora tiroglobulina se yodan por adicion deuno o dos yodos oxidados, lo que da lugar atirosina monoyodada (MIT) o tirosinadiyodada (DIT).
. La combinacion de una mol�ecula MIT y unamol�ecula DIT o bien dos mol�eculas DIT originalas hormonas activas T3 y T4.
. Una vez formadas, T3 y T4 se asocian a laglucoproteına secretora tiroglobulina y seincorporan al material coloidal para sualmacenamiento.
. La exposicion a TSH estimula la liberacion de T3 yT4, que ocupan los receptores para TSH en lamembrana basal de las c�elulas foliculares. Estasc�elulas:
. Emiten filopodios que se extienden hacia elmaterial coloidal para capturar una pequenacantidad de estas hormonas en vesıculasendocıticas.
. Poseen vesıculas endocıticas rellenas de materialcoloidal que vacıan su contenido alcompartimento endosomico en el que se separanlos componentes MIT, DIT, T3 y T4 de latiroglobulina para pasar al citoplasma.
. Secretan T3, aunque su producto principal es T4,que se liberan mediante exocitosis a los lechocapilares del estroma muy vascularizado deltejido conjuntivo de la gl�andula tiroides.
. Estas hormonas se unen a proteınasplasm�aticas en el torrente circulatorio, que lasconducen hasta las c�elulas diana diseminadaspor el organismo.
. La hormona T3 presenta una afinidadmenor por lasproteınas plasm�aticas que T4 y una probabilidadm�as alta de ser captada por endocitosis por la c�eluladiana que T4.. En el citoplasma, T3 forma complejos con mayorfacilidad que T4 con la proteına del receptornuclear de hormonas tiroideas; amboscomplejos pasan al nucleo para poner en marchala transcripcion (tabla 13.4); la actividadfisiologica de T3 es m�as intensa que la de T4.
. T3 y T4 potencian las tasas metabolicas de lasc�elulas diana, incrementan la tasa de crecimientoen los sujetos jovenes, favorecen la agudezamental, estimulan el metabolismo de hidratos decarbono y lıpidos, e incrementan la frecuenciacardıaca, la frecuencia respiratoria y la accionmuscular.
. T3 y T4 reducen la sıntesis de �acidos grasos,colesterol y triglic�eridos, y facilitan eladelgazamiento.
Las c�elulas parafoliculares (c�elulas C, c�elulasclaras) presentan una coloracion p�alida al ser tenidasy se localizan en la periferia de los folıculos, aunquecomparten su l�amina basal. Estas c�elulas fabrican lahormona peptıdico calcitonina, que se secreta direc-tamente a los lechos capilares del estroma conjuntivode la gl�andula, se une a receptores de calcitonina pre-sentes en los osteoclastos e inhibe la resorcion oseapor estas c�elulas (v. tabla 13.4). Las concentracionesplasm�aticas de calcio mayores de los valores normalesinducen la liberacion de calcitonina por las c�elulasparafoliculares.
196
13
SISTEMAENDOCRINO

197
13
SISTEMAENDOCRINO
Figura 13.7 Sıntesis y yodacion de la tiroglobulina (A) y liberacion de la hormona tiroidea (B). (Tomado de Gartner LP, Hiatt JL: Color
Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 315.)
Figura 13.6 Gl�andulas tiroides y paratiroides. (Tomado
deGartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.
Philadelphia, Saunders, 2007, p 313.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Gl�andulas paratiroides
Las gl�andulas paratiroides (fig. 13.8) son cuatrogl�andulas de pequeno tamano (5�4� 2mm) quese localizan en los polos posterosuperior y posteroin-ferior de la gl�andula tiroides. Cada una de las gl�andu-las paratiroides se recubre de una c�apsula de tejidoconjuntivo que puede incluir adipocitos en el adulto.Los tabiques de tejido conjuntivo que atraviesan elpar�enquima glandular conducen a sus nervios, vasossanguıneos y vasos linf�aticos, adem�as de sustentar loscordones de c�elulas parenquimatosas y la profusa redcapilar. Las gl�andulas paratiroides sintetizan la hor-mona paratiroidea (PTH), la cual (v. tabla 13.4):
. Incrementa las concentraciones s�ericas de calcio y,de manera concertada con la calcitonina fabricadapor las c�elulas parafoliculares de la gl�andulatiroides, mantiene una concentracion optima deeste ion en el torrente circulatorio y el lıquidointersticial.
. Se une a los receptores de PTH en los osteoblastospara inducir la liberacion del factor estimulador deosteoclastos, el cual da lugar a una expansion de lapoblacion de estas c�elulas y un aumento de suactividad.
. Favorece la conservacion de calcio y la produccionde vitamina D, el cual favorece la absorcion de esteion en el tubo digestivo, por parte del rinon
El par�enquima de la gl�andula paratiroides englobados poblaciones celulares, las c�elulas principales y lasc�elulas oxıfilas.
. C�elulas principales, son unas pequenasc�elulas eosinofilas redondeadas que formanagregados celulares en el par�enquima bienirrigado de las gl�andulas paratiroides ysintetizan la hormona preproparatiroideaen el retıculo endopl�asmico rugoso. Estaprohormona se procesa en este org�anulo paraconvertirse en hormona proparatiroidea,que pasa al aparato de Golgi en el queescindir�a para dar lugar a la PTH. La hormonaempaquetada se almacena en gr�anulos secretores,de los que se liberar�a posteriormente medianteexocitosis.
. Las c�elulas oxıfilas son c�elulasm�as oscuras, grandesy abundantes que las anteriores. Se agrupan enagregados pequenos y se desconoce cu�al serıa sufuncion, aunque se ha propuesto que representarıanc�elulas principales inactivas.
CONSIDERACIONES CL�INICAS
El hiperparatiroidismo primario, un trastornoque afecta principalmente a la mujer, consiste enla produccion excesiva de PTH. En este caso, el
t�ermino primario indica que la sobreproduccion se
debe a una hiperplasia no neopl�asica de una o
m�as de las gl�andulas paratiroides. Las
concentraciones plasm�aticas excesivas de estahormona se traducen en concentraciones muyaltas de calcio y concentraciones bajas de fosfatoen sangre y lıquido intersticial, lo que origina unadisminucion de la densidad mineral; dolory fracturas oseas; debilidad muscular; parestesia;
fatiga; nefrolitiasis, n�auseas, vomitos, confusiony depresion.
El hipoparatiroidismo se atribuye a una secreciondeficiente de PTH. Suele deberse a la afectacion de
una o m�as de las gl�andulas paratiroides en eltranscurso de una intervencion quirurgica de la
gl�andula tiroides. Esta entidad se distingue por las
concentraciones s�ericas bajas de calcio, la retenciondel calcio enhuesos, y el aumentode la resorcion renalde fosfato. Se manifiesta con espasmos musculares,parestesia, insensibilidad, hormigueo, tetaniamuscular en los musculos faciales y larıngeos,
desarrollo decataratas, confusionmental y p�erdidadememoria. La supervivencia del pacientedependede laadministracion de dosis intravenosas de gluconato
c�alcico y vitamina D, ası como de calcio por vıa oral.
198
13
SISTEMAENDOCRINO

CONSIDERACIONES CL�INICAS
La enfermedad de Graves constituye la variante
m�as frecuente de hipertiroidismo, la cual se debe a la
destruccion de la gl�andula tiroides por el sistemainmunitario que provoca hiperproduccion de la
hormona tiroxina. En los casos m�as graves, sedestruyen los tejidos posteriores al globo ocular,
lo que ocasiona exoftalmia y lesiones cut�aneas enpantorrillas y la cara superior del pie. Asimismo,puede asociarse a un aumento de la tasa metabolicaque deriva en diversas complicaciones, como elincremento de la frecuencia cardıaca. Este trastorno
es m�as prevalente en mujeres mayores de 20 anos.El tratamiento no suprime las crisis, si bien puedesuscitar una mejora de la sintomatologıa y reducirla sıntesis de tiroxina.
El bocio simple es una hipertrofia de la gl�andulatiroides debida a la ingesta insuficiente de yodo.No se asocia ni a hiper-, ni a hipotiroidismo, porlo que es susceptible de tratamiento concomplementos nutricionales de yodo.
El hipotiroidismo, o hipoactividad de la gl�andulatiroides, es un trastorno caracterizado por la sıntesis
insuficiente de hormonas por esta gl�andula. Afecta,
m�as a menudo, a mujeres mayores de 50 anos. Enausencia de tratamiento, esta enfermedad trastoca elequilibrio del organismo y puede originar numerosasalteraciones, como fatiga, obesidad, artralgia,cardiopatıas, ralentizacion mental, alopecia e
insuficiencia multisist�emica. El tratamiento de
eleccion, basado en la hormona tiroidea sint�etica,es eficaz.
El mixedema constituye una forma grave dehipotiroidismo que produce diversas alteraciones,como depresion, ralentizacion mental, debilidad,bradicardia y fatiga. Otras manifestaciones sonel edema facial, la aparicion de bolsas bajo los ojos,edema sin fovea debido a la infiltracion de la matrizextracelular por abundantes glucosaminoglucanos yproteoglucanos. Los pacientes aquejados demixedema han de recibir asistencia sanitariainmediata.
El cretinismo es una variante grave dehipotiroidismo que ocurre durante el desarrollo fetalhasta la infancia provocado por la ausencia
cong�enita de la gl�andula tiroides. Los afectadospresentan retraso mental y del crecimiento.
199
13
SISTEMAENDOCRINO
Figura 13.8 Gl�andulas tiroides y paratiroides. (Tomado
deGartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.
Philadelphia, Saunders, 2007, p 313.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Gl�andulas suprarrenales
Las dos gl�andulas suprarrenales se localizan en elpolo superior del rinon y se rodean de tejido adiposobien desarrollado. Cada una de estas pequenas gl�andu-las pesa menos de 10 g y se recubre de una c�apsulaformada por tejido conjuntivo laxo cuyas proyeccionesconducen los elementos neurales y la profusa red capi-lar hacia en el seno del par�enquima glandular. Lasgl�andulas se subdividen en una corteza externa yuna m�edula interna poco profunda (fig. 13.9), cadauna de las cuales proviene de un tejido embrionariodiferente: la corteza procede del mesodermo, mientrasque la m�edula se forma a partir de la cresta neural.Cada gl�andula suprarrenal recibe su irrigacion de tresarterias: las arterias suprarrenales superior, media einferior. Estos vasos atraviesan la c�apsula y forman elplexo subcapsular, del que parten las arterias corti-cales corta y larga.
. Las arterias corticales cortas originan:. Capilares sinusoidales fenestrados, en los queel di�ametro de las fenestraciones se incrementaconforme avanzan hacia planos m�as profundosde la corteza.
. Capilares sinusoidales, drenados por v�enulaspequenas que atraviesan la m�edula y sontributarias de la vena suprarrenal.
. Las arterias corticales largas no se ramifican en lacorteza, sino que pasan a la m�edula y dan lugar a unplexo capilar drenado por v�enulas pequenas quedesembocan en la vena suprarrenal.
La corteza suprarrenal consta de tres zonas con-c�entricas que se solapan: la zona glomerular externa;la zona fasciculada, la region media m�as ancha; y lazona reticular, la m�as interna (v. fig. 13.9). Estasregiones sintetizan las hormonas basadas en el coles-terol, mineralocorticoides, glucocorticoides y andro-genos como respuesta a la union de la ACTH a susreceptores (v. tabla 13.4).
. Las c�elulas parenquimatosas de la zona glomerular,la capa m�as externa de la corteza suprarrenal,pueden contener gotıculas lipıdicas y un retıculoendopl�asmico liso muy desarrollado. Estas c�elulasproducen las hormonas aldosterona y una pequenacantidad de desoxicorticosterona al serestimuladas por la ACTH y la angiotensina II. Losmineralocorticoides intervienen en la regulaciondel equilibrio hidroelectrolıtico al actuar en lostubulos renales contorneados distales.
. La region m�as amplia de la corteza corresponde a lazona fasciculada, integrada por grandes c�elulas quese organizan en columnas longitudinales y albergan
tantas gotas lipıdicas que remedan esponjas en loscortes histologicos, por lo que reciben el nombre deespongiocitos. Su estimulacion por parte de ACTHprovoca la secrecion de los glucocorticoides cortisoly corticosterona, unas hormonas que controlan elmetabolismo lipıdico, proteico y glucıdico.Asimismo, favorecen la gluconeogenia y laglucogenogenia en el hıgado, y la lipolisisy la proteolisis en los adipocitos y las c�elulasmusculares. Las concentraciones excesivas de estashormonas ejercen una accion inmunosupresora yantiinflamatoria.
. La region m�as delgada e interna de la corteza es lazona reticular, formada por c�elulas semejantes a losespongiocitos de la zona fasciculada, si biencontienen gotıculas lipıdicas de menor tamano. Lasc�elulas parenquimatosas de esta region se disponenen forma de redes de cordones anastomosados ysintetizan androgenos, sobre todo,deshidroepiandrosterona y androstenediona;ninguna de ellas desempena acciones relevantes enlos individuos sanos.
La m�edula suprarrenal (v. fig. 13.9 y tabla 13.4)presenta un tamano relativamente pequeno y suponealrededor del 10%del peso de la gl�andula suprarrenal.Disfruta de una profusa irrigacion, ası como una iner-vacion bien desarrollada; se compone de dos tipos dec�elulas parenquimatosas: las c�elulas cromafines, m�asabundantes, y las c�elulas ganglionares simp�aticas, demayor tamano.
. Las c�elulas cromafines reciben este nombre por sumayor afinidad a las sales de cromafina, lo querefleja la abundancia de catecolaminas en sucitoplasma, especialmente de adrenalina ynoradrenalina. Se distribuyen en toda la m�edulasuprarrenal en forma de estructuras filiformes.Reciben su inervacion de neuronas simp�aticaspreganglionares. Al liberar acetilcolina, esteneurotransmisor se une a receptores de acetilcolinalocalizados en las c�elulas cromafines, lo queprovoca la despolarizacion de su membranaplasm�atica y la secrecion de noradrenalina(ante un estımulo fisiologico) o noradrenalina (anteun estımulo emocional) en los lechos capilares.. La adrenalina da lugar a sendos aumentos de latension arterial y la frecuencia cardıaca, ası comoun descenso de lamotilidad de lamusculatura lisagastrointestinal.
. La noradrenalina incrementa la tension arterial atrav�es de la contraccion del musculo liso vascular.
. Las c�elulas ganglionares simp�aticas se encuentrandispersas en la m�edula suprarrenal y carecen dedendritas y axones.
200
13
SISTEMAENDOCRINO

CONSIDERACIONES CL�INICAS
El sındrome de Cushing (hiperadrenocorticismo)se debe a la presencia de adenomas en laadenohipofisis, los cuales potencian la produccionde ACTH. Las concentraciones altas de esta
hormona estimulan el crecimiento de la gl�andulasuprarrenal, la hipertrofia de la corteza suprarrenal yla produccion excesiva de cortisol. Los pacientespresentan obesidad, en particular en el rostro, el
cuello y el tronco, adem�as de caquexia y
osteoporosis. Los hombres se tornan est�eriles,mientras que las mujeres presentan amenorrea.
La enfermedad de Addison es un tipo deinsuficiencia corticosuprarrenal debida a ladestruccion de la corteza suprarrenal. Suele serconsecuencia de un trastorno autoinmunitario,
aunque tambi�en puede derivar de la tuberculosis yotras enfermedades infecciosas. Se manifiesta a lolargo de varios meses con fatiga, debilidad
muscular, hipotension arterial, n�auseas, vomitos,artralgias, hipoglucemia, adelgazamiento ydepresion. Se trata mediante hormonoterapia desustitucion.
201
13
SISTEMAENDOCRINO
Figura 13.9 Gl�andula suprarrenal y tipos celulares. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia,Saunders, 2007, p 319.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Gl�andula pineal (cuerpo pineal)
La gl�andula pineal, una evaginacion del techo deldienc�efalo (v. tabla 13.4) es una pequena gl�andulaendocrina que pesa menos de 150mg. Est�a recubiertapor la piamadre, la cual forma una c�apsula que emitetabiques dotados de vasos sanguıneos hacia el par�en-quima glandular que lo subdividen en lobulillosincompletos. El par�enquima de esta gl�andula se com-pone de dos tipos celulares: los pinealocitos y lasc�elulas intersticiales.
. Los pinealocitos, las c�elulas m�as destacadas de lagl�andula pineal, poseen una o dos prolongacionestortuosas cuyos extremos se aplanan y dilatanconforme se acercan a los capilares. Estas c�elulasposeen un citoesqueleto bien desarrollado yestructuras tubulares especializadas, los listonessin�apticos, cuya poblacion se amplıa durante elsegmento oscuro del ciclo diurno. Las fibrassimp�aticas posganglionares sinaptan con lospinealocitos, los cuales liberan melatonina a lolargo del perıodo nocturno, no ası en el diurno, paraestablecer el ritmo diurno del organismo. Estasc�elulas controlan algunas funciones corporales alinhibir la liberacion de la hormona del crecimiento ylas gonadotropinas. Las concentraciones s�ericas demelatonina sonm�as altas antes del reposo nocturno.
. Las c�elulas intersticiales similares a la glıa son m�asabundantes en el tallo pineal que en la masa de lagl�andula. Se tinen de color oscuro y presentanproyecciones celulares alargadas con filamentosintermedios, microfilamentos y microtubulos.Junto al tejido conjuntivo, estas c�elulas sustentanlos pinealocitos.
. La gl�andula pineal contiene estructuras calcificadasllamadas cuerpos aren�aceos (arena cerebral) defuncion y origen desconocidos. La calcificacioncomienza en una etapa temprana de la ninez yavanza a lo largo de la vida.
CONSIDERACIONES CL�INICAS
La melatonina puede conferir cierta proteccion alsistema nervioso central al reclutar y eliminar
radicales libres generados por estr�es oxidativo.Algunos sujetos ingieren complementos demelatonina como tratamiento de los trastornos del
estado de �animo y el sueno, ası como la depresion.Se ha descrito que la exposicion a luz artificialintensa podrıa derivar en una disminucion de lasıntesis de melatonina, lo que potenciarıa ladepresion. Por otra parte, muchas personasrefieren que el consumode dosis demelatonina enel momento adecuado puede reducir el jet lag.
202
13
SISTEMAENDOCRINO

203
13
SISTEMAENDOCRINO
Tabla 13.4 HORMONAS Y FUNCIONES DE LAS GLANDULAS TIROIDES, PARATIROIDES, SUPRARRENAL Y PINEAL
Hormona C�elula productora Hormona reguladora Funcion
Gl�andula tiroides
Tiroxina (T4) y
triyodotiroxina (T3)
C�elulas foliculares TSH Facilita la transcripcion nuclear de genes
implicados en la sıntesis de proteınas;
incrementa el metabolismo celular y la
tasa de proliferacion celular;incrementa la actividad de las
gl�andulas endocrinas; estimula el
metabolismo de carbohidratos y
lıpidos; reduce las concentraciones decolesterol, fosfolıpidos y triglic�eridos;
incrementa las concentraciones de�acidos grasos; reduce el peso corporal;incrementa la frecuencias cardıaca y
respiratoria y la accion muscular
Calcitonina (tirocalcitonina) C�elulas parafoliculares Mecanismo de
retroalimentacion conhormona paratifoidea
Reduce las concentraciones plasm�aticas
de calcio mediante la supresion de laresorcion osea
Gl�andula paratiroides
Hormona paratiroidea
(PTH)
C�elulas principales Mecanismo de
retroalimentacion concalcitonina
Incrementa las concentraciones de calcio
en los lıquidos corporales
Gl�andulas suprarrenales y corteza suprarrenal
Mineralocorticoides:aldosterona y
desoxicorticosterona
C�elulas de la zonaglomerular
Angiotensina II y ACTH Control del volumendel lıquido corporaly las concentraciones de electrolitos a
trav�es de los tubulos renales distales al
inducir la excrecion de potasio y la
resorcion de sodioGlucocorticoides: cortisol
y corticosterona
C�elulas de la zona
fasciculada
(espongiocitos)
ACTH Regulacion del metabolismo de hidratos
de carbono, lıpidos y proteınas;
disminucion de sıntesis de proteınas,
aumento de concentraciones s�ericas deamino�acidos; estimulacion de
gluconeogenia mediante la activacion
de la transformacion de amino�acidos
en glucosa por parte del hıgado;liberacion de �acidos grasos y glicerol;
accion antiinflamatoria; reduccion de
permeabilidad capilar; inhibicion derespuesta inmunitaria
Androgenos:
deshidroepiandrosterona
y androstenediona
C�elulas de la zona
reticular
ACTH Induce caracterısticas masculinas d�ebiles
M�edula suprarrenal
Catecolaminas: adrenalinay noradrenalina
C�elulas cromafines Nerviospreganglionares,
simp�aticos y
espl�acnicos
Adrenalina: utiliza un mecanismo de«lucha o huıda» que prepara al
organismo para condiciones de gran
estr�es o miedo; incrementa lafrecuencia cardıaca y el gasto cardıaco,
la irrigacion org�anica y la liberacion de
glucosa por el hıgado para su
utilizacion como fuente de energıaNoradrenalina: provoca un aumento
de la presion arterial debido a
vasoconstriccion
Gl�andula pineal
Melatonina Pinealocitos Noradrenalina Puede incidir en la actividad cıclica de las
gonadas
Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd Ed. Filadelfia, Saunders, p 312
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

14 SISTEMATEGUMENTARIO
El sistema tegumentario, el organo m�as grande ypesado del organismo (supone aproximadamente el15% del peso corporal total y posee un �area de super-ficie media de unos 2 m2), est�a for-mado por la piel, el pelo, las gl�andu-las seb�aceas, las unas y las gl�andulassudorıparas. Recubre la totalidad delorganismo y sigue por la mucosa quereviste los orificios externos de losaparatos gastrointestinal, respira-torio y genitourinario. La piel tapizael conducto auditivo externo, recu-bre la cavidad timp�anica y sigue porla conjuntiva del ojo en el p�arpado.
Piel
La piel consta de dos capas: un epitelio escamosoestratificado queratinizado externo, llamado epider-mis, que recubre una capa de tejido conjuntivodenominada dermis (fig. 14.1). La membrana basalsepara la epidermis de la dermis. La zona de contactono es regular, ya que la dermis forma unas elevacionessimilares a conos y rebordes: los rebordes d�ermicos(papilas d�ermicas). Los rebordes d�ermicos encajanperfectamente con las invaginaciones de la epidermis:los rebordes epid�ermicos (papilas epid�ermicas. Elconjunto de los rebordes epid�ermicos y d�ermicosrecibe el nombre de aparato reticular. Por debajo dela dermis aparece una fascia, la hipodermis (fasciasuperficial), que puede contener un volumen impor-tante de tejido adiposo en personas con sobrepeso; nose considera que esta capa forme parte de la piel.
La piel desempena un gran numero de funciones,entre las que destacan:
. Formacion de una cubierta flexible para elorganismo.
. Proteccion frente a lesiones por impacto y abrasion,invasion de bacterias y deshidratacion.
. Absorcion de radiacion ultravioleta (UV) necesariapara la sıntesis de vitamina D.
. Recepcion de informacion del medio circundante(p. ej., contacto, dolor, temperatura).
. Regulacion de la temperatura.
. Excrecion de sudor.
. Sıntesis de melanina (proteccion de las capasprofundas frente a exposicion excesiva a radiacionUV).
La presencia de rebordes y surcos en forma deasas, remolinos y arcos –dermatoglifos (huellasdactilares)– que se alternan en las yemas de los
dedos de las manos y los pies creauna superficie menos resbaladiza paraasir objetos de pequeno tamano y per-mite captar datos sensoriales que con-ducen a la identificacion de los objetosmanipulados.
EPIDERMIS
La epidermis, la capa m�as externa de lapiel, carece de irrigacion y recibe susnutrientes por difusion desde los capi-lares d�ermicos. La epidermis se com-
pone de un epitelio escamoso estratificado queratini-zado con un espesor medio inferior a 0,1mm, si bienalcanza un grosor de 1mm en la palma de la mano yde 1,4mm en la planta del pie. Se distinguen dos tiposde piel (tabla 14.1; v. fig. 14.1):
. La piel gruesa, presente en la palma de la mano y laplanta del pie, es lampina, carece de musculoserectores del pelo y gl�andulas seb�aceas, aunque est�adotada de gl�andulas sudorıparas.
. La piel delgada, presente en el resto del organismo,posee folıculos pilosos, musculos erectores del pelo,gl�andulas seb�aceas y gl�andulas sudorıparas.
La epidermis est�a formada por cuatro tipos celu-lares diferentes: los queratinocitos, las c�elulas deLangerhans, losmelanocitos y las c�elulas de Merkel,entre los que predominan los queratinocitos, de ori-gen ectod�ermico. Los otros tres tipos se distribuyenentre estos.
Las c�elulas de la superficie epitelial se descaman,por lo que los queratinocitos de las capas m�as profun-das se dividen para sustituirlas. Se cree que el factor decrecimiento epid�ermico y la interleucina-1a inducenla actividad mitotica de los queratinocitos, mientrasque el factor transformador del crecimiento la inhi-birıa. La division celular solamente tiene lugar duranteel perıodo nocturno y las c�elulas reci�en formadasempujan hacia la superficie a las situadas por encimade ellas, que terminan por descamarse. Cada una deestas nuevas c�elulas precisa de unmes para situarse enla capa superficial y desprenderse. Los queratinocitos
TERMINOS CLAVE. Piel. Queratinocitos. No queratinocitos
de la epidermis. Dermis. Gl�andulas de la piel. Pelo. Unas
� 2011 Elsevier Espana, S.L. Reservados todos los derechos204

sufren un proceso de citomorfosis, que permite dis-tinguir cinco capas en la epidermis, conforme migranhacia la superficie libre. En la piel delgada tan solo seaprecian tres de las cinco capas, mientras que la pielgruesa contiene estas cinco capas.
205
14
SISTEMATEG
UMENTARIO
Tabla 14.1 CARACTERISTICAS DE LA PIEL GRUESA Y DELGADA
Tipo de piel Ejemplos Espesor (mm) Estratos Ap�endices
Piel gruesa Palmas de manos,
plantas de pies
400–600 Los cinco estratos Ausencia de folıculos pilosos,
musculos erectores del pelo
y gl�andulas seb�aceas
Piel delgada Resto del organismo 75–150 Carente de estratoslucido y granuloso
definidos
Folıculos pilosos, musculoserectores del pelo y gl�andulas
seb�aceas y sudorıparas
Figura 14.1 Comparacion de la piel gruesa y delgada. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia,
Saunders, 2007, p 328.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Capas de la epidermis
Las cinco capas que integran la epidermis de la pielgruesa son el estrato basal (estrato germinativo), quese encuentra en contacto directo con la membranabasal; el estrato espinoso; el estrato granuloso; elestrato lucido; y el estrato corneo (tabla 14.2 yfig. 14.2). Los queratinocitos de las cinco capas seadhieren a las c�elulas adyacentes a trav�es de desmo-somas. En la piel delgada aparecen c�elulas aisladas delos estratos granuloso y lucido, si bien las c�elulas noforman capas definidas similares a las existentes en lapiel gruesa. La piel delgada unicamente se componede tres de los cinco estratos citados.
. El estrato basal (estrato germinativo) se componede una monocapa de c�elulas cubicas a cilındricasbajas en contacto con la membrana basal. Estasc�elulas sufren divisiones celulares y las c�elulas reci�enformadas empujan a las anteriores situadas porencima de ellas hacia la superficie libre. Las c�elulasdel estrato basal se unen a la l�amina basal mediantehemidesmosomas, mientras que se adhieren a lasc�elulas adyacentes a trav�es de desmosomas. Lasplacas desmosomicas y hemidesmosomicas seasocian a haces de filamentos intermedios(tonofilamentos). El citoplasma contiene escasosorg�anulos, aunque son abundantes los ribosomas.
. El estrato espinoso es una capa gruesa formada porvarias capas de c�elulas de morfologıa poli�edrica enla proximidad del estrato basal se aplanan conformese alejan de la membrana basal. Las c�elulaspoli�edricas presentan actividad mitotica, mientrasque lasm�as superficiales no pueden ya dividirse. Losorg�anulos de estas c�elulas son semejantes a los de lasque integran el estrato basal, si bien lostonofilamentos est�an mejor desarrollados, enparticular en las c�elulas planas m�as superficiales, yoriginan unos haces m�as gruesos llamadostonofibrillas. En esta region, las c�elulas planasalbergan unos gr�anulos secretores que reciben el
nombre de gr�anulos de recubrimiento demembrana (gr�anulos laminares), los cualespresentan undi�ametro inferior a 0,5mmy contienendepositos lipıdicos laminares. Las proyeccionescitopl�asmicas de estas c�elulas remedan espinas,de donde proviene la denominacion de esta capa.Las espinas de las c�elulas adyacentes se interdigitany forman desmosomas para adherirse entre sıy a las c�elulas de los estratos basal y granuloso.
. Las c�elulas del estrato granuloso contienengr�anulos de recubrimiento de membrana ydepositos de queratohialina que no se unen a lamembrana y en los que se insertan los haces detonofilamentos. Los gr�anulos de recubrimientovacıan su carga en el espacio extracelular localizadosobre el estrato espinoso, de modo que se crea unabarrera lipıdica que separa los estratos granuloso ylucido e impide el movimiento de lıquido entreambas capas. La presencia de esta barrera confiereimpermeabilidad frente al agua a la epidermis, loque evita su p�erdida por la dermis suprayacente y supaso hacia esta capa desde el exterior del organismo.
. El estrato lucido corresponde a una capatransparente de c�elulas cuyos org�anulos, incluido sunucleo, han sido eliminados por accion de loslisosomas. Estas c�elulas muertas contienennumerosos tonofilamentos rodeados de eleidina,un derivado de queratohialina. La proteınainvolucrina, de funcion desconocida, abunda en lasuperficie citopl�asmica de las membranasplasm�aticas de estas c�elulas.
. El estrato corneo, la capa m�as superficial, suele serla m�as gruesa de la epidermis de la piel gruesa. Lasmembranas plasm�aticas de estas c�elulas muertas,llamadas escamas, aparecen engrosadas y serellenan de filamentos de queratina. Las c�elulas delas capas m�as superficiales del estrato corneo sonincapaces de mantener el contacto con las c�elulasvecinas a trav�es de los desmosomas y terminan pordesprenderse.
206
14
SISTEMATEG
UMENTARIO

CONSIDERACIONES CL�INICAS
La psoriasis es una enfermedad autoinmunitariacronica no contagiosa que afecta a la piel y lasarticulaciones. Se caracteriza por la presencia delesiones parcheadas, en especial alrededor de lasarticulaciones, que reciben el nombre de placas
psori�asicas, debidas a la acumulacion de c�elulas enel estrato corneo debido a un aumento del numero
de c�elulas proliferativas en el estrato basal.A menudo, las placas se localizan en la piel de codosy rodillas, si bien pueden encontrarse en otrasregiones, como el cuero cabelludo y los genitales;pueden aparecer, incluso, en las unas. La psoriasis
puede originar inflamacion articular, lo que recibe el
nombre de artritis psori�asica. Entre un 10 y un 15%de las personas con psoriasis presenta artritis
psori�asica.
La epidermolisis ampollosa, un trastorno gen�etico,se distingue por la formacion de ampollas en la piel araız de un traumatismo menor. Se debe a laexistencia de anomalıas en los filamentosintermedios de los queratinocitos que losdesestabilizan, ası comoa alteraciones en las fibrillasde anclaje de dermis y epidermis.
207
14
SISTEMATEG
UMENTARIO
Tabla 14.2 ESTRATOS Y CARACTERISTICAS HISTOLOGICAS DE LA PIEL GRUESA
Capa Caracterısticas histologicas
Epidermis Derivada del ectodermo; formada por epitelio escamosco estratificado queratinizado(queratinocitos)
Estrato corneo Muchas capas de c�elulas planas queratinizadas muertas, queratinocitos, sin nucleo ni org�anulos
(escamas o c�elulas corneas) que se desprenden
Estrato lucido* Delgada capa p�alida de queratinocitos sin nucleo ni org�anulos; contienen haces densosde filamentos de queratina y eleidina
Estrato granuloso* Formado por tres a cinco capas. Los queratinocitos son aun nucleados; contienen gr�anulos grandes
y gruesos de queratohialina y gr�anulos de recubrimiento de la membrana
Estrato espinoso Capa m�as gruesa de la epidermis; los queratinocitos, o c�elulas espinosas, se interdigital entre sıy forman puentes intercelulares y abundantes desmosomas; las c�elulas espinosas poseen
numerosos tonofilamentos y gr�anulos de recubrimiento de lamembrana y se dividen demanera
activa; contienen, adem�as, c�elulas de Langerhans.Estrato basal (germinativo) Monocapa de c�elulas cubicas a cilındricas bajas enmitosis activa; se separan de la capa papilar de la
dermis por una membrana basal bien desarrollada; esta capa contiene, asimismo, c�elulas de
Merkel y melanocitos.
Dermis Derivada del mesodermo; formada principalmente por fibras de col�ageno de tipo I y fibrasel�asticas; se subdivide en dos regiones: capa papilar y capa reticular, una capa de tejido
conjuntivo colagenoso irregular denso
Capa papilar Se interdigita con la epidermis y constituye el componente de papilas d�ermicas del aparato
reticular; contiene fibras de col�ageno de tipo III y fibras el�asticas en una organizacion laxa, asıcomo fibrillas de anclaje (col�ageno de tipo VII); los lechos capilares profusos, las c�elulas de
tejido conjuntivo y losmecanorreceptores se localizan en esta capa; en algunas ocasiones, puede
contener melanocitos.Capa reticular Capa m�as profunda de la piel; fibras de col�ageno de tipo I, fibras el�asticas y c�elulas del tejido
conjuntivo; contiene gl�andulas sudorıparas y sus conductos, folıculos pilosos y musculos
erectores del pelo, ası como gl�andulas seb�aceas y mecanorreceptores (p. ej., corpusculos de
Pacini)
*Presentes solamente en la piel gruesa. Todas estas capas suelen presentar un menor espesor en la piel delgada.Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 329.
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

No queratinocitos en la epidermis
Se distinguen tres grupos de no queratinocitos en laepidermis (v. fig. 14.2):
. Las c�elulas de Langerhans, c�elulas presentadoras deantıgenos procedentes de la m�edula osea, sedistribuyen en el estrato espinoso; pueden presentaruna densidad de 800 c�elulas por milımetrocuadrado. Sus nucleos y sus citoplasmas sonnormales, con excepcion de la presencia de losgr�anulos de Birbeck citopl�asmicos (gr�anulosvermiformes), los cuales remedan palas de tenisde mesa y se desconoce su funcion. Las c�elulas sonp�alidas en la microscopia optica y se diferenciande los queratinocitos circundantes por la ausenciade tonofilamentos. De manera similar a otrasc�elulas presentadoras de antıgenos, las c�elulas deLangerhans poseen receptores de Fc y C3, puedenfagocitar antıgenos, forman complejos epıtopo-MHC y migran hacia ganglios linf�aticos proximos,en los que presentan estos complejos a loslinfocitos T.
. Las c�elulas de Merkel, derivadas de la cresta neural,son unas c�elulas p�alidas situadas en el estrato basal,especialmente en la mucosa bucal, los folıculospilosos y las yemas de los dedos de las manos.Poseen un nucleo con surcos profundos y uncitoplasma con abundantes citoqueratinas; seconectan entre sı por fibras sensorialesmielinizadas, lo que da lugar a asociaciones c�elulasde Merkel-neuritas. Las c�elulas de Merkel actuancomo neurorreceptores de contacto suave.
. Al igual que las anteriores, los melanocitosproceden de la cresta neural y se localizan en elestrato basal; estas c�elulas emiten unasprolongaciones delgadas y largas que viajan hacia elestrato espinoso, en el que sus extremos se rodeande extensiones citopl�asmicas de los queratinocitos.Los melanocitos contienen unos gr�anulos ovalados(salvo en las personas pelirrojas, en las que sonredondeados) cargados con la enzima tirosinasaque reciben el nombre de melanosomas. Latirosinasa transforma la tirosina en un pigmentooscuro, llamado melanina, en el seno de estasvesıculas. Los melanosomas migran hacia elextremo de las prolongaciones celulares y acumulangrandes cantidades de este pigmento por medio deun proceso estimulado por la radiacion UV. Losqueratinocitos seccionan los extremos de dichasprolongaciones, lo que recibe el nombre desecrecion citocrina, de modo que las enzimaslisosomicas proceden a atacar los melanosomas delinterior de los queratinocitos del estrato espinoso,cuya degradacion concluir�a al cabo de unos dıas.Hasta entonces, la melanina conferir�a proteccion alos queratinocitos frente a la radiacion UV. Ladensidad poblacional de los melanocitos difiere endistintas regiones de un mismo individuo, aunquesus valores son pr�acticamente los mismos en cadaraza. Las diferencias de color de la piel no obedecena la mayor densidad de poblacion de losmelanocitos, sino a la sıntesis de cantidadesmayores de melanina y la degradacion m�as lentade esta mol�ecula.
208
14
SISTEMATEG
UMENTARIO

CONSIDERACIONES CL�INICAS
La actividad de la tirosinasa aumenta en presencia deradiacion UV, lo que incrementa las concentracionesde melanina. Asimismo, la melanina se oscurece enpresencia de esta radiacion. Por otra parte, lahormona adrenocorticotropa, sintetizada por lahipofisis, influye en la pigmentacion. En algunoscasos, como en los pacientes aquejados de laenfermedaddeAddison, la produccion decortisol esinsuficiente, lo que da lugar a unas concentracionesexcesivas de hormona adrenocorticotropa que setraducen en hiperpigmentacion.
El vitıligo es un trastorno en el que algunas regionesde la piel (a menudo, la cara y las manos) carecen depigmentacion. La destruccion de los melanocitostıpica de esta enfermedad autoinmunitaria origina�areas exentas de pigmentacion con queratinocitosintactos. Esta entidad suele asociarse a otrostrastornos autoinmunitarios.
El albinismo es una anomalıa cong�enita debida aausencia total de sıntesis de melanina. Los sujetos
albinos poseen melanosomas, pero no fabricantirosinasa, por lo que carecen de melanina.
Los nevos son depositos benignos de melanocitosen la epidermis. Su tamano comprende de puntos
pequenos hasta acumulaciones con un di�ametromayor de 3,8 cm. Pueden ser planos o elevados,lisos o rugosos (verrugosos) y presentar folıculospilosos. Por lo general presentan una coloracionmarron o negra, aunque pueden mostrar un colorrosado.
Los rayos UV son de dos tipos. Los rayos UVBproducen quemaduras, mientras que los UVAoriginan el bronceado de la piel. Se ha demostradoque la radiacion UV es un elemento clave en elfotoenvejecimiento y el desarrollo de carcinoma de
c�elulas basales y melanoma en una etapa posteriorde la vida.
209
14
SISTEMATEG
UMENTARIO
Figura 14.2 Melanocitos y sus funciones. RER, retıculo endopl�asmico rugoso. (Tomado de Gartner LP, Hiatt JL: Color Textbook ofHistology, 3rd ed. Philadelphia, Saunders, 2007, p 334.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

DERMIS
La dermis, la capa de tejido conjuntivo situada pordebajo de la epidermis, consta de dos regiones: la capapapilar superficial y la capa reticular m�as profunda.Ambas capas se componen de tejido conjuntivofibroel�astico irregular denso. La capa papilar es laxay alberga haces delgados de fibras de col�ageno detipo I, mientras que la reticular es notablemente m�asdensa y contiene haces gruesos de estas fibras. Pordebajo de la dermis aparece la hipodermis, una fasciasuperficial de anatomıa macroscopica que no perte-nece a la piel y suele incluir una capa variable de tejidoadiposo, el panıculo adiposo, cuyo espesor alcanzavarios centımetros en sujetos obesos. La dermis es unacapa delgada en algunas regiones del organismo,como los p�arpados, en los que presenta un espesorde 0,6mm, mientras que en otras zonas, como laplanta del pie, puede tener 3mm.
. La capa papilar est�a en contacto con la membranabasal y forma unas evaginaciones denominadasrebordes d�ermicos (papilas d�ermicas) que seinterdigitan con los rebordes epid�ermicos. Lasfibras de este tejido conjuntivo laxo (fibras decol�ageno de tipo III y fibras el�asticas delgadas)aparecen entretejidas. Asimismo, las fibras deanclaje, formadas por fibras de col�ageno detipo VII, se unen a las fibras reticulares parareforzar la union de la membrana basal a la capabasal, de tal modo que la epidermis se fija a ladermis. La capa papilar contiene c�elulas del tejidoconjuntivo propiamente dicho, aunque tambi�en
aparecen asas capilares que aportan nutrientes a laepidermis avascular y participan en la regulacionde la temperatura corporal. Asimismo, albergaterminaciones nerviosas encapsuladas, comocorpusculos de Meissner de funcionmecanorreceptora y bulbos terminales deKrause, de funcion posiblemente termorreceptora.Las terminaciones nerviosas desnudas atraviesanla capa papilar en su recorrido hacia laepidermis, en la que actuan como receptoresdel dolor.
. El tejido conjuntivo de la capa reticular es m�asdenso que el que forma la capa papilar, y susfibras corresponden mayoritariamente a hacesgruesos de col�ageno de tipo I entremezcladascon fibras el�asticas gruesas incluidas en unamatriz de sustancia fundamental en la queabunda el dermatano sulfato. Los componentescelulares de esta capa son semejantes a los dela capa papilar, si bien son menos abundantes.Las regiones profundas de las gl�andulassudorıparas, las gl�andulas seb�aceas y losfolıculos pilosos, junto a los musculos erectoresasociados, se hallan en la dermis, la cualcontiene tambi�en un plexo profuso de vasossanguıneos y linf�aticos que se ramifican envasos de menor calibre encargados de irrigar lacapa capilar. Los elementos neuralesencapsulados, como los corpusculos dePacini y los corpusculos de Ruffini,reaccionan ante las fuerzas intensas de presiony de tension.
210
14
SISTEMATEG
UMENTARIO

CONSIDERACIONES CL�INICAS
Los tres tipos de tumores malignos de la piel son el
carcinoma de c�elulas basales, el carcinomaepidermoide y el melanoma maligno. El carcinoma
de c�elulas basales, la neoplasia maligna m�asfrecuente en el ser humano, afecta,aproximadamente, a un millon de estadounidensescada ano. Casi todos los tumores se localizan en�areas del organismo sometidas a una exposicionexcesiva al sol, en especial, la cara, las orejas, elcuello, el cuero cabelludo, los hombros y la espalda.Los sujetos de riesgo presentan piel y cabello claro.Suele afectar a sujetos mayores, si bien a lo largo delos ultimos anos se ha observado un aumento de sufrecuencia en individuos jovenes. Los sujetos quetrabajan o realizan actividades de ocio al aire librepresentan una susceptibilidad mayor. El carcinoma
de c�elulas basales surge de las c�elulas del estratobasal. En el lugar afectado se forma una lesion, lacual puede remedar psoriasis o eczema o bienparecer una llaga de pequeno tamano (p. ej., en la
cara) que sangra y no cicatriza. Un m�edico conexperiencia puede elaborar este diagnostico, quedebe confirmarse con una biopsia. El tratamientohabitual consiste en la reseccion quirurgica. Los
carcinomas de c�elulas basales no suelen formar
met�astasis, aunque la enfermedad puede recidivaren los sujetos afectados.
El carcinoma epidermoide representa la segunda
neoplasia cut�anea m�as prevalente. En EE. UU. se
diagnostican anualmente m�as de 250.000 casosnuevos. Las personas de edad mediana yavanzada, con tez clara y expuestas a radiacion
solar durante un perıodo prolongado presentan unriesgo mayor de afectacion por esta entidad. Sealteran los queratinocitos de la piel; las lesiones
cut�aneas son placas con costra o escamosas conuna base inflamada enrojecida o una ulcera que no
cicatriza. Normalmente se localizan en �areasexpuestas a la radiacion solar, si bien pueden
aparecer tambi�en en los labios, la cavidad bucal,los genitales o cualquier otra region del organismo.Se debe efectuar una valoracion dermatologica decualquier lesion que se hipertrofie, sangre, cambiesu aspecto o no remita. El diagnostico precoz y eltratamiento temprano son de gran importancia, yaque las lesiones pueden extenderse y formar
met�astasis. La reseccion quirurgica suele ser eltratamiento de eleccion.
El melanoma maligno es un tumor maligno muygrave que surge de losmelanocitos. Losmelanocitostransformados proliferan, invaden la dermis, pasan a
los vasos sanguıneos y linf�aticos, y forman
met�astasis a distancia. Esta entidad es m�asprevalente en individuos de tez clara, en particular aaquellos con una exposicion intensa a radiacion UV.Los datos disponibles indican que la radiacion UVutilizada en las cabinas de bronceado podrıa originarmelanoma. La susceptibilidad a la enfermedadpuede ser de tipo hereditario. El melanoma malignoes susceptible de curacion en los pacientes con undiagnostico precoz, aunque puede tenerconsecuencias mortales en caso de diseminarse. Eltratamiento habitual suele ser la reseccion quirurgicaen pacientes con un diagnostico precoz.
211
14
SISTEMATEG
UMENTARIO
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Gl�andulas de la piel
En la piel se distinguen cuatro tipos de gl�andulas(fig. 14.3), si bien en este capıtulo se abordar�an tresde ellos: las gl�andulas sudorıparas ecrinas, las gl�andu-las sudorıparas apocrinas y las gl�andulas seb�aceas. Elcuarto tipo, la gl�andula mamaria, es una gl�andulasudorıparamuy especializada que se tratar�a en el capı-tulo 20, dedicado al aparato reproductor femenino.
. En la mayor parte de la piel que recubre el cuerpo sedistribuyen alrededor de cuatro millones degl�andulas sudorıparas ecrinas. Cada una deestas gl�andulas tubulares espirales sencillasrepresenta una evaginacion ectod�ermica (rodeadade una membrana basal) que atraviesa la epidermisy la dermis y, a menudo, penetra en la hipodermis.En esta fascia se forma la porcion secretoramerocrinamuy enroscada de la gl�andula, de la queparte un conducto estrecho similar a unsacacorchos que perfora el extremo o la cresta de unreborde d�ermico y desemboca en la superficie librede la epidermis a trav�es de un poro sudorıparo.
. El epitelio cubico a cilındrico bajo sencillo de laporcion secretora se compone de c�elulas oscuras yc�elulas claras. Las c�elulas de esta porcion se rodeande c�elulas mioepiteliales, que contienenabundantes filamentos de actina y miosina, quefavorecen la salida del sudor de su luz.. En la microscopia electronica, las c�elulas oscuras(c�elulas mucoides) presentan una morfologıapiramidal cuya base corresponde a la luz y cuyov�ertice puede alcanzar, o no, la l�amina basal. Estasc�elulas fabrican y secretan un producto mucoide.
. Las c�elulas claras presentan una forma similar alas anteriores, aunque sus bases est�an en contactocon la l�amina basal y sus v�ertices rara vez alcanzanla luz. Estas c�elulas presentan grandes cantidadesde glucogeno y una membrana plasm�atica connumerosos repliegues en las im�agenes demicroscopia electronica que reflejan suparticipacion en el transporte epitelial. Fabricanuna secrecion serosa que liberan a su luz.
El epitelio cubico estratificado del conducto de unagl�andula sudorıpara ecrina se compone de una capabasal, la cual alberga un gran numero de mitocon-
drias, y una capa luminal, caracterizada por un cito-plasma escaso y un nucleo de morfologıa irregular.
El sudor sintetizado por la porcion secretora essimilar al suero desde el punto de vista osmotico, sibien las c�elulas del conducto conservan sodio, cloro ypotasio y excretan �acido l�actico, urea y material inge-rido, como algunos f�armacos y la esencia de ajo.
. Las gl�andulas sudorıparas apocrinas sonsemejantes a las ecrinas, aunque su tamano esnotablemente mayor; se localizan en la axila, laareola del pezon y el �area perianal. A pesar de sunombre, la modalidad de secrecion de la mayorıade ellas podrıa ser de tipo merocrino. Comienzana funcionar despu�es de la pubertad, se asocian alos canales de los folıculos pilosos, en los quevacıan sus productos, y sufren alteracionesperiodicas relacionadas con el ciclo menstrual enalgunas mujeres. Aunque sus productos soninodoros, producen olor al ser metabolizadospor la microflora en �acido 3-metil-1,2-hex�anico,el cual podrıa poseer propiedades feromonicas.Las gl�andulas ceruminosas del conductoauditivo externo y las gl�andulas de Moll en losp�arpados son gl�andulas sudorıparas apocrinasmodificadas.
. Las gl�andulas seb�aceas son gl�andulas holocrinasque se asocian a los folıculos pilosos. Desembocanen los canales de los folıculos, donde vierten suproducto secretor, el sebo oleoso; aparecenexclusivamente en la piel glabra (con vello). Lasc�elulas m�as perif�ericas de estas gl�andulas globularesson planas; est�an en contacto con la membranabasal y sufren divisiones celulares para originartanto c�elulas planas como unas c�elulas redondeadasde mayor tamano. Estas ultimas adoptan unaposicion central y acumulan gotıculas lipıdicas quedesplazan, en ultima instancia, los org�anuloscelulares, lo que provoca su degeneracion, necrosisy transformacion en el sebo que recubre el tallo delpelo y la superficie cut�anea. El sebo potencia laflexibilidad del pelo y la textura de la piel. Al igualque las gl�andulas sudorıparas apocrinas, lasgl�andulas seb�aceas est�an sometidas a un controlhormonal y su actividad se intensifica conposterioridad a la pubertad.
212
14
SISTEMATEG
UMENTARIO

213
14
SISTEMATEG
UMENTARIO
Figura 14.3 Gl�andula ecrina y gl�andula seb�acea y sus componentes celulares. (TomadodeGartner LP,Hiatt JL: Color Textbook ofHistology,
3rd ed. Philadelphia, Saunders, 2007, p 337.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Pelo
La superficie de la piel se recubre de pelo (fig. 14.4),un filamento queratınico cuya composicion amino-acıdica determina si ser�a suave y blando o bien rugosoy rizado. En el ser humano se distinguen tres tipos depelo: el lanugo, exclusivo del feto y el neonato; elvello, un pelo fino, blando y corto, como el que recu-bre los p�arpados; y el pelo terminal, el pelo gruesoduro y oscuro que aparece en el cuero cabelludo y lascejas y el rostro en el hombre. Aparentemente, el peloes menos abundante en el ser humano que en otrosprimates; sin embargo, la mayor parte del pelo del serhumano corresponde a pelo velloso, mientras que enprimates lo hace a pelo terminal. El numero de pelospor centımetro cuadrado es equivalente en el serhumano y otros primates.
El pelo se forma en los folıculos pilosos, unasinvaginaciones epid�ermicas que suelen invadir lahipodermis. Se rodean de una membrana basal,denominada membrana vıtrea, que se encuentrainmersa en una membrana conjuntiva derivada de ladermis. El folıculo piloso consta de:
. La raız del pelo, una terminacion expandida huecaen cuya concavidad se localizan elementos de tejidoconjuntivo vascularizado que conforman lallamada papila d�ermica; el conjunto de la raız y lapapila se conoce como el bulbo piloso.
. El nucleo de la raız del pelo est�a formado por unasc�elulas denominadas matriz; el crecimiento delpelo se sustenta en la actividad mitotica de estapoblacion celular.
. Inmediatamente por debajo de la membrana vıtrease encuentra una monocapa de c�elulas en el bulbopiloso cuyo numero aumenta en la proximidad delestrato corneo; esta capa celular recibe el nombre devaina externa de la raız.
. La vaina interna de la raız, envuelta por la vainaexterna, se compone de tres capas celulares: la capade Henley, la capa de Huxley y la cutıcula de lavaina interna de la raız, la cual constituye la capam�as interna. Esta vaina se forma a partir de lasc�elulas m�as perif�ericas de la matriz; se extiendedesde la matriz hasta la desembocadura delconducto de la gl�andula seb�acea en el folıculopiloso. La ausencia de la vaina interna de la raız apartir de ese punto origina un espacio, el canal delfolıculo piloso.
. El tallo del pelo, la porcion del folıculo queatraviesa la epidermis, posee tres capas:. La capa m�as perif�erica es la cutıcula del pelo,que se forma a partir de las c�elulas perif�ericasde la matriz.
. La capa intermedia, la corteza, deriva de lasc�elulas perif�ericas m�as cercanas al centro.
Conforme migran hacia la superficie, las c�elulasde la corteza fabrican filamentos de queratina,que se acumulan y embeben en una matriz detricohialina, un material semejante a laqueratohialina del estrato granuloso, para formarla queratina dura tıpica del tallo del pelo.
. El nucleo central del tallo, lam�edula, se desarrollaa partir de las c�elulas m�as centrales de la matriz.Las c�elulas de la corteza desplazan a las de lam�edula a medida que el pelo crece por encima dela superficie de la piel.
COLOR DEL PELO
El color del pelo se debe a la sıntesis de melanina porparte de los melanocitos intercalados en la matriz a lolargo de la l�amina basal adyacente a las papilasd�ermicas. Las c�elulas de la corteza rodean y seccionanlos extremos de las prolongaciones dendrıticas de losmelanocitos; el color del pelo comprende de rubioclaro a moreno oscuro en funcion de la cantidad demelanina que contengan las c�elulas de la corteza.Como se ha comentado anteriormente, los sujetospelirrojos poseen melanosomas redondeados, enlugar de ovalados. La coloracion gris�acea del cabellode las personas mayores se debe a la menor actividadde la tirosinasa, lo que dificulta la produccion de unacantidad adecuada del pigmento por parte de los me-lanocitos.
MUSCULOS ERECTORES DE PELO
Los musculos erectores del pelo son haces de mus-culo liso que se insertan en la capa papilar de la dermisy, en un �angulo oblicuo, el tejido conjuntivo querodea la raız externa del folıculo. La contraccion delas c�elulas del musculo liso provoca la elevacion deltallo del cabello y la depresion de la piel en el punto deinsercion. Las regiones no deprimidas de la epidermisparecen elevarse, lo que confiere el aspecto de piel degallina.
CRECIMIENTO DEL PELO
El crecimiento del pelo, a un ritmo de 2 a 3mmsemanales, tiene lugar en tres etapas: la fase an�agena,que puede tener una duracion de 6 anos en el pelo delcuero cabelludo y de unos meses en el de la axila;la fase cat�agena, un perıodo breve de involucion;y la fase telogena, en la que el folıculo permaneceen reposo hasta el desprendimiento del tallo y elcomienzo de la formacion de un nuevo tallo pilosoen el lugar del anterior. Los folıculos pilosos de ciertasregiones del organismopueden pasar de pelo velloso apelo terminal por influencia hormonal. En la puber-tad se forma vello pubico y axilar en chicos y chicas,y el pelo facial se torna m�as grueso en los primeros.
214
14
SISTEMATEG
UMENTARIO

CONSIDERACIONES CL�INICAS
El acn�e, una enfermedad cut�anea, constituye el
trastorno m�as frecuente en las consultas de
dermatologıa. Este t�ermino se utiliza para los poros
taponados, espinillas y bultos m�as profundos(quistes o nodulos) que se localizan en la cara, elcuello, el torax, la espalda, los hombros y losbrazos. Afecta, en mayor o menor medida,
pr�acticamente al 100% de los adolescentes. Sinembargo, no se limita a ningun grupo etario, ya que
incluso los sujetos en la quinta d�ecada de vida
pueden presentar acn�e. Las formas graves de
acn�e pueden dar lugar a cicatrices permanentes y
notables. En el desarrollo del acn�e inciden diversosfactores. La enfermedad se debe a obstruccionesque provocan la acumulacion del sebo en losfolıculos pilosos. La bacteria Propionibacterium
acnes sintetiza mol�eculas que provocan
enrojecimiento e inflamacion, adem�as de producirdiversas enzimas que degradan el sebo para formar
mol�eculas irritantes que reagudizan la inflamacion.Los androgenos, unas hormonas masculinaspresentes tanto en el hombre como en la mujer,
estimulan la hipertrofia de las gl�andulas seb�aceas yla produccion de sebo (durante la pubertad), lo quepodrıa provocar obstrucciones que evolucionarıan
hacia acn�e. Los estrogenos, unas hormonas
femeninas, se asocian a una mejora del acn�e en laschicas. El ciclo menstrual femenino obedece afluctuaciones de las concentraciones de
estrogenos, por lo que el acn�e puede mejorar y,posteriormente, empeorar conforme avanza dichociclo. Asimismo, se ha propuesto la existencia de
factores hereditarios en el desarrollo del acn�e,aunque no se han identificado aun.
215
14
SISTEMATEG
UMENTARIO
Figura 14.4 Gl�andula ecrina y gl�andula seb�acea y sus componentes celulares. (TomadodeGartner LP,Hiatt JL: Color Textbook ofHistology,
3rd ed. Philadelphia, Saunders, 2007, p 337.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Unas
Las unas (placas ungueales) (fig. 14.5) se componende placas gruesas de queratina dura y se hallan en lafalange distal de los dedos. La placa ungueal, locali-zada sobre el lecho ungueal, se desarrolla a partir dec�elulas de lamatriz ungueal situadas en esa region dela raız ungueal por debajo del pliegue ungueal proxi-mal, un repliegue sobre la epidermis. El estrato corneodel pliegue ungueal, denominado eponiquio (cutı-
cula), recubre la lunula, la region blanquecina de laplaca de la una. En los laterales de la placa ungueal, laepidermis se dobla para formar las paredes lateralesde la una, en los que cada una se delimita por unadepresion longitudinal, el surco ungueal. La epider-mis se pliega bajo el extremo libre de la placa y suestrato corneo da lugar al hiponiquio, similar a lacutıcula. El crecimiento de las unas de los dedos esmuy lento, de unos 2mmmensuales, y el de las de lospies es aun m�as lento.
216
14
SISTEMATEG
UMENTARIO

CONSIDERACIONES CL�INICAS
La onicomicosis es una micosis frecuente queafecta preferentemente a adultos y,especialmente, ancianos. Se manifiesta conengrosamiento, decoloracion, deformacion o
fractura de las unas. Es m�as frecuente en las unasde los dedos de los pies. En ausencia detratamiento, el roce de las unas muy engrosadascon el calzado puede provocar dolor einflamacion.
217
14
SISTEMATEG
UMENTARIO
Figura 14.5 Estructura de la una del dedo gordo de la mano. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.
Philadelphia, Saunders, 2007, p 343.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

15 APARATORESPIRATORIO
El aparato respiratorio –los pulmones y las vıas respira-torias que conducen a ellos– distribuye oxıgeno (O2)a las c�elulas del organismo, de las que recoge dioxidode carbono (CO2). La realizacion de estas funcionespor parte del aparato respiratorio depende de:
. La ventilacion (respiracion), la cualimpulsa el aire hacia los pulmones ydesde ellos
. La respiracion externa, que consisteen el intercambio del O2 inhaladopor el CO2 liberado por las c�elulas
. Transporte de O2 a las c�elulas yrecogida de CO2
. Respiracion interna, es decir, elintercambio de O2 por CO2 en elcompartimento citopl�asmico.
La ventilacion y la respiracionexterna tienen lugar dentro de los con-fines del aparato respiratorio; el trans-porte de O2 y la recogida de CO2 co-rresponden al aparato circulatorio, y larespiracion interna es un proceso quetiene lugar en todas las c�elulas vivas. Elfuncionamiento adecuado del aparato respiratorio sebasa en el suministro de aire por la porcion conduc-tora a la porcion respiratoria, en la que ocurre elintercambio gaseoso (respiracion externa).
Porcion conductora del aparato respiratorio
La porcion conductora del aparato respiratorio en-globa la cavidad nasal, la boca, la nasofaringe, la fa-ringe, la laringe, la tr�aquea, los bronquios primarios,los bronquios secundarios, los bronquıolos y losbronquıolos terminales. La presencia de elementosde tejido oseo, cartilaginoso y conjuntivo fibroel�asticomantiene la permeabilidad de este sistema de conduc-tos. El di�ametro de los conductos disminuye con-forme se ramifican y aproximan a la porcion respira-toria, si bien su numero aumenta; el di�ametro del �areatransversal aumenta en los niveles m�as profundos, loque da lugar a una disminucion de la velocidad del flujoa�ereo a medida que el aire inspirado se acerca a sudestino final, el alv�eolo. De igual modo, la velocidadde salida del aire espirado se incrementa conforme seaproxima a las narinas y los labios.
La cavidad nasal comienza en lasnarinas y finalizaen la coana; el tabique nasal oseo y cartilaginoso ladivide en dos mitades.
. Su porcion anterior est�a tapizada conpiel delgada (tabla 15.1) con vibrisas,las cuales filtran la materia particuladade gran tamano presente en el aireinspirado.
. La porcion posterior de la cavidadnasal se reviste de un epitelio cilındricoseudoestratificado en el que abundanlas c�elulas caliciformes (v. tabla 15.1).El tejido conjuntivo subyacente poseeuna red profusa de sinusoides de grantamano, gl�andulas seromucosas ynumerosas c�elulas linfoides yanticuerpos.
. La region olfatoria, situada en la caraposterosuperior de la cavidad nasal,presenta una coloracion amarillenta yalberga el epitelio olfatorio encargadode captar los olores (fig. 15.1). Elepitelio olfatorio se compone dec�elulas basales, sustentaculares yolfatorias.
. Las c�elulas basales son c�elulas regenerativaspequenas de dos tipos: horizontales, que danlugar a las siguientes, y globosas, las cuales sedividen para originar c�elulas sustentaculares yolfatorias.
. Las c�elulas sustentaculares fabrican el pigmentoamarillo que confiere esta coloracion al epitelioolfatorio. Estas c�elulas crean complejos de unioncon otras c�elulas sustentaculares y olfatoriascercanas, y prestan apoyo fısico y aislamientoel�ectrico a las c�elulas olfatorias. La vida media deeste tipo celular es de 12 meses.
. Las c�elulas olfatorias, neuronas bipolares delnervio craneal I (nervio olfatorio), se ocupan de lapercepcion de los olores. Sus dendritas seextienden para formar un pequeno bulbo, lavesıcula olfatoria, de la que parten cilios olfatoriosinmoviles hacia la capa mucosa de la cavidadnasal. El axonema de los cilios posee laconfiguracion de nueve dobletes alrededor de dosmicrotubulos sencillos centrales, aunque en suporcion distal se convierten en nueve
� 2011. Elsevier Espana, S.L. Reservados todos los derechos
TERMINOS CLAVE. Cavidad nasal. Epitelio olfatorio. Porcion conductora
del aparatorespiratorio
. Epitelio respiratorio
. Arbol bronquial
. Porcion respiratoriadel aparatorespiratorio
. Alv�eolo
. Intercambiogaseoso
218

219
15
APARATO
RESPIR
ATORIO
Tabla15.1
CARACTER�ISTICASDEL
APA
RATO
RESPIRATO
RIO
Division
Region
Sost� en
Gl� andulas
Epitelio
Tiposcelulares
Otrosrasgos
Conductora
extrap
ulm
onar
Vestıbulo
nasal
Cartılago
hialino
Gl� andulas
seb� aceasy
sudorıparas
Estratificado
escamoso
queratinizad
o
Epidermis
Vibrisas
Cavidad
nasal:
respiratoria
Cartılago
hialinoyhueso
Gl� andulas
seromucosas
Respiratorio
Basales,caliciform
es,ciliadas,
encepillo,serosas,SN
ED
Tejidodetipoer� ectil
Cavidad
nasal:
olfatoria
Hueso
Gl� andulasde
Bowman
Olfatorio
Olfatorias,susten
taculares,y
basales
Vesıcula
olfatoria
Conductora
extrap
ulm
onar
Nasofaringe
Musculo
esquel� etico
Gl� andulas
seromucosas
Respiratorio
Basales,caliciform
es,ciliadas,
encepillo,serosas,SN
ED
Amıgdalas
farıngeas
ytrompas
deEustaq
uio
Laringe
Cartılago
shialinoy
el� astico
Gl� andulas
mucosasy
seromucosas
Respiratorioy
estratificad
o
escamoso
no
queratinizad
o
Basales,caliciform
es,ciliadas,
encepillo,serosas,SN
ED
Epiglotis,pliegues
vocalesy
pliegues
vestibulares
Tr� aquea
ybronquios
primarios
Cartılago
hialinoyTC
colagenoso
irregu
lar
den
so
Gl� andulas
mucosasy
seromucosas
Respiratorio
Basales,caliciform
es,ciliadas,
encepillo,serosas,SN
ED
Anillosen
Cymusculo
traq
ueal(m
usculo
liso)en
adventicia
Conductora
intrap
ulm
onar
Bronquiossecundarios
(intrap
ulm
onares)
Cartılago
hialinoy
musculo
liso
Gl� andulas
seromucosas
Respiratorio
Basales,caliciform
es,ciliadas,
encepillo,serosas,SN
ED
Placasdecartılagohialinoy
dosban
das
demusculo
liso
enespiral
Bronquıolos
(primarios)
Musculo
liso
Ausentes
Cilındrico
simple
acubicosimple
C� elulasciliad
asyc� elulasde
Clara
(yalgu
nas
c� elulas
caliciform
esen
los
bronquıolosm� as
gran
des)
<1mm
dedi� ametro;
conducenaire
aloslobulos;
dosban
das
demusculo
liso
enespiral
Bronquıolos
term
inales
Musculo
liso
Ausentes
Cubicosimple
Algunas
c� elulasciliad
asy
numerosasc� elulasdeClara
(enau
senciadec� elulas
caliciform
es)
<0,5mm
dedi� ametro;
conducenaire
alos� acinos;
algu
nas
fibrasdemusculo
liso
Respiratoria
Bronquıolos
respiratorios
Algunas
fibrasdemusculo
liso
yfibrasdecol� ageno
Ausentes
Cubicosimple
y
escamoso
simple
muyaten
uad
o
Algunas
c� elulascu
bicas
ciliad
as,c� elulasdeClara
y
neu
mocitosdetiposIyII
Alv� eo
losen
pared
es;los
alv� eolosposeen
esfınteres
demusculo
liso
ensus
orificios
Conductosalveolares
Fibrasdecol� agenode
tipoIII(reticulares)
y
esfınteresdemusculo
liso
dealv� eolos
Ausentes
Escam
oso
simple
muyaten
uad
o
Neu
mocitosdetiposIyIIde
alv� eolos
Carecen
depared
espropias,
tansolo
unasecuen
cia
lineardealv� eolos
Sacosalveolares
Fibrasdecol� agenode
tipoIIIyfibrasel� asticas
Ausentes
Escam
oso
simple
muyaten
uad
o
Neu
mocitosdetiposIyII
Agregad
osdealv� eolos
Alv� eo
los
Fibrasdecol� agenode
tipoIIIyfibrasel� asticas
Ausentes
Escam
oso
simple
muyaten
uad
o
Neu
mocitosdetiposIyII
200mm
di� ametro;poseen
macrofago
salveolares
SNED,sistem
aneu
roen
docrinodifuso;TC,tejidoconjuntivo
.Tomad
odeGartner
LP,HiattJL:Color
Textbookof
Histology,3rd
ed.Philadelphia,Saunders,2007,pp
346–347.

microtubulos sencillos que rodean al parde microtubulos sencillos centrales. El extremocontrario del cuerpo de estas c�elulas corresponde asu axon, el cual contacta con los de otras c�elulasolfatorias para asociarse en fibras nerviosasolfatorias que atraviesan la placa cribiforme yestablecen sinapsis con c�elulas del bulboolfatorio.
La l�amina propia vascularizada est�a dotada de ele-mentos linf�aticos y gl�andulas de Bowman que secre-tan un lıquido seroso que contiene la proteına deunion a mol�eculas odorıferas, IgA y compuestos anti-microbianos.
HISTOFISIOLOGIA DE LA CAVIDAD NASAL
La mucosa nasal muy vascularizada alberga numero-sas gl�andulas que sintetizan mucosidad y un lıquidoseroso que mantiene humeda la superficie epitelial.
. La mucosidad atrapa la materia particuladasuspendida en el aire inspirado.
. Los cilios de las c�elulas cilındricas del epitelioseudoestratificado arrastran la mucosidad y laspartıculas atrapadas en ella hacia la porcionposterior de la faringe, en las que ser�an eliminadas.
. La profusa red vascular establece un mecanismo deflujo contracorriente que humidifica y calienta elaire inspirado.
. Los elementos linfoides presentes en la l�aminapropia capturan al�ergenos y antıgenos con el fin deeliminar todas las mol�eculas exogenas, aunque confrecuencia inducen sıntomas asociados a la rinitisal�ergica y el resfriado.
La percepcion de olores por el epitelio olfatorio(v. fig. 15.1) aporta la mayorıa de la informacion utili-zada para discernir sabores. No se conoce bien elmeca-nismo en el que se basa la diferenciacion olfatoria,aunque podrıa parecerse al sistema inmunitario, detal modo que las sustancias olorosas –mol�eculas quese disuelven en la mucosidad– se unen a receptores deolor localizados en la superficie de los cilios olfatorios.
. La union de las sustancias olorosas al numeronecesario de mol�eculas receptoras provoca ladespolarizacion de la membrana de la c�elulaolfatoria; el potencial de accion ası generado viaja alo largo de su axon para estimular a agregados dec�elulas mitrales en unas regiones pequenas,llamadas glom�erulos, situadas en el seno del bulboolfatorio.
. El millar de glom�erulos localizados en el bulboolfatorio reciben informacion procedente de unas2.000 neuronas olfatorias.
. Las permutaciones y las combinaciones de posiblesdatos aferentes hacen posible el reconocimiento de10.000 olores diferentes, lo que supone que cadaglom�erulo participa en el reconocimiento demuchos aromas distintos.
. El abundante lıquido procedente de las gl�andulas deBowman semueve por accion de los cilios olfatoriosy arrastra con rapidez las sustancias olorosas, lo queimpide la generacion de varias respuestas por unasola sustancia olorosa.
LARINGE
La laringe es un conductomusculocartilaginoso cortoque forma parte de la porcion conductora del aparatorespiratorio y se dispone entre la faringe y la tr�aquea.Se ocupa de la fonacion e impide la entrada de ali-mentos o lıquido en el aparato respiratorio durante ladeglucion.
. La laringe consta de varios cartılagos que seconectan entre sı mediante ligamentos y musculoesquel�etico extrınseco e intrınseco.
. La entrada, el orificio superior de la laringe, serecubre de la epiglotis, es una solapa cartilaginosaque permanece abierta durante la fonacion y larespiracion y se cierra en el transcurso de ladeglucion.
En la luz de la laringe aparecen dos tipos de pliegues:
. Los pliegues vestibulares superiores, que soninmoviles
. Los pliegues vocales inferiores, formados por tejidoconjuntivo el�astico denso regular, el ligamentovocal, y elmusculo vocal asociado; ambos ayudan alos dem�as musculos intrınsecos a modificar latension de los ligamentos vocales y moverlos ensentido medial, lo que reduce la anchura de lahendidura glotica, el espacio comprendido entrelos pliegues vocales derecho e izquierdo.
La salida forzada del aire exhalado junto a los plie-gues vocales hace que vibren para generar un sonidoque, al ser modulado por la lengua y los labios, crea elhabla. La longitud del pliegue vocal y el grado detension al que est�a sometido controla el tono delsonido; de este modo, el tono del sonido producidoser�a m�as agudo cuanto m�as corto sea el pliegue ymayor la tension en la cuerda vocal.
Las caras superiores de los pliegues vocales y laepiglotis se recubren de epitelio escamoso estratifi-cado no queratinizado, mientras que las porcionesrestantes se tapizan con epitelio cilındrico seudoes-tratificado ciliado. Los cilios de las c�elulas cilındricasque integran este epitelio transportan la mucosidadhacia la faringe, donde ser�a eliminada.
220
15
APARATO
RESPIR
ATORIO
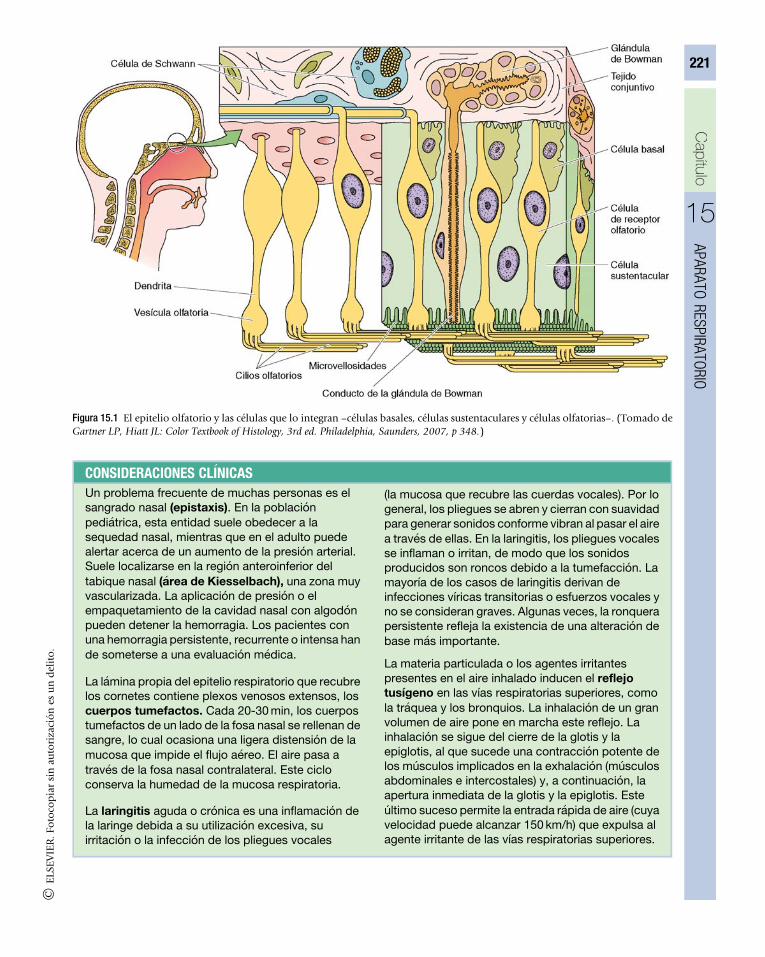
CONSIDERACIONES CL�INICAS
Un problema frecuente de muchas personas es elsangrado nasal (epistaxis). En la poblacion
pedi�atrica, esta entidad suele obedecer a lasequedad nasal, mientras que en el adulto puedealertar acerca de un aumento de la presion arterial.Suele localizarse en la region anteroinferior del
tabique nasal (�area de Kiesselbach), una zona muyvascularizada. La aplicacion de presion o elempaquetamiento de la cavidad nasal con algodonpueden detener la hemorragia. Los pacientes conuna hemorragia persistente, recurrente o intensa han
de someterse a una evaluacion m�edica.
La l�amina propia del epitelio respiratorio que recubrelos cornetes contiene plexos venosos extensos, loscuerpos tumefactos. Cada 20-30min, los cuerpostumefactos de un lado de la fosa nasal se rellenan desangre, lo cual ocasiona una ligera distension de la
mucosa que impide el flujo a�ereo. El aire pasa a
trav�es de la fosa nasal contralateral. Este cicloconserva la humedad de la mucosa respiratoria.
La laringitis aguda o cronica es una inflamacion dela laringe debida a su utilizacion excesiva, suirritacion o la infeccion de los pliegues vocales
(la mucosa que recubre las cuerdas vocales). Por logeneral, los pliegues se abren y cierran con suavidadpara generar sonidos conforme vibran al pasar el aire
a trav�es de ellas. En la laringitis, los pliegues vocalesse inflaman o irritan, de modo que los sonidosproducidos son roncos debido a la tumefaccion. Lamayorıa de los casos de laringitis derivan deinfecciones vıricas transitorias o esfuerzos vocales yno se consideran graves. Algunas veces, la ronquerapersistente refleja la existencia de una alteracion de
base m�as importante.
La materia particulada o los agentes irritantespresentes en el aire inhalado inducen el reflejotusıgeno en las vıas respiratorias superiores, como
la tr�aquea y los bronquios. La inhalacion de un granvolumen de aire pone en marcha este reflejo. Lainhalacion se sigue del cierre de la glotis y laepiglotis, al que sucede una contraccion potente delos musculos implicados en la exhalacion (musculosabdominales e intercostales) y, a continuacion, laapertura inmediata de la glotis y la epiglotis. Este
ultimo suceso permite la entrada r�apida de aire (cuyavelocidad puede alcanzar 150 km/h) que expulsa alagente irritante de las vıas respiratorias superiores.
221
15
APARATO
RESPIR
ATORIO
Figura 15.1 El epitelio olfatorio y las c�elulas que lo integran –c�elulas basales, c�elulas sustentaculares y c�elulas olfatorias–. (Tomado de
Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 348.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

TRAQUEA
La tr�aquea es un tubo de 12 cm de longitud que semantiene permeable gracias a la presencia de 10 a12 anillos de cartılago hialino en forma de C, los ani-llos en C, que refuerzan su estructura. La disposicion deestos anillos es tal que su porcion cerrada est�a orientadahacia la porcion anterior; cada anillo posee su propiopericondrio. Los pericondrios de los anillos en C veci-nos se conectan entre sı por medio de tejido conjuntivofibroel�astico. Los extremos abiertos, orientados hacia lacara posterior, se conectan entre sı por musculo liso, elmusculo traqueal, el cual se contrae para reducir la luzde este conducto y acelerar el flujo de aire a trav�es deeste tubo. La luz de la tr�aquea presenta una morfologıasimilar a la letra D. La tr�aquea est�a formada por trescapas: la mucosa interna, la submucosa intermedia y laadventicia externa. La mucosa traqueal se compone deun epitelio respiratorio, tejido conjuntivo subepitelial(l�amina propia) y una gruesa capa de fibras el�asticasque separa la mucosa de la submucosa. El epiteliorespiratorio, el cual corresponde a un epitelio cilın-drico ciliado seudoestratificado (fig. 15.2), contienec�elulas caliciformes, c�elulas cilındricas ciliadas, c�elulasbasales, c�elulas en cepillo, c�elulas serosas y c�elulas delsistema neuroendocrino difuso (SNED). Estas c�elulas sedisponen sobre la membrana basal, aunque no todasalcanzan la luz.
. Aproximadamente una tercera parte de las c�elulas deeste epitelio son c�elulas caliciformes (fig. 15.3),unas gl�andulas unicelulares que fabrican lasecrecion viscosa denominadamucinogeno, que sealmacena en vesıculas secretoras en la region apicaldilatada de su citoplasma, llamada teca. La mayorıade los org�anulos, como el nucleo, se concentra en laporcion basal delgada, el tallo. El mucinogeno sehidrata al ser secretado a la luz de la tr�aquea y seconvierte en mucina. La mucina se mezcla conmateria particulada en la luz a formar lamucosidad.
. El numero de c�elulas cilındricas ciliadas esequivalente al de c�elulas caliciformes. Estas c�elulasalargadas poseen un gran numero de cilios que
impulsan la mucosidad presente en la luz de latr�aquea hacia la laringe.
. Las c�elulas basales son unas c�elulas indiferenciadascortas; representan algo menos del 30% delcontenido celular del epitelio respiratorio. Estasc�elulas sufren divisiones mitoticas para sustituir alas c�elulas caliciformes, las c�elulas ciliadas y lasc�elulas en cepillo senescentes y muertas.
. Las c�elulas en cepillo, tambi�en llamadas c�elulascon gr�anulos pequenos, constituyen unaproporcion inferior al 3% del contenido celulartotal de dicho epitelio. Estas c�elulas estrechaspueden representar c�elulas caliciformes que hanliberado el mucinogeno o bien desempenar unafuncion neurosensorial a trav�es de lasmicrovellosidades largas que se extienden hacia laluz de la tr�aquea.
. La poblacion de c�elulas serosas es semejante a la delas c�elulas en cepillo. Se trata de c�elulas cilındricasque contienen gr�anulos secretores apicales cargadosde un lıquido seroso de funcion desconocida.
. Las c�elulas SNED suponen alrededor del 3% de lapoblacion celular. Se encargan de fabricar y secretarhormonas paracrinas o endocrinas como respuestaa estımulos tales como la hipoxia. Con ciertafrecuencia est�an inervadas y el complejo formadopor la fibra nerviosa y la c�elula SNED recibe elnombre de cuerpo neuroepitelial pulmonar, elcual puede activar a ciertas neuronas del centrorespiratorio del hipot�alamo en condiciones dehipoxia.
La l�amina propia de la tr�aquea es un tejido conjun-tivo fibroel�astico que alberga c�elulas linfoides y gl�andu-las serosas y mucosas que vierten su contenido a la luz.La capa externa de la l�amina propia, una gruesa l�aminael�astica, separa la mucosa y la submucosa.
La submucosa se compone de tejido conjuntivofibroel�astico denso irregular muy vascularizado. Enella abundan las gl�andulas seromucosas y los elemen-tos linfoides. Los anillos C de la tr�aquea se localizanen el tejido conjuntivo fibroel�astico de la adventicia.Esta capa externa une la tr�aquea a las estructuras cir-cundantes, como el esofago.
222
15
APARATO
RESPIR
ATORIO
CONSIDERACIONES CL�INICAS
La exposicion cronica a agentes irritantes, comohumo de cigarrillos o polvo de carbon, puedeprovocar alteraciones en el epitelio respiratorio, untrastorno conocido como metaplasia. De estemodo, en lugar de aparecer un epitelio cilındrico
seudoestratificado ciliado, la altura de las c�elulas
ciliadas es mucho menor, la poblacion de las c�elulascaliciformes se expande y se fabrica una mayorcantidad de mucosidad para eliminar los
compuestos irritantes. Sin embargo, la disminucion
del numero de c�elulas ciliadas dificulta la eliminacionde la mucosidad, lo que potencia la congestion. Por
otra parte, las gl�andulas seromucosas de la l�aminapropia y la submucosa se hipertrofian y secretancantidades mayores de sus productos. El epiteliorespiratorio recupera su morfologıa normal almodificarse las condiciones ambientales ydesaparecer los compuestos irritantes.
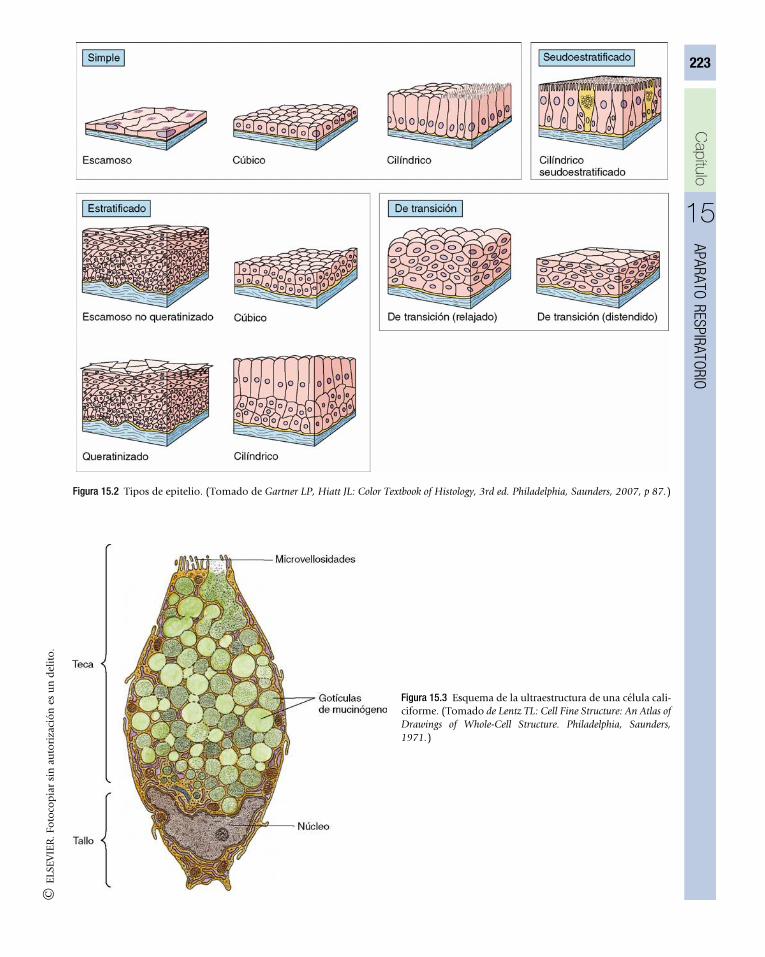
223
15
APARATO
RESPIR
ATORIO
Figura 15.2 Tipos de epitelio. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 87.)
Figura 15.3 Esquema de la ultraestructura de una c�elula cali-ciforme. (Tomado de Lentz TL: Cell Fine Structure: An Atlas of
Drawings of Whole-Cell Structure. Philadelphia, Saunders,
1971.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

ARBOL BRONQUIAL
La bifurcacion de la tr�aquea indica el comienzo del�arbol bronquial, la porcion conductora del aparatorespiratorio, que comprende desde los pulmones hastalos bronquıolos terminales. El di�ametro de las ramas del�arbol bronquial disminuye a medida que se ramifican,su numero aumenta y, como se ha senalado anterior-mente, el �area transversal total tambi�en lo hace, lo queralentiza el flujo de aire exhalado. El �arbol bronquialengloba los bronquios primarios, los bronquios secun-darios y terciarios, los bronquıolos y los bronquıolosterminales (fig. 15.4). Es preciso anadir algunas consi-deraciones generales acerca de la histologıa del �arbolbronquial amedida que se ramifica desde los bronquiosprimarios hacia los bronquıolos terminales:
. Disminuye el tamano del cartılago y las gl�andulas.
. Disminuyen la proporcion de c�elulas caliciformes yel espesor del epitelio.
. Aumenta la cantidad de tejido el�astico y musculoliso con relacion al grosor de la pared.
La tr�aquea se bifurca en los bronquios primarios(extrapulmonares), similares a este tubo si se excep-tuan sus menores di�ametro y espesor de la pared. Elbronquio primario izquierdo es menos recto que elderecho y tambi�en se bifurca. El bronquio derecho sedivide en tres ramas antes de penetrar en el par�en-quima pulmonar para convertirse en los bronquiosintrapulmonares (secundarios). Cada ramificacionde los bronquios primarios izquierdo y derecho per-tenece a uno de los cinco lobulos pulmonares.
Los bronquios secundarios (bronquios intrapul-monares, bronquios lobulares) son similares a losprimarios, si bien el cartılago hialino de refuerzo nopresenta una morfologıa en C. Se compone, por elcontrario, de pequenas piezas cartilaginosas querodean la luz circular, lo que confiere una morfologıaredondeada a la luz en un corte transversal.
. El cartılago no presenta brazos abiertos orientadoshacia la superficie posterior, por lo que el musculoliso migra hacia la luz para asentarse en la zona decontacto de la l�amina propia y la submucosa enforma de dos capas espirales.
. La adventicia fibroel�astica presenta fibras el�asticasque irradian desde esta capa para adoptar unadisposicion m�as o menos perpendicular a unatangente a la superficie de contacto con la pareddel bronquio secundario.
. Estas fibras el�asticas se unen a otras procedentes deotras porciones del par�enquima pulmonar.
En el pulmon, los bronquios secundarios se ramifi-can para formar bronquios terciarios (segmentarios),cada uno de los cuales conduce a 1 de los 10 segmen-
tos broncopulmonares, de tamano relativamentegrande, de cada pulmon. Conforme se subdividen,estos bronquıolos terciarios dan lugar a conductoscilındricos de tamano decreciente, en cuyas paredesllega a desaparecer el cartılago hialino, y poseen unacantidad mayor de c�elulas de musculo liso con relacional espesor de sus paredes que se disponen en doshaces en forma de h�elice. El bronquio menor delos bronquios secundarios se ramifica para convertirseen bronquıolos en los lobulos pulmonares, pequenassubdivisiones de un segmento broncopulmonar.
Una definicion util, aunque no disfruta de acepta-cion generalizada, de un bronquıolo lo considera unconducto de di�ametro menor de 1mm que conduceaire a un solo lobulo pulmonar. Los bronquıolos:
. Carecen de cartılago en sus paredes
. Poseen una envoltura relativamente gruesa demusculo con relacion al grosor de sus paredes
. Presentan fibras el�asticas procedentes del tejidoconjuntivo que envuelven los haces demusculo lisode tal modo que:. Se encuentran en una posicion m�as o menosperpendicular a una tangente situada en la zonade contacto con la pared del bronquıolo
. Establecen contacto con otras fibras el�asticas dedistinto origen, lo que tensa el perımetro delbronquıolo y mantiene su permeabilidad
. Carecen de gl�andulas en la l�amina propia
. Poseen un revestimiento de epitelio cilındricosimple a epitelio cubico simple ciliado
. Carecen de c�elulas caliciformes en el revestimientoepitelial de los bronquıolos de menor calibre
. Contienen c�elulas de Clara cilındricas, queaparecen en los bronquıolos de cualquier calibre, y:. Secretan una sustancia surfactante que ayuda amantener la permeabilidad bronquiolar
. Destruyen toxinas inhaladas y actuan comoc�elulas indiferenciadas en la regeneracion delepitelio
Los bronquıolos se ramifican en bronquıolos ter-minales (v. fig. 15.4), que representan los segmentosm�as pequenos de la porcion conductora del aparatorespiratorio (di�ametro<0,5mm), cada uno de loscuales se dirige a un lobulo pulmonar, el �acino pul-monar. Los bronquıolos terminales son similares alos bronquıolos, si bien su di�ametro es notablementemenor y est�an tapizados por un epitelio simple for-mado por c�elulas cilındricas (algunas de ellas, cilia-das) y c�elulas de Clara. La delgada l�amina propia serodea de varias c�elulas de musculo liso; las fibras el�as-ticas provenientes de la pared de tejido el�astico con-juntivo fibroel�astico mantienen la permeabilidad deltubo. Los bronquıolos terminales dan lugar a bron-quıolos respiratorios.
224
15
APARATO
RESPIR
ATORIO

225
15
APARATO
RESPIR
ATORIO
Figura 15.4 Conductos del aparato respiratorio. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia,
Saunders, 2007, p. 355.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Porcion respiratoria del aparato respiratorio
La porcion respiratoria del aparato respiratorio secompone de bronquıolos respiratorios, conductosalveolares, sacos alveolares y alv�eolos. Los bronquıolosrespiratorios (fig. 15.5) son similares a los bronquıolosterminales si se exceptua la presencia de algunosalv�eolos que se expanden desde sus paredes para llevara cabo el intercambio gaseoso, el cual no puede ocurriren los bronquıolos terminales. Los bronquıolos respi-ratorios se subdividen para formar los conductosalveolares (v. fig. 15.5), que representan una secuencialineal de alv�eolos que se ramifican para originar ungran numero de nuevos conductos alveolares.
. Cadaunode estos conductos alveolares termina enunsaco ciego integradopor dos o tres grupos de alv�eolos.
. Cada uno de estos grupos recibe el nombre de sacoalveolar.
. El espacio a�ereo del que provienen estos grupos sedenomina atrio.
. Los conductos alveolares, los sacos alveolares y losalv�eolos poseen una l�amina basal y mantienen lapermeabilidad a trav�es de fibras el�asticas delgadas quese unen a los escasos elementos del tejido conjuntivode estas estructuras y a otras fibras el�asticas vecinas.
Los alv�eolos (v. fig. 15.5) son unos pequenos espa-cios a�ereos tapizados por dos tipos de c�elulas: neu-mocitos de tipo I muy atenuados (c�elulas alveolaresde tipo I) y neumocitos de tipo II (c�elulas septales).La delgada pared del alv�eolo hace posible el intercam-bio deO2 yCO2. El orificio de cada alv�eolo se rodea dec�elulas de musculo liso y fibras reticulares circun-dantes con el fin de controlar su di�ametro. Se calculaque existen unos 300 millones de alv�eolos en ambospulmones, cada uno de los cuales tendrıa alrededorde 0,002 mm3, lo que supondrıa una superficie totalde 140 m2 dedicada al intercambio de O2 y CO2.
. Los neumocitos de tipo I son c�elulas muyatenuadas que representan una proporcion mayordel 95% de la superficie alveolar. La mayorıa de susorg�anulos se concentran en el �area perinuclear,mientras que el resto de la c�elula tiene unos80nm de di�ametro y contiene, fundamentalmente,citoplasma lıquido. Los neumocitos de tipo Ivecinos establecen uniones oclusivas entre sı paraevitar el paso de lıquido extracelular del tejidoconjuntivo interalveolar al alv�eolo.
. Los neumocitos de tipo II (fig. 15.6) son c�elulascubicas que solamente constituyen el 5% de lasuperficie alveolar a pesar de ser m�as abundantes quelos de tipo I. Estas c�elulas albergan cuerpos laminaresque liberana la luzdelalv�eoloenformadesurfactantepulmonar, una sustancia formada por dipalmitoilfosfatidilcolina y las apoproteınas surfactantes SP-A,
SP-B, SP-C y SP-D, que recubre la pared del espacioa�ereo alveolar y reduce la tension superficial parapotenciar la permeabilidad alveolar. Adem�as desintetizar y reabsorber el surfactante de maneracontinua, estas c�elulas pueden tambi�en dividirse paraoriginar nuevos neumocitos de tipos I y II.
El espacio comprendidoentredos alv�eolos vecinos sedenomina tabique interalveolar; se trata de una estruc-turadelgadaycontinuadetejidoconjuntivo irrigadapornumerosos capilares (v. figs. 15.5 y 15.6) que confierecierta estabilidad a estos delicados elementos. Los tabi-ques albergan monocitos, procedentes de la m�edulaosea, los cuales pasan a la luz alveolar y se conviertenenmacrofagos alveolares (c�elulas de polvo)para fago-citar materia particulada y microorganismos que hanlogrado acceder al alv�eolo a trav�es del aire inhalado,adem�as de surfactante con el fin de garantizar su recam-bioconstante.Las c�elulasdepolvo, cargadasconelmate-rial fagocitado, regresan al tabique interalveolar paraabandonar elpulmona trav�esde las vıas linf�aticasobienmigran a lo largo del �arbol bronquial para pasar a lafaringe, donde ser�an eliminadas junto a la mucosidadal ser ingeridas o expectoradas.
Los tabiques interalveolares pueden ser muy estre-chos y albergar solamente un capilar continuo cuyal�amina basal se fusiona con la del neumocito de tipo I,o bien contener elementos capilares y conjuntivos,como c�elulas linfoides, fibroblastos, y macrofagosrodeados de fibras reticulares, fibras el�asticas, sustanciafundamental y lıquido extracelular. El intercambio deCO2 presente en el alv�eolo por O2 de los capilares tienelugar conmayor facilidad en la zona en la que el tabiquealcanza un espesor m�as pequeno, llamada la barrerahematogaseosa (alveolocapilar), la cual consta de:
. Un endotelio capilar
. L�aminas basales fusionadas del endotelio capilar ylos neumocitos de tipo I
. Neumocitos atenuados de tipo I
. Surfactante
CONSIDERACIONES CL�INICAS
La enfermedad pulmonar obstructiva cronica(EPOC) es una combinacion de trastornospulmonares, entre los que figuran el enfisema y labronquitis. El enfisema es una enfermedad cronicacaracterizada por una destruccion alveolar queoriginadisnea; labronquitiscronicaesunaentidadenla que la obstruccion bronquial impide la respiracion.La EPOC cronica semanifiesta con tumefaccion,inflamacion, sibilancias, tos y reagudizacion de ladisnea. Aunque puede debutar con sıntomas leves,con el paso del tiempo reduce notablemente lacalidad de vida y conduce a una insuficienciarespiratoria que provoca la muerte del paciente.
226
15
APARATO
RESPIR
ATORIO

227
15
APARATO
RESPIR
ATORIO
Figura 15.5 A. Bronquıolo respiratorio, conducto alveolar, saco alveolar y alv�eolos. B. Relacion entre un alv�eolo y los capilarescontinuos. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 358.)
Figura 15.6 Neumocito de tipo II. (TomadodeGartner LP, Hiatt JL:
Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007,p 361.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

INTERCAMBIO GASEOSO ENTRE TEJIDOS Y PULMONES
La respiracion celular en el organismo se traduce en laformacion de unos 200ml de CO2 por minuto quepasan a los capilares para ser transportados a los pul-mones (fig. 15.7AyB), en losque se sustituir�a porO2. Lapresion parcial de CO2 en los tejidos supera la de lasangre, de modo que este gas pasa a los capilares pordifusion simple. La distribucion de esos 200ml de CO2
que ingresanen los capilares cadaminuto es la siguiente:
. 20ml pasan a formar parte del plasma como gasesdisueltos.
. 40ml se unen a la globina de la hemoglobina.
. 140ml entran en el eritrocito, en el que la anhidrasacarbonica cataliza una reaccion con agua cuyoproducto final es H2CO3, el cual se disocia paraoriginar H+ y HCO3
–. El ion bicarbonato difunde alplasma desde el citoplasma del eritrocito, comoconsecuencia de lo cual pasan iones Cl� alcitoplasma de estas c�elulas para compensar eldesequilibrio ionico ası creado, un proceso querecibe el nombre de cambio de cloruro (fig. 15.7C).
En los alv�eolos pulmonares, la presion parcial deloxıgeno es mayor que la de la sangre. Se produce, denuevo, un intercambio de ambos gases por difusionsimple, la cual no precisa de aporte energ�etico. Laliberacion de CO2 y la captacion de O2 tienen lugardel modo que se describe a continuacion (fig. 15.7D):
. Los iones bicarbonato presentes en el plasma pasanal citoplasma del eritrocito, lo que supone la salidade iones cloruro desde este compartimento, unproceso inverso al descrito anteriormente (querepresenta otro cambio de cloruro).
. Se forma �acido carbonico como consecuencia de launion de iones bicarbonato con iones H+.
. El oxıgeno ingresa en el citoplasma eritrocıtico y seune a la porcion hemo de la hemoglobina.
. La anhidrasa carbonica escinde el �acido carbonicopara formar agua y CO2.
. Los 200ml de CO2 que pasan a los capilares desdelos tejidos cada minuto abandonan el torrentecirculatorio, atraviesan la barrera hematogaseosapor difusion, ingresan en los espacios alveolares y seexhalan.
El oxido nıtrico (NO) desempena una funciondestacada en el intercambio gaseoso por su capacidadde union a dos sitios diferentes de la mol�ecula de lahemoglobina.
. En los tejidos, las c�elulas endoteliales liberan NO enel pulmon y este mediador se une a un sitio de la
mol�ecula de la hemoglobina; en los tejidos, elNO selibera y provoca la relajacion del musculo liso de losvasos sanguıneos y, por ende, la dilatacion del vaso,lo que facilita la salida del oxıgeno.
. El NO ocupa el segundo sitio de union de dichamol�ecula al unirse al lugar al que se asocio eloxıgeno liberado; de este modo, el NO se transportahacia los pulmones, donde se libera y pasa alespacio alveolar, de los que saldr�a junto al dioxidode carbono.
MECANISMO DE VENTILACION
Los procesos de inhalacion y exhalacion dependende la relacion anatomica de los pulmones, las mem-branas pleurales, las cavidades pleurales y las fibrasel�asticas del par�enquima pulmonar. La pleura visceral,que se continua con la pleura parietal en la base decada pulmon, recubre a este organo:
. La pleura parietal se adhiere a las paredes de la cajator�acica y a los elementos de tejido conjuntivo delmediastino.
. La pleura visceral se adhiere a la superficiepulmonar.
. El espacio comprendido entre la pleura parietal y lapleura visceral es una cavidad serosa, la cavidadpleural, un espacio hueco lubricado por un lıquidoseroso transparente que reduce la friccion creadapor el movimiento de los pulmones.
La contraccion de la musculatura de la paredtor�acica y el diafragma (musculos respiratorios) faci-lita la inhalacion. A medida que estos musculos secontraen en un proceso dependiente de energıa, lacaja tor�acica se expande y tracciona la pleura parietaladherida, de modo que aumenta el volumen de lascavidades pleurales y, por tanto, disminuye la presionen el interior de las mismas.
. En ese momento, la presion atmosf�erica es mayorque la de las cavidades pleurales, lo que provoca laentrada de aire en los pulmones.
. La entrada de aire en los pulmones distiende estosorganos, lo que incluye a las fibras el�asticas, y reduceel volumen de las cavidades pleurales expandidas, loque incrementa la presion en su seno.
La relajacion de los musculos respiratorios favorecela exhalacion, ya que permite:
. La recuperacion de la longitud normal de las fibrasel�asticas distendidas
. El aumento de presion en el interior de las cavidadespleurales para expulsar el aire de los pulmones.
228
15
APARATO
RESPIR
ATORIO

CONSIDERACIONES CL�INICAS
Los pulmones de los pacientes con congestionpulmonar e insuficiencia cardıaca congestiva seencuentran congestionados debido a la presenciade sangre extravasada. Los eritrocitos acceden a los
alv�eolos, donde son fagocitados por los macrofagosalveolares. Estos macrofagos reciben el nombre de
c�elulas de insuficiencia cardıaca cuando aparecen
en el par�enquima pulmonar y el esputo.
El c�ancer de pulmon es la causa m�as frecuente de
muerte debida al c�ancer tanto en hombres como enmujeres en EE. UU. Se cobra un numero mayor de
vidas que todas las debidas al c�ancer de colon,prostata ymama. Esta neoplasiamaligna consiste en
la proliferacion celular incontrolada del par�enquima
pulmonar, lo que puede producir met�astasis con
invasion del tejido adyacente y otros sistemas
org�anicos. La mayorıa de los casos corresponde acarcinomas pulmonares, los cuales se desarrollan
a partir de c�elulas epiteliales. Alrededor del 90% delos tumores pulmonares se vinculan con laexposicion prolongada a humo de cigarrillos. Los
sıntomasm�as frecuentes son disnea, tos, hemoptisisy adelgazamiento. Se distinguen dos tipos
principales de c�ancer de pulmon: el carcinomapulmonar microcıtico, el cual es susceptible detratamiento mediante quimioterapia, y el carcinoma
pulmonar no microcıtico, frente al cual est�a indicadala reseccion quirurgica. Amenudo, el tratamiento delcarcinoma pulmonar microcıtico incluye radioterapiaradical.
229
15
APARATO
RESPIR
ATORIO
Figura 15.7 A. Bronquıolo respiratorio, conducto alveolar, saco alveolar y alv�eolos.B.Relacion existente entre un alv�eolo y los capilares
continuos. C. Captacion de CO2 de tejidos corporales por eritrocitos y plasma. D. Liberacion de CO2 por eritrocitos y plasma en el
pulmon. (Comp�arese A con el conducto alveolar que aparece en la figura 15.4) (Tomado de Gartner LP, Hiatt JL: Color Textbook of
Histology, 3rd ed. Philadelphia, Saunders, 2007, p 358.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

16 APARATO DIGESTIVO:CAVIDAD BUCAL
El aparato digestivo est�a formado por un conductocontinuo y complejo que se ocupa de las funcionesde la ingestion modificada, la deglu-cion, la digestion, la absorcion denutrientes y lıquido, y la eliminacionde residuos no digeribles y gases. Laporcion glandular del aparato digestivopuede ser intramural o extramural.
Cavidad bucal
La cavidad bucal (boca), tapizada porun epitelio escamoso estratificadohumedo, se divide en dos espacios: elvestıbulo y la cavidad bucal propia-mente dicha. El conjunto formado por el tejido con-juntivo subepitelial y el epitelio constituye lamucosabucal.
. La mucosa recibe el nombre demucosa masticatoriacuando est�a queratinizada o paraqueratinizadadebido a la friccion; se localiza en las encıas, elpaladar duro y la porcion dorsal de la lengua.
. En la mayor parte de la cavidad bucal aparece unamucosa de revestimiento.
. La superficie dorsal de la lengua y algunas zonas delpaladar blando y la faringe poseen corpusculosgustativos, por lo que se conoce como mucosaespecializada.
Los pares de gl�andulas salivales mayores producensaliva, la cual contiene amilasa salival, los compuestosantimicrobianos lactoferrina y lisozima, e IgA; estasecrecion colabora en el mantenimiento de la hume-dad en la cavidad bucal. Al comer, el flujo de salivapermite la transformacion de los alimentos masticadosen el bolo alimentario, que puede ser deglutido.
Labios
Cada labio (fig. 16.1A) posee tres superficies: la caracut�anea externa con folıculos pilosos, la zona berme-llon y la cara mucosa interna humeda. Los capilaresdel aparato reticular alto de la zona bermellon seencuentran proximos a su superficie, lo que confiereuna coloracion rosada. La cara mucosa siempre est�ahumeda y presenta una mucosa de revestimiento contejido conjuntivo muy vascularizado dotado de un
gran numero de gl�andulas salivales menores muco-sas (y algunas serosas).
Dientes
El ser humano posee 20 dientesdeciduos que se sustituyen por ladentadura permanente.
El diente (fig. 16.1B) es unaestructura hueca con una coronarecubierta de esmalte y una raızrodeada de cemento que se unenen el cuello. La cavidad pulparhueca se divide en el conductoradicular y la c�amara pulpar que
contiene la pulpa; se rodea de dentina mineralizada.La raız se encuentra suspendida por el ligamento perio-dontal (LPD) de tejido conjuntivo colagenoso densoen el alv�eolo oseo.
La pulpa posee un nucleo central neurovascularenvuelto por tres capas conc�entricas:
. La zona rica en c�elulas, que se rodea de
. La zona pobre en c�elulas y
. La zona m�as externa, la capa odontobl�astica.
Un plexo de fibras nerviosas sensoriales, el plexo deRaschkow, que se encuentra en la zona de contactodel nucleo pulpar y la zona rica en c�elulas, conduce lassensaciones de dolor hacia el cerebro. Las fibras ner-viosas y los vasos sanguıneos acceden a la pulpa atrav�es del agujero apical del v�ertice radicular.
El esmalte, el tejidom�as duro del organismo, es unmaterial traslucido que envuelve la corona. Se com-pone de un 4% de matriz org�anica y un 96% dehidroxiapatita c�alcica, el cual aparece en formade cristales de gran tamano recubiertos por unamatrizorg�anica (enamelinas y tuftelinas) que originan bas-tones de esmalte (prismas de esmalte). Una c�elulaespecializada, el ameloblasto, sintetiza cada prisma;estas c�elulas mueren antes de que brote el diente, porlo que no se puede reparar el esmalte.
La dentina, de color amarillento, se halla en lacorona y la raız. Se compone de una matriz org�anicacon fibras de col�ageno de tipo I e hidroxiapatita c�alcica(que representa de un 65 a un 70% de la misma).La elasticidad de la dentina confiere cierta proteccional esmalte suprayacente frente a fracturas. La dentina
� 2011 Elsevier Espana, S.L. Reservados todos los derechos
TERMINOS CLAVE. Labios. Dientes. Desarrollo dental. Tejidos que sustentan
los dientes. Lengua. Corpusculos
gustativos
230

es fabricada por los odontoblastos, cuyas extensioneslargas ocupan espacios similares a tuneles, los tubulosdentinales, en el seno de la dentina.
El cemento, formado por un 50-55% de matrizorg�anica con fibras de col�ageno de tipo I y agua y un45-50% de hidroxiapatita c�alcica, aparece exclusiva-mente en la raız. En el cemento de la region apical de
la raız aparecen lagunas ocupadas por cementocitos(cemento celular), mientras que estas c�elulas noest�an presentes en el cemento de la coronal (cementoacelular). Ambos tipos de cemento se rodean de unamonocapa de cementoblastos, encargados de sinteti-zar esta sustancia. Los cementoclastos (odontoclas-tos) se ocupan de la resorcion del cemento.
231
16
APARATO
DIGESTIVO
:CAVID
ADBUCAL
Figura 16.1 A.Representacion de un diente in situ, en el que se aprecia el labio inferior, el vestıbulo y parte de la cavidad bucal.B.Dientey tejidos circundantes. El esmalte de la corona se encuentra con el cemento de la raız en el cuello dental. La dentina se localiza en la
corona y la raız. (A y B. Tomado deGartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, pp 368 and 369.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

232
16
APARATO
DIGESTIVO
:CAVID
ADBUCAL
CONSIDERACIONES CL�INICAS
LABIOLa queilitis angular es un trastorno doloroso en el quelas comisuras de los labios se resecan y agrietan;suele atribuirse a una carencia de vitamina B2,
carencia de zinc, o anemia ferrop�enica. Las lesiones
sonm�as frecuentes en ancianos con dentaduras malajustadas; la zona afectada suele presentarinfecciones por patogenos, como Candida albicans.
CAVIDAD BUCALEn la mucosa de revestimiento de los labios y lasmejillas se pueden formar ulceras, unas lesionespequenas caracterizadas por manchasblanquecinas con un reborde rosado, dolorosas y detamano reducido denominadas ulceras labiales(o aftas). Estas lesiones suelen relacionarse con el
estr�es y remiten al cabo de 7 a 10 dıas. El dolor sealivia mediante la aplicacion de una pomada
anest�esica local.
El carcinoma epidermoide constituye la neoplasia
maligna bucal m�as prevalente. Inicialmente, semanifiesta con una lesion rugosa o lisa de colorblanco o rojo en forma de bulto duro o depresionulcerada que sangra de manera ocasional. En
alrededor del 50% de los pacientes afectados, estoscarcinomas se desarrollan en la mucosa derevestimiento del labio, mientras que en el restopueden hacerlo en la lengua o el suelo de la boca.Con frecuencia, esta neoplasia se relaciona con eltabaquismo, el consumode alcohol o la utilizacion detabaco demascar, si bien puede aparecer en sujetosno fumadores o que no ingieren bebidas alcoholicas.El tratamiento suele consistir en una combinacion decirugıa y radioterapia.
CAMBIOS DENTALES RELACIONADOSCON LA EDADEl esmalte dental es un material inerte y las c�elulasencargadas de fabricarlo desaparecen tras laerupcion de los dientes, por lo que no puederegenerarse. Las fuerzas de friccion y desgaste de lamasticacion destruyen de manera continua elesmalte de las superficies de oclusion e incision, loque reduce su cantidad total. Se anade cemento a lasuperficie apical con el fin de compensar lareduccion del tamano de la corona, de modo que eldiente continua saliendo. Este movimientoposterupcion del diente asegura su contacto con eldiente homologo localizado en el arco contrario.
ODONTOGENIA PREVIA AL ESTADO DE CAMPANA
La odontogenia, o desarrollo dental, se pone enmarcha entre la sexta y octava semana del desarrolloembrionario, en el transcurso de las cuales el epiteliobucal de origen ectod�ermico prolifera para formar unal�amina dental en forma de herradura en el arco ma-xilar y la otra en el mandibular (fig. 16.2). Una mem-brana basal separa a esta l�amina del ectomes�enquimaderivado de la cresta neural
. Se forma una yemadental en 10 puntos diferentes decada l�amina dental, lo que supone el comienzo de laetapa de gemacion de la odontogenia. Cada una deestas 20 yemas anticipa a uno de los 10 dientesdeciduos. La condensacion del ectomes�enquima delextremo de cada yema origina una papila dental.
. Las c�elulas que integran la yema se dividen, demodoque esta crece para formar un organo del esmaltecon tres capas –la etapa de coronilla del desarrollodental–. El epitelio escamoso simple, el epitelioexterno del esmalte (EEE), prosigue en el buclecervical en anillo con el epitelio interno del esmalte(EIE), una capa cubico-escamosa simple concava.Las c�elulas de morfologıa estrellada del retıculoestelar se disponen entre ambos epitelios. Lamembrana basal rodea por completo al organo delesmalte, cuya cavidad ocupa la papila dental,formada por ectomes�enquima bien vascularizado.
El organo del esmalte y la papila dental constituyengermen del diente. El desarrollo del organo delesmalte altera su morfologıa, de modo que da lugar aun molde incisiforme, caniniforme o molariforme.Esta capacidad depende de la protuberancia delesmalte, una acumulacion de c�elulas dispersas entrelas c�elulas del retıculo estelar. Representa el principalcentro de senalizacion en la odontogenia. En la papiladental se sintetiza el factor de crecimientofibrobl�astico 4 (FGF-4) y el factor de crecimientoepid�ermico (EGF-4), necesarios para la supervivenciade dicha protuberancia, la cual sintetiza FGF-4, SonicHedgehog y diversas proteınas morfog�enicas dehueso que dirigen la transformacion del organo delesmalte en un molde molariforme. Al concluir latransformacion, en la papila dental se interrumpe laexpresion de EGF y FGF-4, de modo que laprotuberancia sufre un proceso de apoptosis. Laspapilas dentales de los organos del esmalteincisiformes y caniniformes no expresan FGF-4 niEGF, de modo que sus protuberancias del esmaltemueren durante la etapa de coronilla. La ausencia deesta protuberancia supone la formacion de un dienteexento de cuspide. Las papilas dentales fabrican ladentina y la pulpa de la raız. El ectomes�enquima querodea al germen del diente forma una delgada capade tejido conjuntivo denso, el saco dental, queorigina el alv�eolo, el LPD y el cemento dental. Los

ameloblastos, unas c�elulas diferenciadas a partir delEIE, sintetizan el esmalte. Los dientes permanentessurgen de las l�aminas suced�aneas de las 20 yemasdentales. Los 12 molares permanentes provienen delas extensiones de las dos l�aminas dentales, quecomienzan a alargarse.
. Conforme aumenta el tamano de la coronilla, seforma una cuarta capa, el estrato intermedio, quecaracteriza a la etapa de campana (etapa dehistodiferenciacion y morfodiferenciacion). Alproseguir el desarrollo, las c�elulas del EIE de la zonam�as lejana al asa cervical se diferencian en unasc�elulas alargadas, los preameloblastos, quecomienzan a sintetizar la matriz del esmalte; estasc�elulas maduran para convertirse en ameloblastos(fig. 16.3). La formacion inicial del esmalte induce ladiferenciacion de la capa de c�elulas de las papilasdentales adosadas a la l�amina basal enpreodontoblastos, los cuales se convierten enodontoblastos al comenzar la sıntesis de la matrizde dentina (v. fig. 16.3).
. Se define la union dentina-esmalte y se inicia laetapa aposicional del desarrollo dental. En unprimer momento, la union representa una regionmicroscopica que continua extendi�endose en laconcavidad del organo del esmalte hasta alcanzar el
asa cervical. A lo largo de este proceso prosigue lasıntesis de esmalte y dentina por parte de losameloblastos y los odontoblastos, respectivamente.Ambos tejidos se engrosan y los dos tipos celulares seseparan gradualmente.
. El asa cervical se alarga al concluir la formacion delesmalte y da lugar a una l�amina cilındrica, la vainaepitelial de la raız de Hertwig (VERH), formadaexclusivamente por EEE y EIE, que envuelve a lasc�elulas ectomesenquimatosas y se rodea de ellas. Lasc�elulas ectomesenquimatosas incluidas en esta vainase continuan con la papila dental y forman la pulparadicular y la dentina radicular. Las regiones m�asantiguas de la VERH comienzan a desaparecer yalgunas de las c�elulas ectomesenquimatosas que larecubren migran sobre la superficie de la dentinaradicular, se diferencian en cementoblastos yfabrican el cemento. El desarrollo de la raız prosigueamedida que la VERH se va alargando y, finalmente,se forma la ultima region de la raız, la cual alberga alagujero apical. El diente brota en la cavidad bucalconforme aumenta de tamano de la raız. Elmovimiento de erupcion no depende de laelongacion de la raız, si bien ambos sucesos se dande manera simult�anea. La salida del diente se debe ala actividad de unosmiofibroblastos especializados
233
16
APARATO
DIGESTIVO
:CAVID
ADBUCAL
Figura 16.2 A-G. Odontogenia. (Tomado deGartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 372.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

del saco dental que, al arrastrar las fibras de col�ageno Ide este saco (futuro LPD) ancladas al cemento,empujan al diente en desarrollo hacia la superficie.
ESTRUCTURAS ASOCIADAS A LOS DIENTES
El alv�eolo, el LPD y las encıas est�an asociadas a losdientes y favorecen el mantenimiento de la posicioncorrecta de cada diente en la cavidad bucal (fig. 16.4).
. El LPD se compone de tejido conjuntivo colagenosoirregular denso neurovascular y celular; ocupa elangosto espacio del ligamento (anchura � 0,5mmen una cavidad bucal sana) situado entre el cementoradicular y el alv�eolo (v. fig. 16.4). Las fibras decol�ageno de tipo I del LPD se organizan en gruposde fibras principales, los cuales soportan ycontrarrestan las fuerzas de la masticacion. Estasfibras suspenden al diente en el alv�eolo a trav�es defibras de Sharpey embebidas en el cemento y elhueso alveolar. Las c�elulas m�as numerosas del LPDson los fibroblastos, los cuales sintetizan la matrizextracelular, adem�as de degradar las fibras decol�ageno, lo que explica la extraordinaria tasa de
recambio de este tejido. El LPD cuenta con fibrasautonomas, sensoriales y propiorreceptoras, lasultimas de las cuales aportan informacion acerca dela orientacion espacial.
. El hueso que alberga la raız de cada pieza dental, elalv�eolo (v. fig. 16.4), se divide en tres regiones: elhueso compacto cuneiforme de escaso grosor ynumerosas perforaciones que se encuentra encontacto con el LPD, el hueso alveolarpropiamente dicho; el hueso esponjoso que lorodea, la esponjosa; y el hueso compacto gruesom�as externo, la placa cortical, dispuesta en sentidolingual y labial. Los vasos sanguıneos y las fibrasnerviosas del alv�eolo se encuentran en canalesnutrientes en forma de tunel. Estos vasos y fibrasatraviesan las perforaciones del hueso alveolarpropiamente dicho para irrigar e inervar,respectivamente, tanto el alv�eolo como el LPD.
. El epitelio escamoso estratificado orto- oparaqueratinizado de las encıas soporta las intensasfuerzas de friccion a las que est�a expuesto(v. fig. 16.4). De manera similar al LPD, los haces defibras de col�ageno de tipo I del tejido conjuntivo
234
16
APARATO
DIGESTIVO
:CAVID
ADBUCAL
Figura 16.3 Ameloblasto y odontoblasto. La extension larga del
odontoblasto se acorto al seccionar una porcion considerable de
la misma (espacio blanco). (Tomado de Lentz TL: Cell Fine Struc-
ture: An Atlas of Drawings of Whole-Cell Structure. Philadelphia,Saunders, 1971.)
Figura 16.4 Diente y tejidos circundantes. El esmalte de la corona
se encuentra con el cemento de la raız en el cuello dental. La
dentina se localiza en la corona y la raız. (Tomado deGartner LP,Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders,
2007, p 369.)

235
16
APARATO
DIGESTIVO
:CAVID
ADBUCAL
colagenoso irregular denso se organizan para formargrupos de fibras principales. A medida que elepitelio se acerca al esmalte, realiza un giro brusco yse une a trav�es de hemidesmosomas a la superficiedel esmalte en forma de una banda epitelial querodea al diente en su perımetro y recibe el nombre deepitelio de union. Esta delgada banda cuneiforme de1mm de longitud, que posee un m�aximo de50 c�elulas de ancho en su porcion coronal y menosde 7 en la region apical, impide la invasion del tejidoconjuntivo est�eril gingival por parte de la abundantemicroflora que puebla la cavidad bucal.
Paladar
El paladar, formado por el paladar duro fijo en laporcion anterior y el paladar blando muscular movilen la region posterior, separa la cavidad nasal de labucal.
. En la superficie bucal, el paladar duro est�atapizado por una mucosa masticatoria cuyo tejidoconjuntivo posee tejido adiposo en la regionanterior y gl�andulas salivales mucosas menores
en la posterior. El tejido conjuntivo del paladarduro se adhiere al entrepano oseo en su regioncentral. La cara nasal del paladar duro se componede tejido conjuntivo colagenoso irregular densorecubierto de un epitelio cilındricoseudoestratificado ciliado en el que abundan lasc�elulas caliciformes.
. La superficie bucal del paladar blando se recubre demucosa de revestimiento. En el tejido conjuntivoson numerosas las gl�andulas salivales mucosasmenores que se continuan con las gl�andulas delpaladar duro. El nucleo del paladar blando est�aformado pormusculos esquel�eticos, algunos de loscuales proceden del reborde anterior del entrepanooseo del paladar duro. La superficie nasal del paladarblando es id�entica a la del duro.
. La porcion terminal del paladar blando es la uvulaconica, cuyas superficies se recubren de mucosa derevestimiento y albergan gl�andulas salivalesmucosas menores dispersas entre elementos detejido conjuntivo. La porcion central de la uvulaposee fibras de musculo esquel�etico que intervienenen la elevacion de esta estructura en el transcurso dela deglucion.
CONSIDERACIONES CL�INICAS
El reflejo de sacudida mandibular provoca laapertura de la boca cuando se encuentra un objetoduro al masticar alimentos. Este reflejo comienzacuando la oposicion subita que encuentra el LPDorigina la inhibicion de los musculos implicados en lamasticacion por las fibras propiorreceptoras, lo queconfiere proteccion a la dentadura frente a fracturas.
Los traumatismos oclusales debidos a actividadesatıpicas, como el bruxismo (rechinar de dientesdurante la noche) o el apretado excesivo de losdientes pueden originar trombosis o, en el peor de
los casos, necrosis isqu�emica del LPD. Comoconsecuencia de estas lesiones, el espacio del LPD(es decir, el espacio comprendido entre el cemento
del diente y el alv�eolo oseo) se amplıa, con lo queaumenta la movilidad del diente; en ausencia de
tratamiento, puede ocasionar la p�erdida de esediente.
La accion de fuerzas ortodoncicas excesivamente
r�apidas sobre el diente puede originar danos a los
alv�eolos. Estas fuerzas se transfieren al LPD, el cual
se inflama y se reclutan osteoclastos hacia �el, que
llevan a cabo una resorcion del alv�eolo de mayormagnitud de lo planificado por el odontologo. Laampliacion excesiva del espacio LPD puede
traducirse en la p�erdida del diente comoconsecuencia de su movilidad irreversible.
La halitosis, o mal aliento, suele deberse a lapresencia de partıculas de alimentos atrapadasentre los dientes, las grietas en la lengua o las foveasde las amıgdalas palatinas, en las que los residuoscomienzan a descomponerse y generar un olordesagradable. Asimismo, las personas con unahigiene bucal deficiente o problemas derivados deuna endodoncia que hayan provocado la aparicionde abscesos suelen padecer halitosis. La ingesta deciertos alimentos, como ajo o cebolla cruda, confiereun olor desagradable al aliento que desaparece al
hacerlo los aceites vol�atiles contenidos en estosalimentos del torrente circulatorio. Las infeccionespor algunos patogenos bacterianos, comoHaemophilus influenzae, originan un olor dulce
mohoso caracterıstico que cualquier m�edico conformacion en microbiologıa deberıa reconocer.Otros trastornos menos prevalentes pueden conferirun olor determinado al aliento: olor a acetona en ladiabetes, olor a orina en la insuficiencia renal, y olor
mohoso en las hepatopatıas. Adem�as, algunos
tumores esof�agicos y g�astricos se asocian conhalitosis.
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Lengua
La lengua (figs. 16.5 y 16.6) es un organo musculargrande y dotado de una gran movilidad que no sola-mente participa en la masticacion al colocar los ali-mentos en el plano oclusal, sino que tambi�en inter-viene en la formacion del bolo y su deglucion. Lalengua posee, adem�as, cuatro tipos de papilas lin-guales, la mayorıa de las cuales sobresale por encimade la superficie y presenta una mucosa masticatoriacuyo epitelio escamoso estratificado muy queratini-zado les permite raspar alimento de una superficie.Otras papilas se recubren de epitelio escamoso estra-tificado no queratinizado que contiene corpusculosgustativos implicados en el reconocimiento desabores.
Los musculos de la lengua son voluntarios y seagrupan en dos categorıas:
. Los musculos extrınsecos provienen de otrasregiones, aunque se insertan en la lengua paramoverla.
. Los musculos intrınsecos se encuentran dentro dela lengua y modifican su morfologıa.
Se distinguen tres superficies en la lengua: dorsal,ventral y lateral. La superficie dorsal se divide en dostercios anteriores y un tercio posterior por el surcoterminal en forma de V, cuyo v�ertice posterior sedefine por una concavidad similar a una fosa, el agu-jero ciego. El tercio posterior se caracteriza por lapresencia de una mucosa de revestimiento de super-ficie irregular debido a la abundancia de ganglioslinf�aticos en el tejido conjuntivo subepitelial, conoci-dos en su conjunto como la amıgdala lingual. La raızde la lengua ancla este organomuscular a la base de lacavidad bucal y la faringe.
PAPILAS LINGUALES
Tres de los cuatro tipos de papilas linguales se loca-lizan en el dorso de los dos tercios anteriores de lalengua.
. Las m�as numerosas de ellas, las papilas filiformesmuy queratinizadas, carecen de corpusculosgustativos. Se proyectan por encima de la superficielingual y rascan alimentos de una superficie.
. Las papilas fungiformes son m�as escasas, remedanun champinon, se proyectan por encima de lasuperficie y se entremezclan de maneraaparentemente aleatoria con las papilas filiformes.Est�an tapizadas por un epitelio escamosoestratificado no queratinizado, por lo que parecenpuntos rojos en la superficie de la lengua. El epiteliode su cara dorsal contiene tres o cuatro corpusculosgustativos.
. Las papilas circunvaladas, que aparecen en unnumero cercano a 12, se localizan delante del surcoterminal. Est�an enterradas en la superficie y serodean de un surco cuyo revestimiento epitelialcuenta con corpusculos gustativos. El suelo de estesurco recibe delgados conductos de las gl�andulas devon Ebner.
. La superficie lateral de la cara posterior de los dostercios anteriores de la lengua posee regionessimilares a surcos longitudinales, las papilasfoliadas, que se asemejan a las hojas de un libro. Loscorpusculos gustativos de estas papilas degeneranhacia el tercer ano de vida. En la porcion m�asprofunda del surco desembocan los conductosdelgados de las gl�andulas de von Ebner salivalesserosas menores.
CORPUSCULOS GUSTATIVOS
Los corpusculos gustativos (v. figs. 16.5 y 16.6) sonun conjunto intraepitelial de c�elulas procedentes de lacresta neural que forman una estructura en forma debarril cuyo orificio, el poro gustativo, presentamicro-vellosidades –los llamados pelos gustativos– quesobresalen. El corpusculo gustativo est�a formado por60 a 80 c�elulas fusiformes que se desprenden conti-nuamente y se sustituyen por nuevas c�elulas. Los cercade 3.000 corpusculos gustativos intervienen en la for-macion de cinco (o, quiz�as, seis) sensaciones gustati-vas primarias: amarga, dulce, salada, agria, deliciosa y,para algunos sujetos, grasienta. Cada corpusculo gus-tativo se localiza en el interior del epitelio y est�a for-mado por cuatro tipos de c�elulas, tres de las cualestienen una vida media de 10 dıas. El cuarto tipo celu-lar, la c�elula basal (c�elula de tipo IV), es un tiporegenerativo cuya actividad mitotica da lugar a nuevasc�elulas. Los otros tres tipos son los siguientes:
. C�elula de tipo I (c�elula oscura)
. C�elula de tipo II (c�elula p�alida)
. C�elula de tipo III (c�elula intermedia)
Se cree que las c�elulas basales originan c�elulas detipo I que se diferencian en c�elulas de tipo II, las cualesse convierten en c�elulas de tipo III al degenerar yposteriormente mueren. Las c�elulas de los tres pri-meros tipos poseen microvellosidades (pelos gustati-vos), unas estructuras con capacidad de respuesta aestimulantes del gusto presentes en los alimentosque se disuelven en la saliva. Algunas de estas sustan-cias activan canales ionicos (saladas y agrias), otrasinducen receptores ligados a proteınas G (deliciosas,dulces y amargas), y otras activan transportadores de�acidos grasos (lıpidos). La mayorıa de las sensacionesgustativas que se asocian a los alimentos dependen delolor, enmayormedida que del gusto percibido por loscorpusculos gustativos.
236
16
APARATO
DIGESTIVO
:CAVID
ADBUCAL

237
16
APARATO
DIGESTIVO
:CAVID
ADBUCAL
Figura 16.5 Lengua en la cavidad bucal y corte de la superficie anterior de sus dos tercios anteriores en el que semuestran distintos tipos
de papilas linguales. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 377.)
Figura 16.6 Seccion de la lengua en la que se muestran
distintos tipos de papilas linguales y un diagrama de un
corpusculo gustativo. (Tomado de Gartner LP, Hiatt JL:Color Textbook of Histology, 3rd ed. Philadelphia, Saunders,
2007, p 378.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

17 APARATO DIGESTIVO:TUBO DIGESTIVO
El tubo digestivo consiste en una estructura tubular de9m de largo, compuesta por el esofago, el estomago,los intestinos delgado y grueso y elconducto anal. Digiere los alimen-tos, absorbe los nutrientes y el agua,y compacta y elimina los compo-nentes indigeribles de la comidaingerida.
Organizacion general del tubodigestivo
El tubo digestivo est�a compuesto poruna serie de cilindros conc�entricos entorno a una luz. Estas capas se trans-forman a lo largo de su recorrido,pero antes de pasar a analizar expo-ner sus modificaciones, se describir�asu patron general (fig. 17.1):
. La luz est�a recubierta por una capa epitelial y unestrato de tejido conjuntivo subepitelial, la l�aminapropia, que alberga las gl�andulas y los folıculoslinf�aticos que integran el tejido linf�atico asociado ala mucosa. La l�amina propia est�a rodeada por lamuscular de la mucosa, dos capas de musculo lisoque adoptan una disposicion helicoidal: unacircular interna y otra longitudinal externa. Elepitelio, la l�amina propia y la muscular de lamucosa constituyen en conjunto la mucosa.
. Alrededor de la mucosa hay un tejido conjuntivocol�ageno denso, la submucosa, que representa unaregion vascularizada que contiene gl�andulas, perosolo en el esofago y en el duodeno. El plexosubmucoso de Meissner, un componente delsistema nervioso ent�erico, ocupa la capa m�asperif�erica de esta submucosa, a cargo de lasgl�andulas intraparietales, el riego vascular y lamuscular de lamucosa, con una accion a nivel local.
. La muscular externa est�a compuesta por musculoliso distribuido en dos capas de disposicion espiral:una circular interna y otra longitudinal externa. Elplexo de Auerbach (mient�erico) se situa entre estasdos l�aminas musculares y controla el peristaltismo.Su accion la ejerce a nivel local y tambi�en sobrelargos tramos del tubo digestivo.
. El tubo digestivo est�a recubierto por un tejidoconjuntivo (adventicia), que fija parte de sus
regiones a la pared corporal, o porun epitelio pavimentoso simplehumedo (serosa), que reduce elroce generado por su movimientocon el peristaltismo.
El sistema nervioso ent�erico puedeactuar de manera plenamente inde-pendiente; sin embargo, se ve modu-lado por los sistemas nerviosos sim-p�atico y parasimp�atico.
Esofago
El esofago, un conducto muscularque mide 25 cm de longitud y cuya
luz se contrae cuando no est�a transportando un bolohacia el estomago, presenta la siguiente organizaciongeneral.
. Lamucosa esof�agica est�a compuesta por un epiteliopavimentoso estratificado no queratinizado y unal�amina propia cuyas gl�andulas esof�agicas cardialesproducen moco que facilita la deglucion del bolo.Estas gl�andulas est�an situadas en las regionescercanas a la faringe y al estomago. La capamuscularde la mucosa est�a constituida por una capa demusculo liso con una disposicion longitudinal.
. La submucosa vascular posee las aut�enticasgl�andulas esof�agicas, que generan secrecionesmucosas y serosas. El componente seroso de estagl�andula fabrica pepsinogeno (una proenzima) ylisozima, un producto antibacteriano. Elcomponente mucoso lubrica el epitelio. El esofagopresenta gl�andulas en su submucosa.
. La muscular externa, integrada por una capacircular interna y otra longitudinal externa, esatıpica, porque en el tercio superior del esofago,cerca de la faringe, ambas est�an formadas pormusculo estriado; en el tercio medio lo est�an pormusculo estriado y liso, y en el inferior, junto alestomago, unicamente tienen musculo liso.
. El estrato m�as externo del esofago es la adventiciaen el torax y una serosa nada m�as penetrar en lacavidad abdominal.
� 2011 Elsevier Espana, S.L. Reservados todos los derechos
TERMINOS CLAVE. Esofago. Estomago. Gl�andulas g�astricas. C�elulas del SNED. Produccion de �acido
clorhıdrico. Intestino delgado. Intestino grueso. Recto
238

CONSIDERACIONES CL�INICAS
Los sıntomas m�as frecuentes indicativos de untrastorno del aparato digestivo consisten en disfagia(problemas de deglucion), regurgitacion,estrenimiento, diarrea y hemorragia del tubodigestivo. La disfagia puede tener numerosascausas, como las obstrucciones fısicas de la faringeo el esofago, las alteracionesmusculares o nerviosasy las perturbaciones psicogenas. La regurgitacionno se acompana ni de n�auseas, ni de la violentacontraccion de la musculatura abdominal quesucede durante el vomito. El proceso deregurgitacion puede deberse a una afeccion
nerviosa o muscular ligada al esfınter esof�agico oderivar de una estenosis del esofago a raız de un
c�ancer o un reflujo de �acido sin tratar. Si no existeninguna causa fısica, la regurgitacion se denominamericismo; en este caso, el alimento deglutidovuelve a la boca, donde puedemasticarse y tragarsede nuevo. El mericismo es frecuente entre los
lactantes; tambi�en puede darse en los adultos y
suele guardar una relacion con el estr�es.
El estrenimiento es un cuadro en el que ladefecacion tiene lugar menos de tres veces
semanales. Esm�as habitual entre las mujeres que enlos hombres, en las personas mayores de 65 anos yen las embarazadas. Por regla general, los individuosestrenidos elaboran unas heces pequenas, oscuras,secas y duras, cuya expulsion es difıcil. Entre las
causas del estrenimiento figuran las diet�eticas (p. ej.,un consumo escaso de fibra, la ingestion de
excesivos productos l�acteos, la bebida de pocos
lıquidos); un estilo de vida sedentario; determinadosmedicamentos, contando el abuso de los laxantes; ladesorganizacion de las costumbres diarias, como enlos viajes; y los problemas de salud, tales como unictus y las enfermedades intestinales. La diarreadesigna la produccion de heces sueltas y acuosas unmınimo de tres veces al dıa. Existen dos tipos dediarrea: aguda y cronica. La diarrea aguda esbastante corriente, dura 1 o 2 dıas y desaparece sin
m�as; si supera los 2 dıas, se considera una diarreacronica, y habrıa que acudir a un m�edico para evitar
la deshidratacion y descartar un proceso org�anico.La mayor parte de los casos de diarrea aguda sedeben a infecciones bacterianas, vıricas oparasitarias, mientras que la diarrea cronicanormalmente resulta de alguna alteracion en el tubodigestivo, como el sındrome del intestino irritable.
La sangre de las hemorragias del tubo digestivopuede salir por la boca con el vomito o por el ano conlos excrementos o las heces. Por su aspecto puedeser reciente o estar coagulada. Si la sangre es
reciente, lo m�as f�acil es que tenga un color rojo y
surja cerca de la cavidad bucal o del ano. Si est�acoagulada, se manifiesta como unas partıculas de
materia negra parecidas a los posos de caf�e en elvomito o tine de negro las heces (melena). Lashemorragias del tubo digestivo pueden estar
ocasionadas por ulceras p�epticas, varices querezumen sangre, el empleo de ciertosantiinflamatorios, la enfermedad intestinal
inflamatoria y el c�ancer.
239
17
APARATO
DIGESTIVO
:TUBODIGESTIVO
Figura 17.1 Organizacion general del tubo digestivo. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia,
Saunders, 2007, p 382.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

240
17
APARATO
DIGESTIVO
:TUBODIGESTIVO
Estomago
El estomago tiene una entrada, donde desemboca elesofago, y una salida, que se comunica con el duo-deno. Su capacidad puede pasar desde los 50mlcuando est�a vacıo hasta unos 1.500ml una vez dis-tendido. Al recibir un bolo desde el esofago, segregalos jugos g�astricos para licuarlo en un fluido �acidollamado quimo y empezar a digerirlo a trav�es del�acido clorhıdrico y sus enzimas, renina, pepsina ylipasa g�astrica. La hormona grelina mantiene unapresion intraluminal constante al permitir que lamus-cular externa se adapte al volumen en expansion yconserva la sensacion de hambre mientras aumentael estomago. El quimo �acido se segrega al duodeno enporciones de 1 a 2ml a trav�es del esfınter pilorico, lacapa circular interna de la muscular externa modifi-cada. El estomago tiene:
. Una region cardial en la curvatura menor concava
. Una region pilorica en la curvatura mayor
. Dos regiones anatomicas m�as, el fondo y el cuerpo,id�enticas desde el punto de vista histologico y que sedenominan region fundica (fig. 17.2).
La luz del estomago vacıo muestra pliegues, rugo-sidades de lamucosa y la submucosa, que desaparecenal distenderse. El revestimiento de este organo ofrecenumerosas depresiones cubiertas de epitelio, las crip-tas g�astricas (fov�eolas), cuyo suelo se encuentra per-forado pormultiples gl�andulas g�astricas tubulares quellenan la l�amina propia muy vascularizada. La mus-cular de la mucosa posee tres capas de musculo liso:circular interna, longitudinal externa y una capa obli-cua m�as externa poco definida. La submucosa notiene nada que senalar. La muscular externa englobalas capas circular interna, longitudinal externa y obli-cua m�as interna.
. El epitelio cilındrico simple de la region fundicaest�a compuesto de c�elulas superficiales derevestimiento, muy compactas, y c�elulasregenerativas; las membranas plasm�aticaslaterales de estas c�elulas forman uniones estrechasentre sı. Las c�elulas superficiales de revestimientoproducen el moco visible espeso que protege alepitelio frente al quimo �acido, y las regenerativasproliferan como recambio del revestimientoepitelial del estomago.
. Las criptas g�astricas de las regiones fundica y cardialrecorren un tercio de su longitud en la l�aminapropia, que est�a repleta de gl�andulas g�astricas.
. Cada gl�andula tiene seis tipos celulares, que siguenuna distribucion desigual en sus tres regiones: elistmo, que perfora la cripta g�astrica, el cuello y labase, que linda con la muscular de la mucosa. Lascriptas g�astricas de la region pilorica penetran lamitad de la l�amina propia.
COMPOSICION CELULAR DE LAS GLANDULAS FUNDICAS
V�ease la figura 17.2.
. Las c�elulas mucosas del cuello fabrican un mocosoluble que se anade al quimo y lubrica el tubodigestivo. Las membranas plasm�aticas de estasc�elulas establecen uniones estrechas con sus vecinas.
. La gran velocidad mitotica de las c�elulasregenerativas sustituye todo el revestimientoepitelial cada 5-7 dıas.
. Las c�elulas parietales (oxınticas) no suelen apareceren la base de la gl�andula. Presentan unos profundosconductillos intracelulares recubiertos demicrovellosidades. Una red vesicular intracelular, elsistema tubulovesicular, cuyo componente demembranaposee numerosas bombas de protonesH+,K+-ATPasa, va paralela a los conductillosintracelulares. Las c�elulas parietales producen �acidoclorhıdrico y el factor intrınseco g�astrico.Al elaborarHCl, el sistema tubulovesicular reduce su tamano conel correspondiente aumento en la cantidad demicrovellosidades, lo que indica que la red vesicularse emplea para almacenar membranas destinadas a lageneracion de microvellosidades, lo que permite a lac�elula incrementar su superficie durante su actividadsecretora. Al dejar de generar HCl, las membranas delas microvellosidades regresan al sistema tubulo-vesicular. Este proceso consume energıa, suministradapor las abundantes mitocondrias que contienenlas c�elulas parietales. La glucoproteına factorintrınseco g�astrico se libera a la luz del estomago,donde forma un complejo con la vitamina B12
para su absorcion por las c�elulas del ıleon.. Las c�elulas principales (cimogenas), situadas sobretodo en la base de las gl�andulas fundicas y ausentesdel todo en las gl�andulas cardiales o piloricas,fabrican pepsinogeno, quimosina y lipasa g�astrica.La proenzima pepsinogeno se convierte en laenzima proteolıtica pepsina en el medio �acido delestomago; la quimosina es una enzima proteolıticaque cuaja la leche, y la enzima lipasa g�astrica degradalos lıpidos. Estas enzimas se encuentran encerradasen el interior de vesıculas en el citoplasma apical delas c�elulas principales y se expulsan debido a lainteraccion de acetilcolina y secretina cuando seunen a sus receptores respectivos en la membranaplasm�atica basal de dichas c�elulas para activar sussistemas de segundos mensajeros.
. Las c�elulas del sistema neuroendocrino difuso(SNED) son de dos tipos—abiertas y cerradas—, delas que las primeras llegan a la luz y las ultimas no.Cada c�elula del SNED da lugar a una hormonaconcreta que se desprende a la l�amina propia. Estashormonas son autocrinas si van dirigidas a la c�elulaque las emitio, paracrinas si van dirigidas a unac�elula cercana y endocrinas si tienen que viajar atrav�es de la circulacion sanguınea.

CONSIDERACIONES CL�INICAS
La produccion excesiva de HCl en el estomagopuede deberse a multiples causas, entre ellas a una
neoplasia maligna de las c�elulas G, las c�elulas delSNED que elaboran la hormona paracrina gastrina.
Las personas con este tipo de c�ancer tienen unsındrome de Zollinger-Ellison, y el tumor puedesituarse en diversas zonas del aparato digestivo,
como las vıas biliares, el duodeno o el p�ancreas.Estos pacientes sufren ulceras recidivantes que noresponden a los tratamientos normalesantiulcerosos, como los antibioticos contra elHelicobacter pylori o los antihistamınicos H2. Losinhibidores de la bomba de protones resultaneficaces, pero su curacion supone la extirpacionquirurgica del tumor.
La posicion normal que ocupan el estomago y la
union gastroesof�agica es bajo el diafragma en lacavidad abdominal. Sin embargo, en algunos casosel estomago puede sobresalir hacia la cavidad
tor�acica, cuadro denominado hernia de hiato. Estees un trastorno muy frecuente, y su incidencia crececon la edad. Hay dos tipos de hernia de hiato. En la
hernia de hiato por deslizamiento, que es la m�as
habitual, la union gastroesof�agica y parte delestomago resbalan hacia arriba y hacia abajo a
trav�es de una zona d�ebil en el hiato esof�agico del
diafragma. El segundo tipo, la hernia de hiato
paraesof�agica, es menos corriente pero puedetener mayor importancia. En este proceso, la union
gastroesof�agica permanece en la cavidadabdominal, pero una parte del estomago penetra en
la cavidad tor�acica a trav�es del hiato esof�agico y secoloca al lado del esofago; si queda atrapada en estelugar, existe el riesgo de una posible estrangulacion
de dicha porcion g�astrica. Si el estomago no seretrae por su cuenta, es necesario operar.
La hernia de hiato por deslizamiento suele ser
asintom�atica, aunque puede manifestarse por un
reflujo g�astrico, pirosis y dispepsia, sobre todo en las
personas que se tumban despu�es de comer. Losmismos sıntomas acompanan a la hernia de hiato
paraesof�agica; sin embargo, si se estrangula, elenfermo experimenta un dolor retroesternal. Comoeste sıntoma se asemeja a un posible infarto demiocardio, su aparicion deberıa motivar la busqueda
inmediata de asistencia m�edica. En la mayorıa de loscasos no es necesario operar. El tratamiento de las
hernias de hiato conlleva cambios en los h�abitos
diet�eticos; el paciente deberıa comer m�as a menudo
y cantidades m�as reducidas en cada ocasion.
Asimismo, los anti�acidos y la elevacion de lacabecera de la cama suelen aliviar los sıntomas.
241
17
APARATO
DIGESTIVO
:TUBODIGESTIVO
Figura 17.2 Representacion gr�afica de la mucosa en la region fundica del estomago que muestra las gl�andulas fundicas de la l�amina
propia. Semuestran los diversos tipos de c�elulas que integran el epitelio del estomago y las gl�andulas fundicas. (Tomado deGartner LP,
Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 386.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

HISTOFISIOLOGIA GASTRICA
De manera diaria, las gl�andulas del estomago produ-cen unos 2 o 3 l de secreciones, cuyos principales cons-tituyentes son agua, HCl, enzimas, factor intrınseco ymoco visible y soluble. Al recubrir el revestimiento delestomago, el pH neutro del moco visible lo protege delos jugos g�astricos �acidos. Tambi�en proporciona unmedio favorable para elH. pylori, la bacteria que resideen �el.
. La muscular externa del estomago sirve pararevolver y licuar los alimentos ingeridos, formandoun quimo espeso parecido a una sopa de guisantessecos.
. La muscular de la mucosa garantiza que toda lasuperficie epitelial del estomago entre en contactocon el quimo. Cuando su consistencia es laadecuada, y segun su acidez, osmolalidad ycontenido calorico y lipıdico, las c�elulas del SNEDduodenales liberan la hormona gastrina, que haceque se abra el esfınter pilorico y se contraigan losmusculos longitudinales del pıloro, lo que inyecta1 o 2ml de quimo en el duodeno. Si el quimo no est�alisto para su salida, las c�elulas del SNED emiten lashormonas colecistocinina y polip�eptido g�astricoinhibidor, que inhiben la apertura del esfınterpilorico, y el quimo permanece en el estomago.
La produccion de HCl en el estomago atraviesa tresfases: cef�alica, que se refiere al pensamiento, el olor ola vision de la comida; g�astrica, por su presencia en laluz del estomago; e intestinal, por su presencia en elduodeno. No obstante, el mecanismo seguido por lasc�elulas parietales es el mismo, sea cual sea la fase quelo desencadeno. La cara basal de la membrana plas-m�atica de las c�elulas parietales posee receptores para elneurotransmisor acetilcolina y para las hormonasparacrinas gastrina e histamina 2. Cuando los treslugares se han unido a sus respectivas mol�eculas sena-lizadoras, la elaboracion de HCl y su secrecion hacialos conductillos intracelulares tiene lugar del modosiguiente (fig. 17.3):
1. La anhidrasa carbonica, presente en el citosol,cataliza la obtencion de H2CO3, que se disocia enH+ y HCO3
�.2. El transporte activo intercambia el H+ intracelular
por K+ extracelular situado en el conductillointracelular. A pesar de que el K+ tambi�en sigue untransporte activo en la membrana plasm�atica basalde la c�elula parietal, asimismo sale de ella porcanales de K+ ubicados en esa misma zona.
3. El K+ y el Cl� experimentan un transporte activoque los saca de la c�elula y los introduce en losconductillos intracelulares, donde se combinan elH+ y el Cl� para formar HCl.
4. La circulacion de iones entre la c�elula parietal y ellıquido extracelular de la l�amina propia modificalas presiones osmoticas, lo que provoca el flujo deH2O hacia la c�elula parietal.
5. El movimiento de iones entre la c�elula parietal ylos conductillos intracelulares cambia la presionosmotica de la primera, lo que suscita el paso deH2O hacia los segundos.
6. El HCl y el agua, los principales componentes deljugo g�astrico, se liberan a los conductillosintracelulares, espacios que se continuan con la luzdel estomago.
La presencia de uniones estrechas entre el reves-timiento epitelial de la mucosa g�astrica impide laentrada de HCl desde la luz del estomago a lal�amina propia. Adem�as, el HCO3
� producido porla c�elula parietal se emite hacia la l�amina propia y,por su accion amortiguadora, la resguarda de unafiltracion accidental de HCl desde la luz. Otra posi-ble proteccion procede de la aparicion de prosta-glandinas en zonas cuya barrera epitelial se hayaagrietado de manera fortuita. Las prostaglandinasmultiplican el flujo sanguıneo en ese sector de lal�amina propia, lo que facilita la eliminacion delH+ de la region afectada.
La inhibicion de la fabricacion y secrecion de HClest�a controlada por cuatro hormonas:
. La prostaglandina, el polip�eptido g�astricoinhibidor y la urogastrona actuan directamentesobre las c�elulas parietales, mediante la inhibicionde su actividad secretora.
. La cuarta hormona, la somatostatina, inhibe laliberacion de gastrina por las c�elulas G y dehistamina por las c�elulas de tipo enterocromafın, loque suprime las mol�eculas senalizadoras necesariaspara provocar que las c�elulas parietales elaboren ysegreguen HCl.
Intestino delgado
El intestino delgado (fig. 17.4) mide unos 7m delongitud y suele dividirse en tres regiones. La primeraporcion es el muy corto duodeno, que ocupa unos25 cm; la intermedia corresponde al yeyuno, cuyapared es relativamente gruesa y no llega a los 3m delargo; y la tercera es el ıleon, lam�as estrecha de las tres,la que tiene una pared m�as delgada y se extiende unos4m. El intestino delgado recibe las enzimas digestivasdel p�ancreas y la bilis de la vesıcula biliar, que leayudan a digerir los alimentos en su luz y absorberel agua y los nutrientes generados. Desde el punto devista histologico, las tres regiones son bastante pare-cidas entre sı. Tras una descripcion general de suscaracterısticas en comun, se describen con detalle lasdiferencias que las distinguen.
242
17
APARATO
DIGESTIVO
:TUBODIGESTIVO

243
17
APARATO
DIGESTIVO
:TUBODIGESTIVO
Figura 17.4 Representacion gr�afica del intestino delgado y de los tipos celulares que componen su revestimiento epitelial. (Tomado deGartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 399.)
Figura 17.3 Representacion gr�afica de la produccion de �acido clorhıdrico en las c�elulas parietales.A.C�elula en reposo. B.Mecanismodeliberacion del HCl. C.Numerosas microvellosidades en una c�elula activa. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology,
3rd ed. Philadelphia, Saunders, 2007, p 397.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

CARACTERISTICAS HISTOLOGICAS COMUNES
Con el fin de aumentar la superficie de la luz en elintestino delgado, su submucosa y su mucosa tienen:
. Los pliegues transversales, pliegues circulares(v�alvulas de Kerckring), comienzan en elduodeno y se extienden hasta el ıleon, frenan lavelocidad del quimo y amplıan la superficie aldoble o al triple.
. Las prolongaciones digitiformes de la l�aminapropia, vellosidades, est�an cubiertas por unepitelio cubico simple y dilatan la superficie de laluz 10 veces; estas vellosidades miden 1,5mmde altura en el duodeno, 1mm en el yeyuno y0,5mm en el ıleon; cada una posee un nucleo detejido conjuntivo laxo vascular con un capilarlinf�atico de terminacion ciega, el quilıfero.
. Las c�elulas del epitelio cilındrico simple querecubren cada vellosidad tienen abundantesmicrovellosidades que agrandan la superficie de laluz 20 veces.
. Los espacios intervellosos del intestino delgadopresentan las desembocaduras de las criptas deLieberk€uhn, que multiplican la superficie de la luzpor un factor de 3 o 4.
HISTOLOGIA DEL INTESTINO DELGADO
La mucosa del intestino delgado consta del revesti-miento epitelial, la l�amina propia y la muscular de lamucosa (fig. 17.5). El epitelio cilındrico simple quetapiza la vellosidad est�a compuesto por:
. Las c�elulas absorbentes de la superficie, las m�asnumerosas, funcionan en la ultima fase de ladigestion y en la absorcion de amino�acidos,lıpidos e hidratos de carbono. Estas c�elulasdisponen de 3.000 microvellosidades cubiertasde un glucoc�aliz; el glucoc�aliz abarca sobre todoenterocinasas, aminopeptidasas yoligosacaridasas, enzimas que digierenoligop�eptidos y oligosac�aridos. La cara lateral delas membranas plasm�aticas de estas c�elulasabsorbentes se adhiere a las membranas de lasc�elulas adyacentes creando complejos de union.
. Las c�elulas caliciformes fabrican mucinogeno, unpolisac�arido proteınico complejo que, al entrar encontacto con el agua, se convierte en mucina.Al desprenderse hacia la luz, se mezcla con sucontenido y se transforma en una sustanciaescurridiza llamada moco.
. Las c�elulas del SNED constituyen en torno al 1% delas c�elulas epiteliales de una vellosidad, y cada unaproduce una hormona paracrina o endocrinaespecıfica.
. Las c�elulas con micropliegues (c�elulas M),situadas en el punto de contacto entre los folıculoslinf�aticos de la l�amina propia y el epitelio, poseenunos pliegues profundos: los bolsillosintercelulares. Estas c�elulas fagocitan los antıgenosintraluminales y los traspasan a los linfocitospresentes en sus bolsillos intercelulares, que acontinuacion los entregan a las CPA de la l�aminapropia para poner en marcha una respuestainmunitaria. Una parte de la IgA elaborada porlas c�elulas plasm�aticas sufre una endocitosis por lasc�elulas epiteliales que la enlazan al componentesecretor y liberan el complejo a la luz. La mayorıa sedirige hacia el hıgado; los hepatocitos crean sucomplejo con el componente secretor y lo expulsana la bilis para su transporte hacia la vesıcula biliar(fig. 17.6).
La l�amina propia de la mucosa consiste en untejido conjuntivo laxo con abundantes elementoslinf�aticos y capilares, y, en el centro de la vellosidad,posee quilıferos (v. fig. 17.5). La cara m�as profundade la l�amina propia, entre la base de las vellosidadesy la muscular de la mucosa, est�a bastante vasculari-zada, aunque fundamentalmente queda desplazadapor la profusion de gl�andulas intraparietales intesti-nales, las criptas de Lieberk€uhn. Estas gl�andulas seextienden desde los espacios intervellosos hasta lamuscular de la mucosa, y su epitelio consta de losmismos tipos celulares que los que cubren la vello-sidad, y adem�as:
. Las c�elulas regenerativas proliferan para formarnuevas c�elulas del revestimiento epitelial. Tras ello,emigran a lo largo de la l�amina basal hasta la puntade la vellosidad, donde se desprenden a la luz entre5 y 7 dıas despu�es de su aparicion; el revestimientoepitelial del intestino delgado se renueva una vez ala semana.
. Las c�elulas de Paneth viven m�as tiempo (20 dıas);est�an situadas en la base de las criptas de Lieberk€uhny alojan grandes gr�anulos eosinofilos que contienenlisozima y defensinas, productos antimicrobianosy el factor de necrosis tumoral-a.
La muscular de la mucosa est�a integrada por unacapa circular interna y otra longitudinal externa demusculo liso. Algunos miocitos lisos sueltos penetranen el nucleo de la vellosidad y llegan hasta su extremo.La submucosa del intestino delgado no tiene nadaespecial, y su muscular externa est�a compuesta poruna capa circular interna y otra longitudinal externa.Los plexos de Meissner y de Auerbach ocupan susposiciones normales. La capa m�as externa es unaserosa, salvo en ciertas zonas del duodeno, donde setrata de una adventicia.
244
17
APARATO
DIGESTIVO
:TUBODIGESTIVO

CONSIDERACIONES CL�INICAS
Las c�elulas de Paneth elaboran defensinas comoreaccion al factor de necrosis tumoral-a y a raız dela presencia de subproductos metabolicosliberados por microorganismos. Al introducirse enlas membranas fosfolipıdicas de estos ultimos, lasdefensinas forman canales ionicos que dejan saliriones desde su interior, lo que destruye al agenteinvasor. La lisozima, una enzima proteolıtica
tambi�en fabricada por las c�elulas de Paneth,degrada un componente de la membranabacteriana, el peptidoglucano, lo que acaba con elmicroorganismo.
245
17
APARATO
DIGESTIVO
:TUBODIGESTIVO
Figura 17.5 Representacion gr�afica del intestino delgado y de los tipos celulares que componen su revestimiento epitelial. (Tomado de
Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 399.)
Figura 17.6 Representacion gr�afica de la funcion que cumplen
las c�elulas M y la circulacion de la IgA en el intestino delgado.
(Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd
ed. Philadelphia, Saunders, 2007, p 405.)�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

246
17
APARATO
DIGESTIVO
:TUBODIGESTIVO
Diferencias regionales en la histologıa del intestinodelgado
Las tres regiones del intestino delgado pueden distin-guirse entre sı mediante unas variaciones mınimas desu aspecto histologico.
. Las vellosidades son m�as altas en el duodeno(1,5mm), que en el yeyuno (1mm) y que en elıleon (0,5mm).
. La principal diferencia es la existencia de lasgl�andulas de Brunner en la submucosa del duodeno.
. Las placas de Peyer, que son grupos de folıculoslinf�aticos, est�an presentes en la l�amina propia delıleon. El yeyuno no tiene ni gl�andulas en susubmucosa, ni placas de Peyer en su l�amina propia.
El duodeno recibe el conducto col�edoco y el con-ducto pancre�atico en la papila duodenal (ampollade Vater). Las gl�andulas de Brunner (gl�andulas duo-denales) son tubuloalveolares, est�an ramificadas ydesembocan en la base de las criptas de Lieberk€uhn.Su actividad produce un lıquido seromucoso cargadode bicarbonatos, que sirve para neutralizar el quimo�acido procedente del estomago. Estas gl�andulastambi�en fabrican la hormona urogastrona (factorde crecimiento epid�ermico humano), que inhibe laelaboracion de HCl en las c�elulas parietales y estimulalos procesos mitoticos de las c�elulas epiteliales.
HISTOFISIOLOG�IA DEL INTESTINO DELGADO
Lamotilidad del intestino delgado tiene dos fases: con-tracciones de mezcla, que son fenomenos circuns-critos destinados a dejar expuesto el contenido de laluz intestinal frente a su revestimiento epitelial, y con-tracciones de propulsion (ondas perist�alticas), quelo desplazan a lo largo de su recorrido. Este movi-miento es lento, m�as o menos de 1 a 2 cm/min, y elquimo llegado al duodeno desde el estomago pasa de6 a 12h en el intestino delgado. Las contracciones depropulsion est�an controladas por el plexo de Auer-bach y por las hormonas derivadas de las SNED cole-cistocinina, gastrina, motilina, sustancia P y seroto-nina, que aumentan la motilidad, y secretina y gluca-gon, que la frenan (v. tabla 17.1).
. Las gl�andulas del intestino delgado segregan 2 l deuna secrecion seromucosa al dıa. Este proceso seencuentra b�asicamente dominado por el plexosubmucoso de Meissner, aunque tambi�en sufre lainfluencia de las hormonas secretina ycolecistocinina, originadas en las SNED.
. La digestion de las sustancias nutritivas que ocupanla luz del intestino delgado se debe a la presencia deenzimas, llegadas sobre todo desde el p�ancreas y labilis hep�atica, que se concentra y expulsa a partir dela vesıcula biliar. El mecanismo de la digestioncomienza en la cavidad oral y en el estomago, y
sigue en el duodeno, donde las enzimaspancre�aticas degradan los diversos componentesdel quimo en oligop�eptidos y dip�eptidos, m�asoligosac�aridos y disac�aridos. Las enterocinasas yaminopeptidasas, situadas en el glucoc�aliz de lasmicrovellosidades, completan la digestion delas proteınas en amino�acidos y en dip�eptidos ytrip�eptidos, que acaban captados por las c�elulasabsorbentes de la superficie para transformarlosen amino�acidos mediante las peptidasascitopl�asmicas. El glucoc�aliz tambi�en contieneoligosacaridasas (lactasa, maltasa, sacarasa ya-dextrinasa) que concluyen la digestion de loshidratos de carbono de la alimentacion aldescomponer los oligosac�aridos en monosac�aridospara su incorporacion a trav�es de las c�elulasabsorbentes de la superficie. Las sales biliaresemulsionan en micelas los lıpidos existentes en elquimo, y la lipasa pancre�atica los divide en �acidosgrasos ymonoglic�eridos, que difunden a trav�es de lamembrana plasm�atica de las microvellosidades.
. La absorcion (fig. 17.7) de los productos finales dela digestion y los electrolitos y el agua tiene lugar demanera predominante en el intestino delgado y elgrueso, aunque determinadas sustancias, como elalcohol, tambi�en se absorben en el estomago. Todoslos dıas llegan enormes cantidades a las c�elulasabsorbentes de la superficie en el intestino delgado,contando 1 kg de lıpidos, 0,5 kg de proteınas ehidratos de carbono, unos 35 g de sodio y 7 l delıquido. Los amino�acidos y los monoglic�eridospasan al centro de la vellosidad, penetran en losafluentes de la vena porta hep�atica y van hacia elhıgado para cumplir su accion posterior. Losmonoglic�eridos y los �acidos grasos de cadena largase unen a las proteınas intracelulares ligadoras de�acidos grasos, y se introducen en el retıculoendopl�asmico liso para su esterificacion entriglic�eridos por la acil CoA-sintetasa y lasaciltransferasas. Estos triglic�eridos, ensambladoscon proteınas, se transportan hacia la membranabasolateral para su liberacion hacia el nucleo de lavellosidad como quilomicrones que se incorporana los quilıferos, hasta dejarlos repletos de un lıquidocargado de lıpidos, el quilo. El quilıfero se vacıa porlas contracciones rıtmicas de las tiras de musculoliso derivadas de la capa circular interna de lamuscular de lamucosa, lo que expulsa el quilo haciael plexo linf�atico submucoso. A continuacion, setransporta hacia el conducto tor�acico para verterseen la union entre la vena subclavia y la vena yugularinterna izquierdas. Los �acidos grasos que no lleguena tener 12 carbonos de longitud eluden el procesode reesterificacion; en su lugar, se dirigen a lamembrana plasm�atica basolateral para salir a laparte central de la vellosidad y viajar hasta la venaporta hep�atica y, por ella, hacia el hıgado.

247
17
APARATO
DIGESTIVO
:TUBODIGESTIVO
Figura 17.7 Representacion gr�afica de la absorcion de las grasas en las c�elulas absorbentes de la superficie del intestino delgado.(Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 407.)
Tabla 17.1 CELULAS DEL SISTEMA NEUROENDOCRINO DIFUSO EN EL TUBO DIGESTIVO
C�elula Localizacion Hormona producida Funcion
A Estomago, intestino delgado Glucagon Elevacion de la glucemiaD Estomago, intestinos Somatostatina Inhibicion de la liberacion hormonal en las c�elulas
del SNED
EC Estomago, intestinos Serotonina, sustancia P Aumento de los movimientos perist�alticos
ECL Estomago Histamina Estimulacion de la secrecion de HClG Estomago, intestino delgado Gastrina Estimulacion de la secrecion de HCl; motilidad g�astrica
GL Estomago, intestinos Glicentina Elevacion de la glucemia
I Intestino delgado Colecistocinina Estimulacion de la liberacion de enzimas pancre�aticas y
contraccion de la vesıcula biliarK Intestino delgado Polip�eptido g�astrico
inhibidor
Inhibicion de la secrecion de HCl
Mo Intestino delgado Motilina Aumento del peristaltismo intestinalN Intestino delgado Neurotensina Disminucion del peristaltismo intestinal
PP Estomago, intestino grueso Polip�eptido pancre�atico Estimulacion de la liberacion enzim�atica en las c�elulas
principales; inhibicion de la liberacion de enzimas
pancre�aticasS Intestino delgado Secretina Estimulacion de la liberacion del amortiguador
pancre�atico
VIP Estomago, intestinos P�eptido intestinal
vasoactivo
Aumento del peristaltismo intestinal; estimulacion de
la eliminacion de agua e iones
EC, celula enterocromafın; ECL, celula de tipo enterocromafın; G, celula productora de gastrina; GL, celula productora de glicentina;
HCl, �acido clorhıdrico;MO, celula productora demotilina; N, celula productora de neurotensina; PP, celula productora de polipeptido
pancre�atico; VIP, celula productora de peptido intestinal vasoactivo.Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 392.�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Intestino grueso
El intestino grueso, que mide alrededor de 1,5m delargo, abarca el ciego, el ap�endice, el colon, el recto yel ano. Desde el punto de vista histologico, salvo elap�endice y el ano, estas regiones son muy parecidas yse denominan colon. El colon actua en la absorcionde agua, electrolitos y gases, y en la compactaciondel quimo que recibe del ıleon para generar lasheces.
El colon se asemeja al intestino delgado, exceptoque su di�ametro es mayor y que no tiene vellosidades.
. Las criptas de Lieberk€uhn recuerdan a susequivalentes del intestino delgado, pero carecen dec�elulas de Paneth, poseen pocas c�elulas del SNED ypresentan una cantidad m�as alta de c�elulascaliciformes (fig. 17.8). Aunque a primera vistapodrıa parecer que la mayorıa de las c�elulasepiteliales son caliciformes, en realidad m�as de lamitad son c�elulas absorbentes de la superficie. Aligual que en otras regiones del tubo digestivo, surevestimiento epitelial se renueva por completocomo mınimo una vez a la semana gracias a laactividad mitotica de las c�elulas regenerativas.
. La l�amina propia, la muscular de la mucosa y lasubmucosa no ofrecen nada nuevo.
. La muscular externa est�a modificada en el sentidode que gran parte de las fibras musculares lisas de lacapa longitudinal externa se reunen en tres bandasfinas de musculo, las tenias del colon, quemantienen un tono casi constante, lo que las vuelvem�as cortas que el propio colon.
. El colon forma una secuencia lineal de bolsas,llamadas haustras del colon, a lo largo de sulongitud.
. Todo el colon est�a tapizado por una serosamenos elano, donde se fija a la pared corporal mediante unaadventicia de tejido conjuntivo. Durante elrecorrido del colon, la serosa crea unasevaginaciones llenas de grasa, conocidas comoap�endices epiploicos.
La funcion del colon es la secrecion de un mococon gran cantidad de bicarbonato; tambi�en absorbem�as lıquido y electrolitos del contenido intestinal, loque determina la compactacion de las heces. Cadadıa, el colon recupera en torno a 1,4 l de lıquidosque llevan electrolitos y reduce el volumen diario delas heces m�as o menos a 100ml. El colon absorbeaproximadamente de 6 a 9 l de gases diarios, y noexpulsa m�as que unos 0,5 a 1 l como flato. Las criptasde Lieberk€uhn del recto siguen una distribucion pocodensa y m�as profunda que en el colon; por lo dem�as,este tramo guarda un gran parecido con el colon.
. El conducto analmide de 3 a 4 cm, es m�as estrechoque el recto, y en su mitad inferior ni siquiera poseelas criptas de Lieberk€uhn poco profundas que hayen su mitad superior.
. La mucosa anal presenta unos pliegueslongitudinales, las columnas anales (deMorgagni),que convergen en la lınea pectınea para formar lasv�alvulas anales que albergan los senos analessimilares a bolsas.
. El conducto anal est�a tapizado por un epiteliocubico simple que se convierte en pavimentosoestratificado no queratinizado pasada la lıneapectınea. La l�amina propia fibroel�astica aloja lasgl�andulas perianales en el ano; a este nivel existenfolıculos pilosos y sus gl�andulas seb�aceasacompanantes.
. La muscular de la mucosa est�a representada,pero no se extiende m�as all�a de la lınea pectınea.El tejido conjuntivo fibroel�astico de lasubmucosa del conducto anal tiene un plexovenoso hemorroidal interno por encima de lalınea pectınea y un plexo venoso hemorroidalexterno por debajo, justo antes de llegar alorificio anal.
. Lamuscular externa no ofrece novedades salvo quela capa circular interna est�a engrosada en la lıneapectınea para originar el musculo esfınter internodel ano, y la capa longitudinal externa se vesustituida por una membrana fibroel�astica querodea al esfınter interno del ano. El musculoesfınter externo del ano est�a integrado por unosespesamientos del musculo estriado que hay en elsuelo de la pelvis y envuelve al esfınter interno delano y a la vaina fibroel�astica. Este esfınter demusculo estriado permite el control voluntariosobre el ano.
. La capa m�as externa del colon es una serosa.
El ap�endice es una estrecha evaginacion del ciegoque mide de 5 a 6 cm de longitud y cuya luz estrelladase encuentra revestida por un epitelio cilındrico sim-ple compuesto por c�elulas absorbentes de la super-ficie, c�elulas caliciformes y c�elulas M, que linda conlos folıculos linf�aticos de la l�amina propia. Las criptasde Lieberk€uhn son escasas y poco profundas, y est�anconstituidas por c�elulas absorbentes de la superficie,c�elulas caliciformes, c�elulas regenerativas, numerosasc�elulas del SNED y c�elulas de Paneth aisladas. Lal�amina propia corresponde a un tejido conjuntivolaxo dotado de abundantes linfocitos y folıculoslinf�aticos. La muscular de la mucosa, la submucosay la muscular externa siguen la misma organizaciongeneral del tubo digestivo. La capa m�as externa delap�endice es una serosa.
248
17
APARATO
DIGESTIVO
:TUBODIGESTIVO

CONSIDERACIONES CL�INICAS
La enfermedad de Crohn consiste en unainflamacion cronica de la pared del tubo digestivo
que afecta m�as a menudo al ıleon y al colon. Sepiensa que es un trastorno de naturaleza inmunitaria
que puede tener componentes gen�eticos yambientales. Sigue una distribucion equivalenteentre los hombres y las mujeres, con una primeraaparicion normalmente entre los 15 y los 25 anos,
pero casi siempre antes de los 35. Los sıntomasm�ashabituales son fiebre, diarrea cronica acompanadapor una posible hemorragia, colicos de diversagravedad, adelgazamiento e inapetencia. Suduracion puede ser de varias semanas y a
continuacion resolverse de manera espont�anea,para acabar por recidivar al cabo de diversosintervalos indeterminados con un grado variablede intensidad. El lugar de la inflamacion puede ser elmismo que antes, o propagarse hacia otras regiones
del tubo digestivo. Sus complicaciones m�ascorrientes son la creacion de fıstulas y la obstrucciondel propio tubo digestivo, y en el colon llega a dar
lugar a un c�ancer colorrectal. Aunque la enfermedadde Crohn es incurable, sus tratamientos paliativosconsisten en el empleo de astringentes yantiinflamatorios, antibioticos, inmunomoduladores,
modificaciones diet�eticas y, en caso de que sea
necesario, la reseccion quirurgica de las �areas encuestion.
Las personas con colera han tomado agua o comidainfectados por el Vibrio cholerae, unmicroorganismo
que produce la toxina col�erica. Aunque sea una
enfermedad de muy f�acil tratamiento, est�a difundidapor las regiones tropicales y subtropicales de paısesen desarrollo, donde es la responsable de una granmortalidad. Su prevencion se logra si se recurre a las
condiciones higi�enicas oportunas y se trata con
antibioticos y con una administracion r�apida delıquidos que guarden un equilibrio electrolıtico para
reponer las p�erdidas de lıquidos y electrolitos, hastade 10 l/dıa debido a una diarrea incontenible.
El gas intestinal es un subproducto del
metabolismo bacteriano m�as aire deglutido. Loscomponentes odorıferos de las heces son losmercaptanos, el indol y el sulfuro de hidrogeno, y eldel flato es el metano. Como el gas metano semezcla con O2, CO2 y H2, resulta bastanteinflamable, y a veces durante una cauterizacion en elcurso de una sigmoidoscopia, pueden producirsepequenas explosiones limitadas.
249
17
APARATO
DIGESTIVO
:TUBODIGESTIVOFigura 17.8 Representacion gr�afica del colon y de los tipos celulares que integran su revestimiento epitelial. (Tomado de Gartner LP,
Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 408.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

18 APARATO DIGESTIVO:GLANDULAS
Las gl�andulas del aparato digestivo est�an situadas enla pared del tubo digestivo, en el caso de las gl�andulasintraparietales, y fuera de ella, en el delas gl�andulas extraparietales, como lasgl�andulas salivales mayores (gl�andulasparotida, submandibular y sublingual),el p�ancreas y el hıgado (m�as la vesıculabiliar), cuyas secreciones llegan hasta laluz del tubo digestivo a trav�es de unsistema de conductos.
Gl�andulas salivales mayores
Las gl�andulas salivales mayores(fig. 18.1), la parotida, la submandibular y la sublin-gual, son gl�andulas tubuloalveolares compuestas quesegregan saliva.
. Cada gl�andula salival mayor est�a rodeada por unac�apsula de tejido conjuntivo que manda tabiquesde este mismo tejido hacia el par�enquima glandularpara dividirlo en lobulos y lobulillos.
. Los elementos neurovasculares recorren estostabiques de tejido conjuntivo para irrigar elpar�enquima de la gl�andula. El par�enquima es laporcion secretora, que consta de �acinos, tubulos oambos, y una porcion de conductos que culmina enel conducto principal de la gl�andula.
. La unidad funcional de una gl�andula salival, elsalivon, est�a compuesta de un �acino y susconductos intercalares y estriados.
Tres tipos de c�elulas forman la porcion secretora deuna gl�andula salival: serosas, mucosas y mioepiteliales.
. Las c�elulas serosas se parecen a una pir�amidetruncada, y generan un lıquido acuoso constituidofundamentalmente por agua, electrolitos y enzimas(amilasa y lipasa salival) que comienza la digestionen la cavidad oral. Adem�as, sus secreciones llevancalicreına y los productos antibacterianos lisozima ylactoferrina. El almacenamiento tiene lugar en unosgr�anulos de cimogeno de situacion apical (gr�anulossecretores) hasta que se provoque su liberacion.
. Las c�elulas mucosas son semejantes a las c�elulasserosas, pero su citoplasma apical alberga gr�anulossecretores llenos de mucinogeno, unproteoglucano que, al expulsarse, se hidrata para
formarmucina. Cuando la mucina se mezcla con lasecrecion, se convierte en moco.
. Loa �acinos est�an integradosexclusivamente por c�elulas serosas,c�elulas mucosas o c�elulas mucosaspero coronadas por unas cuantasc�elulas serosas que dan lugar asemilunas serosas. Cada �acinoest�a envuelto por una l�aminabasal, y c�elulas mioepitelialescuya contraccion ayuda a emitir elproducto secretor del �acino haciala luz para incorporarse a losconductos.
Los conductos de las gl�andulas salivales nacencomo unos tubos muy estrechos, tapizados por un epi-telio cubico simple, llamados conductos intercalares.
. La secrecion que penetra en estos conductos y esisotonica respecto a la sangre se denomina salivaprimaria. Los conductos estriados, m�as amplios yrevestidos por c�elulas epiteliales cilındricas bajas,reciben la saliva primaria.
. Las membranas plasm�aticas basales de estas c�elulasexhiben numerosos pliegues cargados demitocondrias que sacan Na+ de la luz del conductopor transporte activo ymetenK+ yHCO3
� en ella, loque modifica la saliva primaria en una salivasecundaria hipotonica. Varios conductos estriadosse reunen para originar los conductosintralobulillares, que confluyen en el inicio de losconductos excretores, que son m�as grandes.
. El conducto excretor principal que deja salir lasaliva hacia la cavidad oral suele estar recubiertopor un epitelio cubico estratificado oseudoestratificado.
Las c�elulas plasm�aticas del estroma de tejido con-juntivo elaboran dımeros de IgA que se mantienenunidos entre sı por una cadena J. Las c�elulas acinares ylas c�elulas de los conductos estriados captan estosdımeros y allı se anade el componente secretor, loque genera la IgA secretora que pasa a la luz del �acinoo del conducto estriado.
Al rev�es que las gl�andulas salivales menores, lasmayores segregan segun las necesidades, y su produc-cion est�a controlada por todo el proceso de oler elalimento, masticar y vomitar. La fabricacion de saliva
� 2011 Elsevier Espana, S.L. Reservados todos los derechos
TERMINOS CLAVE
. Gl�andulas salivales
. P�ancreas exocrino
. P�ancreas endocrino
. Hıgado
. Lobulillos hep�aticos
. Hepatocitos
. Vesıcula biliar
250

251
18
APARATO
DIGESTIVO
:GLANDULAS
est�a en torno a 1 l/dıa y baja con el miedo y el cansan-cio, ası como durante el sueno. La inervacion para-simp�atica provoca el flujo de una saliva eminente-mente acuosa, mientras que la simp�atica causa laemision de una saliva m�as viscosa.
De las gl�andulas salivalesmayores, la gl�andulaparo-tida fabrica una secrecion serosa, alrededor del 30% dela saliva, mientras que la gl�andula submandibular seencarga del 60% y la gl�andula sublingual solo del 5%.Estas dos ultimas gl�andulas liberan una saliva mixta.
Figura 18.1 Descripcion general de una gl�andula salival
mayor. (Tomado de Gartner LP, Hiatt JL: Color Textbook
of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 414.)
CONSIDERACIONES CL�INICAS
El flujo de saliva resulta fundamental para laconservacion de la salud oral porque su accionlimpia los dientes, mantiene humeda la mucosa oraly ofrece la primera lınea de defensa frente a losmicroorganismos invasores. Asimismo, al empaparlos alimentos, permite la formacion de un bolocompacto, pero flexible y deslizante, que puededeglutirse con facilidad. El caudal de saliva es normalen un estado de relajacion; sin embargo, cuando una
persona est�a asustada o nerviosa, la boca se seca.
Esta situacion era de sobra conocida en la �epoca dela Inquisicion, y los tribunales la utilizaron a la hora desus procesamientos. Se daba harina al acusado paraque la tragase presuponiendo que si alguien no eraculpable, no se pondrıa nervioso ni asustado y serıacapaz de elaborar la suficiente cantidad de salivapara hacerlo. Naturalmente, el acusado siempreestaba nervioso y asustado, y por la escasa cantidadde saliva no conseguıa tragar la harina, lo que setomaba como una prueba irrefutable de suculpabilidad de acuerdo con la denuncia.
La parotiditis es una virosis cuya frecuencia essuperior entre los ninos de 5 a 15 anos. Setransmite por el contenido vırico de las gotitas de
saliva que salen en suspension a�erea despu�esde que una persona infectada tosa cerca de otrasana o de que esta entre en contacto con unobjeto sobre el que haya ido a parar saliva dealguien infectado. El perıodo de incubacion de laparotiditis es de 2 a 3 semanas, pasadas lascuales el paciente entra en letargo, tiene cefalea y
sufre inapetencia. El sıntoma m�as frecuente es la
hinchazon dolorosa de las gl�andulas parotidas,acompanada por una fiebre de 39� a 40�. Lamayor parte de los ninos de EE. UU. est�anvacunados contra la parotiditis, por lo que suincidencia es muy baja. Este proceso es mucho
m�as grave en los varones por la posibilidad de quese propague a un testıculo o a ambos, dondepuede ocasionar esterilidad, o a las meninges y al
p�ancreas, pero la infeccion suele resolverse de
manera espont�anea.
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

252
18
APARATO
DIGESTIVO
:GLANDULAS
P�ancreas
El p�ancreas es una gl�andula que mide 25 cm de lon-gitud, pesa unos 150 g y posee un componente exo-crino y otro endocrino. Su fr�agil c�apsula de tejidoconjuntivomanda tabiques hacia el par�enquima glan-dular, que no solo lo subdivide en lobulos y lobuli-llos, sino que tambi�en transportan todo un sistema deconductos y elementos neurovasculares para abastecereste organo. La porcion exocrina ocupa la mayorparte del organo, y el componente endocrino,los islotes de Langerhans, est�a distribuido formandounos conglomerados esf�ericos muy vascularizados dec�elulas endocrinas entre los �acinos secretores(fig. 18.2).
El p�ancreas exocrino, compuesto por unidadestubuloacinares m�as sus sistemas de conductos corres-pondientes, elabora y libera:
. En torno a 1,2 l de un lıquido amortiguado que est�aconcebido para neutralizar el quimo �acidoexpulsado por el estomago hacia el duodeno
. Proenzimas que se activan al llegar a la luz delduodeno para degradar el quimo cargado denutrientes.
Cada �acino est�a integrado por 40 o 50 c�elulasacinares. La luz del �acino aloja unas cuantas c�elulascentroacinares, los constituyentes iniciales del sis-tema de conductos pancre�aticos. La presencia dec�elulas centroacinares es caracterıstica del p�ancreas.
. Las c�elulas acinares parecen pir�amides truncadascuyo v�ertice est�a repleto de gr�anulos de cimogenoque contienen proenzimas. La membranaplasm�atica basal de cada c�elula acinar poseereceptores para la hormona colecistocinina y elneurotransmisor acetilcolina.
. Las c�elulas centroacinares de cada �acino secomunican con los conductos intercalares,muchos de los cuales se unen entre sı y dan origena los conductos intralobulillares, que a su vezconfluyen para generar unos conductosinterlobulillares y de mayor tamano, quefinalmente desembocan en el conductopancre�atico principal. El conducto col�edoco de lavesıcula biliar y el conducto pancre�atico principalse juntan entre sı para perforar la pared delduodeno, formando la ampolla de Vater(papila duodenal).
Las c�elulas acinares actuan en la sıntesis de proen-zimas y enzimas digestivas que se almacenan y liberantras desencadenarse el proceso por la union de la acetil-colina procedente de las fibras posganglionares para-simp�aticas sumada a la colecistocinina derivada de las
c�elulas del sistema neuroendocrino difuso (SNED)del duodeno.
. Las enzimas expulsadas por las c�elulas acinarespancre�aticas son ribonucleasa, desoxirribonucleasa,lipasa y amilasa; y las proenzimas son elastasa,quimotripsinogeno, tripsinogeno yprocarboxipeptidasa.
. Las c�elulas acinares se protegen mediante lasıntesis del inhibidor de la tripsina para impedirla activacion del tripsinogeno en el interior delcitosol.
. Las c�elulas de los conductos intercalares y lasc�elulas centroacinares emiten el amortiguador debicarbonato ante la union a los receptores de susmembranas plasm�aticas basales de la acetilcolinasurgida en las fibras parasimp�aticasposganglionares y de la secretina producida porlas c�elulas del SNED en el duodeno. Elbicarbonato se fabrica en las c�elulas de losconductos estriados al combinarse CO2 con H2O,que generan H2CO3. Esta mol�ecula se disocia enH+ y HCO3
�. El bicarbonato se desprende haciala luz del conducto a la vez que el Na+ paramantener la neutralidad. El H+ se dirige hacia eltejido conjuntivo para penetrar en el lecho capilarperiacinar.
. Como elmecanismo de liberacion de las enzimas yel amortiguador depende de mol�eculassenalizadoras distintas, su salida tiene lugar demodo independiente, aunque a veces seasimult�anea.
PANCREAS ENDOCRINO
El p�ancreas endocrino est�a compuesto m�as o menospor un millon de islotes de Langerhans, cada unoencerrado en una fina vaina fibrosa reticular quemanda fibras hacia �el como medio de soporte parael sistema porta insuloacinar, su abundante irriga-cion vascular particular. Las venas que abandonantodos los islotes vagan entre los �acinos vecinos y llevanmol�eculas senalizadoras expulsadas por las c�elulas delos islotes para regular el funcionamiento de estos�acinos.
Cinco tipos celulares constituyen las 3.000 c�elulaspocom�as omenos de cada islote de Langerhans. Cadauno elabora una hormona concreta: las c�elulas a seencargan del glucagon, las c�elulas b sintetizan insu-lina, las c�elulas d elaboran somatostatina, las c�elulasPP fabrican polip�eptido pancre�atico y las c�elulas Gproducen gastrina. En la tabla 18.1 se recogen lasproporciones de estas c�elulas en los islotes de Langer-hans, las hormonas que fabrican y las funciones quecumplen estas ultimas.

253
18
APARATO
DIGESTIVO
:GLANDULAS
CONSIDERACIONES CL�INICAS
La diabetes mellitus es un proceso en el que la
glucemia est�a por encima de lo normal. Existen dostipos de diabetes mellitus: tipo 1, que comienza en lajuventud porque el paciente no elabora la suficiente
cantidad de insulina, y tipo 2, que se iniciam�as tarde alo largo de la vida, cuando el paciente sı produce esacantidad, pero el organismo se vuelve resistente a susefectos. La principal causa de la diabetes mellitus detipo 2 es la obesidad, aunque el ascenso continuo delas concentraciones de corticoesteroides y lapancreatitis son otros factores que influyen en laaparicion de esta enfermedad. Con el crecienteaumentode la incidencia de laobesidadentre los ninosy los adultos en EE. UU., ha subido la incidencia de ladiabetes mellitus de tipo 2. En un primer momento, no
existen sıntomas patentes de la diabetes de tipo 2,pero pasados unos anos de vida con esta alteracionempiezan a notarse. El cuadro consta de unincremento de la diuresis, sensacion de sed,cansancio, deshidratacion, mareo, confusion, vistaborrosa y convulsiones. A estas alturas es cuando el
paciente suele acudir al m�edico, y los an�alisis desangre muestran una glucemia muy alta. Las secuelasa largo plazo pueden ser muy importantes, como losproblemas circulatorios, hipertension arterial, lesionescardıacas, gangrenade las extremidades, insuficienciarenal y ceguera. El tratamiento consiste en el descensode la glucemia, que en la diabetes de tipo 2 puede
llevarse a cabomediante la dieta y el ejercicio, adem�asde medicamentos en algunos casos.
Tabla 18.1 CELULAS Y HORMONAS DE LOS ISLOTES DE LANGERHANS
C�elulas Frecuencia Hormona producida Funcion de la hormona
C�elula b 70% Insulina Disminucion de la glucemiaC�elula a 20% Glucagon Aumento de la glucemia
C�elula d 5%
C�elula D Somatostatina Inhibicion de la liberacion de hormonas y productos exocrinos
del p�ancreasC�elula D1 P�eptido intestinal vasoactivo Induccion de la glucogenolisis; regulacion de la motilidad intestinal;
control de la secrecion de iones y H2O por los intestinos
C�elula G 2-3% Gastrina Estimulacion de la produccion de HCl por las c�elulas parietales
del estomagoC�elula PP 2-3% Polip�eptido pancre�atico Inhibicion de la secrecion exocrina por el p�ancreas
Modificado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 421.
Figura 18.2 El p�ancreas con la representacion de sus unidades tubuloacinares y su sistema de conductos, ası como sus componentes
endocrinos, los islotes de Langerhans. RE, retıculo endopl�asmico. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology,
3rd ed. Philadelphia, Saunders, 2007, p 418.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

254
18
APARATO
DIGESTIVO
:GLANDULAS
Hıgado
Las c�elulas parenquimatosas del hıgado, la gl�andulam�as grande del organismo, son los hepatocitos, queelaboran la secrecion exocrina –bilis– y forman unamultitud de productos endocrinos que difunden haciala sangre. Casi todo el organo est�a revestido por elperitoneo, bajo el cual hay un tejido conjuntivofibroel�astico d�ebilmente adherido que se llamac�apsula de Glisson. Los componentes del tejido con-juntivo, surgidos de la c�apsula, penetran en el par�en-quima del hıgado a nivel del hilio hep�atico y llevan osacan elementos vasculares, linf�aticos y biliares de suinterior. Las arterias hep�aticas derecha e izquierda apor-tan alrededor del 25%del oxıgeno que recibe el hıgado,mientras que el 75% restante llega a partir de la sangrecargada de nutrientes que conduce la vena portahep�atica, procedente de todo el tubo digestivo y delbazo. Las venas hep�aticas extraen sangre del hıgadodesde su parte posterior, no en el hilio hep�atico, paravaciarla en la vena cava inferior (fig. 18.3).
LOBULILLO HEPATICO CLASICO
El hıgado actua como un deposito central, al recibirsangre que transporta todos los nutrientes absorbidospor el tubo digestivo, salvo los quilomicrones.Tambi�en le llega sangre desde el bazo que porta hierroy productos de degradacion de los globulos rojos viejosdestruidos por dicho organo. Los hepatocitos no soloprocesan estos nutrientes, los almacenan y los convier-ten en compuestos utilizables por las c�elulas del orga-nismo, sino que tambi�en eliminan sustancias toxicas.
. El hıgado est�a organizado en unas figurashexagonales muy vascularizadas, los lobulilloscl�asicos, que miden 2mm de altura y menos de1mm en sentido transversal (v. fig. 18.3). Enalgunos animales, se encuentran delimitados portejido conjuntivo, pero en el hombre su grosor esdemasiado fino como para poder determinar unosbordes con claridad.
. Sin embargo, estos ingredientes de tejido conjuntivoganan espesor, incluso en el hıgado humano, a nivelde la union entre tres lobulillos cl�asicos en unespacio (trıada) portal, que alberga ramas delgadasde la vena porta, la arteria hep�atica, el conductobiliar interlobulillar y un vaso linf�atico (v. fig. 18.3).
. Unicamente tres de los espacios portales asociados alos seis lados longitudinales del lobulillo cl�asico sehallan perfectamente determinados. Est�andispuestos de tal manera que ocupan lados alternosde cada lobulillo.
. Un tabique limitante cilındrico, integrado porhepatocitos modificados, rodea a cada espacioportal pero est�a separado del tejido conjuntivo porel espacio de Moll.
. Cada rama de la arteria hep�atica da origen en elespacio portal a numerosas ramasm�as pequenas, lasarteriolas de distribucion, que parecen las patas deun ciempi�es al envolver las paredes adyacentes dellobulillo hexagonal, hasta alcanzar las arteriolas dedistribucion correspondientes al espacio portalvecino.
. Las arteriolas de entrada todavıa m�as pequenasnacen de las arteriolas de distribucion para abastecerel par�enquima de cada lobulillo cl�asico. Las ramas dela vena porta emulan a las de la arteria hep�atica, yforman venas de distribucion y v�enulas de entrada.
. Los conductos biliares interlobulillares est�an irrigadospor el plexo capilar peribiliar. La bilis, lanzada hacialos conductos biliares, se transfiere a la vesıcula biliarpara su acumulacion y futura liberacion.
El centro de cada lobulillo cl�asico presenta una venacentral de disposicion longitudinal, el nacimiento dela vena hep�atica (v. fig. 18.3). Desde ella irradian capasanastomosadas de c�elulas hep�aticas, generando unoscanales vasculares abiertos entre ellas, que reciben elnombre de sinusoides hep�aticos, y desembocan en lavena central. Las arteriolas y las v�enulas de entradavacıan su sangre en los sinusoides hep�aticos y a con-tinuacion en la vena central. Al salir del lobulillo, lavena central vierte en la vena sublobulillar, que recibemuchas venas centrales m�as procedentes de otros lobu-lillos cl�asicos. Las venas sublobulillares confluyen paracrear las venas colectoras que m�as adelante constitu-yen la vena hep�atica, vaso que extrae la sangre del hıga-do y la dirige hacia la vena cava inferior.
TRES CONCEPTOS SOBRE LOS LOBULILLOS HEPATICOS
. Segun se senalo antes, el lobulillo hep�atico cl�asicoconsiste en una figura hexagonal en la que la sangrecircula desde su periferia hacia el centro, y la bilis lohace en sentido opuesto (fig. 18.4).
. Por regla general, el flujo de los productos desecrecion exocrina va hacia el centro de un lobulillo:de ahı que se haya ideado otro lobulillo en el que, enuna imagen bidimensional del hıgado, se concibieraque tres venas centrales contiguas ocuparan losv�ertices de un lobulillo y el espacio portal pasara asıa ser su centro, lo que se denomina lobulillo portal,donde la bilis avanza hacia el centro, tal como cabrıaesperar de una gl�andula exocrina.
. El tercer modelo est�a basado en el movimiento de lasangre. Se observan tres zonas romboidales yconc�entricas de hepatocitos, lo que se llama el �acinode Rappaport: las m�as proximas a la vena centralcorresponden a la zona 3, las m�as cercanas a laperiferia del �acino de Rappaport son la zona 1, y lasque quedan en la region entre ambas son la zona 2.Los lımites del �acino est�an trazadosmediante cuatro
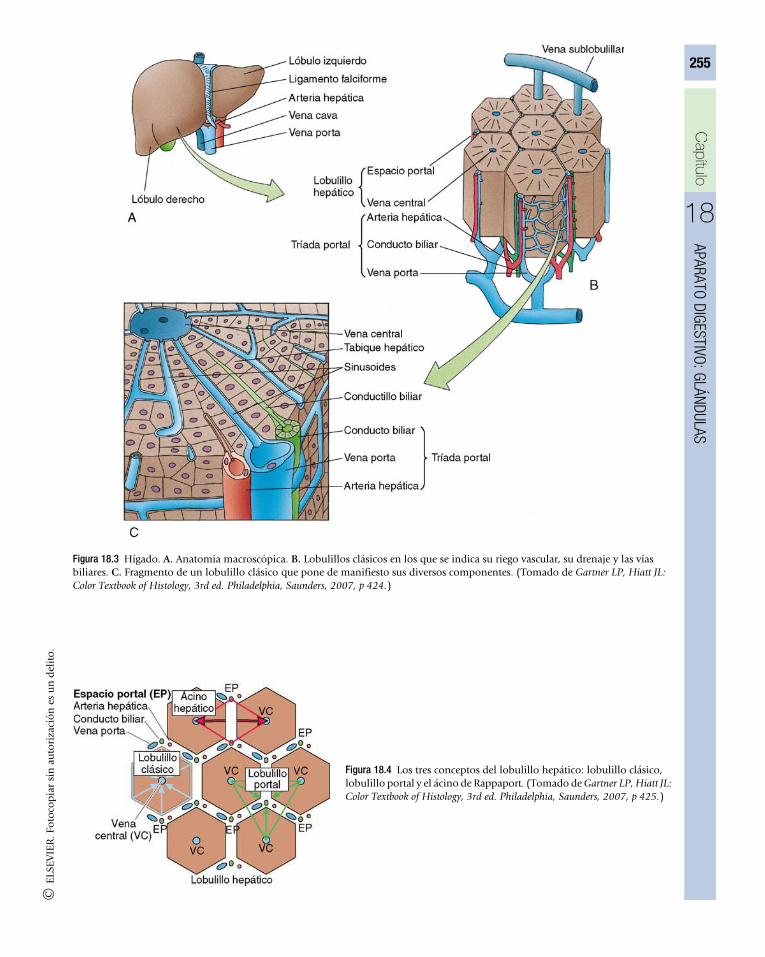
255
18
APARATO
DIGESTIVO
:GLANDULAS
Figura 18.3 Hıgado. A. Anatomıa macroscopica. B. Lobulillos cl�asicos en los que se indica su riego vascular, su drenaje y las vıasbiliares. C. Fragmento de un lobulillo cl�asico que pone de manifiesto sus diversos componentes. (Tomado de Gartner LP, Hiatt JL:
Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 424.)
Figura 18.4 Los tres conceptos del lobulillo hep�atico: lobulillo cl�asico,
lobulillo portal y el �acino de Rappaport. (Tomado deGartner LP, Hiatt JL:
Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 425.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

256
18
APARATO
DIGESTIVO
:GLANDULAS
lıneas imaginarias, que van desde una vena centralhacia el espacio portal, la vena central colindante yel espacio portal opuesto, para volver a la venacentral inicial. El centro del �acino es la arteriola dedistribucion.
SINUSOIDES HEPATICOS, TABIQUES DE CELULAS
HEPATICAS Y HEPATOCITOS
Los tabiques de c�elulas hep�aticas de un hombreadulto, configurados por una sola capa de hepatoci-tos, se anastomosan entre sı en su trayecto desde lavena central hasta alcanzar los bordes de un lobulillocl�asico, de modo semejante a los rayos de una rueda.Los espacios que quedan entre ellos est�an ocupadospor los sinusoides hep�aticos, delimitados por lasc�elulas del revestimiento sinusoidal perforadas. Esteendotelio es poroso porque sus c�elulas tienen huecosintercelulares hasta de 0,5mm. Las c�elulas de Kupffer(macrofagos residentes) est�an situadas en la cara sinu-soidal de las c�elulas endoteliales (fig. 18.5).
. Las c�elulas del revestimiento sinusoidal seencuentran separadas del tabique de hepatocitospor un espacio con una anchura de 0,2 a 0,5mm, elespacio perisinusoidal de Disse, donde tiene lugarel intercambio de sustancias entre el dominiobasolateral de los hepatocitos y la sangre, lo queimpide que los hepatocitos entren en contacto conel torrente circulatorio. Para aumentar su superficie,los hepatocitos adquieren microvellosidades en ellado que linda con el espacio de Disse.
. Asimismo, este espacio contiene fibras de col�ageno,la mayorıa del tipo III, pero tambi�en las hay de lostipos I y IV, y dos tipos de c�elulas: c�elulasgranulosas, que se piensa que son linfocitoscitolıticos naturales, y las escasas c�elulas de Ito(tambi�en llamadas c�elulas almacenadoras de grasao c�elulas estrelladas hep�aticas).. Las c�elulas de Ito guardan grasas y vitamina A,elaboran col�ageno de tipo III y otroscomponentes de la matriz extracelular para suexpulsion hacia el espacio de Disse, ası comofactores de crecimiento.
. Como respuesta a la activacion del factor decrecimiento tumoral-b (TGF-b) por los hepatocitosy las c�elulas de Kupffer, las c�elulas estrelladashep�aticas no solo aumentan su produccion yliberacion de col�ageno, lo que reduce la porosidaddel endotelio, sino que tambi�en se transforman enmiofibroblastos, c�elulas que descienden lacirculacion sanguınea que llega a los senos hep�aticosy facilitan la hipertension portal de origen cirrotico.
Los hepatocitos son c�elulas grandes y poligonales,con un di�ametro de 20 a 30mm, que se encuentranmuy apretados en los tabiques independientes dec�elulas hep�aticas. Cada hepatocito tiene:
. Dominios laterales, donde entra en contacto conotros hepatocitos y constituye estrechos conductosintercelulares, los conductillos biliares, a los quevacıan la bilis primaria por transporte activo.
. Dominios sinusoidales, donde el hepatocito entraen contacto con el espacio de Disse para soltar susecrecion endocrina y endocitar materias desde lossinusoides hep�aticos (fig. 18.6).
Tres de cada cuatro hepatocitos presentan un uniconucleo, mientras que el 25% restante posee dos. El50% de los nucleos de los hepatocitos son pequenos ydiploides, pero los hay demayor tamano y evidencianuna poliploidıa, llegando a 64N.
La bilis que fabrican los hepatocitos es la bilisprimaria, que se concentra y modifica en el interiorde la vesıcula biliar para convertirse en la bilis expul-sada hacia el duodeno.
. Como los hepatocitos sintetizan multitud deproteınas para su propio uso y para su exportacion,su citoplasma tiene gran cantidad de aparato deGolgi, ribosomas y retıculo endopl�asmico (RE)rugoso.
. Estas c�elulas tambi�en poseen abundantesmitocondrias para abastecer sus inmensasnecesidades de trifosfato de adenosina (ATP). Lasmitocondrias de la zona 3 del �acino deRappaport sonm�as pequenas perom�as numerosas que en la zona 1.
. Asimismo, los hepatocitos est�an ampliamentedotados de RE liso porque este org�anulo ejerce sufuncion desintoxicante.
. Los hepatocitos tambi�en presentan multiplesinclusiones como depositos lipıdicos, sobre todoen forma de lipoproteınas de muy baja densidad(VLDL) y glucogeno (partıculas b) en grandesmontones rodeados de RE liso en el caso de lazona 1 y escasos depositos en las c�elulas hep�aticasde la zona 3.
. Los hepatocitos est�an igualmente cargados deperoxisomas, org�anulos que albergan oxidasascapaces de formar H2O2 y catalasa que lodescompone. Estos org�anulos actuan en ladesintoxicacion, la oxidacion b de los �acidos grasos,el metabolismo de las purinas y la gluconeogenia.
Si el hıgado se dana por sustancias toxicas o poruna lesion mec�anica (como la reseccion de una por-cion suya), las c�elulas de Ito sueltan varios factoresde crecimiento, como el TGF-a, el TGF-b, el factor decrecimiento de los hepatocitos, la interleucina 6 y elfactor de crecimiento epid�ermico, que hacen que loshepatocitos existentes experimenten una r�apida acti-vidadmitotica. Si la magnitud del trastorno es grande,los colangıolos y los canales de Hering tambi�en inter-vienen en la regeneracion hep�atica.
Los hepatocitos llevan a cabo la mayor parte de lasmultiples funciones del hıgado.

257
18
APARATO
DIGESTIVO
:GLANDULAS
Figura 18.6 Esquema de un hepatocito que
presenta sus dominios sinusoidal y lateral.RE, retıculo endopl�asmico. (Tomado de
Gartner LP, Hiatt JL: Color Textbook of Histology,
3rd ed. Philadelphia, Saunders, 2007, p 428.)
Figura 18.5 Hıgado. A. Anatomıa macroscopica. B. Lobulillos cl�asicos en los que se indica su riego vascular, su drenaje y las vıas
biliares. C. Fragmento de un lobulillo cl�asico que pone de manifiesto sus diversos componentes. (Tomado de Gartner LP, Hiatt JL:Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 424.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

258
18
APARATO
DIGESTIVO
:GLANDULAS
. Estas funciones consisten en la preparacion de labilis; el metabolismo, almacenamiento y oportunaliberacion de los nutrientes absorbidos por el tubodigestivo; la desintoxicacion de productos nocivos;el paso del colesterol y la IgA secretora a la bilis; lasıntesis de albuminas, globulinas no inmunitarias,protrombina, fibrinogeno, factor VIII, proteınas delcomplemento y proteınas ligadoras para mol�eculassenalizadoras; y la formacion de urea.
. De otras funciones del hıgado se ocupan las c�elulasde Ito (almacenamiento de la vitamina A) y en lasc�elulas de Kupffer (fagocitosis).
BILIS, CONDUCTOS BILIARES Y VESICULA BILIAR
Labilis est�a compuesta de agua, fosfolıpidos, colesterol,sales biliares, pigmentos biliares, lecitina, IgA y electro-litos. Las sales biliares (�acidos biliares) surgen en el REliso del hepatocito al conjugarse la colina, el subpro-ducto metabolico del colesterol, con glicina o taurina,constituyendo el �acido glucocolico o �acido tauroco-lico, respectivamente. La biliverdina, un subproductotras la transformacion del hemoprocedente de la hemo-globina de los eritrocitos destruidos por los macrofagosespl�enicos, se reduce a la bilirrubina (pigmento biliar)insoluble en agua y se lanza hacia el torrente circula-torio, donde se une a la albumina. En los hepatocitosse disuelve el complejo albumina-bilirrubina, y labilirrubina libre, combinada con el transportador pro-teınico citosolico ligandina, penetra en el RE liso,donde vuelve a desacoplarse. La bilirrubina libreentra en el citosol para conjugarse mediante la enzimaglucuronil transferasa en la forma hidrosoluble glu-curonido de bilirrubina (bilirrubina conjugada),que llega a los conductillos biliares para dirigirse haciala vesıcula biliar o expulsarse hacia la circulacion san-guınea (fig. 18.7). Desde la vesıcula biliar, sale hacia elduodeno para eliminarse en las heces, y desde la san-gre, alcanza el rinon para su emision en la orina.
CONDUCTOS HEPATICOS
Los espacios intercelulares delimitados por los hepa-tocitos adyacentes configuran un sistema anastomo-tico de conductillos biliares que vacıan su bilis en loscolangıolos de la periferia de los lobulillos cl�asicos.Estos colangıolos est�an constituidos por hepatocitosen contacto con c�elulas cubicas bajas. Su contenido sevuelca en los canales deHering, ramas delgadas de losconductos biliares, compuestas por c�elulas cubicasbajas que van paralelas a las arteriolas de entrada.Los conductos biliares, integrados por un epiteliocubico simple, se unen entre sı para originar conduc-tos cada vezm�as grandes que acaban en los conductoshep�aticos derecho e izquierdo. Las c�elulas cubicas delos colangıolos, los canales de Hering y los conductosbiliares generan, como respuesta a la secretina emitida
por las c�elulas del SNED en el duodeno, un amorti-guador cargado de bicarbonato, que se conserva en lavesıcula biliar hasta su liberacion al duodeno junto alengendrado por las c�elulas centroacinares y los con-ductos estriados del p�ancreas exocrino.
Los conductos hep�aticos derecho e izquierdo se reu-nen para dar lugar al conducto hep�atico comun, quese une al conducto cıstico de la vesıcula biliar y con-tinua como el conducto col�edoco, de 7u 8 cm delargo. El conducto pancre�atico se junta al conductocol�edoco en la pared del duodeno para formar lapapila duodenal (ampolla de Vater), el orificiocomun en la luz duodenal para la vesıcula biliar y elp�ancreas. Esta desembocadura est�a regulada por ungrupo de fibras musculares lisas, el esfınter de Oddi,con la capacidad de abrir los dos conductos distintoscon independencia entre sı. A no ser que la bilis o lassecreciones pancre�aticas vayan a pasar hacia el duo-deno, ambos conductos est�an cerrados. Esta situacionen el conducto col�edoco permite la entrada de bilis enla vesıcula biliar porque al bajar por �el y no poder saliral duodeno, retrocede, y en la union entre los conduc-tos hep�atico comun y cıstico recorre este ultimo haciaatr�as (el flujo de la bilis en el conducto hep�atico comunchoca con la oposicion del que lleva un sentido contra-rio desde los conductos hep�aticos derecho e izquierdo).
Vesıcula biliar
La vesıcula biliar, sujeta a la c�apsula de Glisson sobrela cara inferior del hıgado, es capaz de guardar unos70ml de bilis; est�a integrada por un cuerpo que pareceun talego cuya abertura, el cuello, se comunica con elconducto cıstico. La funcion de la vesıcula consiste enconcentrar la bilis que almacena. Su luz est�a revestidapor una mucosa muy plegada cuando est�a vacıa, perolisa cuando est�a llena. Su epitelio cilındrico simpleest�a compuesto sobre todo por c�elulas claras, connumerosas microvellosidades, cuya funcion consisteen concentrar la bilis al absorber agua a trav�es de labombaNa+, K+-ATPasa situada en la membrana plas-m�atica basolateral de la c�elula. Al bombear de modoactivo el sodio hacia fuera en el tejido conjuntivosubyacente, le siguen el Cl– y el H2O. La p�erdida deestos iones en la c�elula hace que entren estos mismosdesde la luz, y el cambio osmotico arrastra agua desdela luz hacia la c�elula, lo que reduce el volumen delcontenido luminal y concentra la bilis.
El epitelio tambi�en tiene unas cuantas c�elulas encepillo capaces de producir mucinogeno. La l�aminapropia fibroel�astica es un tejido conjuntivo vascularque aloja pequenas gl�andulas mucosas, pero nadam�as que en el cuello de la vesıcula, cuya secrecionlubrica la estrecha luz de esta region. La vesıculabiliar presenta una tunica muscular lisa mal defi-nida bilaminar, integrada por una capa oblicua

259
18
APARATO
DIGESTIVO
:GLANDULAS
interna y una capa longitudinal externa. Las c�elulasdel SNED (c�elulas I) del duodeno liberan rıtmica-mente la hormona colecistocinina, y la acetil-colina derivada del nervio vago provoca la contrac-cion de estas fibras musculares lisas y el vaciamientointermitente de la bilis desde la vesıcula biliar.
Adem�as, la colecistocinina y la acetilcolina causanuna relajacion simult�anea del esfınter de Oddi, demanera que la bilis puede llegar al duodeno. Lavesıcula biliar tiene una adventicia donde se adhierea la c�apsula de Glisson y una serosa sobre la cara queno est�a pegada.
Figura 18.7 Secrecion de �acidos biliares y bilirrubina por loshepatocitos. REL, retıculo endopl�asmico liso. (Tomado de
Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.
Philadelphia, Saunders, 2007, p 432.)
CONSIDERACIONES CL�INICAS
El hıgado graso (esteatohepatitis) puede ser dedos tipos en EE. UU. y en los paıses occidentales:
alcoholico y no alcoholico. El primero es m�asfrecuente y se debe al consumo excesivo de alcohol.El segundo, la esteatohepatitis no alcoholica,obedece a sındromes del tipo de la diabetesmellitus,la obesidad o las concentraciones altas de
triglic�eridos. Ambas situaciones determinan unaumento del hıgado y pueden desembocar en una
cirrosis hep�atica. La esteatohepatitis por sı sola noes importante, pero deberıa controlarse paraprevenir la formacion de cicatrices en el hıgado y laposterior cirrosis. Su tratamiento consiste en atajarlas causas subyacentes: eliminar el consumo dealcohol en exceso, modificar la alimentacion para
reducir los valores de triglic�eridos y adelgazar en elcaso de la obesidad. El control de la diabetesmellitus, cuando la haya, con insulinoterapia o un
r�egimen diet�etico resulta fundamental.
El proceso que afecta m�as a menudo a la vesıculabiliar y a las vıas biliares es la presencia de cristales
de colesterol, los c�alculos biliares,que se acumulanen dicha vıscera (colelitiasis) o a lo largo de los
conductos biliares extrahep�aticos(coledocolitiasis) y obstruyen el flujo normal de la
bilis. La presencia de c�alculos biliares guarda una
relacion con el sexo y con la edad: es m�as frecuenteentre las mujeres y en las personas mayores de
65 anos. Por regla general, los c�alculos de la vesıcula
biliar son asintom�aticos, pero pueden entrar en losconductos biliares y provocar un cierre con unainflamacion y una infeccion asociadas. Cuando los
conductos biliares extrahep�aticos quedan tapados,el paciente sufre un dolor atroz en la parte superior
derecha del abdomen, con n�auseas y vomitos. Poco
despu�es de la obstruccion, pueden aparecer lainflamacion y la infeccion, con fiebre y escalofrıos, yen un breve plazo desde entones aparecer laictericia. El tratamiento del conducto biliar taponado
es quirurgico o por un m�etodo endoscopico.
Los c�anceres no suelen originarse en los conductos
biliares extrahep�aticos, pero a veces en la union
entre el conducto col�edoco y el hep�atico crece untumor maligno. Cuando la masa obstruya el paso dela bilis, el paciente adquiere una ictericia sin la
existencia de fiebre ni escalofrıos, pero con n�auseas,vomitos, hipersensibilidad o dolor abdominal,adelgazamiento y prurito generalizado. Eltratamiento endoscopico puede permitir lacolocacion de endoprotesis para abrir el conductobiliar, pero las posibilidades de supervivencia no sonfavorables.
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

19 APARATO URINARIOLos dos rinones sirven para eliminar las sustanciastoxicas del torrente circulatorio y conservar el agua,las sales, las proteınas, la glucosa, losamino�acidos y otros productos indis-pensables. Tambi�en prestan ayudapara la regulacion de la presion arterial,la hemodin�amica y el equilibrio acido-b�asico de los lıquidos corporales. Porende, los rinones elaboran hormonas,como la eritropoyetina y las prosta-glandinas, y colaboran en la formacionde la vitamina D.
En la figura 19.1A, una vista hemi-seccionada del rinon pone de mani-fiesto que este organo est�a rodeadopor una c�apsula de tejido conjuntivoque cubre la region externa del par�en-quima renal, llamada corteza, en cuya profundidad sesitua la m�edula con sus pir�amides renales y lascolumnas corticales interpuestas. Cada pir�amiderenal vierte su orina a:
. Un c�aliz menor, y varios c�alices menores lo hacen a
. Un c�aliz mayor, cuya confluencia forma
. La pelvis renal, la region dilatada delur�eter ubicadaen el hilio.
Tambi�en en el hilio, las ramas de la arteria renal pe-netran en el rinon y los afluentes de la vena renal loabandonan.
La unidad b�asica del rinon, denominada tubulourinıfero (fig. 19.1B), tiene una estructura absoluta-mente epitelial y est�a separada de los elementos con-juntivos de este organo por su l�amina basal. Est�acompuesto por una nefrona (cortical o yuxtamedu-lar) y un tubulo colector. Varias nefronas desembo-can en un tubulo colector, y varios tubulos colectoresse reunen para constituir otros tubulos colectores cadavez m�as grandes. Todas las nefronas presentan diver-sos componentes:
. C�apsula de Bowman
. Tubulo proximal
. Asa de Henle
. Tubulo distal
Alrededor del 15% de las nefronas –las yuxtame-dulares– tienen un asa de Henle larga y su corpus-culo renal est�a localizado en la transicion entre lacorteza y la m�edula. En cambio, m�as o menos el 85%de las nefronas –las corticales– tienen un asa de
Henle corta y su corpusculo renal se encuentra m�ascerca de la c�apsula del rinon.
C�apsula de Bowman
La c�apsula de Bowman (fig. 19.1C),la porcion m�as ensanchada de lanefrona, parece un globo duranteel desarrollo embrionario y est�aintegrada por un epitelio pavimen-toso simple que queda invadidopor un conglomerado de capilaresfenestrados, el glom�erulo, cuyosorificios carecen de diafragmas ytienen un di�ametro de 70 a90 nm. De esta manera, el volumenque hay dentro de la c�apsula de
Bowman es escaso y da lugar a una cavidad estrecha,el espacio de Bowman (espacio urinario), situadoentre las capas externa e interna de la c�apsula deBowman (llamadas capas parietal y visceral de lac�apsula de Bowman, respectivamente). Elglom�erulo queda revestido por la capa visceral,cuyas c�elulas, sin excepcion, adquieren una formamodificada y reciben el nombre de podocitos. Enconjunto, el glom�erulo y la c�apsula de Bowman sedenominan corpusculo renal. En el punto en el quelos podocitos entran en contacto con las c�elulasendoteliales del glom�erulo, se fusionan las dosl�aminas basales. Los podocitos (v. fig. 19.1C) lle-van numerosas dilataciones citopl�asmicas largas ycon aspecto de tent�aculo –prolongaciones pri-marias (mayores)– que a su vez poseen muchasprolongaciones secundarias (pedicelos), queadoptan una disposicion ordenada. Los pedicelosenvuelven por completo la mayor parte de los capi-lares glomerulares al interdigitarse con los de lasprolongaciones mayores contiguas de otros podoci-tos diferentes.
CONSIDERACIONES CL�INICAS
Durante la nefrogenia e incluso al nacer, losrinones ofrecen signos de lobulaciones, pero alaparecer las nefronas, adquiere una forma lisa yconvexa. Sin embargo, a veces los lobulos puedenreconocerse en un adulto desde el exterior, y estasituacion recibe el nombre de rinon lobulado. Elcaso es que no tiene unas consecuenciasfuncionales manifiestas.
� 2011 Elsevier Espana, S.L. Reservados todos los derechos
TERMINOS CLAVE. Tubulos urinıferos. Nefronas. Corpusculos renales. Asa de Henle. Aparato
yuxtaglomerular. Tubulos colectores. Ur�eter. Vejiga urinaria
260

261
19
APARATO
URINARIO
Figura 19.1 A. Rinon hemiseccionado. B. Tubulos urinıferos.C.Corpusculo renal. (A-C. Tomado deGartner LP, Hiatt JL: Color Textbookof Histology, 3rd ed. Philadelphia, Saunders, 2007, pp 438 [A and B]and 441 [C].)�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

PROCESO DE FILTRACION
Cuando el lıquido abandona el lecho capilar parapenetrar en el espacio de Bowman, tiene que pasar atrav�es de la barrera de filtracion, compuesta por:
. El endotelio glomerular
. Las l�aminas basales fusionadas (que atrapanmol�eculas mayores de 69.000Da)
. El diafragma de la hendidura de filtracion(fig. 19.2A y recuadro).
El lıquido filtrado que llega al espacio de Bowmanse llama ultrafiltrado glomerular. Como la l�aminabasal retiene las macromol�eculas m�as grandes, aca-barıa taponada si no fueran fagocitadas permanente-mente por las c�elulasmesangiales intraglomerularesy repuestas mediante los esfuerzos conjuntos de lacapa visceral de la c�apsula de Bowman (podocitos)y las c�elulas endoteliales glomerulares.
Tubulo proximal
El tubulo proximal tiene dos regiones:
. La porcion contorneada (tubulo contorneadoproximal) larga y muy sinuosa, situada cerca delcorpusculo renal
. La porcion recta (rama gruesa descendente del asade Henle)m�as corta y rectilınea, que se sumerge enla m�edula, donde empalma con la rama delgadadescendente del asa de Henle (fig. 19.2B).
Las dos regiones del tubulo proximal est�an constitui-dasporunepitelio cilındrico simple conunborde apicalestriado de microvellosidades muy apretadas perfecta-mente formado,unaparatoendocitosicobiensurtidodevesıculasdeendocitosis, yunasprolongaciones celulareslaterales entrelazadas y mezcladas en una marana.
El tubulo proximal es el responsable de reabsorberdel 60 al 80%del agua, el sodio y el cloruro; el 100%delas proteınas, los amino�acidos y la glucosa; y las sustan-cias toxicas del ultrafiltrado que llegan a su luz desde elespacio de Bowman del corpusculo renal. Una bombade sodio alimentada por trifosfato de adenosina (ATP),que est�a presente en la membrana plasm�atica basal,
manda sodio hacia el estroma de tejido conjuntivo, yel cloruro le sigue de forma pasiva para conservar laneutralidad el�ectrica; todo este movimiento va acom-panado por el agua para mantener el equilibrio osmo-tico a trav�es de los canales de acuaporina-I, lo quereduce el volumen sin afectar a la osmolaridad delultrafiltrado. El aparato endocitosico es el responsablede reabsorber las macromol�eculas m�as grandes.
Rama delgada del asa de Henle
La rama delgada del asa de Henle, compuesta por unepitelio pavimentoso simple, tiene tres regiones:
. Rama delgada descendente, recta
. Asa de Henle, con forma de horquilla
. Rama delgada ascendente recta, que desemboca enel tubulo distal (v. fig. 19.2B).
Aunque en las nefronas corticales a veces falte larama delgada, en las yuxtamedulares mide casi 1 cmde longitud y puede llegar lejos en la m�edula, hastaalcanzar la papila renal. La rama delgada descendentees totalmente permeable al agua, la urea, el sodio, elcloruro y otros iones, mientras que la ascendente esrelativamente impermeable al agua, pero es permea-ble a la urea y a la mayorıa de los iones.
Tubulo distal
El tubulo distal, constituido por un epitelio cubicosimple, carece de una dotacion tan abundante demicrovellosidades o unas interdigitaciones lateralestan complejas como las c�elulas del proximal. Su reco-rrido puede dividirse en tres regiones (v. fig. 19.2B):
. Porcion recta (rama gruesa ascendente), querepresenta la continuacion de la rama delgadaascendente del asa de Henle
. M�acula densa muy corta
. Tubulo contorneado distal
La porcion recta del tubulo distal mide casi 1 cm delargo, y sus c�elulas generan unas zonulas de oclusionmuy eficaces con sus vecinas adyacentes, creando una
262
19
APARATO
URINARIO
CONSIDERACIONES CL�INICAS
La enfermedad de cambios mınimos es el
trastorno renal m�as frecuente entre los ninos. Lospedicelos adyacentes parecen fusionarse entre sı, loque se traduce en una proteinuria. En la mayorıa delos casos, los tratamientos con corticoesteroidessolucionan de forma satisfactoria el proceso.
La glomeruloesclerosis focal y segmentariaasociada a la heroına aparece despu�es del consumointravenoso prolongado de heroına, lo que da lugar a
una importante proteinuria con uremia irreversible enun plazo de dos anos. El sındrome afecta sobre todo avarones estadounidenses de raza negra menores de50 anos. La enfermedad ataca a los podocitos,haciendo que parte de ellos degeneren y pierdan el
contacto con la l�amina basal. El mejor tratamientoconsiste en el abandono del uso de la heroına, pero enla mayorıa de los pacientes se produce su evolucionhaciaunanefropatıa terminal que tal vezexija recurrir a
la di�alisis o al trasplante renal.

263
19
APARATO
URINARIO
Figura 19.2 A. Segmento del glom�erulo. B. Tubulo urinıfero y cortes transversales de sus componentes. (Tomado deGartner LP, Hiatt JL:
Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, pp 443 [A] and 446 [B].)�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

barrera eficiente entre la luz y el estroma de tejidoconjuntivo a su alrededor, lo que impide que las sus-tancias tomen el camino paracelular. Esta porcion rectapresenta una gran impermeabilidad al agua y a la urea,pero sus c�elulas cubicas poseen unas bombas de clo-ruro (y tal vez de sodio) de situacion basal que trans-fieren Na+ y Cl– hacia el tejido conjuntivo, y ası dismi-nuyen la concentracion de estos iones en el ultrafiltradode la luz de la porcion recta del tubulo distal hasta talextremo que en el momento de llegar a la union corti-comedular, es bastante hipoosmotico, pero la concen-tracion de urea sigue siendo alta (v. m�as adelante).
La m�acula densa, ubicada entre las arteriolas glo-merulares aferente y eferente, en las inmediaciones delcorpusculo renal de ese mismo tubulo distal, formaparte del aparato yuxtaglomerular. Los tubulos con-torneados distales (fig. 19.3A) midenmenos de 5mmde longitud, son impermeables al agua, y descargan suultrafiltrado en los tubulos colectores. Sus c�elulas cilın-dricas tienen receptores de aldosterona y bombas deintercambio sodio-potasio Na+,K+-ATPasa, ambos delocalizacion basal. La union de la aldosterona a susreceptores activa estas c�elulas para pasar sodio (y, pasi-vamente, cloruro) hacia el intersticio renal, lo que bajaaun m�as la osmolaridad del ultrafiltrado.
Aparato yuxtaglomerular
El aparato yuxtaglomerular (su funcion se describem�as adelante) est�a compuesto de tres partes: lam�aculadensa, las c�elulas yuxtaglomerulares y las c�elulasmesangiales extraglomerulares (fig. 19.3B).
. Los nucleos de las c�elulas estrechas y p�alidas de lam�acula densa est�an muy cerca unos de otros yadoptan el aspecto de una mancha densa, de ahı sudenominacion. No hay l�amina basal entre lam�acula densa y las c�elulas yuxtaglomerulares.
. Las c�elulas yuxtaglomerulares son miocitos lisosmodificados de las arteriolas glomerulares aferentesy (muchas veces) eferentes. Estas c�elulas sintetizan yalmacenan renina, una enzima proteolıtica queconvierte el angiotensinogeno en angiotensina I.Tambi�en contienen enzima convertidora de laangiotensina, angiotensina I y angiotensina II.
. Las c�elulas mesangiales extraglomerulares ocupanel espacio que queda entre las arteriolasglomerulares aferente y eferente. Tambi�en puedenentrar en el corpusculo renal, donde reciben elnombre de c�elulas mesangiales intraglomerulares.
Tubulos colectores
La segunda parte de los tubulos urinıferos, los tubu-los colectores, miden en torno a 2 cm de largo ytienen un origen embrionario diferente a las nefro-
nas. Ambos quedan conectados durante el desarro-llo del embrion. Los tubulos colectores est�an inte-grados por un epitelio cubico simple; presentan tresregiones –cortical, medular y papilar– y durantedeterminados procesos, cambian el ultrafiltradoque reciben de las nefronas.
. Los tubulos colectores corticales est�an situados enlos rayosmedulares, y su epitelio cubico se encuentraconstituido por c�elulas principales e intercaladas.
. Las c�elulas principales no poseen m�as que unascuantas microvellosidades cortas, y sus membranasplasm�aticas laterales son lisas y solo ofrecen unaspocas interdigitaciones con las c�elulas contiguas. Ensu pared hay canales de acuaporina-2 sensibles a lahormona antidiur�etica (ADH o vasopresina).
. Las c�elulas intercaladas son de dos clases, tipo A ytipo B; ambas llevan numerosos micropliegues yvesıculas de distribucion apical.. Las c�elulas de tipo A transportan H+ hacia la luz atrav�es de una H+-ATPasa de localizacion apical yacidifican la orina.
. Las c�elulas de tipo B desplazan su H+-ATPasa a lazona basolateral, y reabsorben H+ y segreganHCO3
–.. Varios tubulos colectores corticales se reunen paraformar tubulos colectores medularesm�as grandes,que crecen de di�ametro a medida que profundizanen la m�edula. Los de la zona externa de la m�edulaest�an compuestos por c�elulas principales eintercaladas, mientras que en los de la zona internasolo hay c�elulas principales.
. Varios tubulos colectores medulares confluyenentre sı para dar lugar a los grandes tubuloscolectores papilares (conductos de Bellini) (conun di�ametro de 200 a 300mm) que desembocan enel �area cribosa de la papila renal para vaciar su orinaen el c�aliz menor. Los tubulos colectores papilaresest�an dotados de un epitelio cilındrico simpleintegrado unicamente por c�elulas principales. Enellas existen receptores de ADH, y si se produce launion de estamol�ecula con ellos, las c�elulas colocanlos canales de acuaporina-2 en su membrana y sevuelven permeables al agua y a la urea; cuando elagua abandona las luces de estos tubulos y penetraen el intersticio renal, la orina pasa a serhiperosmotica y baja de volumen.
Intersticio renal
Los tubulos urinıferos y la gran red vascular del rinonse hallan totalmente rodeados por finos elementos detejido conjuntivo, denominados intersticio renal.Nada m�as que el 7% del volumen cortical y menosdel 30% del volumen medular est�an compuestos portejido conjuntivo.
264
19
APARATO
URINARIO

CONSIDERACIONES CL�INICAS
La necrosis tubular aguda, puesta de manifiestopor la presencia de unos rinones congestionados ynecrosis focal de los tubulos renales, puede debersea la accion de sustancias nefrotoxicas o a unaisquemia, lo que deriva en una insuficiencia renalaguda. La correccion inmediata del problema
permite una r�apida recuperacion, senalada por elascenso en la produccion de orina y la disminucion
de la creatinina s�erica.
Se ha demostrado que la insuficiencia renalirreversible, r�apida y progresiva est�a ocasionada
por el empleo de un r�egimen de adelgazamiento
que lleva la planta arom�atica china Aristolochia
fangchi, perteneciente a una familia de plantas que
contienen �acido aristoloquico. En los pacientes que
consumen esta hierba para perder peso tambi�en se
ha comprobado la aparicion de c�anceres renales.La mayor parte de los afectados eran mujeresobesas de mediana edad. Muchas personaspreocupadas por su salud recurren acomplementos o remedios medicinales sin avisar a
su m�edico o a otros profesionales sanitariosporque los compuestos que toman se venden sinreceta, y piensan erroneamente que son«naturales». Siempre es prudente que losprofesionales sanitarios indaguen si sus enfermosutilizan suplementos de venta sin receta.
265
19
APARATO
URINARIO
Figura 19.3 A. Tubulo urinıfero y cortes transversales de sus componentes. B. Corpusculo renal y aparato yuxtaglomerular. (Tomado
de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, pp 446 [A] and 450 [B].)
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

Circulacion renal
La arteria renal (fig. 19.4), procedente de la aortaabdominal, se bifurca en sus divisiones anterior yposterior, que a su vez se ramifica para formar lascinco arterias segmentarias que entran en el rinonpor el hilio. Estas arterias segmentarias no se anasto-mosan entre sı; en caso de obstruirse, se interrumpe lacirculacion sanguınea dirigida a la region renal irri-gada por ese vaso.
Cada arteria segmentaria da origen a las arteriaslobulares, que se separan en dos o tres arterias in-terlobulares cuyo trayecto pasa entre las pir�amidesrenales para subir hacia la union corticomedular,donde constituyen las arterias arqueadas. Estos vasospermanecen en el lımite entre la corteza y la m�edulasegun se distribuyen sobre la base de las pir�amidesrenales para generar numerosas arterias interlobuli-llares. Las ramas terminales de las arterias arqueadastambi�en se convierten en arterias interlobulillares.
Las arterias interlobulillares ascienden hacia lacorteza m�as o menos a mitad de camino entre cadados rayos medulares contiguos y suministran multi-ples vasos que abastecen a los corpusculos renales.Estos elementos son las arteriolas glomerulares afe-rentes, responsables de producir el lecho capilar, oglom�erulo, del corpusculo renal (v. fig. 19.4). Lasramas terminales de algunas arterias interlobulillarespasan a ser arteriolas glomerulares aferentes, mientrasque otras acaban justo en la profundidad de la c�apsularenal para participar en la creacion delplexo capsular.El glom�erulo se vacıa por las arteriolas glomerulareseferentes (AGE), razon por la que la presion arterial esm�as alta en su interior que en la mayor parte de losdem�as lechos capilares (v. fig. 19.4).
. Las AGE de las nefronas corticales son lasresponsables de la aparicion de la red capilarperitubular que surte a los tubulos del laberintocortical. Estos capilares, cuyas c�elulas endotelialeselaboran eritropoyetina, desembocan en la venaarqueada (v. fig. 19.4).
. Las AGE de las nefronas yuxtamedulares seramifican para originar 25 capilares largos conforma de horquilla que se extienden hacia lam�edulahasta llegar a la papila renal. Los brazosdescendentes de estos capilares, las arteriolasrectas, tienen una luz estrecha, mientras que losascendentes, las v�enulas rectas, disponen de unaluz m�as amplia, y vierten su sangre en las venasarqueadas. En conjunto, las arteriolas y las v�enulasrectas reciben el nombre de vasos rectos, yenvuelven las regiones medulares de los tubulosurinıferos. Su funcion en la concentracion de laorina se describe m�as adelante.
Las venas arqueadas devuelven su sangre hacialas venas interlobulillares, que la transfieren a lasvenas interlobulares y desde ellas a las venas renales(v. fig. 19.4), que acaban en la vena cava inferior.
Mecanismo de formacion de la orina
Cada 5min, todo el volumen de la sangre atraviesalos dos rinones; unos 1.250ml de sangre penetranen los glom�erulos por minuto. Como esta estruc-tura es un lecho capilar arterial, la presion arteriales mucho m�as alta que en la mayor parte de losdem�as lechos capilares. Este y otros factores ejercenun promedio de 25mmHg (fuerza de filtracion),que hace salir el componente lıquido de la sangrefuera de los capilares hacia el espacio de Bowman,donde recibe la denominacion de ultrafiltrado glo-merular; cada minuto llegan 125ml del ultrafil-trado a los espacios de Bowman. Para ello, estelıquido ha de pasar por la barrera de filtracion, com-puesta por:
. Las c�elulas endoteliales del glom�erulo (detienenlas partıculas con un di�ametro >90nm)
. La l�amina basal fusionada (detiene lasmacromol�eculas >69 kDa)
. Las hendiduras de filtracion de los podocitos.
Las l�aminas basales (semejantes a un papel de filtro enun embudo de B€uchner) quedarıan r�apidamenteobstruidas, pero como las c�elulas mesangiales intra-glomerulares se encargan de fagocitarlas, los podoci-tos y las c�elulas endoteliales las sustituyen.
CONSIDERACIONES CL�INICAS
Los infartos renales son frecuentes entre lospacientes con anemia drepanocıtica, en los que
los vasos m�as pequenos quedan ocluidos por loseritrocitos mal formados. La magnitud de la lesionviene determinada por el vaso que haya quedadocerrado.
La fibrodisplasia (displasia fibromuscular) es unproceso de causa desconocida que afecta amujeres jovenes. La arteria renal sufre unestrechamiento debido al deposito de tejidoconjuntivo fibroso en varios puntos de la paredarterial. La estenosis es la responsable de lahipertension y deberıa sospecharse en mujeresjovenes con una presion arterial elevada. Estetrastorno responde bien a la angioplastia y nosuele recidivar.
266
19
APARATO
URINARIO
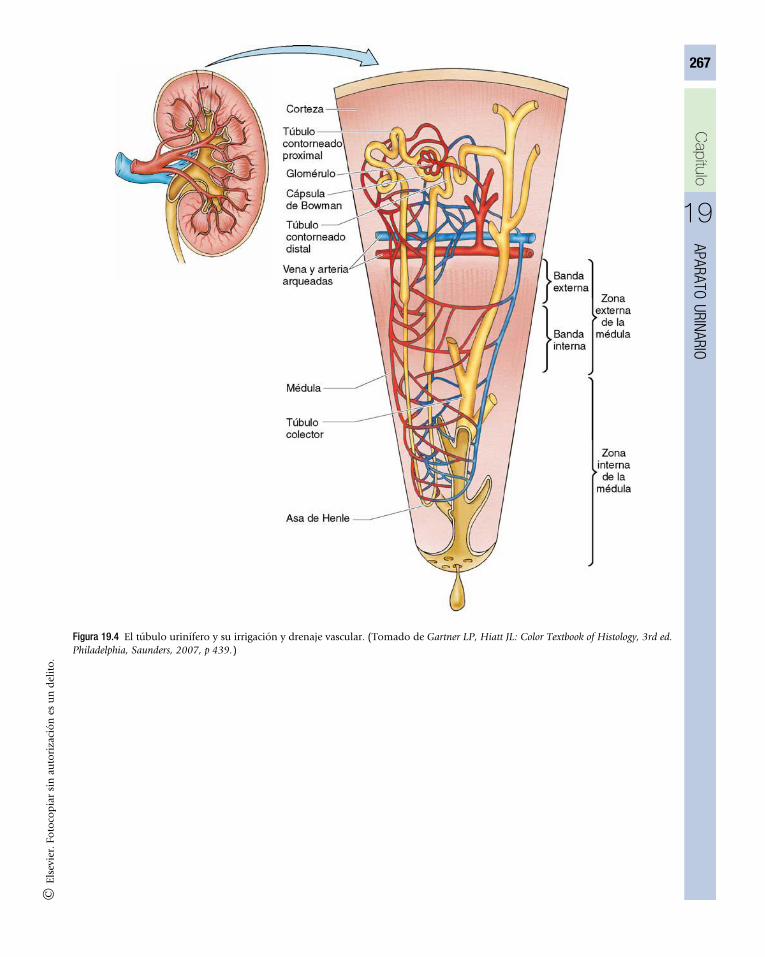
267
19
APARATO
URINARIO
Figura 19.4 El tubulo urinıfero y su irrigacion y drenaje vascular. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.
Philadelphia, Saunders, 2007, p 439.)
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

La mayor proporcion de la reabsorcion tiene lugaren el tubulo proximal, y las sustancias recuperadasentran en los capilares del intersticio renal para devol-verlos al torrente circulatorio (fig. 19.5A y B). Lamayorıa de los iones se reabsorben debido a la accionde las bombas de sodio situadas en las membranasplasm�aticas basolaterales de los tubulos proximales;del 67 al 80% del Na+, Cl� y H2O se reabsorbe en eltubulo proximal, lo que reduce el volumen sin reper-cutir sobre la osmolaridad del ultrafiltrado. Adem�as,se reabsorbe casi el 100%del HCO3
�; y el 100%de lasproteınas, glucosa, creatinina y amino�acidos vuelvena la sangre.
Las nefronas yuxtamedulares poseen un asa deHenle larga y, por medio del sistema multiplicadorpor contracorriente, establecen un gradiente osmo-tico creciente desde la union corticomedular hasta lapapila renal.
. El epitelio pavimentoso simple de la rama delgadadescendente del asa de Henle es permeable al aguay en parte a las sales. A medida que baja elultrafiltrado, pierde agua, lo que aumenta suosmolaridad (v. fig. 19.5A y B).
. La relativamente corta rama delgada ascendentedel asa de Henle b�asicamente es impermeable alagua. Segun avanza el ultrafiltrado hacia la ramagruesa ascendente, entra urea en la luz y las sales laabandonan.
. La rama gruesa ascendente del asa de Henle est�acompuesta por un epitelio cubico simple cuyasc�elulas tienen un cotransportador Na+/K+/2Cl�
apical y una bomba Na+,K+-ATPasa de localizacionbasal m�as bombas de cloruro y tal vez de sodio,que lanzan Cl� y Na+ hacia el intersticio renaldesde la luz, lo que instaura el gradiente deconcentracion de sales, que es m�as alto en la parteprofunda de la m�edula y m�as bajo hacia la corteza(v. fig. 19.5A y B). Por consiguiente, el volumendel ultrafiltrado permanece constante, pero suosmolaridad disminuye a medida que el lıquidode la luz se acerca a la corteza.
CONTROL DEL FILTRADO EN EL APARATO
YUXTAGLOMERULAR
Cuando el ultrafiltrado glomerular llega a la m�aculadensa, cuyas c�elulas est�an cargadas de las enzimas dela ciclooxigenasa (COX-2) y oxido nıtrico sintasa, sevigila su concentracion de sodio (o de cloruro) y suvolumen. Si la cantidad de sodio es pequena, la oxidonıtrico sintasa sintetiza oxido nıtrico, que al liberarse,provoca dilatacion de la arteriola glomerular aferente,lo que incrementa el flujo de sangre hacia elglom�erulo. Simult�aneamente, el oxido nıtrico y la
prostaglandina E2, un producto de la COX-2, hacenque las c�elulas yuxtaglomerulares expulsen renina altorrente circulatorio. Esta enzima escinde dos ami-no�acidos del angiotensinogeno circulante, trans-form�andolo en angiotensina I. Las c�elulas endote-liales de la mayorıa de los capilares en el organismo,pero sobre todo en el pulmon, poseen una gran can-tidad de enzima convertidora de la angiotensina,que separa dos amino�acidos de la angiotensina I paraoriginar angiotensina II. Esta mol�ecula ocasiona lacontraccion de los vasos sanguıneos, entre ellos la ar-teriola glomerular eferente (lo que eleva la presionarterial dentro del glom�erulo con un aumento para-lelo de la filtracion glomerular), y determina que lacorteza suprarrenal descargue aldosterona, una hor-mona que facilita unamayor reabsorcion deNa+ y Cl�
desde la luz del tubulo contorneado distal, lo quevuelve m�as hipotonico el ultrafiltrado.
MOVIMIENTO DE AGUA Y UREA DESDE Y HACIA
EL FILTRADO EN LOS TUBULOS COLECTORES
Como el tubulo colector atraviesa toda la longitud dela m�edula, tropieza con unos gradientes osmoticosid�enticos a los de las ramas del asa de Henle (v. fig.19.5A y B). Sus c�elulas son impermeables al agua si nohay ADH (v. fig. 19.5A); sin embargo, cuando estahormona se une a los receptores deADHde las c�elulascubicas del tubulo colector, hace que coloquen loscanales de acuaporina en su membrana plasm�atica.A medida que el ultrafiltrado desciende hacia el �areacribosa, pierde agua de manera pasiva, y en la parteinterna de la m�edula pierde urea, tambi�en de manerapasiva. La orina baja de volumen y queda m�as concen-trada (v. fig. 19.5B). Por ende, sube la concentracion dela urea en el intersticio de la parte interna de la m�edula,lo que es responsable del gran gradiente de concentra-cion existente en esta porcion (v. fig. 19.5B).
Vasos rectos
Los vasos rectos, compuestos por las arteriolas rec-tas descendentes estrechas y las m�as anchas v�enulasrectas ascendentes, son absolutamente permeablesal agua y las sales, y la sangre de ambas ramas deestos vasos con forma de horquilla reacciona frenteal gradiente de concentracion en la m�edula renal(fig. 19.5C) y genera un sistema de intercambiopor contracorriente. Los vasos rectos no solo man-tienen el gradiente osmotico de la m�edula renal sinoque tambi�en lo aprovechan al extraer m�as agua ysales en el mayor volumen de salida por las v�enulasrectas que el que entra por las arteriolas rectas demenor calibre (v. fig. 19.5C).
268
19
APARATO
URINARIO

CONSIDERACIONES CL�INICAS
El sındrome de Fanconi se da en ninos cuyostubulos proximales no reabsorben la cantidadapropiada de glucosa, fosfato, bicarbonato y
amino�acidos, lo que se traduce en una acidosis,deshidratacion, desequilibrio hidroelectrolıtico,proteinuria, raquitismo, osteomalacia y retraso delcrecimiento. Este sındrome podrıa tener diversascausas, entre ellas un componente hereditario,
como la intolerancia hereditaria a la fructosa. La
aparicion del sındrome de Fanconi tambi�en se haatribuido al empleo de tetraciclina caducada, cuyoproducto de descomposicion anhidro-4-tetraciclinaprovoca una disfuncion tubular reversible, laintoxicacion por metales pesados y la inhalacion depegamento, ası como determinadas plantasmedicinales chinas de venta sin receta.
269
19
APARATO
URINARIO
Figura 19.5 Histofisiologıa del tubulo urinıfero. A. Sin ADH. B. Con ADH. C. Vasos rectos. (Tomado de Gartner LP, Hiatt JL: Color
Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, pp 456 [A and B] and 460 [C].)
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

Vıas excretoras
Las vıas excretoras del aparato urinario constan de losc�alices menores ymayores, la pelvis del rinon, el ur�eter,la vejiga urinaria y la uretra, estos dos ultimos impares.
CALICES Y URETER
. Cada c�aliz menor, revestido por un epitelio detransicion, recibe la orina desde el �area cribosa de lapapila renal. En la profundidad del epitelio seencuentra la l�amina propia rodeada por una capadelgada de musculo liso, cuya contraccion empujala orina hacia un c�aliz mayor. Salvo por su tamano,los c�alices mayores se parecen a los c�alices menoresy a la pelvis renal.
. El ur�eter, quemide alrededor de 30 cm de longitud y0,5 cm de di�ametro, est�a tapizado por una mucosacuyo epitelio de transicion se halla sobre una l�aminapropia compuesta por tejido conjuntivo fibroel�astico(figs. 19.6 y 19.7). Una capa longitudinal interna demiocitos lisos y otra circular externa forman la tunicamuscular del ur�eter, y sus contracciones perist�alticasvacıan la orina en la vejiga urinaria.
VEJIGA URINARIA
Cuando la vejiga urinaria se distiende por la orinaacumulada, su epitelio de transicion impermeable seadelgaza y se aplana (v. figs. 19.6 y 19.7). Sus grandesc�elulas cupuliformes tambi�en se aplanan y las mem-branas plasm�aticas se despliegan, de manera que lasgruesas regiones de las placas dejan de estar plegadashacia el citoplasma, y en su lugar pasan a formar unmosaico de placas engrosadas y regiones entre lasplacas m�as delgadas, lo que sirve para alojar la cre-ciente cantidad de orina de la vejiga.
El trıgono, la zona triangular lisa de la mucosavesical, tiene tres v�ertices:
. Las desembocaduras de los dos ur�eteres
. El orificio uretral, donde a veces asientan unasgl�andulas mucosas en el tejido conjuntivofibroel�astico.
La tunica muscular de la vejiga urinaria est�a inte-grada por una capa longitudinal interna, otra circu-larmedia y otra longitudinal externa demusculo lisoque amenudo se encuentran entrelazadas y son indis-tinguibles como estratos diferentes. El musculo esfın-ter interno de la uretra est�a constituido por la gruesacapa circular media. Una adventicia fibroel�asticarodea a la tunica muscular.
URETRA
La uretra lleva la orina desde la vejiga urinaria para suvaciamiento. La uretra femenina es m�as corta que lamasculina.
. La uretra femenina mide unos 5 cm de largo(v. fig. 19.6); est�a revestida por un epiteliode transicion en las inmediaciones de lavejiga urinaria y por un epitelio pavimentosoestratificado no queratinizado a lo largo delresto de su longitud. Las gl�andulas mucosasde Littre est�an en su l�amina propiafibroel�astica.
. La uretra masculina tiene m�as o menos 20 cm delongitud y presenta tres regiones: la uretraprost�atica, la uretra membranosa y la uretrapeniana (o esponjosa) (v. fig. 19.7).. La uretra prost�atica atraviesa la prostata y est�atapizada por un epitelio de transicion.
. La uretra membranosa cruza el diafragmaurogenital y est�a revestida por un epiteliocilındrico estratificado (con zonas deepitelio cilındrico seudoestratificado).
. La uretra peniana recorre toda la longitud delpene y est�a recubierta por un epitelio cilındricoestratificado (con zonas de epitelio cilındricoseudoestratificado) hasta el glande del pene,donde su epitelio es pavimentoso estratificadoqueratinizado.
La l�amina propia est�a compuesta de un tejidoconjuntivo fibroel�astico con gl�andulas de Littre pro-ductoras democo en las tres regiones de la uretra mas-culina.
270
19
APARATO
URINARIO
CONSIDERACIONES CL�INICAS
El 10% de los c�anceres renales son carcinomas de
c�elulas de transicion de los c�alices y la pelvis
ureteral. Los c�anceres renales a menudo est�an
asociados a los carcinomas de c�elulas de
transicion de la vejiga urinaria, que son mucho m�ashabituales.
La infeccion de las vıas urinarias (IVU) agudasin complicaciones se da con frecuencia entrelas mujeres, afectando cada ano a 11 millones en
EE. UU., pero ni siquiera a 10 de cada 10.000
hombres menores de 50 anos. Lo m�as corriente esque, entre las mujeres jovenes sanas, la causa de lainfeccion sean las bacterias intestinales (por reglageneral, Escherichia coli del recto), que penetran en
la vejiga urinaria a trav�es de la uretra. Sus sıntomasclınicos consisten en disuria, sensacion dequemazon y dolor con la miccion, aumento en lafrecuencia de la necesidad de orinar, y dolor en lasregiones suprapubicas.

271
19
APARATO
URINARIO
Figura 19.7 Vıa urinaria masculina. (Tomado deNetter FH: Atlas of Human Anatomy, 3rd ed. Teterboro, NJ, ICON Learning System, 2003.
� Elsevier Inc. Todos los derechos reservados.)
Figura 19.6 Vıa urinaria femenina. (Tomado de Netter FH: Atlas of Human Anatomy, 3rd ed. Teterboro, NJ, ICON Learning System, 2003.
� Elsevier Inc. Todos los derechos reservados.)
�Elsevier.Fo
tocopiarsinau
torizaciones
undelito.

20 APARATO GENITALFEMENINO
El aparato genital femenino comprende los ovarios,los oviductos, el utero, la vagina y los genitales exter-nos, ası como las gl�andulas mamarias.
Antes de la pubertad, las c�elulas ger-minativas del ovario se encuentran enfase de reposo. Una vez que la hipofisiscomienza a liberar las gonadotropinas,se activa el aparato genital, alrededor delos 12 anos de edad, y tiene lugar lamenarquia, la primera menstruacion. Apartir de entonces, el ciclo menstrual de28 dıas se conserva durante toda la vidareproductiva de la mujer hasta llegar a lamenopausia.
Ovarios
Cada ovario (3 cm� 2 cm� 1 cm) est�a sujeto al liga-mento ancho del utero por elmesoovario (fig. 20.1).Adem�as, se encuentra cubierto por el epitelio germi-nativo pavimentoso simple, que se halla sobre suc�apsula de tejido conjuntivo, la tunica albugınea. Elovario posee una corteza externa y una m�edulainterna.
El estroma de tejido conjuntivo (compartimentointersticial) de la corteza est�a poblado por los folıcu-los ov�aricos y las c�elulas del estroma (intersticiales)que parecen fibroblastos y forman las tecas interna yexterna de los folıculos que albergan el ovocito pri-mario.
. En el momento de la fecundacion, el sexo delembrion queda determinado por la existencia ono del cromosoma Y. Si est�a presente, el factorde transcripcion del gen SRY (regiondeterminante del sexo en el cromosoma Y),el factor determinante de los testıculos,induce la aparicion de los testıculos.
. La falta del gen SRY produce el estado por omision:la constitucion de las gonadas femeninas.
. Al principio del desarrollo, surgen un par decrestas cubiertas de epitelio, las crestas gonadales,y las c�elulas epiteliales que las recubren emigranhacia su interior para configurar los cordonessexuales primitivos.
. Al cumplirse 6 semanas desde la fecundacion, lasc�elulas germinativas primordiales emigran desde
el saco vitelino hacia las crestasgonadales, provocan una mayorevolucion de las gonadas, y siguendividi�endose para generarnumerosas c�elulas germinativas.A estas alturas, las gonadasmasculina y femenina sonid�enticas y se denominangonadas indiferenciadas.
. En lamujer, los cordones sexualesprimitivos emigran hacia lam�edula de las crestas gonadales yforman agregados celulares quedegeneran pronto, para quedar
sustituidos por tejido conjuntivo. Durante lasemana 7, otras c�elulas del recubrimiento epitelialemigran hacia la corteza para constituir loscordones sexuales corticales.
. A lo largo del cuarto mes, estos cordones sexualescorticales se disocian, y sus c�elulas rodean a lasc�elulas germinativas primordiales, reciben elnombre de c�elulas foliculares (aunque las c�elulasfoliculares tambi�en pueden nacer a partir delepitelio germinativo) y las c�elulas germinativasprimordiales se convierten en ovogonias.
. A finales del quintomes de la vida fetal, cada cortezaov�arica aloja un mınimo de tres millones deovogonias, pero tan solo 500.000 se ven rodeadospor las c�elulas foliculares; los dem�as degeneran. Losovogonias restantes entran en la primera divisionmeiotica y pasan a denominarse ovocitos primarios.Cada ovocito primariom�as las c�elulas foliculares quelo rodean se llama folıculo primordial.
. Las c�elulas foliculares que hay alrededor delovocito primario segregan el factor bloqueador dela meiosis, que detiene al ovocito primario en lafase de diploteno de la meiosis I justo hasta antes dela ovulacion del ovocito. En los 10 primeros anosde vida, dos tercios de los folıculos primordialesexperimentan una atresia.
. Los ovocitos primarios restantes puedenmantenerse en este estado detenido de la meiosishasta la pubertad o durante los 30 o 40 anossiguientes.
� 2011 Elsevier Espana, S.L. Reservados todos los derechos
TERMINOS CLAVE. Corteza del ovario. Folıculos ov�aricos. Ovulacion. Trompas de Falopio. Utero. Ciclo menstrual. Placenta. Gl�andulas mamarias
272

CONSIDERACIONES CL�INICAS
Durante el desarrollo, como respuesta a losestrogenos, se forman las trompas de Falopio, elcuerpo y el cuello del utero, y la porcion superior de lavagina, mediante la fusion de los conductos de
M€uller (parameson�efricos) derecho e izquierdo. En
torno a la semana 9 despu�es de la fecundacion, sedesintegra la pared que divide los dos conductosfusionados, lo que crea una sola luz. A veces, esta
fusion no sucede, y la persona tiene dos uteros m�aspequenos, llamado utero didelfo, y dos cuellos condos vaginas independientes, cada una con un orificio
hacia su propio utero. M�as frecuente es que la paredentre los dos conductos de M€uller permanezcaıntegra, pero las luces de los dos cuernos uterinosdesemboquen en una sola vagina comun; estasituacion recibe el nombre de utero bicorne.
El gen WNT4 regula otros genes que actuan en laaparicion de los ovarios al suprimir el gen
responsable de la produccion de los testıculos(SOX9) y activar otra serie de genes, como DAX1, unmiembro de la familia de receptores nucleares dehormonas que dirige la evolucion del ovario. Aunqueno se ha demostrado en el hombre, en ratonesDAX1controla un gen que codifica una parte de la proteınaligadora de la secuencia TATA para la polimerasade ARN. Sin esta proteına, los ratones son incapacesde constituir unos ovarios.
A lo largo de la vida fecundade unamujer, ovula cada28 dıas y expulsa unos 450 ovocitos. Como elembrion femenino no fabrica testosterona ni la
hormona antim€ulleriana, el conducto meson�efricodegenera. Los estrogenos elaborados hacen que losconductos de M€uller (conductos
parameson�efricos) generen las trompas deFalopio, el cuerpo y el cuello del utero, y parte de lavagina.
273
20
APARATO
GENITA
LFEM
ENINO
Figura 20.1 Aparato genital femenino. El ovario est�a cortado para visualizar los folıculos en desarrollo, y la trompa de Falopio, el utero
y la vagina est�an abiertos para mostrar la continuidad de su luz. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology,
3rd ed. Philadelphia, Saunders, 2007, p 464.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

FOLICULOS OVARICOS
Existen dos categorıas de folıculos ov�aricos, los folı-culos primordiales (en reposo) y los folıculos encrecimiento; estos ultimos presentan cuatro etapas:folıculo primario unilaminar, folıculo primario mul-tilaminar, folıculo secundario (antral) y folıculo deGraaf (maduro) (fig. 20.2 y tabla 20.1). De ellos, losfolıculos secundarios y de Graaf (pero no el folıculodominante) necesitan la hormona foliculoestimu-lante (FSH) para su desarrollo, mientras que el pri-mordial y los dos tipos de folıculos primarios se for-man debido a factores locales desconocidos tal vezelaborados por las c�elulas foliculares. Durante cual-quier ciclo menstrual concreto, alrededor de 50 folı-culos primordiales empiezan a evolucionar; sin embar-go, lo hacen en diversos momentos a lo largo del ciclo,por lo que en el ovario hay folıculos en distintas fasesde su produccion.
. El folıculo primordial (en reposo), el m�aspequeno y menos maduro de todos, es unconglomerado esf�erico de c�elulas compuesto por unovocito primario pequeno con un di�ametro de25mm (en la profase de la meiosis I) rodeado poruna unica capa de c�elulas foliculares planasadheridas entre sı por desmosomas. El folıculo est�aenvuelto por una l�amina basal que lo aısla delestroma de tejido conjuntivo.
. Los folıculos primarios unilaminares est�anconstituidos por un ovocito primario (con undi�ametro de 100 a 150mm) cuyo nucleo es grande ytiene un aspecto vesicular (designado vesıculagerminativa), rodeado por una sola capa de c�elulasfoliculares cubicas (v. fig. 20.2 y tabla 20.1). Losfolıculos primarios multilaminares se parecen alos folıculos primarios unilaminares, salvo en que laactividadmitotica de las c�elulas foliculares derivo enla formacion de varias capas suyas (c�elulas de lagranulosa) alrededor del ovocito. Adem�as, surgeotra capa de material amorfo, la zona pelucida,integrada por las glucoproteınas ZP1,ZP2 y ZP3, entorno al ovocito primario. No solo los filopodios delas c�elulas de la granulosa invaden la zona pelucida,entran en contacto con la membrana plasm�atica delovocito y crean uniones gap con el ovocitoprimario, sino que dichas c�elulas tambi�enestablecen uniones gap entre ellas. El estroma quecircunda las c�elulas de la granulosa, aunqueseparado de ellas por la l�amina basal, se reorganizapara originar una capa celular vascularizada internay otra capa fibrosa externa, llamadas teca interna yteca externa (v. fig. 20.2 y tabla 20.1). Las c�elulas dela teca interna presentan receptores para lahormona luteinizante (LH) ligados a sumembrana, y como sintetizan y liberan la hormonamasculina androstenediona, poseen los rasgos
estructurales ıntimos de las c�elulas que fabricanhormonas esteroideas. La androstenedionaexpulsada atraviesa la l�amina basal para penetrar enlas c�elulas de la granulosa, donde la enzimaaromatasa la convierte en estradiol, un estrogeno.
. Los folıculos secundarios (antrales) sonsemejantes a los folıculos primariosmultilaminares, excepto que los ovocitos primariosson m�as grandes (con 200mm de di�ametro [v. fig.20.2 y tabla 20.1]), la capa de c�elulas de la granulosaaumenta de espesor y entre ellas aparecen espaciosllenos de lıquido (cuerpos de Call-Exner). Sucontenido, el lıquido folicular, consiste en lıquidoextracelular al que se anaden proteınas fijadoras deesteroides procedentes de las c�elulas de la granulosa,glucosaminoglucanos, proteoglucanos y hormonascomo progesterona, estradiol, inhibina,folistatina (foliculoestatina) y activina. La activinaes una hormona que hace que las c�elulas basofilasde la adenohipofisis liberen LH y FSH. Elcrecimiento constante del folıculo secundariodepende de la FSH.
. Los folıculos de Graaf (maduros) son reconociblespor su gran tamano y por el hecho de que el lıquidofolicular, que en el folıculo secundario adoptaba elaspecto de pequenos lagos aislados, se combina enun unico gran compartimento esf�erico cargado delıquido: el antro. Su pared est�a compuesta porvarias capas de c�elulas foliculares, denominadasmembrana granulosa, y en algun punto otroagregado de c�elulas de la granulosa, el cumuloooforo, salientes hacia el antro, que recuerdan a laslenguas de Dinamarca que se adentran en el Mar delNorte. El cumulo se parece a un polo helado en elque su tallo, que nace en la membrana granulosa, escilındrico, y su extremo libre ensanchado es esf�erico.El centro del extremo libre alberga el ovocitoprimario, rodeado por numerosas capas de c�elulasde la granulosa, de las cuales la m�as interna, quelinda con la zona pelucida, recibe el nombre decorona radiada. Pueden distinguirse dos tiposdiferentes de c�elulas de la granulosa: las que formanla pared del antro, conocidas como c�elulas de lamembrana granulosa, y las del cumulo ooforo, oc�elulas del cumulo de la granulosa. Las tecasinterna y externa siguen desarroll�andoseconstituyendo capas m�as gruesas (v. fig. 20.2y tabla 20.1).
. El folıculo dominante designa aquel folıculo deGraaf que continua su evolucion, se vuelveindependiente de la FSH, y produce la hormonainhibina, que detiene la liberacion de FSH por laadenohipofisis, lo que suscita la degeneracion, oatresia, de los dem�as folıculos en progresion.Normalmente, tan solo el folıculo dominante es elque expulsa su ovocito en la ovulacion.
274
20
APARATO
GENITA
LFEM
ENINO

275
20
APARATO
GENITA
LFEM
ENINO
Figura 20.2 A y B. Desarrollo del folıculo en el ovario. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.Philadelphia, Saunders, 2007, p 465)
Tabla 20.1 FASES DEL DESARROLLO DEL FOL�ICULO OVARICO
FaseDependenciade la FSH Ovocito Zona pelucida
C�elulasfoliculares o dela granulosa
Lıquidofolicular
Tecainterna
Tecaexterna
Folıculo
primordial
No Primario Ausente Capa unica de
c�elulas planas
Ausente Ausente Ausente
Folıculo
primario
unilaminar
No Primario Presente Capa unica de
c�elulas cubicas
Ausente Ausente Ausente
Folıculo
primario
multilaminar
No Primario Presente y las
microvellosidades
del ovocito
primario forman
uniones gap con
los filopodios de
las c�elulas de la
corona radiada
Varias capas
de c�elulas
foliculares
(ahora
llamadas
c�elulas de la
granulosa)
Ausente Presente Presente
Folıculo
secundario
Sı Primario Presente con
uniones gap
Aparicion de
espacios entre
las c�elulas de la
granulosa
Acumulacion en
espacios
entre las
c�elulas de la
granulosa
Presente Presente
Folıculo
de Graaf
Sı, hasta
convertirse
en el folıculo
dominante
Primario,
rodeado por
la corona
radiada en
el cumulo
ooforo
Presente con
uniones gap
Formacion de la
membrana
granulosa y el
cumulo ooforo
Relleno del
antro
Presente Presente
Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 467.�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

OVULACION
La ovulacion sucede el dıa 14 antes de la menstrua-cion y tiene lugar en funcion de un descenso de la FSHy un subito incremento en las concentraciones sanguı-neas de LH. Estos cambios hormonales se deben a lasgrandes cantidades de estrogenos elaboradas por losfolıculos de Graaf y secundario (figs. 20.3 y 20.4). Losfolıculos dependientes de la FSH que han estadocreciendo a lo largo de su maduracion pierden suacceso a la FSH; degeneran y pasan a llamarse folıcu-los atr�esicos. La oleada en los valores de LH es laresponsable de los siguientes fenomenos:
. Aumenta la circulacion sanguınea de los ovarios,lo que lleva a la aparicion de un edema en la tecaexterna y al desprendimiento de colagenasa,histamina y prostaglandina en las inmediacionesdel folıculo dominante.
. La membrana granulosa del folıculo dominantesufre una proteinolisis debido a que se da laformacion de plasmina provocada por la LH, lo quepermite al folıculo dominante soltar su ovocito.
. Las c�elulas foliculares fabrican y emiten sustanciainductora de la meiosis, que estimula lafinalizacion de la primera division meiotica en elovocito primario del folıculo dominante, lo queorigina el ovocito secundario y el primer cuerpopolar. El ovocito secundario inicia su segundadivision meiotica pero no logra pasar de lametafase.
. Las c�elulas de la granulosa continuan sintetizandoglucosaminoglucanos y proteoglucanos, lo quedetermina una acumulacion permanente de agua yun crecimiento en el tamano del folıculodominante, que acentua su presion sobre la tunicaalbugınea del ovario, cortando el aporte de sangreen el punto de mayor presion. Esta region avascularde la c�apsula ov�arica, denominada estigma,experimenta una necrosis y, sumada a laproteinolisis de la membrana granulosa en suproximidad, abre un canal que conduce desde la luzdel antro hacia la cavidad peritoneal.
. El ovocito secundario, rodeado por la coronaradiada y en companıa del primer corpusculo polar,abandona el ovario, proceso que recibe el nombrede ovulacion, y, en condiciones normales, penetraen el infundıbulo de la trompa de Falopio(oviducto).
. El resto del folıculo dominante se convierte en elcuerpo amarillo (v. fig. 20.3).
CUERPO AMARILLO Y CUERPO BLANCO
Cuando sucede la ovulacion, el resto del folıculo deGraaf dominante almacena una pequena cantidad de
sangre procedente de los vasos sanguıneos afectadosen la zona, se pliega sobre sı mismo, y pasa a llamarsecuerpo hemorr�agico. En el plazo de un par de dıas,los macrofagos reabsorben la sangre y, debido a lainfluencia de la LH, esta estructura se transforma enuna gl�andula endocrina transitoria, el cuerpo ama-rillo (v. fig. 20.3). El cuerpo amarillo consta de dostipos celulares principales: el 80% son c�elulas luteıni-cas de la granulosa (derivadas de las c�elulas de lagranulosa) y el 20% son c�elulas luteınicas de la teca(derivadas de la teca interna). La membrana basalentre la antigua teca interna y la teca externa se desin-tegra, lo que permite la vascularizacion de la regioncorrespondiente a las c�elulas luteınicas de la granu-losa.
. Las c�elulas luteınicas de la granulosa son las m�asgrandes de las dos, y sintetizan progesteronaadem�as de modificar la androstenediona(un androgeno) en estradiol (un estrogeno).
. Las c�elulas luteınicas de la teca ocupan unasituacion perif�erica y sintetizan progesterona,androstenediona y varios estrogenos.
La progesterona y los estrogenos segregados porel cuerpo amarillo inhiben la liberacion de FSH y LHpor las c�elulas basofilas de la hipofisis, las hormonasnecesarias para la evolucion de los folıculos ov�aricosy para la conservacion del cuerpo amarillo (v. fig.20.4). Si no tiene lugar la gestacion, el cuerpo ama-rillo empieza a degenerar y se le designa cuerpoamarillo de la menstruacion. En caso de haber unembarazo, la placenta en formacion (pero al princi-pio los trofoblastos del embrion en desarrollo)suelta gonadotropina corionica humana (hCG),que hace crecer al cuerpo amarillo y lo preserva unos3 meses, perıodo en el que recibe la denominacionde cuerpo amarillo de la gestacion. Pasado esetiempo, el cuerpo amarillo ya no hace falta porquela placenta asume la elaboracion de las hormonasprecisas para mantener el embarazo, pero perma-nece siendo funcional varios meses.
A la larga, el cuerpo amarillo (de la menstruacion yde la gestacion) pierde tamano debido a su luteolisis(regresion), y los fibroblastos que lo invaden fabricanfibras de col�ageno de tipo I para convertirlo en uncuerpo blanco, un tejido conjuntivo fibroso que siguesu reabsorcion hasta quedar reducido a una cicatrizsobre la superficie del ovario. La luteolisis se pone enmarcha por la hipoxia ocasionada al bajar el flujosanguıneo, lo que suscita la llegada de linfocitos Tcuyo producto, el interferon-g , es el responsable derecolectar macrofagos. Los macrofagos descargan elfactor de necrosis tumoral-a, que empuja a las c�elu-las del cuerpo amarillo a la apoptosis.
276
20
APARATO
GENITA
LFEM
ENINO

277
20
APARATO
GENITA
LFEM
ENINO
Figura 20.3 A y B. Formacion del cuerpo amarillo y el cuerpo blanco. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology,
3rd ed. Philadelphia, Saunders, 2007, p 465.)
Figura 20.4 Interaccion entre las hormonas de la hipofisis y del ovario. LHRH, hormona liberadora de la hormona luteinizante.
(Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 472.)�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

278
20
APARATO
GENITA
LFEM
ENINO
Oviductos (trompas de Falopio)
Cada oviducto consiste en un tubulo estrecho abiertopor ambos extremos: en su extremo libre peritoneal,donde se acerca al ovario, y en su otro extremo, dondese fija a la pared del cuerpo uterino y la perfora. En sutrayecto presenta cuatro regiones nıtidamente delimi-tadas, identificables por rasgos especıficos de cada una(fig. 20.5).
. El infundıbulo es el extremo libre del oviducto, ysus fimbrias se aprietan contra el ovario durante laovulacion para captar el ovocito secundario, susc�elulas foliculares acompanantes y el primer cuerpopolar.
. La ampolla es la continuacion dilatada delinfundıbulo, habitualmente el lugar de lafecundacion.
. La region intraparietal del oviducto atraviesa lapared del utero y desemboca en su luz.
. El istmo es la parte estrecha que queda entre laregion intraparietal y la ampolla.
El oviducto posee tres capas: la mucosa interna, lamuscular intermedia y la serosa externa (ausente en laregion intraparietal).
La mucosa del oviducto est�a muy plegada, lo quevuelve su luz muy intrincada. Tiene un revestimientopor un epitelio cilındrico simple, compuesto dec�elulas en clavija y c�elulas cilındricas ciliadas.
. Las c�elulas en clavija son c�elulas cilındricas sin cilios;segregan un lıquido que no solo lleva abundanteselementos nutritivos, sino que tambi�en contienefactores necesarios para la capacitacion de losespermatozoides. Sin su presencia, estos ultimos sonincapaces de fecundar al ovocito secundario. El lıquidolleno de nutrientes alimenta no solo al embrion endesarrollo en su recorrido a trav�es del oviducto, sinotambi�en a los propios espermatozoides.
. Las c�elulas ciliadas tambi�en tienen una formacilındrica y est�an dotadas de numerosos cilios queimpulsan al ovulo fecundado hacia el utero.
La l�amina propia est�a constituida por un tejidoconjuntivo col�ageno, denso y vascular, rodeado porlamuscular, integrada por musculo liso y organizadaen una capa circular internamal delimitada y una capalongitudinal externa. Salvo en la region intraparietal,la muscular se encuentra recubierta por una serosa.
Utero
El utero es un organo muy muscular que acoge alembrion y al feto en desarrollo hasta el parto. Mide7� 4�2,5 cm y est�a dividido en tres regiones: elcuerpo, el fondo y el cuello (v. fig. 20.5). Su paredpresenta tres capas: el endometrio, el miometrio y laserosa o adventicia.
. El endometrio corresponde a la mucosa uterina yconsiste en un epitelio cilındrico simple con c�elulassecretoras sin cilios y c�elulas ciliadas que cubren altejido conjuntivo vascular en el que existenfibroblastos, c�elulas deciduales y gl�andulasuterinas tubulares y ramificadas. El endometrioposee dos capas: la capa funcional externa m�asgruesa, que se desprende durante la menstruacion yla capa basal m�as profunda que se conserva en esemomento y cuyo tejido est�a ocupado por la base delas gl�andulas uterinas. La capa basal se encuentravascularizada por las arterias rectas, mientras que lafuncional lo est�a por las arterias helicoidales; ambosvasos nacen en las arterias arqueadas del miometrio.
. El miometrio consta de musculo liso dispuesto entres capas: la interna y la externa, longitudinales, y laintermedia circular. La capa circular intermedia est�amuy vascularizada por las arterias arqueadas ymuchas veces recibe el nombre de estrato vascular.Las capas musculares quedan sustituidas en el cuellopor tejido conjuntivo fibroel�astico denso. El tamanoy el numero de los miocitos lisos miometriales sehalla bajo la influencia directa de las concentracionessanguıneas de estrogenos; cuantomayores sean estas,m�as grandes ym�as abundantes ser�an. Sin estrogenos,los miocitos lisos del utero se atrofian. Durante lagestacion, sucede lo contrario: hay una hipertrofia yuna hiperplasia de los miocitos lisos uterinos. En elmomento del parto, la hormona corticotropa haceque ellos y las c�elulas de las membranas que rodeanal feto liberen prostaglandinas, que, junto a laoxitocina procedente de la neurohipofisis,determinan las contracciones que sufre el miometriopara expulsar al feto. La salida continua de oxitocinaprovoca la contraccion de los vasos sanguıneosuterinos para reducir al mınimo la p�erdida de sangreque si no derivarıa del desprendimiento de laplacenta del revestimiento uterino.
. Gran parte del utero est�a recubierto por una serosaexcepto la zona en la que se pega a la vejiga urinaria.En esa region, existe una adventicia tapiz�andolo.
El cuello es una estructura muy fibrosa que sobre-sale en la vagina como el final del utero. Su luz est�arevestida por un epitelio cilındrico simple integradopor c�elulas secretoras de moco. Cuando penetra en lavagina, queda recubierto por un epitelio pavimentosoestratificado no queratinizado. El tejido conjuntivosubepitelial del cuello alberga gl�andulas cervicalessecretoras de moco cuyo producto puede facilitar laentrada de los espermatozoides –como sucede du-rante la ovulacion– o generar un taponmucoso espeso–como sucede durante la gestacion– para impedir lallegada de espermatozoides a la luz del utero. El cam-bio de consistencia en el moco elaborado por estasgl�andulas est�a controlado por la concentracion san-guınea de progesterona.
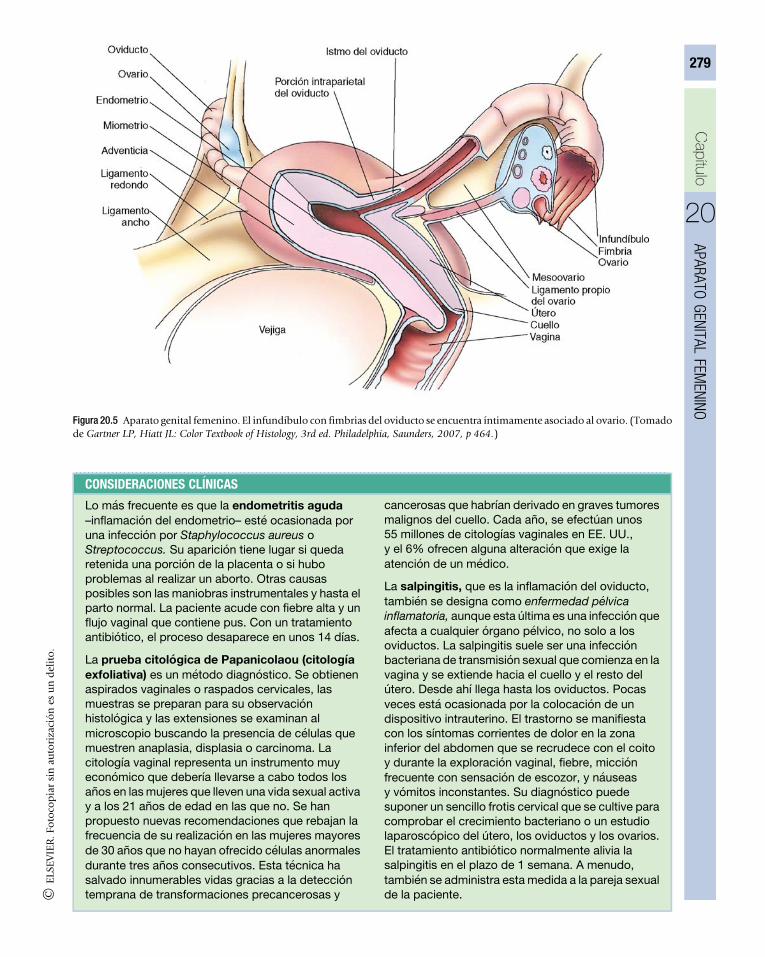
CONSIDERACIONES CL�INICAS
Lo m�as frecuente es que la endometritis aguda–inflamacion del endometrio– est�e ocasionada poruna infeccion por Staphylococcus aureus oStreptococcus. Su aparicion tiene lugar si quedaretenida una porcion de la placenta o si huboproblemas al realizar un aborto. Otras causasposibles son las maniobras instrumentales y hasta elparto normal. La paciente acude con fiebre alta y unflujo vaginal que contiene pus. Con un tratamientoantibiotico, el proceso desaparece en unos 14 dıas.
La prueba citologica de Papanicolaou (citologıaexfoliativa) es un m�etodo diagnostico. Se obtienenaspirados vaginales o raspados cervicales, lasmuestras se preparan para su observacionhistologica y las extensiones se examinan al
microscopio buscando la presencia de c�elulas quemuestren anaplasia, displasia o carcinoma. Lacitologıa vaginal representa un instrumento muyeconomico que deberıa llevarse a cabo todos losanos en lasmujeres que lleven una vida sexual activay a los 21 anos de edad en las que no. Se hanpropuesto nuevas recomendaciones que rebajan lafrecuencia de su realizacion en las mujeres mayores
de 30 anos que no hayan ofrecido c�elulas anormales
durante tres anos consecutivos. Esta t�ecnica hasalvado innumerables vidas gracias a la detecciontemprana de transformaciones precancerosas y
cancerosas que habrıan derivado en graves tumoresmalignos del cuello. Cada ano, se efectuan unos55 millones de citologıas vaginales en EE. UU.,y el 6% ofrecen alguna alteracion que exige la
atencion de un m�edico.
La salpingitis, que es la inflamacion del oviducto,
tambi�en se designa como enfermedad p�elvicainflamatoria, aunque esta ultima es una infeccion que
afecta a cualquier organo p�elvico, no solo a losoviductos. La salpingitis suele ser una infeccionbacteriana de transmision sexual que comienza en lavagina y se extiende hacia el cuello y el resto delutero. Desde ahı llega hasta los oviductos. Pocas
veces est�a ocasionada por la colocacion de undispositivo intrauterino. El trastorno se manifiestacon los sıntomas corrientes de dolor en la zonainferior del abdomen que se recrudece con el coitoy durante la exploracion vaginal, fiebre, miccion
frecuente con sensacion de escozor, y n�auseasy vomitos inconstantes. Su diagnostico puedesuponer un sencillo frotis cervical que se cultive paracomprobar el crecimiento bacteriano o un estudiolaparoscopico del utero, los oviductos y los ovarios.El tratamiento antibiotico normalmente alivia lasalpingitis en el plazo de 1 semana. A menudo,
tambi�en se administra estamedida a la pareja sexualde la paciente.
279
20
APARATO
GENITA
LFEM
ENINO
Figura 20.5 Aparato genital femenino. El infundıbulo con fimbrias del oviducto se encuentra ıntimamente asociado al ovario. (Tomado
de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 464.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

280
20
APARATO
GENITA
LFEM
ENINO
Ciclo menstrual
En condiciones normales, el ciclo menstrual dura28 dıas y se divide en tres fases continuas: menstrual,proliferativa (folicular) y secretora (luteınica). Su apari-cion tiene lugar cuando la mujer no est�a embarazada;el ciclo se interrumpe al comenzar una gestacion.
. La fasemenstrual supone de 3 a 4 dıas. Empieza conel comienzo de la menstruacion (hemorragiavaginal) y es el resultado de un descenso en lasconcentraciones sanguıneas de estrogenos yprogesterona que suscita una contraccion de lasarterias helicoidales, al principio espor�adica, eininterrumpida en cuestion de 24 a 48h (figs. 20.6 y20.7). La privacion de riego sanguıneo en la capafuncional ocasiona una necrosis y, cuando serompen las arterias helicoidales, su desprendimientocorrespondiente. La capa basal sigue recibiendo lallegada de sangre desde las arterias rectas y, aunquequede despojada de su cubierta epitelial, permanecesana. Las c�elulas epiteliales situadas en la base de lasgl�andulas uterinas ponen en marcha lareepitelizacion de la superficie perdida.
. La fase proliferativa (folicular), de unos 10 dıas deduracion, se inicia al acabar la menstruacion y seextiende hasta la ovulacion. Esta fase se caracterizapor la reepitelizacion de la superficie desprendida,la reconstruccion del endometrio y las arteriashelicoidales en su espesor, y el restablecimiento delas gl�andulas, por lo que se recupera la capafuncional hasta completar unos 2 o 3mm de grosor(v. fig. 20.6). En el momento de la ovulacion, lasgl�andulas, aunque no formen espirales, empiezan aacumular glucogeno en sus c�elulas. Las arteriashelicoidales apenas han comenzado a enroscarse enuna h�elice cerrada y han cubierto alrededor de lasdos terceras partes de su trayecto hacia la capafuncional. Justo antes de la ovulacion, lasconcentraciones sanguıneas de FSH, LH yestrogenos llegan a su m�aximo.
. La fase secretora (luteınica), que se prolonga unas2 semanas, arranca tras la ovulacion y finaliza alaparecer la menstruacion. A lo largo de esta fase, lasgl�andulas del endometrio adquieren muchasespirales; los vasos helicoidales ocupan toda ladistancia que hay hasta la cara de la capa funcional,que ha crecido hasta unos 5mm de anchura,cubierta por epitelio; y la luz de las gl�andulasendometriales est�a rellena con su producto desecrecion (v. fig. 20.6). Para el dıa 20 del ciclomenstrual, la concentracion sanguınea deprogesterona ha alcanzado su m�aximo, y la de losestrogenos tambi�en es bastante alta, aunque notanto como durante la fase proliferativa (v. fig.20.7). Las arterias helicoidales se ven rodeadas porc�elulas del estroma que aumentan y se transformanen c�elulas deciduales (reaccion decidual) que
almacenan glucogeno y lıpidos en prevision de laimplantacion del embrion. La funcion de estasc�elulas deciduales anticip�andose a la implantacion ya lo largo de este proceso se comenta m�as adelanteen el apartado dedicado a la implantacion.
Fecundacion
Cuando el ovocito secundario, sus c�elulas foliculares yel primer cuerpo polar se expulsan del folıculo de Graafdominante, se introducen en el infundıbulo con fim-brias del oviducto y son transportados por la actividadmuscular de la capa correspondiente de este organo ymediante las acciones coordinadas de los cilios perte-necientes a las c�elulas cilındricas ciliadas que hay en ella(fig. 20.8). Si han entrado espermatozoides en el apa-rato genital de la mujer, han sufrido su maduracion enel epidıdimoy su capacitacion en el istmo, y han llegadohasta la ampolla del oviducto, comienzan a abrirse pasoen su recorrido entre las c�elulas de la corona radiadapara entrar en contacto con la zona pelucida.
Cuando los receptores de ZP3 de la membranaplasm�atica de los espermatozoides se unen a estamol�ecula en la zona pelucida, el espermatozoideexperimenta la reaccion acrosomica, lo que liberalas enzimas presentes en su acrosoma –en especial,acrosina– como medio que le deja traspasar la zonapelucida y ponerse en contacto con lamembrana plas-m�atica del ovocito. Este hecho hace que el ovocitosuelte sus enzimas lisosomicas, quemodifican la zonapelucida en la reaccion de zona, y la impermeabilizafrente a los dem�as espermatozoides. La membranaplasm�atica del espermatozoide posee fertilina, queinteracciona con las integrinas y el CD9 de la mem-brana plasm�atica del ovocito, lo que permite que lafusion entre ambas membranas. El espermatozoidepenetra en el ovocito, un fenomeno denominadofecundacion. En ese momento, el ovocito:
. Atraviesa la reaccion cortical, que impide lairrupcion de m�as espermatozoides en el ovocito.
. Reanuda su segunda divisionmeiotica, lo que formael ovulo y el minusculo segundo cuerpo polar.
. Reconfigura su nucleo haploide, el pronucleofemenino, que se desplaza hacia el nucleo haploidedel espermatozoide (pronucleo masculino).
Los dos pronucleos duplican su ADN, y sus mem-branas nucleares se disuelven. El complejo del husoque surge cuando los centrıolos emigran hacia lospolos opuestos del ovulo ahora es ya una c�eluladiploide, llamada cigoto. Este ultimo efectua unamitosis, que recibe el nombre de segmentacion, ylas c�elulas resultantes avanzan a lo largo del oviductohacia el utero (v. fig. 20.8) y siguen dividi�endose,hasta constituir un conglomerado celular conocidocomo morula. Si el ovocito no se fecunda, degeneraen un plazo de 24h.

281
20
APARATO
GENITA
LFEM
ENINO
Figura 20.6 Las dos capas del endometrio uterino y su
vascularizacion. (Tomado de Gartner LP, Hiatt JL:Color Textbook of Histology, 3rd ed. Philadelphia,
Saunders, 2007, p 477.)
Figura 20.7 Relaciones entre los acontecimientos
ov�aricos, las fases del endometrio uterino y el ciclohormonal. (Tomado de Gartner LP, Hiatt JL:
Color Textbook of Histology, 3rd ed. Philadelphia,
Saunders, 2007, p 479.)
Figura 20.8 Fecundacion y mitosis en el cigoto.
(Modificado de Gartner LP, Hiatt JL: Color Textbook of
Histology, 3rd ed. Philadelphia, Saunders, 2007, p 481.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Implantacion
Cuando lamorula viaja a trav�es del oviducto, todavıa seencuentra rodeada por la zona pelucida, y sus c�elulas,los blastomeros, no han dejado de dividirse. De 4 a5 dıas despu�es de la fecundacion, la morula llega alutero (fig. 20.9). El lıquido uterino accede a la zonapelucida y reorganiza las c�elulas de la morula paraoriginar el blastocisto, cuya luz, el blastocele, contienelıquido uterino y un pequeno agregado de c�elulas, losembrioblastos (masa celular interna). Las c�elulasperif�ericas que integran la pared del blastocisto sedenominan trofoblastos (v. fig. 20.9). La zona pelucidase desintegra, y los trofoblastos expresan L-selectinas eintegrinas sobre sus superficies, que entran en contactocon receptores del epitelio uterino, poniendo enmarchael proceso de la implantacion. El endometrio, en la fasesecretora (luteınica), est�a preparado para alimentar alembrion al hundirse en la pared uterina.
. Las c�elulas de los embrioblastos dan lugar alembrion y al amnios.
. Los trofoblastos generan la porcion embrionaria dela placenta y hacen que el endometrio uterinoproduzca la porcion materna de la placenta.
Cuando los trofoblastos proliferan, configuran unacapa interna de citotrofoblastos cuyas c�elulas experi-mentan una en�ergica fase de division y una capaexterna de sincitiotrofoblastos sin mitosis. A medidaque se dividen las c�elulas del citotrofoblasto, las reci�enformadas se incorporan a la capa del sincitiotrofo-blasto, que crece, sufre una vacuolizacion para crearlagunas interconectadas, y traspasa el revestimientoendometrial. Al concluir el dıa 11 tras la fecundacion,el embrion y sus capas han quedado sumergidas en elendometrio vascularizado (fig. 20.10; v. fig. 20.9).
Formacion de la placenta
Amedida que los sincitiotrofoblastos continuan infil-trando el endometrio vascular, atraviesan las paredesde muchos de los vasos sanguıneos y la sangre salehacia las lagunas, para abastecer al embrion incipientecon alimentos y oxıgeno. Segun evoluciona la pla-centa, las c�elulas de los trofoblastos constituyen elcorion. Enmomentos posteriores, el corion engendrala l�amina corionica, de la que surgen las vellosidadescorionicas (v. fig. 20.10). Como respuesta a la apari-cion del corion, el endometrio uterino pasa a llamarsedecidua. La decidua posee tres regiones:
. La decidua basal es la region de la decidua quecorresponde a la porcion materna de la placenta.
. La decidua capsular es la parte de la decidua situadaentre el embrion y la luz del utero; no aporta nada ala placenta y recibe el nombre de corion leve.
. La decidua parietal es la zona de la decidualocalizada entre el miometrio y la luz del utero: lasregiones endometriales que no se hallan en ıntimaasociacion con el embrion ni con la placenta.
La decidua basal se ve invadida por el aporte vas-cular materno para configurar la porcion materna dela placenta, y los sincitiotrofoblastos y citotrofoblas-tos de la l�amina corionica responden mediante lageneracion de las vellosidades corionicas, conocidascomo vellosidades primarias (del corion frondoso).Cuando el nucleo de las vellosidades primarias sepuebla de c�elulas mesenquimatosas y por los vasossanguıneos embrionarios, estas pasan a denominarsevellosidades secundarias (v. fig. 20.10). Los citotro-foblastos de las vellosidades secundarias bajan denumero al comenzar a formar parte del sincitio enexpansion. A la vez que sucede esto, aparecen lagunasllenas de sangre en la decidua basal, y las vellosidadessecundarias sobresalen en estos grandes espacios vas-culares. Las lagunas reciben sangre desde las arteriolasy v�enulas maternas y allı la vierten. Parte de las vello-sidades est�an sujetas a la decidua basal y se las designavellosidades de fijacion, mientras que otras no loest�an y se las conoce como vellosidades libres.
Los lechos capilares de las vellosidades se acercan alos sincitiotrofoblastos, y los nutrientes y el oxıgeno dela sangre materna se adentran en los tejidos de estasvellosidades para atravesar sus lechos capilares. Los pro-ductos de desecho y el dioxido de carbono salen de loscapilares fetales para recorrer su trayecto desde las ve-llosidades hacia las lagunas con el fin de que los retire lasangrematerna. El riego sanguıneo fetal y elmaterno noentran en contacto; el tejido interpuesto entre ambascirculaciones se llama barrera placentaria (tabla 20.2).
La mayorıa de las pequenas mol�eculas pueden cru-zar esta barrera, pero solo unas cuantas macro-mol�eculas son capaces de hacerlo. Los antibioticosmaternos se transportan a trav�es de la placenta segununa endocitosis mediada por receptores, y los ionesy la glucosa emplean el transporte activo (iones) y ladifusion facilitada (glucosa). Los sincitiotrofoblastosno solo intervienen en la constitucion de la barreraplacentaria, sino que tambi�en segregan hCG (paraconservar el cuerpo amarillo), estrogenos, progeste-rona, tirotropina corionica y somatomamotropinacorionica. Las c�elulas deciduales del estroma deci-dual sintetizan prostaglandinas y prolactina.
282
20
APARATO
GENITA
LFEM
ENINO
Tabla 20.2 COMPONENTES DE LA BARRERAPLACENTARIA
Endotelio de los capilares fetalesL�amina basal de los capilares fetales
L�amina basal de los citotrofoblastos
Citotrofoblastos
Sincitiotrofoblastos

283
20
APARATO
GENITA
LFEM
ENINO
Figura 20.9 Proceso de implantacion y formacion de los citotrofoblastos y sincitiotrofoblastos. (Tomado de Gartner LP, Hiatt JL: ColorTextbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 481.)
Figura 20.10 Proceso de formacion del corion y la decidua.
(Tomado de Gartner LP, Hiatt JL: Color Textbook ofHistology, 3rd ed. Philadelphia, Saunders, 2007, p 483.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Vagina y genitales externos
La vagina (fig. 20.11), una cubierta fibromuscular de8 a 9 cm de longitud, dotada de tres capas y situadaentre el vestıbulo y el utero, est�a compuesta de:
. Mucosa, revestida por un epitelio pavimentosoestratificado no queratinizado cuyas c�elulasposeen receptores para los estrogenos, que alocuparse, hacen que estas c�elulas formen yalmacenen glucogeno. El glucogeno descargado enla luz vaginal lo metaboliza la flora bacterianaendogena para producir �acido l�actico, y al bajar elpH vaginal, protege a la vagina de las bacteriaspatogenas.
. L�amina propia, un tejido conjuntivo fibroel�asticomuy vascular con una dotacion abundante delinfocitos y neutrofilos. Durante la excitacionsexual, los capilares liberan plasma que difundehacia la luz y se combina con las secrecionescervicales como ayuda para lubricar la paredvaginal.
. Los miocitos lisos de lamuscular, que adoptan unadisposicion longitudinal entremezclados conalgunas fibras de orientacion circular. El orificioexterno de la vagina tiene fibrasmusculares lisas quese despliegan en sentido circular para configurarun esfınter.
. Adventicia, un tejido conjuntivo fibroel�asticodenso que presenta un plexo venoso muydesarrollado y una profusion de fibras nerviosassimp�aticas procedentes de los nervios espl�acnicosp�elvicos.
. El himen, una cubierta de tejido conjuntivodelgado, tapizada por un epitelio por ambas caras,que limita el orificio vaginal en las vırgenes.
GENITALES EXTERNOS
Los genitales externos (vulva) constan de:
. Los labios mayores, el equivalente al escrotomasculino, un par de pliegues cut�aneos rellenos degrasa, ampliamente surtidos de gl�andulassudorıparas y seb�aceas. Su cara interna carece depelo, mientras que la externa est�a cubierta por elvello pubico (v. fig. 20.11).
. Los labios menores, tendidos entre los labiosmayores, un par de pliegues de piel sin pelo m�aspequenos, cuyo nucleo de tejido conjuntivo est�amuy inervado y abundantemente vascularizado(v. fig. 20.11).
. El espacio que queda entre los dos labiosmenores, llamado vestıbulo. Est�a humedecidopor las gl�andulas vestibulares menores situadasen sus paredes y por las gl�andulas de Bartholin.
La vagina y la uretra desembocan en el vestıbulo(v. fig. 20.11).
. Los dos labios menores se reunen por su partesuperior para formar el prepucio sobre el reducidoglande del clıtoris, la cara externa del pequenoclıtoris, que es el equivalente al pene (v. fig. 20.11).Los dos cuerpos er�ectiles que componen el clıtorisreciben un gran aporte neurovascular y son muysensibles a la estimulacion sexual.
Mama
Las mamas masculinas y femeninas son id�enticashasta la pubertad, cuando la progesterona y los estro-genos ov�aricos inducen un mayor desarrollo en lamujer por la aparicion de los lobulillos y los conduc-tillos terminales (fig. 20.12). Las mamas femeninascrecen de tamano al acumular tejido adiposo y tejidoconjuntivo hasta alrededor de los 20 anos de edad.Durante la gestacion, la placenta produce estrogenos yprogesterona m�as prolactina por la adenohipofisis yglucocorticoides y somatotropina, que provocan unnuevo aumento para preparar las gl�andulas mamariaspara la elaboracion de leche con la que alimentar alreci�en nacido. Tras el parto, los estrogenos y la pro-gesterona proceden de los ovarios. La gl�andulamamaria, una gl�andula tubuloalveolar compuesta,est�a constituida por 15 a 20 lobulos, en los que laporcion dilatada del conducto galactoforo de cadalobulo, denominado seno galactoforo, se estrechaal atravesar y desembocar en la superficie del pezon.El conducto y el seno galactoforos est�an revestidos porun epitelio cubico estratificado, y los conductos m�aspequenos lo est�an por un epitelio cubico simple.Todos los conductos se encuentran rodeados por va-rias c�elulas mioepiteliales que est�an instaladas entrela l�amina basal y las c�elulas de los conductos. Lasgl�andulas mamarias de las mujeres pospuberales per-manecen en el estado de reposo (inactivo), a no serque la mujer est�e embarazada, cuando se dice queest�an en el estado lactante (activo). Las gl�andulasen reposo poseen un pequeno punado de c�elulas,llamado yemas alveolares, al final de los conductosgalactoforos y sus ramas m�as pequenas. Bajo la in-fluencia de la progesterona, las yemas alveolares seextienden, y durante la gestacion la estimulacionsuplementaria facilitada por los lactogenos y estro-genos placentarios ocasiona la formacion de alv�eolosque generan calostro (una secrecion espesa cargada deproteınas e inmunoglobulinas), y transcurridos unosdıas desde el parto, la prolactina y los estrogenosmaternos desencadenan la fabricacion de leche. Esteproceso es constante, y su extraccion por el lactante, elreflejo de emision de la leche, est�a causado por laoxitocina de la neurohipofisis.
284
20
APARATO
GENITA
LFEM
ENINO
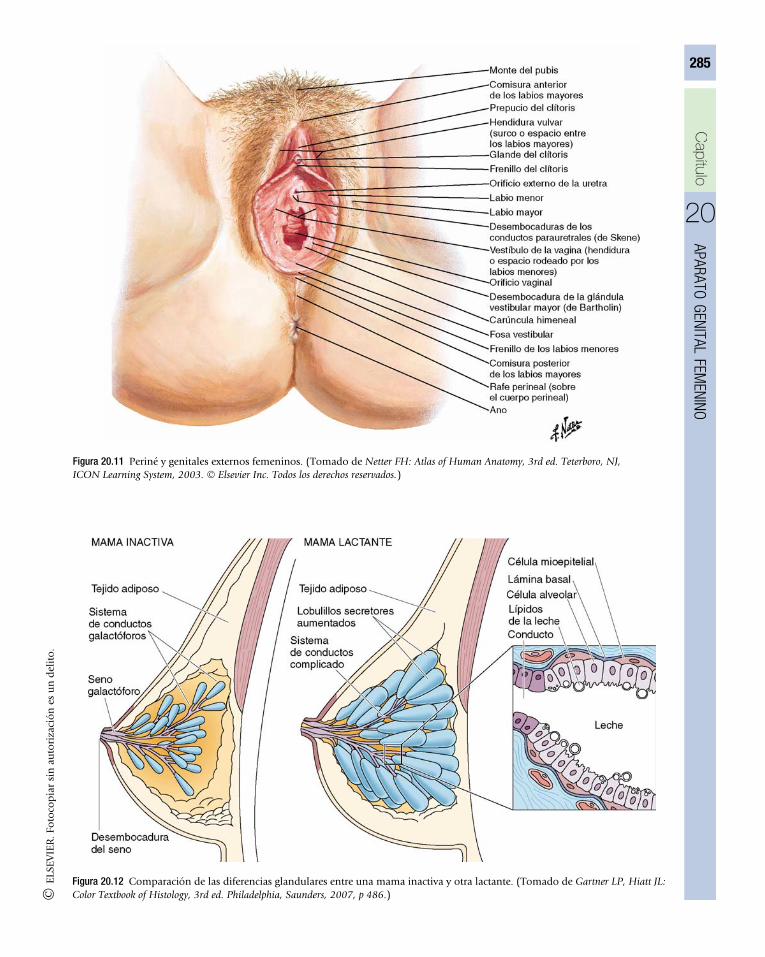
285
20
APARATO
GENITA
LFEM
ENINO
Figura 20.11 Perin�e y genitales externos femeninos. (Tomado de Netter FH: Atlas of Human Anatomy, 3rd ed. Teterboro, NJ,ICON Learning System, 2003. � Elsevier Inc. Todos los derechos reservados.)
Figura 20.12 Comparacion de las diferencias glandulares entre una mama inactiva y otra lactante. (Tomado de Gartner LP, Hiatt JL:
Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 486.)�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

21 APARATO GENITALMASCULINO
El aparato genital masculino comprende los testıcu-los, las vıas genitales, el escroto, el pene y las gl�andu-las accesorias: las vesıculas seminales, la prostata ylas gl�andulas bulbouretrales (deCowper). Estas gl�andulas segreganlos componentes no celulares delsemen que alimentan a los esper-matozoides y ofrecen un mediolıquido para la salida del semen.El pene no solo es el organo deexpulsion del semen hacia el apa-rato genital femenino, sino quetambi�en saca la orina fuera delorganismo. Los testıculos formanlos espermatozoides (los gametosmasculinos) y sintetizan, almace-nan y liberan testosterona, la hor-mona sexual masculina (fig. 21.1).
Testıculos
Los testıculos (fig. 21.2) son losorganos sexuales masculinos paressituados en el escroto, que producenespermatozoides y testosterona. Cada testıculo de unadulto mide en torno a 4 cm de largo, 2 o 3 cm deancho y 3 cm de espesor. Durante la embriogenia,estos organos aparecen en la pared abdominal poste-rior detr�as del peritoneo y descienden a trav�es de lapared del abdomen hacia el escroto, arrastrando lascubiertas de dicha pared con ellos, que se conviertenen las tunicas testiculares.
. La tunica vaginal es un saco seroso derivado delperitoneo que pr�acticamente encierra el testıculo yle permite un cierto grado de movilidad en elinterior del escroto.
. La tunica albugınea es la c�apsula de tejidoconjuntivo col�ageno del testıculo.
. La tunica vascular es la c�apsula vascular de lostestıculos.
. El mediastino testicular representa la porcionposterior engrosada de la tunica albugınea quealberga la red testicular.
. Los lobulillos testiculares constan deunos 250 compartimentospiramidales que alojan un tejidoconjuntivo vascular que envuelve alos tubulos seminıferos, donde seoriginan los espermatozoides, y a lasgl�andulas intersticiales (de Leydig),que elaboran testosterona.
En cada lobulillo hay de uno a cuatrotubulos seminıferos muy intrinca-dos y con un extremo ciego, cadauno revestido por un epitelio semi-nıfero cuya funcion consiste en lafabricacion de espermatozoides. Unavez generados y desprendidos del epi-telio seminıfero, los espermatozoidespenetran en los tubulos rectos queconectan los extremos abiertos delos tubulos seminıferos con una seriede espacios laberınticos dentro delmediastino llamados red testicular.
Los espermatozoides llegan a 10 o 20 tubulos cor-tos, los conductillos eferentes, y desde ahı pasan alepidıdimo.El par de arterias testiculares, procedentes de la
aorta, siguen a los testıculos y al conducto deferentehacia el escroto, para suministrar su aporte vascular acada testıculo. Al acercarse a ellos, las arterias testicularesse vuelven muy enrevesadas y quedan rodeadas por elplexo venoso pampiniforme. Todas estas vueltas m�asel plexo de venas constituyen un sistemade intercambiode calor por contracorriente entre estos vasos, que bajala temperatura de la sangre arterial a 35 �C, algo impres-cindible para la produccion de espermatozoides viables.En conjunto, la arteria testicular, el plexo venoso pam-piniforme y el conducto deferente integran el cordonesperm�atico, que recorre el conducto inguinal.
� 2011 Elsevier Espana, S.L. Reservados todos los derechos
TERMINOS CLAVE. Tubulos seminıferos. C�elulas de Sertoli. Espermatogonias. Formacion de los
espermatozoides. C�elulas intersticiales
de Leydig. Conductos genitales
masculinos. Gl�andulas genitales
accesoriasmasculinas
. Mecanismo de laereccion
286

CONSIDERACIONES CL�INICAS
La hipertermia es un factor principal que deriva en
una esterilidad masculina. Los estudios m�asrecientes llevados a cabo con hombres mientras
usaban un port�atil con el ordenador situado sobre suregazo durante 1 h como mınimo, observaron queeste contacto ocasionaba un ascenso de 2,8 �C en latemperatura intraescrotal. Estos trabajos no sonconcluyentes, pero resulta recomendable que losninos y los jovenes eviten el empleo de ordenadores
port�atiles sobre su regazo durante un perıodoprolongado.
La criptorquidia es un defecto cong�enito en el queno baja hasta el escroto un testıculo o los dos.
Cuando no falla m�as que uno, el esperma deltestıculo descendido suele ser normal y fecundo.
Cuando fallan los dos, el paciente es est�eril porque latemperatura normal de su organismo inhibe la
espermatogenia. Puede recurrirse a t�ecnicasquirurgicas para corregir este problema, pero aveces el esperma es anormal. La mutacion de dos
genes, el factor similar a la insulina 3 yHOXA10, est�arelacionada con la criptorquidia bilateral. Existe unagran incidencia de tumores testiculares asociados ala criptorquidia testicular no tratada. Laadministracion de hormonas es capaz de inducirel descenso, pero si fracasa, se recomienda lacirugıa.
287
21
APARATO
GENITA
LMASCULIN
O
Figura 21.2 Testıculo y epidıdimo. Los lobulillos y su contenido noest�an dibujados a escala. (Tomado de Gartner LP, Hiatt JL: Color
Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 490.)
Figura 21.1 Aparato genital
masculino. (Tomado de Gartner
LP, Hiatt JL: Color Textbook ofHistology, 3rd ed. Philadelphia,
Saunders, 2007, p 490.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

TUBULOS SEMINIFEROS
Cada testıculo posee alrededor de 500 tubulos semi-nıferos que forman espermatozoides (30 a 70 cm delongitud y 150 a 250mm de di�ametro) sumergidos enun tejido conjuntivo vascular laxo. La pared de tejidoconjuntivo de cada tubulo seminıfero, denominadatunica propia, est�a rodeada por una l�amina basal. Elgrueso epitelio seminıfero (epitelio germinal) est�acompuesto por dos tipos epiteliales diferentes: lasc�elulas de Sertoli (de sost�en) y las espermatogoniasque se encuentran en proceso de diferenciacion paraconfigurar espermatozoides.
Las c�elulas de Sertoli (fig. 21.3) son c�elulas cilın-dricas altas que presentan grandes nucleos transpa-rentes y hendidos, abundantes mitocondrias, un retı-culo endopl�asmico liso bien desarrollado, aparato deGolgi, endolisosomas y numerosos elementos citoes-quel�eticos. Las zonas de oclusion establecidas entrelas c�elulas de Sertoli adyacentes subdividen la luz deltubulo seminıfero en:
. Un compartimento basal, basal respecto a lasuniones estrechas, que se halla expuesto al tejidoconjuntivo vascular subyacente.
. Un compartimento yuxtaluminal, aislado deltejido conjuntivo vascular, que crea una barrerahematotesticular y protege los gametosen progresion frente a su contacto conel sistema inmunitario, que si no crearıa unarespuesta inmunitaria contra ellos.
Las funciones de las c�elulas de Sertoli son lassiguientes:
. Sostener, proteger y alimentar las espermatogoniasen crecimiento
. Fagocitar los restos celulares (cuerpos residuales)desechados durante el proceso de la espermiogenia
. Facilitar la salida de las esperm�atidas maduras haciala luz de los tubulos seminıferos a trav�es de sucontraccion mediante la actina (espermiacion)
. Segregar:. Proteına fijadora de androgenos (ABP) hacia laluz del tubulo seminıfero, lo que eleva laconcentracion de testosterona en su interior
. Inhibina, que dificulta la liberacion de FSH
. Lıquido cargado de fructosa, que alimenta losespermatozoides y los transporta a lo largo de lasvıas genitales
. Transferrina testicular como refuerzo parasuministrar hierro a los gametos en maduracion
. Hormona antim€ulleriana, durante el desarrolloembrionario, que impide la formacion delaparato genital femenino y permite la apariciondel masculino.
Espermatogonias
La mayor parte de las c�elulas que componen el epitelioseminıfero son espermatogonias (fig. 21.4; v. fig. 21.3)que llevan a cabo el mecanismo de generacion de losespermatozoides a trav�es de las tres fases siguientes: es-permatocitogenia, la creacion de los espermatocitos;meiosis, la configuracion de esperm�atidas haploides apartir de espermatocitos primarios diploides; y esper-miogenia, la transformacion de las esperm�atidas enespermatozoides maduros.
. La espermatocitogenia, la produccionde espermatocitos primarios a partir de lasespermatogonias, sucede en el compartimentobasal del tubulo seminıfero. Las espermatogoniasdiploides asientan sobre la l�amina basal, y lapresencia de la hormona testosterona les haceponer en marcha su actividad mitotica.Los tres tipos diferentes de espermatocitos sonlos siguientes:. Las espermatogonias oscuras de tipo A,las menos maduras, son c�elulas de reservacuyos nucleos ovalados tienen muchaheterocromatina, lo que otorga a la c�elulaun aspecto oscuro. Estas c�elulas entran en elciclo mitotico para originar m�as c�elulas oscurasde tipo A.
. Las espermatogonias p�alidas de tipo A,cuyos nucleos ovalados poseen muchaeucromatina, que les confiere un aspectom�as p�alido; la testosterona empuja a estasc�elulas a sufrir una r�apida division celularpara engendrar m�as espermatogonias de tipo A.
. Las espermatogonias de tipo B, cuyos nucleosredondos les distinguen de sus precursores. Estasc�elulas tambi�en inician el ciclo mitotico para darlugar a espermatocitos primarios.
Mientras estas c�elulas experimentan sus divisionescelulares, mantienen el contacto entre sı por medio deprolongaciones citopl�asmicas, que generan un gransincitio.
288
21
APARATO
GENITA
LMASCULIN
O

289
21
APARATO
GENITA
LMASCULIN
O
Figura 21.4 Espermatogenia que ofrece los puentes intercelulares que contiene el sincitio durante la diferenciacion y la maduracion.
(Modificado de Ren X-D, Russell L: Clonal development of interconnected germ cells in the rat and its relationship to the segmental and
subsegmental organization of spermatogenesis. Am J Anat 192:127, 1991.)
Figura 21.3 Epitelio seminıfero. (Tomado de Gartner LP, Hiatt JL: Color Textbookof Histology, 3rd ed. Philadelphia, Saunders, 2007, p 492.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

. Lameiosis es una division reductora que hace surgirc�elulas haploides.. El gran sincitio de los espermatocitos primariosdiploides emigra desde el compartimento basalhacia el yuxtaluminal del tubulo seminıfero yatraviesa la primera division meiotica.
. Los espermatocitos secundarios est�an en elcompartimento yuxtaluminal, y se someten a lasegunda division meiotica, que deriva en lasesperm�atidas.
. Las esperm�atidas, c�elulas haploides, est�ansostenidas por las c�elulas de Sertoli mientrasrecorren la fase final de la espermatogenia.
. La espermiogenia es la fase de la espermatogenia enla que las esperm�atidas pierden gran parte de sucitoplasma y se transforman en espermatozoides. Lascuatro fases de la espermiogenia son la fase de Golgi,la del capuchon, la acrosomica y la de maduracion.. La fase de Golgi, como su nombre indica,conlleva la introduccion de las enzimashidrolıticas en vesıculas por el aparato de Golgi,que se fusionan entre sı para formar lavesıcula acrosomica que contiene el gr�anuloacrosomico; adem�as, el axonema flagelar yla pieza de conexion est�an en procesode configuracion.
. La vesıcula acrosomica no solo crece durante lafase de capuchon, sino que tambi�en se fija a lamembrana nuclear y la envuelve en parte,pasando a llamarse acrosoma.
. El nucleo de la esperm�atida se aplana y se reducede tamano, la c�elula en su integridad se alarga y lasmitocondrias se reunen en un punto durante lafase acrosomica. Adem�as, aparece un manguito,una acumulacion cilındrica transitoria demicrotubulos, lo que provoca que la esperm�atidaaumente m�as de longitud a medida que seconvierte en un espermatozoide. Cuando elmanguito se disocia, se crea el anillo, que senalala union entre las piezas principal y media delespermatozoide en desarrollo. Las mitocondriasse congregan en la pieza media, y nacen las fibrasdensas externas y la vaina fibrosa.
. La fase final de la espermiogenia es la fase demaduracion, cuando las esperm�atidas descargansu exceso de citoplasma, lo que liberaespermatozoides independientes del sincitio.Los espermatozoides son inmoviles hasta que
sufren la capacitacion en el aparato genitalfemenino. Las c�elulas de Sertoli fagocitan losrestos celulares; este fenomeno se denominaespermiacion.
Espermatozoides
Los espermatozoides son c�elulas haploides quemiden unos 65mm de largo y constan de una cabezay una larga cola. La cabeza del espermatozoide (fig.21.5) no llega a 5mm de longitud y alberga el nucleohaploide y el acrosoma lleno de enzimas que comu-nica con las membranas nuclear y plasm�atica. Segunse describio en el capıtulo 20, cuando el espermato-zoide entra en contacto con lamol�ecula ZP3 en la zonapelucida que rodea al ovulo, experimenta la reaccionacrosomica, con lo que suelta las enzimas neuramini-dasa, hialuronidasa, arilo sulfatasa, acrosina (unaenzima similar a la tripsina) y fosfatasa �acida, alojadasen el acrosoma. Estas enzimas degradan la zona pelu-cida en el trayecto del espermatozoide, lo que le faci-lita alcanzar el ovulo y fecundarlo.
Cola del espermatozoide
Cuatro regiones distintas constituyen la cola del esper-matozoide (v. fig. 21.5):
. El cuello, interpuesto entre la cabeza y la cola, est�aintegrado por la pieza de conexion cuyas nuevecolumnas, alrededor de los centrıolos, persistencomo las nueve fibras densas externas.
. La pieza media conecta el cuello con la piezaprincipal y acaba en el anillo. Est�a compuesta por lavaina mitocondrial que envuelve a las fibrasdensas externas y el axonema central.
. La pieza principal empieza en el anillo y acaba en lapieza terminal. Est�a formada por el axonemaencerrado por las siete fibras densas externas que seencuentran cercadas por una vaina fibrosa. Cercade su terminacion distal, la pieza principal de 45mmde largo reduce su di�ametro porque desaparecen lasfibras densas externas y la vaina fibrosa.
. La pieza terminal representa el extremo caudal delespermatozoide. Corresponde al axonema central,con sus nueve dobletes y dos singletesacostumbrados, pero en su final quedadesorganizado en un conglomerado demicrotubulos sueltos.
290
21
APARATO
GENITA
LMASCULIN
O

CONSIDERACIONES CL�INICAS
La parotiditis, una virosis sist�emica, produceuna incidencia del 20 al 30% de orquitis aguda(inflamacion de los testıculos) en los hombrespospuberales. Por regla general, laespermatogenia no se ve afectada por estaenfermedad.
El sındrome de Klinefelter deriva de unaalteracion denominada no disyuncion que sucededurante la meiosis, en la que los homologos XX nose separan, lo que genera una persona con ungenoma XXY (un cromosoma X supernumerario).Estos individuos tienen retraso mental, son
est�eriles, altos y delgados, y exhiben unascaracterısticas masculinas atenuadas, entre ellasunos testıculos pequenos.
291
21
APARATO
GENITA
LMASCULIN
O
Figura 21.5 Espermatogenia y un espermatozoide maduro. (Tomado de Kessel RG: Tissue and Organs: A Text Atlas of Scanning Electron
Microscopy. San Francisco, Freeman, 1979.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Ciclo del epitelio seminıfero
Las c�elulas germinativas derivadas de una espermato-gonia p�alida de tipo A semantienenmuy juntas comoun sincitio cuyos miembros se comunicaran entre sı ysincronizaran su desarrollo en las seis etapas de la es-permatogenia (fig. 21.6) al evolucionar hacia losespermatozoides. Cada etapa dura 16 dıas y recibe elnombre de ciclo del epitelio seminıfero. La finaliza-cion de la espermatogenia exige recorrer cuatro ciclos(64 dıas).
CELULAS INTERSTICIALES DE LEYDIG
El tejido conjuntivo laxomuy vascularizado que rodeaa los tubulos seminıferos tambi�en alberga pequenosgrupos de grandes c�elulas endocrinas poli�edricas, lasc�elulas intersticiales de Leydig (fig. 21.7), que ela-boran testosterona, la hormona sexual masculina.Las c�elulas de Leydig son tıpicas c�elulas productoras
de esteroides, que contienen un abundante retıculoendopl�asmico liso, numerosas mitocondrias con cres-tas tubulares, y cristales de Reinke, cuya funcion sedesconoce. Se cree que la testosterona se libera segunse sintetiza; estas c�elulas no muestran vesıculas secre-toras.
CONSIDERACIONES CL�INICAS
Los tratamientos contra el c�ancer conquimioterapia en varones jovenes puedenprovocarles una aspermatogenia por el riesgo deafectar a las espermatogonias que sufrenmitosis ya los espermatocitos que sufren meiosis. Las
c�elulas madre latentes cuya participacion no est�aprevista en la sıntesis de ADN y en el ciclo celular
ser�an capaces de repoblar el epitelio seminıferocuando se interrumpa la quimioterapia
antineopl�asica.
292
21
APARATO
GENITA
LMASCULIN
O

293
21
APARATO
GENITA
LMASCULIN
O
Figura 21.6 Las seis etapas de laespermatogenia en el tubulo seminıfero
humano.(Reproducido a partir de Clermont Y:
The cycle of the seminiferous epithelium in man.Am J Anat 112:35-52, 1963.)
Figura 21.7 Sıntesis de testosterona en las c�elulas
intersticiales de Leydig. AMPc, monofosfato deadenosina cıclico; ATP, trifosfato de adenosina;
CoA, coenzimaA; LH, hormona luteinizante; PPi,
pirofosfato; REL, retıculo endopl�asmico liso.(Tomado de Gartner LP, Hiatt JL: Color Textbook of
Histology, 3rd ed. Philadelphia, Saunders, 2007,
p 500.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

HISTOFISIOLOGIA DE LOS TESTICULOS
Cada testıculo genera alrededor de 100 millones deespermatozoides diarios, que se alimentan y trans-portan hasta las vıas genitales en el medio cargadode fructosa originado por las c�elulas de Sertoli(fig. 21.8). El proceso requiere la accion de la hor-mona luteinizante (LH) y de la hormona foliculo-estimulante (FSH). El mecanismo de control hor-monal sobre la espermatogenia se representa en lafigura 21.9.
. La LH procedente de la adenohipofisis activa lasc�elulas intersticiales de Leydig para formar elandrogeno masculino testosterona. El mecanismode la sıntesis de testosterona y su salida se explicaen las figuras 21.8 y 21.9.
. La FSH, tambi�en derivada de la adenohipofisis,estimula la fabricacion y expulsion de proteınafijadora de androgenos (ABP), que se une a latestosterona y le impide abandonar el tubuloseminıfero. La elevada concentracion detestosterona en la region de la espermatogeniaestimula el fenomeno de la obtencion deespermatozoides.
. Las hormonas testosterona e inhibina, segregadaspor las c�elulas de Sertoli, activan un circuito deretroalimentacion que inhibe la preparacion de LH.La testosterona tambi�en es:
. Necesaria para el correcto funcionamiento de lasvesıculas seminales, la prostata y las gl�andulasbulbouretrales.
. Responsable de las caracterısticas sexualesmasculinas y su aspecto.
CONSIDERACIONES CL�INICAS
El c�ancer testicular es mucho m�as habitual entrelos hombres blancos que en los de origen africano o
asi�atico. Aunque sea una enfermedad rara, es
la forma m�as frecuente de c�ancer en los hombres
de 20 a 34 anos. Los c�anceres testiculares nacen de
las c�elulas germinativas del epitelio seminıfero el
95% de las veces y de las c�elulas intersticiales de
Leydig m�as o menos en el 5% de los casos. No seconoce su causa, pero las siguientes situacionesparecen predisponer a que una persona sufra esteproceso: criptorquidia, sındrome de Klinefelter y
antecedentes familiares de c�ancer testicular. Sussıntomas consisten en un cambio de tamano en untestıculo o en ambos, acompanado de dolor o no;sensacion de peso en el escroto; y una presion o undolor sordo en la parte inferior de la espalda, en elestomago o en la ingle. Su diagnostico se efectua
mediante an�alisis de sangre y pruebasde imagen. Eltratamiento comprende la cirugıa, la quimioterapia y
la radioterapia. El c�ancer de testıculo puede curarsesi se trata pronto.
294
21
APARATO
GENITA
LMASCULIN
O

295
21
APARATO
GENITA
LMASCULIN
O
Figura 21.8 Sıntesis de testosterona en las c�elulas intersticiales de Leydig. AMPc, monofosfato de adenosina cıclico; ATP, trifosfato deadenosina; CoA, coenzima A; LH, hormona luteinizante; PPi, pirofosfato; REL, retıculo endopl�asmico liso. (Tomado de Gartner LP,
Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 500.)
Figura 21.9 Control hormonal de la espermatogenia. LHRH, hormona liberadora de la hormona luteinizante. (Adaptado de Fawcett
DW: Gloom and Fawcett’s A Textbook of Histology, 10th ed. Philadelphia, Saunders, 1975.)�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Vıas genitales
El sistema de las vıas genitales masculinas comienzaen el testıculo, con los conductos intratesticulares, ysigue con los conductos extratesticulares, que acabanen la uretra prost�atica. Los conductos intratesticu-lares son los tubulos rectos y la red testicular (fig.21.10 y tabla 21.1).
. Los tubulos rectos son unos tubulos cortos yrectilıneos que llevan los espermatozoides desde lostubulos seminıferos hasta la red testicular. Su mitadproximal est�a tapizada por las c�elulas de Sertoli, y ladistal lo est�a por un epitelio cubico simple cuyasc�elulas poseen microvellosidades bajas ynormalmente un solo cilio.
. La red testicular ocupa el mediastino testicular yconsiste en un sistema laberıntico de espaciosrevestidos por un epitelio cubico simple. Las c�elulasde este epitelio son semejantes a las que recubren lamitad distal de los tubulos rectos.
Los conductos extratesticulares son el epidıdimo,el conducto deferente y el conducto eyaculador(v. fig. 21.10 y tabla 21.1).
. El epidıdimo est�a compuesto por dos partes, losconductillos eferentes y el conducto del epidıdimo.. Los 10 a 20 conductillos eferentes son unostubulos cortos que transportan losespermatozoides desde la red testicular hasta elconducto del epidıdimo. Estos conductos est�anrevestidos por un epitelio simple cuyas c�elulasadoptan un aspecto festoneado debido a los focosalternos de c�elulas cubicas simples y cilındricassimples, y reabsorben parte del lıquido en el quese encuentran suspendidos los espermatozoides.En la profundidad del epitelio hay unamembranabasal que lo separa del tejido conjuntivo, que a suvez est�a envuelto por una capa delgada demiocitos lisos con una disposicion circular.
. El conducto del epidıdimo (epidıdimo) es untubo que mide unos 4 a 6m de largo y est�atapizado por un epitelio estereociliado (conmicrovellosidades largas e inmoviles)seudoestratificado. Su pared aloja unas capascirculares de musculo liso cuyas contracciones
perist�alticas facilitan la descarga de losespermatozoides en el conducto eferente. Suepitelio est�a constituido por dos tipos celulares,las c�elulas basales regenerativas y las c�elulasprincipales estereociliadas que reabsorbenlıquido desde la luz, y segrega glicerofosfocolina,que vuelve est�eriles a los espermatozoides hastaque tenga lugar la capacitacion en el aparatogenital femenino.
. El conducto deferente tiene una luz pequena y deforma irregular y una pared muscular gruesa;conduce los espermatozoides desde el conducto delepidıdimo hasta el conducto eyaculador. Surevestimiento epitelial es seudoestratificado yparece el del conducto del epidıdimo con c�elulasprincipales m�as bajas. La tunica de musculo lisoposee una capa longitudinal interna, otra circularintermedia y otra longitudinal externa. La porcionterminal del conducto deferente, la ampolla, est�adilatada y recibe el conducto de la vesıcula seminalpara originar el conducto eyaculador.
. El corto conducto eyaculador, tapizado por unepitelio cilındrico simple, carece de c�elulasmusculares en su pared. Perfora el par�enquima de laprostata y da salida al contenido de su luz en elcolıculo seminal de la uretra prost�atica.
CONSIDERACIONES CL�INICAS
M�as de 600.000 estadounidenses eligen lavasectomıa al ano como medio anticonceptivo.Esta breve intervencion quirurgica presenta una
eficacia pr�acticamente del 100% y se pretende
que sea permanente. No resulta un m�etodocomplicado y en general se lleva a cabo en un
consultorio m�edico. La m�as empleada es la
t�ecnica sin bisturı, en la que se punciona elescroto, y se tira de un asa del cordon
esperm�atico, se le corta y se cauteriza, paradevolverlo a continuacion al escroto. El semen serecoge y se examina al microscopio pasadas unas
6 semanas de la cirugıa y tal vez m�as tarde, paragarantizar que no quedan espermatozoides.Pueden sufrirse unas cuantas complicaciones,pero la mayorıa son transitorias.
296
21
APARATO
GENITA
LMASCULIN
O

297
21
APARATO
GENITA
LMASCULIN
O
Tabla 21.1 CARACTERISTICAS HISTOLOGICAS Y FUNCIONES DE LAS VIAS GENITALES MASCULINAS
Conducto Revestimiento epitelial Tejidos de sost�en Funcion
Tubulos rectos C�elulas de Sertoli en su mitad
proximal; epitelio cubico simple
en su mitad distal
Tejido conjuntivo laxo Transporte de los
espermatozoides desde los
tubulos seminıferos hasta
la red testicularRed testicular Epitelio cubico simple Tejido conjuntivo vascular Transporte de los
espermatozoides desde los
tubulos rectos hasta los
conductillos eferentesConductillos
eferentes
Focos de c�elulas cubicas no
ciliadas alternos con c�elulas
cilındricas ciliadas
Tejido conjuntivo laxo fino
rodeado por una capa delgada
de miocitos lisos con unadisposicion circular
Transporte de los
espermatozoides desde la red
testicular hasta el epidıdimo
Epidıdimo Epitelio seudoestratificado
compuesto por c�elulas basales
bajas y c�elulas principales altas(con estereocilios)
Tejido conjuntivo laxo fino
rodeado por una capa de
miocitos lisos con unadisposicion circular
Transporte de los
espermatozoides desde los
conductillos eferentes hastael conducto deferente
Conducto
deferente
Epitelio cilındrico
seudoestratificado
estereociliado
Tejido conjuntivo fibroel�astico
laxo; tunica gruesa de musculo
liso en tres capas; longitudinalinterna y externa, circular
intermedia
Transporte de los
espermatozoides desde la cola
del epidıdimo hasta elconducto eyaculador
Conducto
eyaculador
Epitelio cilındrico simple Tejido conjuntivo subepitelial
plegado, que da a la luz unaspecto irregular; sin musculo
liso
Transporte de los
espermatozoides desde elsemen hasta la uretra prost�atica
en el colıculo seminal
Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p. 502.
Figura 21.10 Testıculo y epidıdimo. Los lobulillos y su contenido no est�an dibujados a escala. (Tomado de Gartner LP, Hiatt JL: Color
Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 490.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Gl�andulas genitales accesorias
Las gl�andulas genitales accesorias masculinas son elpar de vesıculas seminales y de gl�andulas bulboure-trales m�as la prostata.
. Las vesıculas seminales derecha e izquierda, dosconductos largos y enrollados situados detr�as de lavejiga, unen sus respectivos conductos excretorescon el deferente para generar el conductoeyaculador. Su epitelio cilındricoseudoestratificado est�a compuesto de c�elulasbasales regenerativas y c�elulas cilındricas,cuyo peso depende de la concentracion local detestosterona. Cada c�elula cilındrica presenta unasmicrovellosidades cortas y un unico flagelo. Eltejido conjuntivo fibroel�astico que subyace alepitelio est�a envuelto por una capa circular internay otra longitudinal externa de la tunica muscularlisa. Estas gl�andulas fabrican un lıquido amarillocargado de fructosa que tambi�en contieneamino�acidos, proteınas, citrato y prostaglandinas.La secrecion de las vesıculas seminales representael 70% del volumen del semen y aporta nutrientesa los espermatozoides.
. Las dos pequenas gl�andulas bulbouretrales(gl�andulas de Cowper), revestidas por epitelioscubicos simples o cilındricos simples, se hallancerca de la uretra membranosa y vuelcan susecrecion resbaladiza y viscosa con abundantegalactosa y �acido si�alico hacia su luz, lo que lalubrica. La c�apsula de tejido conjuntivofibroel�astico de la gl�andula lleva fibras musculareslisas y estriadas.
. La prostata (fig. 21.11), gl�andula impar,normalmente del tamano de una castana, rodeapor completo a los conductos eyaculadores y a laporcion prost�atica de la uretra. Su c�apsula
fibroel�astica, intercalada con los miocitos lisos,invade el par�enquima de la gl�andula para formar elestroma, que tambi�en est�a dotado demiocitos lisos.Este par�enquima est�a constituido por 50 gl�andulastubuloalveolares compuestas, organizadas en trescapas conc�entricas:. La m�as interna o mucosa (justo alrededorde la uretra), con las gl�andulas m�as cortas
. Las gl�andulas principales, que son las m�asexternas y ocupan la mayor parte de la prostata
. Las gl�andulas submucosas, cuyo tamano ylocalizacion son intermedios, y quedan en laregion entre las gl�andulas mucosas y lasprincipales.
El par�enquima de las tres gl�andulas consta de unepitelio simple a seudoestratificado cuyas c�elulasest�an perfectamente dotadas de retıculo endopl�as-mico rugoso, aparato de Golgi, lisosomas y vesıculassecretoras. Estas c�elulas elaboran una secrecion acuosacuya liberacion es promovida por la dihidrotestoste-rona, y que est�a integrada por fosfatasa �acida, citrato,lıpidos, enzimas proteolıticos y fibrinolisina. A me-nudo, sobre todo en los ancianos, su luz contieneglucoproteınas calcificadas, concreciones prost�aticas(cuerpos amil�aceos).
La emision de las secreciones procedentes de lasgl�andulas genitales accesorias tiene lugar tras la erec-cion. Las gl�andulas bulbouretrales son las primerasen soltar su lubricante resbaladizo poco despu�es dela ereccion, mientras que la salida de los espermato-zoides desde la ampolla del conducto deferente y delas secreciones de las vesıculas seminales y la pros-tata sucede directamente antes de la eyaculacion.El semen eyaculado supone un volumen aproxi-mado de 3ml y de 200 a 300 millones de esperma-tozoides suspendidos en las secreciones de las gl�an-dulas accesorias.
298
21
APARATO
GENITA
LMASCULIN
O

299
21
APARATO
GENITA
LMASCULIN
O
Figura 21.11 Prostata humana. (Tomado deGartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 505.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Pene
El pene cumple una funcion doble: la expulsion de laorina hacia el exterior y el transporte del semen haciael aparato genital femenino. Este es el organo de lacopula y est�a compuesto de tres masas de cuerposer�ectiles (fig. 21.12):
. Dos cuerpos cavernosos de ubicacion dorsal
. Un unico cuerpo esponjoso de la uretra de situacionventral, cuya terminacion distal es la cabeza del pene,llamada glande del pene, que ofrece una hendiduravertical, el orificio externo de la uretra.
Los tres cuerpos er�ectiles poseen su propia tunicaalbugınea, una c�apsula de tejido conjuntivo fibroso, ylas tres estructuras est�an recubiertas por una vainatubular de piel fina que se extiende sobre el glandedel pene como una funda retr�actil suelta, el prepucio.Los tejidos er�ectiles del pene est�an constituidos porunos espacios vasculares laberınticos e irregularesrevestidos de endotelio, que se encuentran delimita-dos por trab�eculas de tejido conjuntivo dotadas demiocitos lisos. Los dos cuerpos cavernosos:
. Presentan espacios vasculares de dimensionesvariables, que son m�as pequenos en la periferia ym�as grandes cerca del centro.
. Tienen menos fibras el�asticas y m�as miocitos lisosen sus trab�eculas.
. Reciben su riego sanguıneo de la arteria profunda yla arteria dorsal del pene, que perforan las trab�eculasy forman lechos capilares y arterias helicoidales. Lasarterias helicoidales desempenan una misionprimordial en la ereccion del pene.
El cuerpo esponjoso de la uretra:
. Posee espacios vasculares de tamano comparable enzonas centrales y perif�ericas
. Tiene menos miocitos lisos y m�as fibras el�asticasque los cuerpos cavernosos
. Est�a rodeado en su extremo proximal por el potentemusculo bulboesponjoso (musculo estriado).
El drenaje venoso de los tejidos er�ectiles correspon-dientes a los tres cuerpos er�ectiles y al glande del penesigue tres conjuntos de venas que son afluentes de lavena dorsal profunda.
300
21
APARATO
GENITA
LMASCULIN
O

CONSIDERACIONES CL�INICAS
Un unico eyaculado normal contiene alrededor de70 a 100millones de espermatozoides pormililitro.Un hombre con un numero inferior a 20millones de
espermatozoides/ml se considera est�eril.
301
21
APARATO
GENITA
LMASCULIN
O
Figura 21.12 Pene en un corte transversal. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders,
2007, p 507.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

MECANISMOS DE LA ERECCION, EYACULACION
Y DETUMESCENCIA
La circulacion sanguınea por el pene fl�acido se redirigehacia anastomosis arteriovenosas entre el riego arte-rial y el drenaje venoso, lo que obstaculiza el paso desangre hacia los espacios vasculares de los cuerposer�ectiles (fig. 21.13). Cuando el flujo sanguıneo est�amodificado y en vez de entrar en las anastomosisarteriovenosas va hacia los espacios vasculares de lostejidos er�ectiles, el pene se pone en ereccion y latunica albugınea de los cuerpos er�ectiles queda esti-rada. La ereccion se alcanza mediante:
. Estimulacion sexual, de tipo t�actil, visual, olfatoriao cognitiva, lo que implica al sistema nerviosoparasimp�atico
. La provocacion de la liberacion de oxido nıtricodesde el endotelio de las arterias profunda y dorsaldel pene
. La relajacion de los musculos lisos que constituyenla tunica media de estos vasos, lo que aumenta lallegada de sangre hacia ellos.
Simult�aneamente, las anastomosis arterioveno-sas se contraen y la sangre penetra en las arteriashelicoidales, que la envıan hacia los tejidos er�ectiles.Estos ultimos se ponen turgentes, comprimen a lasvenas e impiden la salida de la sangre, con lo que laereccion se mantiene.
Si el glande del pene sigue estimulado, las gl�andu-las bulbouretrales descargan su secrecion resbaladiza,los productos sintetizados por las vesıculas seminales,junto a los espermatozoides que hay en la ampolla delconducto deferente, hacia los conductos eyacula-dores. La prostata emite su secrecion hacia la uretraprost�atica, y el semen se eyacula, proceso que est�asometido al control del sistema nervioso simp�atico,de la siguiente manera:
. Los musculos lisos de las gl�andulas accesorias y lasvıas genitales se contraen y conducen el semen haciala uretra.
. El musculo del esfınter de la vejiga urinaria secontrae para evitar la filtracion de orina.
. El musculo bulboesponjoso se contrae de manerarıtmica, y expulsa el semen de la uretra.
La detumescencia sucede tras la eyaculacion por-que:
. Los nervios parasimp�aticos dejan de ocasionar laliberacion de oxido nıtrico en las arterias profunday dorsal del pene.
. El descenso de la circulacion sanguınea en estosvasos permite la apertura de las anastomosisarteriovenosas.
. La sangre evacuada desde los espacios vasculares delos tejidos er�ectiles hace que el pene regrese a suestado fl�acido.
302
21
APARATO
GENITA
LMASCULIN
O

CONSIDERACIONES CL�INICAS
La disfuncion er�ectil tiene una participacion en elsistema nervioso, con las alteraciones de las vıas
correspondientes a la corteza cerebral–hipot�alamo,
m�edula espinal, sistema nervioso autonomo– y losproblemas derivados de las enfermedadesvasculares. Otras causas son el ictus, los
traumatismos craneales, las lesiones de la m�edulaespinal y los trastornos de ansiedad. Numerosos
procesos sist�emicos, como la enfermedad deParkinson, la diabetes y la esclerosis multiple,
tambi�en pueden dar lugar a una disfuncion er�ectil.
El sildenafilo se ideo en un principio como un
f�armaco para tratar la insuficiencia cardıaca. Sinembargo, los pacientes que antes eran impotentesy recibieron este producto senalaron que estabanexperimentando erecciones. A partir de estasobservaciones y los posteriores estudios clınicos, el
sildenafilo se convirtio en un f�armaco de eleccionpara el tratamiento de la impotencia. Han aparecido
numerosas sustancias semejantes que tambi�en soncapaces de restablecer la capacidad de lograr unaereccion en pacientes que eran impotentes.
303
21
APARATO
GENITA
LMASCULIN
O
Figura 21.13 Circulacion de la sangre por el pene fl�acido y en ereccion. La anastomosis arteriovenosa (flechas) del pene fl�acido es ancha,
y desvıa la sangre hacia el drenaje venoso. En el pene en ereccion, las anastomosis arteriovenosas est�an contraıdas y aumenta el paso
de sangre hacia los espacios vasculares del tejido er�ectil, lo que pone turgente el pene con la sangre. (Adaptado deConti G: [The erectionof the human penis and its morphologico-vascular basis.] Acta Anat (Basel) 5:217, 1952.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

22 ORGANOSDE LOS SENTIDOS
Las terminaciones sensitivas son receptores especiali-zados en las terminales de las dendritas que detectanestımulos transmitidos hacia el sis-tema nervioso central para su proce-samiento. Este capıtulo estudia estosreceptores especializados que soncomponentes de las vıas aferentessom�aticas general o especial y vis-ceral. Segun el estımulo recibido seidentifican tres clases diferentes dereceptores:
. Los exterorreceptores est�ansituados sobre la superficie delcuerpo y reciben estımulos como latemperatura, la presion, el tacto y eldolor (aferente som�atico general);la luz que permite la vision y lasondas sonoras que permiten la audicion (aferentesom�atico especial); y el gusto y el olfato (aferentevisceral especial; descritos en los capıtulos 16 y 15).
. Los propiorreceptores est�an distribuidos por lostendones, en las c�apsulas articulares y en los husosmusculares del musculo estriado, y recibenestımulos acerca de la actitud alerta de la posicioncorporal en el espacio.
. Los interorreceptores est�an presentes en losorganos del cuerpo y transmiten informacion sobreellos, formando parte de la modalidad aferentevisceral general.
Receptores perif�ericos especializados
Las especializaciones dendrıticas ubicadas en los ten-dones, la piel, los musculos, las fascias y las c�apsulasarticulares reaccionan frente a ciertos estımulos espe-cıficos y se clasifican como mecanorreceptores, ter-morreceptores y nocirreceptores; sin embargo,cuando un estımulo concreto alcanza una intensidadespecıfica, puede activar cualquier receptor (fig. 22.1).
Los mecanorreceptores se deforman por un estı-mulo o por el tejido que los rodea y responden alestiramiento, las vibraciones, el tacto y la presion.Pueden estar encapsulados o no. Hay dos tipos demecanorreceptores no encapsulados:
. Las terminaciones nerviosas peritriquiales (v. fig.22.1D) no tienen ni vaina mielınica, ni c�elulas de
Schwann, y penetran en la epidermis de la cara y lacornea en el ojo, lo que aporta una gran sensibilidad
al tacto y a la presion a dichasregiones. Otras terminacionesnerviosas peritriquiales est�anvinculadas a los folıculos pilosos yfuncionan ante losmovimientos delpelo. Algunos de los estımulos seinterpretan como un cosquilleo,dolor o hasta temperatura alta.. Los discos deMerkel (v. fig. 22.1A)son mecanorreceptores estudiadosen el capıtulo 14.
Los mecanorreceptores encapsu-lados (v. fig. 22.1B, C, E, F, G y H)est�an integrados por fibras nerviosasen el interior de una c�apsula de teji-do conjuntivo.
. Los corpusculos de Meissner, abundantes en lascrestas epid�ermicas de la yema de los dedos, losp�arpados, los labios, la lengua, los pezones y lapiel de los pies y el antebrazo, est�an especializadosen la discriminacion t�actil. Tres o cuatroterminales nerviosas m�as sus c�elulas de Schwannest�an encapsuladas por elementos de tejidoconjuntivo.
. Los corpusculos de Pacini est�an compuestos por ununico axon amielınico rodeado por un complejo devainas de tejido conjuntivo con capas conc�entricasde c�elulas aplanadas. Estos corpusculos, situados enla dermis, la hipodermis, el mesenterio y elmesocolon, reaccionan frente a la presion, el tactoy la vibracion.
. Las terminaciones de Ruffini son terminacionesnerviosas muy ramificadas rodeadas por unas pocascapas de fibroblastos modificados, localizadas en ladermis cut�anea, los lechos ungueales, losligamentos periodontales y las c�apsulas articulares.Estas terminaciones perciben el estiramiento y lapresion.
. Los bulbos terminales de Krause, cuya funcion nose conoce, son terminaciones encapsuladas esf�ericasen los tejidos conjuntivos subepiteliales de lascavidades oral y nasal, el peritoneo, la dermispapilar, las articulaciones, la conjuntiva y lasregiones genitales.
� 2011 Elsevier Espana, S.L. Reservados todos los derechos
TERMINOS CLAVE
. Receptores perif�ericosespecializados
. Ojos
. Retina
. Bastones y conos
. Oıdos
. Laberintos oseoy membranoso
. Organo de Corti
. Funcion vestibular
304

305
22
ORGANOSDELOSSEN
TIDOS
Figura 22.1 Diversos mecanorreceptores. A. Disco de Merkel. B. Corpusculo de Meissner. C. Corpusculo de Pacini. D. Terminaciones
nerviosas peritriquiales (desnudas). E.Corpusculo de Ruffini. F.Bulbo terminal de Krause.G.Husomuscular.H. Organo tendinoso de
Golgi. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 512.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

. Los husos musculares y los organos tendinososde Golgi son mecanorreceptores encapsuladosespecializados en la propiocepcion. Los husosmusculares detectan cambios en la longitudmuscular y la velocidad que siguen,mientras que losorganos tendinosos de Golgi comprueban latension y su ritmo de aplicacion a la articulacion.
No se han identificado termorreceptores, pero sesupone que las terminaciones nerviosas desnudas dela epidermis responden al frıo y al calor. Los nocirre-ceptores son terminaciones nerviosas desnudas muyramificadas en la epidermis que captan el dolor. Fun-cionan en alguna de las tres maneras siguientes;actuan frente a:
. Una agresion o un dano mec�anico
. Un grado extremo de calor o de frıo
. Compuestos quımicos como bradicinina,serotonina e histamina.
Ojo
Los ojos son los organos fotosensibles del cuerpo yest�an alojados en las orbitas oseas del cr�aneo. Elglobo ocular (bulbo del ojo) (fig. 22.2) y susestructuras asociadas sirven para recibir los rayosluminosos a trav�es de la cornea y otros medios derefraccion y enfocarlos sobre su pared posterior,donde est�a situada la retina con sus bastones yconos fotosensibles. Al estimularse con la luz, setransmite una senal hacia el cerebro para su proce-samiento en una compleja imagen visual que per-cibe el individuo.
El ojo se forma a partir de tres orıgenes. La retina yel nervio optico son excrecencias del prosenc�efalo y selas puede observar a las 4 semanas de desarrollo. Elcristalino y parte de las estructuras auxiliares existentesen la porcion anterior del ojo proceden del ectodermosuperficial de la cabeza. Las estructuras vinculadas conel globo ocular y sus tunicas (cubiertas) surgen a partirde los tejidos mesenquimatosos adyacentes. Estas trescapas son la tunica fibrosa externa, la tunica vascularintermedia y la tunica nerviosa interna.
Los componentes de la tunica fibrosa son la escle-rotica opaca, la esclerotica blanca y la cornea trans-parente.
. La esclerotica, el blanco opaco del globo ocular,est�a constituida por fibras de col�ageno de tipo Ientremezcladas con fibras el�asticas, lo que crea unapotente capa fibrosa que resiste la presion aplicadasobre ella por los humores vıtreo y acuoso. En sussuperficies superior, inferior, medial y lateral recibelas inserciones de los musculos extrınsecos del ojo.La cara profunda de la esclerotica manifiesta la
presencia de melanocitos, y su territorio posteriorest�a perforado por el nervio optico.
. La porcion transparente anterior del globo, lacornea, sobresale hacia delante, es avascular y tieneuna inervacion abundante a cargo de fibrasnerviosas sensitivas. La cornea est�a integrada porcinco capas:. El epitelio corneal, un epitelio pavimentosoestratificado no queratinizado, es lacontinuacion de la conjuntiva. Sus capassuperficiales presentan zonas de oclusion,mientras que las c�elulas de la capa m�asprofunda se interdigitan y est�an unidas entre sımediante desmosomas. Las fibras del doloratraviesan la cara basal del epitelio corneal y searborizan cerca de su superficie. Las c�elulasepiteliales de la periferia de la cornea poseenactividad mitotica, y las reci�en formadas tardanuna semana en descamarse. El agua y los ionesdel estroma subyacente penetran en la cornea yllegan al saco conjuntival.
. Lamembrana de Bowman, una capa fibrosa, est�acompuesta por col�ageno de tipo I que separa elepitelio del estroma subyacente.
. El estroma, tambi�en transparente, es la capa m�asgruesa de la cornea. Contiene de 200 a250 laminillas con haces de col�ageno de tipo Isegun una disposicion regular, donde las fibras decol�ageno en cada una son paralelas entre sı perono con respecto a las de la laminilla m�assuperficial o m�as profunda. Estas fibras decol�ageno, junto a las fibras el�asticas y losfibroblastos asociados, se encuentran sumergidosen una sustancia fundamental cargada de sulfatode condroitina y sulfato de queratano. La mallatrabecular del espacio revestido de endotelio,llamado limbo, est�a situada en el lımite entre laesclerotica y la cornea. Estos espacios se drenanpor el canal de Schlemm, el conducto que vacıa elhumor acuoso desde la c�amara anterior del ojo enun plexo venoso.
. Lamembrana deDescemet, unamembrana basalgruesa y muy desarrollada que divide el estromadel endotelio corneal, gana espesor y se vuelvem�as fibrosa con la edad.
. El endotelio corneal, un epitelio pavimentososimple que reviste la cara profunda de la cornea,elabora la membrana de Descemet. Asimismo,este endotelio lleva a cabo el transporte activo deiones de sodio (a los que siguen de manera pasivalos iones de cloruro y el agua) desde el estromahacia la c�amara anterior, lo que deriva en ladeshidratacion del estroma. Esta situacionmantiene la transparencia caracterıstica delestroma.
306
22
ORGANOSDELOSSEN
TIDOS

CONSIDERACIONES CL�INICAS
El glaucoma, la principal causa de ceguera a nivelmundial, deriva de una presion intraocularprolongada secundaria a la obstruccion de la
salida del humor acuoso desde la c�amara anteriordel ojo. Como la produccion de humor acuoso esconstante, con el tiempo, el bloqueo de su
evacuacion desde la c�amara anterior aumenta lapresion por todo el ojo, afectando enprimer lugar a
la retina, lo que provoca una p�erdida de la vision
perif�erica, que a la larga genera una grave lesionen el nervio optico y, si no se cuida, ceguera.
307
22
ORGANOSDELOSSEN
TIDOS
Figura 22.2 Anatomıa del ojo (globo). (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders,
2007, p 515.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

TUNICA VASCULAR (UVEA)
Los integrantes de la tunica vascular son la coroides,el cuerpo ciliar y el iris (fig. 22.3).
. La coroides, la capa pigmentada y muyvascularizada de la pared posterior del globo ocular,est�a sujeta de manera laxa a la tunica fibrosa. Secompone de tejido conjuntivo laxo que albergaabundantes fibroblastos, multiples vasossanguıneos y numerosos melanocitos que leotorgan su tıpico color negro. La region interna de lacoroides, la l�amina coroidocapilar, tiene unacantidad especial de capilares y alimenta a la retina,de la que se encuentra separada por la membranade Bruch, cuyo nucleo de fibras el�asticas est�arecubierto de fibras de col�ageno por ambos lados.
. El cuerpo ciliar, localizado a la altura delcristalino, es una continuacion cuneiforme de lacoroides que rodea la pared interna del ojo. Laprolongacion m�as anterior del cuerpo ciliar lindacon la esclerotica en el limbo, mientras que la m�asposterior lo hace con el cuerpo vıtreo. La porcioncentral (media) sobresale hacia el cristalino, ydesde ella surgen unos relieves digitiformes, losprocesos ciliares. La superficie interna del cuerpociliar y de los procesos ciliares est�a revestida por laporcion ciliar de la retina (una capa de la retinainsensible a la luz), constituida por dos estratos: elque da a la luz del globo, que carece de pigmento,y el interno pigmentado con melanina. Las fibraszonulares, formadas por fibrilina, irradian desdelos procesos ciliares de la porcion anterior delcuerpo ciliar para insertarse en la c�apsula delcristalino y crear el ligamento suspensoriodel cristalino. La capa interna sin pigmentos de laporcion ciliar transporta un filtrado plasm�atico, elhumor acuoso, que aporta oxıgeno y nutrientes alcristalino y a la cornea, en la c�amara posterior delojo. A continuacion, el humor acuoso sale a trav�esde la abertura pupilar hacia la c�amara anteriordel ojo y en ultima instancia abandona el ojo paraentrar en el canal de Schlemm a nivel del limbocon el fin de ser desaguado hacia el sistemavenoso.. Elmusculo ciliar (tres fascıculos demusculo liso)est�a situado en el cuerpo ciliar. Debido a suposicion, las contracciones de uno de losfascıculos contribuyen a abrir el canal deSchlemm. Al contraerse los dos fascıculosrestantes, liberan la tension de los ligamentossuspensorios del cristalino, haciendo queaumente la convexidad y el grosor del cristalino,lo que le permite enfocar los elementos que est�encerca, proceso denominado acomodacion.La relajacion de los musculos ciliares tensa el
cristalino, lo que se traduce en su aplanamiento;en estas condiciones, el enfoque m�as agudo caesobre los objetos alejados.
. La porcion coroidea de la tunica vascular continuahacia delante con el iris, que queda entre lasc�amaras anterior y posterior del ojo y cubre todo elcristalino excepto a nivel de la pupila. Su superficieanterior posee dos anillos: la zona pupilar y la zonaciliar. La cara anterior es irregular con surcos en lospuntos de retraccion. Su cara posterior es lisa y seencuentra revestida por el mismo epitelio bilaminarque tapiza al cuerpo ciliar. La superficie posteriorque mira hacia el cristalino est�a muy pigmentada,por lo que no deja pasar la luz m�as que por lapupila. El iris tiene dos musculos intrınsecos:. Elmusculo dilatador de la pupila, que nace en elborde del iris e irradia hacia la pupila, y est�ainervado por el sistema nervioso simp�atico. Sucontraccion dilata la pupila cuando el grado deiluminacion es bajo.
. El musculo esfınter de la pupila, que forma unanillo conc�entrico alrededor de la pupila y est�ainervado por el sistema nervioso parasimp�atico atrav�es del nervio oculomotor (NC III). Suscontracciones contraen la pupila si la luz es fuerte.
El color del iris depende de la cantidad de melano-citos que haya en su epitelio. Los ojos oscuros sedeben a que su numero es abundante, mientras quecuando hay pocos los ojos son de color azul claro.
CRISTALINO
El cristalino, un disco biconvexo flexible y transpa-rente, est�a integrado por varias capas de c�elulas apla-nadas m�as sus productos de secrecion (v. fig. 22.3).Esta estructura tiene tres partes:
. La c�apsula del cristalino representa una l�aminabasal transparente que contiene col�ageno de tipo IVy glucoproteınas. La c�apsula envuelve a todo elcristalino y es m�as gruesa por su parte anterior.
. El epitelio subcapsular se halla en la inmediataprofundidad de la c�apsula y solo en sus porcionesanterior y lateral. Est�a compuesto de una unica capade c�elulas cubicas conectadas por uniones gap.Los v�ertices de las c�elulas van dirigidos hacia lasfibras del cristalino y se interdigitan con ellas.
. Las fibras del cristalino, unas 200 c�elulas alargadaso m�as, se originan en el epitelio subcapsular yadquieren una gran especializacion al perder susnucleos y org�anulos y convertirse en c�elulashexagonales largas (de 7 a 10mm), proceso quecontinua toda la vida. Estas c�elulas se llenan de unasproteınas del cristalino llamadas cristalinas, queaumentan su ındice de refraccion.
308
22
ORGANOSDELOSSEN
TIDOS

CONSIDERACIONES CL�INICAS
La presbicia es un proceso relacionado con la edaden el que el ojo manifiesta una disminucionprogresiva de la capacidad para enfocar los objetosproximos. Su causa exacta se desconoce; sinembargo, existen pruebas de que el cristalino pierdeelasticidad con los anos y esto puede combinarse aun descenso en la fuerza de contraccion de losmusculos ciliares. Aunque no tiene remedio, lamayorıa de las personas consiguen adaptarsemediante la prescripcion de unas gafas que realicenla acomodacion de la vision cercana.
La catarata es una opacidad que surge en elcristalino del ojo, de diverso grado, desde ligerahasta completa, y que obstruye el paso de la luz.
Lo normal es que evolucione con lentitud hastaprovocar una disminucion de la vision y laposibilidad de acabar en una ceguera si se deja sintratar. El trastorno suele afectar a ambos ojos,pero casi siempre a uno antes que al otro. Lascataratas se forman a partir de distintas causas,como la exposicion prolongada a la luz ultravioletao a las radiaciones, una edad avanzada y comoefecto secundario de enfermedades del tipo de la
diabetes y la hipertension. Tambi�en puedenproducirse a partir de una lesion ocular o untraumatismo fısico. Las cataratas no responden alos medicamentos, pero el cristalino danadopuede retirarse y sustituirse por una lentecorrectora artificial.
309
22
ORGANOSDELOSSEN
TIDOS
Figura 22.3 Anatomıa del ojo (globo). (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007,
p 515.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

CUERPO VITREO
La estructura semigelatinosa y transparente que llenala concavidad posterior tras el cristalino recibe elnombre de cuerpo vıtreo. Est�a constituido en un99% por agua que lleva una pequena cantidadde electrolitos, unas cuantas fibras de col�ageno y�acido hialuronico. El cuerpo vıtreo se adhiere a laretina sobre todo en la ora serrata (el borde anteriorde la retina fotosensible). Unas c�elulas pequenas,denominadas hialocitos, que se piensa que sinteti-zan col�ageno y �acido hialuronico, est�an localizadasen la periferia del cuerpo vıtreo. Un pequeno con-ducto, el conducto hialoideo, situado en la lıneamedia del cuerpo vıtreo, se extiende desde la caraposterior del cristalino hasta el disco optico; albergala arteria hialoidea en el feto, pero en el adulto seencuentra ocupado por lıquido.
RETINA (TUNICA NERVIOSA)
La tunicam�as interna del ojo, la retina (fig. 22.4), es laporcion nerviosa que contiene los bastones y losconos, las c�elulas fotorreceptoras especializadas. Laretina procede del tejido nervioso de la vesıcula opticaque se origina en el dienc�efalo cerebral. M�as adelanteen el curso del desarrollo, la vesıcula optica se hundepara formar la cupula optica. Esta copa consta de doscapas y est�a conectada al cerebro por el tallo optico.
La externa se convierte en la capa pigmentaria de laretina, la interna se diferencia en la capa neuronal dela retina (retina propiamente dicha) y el tallo opticose transforma en el nervio optico (NC II). La capapigmentada de la retina cubre la superficie interna delglobo, contando el cuerpo ciliar y la pared posteriordel iris; sin embargo, la retina propiamente dichaacaba en la ora serrata.
. El disco optico de la pared posterior representa elpunto de salida del nervio optico, y como carece debastones y de conos, se le considera lamancha ciegade la retina.
. A unos 2,5mmde lamancha ciega en sentido lateralhay una zona pigmentada de amarillo llamadam�acula lutea, que posee una depresion en su centrodenominada fovea central, donde no existen m�asque conos.. Los conos est�an tan apretados en la fovea que lasdem�as capas de la retina quedan amontonadas aun lado. La agudeza visual es mayor en la foveacentral.
. Segun crece la distancia desde la fovea, cada vezhay menos conos, mientras que los bastonespredominan.
La region de la retina que actua en la fotorrecepcionest�a integrada por 10 capas que miran hacia la super-ficie interna de la coroides.
310
22
ORGANOSDELOSSEN
TIDOS

CONSIDERACIONES CL�INICAS
Las moscas volantes que surgen en la vision,sobre todo en las personas mayores, yaparentemente se trasladan en realidad son lassombras de pequenos fragmentos de partıculasen el cuerpo vıtreo lanzados sobre la retina.Cuando el ojo se mueve de un lado a otro o de
arriba abajo, estas moscas tambi�en cambian deposicion en el interior del cuerpo vıtreo, haciendoque lasmoscas se desplacen y parezca que floten.
Las moscas flotantes est�an relacionadas con la
retinopatıa diab�etica, los desgarros de retina, el
desprendimiento de retina y la miopıa. Se danm�asamenudo en aquellos casos en que ha habido unalesion ocular o una operacion de cataratas. Lamayor parte de las moscas volantes bajan detamano e intensidad con el tiempo debido a quepueden disolverse. A la larga el cerebro las ignora,y el paciente deja de sentirlas.
CONSIDERACIONES CL�INICAS
El desprendimiento de retina puede deberse aun golpe subito en el ojo, como el impacto de unapelota de tenis, o a las sacudidas tras una caıda
sobre la cabeza. M�as habitual es el desecamientodel cuerpo vıtreo y su traccion sobre la retina, loque la provoca un desgarro al tirar de la capapigmentada. El cuerpo vıtreo puede rezumar
lıquido detr�as de la retina y desprenderla m�as. Laspersonas con un desprendimiento de retina hande acudir a un oftalmologo de inmediato porque sudiagnostico precoz y su reparacion brindan losmejores resultados para la vision. Cualquierretraso deja que el desprendimiento se extiendahasta abarcar toda la retina. Si no se trata, laceguera llega a ser completa en el ojo afectado.
Los m�etodos actuales para abordar undesprendimiento de retina consisten en la cirugıa
por l�aser y la crioterapia.
311
22
ORGANOSDELOSSEN
TIDOS
Figura 22.4 Capas celulares de la retina. El espacio obser-
vado entre la capa pigmentada y el resto de la retina cons-tituye un artefacto de obtencion y no existe en el adulto
excepto durante un desprendimiento de retina. (Tomado
de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.Philadelphia, Saunders, 2007, p 520.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Capas de la retina
Las 10 capas de la retina, desde el epitelio pigmen-tario m�as interno hasta la membrana limitanteinterna m�as externa, guardan una distribucion muyprecisa (fig. 22.5).
. El epitelio pigmentario est�a compuesto por c�elulascubicas o cilındricas, derivadas de la capa externa dela cupula optica, y se encuentra unido a lamembrana de Bruch. Los nucleos de las c�elulaspigmentarias ocupan una posicion basal; en lospuntos de invaginacion en la membrana de Bruch,las mitocondrias son numerosas, lo que indica untransporte activo. Las microvellosidades que saltandesde la superficie libre de estas c�elulas seinterdigitan con la punta de los bastones y losconos. La cara apical de estas c�elulas est�a llena degr�anulos de melanina, lo que favorece una mayoragudeza visual. Asimismo, el citoplasma apicalposee cuerpos residuales que contienen el extremofagocitado perdido por los bastones. El epiteliopigmentado sirve para:. Evitar los reflejos de luz al absorberla tras haberactivado los bastones y los conos.
. Fagocitar la punta agotada de los bastones y losconos.
. Esterificar la vitamina A.. En la capa de bastones y conos de la retina haydos tipos distintos de c�elulas fotorreceptoras(fig. 22.6; v. fig. 22.5). El numero de bastonesasciende a unos 100 a 120 millones, y el de conos aunos 6 millones. Las porciones apicales de estasc�elulas polarizadas y muy especializadas, llamadassegmentos externos, se interdigitan con lasregiones apicales de las c�elulas de la capapigmentada. Las caras basales de los bastones y losconos establecen sinapsis con las c�elulas de la capabipolar de la retina. Los bastones est�anespecializados en percibir objetos con poca luz,mientras que los conos lo est�an en percibirlos aplena luz y en diferenciar los colores.
. La fotosensibilidad de los bastones (v. fig. 22.6) estan aguda que son capaces de generar una senal apartir de un solo foton de luz, pero no lo hacencon una luz fuerte, ni son capaces de detectar elcolor. Estos bastones son c�elulas alargadas (50 mmde largo�3 mm de di�ametro) que se situanalineadas paralelas entre sı y perpendiculares a laretina. Se dividen en un segmento externo, otrointerno, una region nuclear y una region sin�aptica.El extremo sensible a la luz con forma de baston(segmento externo) est�a integrado por 600 a1.000 discos membranosos planos apilados, quecada uno representa una invaginacion de lamembrana plasm�atica (despegada de la superficie
celular). Las membranas llevan un pigmentosensible a la luz denominado rodopsina (purpuravisual). La velocidad de respuesta a la luz es m�aslenta en los bastones que en los conos, pero soncapaces de ir agregando su recepcion a unconjunto. Los discos emigran poco a poco hacia elextremo apical del segmento externo y se pierdenpara ser fagocitados por las c�elulas epitelialespigmentadas. El segmento interno est�a separadodel externo por un estrechamiento, el pedıculo deconexion. Un cilio modificado nace del cuerpobasal situado en la porcion apical del segmentointerno y atraviesa el pedıculo de conexiondirigido hacia el segmento externo. Lasmitocondrias que aportan energıa para elprocesamiento visual est�an apinadas en torno allımite entre los segmentos interno y externo. Lasproteınas elaboradas en el segmento internoemigran para integrarse en los discos del segmentoexterno. En la fotorrecepcion sucede lo siguiente:. La rodopsina (opsina ligada a cis retinal) absorbela luz en el baston.
. La absorcion de la luz convierte el retinal entodo-trans retinal, que se disocia de la opsina.
. Su descoloracion produce opsina activada, quefacilita la union del trifosfato de guanosina a lasubunidad a de la transducina, una proteına Gtrim�erica que cataliza la degradacion del3’,5’-monofosfato de guanosina cıclico (GMPc).
. El descenso de la concentracion de GMPc en elcitosol da lugar al cierre de los canales de Na+
en la membrana plasm�atica del baston.. La hiperpolarizacion del baston conduce a unainhibicion de la salida de neurotransmisores en lasinapsis con las c�elulas bipolares.
. En la proxima fase de oscuridad, se regenera lacantidad deGMPc, vuelven a abrirse los canales deNa+ y se reanuda el paso de este ion.
. El todo-trans retinal sobrante se difunde ytransporta al epitelio pigmentario de la retina atrav�es de las proteınas fijadoras de retinal. Estamol�ecula se recicla a su forma 11-cis retinal.
. Por ultimo, el cis retinal vuelve al baston y se unede nuevo a la opsina, originando rodopsina.
Los canales de Na+ de la membrana plasm�atica semantienen abiertos cuando los bastones no est�an acti-vados por la luz. Durante la fase de oscuridad, losiones de sodio se bombean desde el segmento internohacia el externo, lo que desencadena la liberacion de lasustancia neurotransmisora a las sinapsis con lasc�elulas bipolares. La senal no surge m�as que con lahiperpolarizacion ocasionada por la luz, y se transmitea trav�es de las capas celulares hasta las c�elulas gang-lionares, donde dispara un potencial de accion a lolargo de los axones en su trayecto hacia el enc�efalo.
312
22
ORGANOSDELOSSEN
TIDOS

313
22
ORGANOSDELOSSEN
TIDOS
Figura 22.5 Capas celulares de la retina. El espacio obser-
vado entre la capa pigmentada y el resto de la retina cons-tituye un artefacto de obtencion y no existe en el adulto
excepto durante un desprendimiento de retina. (Tomado
de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed.Philadelphia, Saunders, 2007, p 520.)
Figura 22.6 Estructura de un baston y un cono. C, pedıculo de conexion;
CB, cuerpo basal; Ce, centrıolo; M, mitocondrias; RN, region nuclear;RS, region sin�aptica; SE, segmento externo; SI, segmento interno;
VS, vesıculas sin�apticas. (Modificado de Lentz TL: Cell Fine Structure:
An Atlas of Drawings of Whole-Cell Structure. Philadelphia, Saunders, 1971.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

. Los conos, c�elulas alargadas quemiden unos 60mmde largo� 1,5mm de di�ametro (fig. 22.7),funcionan de manera similar a los bastones exceptoque rinden mucho m�as con la luz fuerte que en laoscuridad, y contienen el fotopigmento yodopsina,del que existen tres variedades diferentes. Cada unaposee distintas regiones de la opsina, y unasensibilidad m�axima frente a alguno de los trescolores del espectro: rojo, verde y azul. La estructurade los conos es parecida a la de los bastones, salvo enlos siguientes aspectos:. El segmento externo tiene forma de cono.. Los discos est�an sujetos a la membranaplasm�atica.
. La proteına producida en el segmento internose introduce en todos los discos.
. Los conos son sensibles a los colores.
. El reciclado del fotopigmento no necesita a lasc�elulas epiteliales pigmentarias.
. La membrana limitante externa no es unamembrana; en cambio, es la region de las zonas deadhesion creadas entre las c�elulas de M€uller (v. m�asadelante) y las c�elulas fotorreceptoras.
. La capa nuclear externa es una region ocupada porlos nucleos de los bastones y los conos.
. La capa plexiforme externa consta de sinapsis entrelos axones de las c�elulas fotorreceptoras y dendritasde las c�elulas bipolares y horizontales. Hay dos tiposde sinapsis: sinapsis planas y sinapsis invaginadas.En estas ultimas, una dendrita de una c�elula bipolarm�as una dendrita de cada una de dos c�elulashorizontales constituyen una trıada. En las sinapsisinvaginadas se observan bandas sin�apticas quecapturan y colaboran en la distribucion de lassustancias neurotransmisoras.
. La capa nuclear interna alberga las regionesnucleares de cuatro tipos de c�elulas:. Cada neurona bipolar puede recibir senalesdesde docenas de bastones que ofrecen laposibilidad de sumar sus datos, lo que permitepotenciar la informacion correspondiente a unaintensidad lumınica baja. Sin embargo, cadacono tambi�en aporta estımulos a varias
neuronas bipolares, lo que multiplica lainformacion visual. Los axones de las c�elulasbipolares hacen sinapsis sobre las dendritas delas c�elulas ganglionares.
. Las c�elulas horizontales controlan ymodulan lasrelaciones sin�apticas entre las c�elulasfotorreceptoras y las c�elulas bipolares.
. Las dendritas de las c�elulas amacrinasmantienenun ıntimo contacto con las sinapsis entre lasc�elulas ganglionares y las c�elulas bipolares,y transmiten su informacion a las c�elulasinterplexiformes, que influyen sobre la actividadde las c�elulas horizontales y bipolares.
. Las c�elulas de M€uller se extienden entre el cuerpovıtreo y el segmento interno de los bastones y losconos, donde crean zonas de adhesion con lasc�elulas fotorreceptoras en la membrana limitanteexterna. Estas c�elulas actuan como c�elulas desost�en.
. La capa plexiforme interna es una region complejadonde los axones y las dendritas de las c�elulasbipolares, ganglionares y amacrinas se mezclan yconectan entre sı, formando sinapsis planas einvaginadas. Las sinapsis invaginadas constan delaxon de una c�elula bipolar y dos dendritas de unac�elula amacrina y una c�elula ganglionar o unadendrita de cada una de estas dos c�elulas diferentes,constituyendo una dıada.
. Los somas celulares de las grandes c�elulasganglionares multipolares est�an situados en lacapa de las c�elulas ganglionares. Lahiperpolarizacion de los bastones y los conos lasactiva para generar un potencial de accion que sepropaga a lo largo de sus axones hasta las �areasvisuales del enc�efalo.
. La capa de fibras del nervio optico es la region de laretina donde los axones amielınicos de las c�elulasganglionares se combinan para originar fibrasnerviosas. Cuando estos axones perforan laesclerotica, se vuelven mielınicos.
. La membrana limitante interna es la capa m�asinterna de la retina y est�a integrada por la l�aminabasal de las c�elulas de M€uller.
314
22
ORGANOSDELOSSEN
TIDOS

CONSIDERACIONES CL�INICAS
Existen dos tipos b�asicos de degeneracionmacular: humeda y seca. Alrededor del 10 al 15%delos casos de degeneracion macular son del tipohumedo, que al principio se manifesto como el tiposeco. En el tipo humedo de degeneracion macular,crecen vasos sanguıneos anormales en la
profundidad de la retina y de la m�acula, con el riesgode que sangren o dejen salir lıquido que haga que la
m�acula se abombe, lo que se traduce en unadistorsion o una destruccion de la vision central
r�apida y grave. Se han empleado distintos tipos de
tratamiento con l�aser para su resolucion, tan solo
con un �exito parcial al frenar el procesodegenerativo. Asimismo, las cicatrices que deja su
aplicacion pueden afectar a la m�acula, y provocar
una mayor p�erdida de vision. M�as recientemente, seha descubierto una proteına en el ojo, llamada factorde crecimiento endotelial vascular (VEGF). Su accionfomenta la produccion de vasos sanguıneos. Se
est�an creando f�armacos para inhibir el VEGF albloquearlo o evitar que se una a los elementos queestimularıan el crecimiento. En este momento, seadministran tres tipos de inhibidores del VEGF parasu tratamiento mediante inyecciones intraocularesdurante un perıodo prolongado. Su utilizaciontemprana ha ofrecido resultados positivos pararetardar la degeneracion, y en algunos casos hamejorado la agudeza visual.
315
22
ORGANOSDELOSSEN
TIDOS
Figura 22.7 Estructura de un baston y un cono. C, pedıculo de conexion;
CB, cuerpo basal; Ce, centrıolo; M, mitocondrias; RN, region nuclear;
RS, region sin�aptica; SE, segmento externo; SI, segmento interno;
VS, vesıculas sin�apticas. (Modificado de Lentz TL: Cell Fine Structure: AnAtlas of Drawings of Whole-Cell Structure. Philadelphia, Saunders, 1971.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

ANEJOS OCULARES
Los anejos oculares son la conjuntiva, los p�arpados yel aparato lagrimal.
. La conjuntiva es una mucosa transparente que est�acompuesta por un epitelio cilındrico estratificadocon c�elulas caliciformes sobre un tejido conjuntivolaxo. Reviste la cara interna de los p�arpados comoconjuntiva palpebral, y se refleja sobre laesclerotica de la superficie anterior del globo ocularcomo conjuntiva ocular. Cuando la conjuntivaocular llega al limbo de la cornea, deja de tenerc�elulas caliciformes y se convierte en el epiteliopavimentoso estratificado de la cornea.
. Los p�arpados son repliegues de piel fina quecierran la superficie anterior del ojo. Sus bordescontienen las pestanas, que est�an dispuestas enfilas de tres o cuatro, y carecen del musculo erectordel pelo. Las gl�andulas de Moll, gl�andulassudorıparas modificadas, desembocan en losfolıculos de las pestanas. Las gl�andulas deMeibomio, gl�andulas seb�aceas modificadas, est�anen el interior de las l�aminas tarsales. Estas ultimasson unas cubiertas de tejido conjuntivo engrosadoque sustentan cada p�arpado, y las gl�andulas deMeibomio producen una secrecion aceitosa que semezcla con las l�agrimas y retrasa su evaporacion.Otras gl�andulas seb�aceas modificadas m�aspequenas, las gl�andulas de Zeis, est�anrelacionadas con las pestanas, y vierten sussecreciones sobre sus folıculos.
. El aparato lagrimal est�a integrado por las gl�andulaslagrimales, los conductillos lagrimales, el sacolagrimal y el conducto nasolagrimal.. La gl�andula lagrimal es una gl�andula serosatubuloalveolar compuesta cuyos �acinos desecrecion est�an rodeados de c�elulasmioepiteliales. La gl�andula est�a localizada fueradel saco conjuntival; sin embargo, el lıquidosegregado (l�agrimas) se vuelca en este a trav�es de6-12 conductos secretores. Las l�agrimas, formadasprincipalmente por agua con lisozima, unasustancia antibacteriana, recorren los conductossecretores hacia el saco conjuntival.
. Cuando baja el p�arpado superior, barre lasl�agrimas en sentido medial para introducirlas en
el punto lagrimal, una pequena abertura cerca delos bordes mediales de los p�arpados superior einferior.
. Cada punto lleva hacia los conductilloslagrimales que confluyen en un conducto comunencaminado hacia el saco lagrimal, la porcionsuperior dilatada del conducto nasolagrimal queacaba en el suelo de la cavidad nasal bajo el meatoinferior.
Oıdo (aparato vestibulococlear)
Eloıdo actua comoel organode la audicion ydel equili-brio, y est�a dividido en tres partes: oıdo externo, oıdomedio (cavidad timp�anica) y oıdo interno (fig. 22.8).
El oıdo externo est�a constituido por la oreja(pabellon auricular), el conducto auditivo externo yla membrana timp�anica (v. fig. 22.8).
. Unas l�aminas irregulares de cartılago el�asticoconfiguran el armazon de la oreja, que sigue por elcartılago del conducto auditivo externo. El pabellonauricular est�a cubierto por una piel fina y muyadherida.
. El conducto auditivo externo se encuentrarevestido por una piel fina que posee folıculospilosos, gl�andulas seb�aceas y gl�andulasceruminosas (gl�andulas sudorıparasmodificadas) que elaboran cerumen (cera). El peloy el cerumen contribuyen a frustrar la entrada de losobjetos hacia los territorios profundos delconducto.
. La membrana timp�anica, que tapa el sector m�asprofundo del conducto auditivo externo,representa el tabique de cierre entre la primerahendidura farıngea y la primera bolsa farıngea. Susuperficie externa est�a compuesta por un epitelioderivado del ectodermo, mientras que la internaest�a tapizada por un epitelio derivado delendodermo. Unos cuantos elementos dispersos detejido conjuntivo mesod�ermico se hallan situadosentre estas dos caras. Las ondas sonoras setransmiten a trav�es del conducto auditivo externo,haciendo vibrar la membrana timp�anica. Estasvibraciones se trasmiten a la cadena de huesecillosdel oıdo medio.
316
22
ORGANOSDELOSSEN
TIDOS

CONSIDERACIONES CL�INICAS
La conjuntivitis es una inflamacion de laconjuntiva que puede proceder de multiplesorıgenes, como las infecciones bacterianas y
vıricas (en cuyo caso la situacion tambi�en sedenomina conjuntivitis aguda bacteriana) y laslesiones oculares, pero en la mayorıa de lascircunstancias se debe a la exposicion a
al�ergenos. Sus sıntomas consisten enenrojecimiento de la esclerotica, irritacion,prurito y lagrimeo con descarga inconstante de
los p�arpados. Los casos de conjuntivitis vırica ybacteriana son contagiosos y requieren un
tratamiento m�edico, mientras que las dem�ascausas pueden solucionarse en unos cuantosdıas o en cuestion de una o dos semanas.
Cuando el cuadro persiste, un m�edico deberıaevaluar al paciente porque algunas formas deconjuntivitis llegan a ocasionar una ceguerasi no se las trata.
CONSIDERACIONES CL�INICAS
La conexion con la faringe est�a abierta al tragar,bostezar y sonarse la nariz, lo que permite igualar
la presion a�erea a ambos lados de la membrana
timp�anica. El diferencial de presiones puede
sentirse durante un descenso r�apido al aterrizar enun avion. La deglucion normalmente descargaesta presion sobre el oıdo al abrir la trompaauditiva en la faringe.
317
22
ORGANOSDELOSSEN
TIDOS
Figura 22.8 Anatomıa del oıdo. (Tomado deGartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 527.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

OIDO MEDIO
El oıdo medio (cavidad timp�anica) (fig. 22.9) est�alocalizado en la porcion petrosa del hueso temporal, yes un espacio lleno de aire entre la membranatimp�anica y la membrana que cubre la ventana oval.Hacia atr�as comunica con las celdillas neum�aticasmastoideas y hacia delante con la faringe a trav�es dela trompa auditiva (trompa de Eustaquio). Los treshuesecillos ocupan este espacio, que est�a tapizadopor un epitelio pavimentoso simple, que continua elde la cara interna de lamembrana timp�anica. La paredosea de la cavidad timp�anica se encuentra sustituidapor cartılago al acercarse hacia la trompa auditiva, y elrevestimiento epitelial cambia a un epitelio cilındricociliado seudoestratificado. La l�amina propia contieneen esta region numerosas gl�andulas mucosas que aca-ban en la luz de la cavidad timp�anica; y, cerca de sudesembocadura en la faringe, aparecen c�elulas calici-formes y tejido linf�atico.
En la paredmedial de la cavidad timp�anica hay doshuecos en la pared osea cerrados por una membrana:las ventanas oval y redonda, que conectan la cavidaddel oıdo medio con el oıdo interno.
. La superficie interna de lamembrana timp�anica est�aunida a la membrana de la ventana oval por los treshuesecillos: elmartillo, el yunque y el estribo. Estoselementos trasmiten las vibraciones de lamembrana timp�anica a la membrana de la ventanaoval y las amplifican.
. Dos pequenos musculos estriados –el musculotensor del tımpano inervado por el nerviotrig�emino (NC V) y el musculo del estriboinervado por el nervio facial (NC VII)– sirven paramodular las vibraciones de la membranatimp�anica y los movimientos de las articulacionesentre los huesos.
OIDO INTERNO
El oıdo interno (v. fig. 22.9) est�a integrado por ellaberinto oseo y el laberinto membranoso que seencuentra suspendido en su interior.
. El laberinto oseo (fig. 22.10), alojado en laporcion petrosa del hueso temporal, est�a
recubierto por endostio y queda separado dellaberinto membranoso por el espacioperilinf�atico lleno de perilinfa. Su porcion centrales el vestıbulo, tras el cual se halla el aparatovestibular, que consta de los tres conductossemicirculares (oseos) (superior, posterior ylateral), que nacen y regresan al vestıbulo. Encada caso, un extremo est�a ensanchado y sedenomina ampolla. Suspendidos dentro de ellosest�an los conductos semicirculares(membranosos), que forman parte del laberintomembranoso. La pared lateral del vestıbulocontiene las ventanas oval y redonda cerradaspor una membrana. Tambi�en se originan en elvestıbulo otras regiones especializadas dellaberinto membranoso, el utrıculo y el s�aculo.Por delante del vestıbulo est�a la coclea, unespacio en espiral vaciado en el penasco delhueso temporal, que gira sobre sı mismo dosveces y media alrededor de una columna oseacentral llamada modıolo, y su cornisa de hueso,la l�amina espiral osea, que facilita un modo deentrada a los vasos sanguıneos y al ganglioespiral del componente coclear del nerviovestibulococlear.
. El laberinto membranoso (v. fig. 22.10),constituido por un epitelio de origen ectod�ermico,se encuentra colgando del laberinto oseo por unasfibras de tejido conjuntivo. Da lugar al s�aculo, elutrıculo, los conductos semicirculares(membranosos) y el conducto coclear. Laendolinfa, un lıquido viscoso, circula dentro dellaberinto membranoso. El s�aculo y el utrıculoest�an conectados entre sı por un pequenoconducto. Asimismo, cada uno posee pequenosconductos que se reunen para generar el conductoendolinf�atico, cuyo extremo ciego recibe elnombre de saco endolinf�atico. Otro pequenoconducto que va del s�aculo al conductococlear es el ductusreuniens. Las regionesespecializadas del s�aculo (m�acula sacular)y del utrıculo (m�acula utricular) son receptoresque detectan la orientacion de la cabeza en elespacio y su aceleracion. Las dos m�aculastienen c�elulas receptoras neuroepiteliales y otrasque no lo son.
318
22
ORGANOSDELOSSEN
TIDOS

319
22
ORGANOSDELOSSEN
TIDOS
Figura 22.10 Coclea del oıdo interno. A. Anatomıa del laberinto oseo. B. Anatomıa del laberinto membranoso. C. Anatomıa del
laberinto sensitivo. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 528.)
Figura 22.9 Anatomıa del oıdo. (Tomado deGartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 527.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

. Hay dos tipos de c�elulas en ambasm�aculas que noson receptores, c�elulas claras y c�elulas oscuras,cuya funcion se desconoce; aunque se piensa quelas c�elulas claras tal vez absorban la endolinfa,mientras que las oscuras controlen sucomposicion.
. Dos tipos de c�elulas receptoras (fig. 22.11)est�an presentes en ambas m�aculas: las c�elulasciliadas (c�elulas neuroepiteliales) de tipo I yde tipo II. Ambas clases poseen un unicocinocilio y entre 50 y 100 estereociliosalineados en hileras. Las c�elulas de sost�en seapoyan sobre la l�amina basal y se cree quemantienen a las c�elulas ciliadas o producenendolinfa. El componente vestibular del nerviovestibulococlear se ocupa de las c�elulas ciliadas(v. fig. 22.11). Los estereocilios de las c�elulasciliadas est�an sumergidos en una masagelatinosa espesa, la membrana otolıtica, cuyasuperficie contiene los otolitos o estatoconias(cristales de carbonato c�alcico).
. El laberinto membranoso se prolonga desde elutrıculo por los tres conductos semicircularesmembranosos (fig. 22.12) alojados en susrespectivos conductos semicirculares oseos. Losextremos laterales dilatados de los tres conductosse denominan ampollas y albergan unos puntosreceptores especializados llamados crestasampulares. Cada cresta ampular manifiesta unrelieve dotado de c�elulas ciliadas neuroepitelialesapretadas en cuna entre las c�elulas de sost�en,todas ellas sobre la l�amina basal. Las c�elulasciliadas son parecidas a las que hay en el utrıculo yen el s�aculo. La masa gelatinosa situada sobre lascrestas ampulares es la cupula, pero en este casocarece de otolitos.
. El conducto coclear (rampa media), que naceen el laberinto membranoso del s�aculo, es unorgano receptor que se encuentra dentro de lacoclea osea. Tiene aspecto de cuna y est�arodeado por sus dos lados por perilinfa. Dosmembranas del conducto coclear son las queconstituyen la cuna. La que forma el techo es lamembrana vestibular, mientras que la queforma el suelo es la l�amina basilar. Entre ambasel conducto coclear queda aislado de la perilinfaque lo rodea. El compartimento lleno deperilinfa que hay por encima de la membranavestibular es la rampa vestibular, y el que est�abajo la l�amina basilar es la rampa timp�anica.La comunicacion entre ambos tiene lugar en elhelicotrema.
. La membrana vestibular consta de dos capas deepitelios pavimentosos separadas por una l�aminabasal. La l�amina basilar sostiene el organo deCorti (fig. 22.13) y posee varios tipos de c�elulas,algunas de cuyas funciones no se conocen, y otras,como en el caso de las c�elulas interdentales,segregan la membrana tectoria, una masagelatinosa que est�a sobre el organo de Corti.Los estereocilios de las c�elulas receptorasespecializadas est�an inmersos en la membranatectoria. Las c�elulas neuroepiteliales (ciliadas)del organo de Corti transfieren los impulsosauditivos. Se trata de las c�elulas ciliadas internasy externas.
. Las c�elulas ciliadas internas est�an organizadas enuna sola fila y rodeadas por c�elulas de sost�en. Sonpequenas y contienen un nucleo de posicioncentral, abundantes mitocondrias, retıculoendopl�asmico rugoso, retıculo endopl�asmico liso ypequenas vesıculas. En la zona basilar haymicrotubulos. Los estereocilios, de 50 a60 adoptando una forma de «V», emanan desde lasuperficie apical. Su centro lleva microfilamentosentrecruzados con fimbrina. Asimismo, losmicrofilamentos de los estereocilios se combinancon la red terminal. Un cuerpo basal y un centrıoloest�an presentes en la region apical de las c�elulasciliadas internas. Sus membranas basales hacensinapsis con las fibras aferentes del componentecoclear del nervio vestibulococlear.
. Las c�elulas ciliadas externas, situadas cerca dellımite externo del organo de Corti, est�an alineadasen hileras de tres a lo largo de toda su extension. Setrata de unas c�elulas cilındricas alargadas cuyosnucleos ocupan una localizacion basal. Sucitoplasma presenta retıculo endopl�asmico rugoso ynumerosas mitocondrias de distribucion basal. Enla parte inmediatamente interna a la membranaplasm�atica lateral hay una estructura llamadaentramado cortical compuesta por filamentos de5 a 7nm entrelazados con filamentosm�as delgados.Se supone que este elemento sirve para sostener lasc�elulas ciliadas y oponerse a su deformacion. Unos100 estereocilios colocados en forma de «W» surgende la superficie apical de las c�elulas ciliadas externas.Adem�as, como su longitud varıa, est�an ordenadosde manera escalonada segun su extension. Estasc�elulas carecen de cinocilio pero sı que tienen uncuerpo basal. Las fibras aferentes y eferentes delcomponente coclear del nervio vestibulococlearhacen sinapsis en las porciones basilares de lasc�elulas ciliadas.
320
22
ORGANOSDELOSSEN
TIDOS

321
22
ORGANOSDELOSSEN
TIDOS
Figura 22.11 Morfologıa de las c�elulas neuroepiteliales (ciliadas) de tipo I y de tipo II de lasm�aculas del s�aculo y el utrıculo. (Modificado
de Lentz TL: Cell Fine Structure: An Atlas of Drawings of Whole-Cell Structure. Philadelphia, Saunders, 1971.)
Figura 22.12 C�elulas ciliadas y c�elulas de sost�en en una de las crestas ampulares de los conductos semicirculares membranosos.
(Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 531.)�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

FUNCIONES DEL OIDO
La doble funcion del oıdo consiste en detectar la posi-cion del cuerpo y su movimiento en el espacio tridi-mensional (funcion vestibular) y discernir el sonido(funcion coclear).
. La funcion vestibular del oıdo interno identificalos cambios en la velocidad de los movimientosrectilıneos o circulares de la cabeza, aspecto quedepende del aparato vestibular: el utrıculo,el s�aculo y los conductos semicircularesmembranosos.. La endolinfa de las ampollas del utrıculo y els�aculo responde a losmovimientos rectilıneos dela cabeza desplazando los otolitos y la membranaotolıtica. A raız de este fenomeno, se doblan losestereocilios de las c�elulas ciliadas y se despolarizala membrana plasm�atica. La modificaciondel potencial en reposo de la membrana disparapotenciales de accion que se transmiten hacia elcomponente vestibular del nerviovestibulococlear, encargado de conducir losimpulsos hacia el enc�efalo para su procesamiento.
. Las c�elulas ciliadas neuroepiteliales de las crestasampulares de la cupula en los conductossemicircularesmembranosos reaccionan frente alos movimientos circulares de la cabeza de unmodo semejante a como responden el utrıculo yel s�aculo frente a los movimientos rectilıneos. Losestereocilios de las c�elulas ciliadas en las crestasampulares se deforman ante la oscilacion de laendolinfa en los conductos semicirculares.La inclinacion de los estereocilios deriva en eldesencadenamiento de los potenciales de accionen las c�elulas ciliadas, que se trasladan alcomponente vestibular del nerviovestibulococlear para su envıo hacia el enc�efalocon el fin de procesarlo.
. Los movimientos rectilıneos y circulares de lacabeza requieren la contraccion de los musculos
estriados responsables de mantener el equilibrio.Para ello, el cerebro debe interpretar lainformacion recibida desde las c�elulas ciliadas delaparato vestibular y preparar una respuesta casiinstant�anea para evitar que la persona pierda elequilibrio y se caiga.
. La funcion coclear (v. fig. 22.13) es responsabilidadde las tres regiones del oıdo: oıdo externo, medio einterno.. Las ondas sonoras que llegan al oıdo y recorren elconducto auditivo externo acaban en lamembrana timp�anica, y la ponen enmovimiento.
. Esta oscilacion se transforma en energıa mec�anicaque moviliza el martillo y los otros doshuesecillos de la cavidad timp�anica.
. Las vibraciones de la membrana timp�anica seamplıan unas 20 veces cuando la energıa alcanzala base del estribo, donde golpea la membrana dela ventana oval.
. Dos pequenos musculos estriados en la cavidaddel oıdo medio modulan los desplazamientos delmartillo y del estribo.
. Las oscilaciones de la membrana de la ventanaoval crean ondas de presion en la perilinfacontenida en la rampa vestibular, que atraviesanel helicotrema y siguen por la rampa timp�anica,provocando unos movimientos ondulares en lal�amina basilar.
. Este balanceo origina unas fuerzas de arrastrelaterales sobre los estereocilios de las c�elulasciliadas sumergidos en la membrana tectoria.
. Al desviarse los estereocilios, la c�elula sedespolariza y genera un impulso que se transmitehacia las fibras nerviosas aferentes delcomponente coclear del nervio vestibulococlearpara su procesamiento en el enc�efalo.
. Los sonidos de alta frecuencia se detectan en elextremo inferior del organo de Corti (v. fig. 22.13),mientras que los de baja frecuencia lo hacen en elsuperior, cerca de su v�ertice.
322
22
ORGANOSDELOSSEN
TIDOS

CONSIDERACIONES CL�INICAS
El sındrome de M�eni�ere es una anomalıa episodicadel oıdo interno que provoca una multitud de
sıntomas, como v�ertigo intenso, acufenos (zumbidode oıdos), hipoacusia fluctuante y sensacion depresion o de dolor en el oıdo afectado. El trastornosuele darse en un solo lado y es una causa frecuentede sordera parcial. Sus sıntomas van ligados a unaumento en el volumen de endolinfa dentro de unaporcion del oıdo interno, lo que hace que el laberintomembranoso se hinche o se dilate, proceso
denominado edema o hidropesıa endolinf�atica.Muchos expertos piensan que la rotura del laberintomembranoso deja que la endolinfa se mezcle con laperilinfa, situacion capaz de producir los sıntomas
del sındrome de M�eni�ere. Otros especialistas est�aninvestigando varios posibles orıgenes del sındrome,
entre ellos factores ambientales y diet�eticos. Aunque
no hay remedio para el sındrome de M�eni�ere, sussıntomas pueden corregirse satisfactoriamentemediante una reduccion en la retencion de loslıquidos corporales y con cambios alimentarioscomo los regımenes hiposodicos o sin sal, aparte deabstenerse de la cafeına o el alcohol.
La hipoacusia neurosensitiva (sordera nerviosa)se da especıficamente en el organo de Corti cuando
se alteran o se destruyen las c�elulas ciliadas. Lahipoacusia neurosensitiva puede deberse a distintascausas, como la herencia, el envejecimiento, algunaenfermedad, las infecciones o la exposicion
prolongada a ruidos intensos. Pocas veces resultadanado el tronco nervioso que se dirige hacia el
enc�efalo. En cambio, lo m�as habitual es que se
afecten las c�elulas ciliadas del organo de Corti, quesirven para enviar informacion hacia el nervio
coclear, en forma de senales el�ectricas. Cuando se
ha lesionado una cantidad importante de c�elulasciliadas, la persona sufre una hipoacusia profunda ograve, y los audıfonos son incapaces de paliar elproblema. En los casos de una hipoacusia acusada,puede estar indicado el implante coclear.
La sordera de conduccion surge cuando sedificulta o se bloquea la transmision de las ondas
sonoras a trav�es del oıdo externo o el oıdo medio, loque impide su recepcion en el oıdo interno. Entre lospadecimientos que dan lugar a una sordera deconduccion figura la entrada de cuerpos extranos, la
rotura de la membrana timp�anica, los tapones decerumen, la otitis media y la otoesclerosis (en la quela base del estribo queda fija en la ventana oval).
La otitis media es una infeccion frecuente quesucede en la cavidad del oıdo medio, sobre todo enlos ninos pequenos, a partir de una infeccionrespiratoria que se haya extendido a la trompaauditiva. En este cuadro, existe una acumulacionlıquida en la cavidad del oıdo medio que restringeel movimiento de la cadena de huesecillos, lo quelimita la capacidad para oır en el lado afectado.
Su tratamiento m�as habitual es con antibioticos.
323
22
ORGANOSDELOSSEN
TIDOS
Figura 22.13 Organo de Corti. (Tomado de Gartner LP, Hiatt JL: Color Textbook of Histology, 3rd ed. Philadelphia, Saunders, 2007, p 532.)
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.


�Indice alfab�etico
AAV. Vease Anastomosis auriculoventricular(AAV).
ABP. Vease Proteına(s) de union a androgenos(ABP).
Absorcion, intestino delgado, 246, 247fAcetil-coenzima A (CoA), 20Acetilcolina (ACh), 100, 121t, 124, 200interaccion con secretina, 240presion arterial, 154produccion de �acido clorhıdrico, 242vesıcula biliar, 259
Acetilcolinesterasa, 100Acetiltransferasa de colina-O, 100ACh. Vease Acetilcolina (ACh).Acido(s)araquidonico, 66gamma-aminobutırico (GABA), 121tglicocolico, 258glut�amico, 121tgraso, 66de cadena larga, intestino delgado, 246
hialuronico, 40, 41f, 41t, 47c, 70, 76, 92hipocloroso, 138peryodico (Schiff), 3ttaurocolico, 258uronico, 40–42
Acidofilos, 190mamotropos, 190
Aciltransferasas, 246Acinos, 224pulmonares, 224
Acn�e, 215cAcomodacion visual, 308Acoplamiento, 90Acromegalia, 89cAcrosina, 280Acrosoma, 290ACTH. Vease Hormona(s) adrenocorticotropa
(ACTH).Actina, 96, 140Actina F, 98Actina G, 24, 98a-actininas, 46, 96, 104, 106Activacion plaquetaria, 140Activador de plasminogeno, 140Activina, 274Acuaporinas, 10Addison, enfermedad, 201c, 209cAdenocarcinomas, 57cAdenohipofisis, 189, 189f, 189t, 189, 191fAdenoides, 186Adenomas hipofisarios, 192cADH. Vease Hormona(s) antidiur�etica (ADH).Adhesion(es)de matriz celular ligadas a actina, 52plaquetaria, 140
Adipocitos, 64–66, 65falmacenamiento y liberacion de grasa, 66,
67fmultiloculares, 62uniloculares, 62
ADN, 28, 30circular, 20espaciador, 28
ADP. Vease Difosfato de adenosina (ADP),plaquetas.
Adrenalina, 66, 200, 203tAdventiciadel aparato digestivo, 238esof�agica, 238del intestino delgado, 244de la l�amina propia traqueal, 222uterina, 278vaginal, 284de vejiga urinaria, 270de la vesıcula biliar, 259
Aftas, 232cAGE. Vease Arteriolas glomerulares eferentes
(AGE).Agranulocitos, 136Agrecanos, 40, 76Agregacion plaquetaria, 140Aguaflujo hacia c�elula parietal, 242resorcion, calcificacion osea, 88
Agujeroapical de raız del diente, 230, 233ciego, 236
Albinismo, 23c, 209cAlbumina en plasma, 133tAldosterona, 104, 200, 203tunion, 264
Aletas nasales, 218Alport, sındrome, 47cAlv�eolodental, 230, 232pulmonar, 226, 227f, 234, 234f
Alzheimer, enfermedad, 129cAmeloblastos, 230, 233, 233fAmıgdalas, 168, 185-6farıngeas, 186linguales, 186, 236palatinas, 186
Amilasa salival, 230, 250Amino�acidos, intestino delgado, 246Aminoacilo ARNt, 14Aminoazucares, 40–42Aminopeptidasas, 246del intestino delgado, 244
AMP. VeaseMonofosfato de adenosina (AMP).AMPc. VeaseMonofosfato de adenosina cıclico
(AMPc).Ampolla(s)del conducto semicircular, 318del oviducto, 278del s�aculo, 322del utrıculo, 322del vaso deferente, 296
Anafase, 36, 37fAnafase I, meiosis, 38, 39fAnafilaxia sist�emica, 70cAnastomosisarteriovenosas, pene, 302auriculoventricular (AAV), 158, 159f
Androgenos, 200, 203treparacion osea, 91c
Androstenediona, 200, 203t, 274, 276Anemia drepanocıtica, 15cAnencefalia, 109cAneurismas, 153cAngiotensina I, 268presion arterial, 156
Angiotensina II, 158, 268presion arterial, 156
Angiotensinogeno, 268presion arterial, 156
Anhidrasa carbonica, 134g�astrica, 242intercambio gaseoso, 228
Anillode actina, 80en «C», 222citopl�asmico, 26, 27fcontr�actil, 36distal, 26, 27fespermatogenia, 290luminal, 26, 27fnuclear, 26, 27fnucleopl�asmico, 26, 27f
Anticodones, 14Anticuerpos, 68, 170, 171f, 172tde membrana, 170
Antıgeno(s), 169, 182casociado a funcion leucocitaria-1 (LFA-1),
139cdel complejo principal de
histocompatibilidad II, 136independientes del timo, 185leucocitario humano de clase II
(HLA clase II), 136del Rh, 134, 135c
Antıgenos A, 134Antıgenos B, 134Antıgenos C, 134Antıgenos D, 134Antıgenos E, 134Antro ov�arico, 274Apaf1.Vease Proteına(s) adaptadora activadora
de procaspasas apoptosicas (Apaf1).Aparatocardiovascular, 152–164arterias, 152, 154–156, 155t
estructuras sensoriales, 154regulacion de presion sanguınea,
154–156capilares, 152, 156–158, 157f
histofisiologıa, 158, 159firrigacion, regulacion, 158, 159f
corazon, 162–164, 163f, 165fenvejecimiento, 157ctunicas de vasos, 152–154, 153f, 155fvenas, 152, 160, 161t
circulatorio. Veanse Aparato cardiovascular;Sistema linf�atico.
digestivo. Veanse tambien Cavidad bucal;Tubo digestivo; organos especıficos.
gl�andulas, 250–258. Vease tambienGl�andula(s).
hemorragia, 239cgenitalfemenino, 272–284. Veanse tambien
organos especıficos.ciclo menstrual, 280, 281fdesarrollo de la placenta, 282, 282tfecundacion, 280, 281fimplantacion, 282, 283fovulacion, 276, 277f
masculino, 286–302, 287f. Veanse tambienorganos especıficos.
del huso mitotico, 36lacrimal, 316respiratorio, 218–228porcion
conductora, 218–224, 219t, 221frespiratoria, 218, 226–228, 227f
urinario, 260–270aparato yuxtaglomerular, 264, 265fasa de Henle, 260, 262c�apsula de Bowman, 260–262, 261fcirculacion renal, 266, 267fformacion de orina, mecanismo, 266–268,
269fintersticio renal, 264tubulo(s)
colectores, 264distal, 260, 262–264, 265fproximal, 260, 262, 263f
vasos rectos, 268, 269fvıas excretoras, 270
yuxtaglomerular, 264, 265ffiltrado, control, 268
Ap�endice(s), 248epiploicos, 248
Apertura papilar, 308Aponeurosis, 94Apoptosis, 38, 170, 175, 178, 180, 182cApoptosomas, 39Aracnoides, 126, 127f
325� 2011 Elsevier Espana, S.L. Reservados todos los derechos

326
INDICEALFA
BETIC
O
Arbol bronquial, 224, 225fArcos reflejos simples, 103cArea cribosa, 268Arena cerebral, 202ARNmensajero (ARNm), 12, 30precursor (ARNm-pre), 30
ribosomico (ARNr), 12, 30de transferencia (ARNt), 30
ARNm. Vease ARN mensajero (ARNm).ARNm-pre. Vease ARN mensajero precursor
(ARNm-pre).ARNr. Vease ARN ribosomico (ARNr).ARNt, 14iniciador, 14. Vease tambien ARN de trans-
ferencia (ARNt).Aromatasa, 274Arrugas, 240Arteria(s), 152, 154–156, 155tarqueadas, 266, 267f, 278central, 183corticales largas, 200espl�enica, 183, 184festructuras sensoriales, 154helicoidales, 278, 302hipofisarias inferiores, 190interlobulares, 266lobulares, 266en m�edula osea, 142peniciliares, 184rectas, 278regulacion de la presion sanguınea,
154–156renal, 260, 266, 267fsegmentarias, 266testicular, 286trabeculares, 183
Arterioesclerosis, 157cArteriolasde distribucion, hep�aticas, 254glomerulares eferentes (AGE), 266hep�aticas de la entrada, 254rectas, 266, 268, 269f
Articulacion(es), 92, 93fde diartrosis, 92, 93fde sinartrosis, 92, 93f
Artritisartrosis, 93cpsori�asica, 207creumatoide, 93c
Artrosis, 93cAsa cervical, 232Asma, 70cAstasdorsales, 126ventrales, 126
Astrocitos, 112, 128fibrosos, 112protopl�asmicos, 112
Ateromas, 157cAterosclerosis, 157cATP. Vease Trifosfato de adenosina (ATP).ATPasabomba de Na+ K+, 10de tubulo distal, 264de la vesıcula biliar, 258
ATPasa H+ K+, 240Auerbach, plexo, 109c, 124, 238, 244, 246Aurıcula(s)cardıacas, 164, 165fderecha del corazon, 164, 165fizquierda, 164, 165fpulmonar, 226
Autoinjertos, 91cAutorradiografıa, 5Autosomas, 28Axolema, 114Axonema, 50, 290Axonesmielinizados, 100de tipo Ib, 102
Bacterias fagocitadas, destruccion asistida porlinfocitos T, 178, 178f
Balsas lipıdicas, 8, 106BALT. Vease Tejido(s) linfoide asociado a los
bronquios (BALT).Banda 4.1, 134Banda A, 96Banda H, 94, 96, 97fBanda I, 94, 96, 97fBarorreceptores, 154Barr, cuerpos, 138Barrasdensas, 100terminales, 52
Barrerahematoencef�alica, 127c, 128hematogaseosa, 226hematotesticular, 288hematotımica, 181clipıdica, estrato granuloso, 206placentaria, 282, 282t
Basofilos, 70, 137t, 138, 190gonadotropos, 190
Bastonesdel esmalte (prismas), 230retinales, 312, 313f
Bazo, 168, 183–185, 184ffunciones, 183, 184firrigacion, 183, 184f, 185pulpablanca, 183, 185froja, 183, 185f
zona marginal, 183, 184fBellini, conductos, 264BFU-E. Vease Unidad formadora de brote-
eritrocitos (BFU-E).BHE. Vease Barrera hematoencef�alica.Bicarbonatog�astrico, 242pancre�atico, 252salival, 250
Bilirrubina, 258conjugada, 258libre, 258
Bilis, 258primaria, 256
Biliverdina, 258Birbeck, gr�anulos, 208Blastocele, 282Blastocisto, 282Blastomeros, 282Bloques de parafina, 2Boca, 230–236dientes, 230–234, 231c, 231flabios, 230, 232c, 231flengua, 236, 237fpaladar, 235
Bociono toxico, 195csimple, 199c
Bolsillos intercelulares, intestinodelgado, 244
Bombasde calcio, 88de Na+ K+, 116
Borde irregular, osteoclastos, 80Boton(es)terminal, 128de union, 96
Bowmanc�apsula, 260–262, 261fespacio, 260gl�andulas, 220membrana, 306
Bradicininas, 68, 69tpermeabilidad capilar, 158
Brazos de dineına, 50Broncoespasmo, 70cBronquıolos, 224respiratorios, 226, 227fterminales, 224, 225f
Bronquios, 224extrapulmonares, 224intrapulmonares, 224lobulares, 224primarios, 224secundarios, 224segmentarios, 224terciarios, 224
Brotes axonales, 130Bruch, membrana, 308Brunner, gl�andulas, 246Bulboolfatorio, 220piloso, 214
Burkitt, linfoma, 37c
C3c, 138Cabeza de sintetasa de ATP, 20Cadena(s)alfa de tropocol�ageno, 42ligeras, 171pesadas, 171de preprocol�ageno, 44
Cadenas J, 250Cadherinas, 54Calcificacionosea, 88zona, 88
Calcio, concentraciones s�ericas, manteni-miento, 92
Calcitonina, 80, 90, 92, 196, 198, 203tCaldesmona, 106C�alices renales, 270Call-Exner, cuerpos, 274Callo, 90externo, 90interno, 90
Calmodulina, 50Calostro, 284C�amaraanterior del ojo, 308ocular anterior, 308pulpar del diente, 230
Cambio de cloruro, 134, 228, 229fCanal(es)de acuaporinas, 268acuaporina-I, 262acuaporina-2, 264
de calcio, compuerta de voltaje, 100de calcio-sodio, 104central(es), 126, 158con compuerta, 10de ligando, 10de nucleotidos, 10de voltaje, 10
controlados con neurotransmisores, 10de desag€ue, 158de escapede potasio, 10, 116
ionicos, 10asociados a receptores, 118con compuerta de proteına G, 10
lentos de sodio, 104de liberacion de calcio, 96, 104de nutrientes, 142del hueso alveolar, 234
de potasio, compuerta de voltaje, 116, 118de la raız, 230r�apidos de sodio, 104semicirculares, 318sin compuerta, 10de sodiocon compuerta
de ligandos, 100lentos, 104
de voltaje, 116–118r�apidos, 104
Canalıculos, 82, 84biliares, 256intracelulares, 240lacrimales, 316

Canasta nuclear, 26, 27fC�ancerbiliar, 259cdiseminacion linf�atica, 167cde piel, 211cpulmonar, 229crenal, 270ctesticular, 294cvesical, 37c
Cap Z, 96Capabasalde gl�andulas sudorıparas ecrinas, 212uterina, 278
celular internadel cartılago, 76del periostio, 78, 82
coriocapilar, 308de c�elulasganglionares de la retina, 314limıtrofes de la duramadre craneal, 126
epitelial de la luz del aparato digestivo, 238espumosa, 132de fibras nerviosas opticas de la retina, 314fibrosa, 92externa
del cartılago, 76del periostio, 78, 82
funcional uterina, 278granulosa de la corteza cerebelosa, 130de hidratacion, 78luminal de gl�andulas sudorıparas ecrinas,
212menıngea, duramadre craneal, 126molecular de la corteza cerebelosa, 130muscularexterna
del ap�endice, 248esof�agica, 238g�astrica, 240, 242del intestino
delgado, 244grueso, 248
de luz del aparato digestivo, 238del oviducto, 278de la mucosa
del aparato digestivo, 238del ap�endice, 248g�astrica, 240, 242del intestino
delgado, 244grueso, 248
vaginal, 284neuronal de la retina, 310nuclearexterna de la retina, 314interna de la retina, 314
odontobl�astica de la pulpa dental, 230papilar de la dermis, 207t, 210periostica de la duramadre craneal, 126pigmentada de la retina, 310plexiformeexterna de la retina, 314interna de la retina, 314
reticular de la dermis, 207t, 210subendoc�ardica, 162
Capa/membrana sinovial, 92Capacitacion de espermatozoides, 278Caperuzadel cr�aneo, 82del microtubulo, 22
Capilares, 152, 156–158, 157fcontinuos, 94, 128, 156fenestrados, 156histofisiologıa, 158, 159firrigacion, regulacion, 158, 159flinf�aticos, 166, 167fsinusoidales, 156, 200
C�apsulaarticular, 92
del cristalino, 308pericelular, 76
Cara(s)cis, aparato de Golgi, 16intermedias, aparato de Golgi, 16trans, aparato de Golgi, 16
Cara E, 8, 9fCara P, 8, 9fCarbaminohemoglobina, 134Carcinoma(s), 57cadenocarcinoma, 57cc�elulasbasales, piel, 211cescamosas
de la cavidad bucal, 232cde piel, 211c
de transicion renal, 270cpulmonar, 229c
Cardiodilatina, 162Cardionatrina, 162Cardiopatıacoronaria, 165creum�atica, 163c
Cargo, 16Cariocinesis, 36Carioplasma, 8Cartılago, 74–76, 75f, 75tarticular, 74, 75f, 75t, 92condrocitos, 76cefectos de hormonas, 76, 77thistogenia y crecimiento, 76, 77f, 77tmatriz, 76
condrocitos, 76cefectos de hormonas, 76, 77tel�astico, 74, 75f, 75thialino (articular), 74, 75f, 75t, 92condrocitos, 76cefectos de hormonas, 76, 77thistogenia y crecimiento, 76, 77f, 77tmatriz, 76
histogenia y crecimiento, 76, 77f, 77tmatriz, 76
Caspasas, 38Catalasade hepatocitos, 256en peroxisomas, 18
Cataratas, 309cCat�astrofe, 22Catecolaminas, 200, 203tCatepsina K, 80Cav�eolas, 106Caveolina, 106Cavidadbucal, 230–236dientes, 230–234, 231c, 231flabios, 230, 232c, 231flengua, 236, 237fpaladar, 235propiamente dicha, 230
medular, 78, 82nasal, 218histofisiologıa, 220
peric�ardica, 164pleural, 228pulpar del diente, 230de resorcion, 90timp�anica, 318, 319f
CD9, 280CD95, 175, 178CDK. Vease Cinasa(s) dependientes de ciclinas
(CDK).C�elula(s), 2, 8acinares, 252adiposas. Vease Adipocitos.de almacenamiento de lıpidos, 256alveolares, tipo I, 226amadrinas de retina, 314APUD, 60argentafines, 60argirofilas, 60
basalesde corpusculos gustativos, 236del epidıdimo, 296del epitelio
olfatorio, 218respiratorio, 222
de la vesıculas seminales, 298caliciformesdel epitelio traqueal, 222, 223fcomo gl�andulas exocrinas unicelulares, 58,
59fdel intestino delgado, 244
centroacinares, 252en cepillo del epitelio respiratorio, 222ciliadasendometriales, 278del oviducto, 278
cilındricasciliadas del epitelio respiratorio, 222de la vesıculas seminales, 298
cimogenas, 240citoesqueleto, 22–24, 23f, 25fclaras, 196de gl�andulas sudorıparas ecrinas, 212, 213fde la vesıcula biliar, 258
condrogenas, 76, 78, 90de cromafines, 200deciduales (reaccion) endometriales, 278,
280dendrıticas, 182cfoliculares, 182c
de ganglios linf�aticos, 183de ganglios linf�aticos, 183interdigitantes
espl�enicas, 185en zona marginal espl�enica, 185
migratorias de ganglios linf�aticos, 183,182c
del timo, 180diana, 10, 58diploides, 280efectoras, 170endotelialesde ganglios linf�aticos, 183de glom�erulo, 266
ependimarias, 114, 126espermatog�enicas, 288–290, 289festromales ov�aricas, 272externas del pelo, 320foliculares, 272, 276foliculoestrelladas, 190ganglionares de la retina, 314de ganglios simp�aticos, 200germinales primitivas, 272gigantes de cuerpo extrano, 68, 136globosas del epitelio olfatorio, 218de glomo, 154de gr�anulo pequeno del epitelio respiratorio,
222de la granulosa(s), 256, 274, 276del cumulo, 274
hep�aticas estrelladas, 256horizontales de la retina, 314inclusiones, 22de insuficiencia cardıaca, 229cintercaladas de tubulos colectores, 264interdentales, 320intermedias de los corpusculos gustativos,
236internas del pelo, 320interplexiformes, 314intersticialesde gl�andula pineal, 202ov�aricas, 272
ligerasde corpusculos gustativos, 236del oıdo interno, 320
linfoides, interaccion, 176de luteına de la teca, 276madre, 136, 144, 145t, 146–148, 147f
327INDICEALFA
BETIC
O
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

C�elula(s) (cont.)hematopoy�eticas
multipotenciales (CMHM), 146, 148pluripotenciales (CMHP), 146–148,
147fneuronales, 131c
de membrana granulosa, 274de memoria, 170mesangialesextraglomerulares, 264intraglomerulares, 262
mesenquimatosas, 62, 70microgliales, 112de micropliegues, intestino delgado, 244mioepitelialesde gl�andulas
mamarias, 284salivales, 60, 250sudorıparas ecrinas, 60, 212, 213f
mitrales, olfato, 220mucoides de gl�andulas sudorıparas ecrinas,
212mucosasdel cuello, 240de gl�andulas salivales, 250
de musculo liso arterial, 152neuroepiteliales del oıdo interno, 320, 321fde la neuroglıa, 108, 112–114, 113f, 115fnulas, 136olfatorias, 218oscurasde corpusculos gustativos, 236de gl�andulas sudorıparas ecrinas, 212, 213fdel oıdo interno, 320
osteogenas, 78osteoprogenitoras, 76, 78, 88, 90oxıfilas, 198oxınticas, 240parafoliculares, 196parietales, 240peg, 278pilosas del oıdo interno, 320, 321fplasm�aticas, 68, 69f, 136, 170, 176, 183, 182cen zona marginal espl�enica, 185
de polvo, 226precursoras, 144, 145t, 146presentadoras de antıgenos (CPA)como c�elulas dendrıticas migratorias, 182cmacrofagos, 68, 136tipos, 176en zona marginal espl�enica, 185
principales, 198, 240, 264del epidıdimo, 296
progenitoras, 144, 145t, 146regenerativas, 240de capilares, 156g�astricas, 240del intestino delgado, 244
reticularesadventicia, 142del bazo, 185epiteliales, 180
medulares, 180estrelladas
espl�enicas, 185de ganglios linf�aticos, 183
de ganglios linf�aticos, 183del revestimientode hueso, 78sinusoidal, 256superficial g�astrico, 240
sat�elite, 94secretoras no ciliadas endometriales, 278de senalizacion, 10, 58septales, 226, 227fserosasdel epitelio respiratorio, 222de gl�andulas salivales, 250
del sistema neuroendocrino difuso (SNED),60, 188
del epitelio respiratorio, 222g�astricas, 240, 242del intestino delgado, 244, 259
superficiales de absorcion, intestino delgado,244
sustentaculares del epitelio olfatorio, 218de tipo Ide corpusculos gustativos, 236de cuerpo carotıdeo, 154
de tipo IIde corpusculos gustativos, 236de cuerpo carotıdeo, 154
de tipo III de corpusculos gustativos, 236de tipo IV de corpusculos gustativos, 236de tipo Aen capa sinovial de articulaciones de dia-
rtrosis, 92de tubulos colectores, 264
de tipo Ben capa sinovial de articulaciones de dia-
rtrosis, 92de tubulos colectores, 264
trabeculares de la aracnoides, 126transformadas por virus, destruccion por li-
nfocitos TH, 178, 177fde la vaina, 154vırgenes, 170yuxtaglomerulares, 264, 268
C�elulas C, 196C�elulas CR7+, 172C�elulas CR7�, 172C�elulas L, 71cC�elulas Mintestino delgado, 244tejido linfoide asociado a mucosas, 185
C�elulas P/D1, 71cC�elulas SNED. Vease C�elula(s) del sistema ne-
uroendocrino difuso (SNED).Cemento, 231, 233acelular, 231celular, 231
Cementoblastos, 231, 233Cementocitos, 231Cementoclastos, 231Centrıolos, 9f, 22, 23f, 36Centrodel apetito, 71cfibrilar de tincion p�alida, 32vasomotor, 154–156
Centroblastos, 183Centrocitos, 183Centromeros, 36Centrosde condrificacion, 76germinalesde ganglios linf�aticos, 183, 182cen la pulpa blanca espl�enica, 185
organizadores de microtubulos (COMT), 22,36
de osificacion, 84, 86, 86tprimarios, 84, 86, 86tsecundarios, 86, 86t
Centrosomas, 22, 36Cerebelo, 130Cerumen, 316CFU-basofilos, 146CFU-E, 146CFU-eosinofilos, 146CFU-G, 146CFU-GM, 146, 150CFU-linf. Vease Unidad formadora de colonias
de linfocitos (CFU-linf).CFU-M, 146, 150CFU-Meg, 146, 150Choque anafil�actico, 70cCicatrices gliales, 131cCiclasa de adenilato, 66Ciclinas G1, 34Ciclinas G1/S, 34Ciclinas M, 34
Ciclinas S, 34Ciclocelular, 34–38, 35fapoptosis, 38
menstrual, 280, 281ffase
lutea, 280, 282menstrual, 280secretora, 280, 282
Cigoteno, 38Cigoto, 280, 281fCilio(s), 50, 51fmovimiento, 52primarios, 52retinales, 312
Cinasa(s)de la cadena ligera de la miosina (MLCK),
106dependientes de ciclinas (CDK), 34
Cinasa A, 12Cinesina, 22Cinetocoros, 36Circunvoluciones, 128Cisterna(s), 16terminales, 96, 97f
Cistinuria, 11cCitocinas, 58hematopoyesis, 144origen y funciones, 174tsistema inmunitario innato, 168
Citocinesis, 36, 37f, 150Citoesqueleto, 22–24, 23f, 25fCitomorfosis, 205Citoplasma, 8Citoquımica, 4Citotrofoblasto, 282Clatrina, 16Claudinas, 54, 114CLIP. Vease Proteına(s) invariante asociada a
clase II (CLIP).Clones, 170, 182cCloruro g�astrico, 242Clostridium botulinum, 119cClostridium tetani, 101cCMHM. Vease C�elula(s) madre hemato-
poy�eticas multipotenciales (CMHM).CMHP. Vease C�elula(s) madre hemato-
poy�eticas pluripotenciales (CMHP).CoA. Vease Acetil-coenzima A (CoA).Coagulacion, 132pulpa roja espl�enica, 185
Co�agulos sanguıneos, 140Coanas, 218Coatomero I (COP I), 16Coatomero II (COP II), 16Coclea, 318Codones, 14de interrupcion, 14
Cofilina, 24Cohesina, 36Col�ageno(s), 42adhesion plaquetaria, 140asociados a fibrillas, 42, 43fdel bazo, 183formadores de redes, 42, 44sıntesis, 44, 45ftipo I, 63t, 72tipo II, 63t, 76sıntesis por capilares, 158
tipo III, 63t, 70, 72tipo IV, 46, 47f, 56, 63tsıntesis por capilares, 158
tipo Vease 63tsıntesis por capilares, 158
tipo VII, 63tColangiolos, 258Colchicina, 35cColecistocinina, 242secrecion por duodeno, 259
Coledocolitiasis, 259c
328
INDICEALFA
BETIC
O

Colelitiasis, 259cColera, 249cColıculo seminal, 296Colitis colagenosa, 43cColor del pelo, 214Columnas anales, 248Compartimentohematopoy�etico, 142subosteocl�astico, 80
Complejo(s)afadina-nectina, 54de agrecano, 76, 78de ataque a membrana, 168epıtopo-MHC II, 178hormona-receptor, 188MHC propias-epıtopos propios, 180del poro nuclear, 26, 27fsinapton�emicos, 38tubulovesicular (CTV), 16de union, 52, 53f, 55f, 57f
Complejo TCR, 172Complemento, 168Componentesaferentes del sistema nervioso perif�erico, 108eferentes del sistema nervioso perif�erico, 108motores del sistema nervioso perif�erico, 108sensoriales del sistema nervioso perif�erico, 108
Compuerta(s), 10de inactivacion, 116
COMT. Vease Centros organizadores de micr-otubulos (COMT).
Concreciones prost�aticas, 298Condroblastos, 74, 76Condrocitos, 74, 76, 76cCondroitina sulfato, 66, 69tCondroitina, 4-sulfato, 41tCondroitina, 6-sulfato, 41tCondronectina, 42, 62, 76Conduccioncontinua, 122saltatoria, 122
Conductilloseferentes, 286, 296, 297tterminales, 284
Conducto(s)alveolares, 226, 227fbiliar(es), 246, 258, 259fcomun, 258
coclear, 320deferente, 286, 296, 297tendolinf�atico, 318–320epididimario, 296estriados de gl�andulas salivales, 250extratesticulares, 296, 297f, 297teyaculador, 296, 297tgalactıfero, 284genitales masculinos, 296, 297f, 297thep�aticos, 258intercaladosde gl�andulas salivales, 250pancre�aticos, 252
interlobulares pancre�aticos, 252intratesticulares, 296, 297f, 297tlinf�atico(s), 166derecho, 166
m€ullerianos, 273cnasolagrimal, 316ov�aricos, 276, 278, 279fpancre�atico, 246, 258principal, 252
parameson�efricos, 273cquıstico, 258semicirculares, 318, 320, 321f, 322tor�acico, 166
Conexinas, 56Conexones, 56, 57fConjuntiva, 316bulbar, 316palpebral, 316
Conjuntivitis, 317c
Conos retinianos, 312, 313f, 314, 315fContracciones de propulsion, 246Control del pesoa corto plazo, 71ca largo plazo, 71c
COP I. Vease Coatomero I (COP I).COP II. Vease Coatomero II (COP II).Copa optica, 310Corazon, 162, 164, 163f, 165fesqueleto fibroso, 162
Cordonesespl�enicos, 185hematopoy�eticos, 142medulares de ganglios linf�aticos, 173, 182csexuales primitivos, 272
Corion, 282frondoso, 282liso, 282
Cornea, 306Coroides, 308Coronadel diente, 230de ganglios linf�aticos, 183radiada, 274
Corpusculo(s)gustativos, 236, 237frenal, 260tımicos, 180
Cortesen microscopia optica, 2microscopicos, interpretacion, 4, 5f
Cortezacerebelosa, 130cerebral, 128, 129tsuprarrenal, 200, 201f, 203tde tallo del pelo, 214tımica, 179
Corti, organo, 320, 323fCorticosterona, 200, 203tCorticotropina, 190Cortisol, 200, 203tCost�amero, 96Cowper, gl�andulas, 298COX-2. Vease Enzimas de ciclooxigenasa 2
(COX-2).CPA. Vease C�elula(s) presentadoras de
antıgenos (CPA).CRE. Vease Elementos de respuesta a AMPc.Creatina cinasa, 96, 105cCrecimientoaposicional, 76intersticial, 76
Cresta(s)ampulares, 320, 322neural, 108
Cretinismo, 199cCriptasamigdalinas, 186g�astricas (fov�eolas), 240
Criptorquidia, 287cCrisis de miastenia, 103cCristales, 22de carbonato c�alcico, oıdo interno, 320de hidroxiapatita, 78
Cristalinas, 308a b–cristalina, 96
Cristalino del ojo, 308, 309fligamentos suspensores, 308
Crohn, enfermedad, 249cCrom�atidas hermanas, 36Cromatina, 28–32, 29fasociada al nucl�eolo, 32
Cromatolisis, 130Cromofilos, 190Cromofobos, 190Cromosomas, 28, 29fsexuales, 28X segundos, 138
CSF-1, 90, 92sistema inmunitario innato, 168
CSF-G. Vease Factor(es) estimulador de colo-nias de granulocitos (CSF-G).
CSF-GM. Vease Factor(es) estimulador de col-onias granulocitos-macrofagos(CSF-GM).
CSF-M. Vease Factor(es) estimulador de colo-nias de monocitos (CSF-M).
CTV. Vease Complejo(s) tubulovesicular(CTV).
Cubierta de musculo liso, vesıcula biliar, 258Cuellodel diente, 230uterino, 278
Cuerpo(s)amil�aceos, 298aorticos, 154apoptosicos, 38aren�aceos, 202basales, 22, 52, 53fblanco, 276carotıdeo, 154cavernosos, 300, 303fcelulares de neuronas motoras
posganglionares, 124ciliar, 308densos, 106, 107fer�ectiles del pene, 284, 300, 301fesponjoso, 300uretra, 300
hemorr�agico, 276hialoideo, 310laminares, 226luteo, 276, 277fdel embarazo, 276
medio, 36neuroepiteliales pulmonares, 222polares, primeros, 276residuales, 18tumefactos, 221cvıtreo, 310
Cumulo ooforo, 274Cupula, 320, 322Cushing, sındrome, 201cCutıculadel pelo, 214de unas, 216, 217fde vaina interna de la raız, 214
Dano alveolar, 235cDeciduabasal, 282, 283fcapsular, 282parietal, 282
Decorina, 40Defensinas, 168del intestino delgado, 244, 245c
Deficiencia de adhesion leucocitaria I, 139cDegeneracionmacular, 315ctransneuronal, 130walleriana, 130
Delecion clonal, 170, 180Denticion permanente, 230Dentina, 230, 232radicular, 233
Depositos lipıdicos hepatocitos, 256Dermatano sulfato, 41tDermatoglifos, 204Dermis, 204, 210Descemet, membrana, 306Deshidratacion, microscopia optica, 2Deshidroepiandrosterona, 200, 203tDesmina, 96, 106, 156union, 24
Desmocolina, 54Desmogleına, 54Desmoplaquinas, 54Desmosomas, 52, 53f, 54, 55f, 57fDesoxicorticosterona, 200, 203tDesoxihemoglobina, 134
329INDICEALFA
BETIC
O
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Despolarizacion, 116–118Despolimerizacion, 36Desprendimiento de retina, 311cDetumescencia, 302, 303fa-dextrinasa, 246Diabetesinsıpida, 192cmellitus, 253ctipo, 1, 253ctipo, 2, 253c
Diacinesis, 38Di�afisis, 82Diap�edesis, 136, 138, 158Diarrea, 239caguda, 239ccronica, 239c
Diente(s), 230–234, 231fcambios relacionados con la edad, 231cdeciduos, 230estructuras asociadas, 234, 234fgerminal, 232odontogenia previa a la etapa de campana,
233, 233fpermanentes, 230
Difosfato de adenosina (ADP), plaquetas, 1402,3-difosfoglicerato, 134DiGeorge, sındrome, 181Digestion, 246Dımeros de IgA, 250Dineına, 22, 36Dioxido de carbonoliberacion por eritrocitos, 134suministro, 218transporte por eritrocitos, 134
Dıploe, 82Diploteno, 38Disco(s)intercalados, 104, 105fZ (lınea), 94, 97f
Disfagia, 239cDisfuncion er�ectil, 303cDisplasia fibromuscular, 266cDisposicion helicoidal, 82–84Disse, espacio perisinusoidal, 256Distrofina, 46, 96Distroglucanos, 46DIT. Vease Tirosina diyodada (DIT).Division vestibular del nervio vestibulococlear,
320, 321fDobletes, axonemas, 50Dolicol fosfato, 12Dominio(s)apical de c�elulas epiteliales, 50, 51fbasolateral, 52–56de c�elulas epiteliales, 52–56especializaciones
de la membrana lateral, 52–56, 53f, 55fde la superficie basal, 56, 57f
de hepatocitos, 256de c�elulas epiteliales, 50sinusoidales, 256, 257f
Dopamina, 121tDown, sındrome, 39cDuctus reuniens, 318–320Duramadre, 82, 126, 127fcraneana, 126raquıdea, 126
ECP. Vease Proteına(s) cationica de eosinofilos(ECP).
Ectodermo, 48Edemamecanismo, 70cpermeabilidad capilar, 158
EEE. Vease Epitelio del esmalte externo (EEE).Efectosautocrinos, 58endocrinos, 58paracrinos, 58
Eferencia inhibidora, c�elulas de Purkinje, 130
EGF. Vease Factor(es) de crecimiento epid�er-mico (EGF).
Ehlers-Danlos, sındrome, 63cEIE. Vease Epitelio interno del esmalte (EIE).Ejecutor, 38Elastina, 44, 45f, 62Elastosis solar, 45cElectronesretrodispersos, 7secundarios, 7
Eleidina, 206Elementosformes de la sangre, 132–140, 133fde respuesta a AMPc, 12
Embarazo, cuerpo luteo, 276Embrioblastos, 282Eminencia mediana de la neurohipofisis, 192,
193fEnamelinas, 230Enanismo, 89cEncefalinas, 121tEncefalomiopatıa mitocondrial, 21cEncıas, 234, 234fEndocardio, 162–164Endocitosis, 18, 19fmediada por receptores, 282
Endodermo, 48Endolinfa, 322Endolisosomas, 18, 19fEndometrio, 278implantacion, 282uterino, implantacion, 282
Endometritis, aguda, 279cEndomisio, 94, 95fEndomitosis, 150Endoneuro, 120, 123fb-endorfina, 190Endorfinas, 121tEndosomas, 18, 19fde reciclaje, 18tardıos, 18tempranos, 18
Endostio, 78, 84Endotelina, 140Endotelioarterial, 152corneo, 306
Enfermedadde cambio mınimo, 262chemorr�agica del neonato, 141cpulmonar obstructiva cronica (EPOC),
226cEnlaces cruzados de desmosina, 44Entactina, 42, 46Enterocinasas, 246del intestino delgado, 244
Entrada de laringe, 220Entramado cortical, 320Entrepano oseo del paladar, 235Envolturade clatrina, 118nuclear, 8, 9f, 26–28, 27f
Enzima(s)calcificacion osea, 88de ciclooxigenasa 2 (COX-2), 268convertidora de angiotensina, 268presion arterial, 156sıntesis por capilares, 158
hidrolıticas, 140pancre�aticas, 252sıntesis por capilares, 158
Eosinofilos, 70, 137t, 138Epicardio, 162, 164Epidermis, 204–208, 205tcapas, 206, 207t, 209fcomo defensa, 168no queratinocitos, 208, 209fqueratinocitos, 204, 205f
Epidermolisis ampollosa, 207cEpidıdimo, 286, 296, 297t
Epıfisis, 82Epiglotis, 220Epilepsia, 109cmioclonıa, 21c
Epimisio, 94, 95fEpineuro, 120, 123fEpistaxis, 221cEpiteliocilındrico, 49f, 49tseudoestratificado, 49f, 49t
ciliado de la laringe, 220simple cervicouterino, 278
corneo, 306cubico, 49f, 49tcomo defensa, 168escamoso, 49f, 49testratificado
cavidad bucal, 230no queratinizado de la laringe, 220
del esmalte externo (EEE), 233estratificado, 48, 49tg�astrico, 240germinalov�arico, 272de tubulos seminıferos, 288
interno del esmalte (EIE), 232larıngeocilındrico seudoestratificado ciliado,
220escamoso estratificado no queratinizado,
220no queratinizado escamoso estratificadovaginal, 284
olfatorio, 220, 221fc�elulas
basales, 218globosas, 218sustentaculares, 218
pigmentado, retina, 312, 313fqueratinizado, 49f, 49trespiratorio, 222, 223fseminıfero, 286ciclo, 292–294, 293fgerminal, 288
seudoestratificadocilındrico, 49f, 49testereociliado, epidıdimo, 296
simple, 48, 49tsubcapsular, cristalino, 308de transicion, 49f, 49tde c�alices renales, 270ureteral, 270
de union gingival, 235ureteral, 270
Epıtopos, 18, 136, 171, 175EPOC. Vease Enfermedad pulmonar obstruc-
tiva cronica (EPOC).Eponiquio, 216Eritroblastos, 144basofilos, 150tortocromatofilos, 150tprocromatofilos, 150t
Eritroblastosis fetal, 135cEritrocitos, 132, 134, 144, 150tmembrana celular, 134, 135f, 135ttransporte de dioxido de carbono y oxıgeno,
134, 135tEritropoyesis, 150, 150tEritropoyetina, 148, 149t, 266Eritroqueratodermia variable, 57cEscamas, 206Escherichia coli, infecciones del tracto urinario,
270cEsclerosisglomerular segmentarıa focal asociada a la
heroına, 262chialina central, 43cmultiple, 115c
Esclerotica, 306Escorbuto, 45c, 93c
330
INDICEALFA
BETIC
O

331INDICEALFA
BETIC
OEsferocitosis hereditaria, 135cEsfingomielina, 114Esfınter(es)anal, 248externo, 248
pilorico, 240vaginal, 284
Esmalte, 230, 233cambios relacionados con la edad, 232c
Esofago, 238Espacio(s)epidural, 126extracelular, 114intercrestal, 20, 21fintermembrana, 20, 21fde la matriz, 20, 21fperiaxial, 102periosteocıticos, 79subaracnoideo, 126subdural, 127curinario, 260
Esperm�atidas, 290Espermatocitogenia, 288Espermatocitos, 288primarios, 290secundarios, 290
Espermatogenia, 292, 293fEspermatozoide, 290, 291fcapacitacion, 278cola, 290, 291f
Espermiacion, 288Espermiogenia, 290Espıculas, 82Espina bıfida, 109canterior, 109c
Espliceosomas, 30Espongiocitos, 200Esponjosa, 234Estallido respiratorio, 138Est�ator de sintetasa de ATP, 20Esteatohepatitis, 259cno alcoholica, 259c
Esteatosis hep�atica, 259cEstereocilios, 50, 320, 322Esterilidad masculina, 301cEstigma, c�apsula ov�arica, 276Estimulacion encef�alica profunda, 121cEstomago, 240–242, 241fanatomopatologıa, 242, 243fgl�andulas fundicas, composicion celular,
240–242, 241fEstradiol, 274, 276Estratobasal, 206, 207tcorneo, 206, 207tespinoso, 206, 207tgerminativo, 206, 207tgranuloso, 206, 207tintermedio, 233lucido, 206, 207tvascular, 278
Estrenimiento, 239cEstribo, 318Estrogenosacn�e, 215cgl�andulas mamarias, 284reparacion osea, 91csecrecion por secrecion por sincitiotrofo-
blastos, 282Estroma, 58, 306salival, 250
Estructura(s)funcion, 2oculares accesorias, 316
Etapaaposicional de la odontogenia, 233coronilla de la odontogenia, 232de gemacion de la odontogenia, 232de lactancia, 284de morfodiferenciacion de la odontogenia,
233
Eucromatina, 28Exhalacion, 228Exocitosis, 18continua, 16discontinua, 16
Exones, 30Expansion clonal, 170Exportinas, 28, 29fExterorreceptores, 304Extremoafilado de microfilamentos, 24negativode filamento delgado, 96de microfilamentos, 24de microtubulo, 22
positivode filamento delgado, 96de microfilamentos, 24de microtubulo, 22
Eyaculacion, 302
Factor(es)de acoplamiento a receptores, 66activador de plaquetas, 68, 69tde c�elulas madre, 148, 149tde coagulacion, 140de corte y empalme no RNPnp, 30de crecimientodel endotelio vascular, 88epid�ermico (EGF), 204, 232
humano, 246fibrobl�astico-4 (FGF-4), 232hematopoyesis, 144hematopoy�eticos, 148, 149tinsulinoides, 92tumoral b (TGF-b), 256
determinante de testıculos, 272estimuladorde colonias (CSF), 148, 149t
de granulocitos (CSF-G), 148, 149tde granulocitos-macrofagos (CSF-GM),
148, 149tdemonocitos (CSF-M), 78, 80, 148, 149t
receptores, 80de osteoclastos, 79, 92, 198
inhibidor, meiosis, 272intrınseco g�astrico, 240liberador de prolactina, 190natriur�eticoauricular, 104cerebral, 158
de necrosis tumoral, 90, 92, 138de necrosis tumoral-a (TNF-a), 68cuerpo luteo, 276del intestino delgado, 244origen y funciones, 174t
plaquetario, 3, 140quimiot�acticode eosinofilos, 66, 68, 69tde neutrofilos, 66, 68, 69t
de transcripcion Cbfa1, 84, 88de transcripcion Runx2, 84, 88transformador del crecimiento, 204transformador del crecimiento-b (TGF-b),
78, 90, 92Factor XIII, 140FADH2, 20Fagocitosis, 18por monocitos, 136por neutrofilos, 138
Fagosomas, 18Falopio, trompas, 276, 278, 279fFanconi, sındrome, 269cFaringe bucal, amıgdalas, 186Fasciaadherente, 54oclusiva, 156superficial, 204
Fascıculos, 94, 120Faseacrosomica, espermiogenia, 290
an�agena del crecimiento del pelo, 214de capuchon de espermiogenia, 290cat�agena del crecimiento del pelo, 214espl�enica de la hematopoyesis, 144folicular del ciclo menstrual, 280hep�atica de la hematopoyesis, 144de histodiferenciacion de la odontogenia, 233luteınica del ciclo menstrual,
280, 282de maduracion de la espermiogenia, 290de m�edula osea, hematopoyesis, 144mesobl�astica de la hematopoyesis, 144mieloide de la hematopoyesis, 144proliferativa del ciclo menstrual, 280secretora del ciclo menstrual, 280, 282telogena del crecimiento del pelo, 214
Fase G0, 34Fase G1 (gap), 34, 35fFase G2, 34, 35fFase S (sint�etica), 34, 35fde la meiosis, 38, 39f
Fecundacion, 280, 281fFertilidad, hombre, 287c, 301cFGF-4. Vease Factor(es) de crecimiento
fibrobl�astico-4 (FGF-4).Fibrasamarillas, 44, 45fblancas, 42, 94de col�ageno, 62, 63tdel cristalino, 308el�asticas, 44, 45f, 62intermedias, 94musculares, 94de la bolsa nuclear, 102de la cadena nuclear, 102din�amicas, 102est�aticas, 102extrafusales, 102intrafusales, 102
nerviosasaferentes, 120eferentes, 120est�aticas, 102, 103fdel grupo Ia, 102, 103fdel grupo II, 102, 103fcon terminaciones sensoriales din�amicas,
102, 103fnerviosas Ia, 102, 103freticulares, 44, 70, 106en pulpa roja espl�enica, 185
rojas, 94de tipo A, 122tde tipo B, 122tde tipo C, 122tde la zonula, 308
Fibrilina, 44Fibrina, 140Fibrinogeno, 140Fibroblastos, 64, 64c, 65f, 94activos, 64inactivos, 64
Fibrocartılago, 74, 74c, 75f, 75tFibrodisplasia, 266cFibronectina, 42, 62sıntesis por capilares, 158
Fijacion, microscopia optica, 2Filagrinas, 24Filamentosde anclaje linf�aticos, 166delgados, 22–24, 23f, 25f, 98, 99fdel musculo liso, 106
gruesos, 98, 99fdel musculo liso, 106
intermedios, 22–24, 23f, 25f, 54de queratina, 56
de queratina, 206, 214Filopodios tiroideos, 196Fimbrias, 278Fimbrina, 24, 50, 320Flagelos, 525-fluorouracilo, 35c�
ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Folıculo(s)antrales, 275f, 275tatr�esicos, 276dominante ov�arico, 274linfoides, 185maduros, ov�aricos, 274ov�aricos, 272, 274, 275f, 275tatresia, 274atr�esicos, 276dependientes de FSH, 276primordiales, 272
pilosos, 214primarios, 272, 274, 275tmultilaminares, 274, 275f, 275tov�aricos, 274, 275f, 275tunilaminares, 274, 275t
secundarios, ov�aricos, 275f, 275tFoliculostatina, 274Folistatina, 274Folitropina (FSH), 190efectos fisiologicos, 194tfolıculosdependientes, 276ov�aricos, 274
testıculos, 294Fontanelas, 84Formacionde hueso endocondral, 74, 84–88, 84t, 87f,
89f, 90osea intramembranosa, 74, 84, 85f
Fosfatasaalcalina, osteoblastos, 80cde creatina, 96de miosina, 106
Fosfolipasa A2, 66Fosfolipoproteınas, 20Fosforilacion oxidativa, 20Fovea central, 310Fov�eolas, 240FragmentoFc, 171Fab, 171FSH. Vease Folitropina (FSH).Fuerzade filtracion, 266motriz de protones, 20
Funcioncoclear, 322, 323festructura, 2vestibular del oıdo, 322
Fundus, estomago, 240
GABA. Vease Acido(s) gamma-aminobutırico(GABA).
GAG. Vease Glucosaminoglucanos (GAG).GALT. Vease Tejido(s) linfoide asociado al in-
testino (GALT).Ganglios, 124, 125fautonomos, 124linf�aticos, 166, 168, 181, 182c, 181fraız dorsal, 124sensorial, 124
Gas intestinal, 249cGastrina, 242, 252, 253tsıntesis de �acido clorhıdrico, 242
Gelatinasa, 138Gelsolina, 24Gen SRY, 272Genitales externosfemeninos, 284, 285fmasculinos, 300–302, 301f
Genoma, 28Giemsa, tincion, 3t, 133Gigantismo hipofisario, 89cGlandedel clıtoris, 284, 285fdel pene, 300, 302
Gl�andula(s), 58–60, 188bulbouretrales, 298cardıacas esof�agicas, 238
ceruminosas, 316cervicales, 278duodenales, 246endocrinas, 58, 60, 58–60, 59fsistema neuroendocrino difuso, 60
esof�agicas propiamente dichas, 238exocrinas, 58–60, 59facinares, 60, 61falveolares, 60, 61ffoliculares, 60multicelulares, 58, 60, 61f
compuestas, 60simples, 60
tubulares, 60, 61funicelulares, 58, 59f
extramurales, 250fundicas, composicion celular, 240–242, 241fg�astricas, 240genitales accesorias, masculinas, 298, 299fholocrinas, 212del intestino delgado, 246intramurales, 250lacrimales, 316mamarias, 284, 285fmulticelulares, 58, 60, 61fparatiroideas, 196, 197f, 198, 199f, 203tparotidas, 251perianales, 248de la piel, 212, 213fpineal (cuerpo), 202, 203tsalivalesmayores, 250, 251fmenores, 230mucosas, 234
posteriores, 186seb�aceas, 212, 213fsistema neuroendocrino difuso, 60sublinguales, 251submandibulares, 251sudorıparasapocrinas, 212ecrinas, 212, 213f
suprarrenales, 200, 201ftiroides, 196, 197f, 203ttubularesenroscadas, sencillas, 212espirales simples, 212
tubuloalveolaresexocrinas, 60, 61fgl�andula mamaria como, 284
unicelulares, 58, 59futerinas, 278
Glaucoma, 307cGlıa limitante perivascular, 128Glicerofosfocolina, 296Glicerol, 66Glicoforina A, 134Glisson, c�apsula, 254Globina, 134Globo ocular, 306, 307fGlobulinas, plasma, 133tGlobulos blancos (leucocitos), 136–138, 137t,
144basofilos, 137t, 138eosinofilos, 137t, 138linfocitos, 136, 137tmonocitos, 136, 137tneutrofilos, 137t, 138, 139f
Glom�eruloen bulbo olfatorio, 220renal, 260
Glomerulonefritis por anticuerpos frente amembrana basal glomerular, 47c
Glucagon, 252, 253tGlucina, 42, 62, 121tGlucoc�aliz, 8Glucocorticoides, 203tGlucogeno, 22de hepatocitos, 256
Glucolıpidos, 8
Glucoproteınas, 8, 42, 114adhesivas, 42, 62, 78
Glucosaminoglucanos (GAG), 40–42, 41t, 62,74
Glucuronido de bilirrubina, 258Golgiaparato (complejo), 9f, 14, 16, 17f, 256cis, red (RGC), 16fase de la espermiogenia, 290organos tendinosos, 102, 306, 305ftrans, red (RGT), 16
Golpe de fuerza, contraccion muscular, 94Gonadas indiferenciadas, 272Gonadotropina corionica humana (hCG),
276secrecion por sincitiotrofoblastos, 282
Goodpasture, sındrome, 47cGraaf, folıculos, 274, 275tGranulocito(s), 136Granulocitopoyesis, 150, 151tGranulomeros, 140Gr�anulosalfa, 140auriculares, 104azurofilos, 138de cimogeno, 250delta, 140de envoltura de membrana, 206especıficos, 138gamma, 140de intercromatina, 32lambda, 140laminares, 206de la matriz, 20de pericromatina, 32de secrecion, 9fsecretores, 58terciarios, 138vermiformes, 208
Granzimas, 175Graves, enfermedad, 175c, 199cGrelina, 71c, 240Gruposde fibras principalesgingivales, 234del ligamento periodontal, 234
hemo, 134isogenos, 76
GTP, 12Guillain-Barr�e, sındrome, 115c
H-E. Vease Hematoxilina y eosina (H-E).Habla, 220Hacescontr�actiles, 24paralelos, 24
Haemophilus influenzae, halitosis, 235cHalitosis, 235cHassall, corpusculos, 180Haustras, colon, 248Haversconducto, 82–84, 85fsistemas de canales, 82–84
Haz auriculoventricular, 162hCG. Vease Gonadotropina corionica humana
(hCG).HCO3
�
g�astrico, 242salival, 250
Hebras de fusion, 54Helicotrema, 320Hematocrito, 132Hematopoyesis, 78, 132, 142, 144–150, 145tc�elulas, 144, 145tposnatal, 144prenatal, 144
Hematoxilina y eosina (H-E), 2, 3tHematoxilina-hierro, 3tHemidesmosomas, 52, 53f, 55f, 56, 57fHemoglobina, 134, 135t
332
INDICEALFA
BETIC
O

Hemopoyesis, 78, 132, 142, 144–150, 145tc�elulas, 144, 145tposnatal, 144prenatal, 144
Hemorragianasal, 221csubdural, 127cde tubo digestivo, 239c
Hendidura(s)glotica, 220sin�apticas, 100, 118, 119fprimarias, 100secundarias, 100
Henle, asa, 260ramadelgada, 262
ascendente, 268descendente, 268
gruesaascendente, 262, 268descendente, 262
Henley, capa, 214Heparano sulfato, 41t, 46Heparina, 41t, 66, 69tHepatitis alcoholica, 43cHepatocitos, 254, 256domino basolateral, 256
Hering, canales, 258Hernia de hiato, 241cdeslizante, 241cparaesof�agica, 241c
Herring, cuerpos, 192Hertwig, raız, vaina epitelial (VERH), 233Heterocromatina, 28Heteroinjertos, 91cHialocitos, 310Hidrocefalia, 129c�acido clorhıdrico, 240produccion g�astrica, 242
Hidroxilisina, 42Hidroxiprolina, 42Hıgado, 254–258, 255fbilis, 258conductosbiliares, 258hep�aticos, 258
funciones, 256hepatocitos, 254, 256lobuloscl�asicos, 254conceptos, 254, 255f
regeneracion, 256sinusoides, 256, 257fvesıcula biliar, 258
Himen, 284Hiperparatiroidismo primario, 198cHipertermia, esterilidad masculina, 287cHipervitaminosis A, desarrollo oseo, 93tHipoacusiaconductora, 323cnerviosa, 323cneurosensorial, 323cno sindromica, 57c
Hipocalcemia, sındrome de DiGeorge, 181Hipodermis, 204, 210Hipofisis, 188–192, 189f, 189t, 191tanterior, 188, 189f, 189t, 189, 191fhormonas, efectos fisiologicos, 194tposterior, 188, 189f, 189t, 189, 192, 193f,
195fpresion arterial, 156
Hiponiquio, 216Hipoparatiroidismo, 198cHipot�alamo, 71chormonas liberadoras, 191tnucleos, 192
Hipotiroidismo, 199cHipoxia, luteolisis, 276Hirschsprung, enfermedad, 109cHis, haz, 162
Histamina, 66, 68, 69tpermeabilidad capilar, 158
Histamina, 2, sıntesis de �acido clorhıdrico, 242Histona, 28Histona H1, 28Histoquımica, 4Homoinjertos, 91cHormona(s), 188adrenocorticotropa (ACTH), 190efectos fisiologicos, 194t
antidiur�etica, 192efectos fisiologicos, 194tmovimiento de agua y urea fuera y dentro
de tubulos colectores, 268presion arterial, 156
antim€ulleriana, 288autocrinas, 240clasificacion, 188corticotropa, 190, 278derivadasde �acidos grasos, 188de amino�acidos, 188
endocrinas, 240esteroideas, 188estimuladorade c�elulas intersticiales
efectos fisiologicos, 194tde melanocitos a (MSH-a), 190de la tiroides
efectos fisiologicos, 194tunion, 196
formacion de hueso, 91c, 92, 93tg�astricas, 240, 242gonadotropas, 272hipofisarias, efectos fisiologicos, 194tque influyen en el cartılago hialino, 76, 77tinhibidoras hipofisarias, 190liberadoras hipofisarias, 190lipotropa, 190luteinizante (LH), 190efectos fisiologicos, 194treceptores, 274testıculos, 294
neurosecretoras hipotal�amicas, 190paracrinas, 240paratiroidea, 80, 90, 198, 203tpreproparatiroidea, 198proparatiroidea, 198proteicas, 188union a receptores, 188
Howship, lagunas, 80Huellas dactilares, 204Hueso(s), 78–92alveolar propiamente dicho, 234amorfo, 82articulaciones, 92, 93faspecto macroscopico, 82, 83fcalcificacion, 88c�elulas, 78–80, 79f, 81fcompacto, 82sistemas laminares, 82–84, 85f
concentraciones s�ericas de calcio, 92cortos, 82efectos de hormonas, 92, 93tesponjoso, 82formacionendocondral, 74, 86–88, 86t, 87f, 90intramembranoso, 74, 84, 85f, 88
histogenia, 84–88, 85finmaduro, 82intramembranoso subperiosticoformacion, 88
irregular, 82laminar, 82largos, 82maduro, 82plano, 82primario (inmaduro, amorfo), 82remodelacion, 90reparacion, 90, 91f
resorcion, mecanismo, 80secundarios (maduros, laminares), 82sesamoideos, 82tipos microscopicos, 82–84
Humor acuoso, 308Huntington, corea, 121cHusos musculares, 102, 103f, 306, 305fHuxleycapa, 214teorıa del filamento deslizante, 98
ICAM. Vease Mol�ecula(s) de adhesion intrace-lular (ICAM).
IgA, 230de superficie, 170
Igs. Vease Inmunoglobulinas de superficie(Igs).
IL-1, origen y funciones, 174tImplantacion, 282, 283fImportinas, 28, 29fImpulsos nerviososgeneracion y conduccion, 116–118,
117f, 119fpropagacion, 116–118sinapsis, 118–120, 119f, 119t
Inactivacion de canales de Na+ con compuertade voltaje, 116
Inclusion(es)de c�elulas, 22para microscopia optica, 2
Indian hedgehog, 88Inervacionmotoraautonoma, 122som�atica, 122
del musculo esquel�etico, 100Inestabilidad din�amica, 22Infarto(s)de miocardio, 105crenales, 266c
Infecciones de las vıas urinarias (IVU), 270cInflamacionaguda, 136ccronica, 136c
Infundıbulode neurohipofisis, 192, 193fde trompa de Falopio, 276, 278
Inhalacion, 228Inhibidor de tripsina, 252Inhibina, 274, 288, 294Iniciador, 38Inmunocitoquımica, 4Inmunogenos, 169Inmunoglobulinas, 171, 171f, 172tde superficie (Igs), 171
Insuficiencia renal, 265cInsulina, 71c, 252, 253tIntegrinas, 46, 78, 138, 280implantacion, 282
Integrinas av b3, 80Interfase, 34, 35fInterferon(es)IFN-a, origen y funciones, 174tIFN-b, origen y funciones, 174tIFN-g , origen y funciones, 174tIFN-t, 92, 149tluteolisis, 276sistema inmunitario innato, 168
Interleucina(s)IL-1, 68, 90, 92, 138IL-1a, 204IL-2, 148, 149torigen y funciones, 174t
IL-3, 148, 149tIL-4, origen y funciones, 174tIL-5, 148, 149torigen y funciones, 174t
IL-6, 92, 148, 149torigen y funciones, 174t
IL-7, 148, 149t
333INDICEALFA
BETIC
O
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Interleucina(s) (cont.)IL-8, 139cIL-10, origen y funciones, 174tIL-11, 148, 149tsistema inmunitario innato, 168
IL-12, 148, 149torigen y funciones, 174t
Interneuronas, 112, 126Interorreceptores, 304Intersticiointraperıodo, 114renal, 266
Intestinodelgado, 242–246, 243fanatomopatologıa, 244–246, 245fcaracterısticas histologicas frecuentes, 244histopatologıa, 246, 247f, 247tmotilidad, 246
grueso, 248, 249ffuncion, 248
Intrones, 30Involucrina, 206Ion(es)de calcio, 66hidrogeno g�astrico, 242movimiento, 242potasiog�astrico, 242salival, 250
de sodioconcentracion extracelular, 10salival, 250
Irrigacion de lechos capilares, regulacion, 158,159f
Islasde c�elulas hematopoy�eticas, 142sanguıneas, 144
Islotes hematopoy�eticos, 142Isoenzima MB, creatina cinasa, 105cIstmo, 240c�elulas Ito, 256del oviducto, 278
IVU.Vease Infecciones de las vıas urinarias (IVU).
Kearns-Sayre, sındrome, 21cKerckring, v�alvulas, 244Kiesselbach, �area, 221cKlinefelter, sındrome, 291cKrause, bulbos terminales, 304, 305fKupffer, c�elulas, 256
L-selectinas, 138implantacion, 282
Laberintomembranoso, 318–320, 319foseo, 318, 319f
Labios, 230, 231c, 231fmayores, 284, 285fmenores, 284, 285f
Lactasa, 246Lactoferrina, 230Lactogenos, 284Lagunasdel cemento, 231condroblastos, 76, 77fcondrocitos, 74en decidua basal, 282osteoblastos, 78, 84osteocitos, 82en sincitiotrofoblastos, 282
L�amina(s), 82–84, 85fbasal(es)de capilares, 156de c�apsula de Bowman, 260de c�elulas epiteliales, 46, 56de matriz extracelular, 46, 47fde m�edula osea, 142en pulpa roja espl�enica, 185
densa, 46, 47fdental, 232, 233f
el�asticaexterna arterial, 152interna arterial, 152de la l�amina propia traqueal, 222
espiral osea, 318externade c�elulas de musculo liso, 106de membrana
basal de la matriz extracelular, 46postsin�aptica, 100
intersticiales, 82–84lucida, 46, 47fdel musculo cardıaco, 104nuclear, 26propiade la cavidad nasal, 220g�astrica, 240del intestino
delgado, 244grueso, 248
de la luz del aparato digestivo, 238del oviducto, 278de la tr�aquea, 222vaginal, 284de la vesıcula biliar, 258
reticular, 46, 47fsuced�aneas, 233de la tunica venosa, 161t
L�aminas A, B, y C, 26Laminina, 42, 46, 56, 62adhesion plaquetaria, 140sıntesis por capilares, 158
Langerhansc�elulas, 183, 204, 205f, 208islotes, 188, 252
Lanugo, 214Laringe, 220Laringitis, 221cLazos sin�apticos, 202, 314LCR. Vease Lıquido cefalorraquıdeo (LCR)Leche, formacion, 284Lecho(s) capilar(es), 156irrigacion, regulacion, 158, 159fsecundario, 190
Leiomiomas, 107cLeiomiosarcomas, 107cLengua, 236, 237fcorpusculos gustativos, 236, 237fpapilas linguales, 236raız, 236
Lente(s)condensadoras, 3del objetivo, 3del ocular, 3
Leptina, 71cLeptoteno, 38Leucemia mielogena aguda, 37cLeucocitos, 70, 132, 136–138, 137t, 144. Vease
tambien Globulos blancos (leucocitos).basofilos, 137t, 138eosinofilos, 137t, 138linfocitos, 136, 137tmonocitos, 136, 137tneutrofilos, 137t, 138, 139fpolimorfonucleares, 136, 137t, 138, 139f
Leucotrieno, 70cLeucotrieno C, 4, 68, 69tLeucotrieno D, 4, 68, 69tLeucotrieno E, 4, 68, 69tLey de «todo onada», contraccionmuscular, 98Leydigc�elulas intersticiales, 292, 293fgl�andulas intersticiales, 286
LFA-1. Vease Antıgeno(s) asociado a funcionleucocitaria-1 (LFA-1).
LH. Vease Hormona(s) luteinizante (LH).Lieberk€uhn, cripta, 244, 248Ligamento(s)Periodontal (LPD), 230, 232, 234vocal, 220
suspensores del cristalino, 308Ligando(s), 10, 18de apoptosis, 175, 178de CD40, 174cFas, 178de osteoprotegerina (OPGL), 90, 92
Limbo, 306, 308Linaje de linfocitos T, 180Lınea(s)de cementacion, 82–84densa principal, 114intraperıodo, 114pectinada, 248
Lınea M, 94, 96, 97fLinfa, 166Linfedema, 167cLinfocitos, 70, 136, 137tLinfocitos B, 136, 171, 173tactivados, 182cespl�enicos, 185de memorias, 176, 183, 182cvırgenes, 146en zona marginal espl�enica, 185
Linfocitos NK, 136, 168Linfocitos T, 136, 171–174, 173t, 185citotoxicos, 174naturales, 175
efectores, 172, 174, 174tluteolisis, 276de memoria, 172centrales (LTMC), 172efectores (LTME), 172
reguladores, 170, 175inducibles, 175naturales, 175
reguladores CD, 170, 175vırgenes, 146, 172, 180en zona marginal espl�enica, 185
Linfocitos TH, 174, 174tTH, 170TH2, 170TH17, 170
Linfopoyesis, 150Linfopoyetina estromal tımica, 180Lipasag�astrica, 240de lipoproteınas, 66, 67fsıntesis por capilares, 158
pancre�atica, 66salival, 250sensible a hormonas, 66, 67f
Lıpidos, 22Lipofuscina, 22Lipomas, 71cLipoproteınasde baja densidad plasm�aticas, 133tde densidad muy baja (VLDL)de hepatocitos, 256plasm�aticas, 133t
plasm�aticas, 133tLiposarcomas, 71cbien diferenciados, 71cde c�elulas redondeadas, 71cdesdiferenciados, 71cmixoides, 71c
Lipotropina, 190Lıquidocefalorraquıdeo (LCR), 114, 126, 128, 129textracelular, 2, 72, 132sinovial, 92
Lisosomas, 9f, 18, 19f, 138, 140Lisozimadel intestino delgado, 244, 245cen l�agrimas, 316como p�eptido antimicrobiano, 168produccion salival, 230sıntesis por gl�andulas esof�agicas, 238
Litiasis biliar, 259cLobulillosde gl�andulas exocrinas, 60, 61f
334
INDICEALFA
BETIC
O

pulmonares, 224secretores de gl�andulas mamarias, 284, 285ftesticulares, 286
Lobulo(s)de gl�andulas exocrinas, 60portal hep�atico, 254
LPD. Vease Ligamento(s) periodontal (LPD).LTC, 170, 174, 176LTMC. Vease Linfocitos T de memoria centrales
(LTMC).LTME. Vease Linfocitos T de memoria efectores
(LTME).Lubricina, 92Lunula, 216, 217fLuteına de la granulosa, 276Luteolisis, 276Luzdel aparato digestivo, 238tubo de Schwann, 130
Mac-1. Vease Macrofago-1 (Mac-1).Macrofago(s), 68, 88, 136, 142, 183alveolares, 226c�elulas reticulares estrelladas, 183cuerpo luteo, 276fijos, 68inducidos, 68libres, 68en pulpa roja espl�enica, 185residentes, 68sistema inmunitario innato, 168transeuntes, 68
Macrofago-1 (Mac-1), 139cM�aculaadherente, 52, 53f, 54, 55f, 57fdensa, 262, 264, 268lutea, 310de s�aculo, 318–320de utrıculo, 318–320
MALT. Vease Tejido(s) linfoide asociado a lasmucosas (MALT).
Maltasa, 246Mamas, 284, 285fManguito, 290Manto de ganglios linf�aticos, 183MAP2, 22Marcadores de linfocitos T, 180Marcapasos artificiales, 105cMarfan, sındrome, 63cMartillo, 318Masson, tricoma, 3tMastocitos, 66–68, 67f, 69tactivacion y desgranulacion, 66respuesta inflamatoria, 68, 69tsensibilizacion, 66
Matrizdel esmalte, 233extracelular (MEC), 2, 8, 40–46, 41f,
62, 74fibras, 42–44integrinas y distroglucanos, 46membrana basal, 46, 47fsustancia fundamental, 40–42, 41t
interterritorial, 76nuclear, 32osea, 78, 84de la raız del pelo, 214territorial, 76ungueal, 216
MBP. Vease Proteına b�asica de la mielina;proteına b�asica principal.
Meato auditivo externo, 316MEC. Vease Matriz extracelular (MEC).Mecanismode coagulacion, 158de retroalimentacion, 188vestibular, 318
Mecanorreceptores, 208, 304, 305fencapsulados, 304, 305fno encapsulados, 304, 305f
Mediadorespreformados, 66primarios, 66secundarios, 66
Mediastino testicular, 286M�edulaamarilla, 82, 142osea, 78, 84, 142–150, 143f, 182camarilla, 82, 142hematopoyesis, 142, 144–150, 145trespuesta inmunitaria celular, 170roja, 82, 142
roja, 82, 142suprarrenal, 200, 201f, 203tde la vaina del pelo, 214
Megacarioblastos, 146, 150Megacariocitos, 150Megacolon cong�enito, 109cMeibomio, gl�andulas, 316Meiosis, 38, 290divisionecuatorial, 38, 39f
no separacion, 39creduccional, 38, 39f
Meissnercorpusculos, 304, 305fplexo, 124, 244submucoso, 238
Melanina, 22, 209calteraciones, 23c
Melanocitos, 204, 205fepidermis, 208
Melanoma maligno, 211cMelatonina, 202, 202c, 203tMembrana(s)anticuerpos, 170basalde matriz extracelular, 46, 47fde paredes de vasos sanguıneos, 152de piel, 204
basilar, 320celular, 8, 9fcara E y cara P, 8, 9fde eritrocitos, 134, 135f, 135thojuela
externa, 8interna, 8
modelo de mosaico fluido, 8, 9fpolarizacion, 116
el�asticas, 156externa de la mitocondria, 20, 21ffenestradas, 156granulosa, 274, 276limitanteexterna de la retina, 314interna de la retina, 314
nuclear(es), 26, 27fexterna, 26, 27finterna, 26, 27f
otolıtica, 320piamadre-glial, 112plasm�atica. Vease Membrana(s) celular.postsin�apticas, 100, 118, 119fpresin�apticas, 100, 118, 119ftectorial, 320timp�anica, 316equilibrio de presion, 317c
vestibular, 320vıtrea, 214
M�eni�ere, enfermedad, 323cMeninges, 126, 127fMeningiomas, 127cMeningitis, 127cMenopausia, 272Menstruacion, 276, 280Merkelc�elulas, 204, 205f, 208asociaciones de neuritas, 208
discos, 304, 305fMeromiosina ligera, 106
Mesaxonexterno, 114interno, 114
Mes�enquima, 62Mesodermo, 48, 62Mesoovario, 272, 273fMetaarteriolas, 158Metacromasia, 66Metafase, 36, 37f, 276Metafase I de la meiosis, 38, 39fMet�afisis, 82Metaplasia, 57cdel epitelio respiratorio, 222cescamosa, 57c
M�etododirecto de inmunocitoquımica, 4, 5findirecto de inmunocitoquımica, 4, 5f
Metotrexato, 35cMiastenia grave, 103cMicelas, 246Microfibrillas, 44Microfilamentos, 9f, 24, 25fMicroglıa, 131cMicroscopiaconfocal, 5, 7felectronica, 5, 7fde transmision, 3f, 7f
optica, 2–5m�etodos visuales avanzados, 4, 5finterpretacion of cortes microscopicos,
3, 5fpreparacion del tejido, 2–5, 3f, 3t
Microscopiocompuesto, 2, 3felectronicode barrido, 3fde transmision, 3f
optico, 3fMicrotubulos, 9f, 22–24, 23fdel huso mitotico, 36polares, 36solitarios en axonema, 50
Microvellosidades, 9f, 50, 51fMielinizacion, 114Mieloblastos, 146, 150, 151tMielocitos, 150, 151tbasofilos, 150, 151teosinofilos, 150, 151tneutrofilos, 150, 151t
Mielofibrosis, 147cMioblastos, 94Miocardio, 162–164Miofibrillas, 94, 97forganizacion estructural, 96–98, 97f
Miofibroblastos, 64, 233, 256Miofilamentos, 94, 104, 106, 140delgados, 96gruesos, 96
Mioglobina, 94, 96, 104Miomesina, 96Miometrio, 278Miopatıasinflamatorias, 95cmitocondriales, 21c
Miosina, 140Miosina II, 96, 98, 106Miositis, 95ctemporal, 95c
Miotubos, 94MIT. Vease Tirosina monoyodada (MIT).Mitocondrias, 9f, 20, 21fde hepatocitos, 256membrana externa, 20, 21f
Mitosis, 34, 35f, 36, 37fMixedema, 199cMLCK. VeaseCinasa(s) de la cadena ligera de la
miosina (MLCK).Modalidad aferente visceral general, 304Modelode cartılago hialino, formacion osea, 86
335INDICEALFA
BETIC
O
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Modelo (cont.)modificado del mosaico fluido, 8del mosaico fluido de la membrana celular,
8, 9fModıolo, 318Mol�ecula(s)de adhesion intracelular (ICAM)ICAM-1, 138, 139cICAM-2, 138, 139c
de ARN, 30del complejo principal de histocompatibili-
dad, 172, 175carga, 175MHC I, 174c
hidrofobas no polares, 10odorıferas, 220polares, 10receptoras de olores, 220de senalizacion, 10liberacion por monocitos, 136
similar a la heparina, 140de tropomiosina, 98
Mol�eculas B7, 178Mol�eculas CD, 172Mol�eculas CD4, SIDA, 176cMol�eculas CD95L, 178Mollespacio, 254gl�andulas, 316
Monocitos, 68, 70, 136, 137ten tabiques alveolares, 226
Monocitopoyesis, 150Monofosfato de adenosina (AMP), 66cıclico (AMPc), 12, 66, 188
Monoglic�eridos, intestino delgado, 246Moscas volantes, 311cMotoneuronas, 112motoneuronas a, 100motoneuronas t, 102est�aticas, 102
preganglionares, 124Movimientosde la cabeza, funcion vestibular, 322circulares de la cabeza, funcion vestibular, 322lineares de la cabeza, funcion vestibular, 322
MPZ. Vease Proteına(s) cero de mielina (MPZ).MSH-a. Vease Hormona(s) estimuladora de
melanocitos a (MSH-a).Mucina, 222Mucinogenos, 222, 223f, 250del intestino delgado, 244
Mucosa(s)del aparato digestivo, 238bucal, 230como defensa, 168esof�agica, 238especializada, 230de la cavidad bucal, 230
del intestino delgado, 244, 245fmasticatoria, 230del oviducto, 278del paladar blando, 235de revestimiento, 230de la cavidad bucal, 230
ureteral, 270vaginal, 284
Mucosidadde c�elulas caliciformes, 58esof�agica, 250del intestino delgado, 244traqueal, 222
Muerte celular programada, 38, 170, 175, 178,180, 182c
M€uller, c�elulas, 314Musculo(s), 94–106, 95fbulboesponjoso, 300cardıaco, 94, 95f, 105f, 162–164c�elulas, 104
ciliar, 308dilatador de la pupila, 308
erectores del pelo, 214del esfınter de la pupila, 308esquel�etico, 94–102, 95fcontraccion, 98inervacion, 100microscopia
electronica, 96, 97foptica, 94
miofibrilar organizacion, mutaciones, 99corganizacion estructural de las miofibrillas,
96–98, 97fdel paladar blando, 235relajacion, 100sistema sensorial, 102
estriado, 94del estribo, 318de lengua, 236liso, 94, 106contraccion, 106del epidıdimo, 296microscopia
electronica, 106optica, 106, 107f
multiunitario, 106de unidad unica vascular, 106unitario, 106vascular, 106
tensor del tımpano, 318traqueal, 222vocal, 220
Mutacionesorganizacion miofibrilar del musculo esqu-
el�etico, 99coxidasa de NADPH, 139c
NADH, 20Narinas, 218Nebulina, 96Necrosis tubular aguda, 265cNefritis hereditaria, 47cNefronas corticales, arteriolas glomerulares e-
ferentes, 266, 267fNervio(s)motores craneales, 122optico, 310raquıdeos motores, 122vago, 259
Neumocitos, 226de tipo I, 226de tipo II, 226, 227f
Neuroepitelio, 108Neuroglia, 108Neurohipofisis, 189, 189f, 189t, 189, 192,
193f, 195fNeurohormonas, 120Neuromoduladores, 120Neuronas, 108, 110aferentes, 112bipolares, 112, 113f, 314eferentes, 112estructura y funcion, 110, 111fmotoraseferentes, 112preganglionares, 124
multipolares, 112, 113fparasimp�aticas, 124posganglionares, 108, 122preganglionares, 108, 122sensoriales (aferentes), 112simp�aticas, 124unipolares, 112, 113f, 124yuxtamedulares, 268arteriolas glomerular eferentes, 266
Neuropilo, 126Neurotransmisores, 118, 120, 121tNeurotrofinas, 131cNeutrofilos, 70, 137t, 138, 139fNevos, 209cNidogeno, 42Nidos de cristalizacion, 88
NO. Vease Oxido nıtrico (NO).Nocirreceptores, 306Nodulo(s)auriculoventricular, 162linfoides, 182cde amıgdalas, 186primarios, 183, 182cen pulpa blanca espl�enica, 184secundarios, 183, 182c
sinoauricular (SA), 162Nodulo SA. VeaseNodulo(s) sinoauricular (SA).Noradrenalina, 66, 121t, 124, 200, 203tvasos sanguıneos, 154
Notch-1, receptores, 180Nucleo, 8, 26–38bilobulado, 138cromatina, 28–32, 29fenvoltura nuclear, 26–28, 27fparaventricular del hipot�alamo, 192supraoptico del hipot�alamo, 192
Nucl�eolo, 9f, 32, 33fNucleoplasma, 26, 32, 33f
Obesidaden adultos, 71chipercelular (hiperpl�asica), 71chiperpl�asica, 71chipertrofica, 71c
Ocludinas, 54Oddi, esfınter, 258Odontoblastos, 231, 233, 233fOdontoclastos, 231Odontogenia, 232, 233fOıdo, 316–322, 317fexterno, 316, 317ffunciones, 322interno, 318–320, 319f, 321f, 323fmedio, 318, 319f
Ojos, 306–316, 307fcristalino, 308, 309fcuerpo vıtreo, 310estructuras accesorias, 316retina, 310–314, 311ftunica vasculosa, 308, 309f
Oligodendrocitos, 112interfascicular, 112sat�elite, 112
Oligosacaridasas, 246del intestino delgado, 244
Oncogenes, 37cOnda(s)de despolarizacion, 116–118perist�alticas, 246
Onicomicosis, 217cOPG. Vease Osteoprotegerina (OPG).OPGL. Vease Ligando(s) de osteoprotegerina
(OPGL).Opistotonos, 101cOra serrata, 310Organizadores del procentrıolo, 52Organo del esmalte, 232Organos, 8efectores, 108, 122fotosensoriales. Vease Ojos.linf�aticoscentrales, 170, 179perif�ericos, 170, 179primarios, 170, 179secundarios, 182c
linfoides, 178–185primarios (centrales), 170, 179secundarios (perif�ericos), 170, 179
Org�anulos, 8, 9fOrina, formacion, mecanismo, 266–268, 269fOrquitis, parotiditis, 291cOsıculos, 318Osteoblastos, 78, 79f, 84formacion, 78fosfatasa alcalina, 80c
Osteocalcina, 78
336
INDICEALFA
BETIC
O

Osteocitos, 78, 79, 79f, 82, 84Osteoclastos, 68, 79, 79f, 80Osteoclastogenia, 80Osteoide, 78Osteomalacia, 93cOsteonas, 82–84Osteonectina, 42, 62, 88Osteopetrosis, 81cOsteopontina, 42, 78Osteoporosis, 91cOsteoprotegerina (OPG), 80, 90, 92Osterix, 84Otitis media, 323cOtoconia, 320Otolitos, 320Ovarios, 272–276, 273ffolıculos, 272, 274, 275f, 275t
Ovocitosfecundacion, 280primarios, 272secundarios, 276
Ovulacion, 276, 277fOxidasa de NADPH, carencia hereditaria, 139cOxido nıtrico (NO), 140ereccion del pene, 302intercambio gaseoso, 228presion arterial, 154
Oxıgenoliberacion por eritrocitos, 134transporte por eritrocitos, 134
Oxihemoglobina, 134Oxitocina, 192, 278efectos fisiologicos, 194t
Pacini, corpusculos, 210, 304, 305fPacoglobinas, 54Paladar, 235blando, 235duro, 235
P�ancreas, 252, 253fendocrino, 252, 253texocrino, 252
Paneth, c�elulas, 244Panıculo adiposo, 210Papanicolaou, frotis, 279ct�ecnica, 279c
Papila(s)circunvaladas, 236dental, 232d�ermicas, 214duodenal, 246, 258filiformes, 236foliadas, 236fungiformes, 236linguales, 236
Paquiteno, 38Paredes ungueales, 216Par�enquima, 58Parkinson, enfermedad, 121cParotiditis, 251corquitis, 291c
P�arpados, 316Partıcula(s)beta de hepatocitos, 256de reconocimiento de senales (PRS), 14, 15fde ribonucleoproteınas nuclearesheterog�eneas (RNPnh), 30pequenas (RNPnp), 30
Pecas, 209cPelo(s), 214, 215fcolor, 214crecimiento, 214gustativos, 236musculos erectores del pelo, 214terminales, 214
Pelvis renal, 260, 270Pene, 300–302, 301f, 303fP�enfigo vulgar, 55cPepsina, 240Pepsinogeno, 238, 240
Peptidasa(s)citopl�asmicas, 246de procol�ageno, 44de senales, 12
P�eptido(s)antimicrobianos, 168inhibitorio g�astrico, 242intestinal vasoactivo, 253tsenal, 14
P�eptido YY, 71cPerforinas, 175Pericardioparietal, 164visceral, 164
Pericarditis, 163cPericitosde tejido conjuntivo, 64, 65fde vasos sanguıneos, 152, 156
Pericondrio, 74, 76Pericr�aneo, 82Perilinfa, 320Perimisio, 94, 95fPerineuro, 120, 123fPerıodo resistente, 116–118Periostio, 78, 82, 83f, 84Peristaltismo, 238Peritoneo, 254Perlecano, 46Peroxidasa tiroidea, 196Peroxido de hidrogeno, 138Peroxisomas, 18de hepatocitos, 256
Peyer, placas, 186, 187f, 246Piamadre, 126, 127f, 202Piel, 204–210, 205fdelgada, 204, 205f, 205t, 206dermis, 204epidermis, 204–208, 205tde «gallina», 214gl�andulas, 212, 213fgruesa, 204, 205f, 205t, 206, 207t, 209ftrastornos, 209c, 211c
Piezade conexion, 290terminal del espermatozoide, 290
Pigmentos, 22de la bilis, 258
Pinealocitos, 202Pinocitosis, 18Pir�amides renales, 260Pirimidina, 30Piruvato, 20Pituicitos, 192Placa(s)de anclaje, 56corionica, 282cortical de hueso alveolar, 234epifisaria, 82, 89fmetaf�asica, 36neural, 108psori�asicas, 207cdel tarso, 316terminales motoras, 100
Placenta, desarrollo, 282, 282tPlaquetas, 140, 141f, 141tPlaquinas, 24Plasma, sangre, 132, 133tPlasmalema. Vease Membrana(s) celular.Plasmalogeno, 18Plasmina, 140, 276Plasminogeno, 140Plasticidad neuronal, 131cPlectina, 24, 96Pleuraparietal, 228visceral, 228
Plexocapilarperibiliar, 254primario, 190
capsular, 266coroideo, 114, 128mient�erico, 238subcapsular suprarrenal, 200venosohemorroidal, 248pampiniforme, 286
Pliegue(s)circulares, 244marginal de los capilares, 156de la membrana plasm�atica basal, 56ungueal proximal, 216de union, 100vestibulares de la laringe, 220vocales, 220
Podocitos, 260, 261fPolarizacion de membranas celulares, 116Poli, 136, 137t, 138, 139fPolip�eptidonatriur�etico auricular, 162pancre�atico, 252, 253t
Polisomas, 14Porcionconductora del aparato respiratorio,
218–224, 219t, 221fcontorneada, 262distal, 190, 191fF0 de sintetasa de ATP, 20F1 de sintetasa de ATP, 20fibrosa, 32granulosa, 32intermedia, 190, 191fnerviosa de la neurohipofisis, 191t, 192,
193f, 194trecta, 262respiratoria del aparato respiratorio, 218,
226–228, 227fsecretoramerocrina de gl�andulas sudorıparas
ecrinas, 212tuberosa, 190, 191f
Porinas, 20Porosde capilares, 156grandes, 158de los capilares, 158
gustativos, 236nucleares, 26, 27ffuncion, 28, 29f
pequenos, 158de capilares, 158
en retıculo endopl�asmico rugoso, 14Porosomas, 16Porta hep�atica, 254Potencial(es)de accion, 116–118, 117f, 119folfato, 220
de membrana, 116postsin�apticosexcitatorios, 118inhibidores, 118
de reposo, 116, 117fPreameloblastos, 233Precursores de linfocitos T, 180Preosteoclastos, 79Preparacion del tejido, 2–5, 3f, 3tPrepucio, 300Presbicia, 309cPresion(es)arterial, regulacion, 154–156osmoticas g�astricas, 242
Primer cuerpo polar, 276Procaspasas, 38Procol�ageno, 44, 45fProenzimas, 252Proeritroblastos, 146, 150, 150tProfase, 36, 37fProfase I, meiosis, 38, 39fProgesterona, 274, 276desarrollo de la gl�andula mamaria, 284secrecion por sincitiotrofoblastos, 282
337INDICEALFA
BETIC
O
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Prolactina, 190efectos fisiologicos, 194tsıntesis por c�elulas deciduales, 282
Prolina, 62Prometafase, 36, 37fPromielocitos, 150, 151tPromonocitos, 146Proopiomelanocortina, 190Propagacion retrograda, 116–118Propionibacterium acnes, 215cProprioceptores, 304Prostaciclinas, 140sıntesis por capilares, 158
Prostaglandinas, 278g�astricas, 242prostaglandina D2, 68, 69tprostaglandina E2, 268sıntesis por c�elulas deciduales, 282
Prostata, 298, 299fProteasas neutras, 66, 69tProteasomas, 18, 175Proteına(s)acıdica fibrilar glial, 112adaptadora activadora de procaspasas apop-
tosicas (Apaf1), 39de anclaje, 14, 15fbanda, 3, 134b�asicade la mielina, 114principal, 138
de canal, 10cationica de eosinofilos (ECP), 138cero de mielina (MPZ), 114cinasas, 34de coagulacion, 133tdel complemento, 133tde coronamiento, 24de desacoplamiento-1 (UPC-1), 73cdiana, 38endogenas, 175de envoltura de vesıculas AP-2, 118exogenas, 175del grupo de diferenciacion (mol�eculas CD),
172inhibidora de macrofagos-a, 148integrales, 8invariante asociada a clase II (CLIP), 176ligadoras transmembrana, 54, 56morfogen�eticas de hueso, 78, 232N-glucosiladas, 14perif�ericas, 8en plasma, 133tplasm�aticas, union de c�elula folicular, 196plegadas incorrectamente, 14polipeptıdicas, 188poliubiquinada, 18del receptor tiroideo nuclear, 196receptoras de cargo, 18relacionada con PTH, 88en ribosomas, 12secretorasvıa
constitutiva, 16regulada, 16
similares al col�ageno, 42soluble de union a la proteına de fusion
sensible a N-etilmaleimida-25, 118translocadoras, 12, 14transmembrana, 8transportadoras, 10, 176de transporte de membrana, 10, 11fde uniona androgenos (ABP), 288, 294a filamentos intermedios, 24
de union a CRE, 12Proteına C, 96Proteına Fas, 178Proteına G, 12, 188Proteına GS, 12Proteına SNARE, 16
Proteoglicanos, 40, 41f, 62, 74, 88, 100Protofilamentos, 22Protooncogenes, 34, 37cProtoplasma, 8Protrombina, 140Protuberancia del esmalte, 232Proyeccion(es)ciliares, 308neurogliales, 126osteocıticas, 80, 82radial en axonemas, 50
PRS. Vease Partıcula(s) de reconocimiento desenales (PRS).
Puente de nexina, axonemas, 50Pulpablanca del bazo, 183del diente, 230, 232radicular, 232
radicular, 233rojadel bazo, 185espl�enica, 185
Punto(s)ciego, 310de control, 34de metafase/anafase, 34
de control G2/M, 34de inicio/restriccion, 34lacrimal, 316
Pupila del ojo, 308Purinas, 30Purkinjecapa de c�elulas de la corteza cerebelosa, 130c�elulas, eferencia inhibidora, 130fibras, 162
Purpura visual, 312Pus, 138
Quanta, 100Queratano sulfato, 41tQueratinocitos, 204, 205fausencia, epidermis, 208, 209f
Queratohialina, 206Quiasmas, 38Quilıferos, 66del intestinodelgado, 244, 245fgrueso, 244
Quilo, 246Quilomicrones, 66, 246en plasma, 133t
Quimiocinas, sistema inmunitario innato, 168Quimioterapia, 115c, 292cciclo celular, 35c
Quimo, 240
Rab, 16Rab3A, 118Radioterapia, 115cRaızde la lengua, 236del pelo, 214ungueal, 216, 217f
Rampatimp�anica, 320vestibular, 320
RANK, 80RANKL, 78, 80Ranvier, nodulo, 114, 115fRappaport, �acinos, 254Raquitismo, 93cRaschkow, plexo, 230Raynaud, fenomeno, 163cRayos ultravioletas (UV), 209cRayos UV. Vease Rayos ultravioleta (UV).RE. Vease Retıculo endopl�asmico (RE).Reaccionacrosomica, 280anafil�actica, 66axonal, 130, 131f
de hipersensibilidad inmediata, 66Rebordesd�ermicos (papilas), 204, 210epid�ermicos (papilas), 204gonadales, 272
Receptor(es), 108activadores de linfocitos T citotoxicos, 168de aldosterona, tubulo distal, 264de apoptosis, 38, 175, 178de calcitonina, 80del complemento, 138del factor estimulador de osteoclastos-1, 80de hormona paratiroidea, 78inhibidores de linfocitos T citotoxicos, 168intracelulares, 188ligados a proteınas G, 12, 13fde linfocitos T (TCR), 170, 172de partıculas de reconocimiento de senales,
12, 14, 15fde prelinfocitos T, 180de rianodina, 96sensibles a dihidropiridina (RSDH), 96de superficie celular, 188tipo Toll, 168, 170tde transferrina, 127c
Receptores CSF-M, 80Receptores Fc, 138Receptores PRS, 12, 14, 15fReceptores RANK, 80Recto, 248Red(es)similares a gel, 24testicular, 286, 296, 297ttrabecular, 306
Reflejo(s)arcos reflejos sencillos, 103cautonomos, comparacion con reflejos som-
�aticos, 109fde distension, 102de estiramiento, 102rotuliano, 103cde sacudida mandibular, 235csom�aticosy autonomos, 109fcomparados con reflejos autonomos, 109f
tusıgeno, 221cRegeneracion nerviosa, 130, 131freaccion axonal, 130
Region(es)del cardias del estomago, 240externa, 138fundica del estomago, 240interplaca de la vejiga urinaria, 270intramural del oviducto, 278olfatoria de la cavidad nasal, 218organizadoras nucleolares, 36pilorica del estomago, 240de la placa de la vejiga urinaria, 270porta insuloacinar, 252
Regurgitacion, 239cRehidratacion, microscopia optica, 2Reinke, cristales, 292REL. Vease Retıculo endopl�asmico liso (REL).Remodelacion de superficie, 90Renina, 104, 240, 268Repliegues de amıgdalas, 186Repolarizacion, 116–118RER. Vease Retıculo endopl�asmico rugoso
(RER).Rescate, 22Resolucion de microscopios, 5Respiracion, 218celular, 228, 229fexterna, 218interna, 218mecanismo, 228
Respuesta(s)anamn�esica, 170inmunecelular, 136, 170
338
INDICEALFA
BETIC
O

humoral, 136, 170primaria, 170secundaria, 170TH1, 178, 177fTH2, 176, 177f
inmunitariacelular, 136, 170
TH1, 178, 177fTH2, 176, 177f
humoral, 136, 171mediada por
linfocitos TH1, 176, 177flinfocitos TH2, 176, 177f
primaria, 170secundaria, 170
inflamatorias, 138inhibidoras, 188
Rete, aparato, 204Retıculodel co�agulo, 140endopl�asmico (RE), 12liso (REL), 9f, 12
de hepatocitos, 256rugoso (RER), 9f, 12
de hepatocitos, 256de transicion, 16, 17f
estrellado, 232sarcopl�asmico, 94, 104
Reticulocitos, 150tRetina, 310–314, 311fcapas, 312–314, 313f, 315fdesprendimiento, 311cporcion ciliada, 308propiamente dicha, 310
Retroalimentacion positiva, 188RGC. Vease Golgi cis, red (RGC).RGT. Vease Golgi trans, red (RGT).Riboforinas, 12Ribonucleoproteına mensajera (RNPm), 30Ribosomas, 12Ribozimas, 12Rigidez cadav�erica, 99cRinitis al�ergica, 70cRinones, 260, 261flobulacion, 260cpresion arterial, 156
RNPm. Vease Ribonucleoproteına mensajera(RNPm).
RNPnh. Vease Partıcula(s) de ribonucleopro-teınas nucleares heterog�eneas (RNPnh).
RNPnp. Vease Partıcula(s) de ribonucleopro-teınas nucleares pequenas (RNPnp).
Rodopsina, 312Romanovsky, tincion, 133Rotor de la sintetasa de ATP, 20RSDH. Vease Receptor(es) sensibles a dihidro-
piridina (RSDH).Ruffinicorpusculos, 210terminaciones, 304, 305f
Ruminacion, 239c
Sacarasa, 246Saco(s)alveolares, 226dental, 232endolinf�atico, 318–320lacrimal, 316
S�aculo, 318Sales biliares (�acidos), 258Saliva, 230flujo, 251cmezclada, 250mixta, 251primaria, 250secundaria, 250
Salivon, 250Salpingitis, 279c
Sangre, 132–140coagulacion, 132elementos formes, 132–140, 133fplasma, 132, 133t
Sarcolema, 94Sarcomeros, 94, 97fSarcosomas, 94Schlemm, canal, 306, 308Schmidt-Lanterman, incisuras, 114Schwannc�elulas, 100, 114, 115ftubo, 130luz, 130
Sebo, 212Secrecion(es)citocrina, 208glandular(es)holocrina, 58, 59fmerocrina, 58, 59fmixtas, 58mucosas, 58serosas, 58
de gl�andulas apocrinas, 58, 59fSecretina, 240Segmentacion, 280Segmento(s)broncopulmonares de los pulmones, 224externos de la retina, 312interganglionar, 114interno de la retina, 312
Segundos mensajeros, 188Seno(s)anales, 248carotıdeo, 154galactıfero, 284marginales espl�enicos, 185medulares de ganglios linf�aticos, 183paratrabeculares de ganglios linf�aticos, 183subcapsular de ganglios linf�aticos, 183
Senalescoestimuladoras, 176de exportacion nuclear, 28de localizacion nuclear, 28
Senalizacion celular, 10, 12Separacion, ausencia, 39cSerosadel aparato digestivo, 238del ap�endice, 248del intestinodelgado, 244grueso, 248
esof�agica, 238semilunares, 250uterina, 278
Serotonina, 121tSertoli, c�elulas, 288, 289fSharpey, fibras, 82, 83f, 234Sialoproteınas oseas, 78, 88SIDA, mol�eculas CD4, 176cSildenafilo, 303cSinapsina-I, 118Sinapsina-II, 118Sinapsis, 116, 118–120, 119f, 119tel�ectrica, 118quımica, 118
Sinaptobrevina, 118Sinaptofisina, 118Sinaptotagmina, 118Sincitiotrofoblastos, 282Sincondrosis, 92Sindesmosis, 92Sındromecarcinoide, 60cde hiper-IgM, 174cde inmunodeficiencia adquirida, mol�eculas
CD4, 176cSinemina, 24Sinostosis, 92
Sintaxina, 118Sıntesis de proteınas, 12, 13faparato de Golgi, 16, 17fde proteınasempaquetadas, 14, 15fno empaquetadas, 14, 15f
Sintetasade acetil-CoA, intestino delgado, 246de ATP, 20de oxido nıtrico, 268
Sinusoides, 156hep�aticos, 254, 255f, 256marginales espl�enicos, 185en m�edula osea, 142en pulpa roja espl�enica, 185
Sistema(s)de la ciclasa de adenilato, 66endocrino, 188–202. Veanse tambien
Gl�andula(s); hormona(s)fagocıtico-mononuclear, 68, 80, 136inmunitario, 168–186acronimos asociados, 170tadaptativo (adquirido), 169
c�elulas, 168, 170t, 172–178humoral, 172innato (natural), 168, 170t
c�elulas, 168, 170t, 172–178organos linfoides, 178–186
bazo, 168, 183–185, 184fganglios linf�aticos, 168, 181, 181ftejido linfoide asociado a mucosas, 185timo, 168, 179, 179f
seleccion y expansion clonal, 170de intercambio contracorriente, 268laminarcircular, 82–84perimetral interno, 82, 84
linf�atico, 166, 167fcapilares y vasos, 166, 167fdifuso, 168
multiplicador contracorriente, 268nervioso, 108–130autonomo, 108, 122, 123fc�elulas, 110–114central (SNC), 108, 126–130
barrera hematoencef�alica, 128corteza
cerebelosa, 130cerebral, 128, 129t
meninges, 126, 127fplexo coroideo, 128
desarrollo, 108ent�erico, 238ganglios, 124, 125fimpulso nervioso generacion y conduc-
cion, 116–118, 117f, 119fsinapsis, 118–120, 119f, 119t
motor som�atico, 122, 123fparasimp�atico, 124, 125f
ereccion del pene, 302presion arterial, 154sistema nervioso ent�erico, 238
perif�erico (SNP), 108, 120–122clasificacion funcional de los nervios, 120revestimientos de tejido conjuntivo,
120, 123fvelocidad de conduccion, 122, 122t
propagacion del impulso nervioso, 116,118
regeneracion nerviosa, 130, 131freaccion axonal, 130
simp�atico, 124, 125fent�erico sistema nervioso, 238eyaculacion, 302presion arterial, 154
som�atico, 108, 122–124, 123f, 125fmotor, 122, 123f
org�anicos, 8
339INDICEALFA
BETIC
O
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Sistema(s) (cont.)de segundos mensajeros, 10, 240tegumentario, 204–216. Veanse tambien
Pelo(s); Piel; Unas.tubularde apertura en superficie, 140denso, 140
tubulovesicular, 240Sitio A, 12, 14Sitio E, 12formacion, 32, 33fde hepatocitos, 256
Sitio P, 12, 14, 15fSitios activos, 118de mol�ecula de actina G, 98de las vesıculas presin�apticas, 100, 101f
Sj€ogren, sındrome, 59cSNAP-25 (proteına soluble de union a la
proteına de fusion sensible aN-etilmaleimida-25), 118
SNC. Vease Sistema(s) nervioso central (SNC).SNP. Vease Sistema(s) nervioso perif�erico
(SNP).Somatomamotropina corionica, secrecion por
sincitiotrofoblastos, 282Somatomedinas, 92Somatostatina, 242, 253tSomatotrofos, 190Somatotropina, 92, 190efectos fisiologicos, 194t
Sonic hedgehog, 232Soriasis, 207cSox9, 76Staphylococcus aureus, endometritis, 279cSteel, factor, 148, 149tStreptococcus, endometritis, 279cSubmucosadel aparato digestivo, 238del ap�endice, 248esof�agica, 238g�astrica, 240del intestinodelgado, 244grueso, 248
de la l�amina propia traqueal, 222Subunidad Aen axonema, 50del cilio, 52
Subunidad Ben axonema, 50del cilio, 52
Subunidad C, cilio, 52Sujetos hiperal�ergicos, 70cSulfatasa de arilo, 69tSulfato de arilo, 66Suministro de oxıgeno, 218Superoxidos, 138Surco(s), 128neural, 108de segmentacion, 36terminal, 236ungueal, 216
Surfactante pulmonar, 226Sustanciablanca, 126fundamental, 40–42, 41t, 70, 72gris, 126inductora, meiosis, 276
T3. Vease Triyodotironina.T4. Vease Tiroxina.Tabique(s)de gl�andulas exocrinas, 60interalveolares, 226, 227fnasal, 218
Tablaexterna del hueso compacto, 82interna del hueso compacto, 82
Talinas, 46Tallode c�elulas caliciformes, 58, 59fconector de la retina, 312optico, 310de sintetasa de ATP, 20
TATA, caja, 273cTau, 22TCR. Vease Receptor(es) de linfocitos T (TCR).Teca, 222, 223fde c�elulas caliciformes, 58, 59fexterna, 274, 275f, 275t
Tejido(s), 8adiposo, 72, 73fblanco (unilocular), 72, 73fganglios linf�aticos, 181pardo (multilocular), 72, 73c
conjuntivo, 62–72areolar, 72c�elulas, 62, 64–70, 64t
fijas, 64–68, 65ftransitorias, 64, 68–70
clasificacion, 70–72, 71tcolagenoso
irregular denso, 72regular denso, 72
denso, 72el�astico regular denso, 72embrionario, 70funciones, 62laxo, 72mesenquimatoso, 70mucoide, 70propiamente dicho, 62, 72
composicion, 62, 63tdenso, 72laxo (areolar), 72reticular, 72tejido adiposo, 72
reticular, 72subendotelial, 152subepitelial, cavidad bucal, 230tejido adiposo, 72, 73f
epitelial, 48–56clasificacion, 48, 49f, 49tfunciones, 48polaridad y especializaciones de superficie,
50–56de granulacion, 90linfoide asociadoa los bronquios (BALT), 18a las mucosas (MALT), 185, 187f, 238al intestino (GALT), 185
reticular, 72Telofase, 36, 37fTelofase I de la meiosis, 38, 39fTenascina, 42Tendones, 94Tenia del colon, 248Terminaciones nerviosas peritriquiales, 304,
305fTerminales axonales, 100, 116–118Termogeninas, 20, 73cTermorreceptores, 306Testıculos, 286–290, 287fhistofisiologıa, 294, 295f
Testosterona, 292, 294Tetania, 101cpor calcio, sındrome de DiGeorge, 181
Tetr�ameros de espectrina, 134Tetrayodotironina, 196, 203tTGF-b. Vease Factor(es) de crecimiento tumo-
ral-b (TGF-b); Factor(es) transformadordel crecimiento-b (TGF-b).
Timo, 168, 180, 179fTimocitos, 180doblesnegativos, 180
positivos, 180sencillos positivos, 180
Tincionel�astica de orceına, 3ten microscopia optica, 2, 3tde plata, 3t
Tirocalcitonina, 203tTiroglobulina, 196Tirosinadiyodada (DIT), 196monoyodada (MIT), 196
Tirotrofos, 190Tirotropina, 190corionica, secrecion por sincitiotrofoblastos,
282Tiroxina, 196, 203tTitina, 96TnC. V�ease Troponinas.TnI. V�ease Troponinas.TnT. V�ease Troponinas.Todo-trans retinal, 312Tonofibrillas, 206Tonofilamentos, 56, 206Toxina(s)antibotulınicas, 119cbotulınica, 119c
Trab�eculas, 82, 84, 126del bazo, 183de ganglios linf�aticos, 182
Tr�aficode membrana, 18, 19fendocitosis, 18endosomas, 18, 19flisosomas, 18peroxisomas, 18proteasomas, 18
de proteınas, 16, 17fTranscitosis, 158Transcripcion, 30, 31fTransduccion, 10, 12de senales, 188
Transferasa de glucuronilo, 258Transferrina, testicular, 288Transmision de volumen de
neurotransmisores, 120Transportador, 26Transporteacoplado, 10activo, 10, 11fg�astrico, 242secundario, 10
anterogrado, 16antiporte, 10mediado por receptores, 28, 128pasivo, 10, 11fretrogrado, 16simporte, 10uniporte, 10
Tr�aquea, 222, 223fTrastornosdel almacenamiento de glucogeno, 23cautoinmunitarios, 175c
Traumatismo oclusal, 235cTrıadas, 96, 97fretinianas, 314
Tricelulina, 54Tricohialina, 214Trifosfato de adenosina (ATP), 96, 100sıntesis, 20
Trıgono de vejiga urinaria, 270Trimerizacion, 80Tripletes de microtubulos, 52, 53fTrismo, 101cTrisomıa del cromosoma, 21, 39cTriyodotironina, 196, 203tTrofoblastos, 276, 282Trombina, 140Trombomodulina, 140
340
INDICEALFA
BETIC
O

Tromboplastinaplaquetas, 140tisular, 140
Trombopoyetina, 148, 149tTrombos, 140Tromboxano A2, 68, 69tplaquetas, 140
Tronco pulmonar, 164Tropocol�ageno, 42, 43f, 44, 45fTropomodulina, 96Troponina, 98troponina C, 98troponina I, 98especıfica de cardiocitos (Tnl especıfica de
cardiocitos), 105ctroponina T, 98
TSH. Vease Tirotropina.Tubodigestivo, 238–248. Veanse tambien organos
especıficos.organizacion general, 238, 239f
neural, 108Tubulina a, 22, 23fTubulina b, 22, 23fTubulina t, 22Tubulo(s)colectores, 264corticales, 264lıquido filtrado, movimiento de agua y
urea dentro y fuera, 268papilares, 264
contorneadodistal, 262, 264, 265fproximal, 262
dentinales, 231distal, 260, 262, 264, 265fproximal, 260, 262, 263f, 268rectos, 286, 296, 297tseminıferos, 286, 288–290, 289furinıferos, 260, 261f
Tubulos T, 96, 97f, 104Tuftelinas, 230Tumor(es). Veanse tambien C�ancer.carcinoides, 60cneuroendocrinos, 60c
Tunicaadventicia, 152–154, 153f, 155farterial, 155tvenosa, 161t
albugıneade cuerpos er�ectiles del pene, 300ovarios, 272testıculos, 286
fibrosa del ojo, 306ıntima, 152–154, 153f, 155farterial, 155t
media, 152–154, 153f, 155farterial, 155tvenosa, 161t
propia, 288vaginal, 286vasculosadel ojo, 308, 309ftesticular, 286
Ubiquinona, 18Ultrafiltracion glomerular, 262, 266Unidadformadorade brote-eritrocitos (BFU-E), 146de colonias de linfocitos (CFU-linf), 146
de membranas, 8Union(es)de anclaje, 52dentina-esmalte, 233estrechas, 52, 53f, 54, 55f, 114, 156gap, 52, 53f, 55f, 56, 57f, 79miotendinosa, 94neuromusculares, 100, 101f, 103foclusivas, 288osea, 91c
Unas, 216, 217fUPC-1. Vease Proteına(s) de desacoplamiento-
1 (UPC-1).Uracilo, 30Ur�eteres, 270, 271fUretramasculina, 300prost�atica, 296
Urogastrona, 242, 246Utero, 278, 279fbicorne, 273cdidelfo, 273c
Utrıculo, 318Uvula, 235
Vainacentral, axonemas, 50externa de la raız, 214linf�atica periarterial (VLPA), 183mitocondrial, 290del pelo, 214de la raız interna, 214
V�alvula(s)anales, 248auriculoventricularderecha, 164izquierda, 164
bicuspide, 164cardıacas, 164mitral, 164semilunar, 164tricuspıdea, 164venosas, 160
Vasa vasorum, 154Vasectomıa, 296cVasopresina, 192efectos fisiologicos, 194tpresion arterial, 156
Vasosdeferentes, 286, 296, 297tlinf�aticosaferentes, 166eferentes, 166, 182
rectos, 266, 268, 269fVater, papila, 246, 252, 258VEA. Vease V�enulas endoteliales altas (VEA).Vejiga urinaria, 270Vello, 214Vellosidadesde anclaje, 282aracnoideas, 126corionicas, 282, 283fdel intestino delgado, 244libres, 282primarias, 282secundarias, 282
Velocidad de conduccion nerviosa,122, 122t
Vena(s), 152, 160, 161tarqueada, 266, 267fcava inferior, 254, 255fcentral, 254, 255fcolectoras, 254de distribucion, hep�aticas, 254dorsal profunda, 300hep�aticas, 254, 255flongitudinal central, 142en m�edula osea, 142porta hep�atica, 254de pulpa espl�enica, 184, 185frectas, 266, 268renal, 260sublobulillar, 254, 255fsuprarrenal, 200varicosas, 161c
Ventanaoval, 318redonda, 318
Ventilacion, 218mecanismo, 228
Ventrıculo(s)cardıacos, 164, 165fderecho, 164, 165fizquierdo, 164, 165f
V�enulas, 160, 200endoteliales altas (VEA), 182hep�aticas de la entrada, 254
Vermis, 130Vesıcula(s)acrosomicas, 290biliar, 258de la matriz, 88optica, 310pinocıticas, 18, 156, 158recubiertas, 16de clatrina, 16, 158
seminales, 298sin�apticas, 100, 118de transferencia, 14de transporte, 16
Vestıbulode la cavidad bucal, 230de laberinto oseo, 318entre labios menores, 284, 285f
Vıa(s)constitutiva de proteınas secretoras, 16extrınsecas de la apoptosis, 38hipotalamohipofisaria, 192intrınseca de la apoptosis, 38reguladas de proteınas secretoras, 16
Villina, 24, 50Vimentina, 106, 156union, 24
Vincristina, 35cVinculina, 104Vitamina(s)cartılago hialino, 77tdesarrollo oseo, 93t
Vitamina A, desarrollo oseo, 93tVitamina B12, 240Vitamina C, desarrollo oseo, 93tVitamina D, 198desarrollo oseo, 93t
Vitıligo, 209cVLDL. Vease Lipoproteınas de densidad muy
baja (VLDL).VLPA. Vease Vaina linf�atica periarterial
(VLPA).Volkmann, canales, 82–84Von Ebner, gl�andulas, 236Von Willebrand, factor, 140, 153c
Weigert, tincion el�astica, 3tWharton, gelatina, 70Wright, tincion, 3t, 133
Yemas alveolares, 284Yodofuncion tiroidea, 196oxidacion, 196
Yunque, 318
Zeis, gl�andulas, 316Zellweger, sındrome, 19cZollinger-Ellison, sındrome, 241cZonabasal de osteoclastos, 80de calcificacion, 88, 89fde cartılago de reserva, 88, 89fciliar, 308clara de osteoclastos, 80de estimulacion m�axima, 116–118fasciculada suprarrenal, 200glomerular suprarrenal, 200intermedia, 190, 191fde maduracion e hipertrofia, 88, 89fmarginal del bazo, 183
341INDICEALFA
BETIC
O
�ELSE
VIER.Fo
tocopiarsinau
torizaciones
undelito.

Zona (cont.)oclusiva, 114de osificacion, 88, 89fpapilar, 308pelucida, 274, 275fpobre en c�elulas de la pulpa dental, 230
de proliferacion, 88, 89fde reaccion, 280reticular suprarrenal, 200rica en c�elulas de la pulpa dental, 230de sellado de los osteoclastos, 80vesicular de osteoclastos, 80
Zonulaadherente, 52, 53f, 54, 55f, 57foclusiva, 52, 53f, 54, 55f, 57f
ZP1, 274ZP2, 274ZP3, 274
342
INDICEALFA
BETIC
O


















