
heT usnGe lac cBa The Genus Bacillus—Nonmedicalsaltikov/migrated/bio119l/...The other genera of...
33
CHAPTER 1.2.16 T h e G e n u s B a c i l l u s - - N o n m e d i c a l The Genus Bacillus—Nonmedical RALPH A. SLEPECKY AND H. ERNEST HEMPHILL History One of the earliest bacteria to be described was “Vibrio subtilis” by Ehrenberg in 1835. In 1872, Cohn renamed the organism Bacillus subtilis (Gordon, 1981). That organism was a charter member of a large and diverse genus , initiated by Cohn, that is part of the family Bacillaceae. This family’s distinguishing feature is production of endospores , which are round, oval, or cylindrical highly refractile structures formed within bacte- rial cells. Spores were first described by Cohn in subtilis and later by Koch in the pathogen, B. anthracis (the only major pathogen of verte- brates in the genus). Cohn demonstrated the heat resistance of spores of B. subtilis and Koch first described in B. anthracis the developmental cycle of sporeformers , vegetative cell to spore and spore to vegetative cell (Keynan and San- dler, 1983). For the reasons of unusual spore resistance to chemical and physical agents; the developmental cycle; ubiquity of its members; and B. anthracis pathogenicity, the genus Bacillus attracted early interest which has continued since. The endospore, either as the free spore or as the structure within the vegetative cell, in which case the whole entity is referred to as a spo- rangium, is readily detected using the phase con- trast microscope (see Fig. 1). This is because the spore at a point in the life cycle (to be detailed later) becomes highly refractile. Early workers used stains and special conditions (such as pro- longed heating) to colorize the chemically im- permeable spore (Doetsch, 1981). However, a Gram-stain is sufficient to determine the pres- ence of spores because the spore remains un- stainable while the vegetative cells or the vegetative part of the sporangia will stain. Because of this ease of microscopic detection of the spore and its heat resistance, many different endosporeformers can be easily found. Using any habitat—soil, water, food, etc.—as the source, sporeformers can be readily isolated by suspending a sample in water and heating at 80°C for 10 to 30 min. Vegetative cells and other resting forms such as cysts and exospores are usually killed at that temperature. The heat- resistant endospore can then be plated on appro- piate media and isolates recovered in 24 to 48 h. An idea of the kinds of habitats from which Ba- cillus species have been isolated can be obtained from Table 1. Heating the inoculum, when used in conjunction with cultivation at different tem- peratures , hydrogen ion concentrations , degrees of aeration, and substrates , has resulted in isolat- ing many different species of endosporeformers. The media used for the isolation and cultivation of Bacillus species are listed in Table 2. More often than not since the discovery of bacteria (and in every case since 1913), the pos- session of an endospore has been used as a pre- mier characteristic in keys for the classification of bacteria. The family Bacillaceae was first for- mulated by Fisher in 1895 (Gordon, 1981). The features of the members of the genus Bacillus that distinguish it from other Bacillaceae (all endosporeformers) are their aerobic nature, which may be strict or facultative, rod shape, and catalase production. The other genera of spore- formers include Sporolactobacillus, which is microaerophilic and catalase-negative; Clostrid- ium, anaerobic but does not reduce sulfate; Desulfotomaculum, anaerobic but does reduce sulfate; Sporosarcina, a coccus; and Thermoacti- nomycetes, which while forming endospores dis- plays typical actinomycete characteristics. General Taxonomic Considerations Like the sirens of Greek mythology enticing the unsuspecting sailors , Bacillus species have cap- tured the curiosity of many microbiologists. The first 107 years of the efforts to classify and iden- tify members of the genus Bacillus is chronicled by R.E. Gordon (1981) who with her colleagues (Gordon et al., 1973; Smith et al., 1946, 1952) made many significant contributions on which the current classification (Claus and Berkeley, 1986) was built. The early attempts were “on This chapter was taken unchanged from the second edition. Prokaryotes (2006) 4:530–562 DOI: 10.1007/0-387-30744-3_16
Transcript of heT usnGe lac cBa The Genus Bacillus—Nonmedicalsaltikov/migrated/bio119l/...The other genera of...

CHAPTER 1.2.16The Genus Bacillus--Nonmedical
The Genus Bacillus—Nonmedical
RALPH A. SLEPECKY AND H. ERNEST HEMPHILL
History
One of the earliest bacteria to be described was“Vibrio subtilis” by Ehrenberg in 1835. In 1872,Cohn renamed the organism Bacillus subtilis(Gordon, 1981). That organism was a chartermember of a large and diverse genus, initiated byCohn, that is part of the family Bacillaceae. Thisfamily’s distinguishing feature is production ofendospores, which are round, oval, or cylindricalhighly refractile structures formed within bacte-rial cells. Spores were first described by Cohn insubtilis and later by Koch in the pathogen, B.anthracis (the only major pathogen of verte-brates in the genus). Cohn demonstrated theheat resistance of spores of B. subtilis and Kochfirst described in B. anthracis the developmentalcycle of sporeformers, vegetative cell to sporeand spore to vegetative cell (Keynan and San-dler, 1983). For the reasons of unusual sporeresistance to chemical and physical agents; thedevelopmental cycle; ubiquity of its members;and B. anthracis pathogenicity, the genus Bacillusattracted early interest which has continuedsince.
The endospore, either as the free spore or asthe structure within the vegetative cell, in whichcase the whole entity is referred to as a spo-rangium, is readily detected using the phase con-trast microscope (see Fig. 1). This is because thespore at a point in the life cycle (to be detailedlater) becomes highly refractile. Early workersused stains and special conditions (such as pro-longed heating) to colorize the chemically im-permeable spore (Doetsch, 1981). However, aGram-stain is sufficient to determine the pres-ence of spores because the spore remains un-stainable while the vegetative cells or thevegetative part of the sporangia will stain.Because of this ease of microscopic detection ofthe spore and its heat resistance, many differentendosporeformers can be easily found. Usingany habitat—soil, water, food, etc.—as thesource, sporeformers can be readily isolated by
suspending a sample in water and heating at80
°C for 10 to 30 min. Vegetative cells and otherresting forms such as cysts and exospores areusually killed at that temperature. The heat-resistant endospore can then be plated on appro-piate media and isolates recovered in 24 to 48 h.An idea of the kinds of habitats from which Ba-cillus species have been isolated can be obtainedfrom Table 1. Heating the inoculum, when usedin conjunction with cultivation at different tem-peratures, hydrogen ion concentrations, degreesof aeration, and substrates, has resulted in isolat-ing many different species of endosporeformers.The media used for the isolation and cultivationof Bacillus species are listed in Table 2.
More often than not since the discovery ofbacteria (and in every case since 1913), the pos-session of an endospore has been used as a pre-mier characteristic in keys for the classificationof bacteria. The family Bacillaceae was first for-mulated by Fisher in 1895 (Gordon, 1981). Thefeatures of the members of the genus Bacillusthat distinguish it from other Bacillaceae (allendosporeformers) are their aerobic nature,which may be strict or facultative, rod shape, andcatalase production. The other genera of spore-formers include Sporolactobacillus, which ismicroaerophilic and catalase-negative; Clostrid-ium, anaerobic but does not reduce sulfate;Desulfotomaculum, anaerobic but does reducesulfate; Sporosarcina, a coccus; and Thermoacti-nomycetes, which while forming endospores dis-plays typical actinomycete characteristics.
General Taxonomic Considerations
Like the sirens of Greek mythology enticing theunsuspecting sailors, Bacillus species have cap-tured the curiosity of many microbiologists. Thefirst 107 years of the efforts to classify and iden-tify members of the genus Bacillus is chronicledby R.E. Gordon (1981) who with her colleagues(Gordon et al., 1973; Smith et al., 1946, 1952)made many significant contributions on whichthe current classification (Claus and Berkeley,1986) was built. The early attempts were “onThis chapter was taken unchanged from the second edition.
Prokaryotes (2006) 4:530–562DOI: 10.1007/0-387-30744-3_16

CHAPTER 1.2.16 The Genus Bacillus—Nonmedical 531
rocky shoals” because a classification based ononly the two characteristics of aerobic growthand endospore formation resulted in groupingtogether many bacteria possessing differentkinds of physiology and occupying a variety ofhabitats. This heterogeneity in physiology, ecol-ogy, and genetics makes it difficult to categorizethe genus or to make generalizations about it.The range of physiological life styles is impres-sive: degraders of most all substrates derivedfrom plant and animal sources including cellu-lose, starch, proteins, agar, hydrocarbons, andothers; antibiotic producers; heterotrophic nitri-fiers; denitrifiers; nitrogen fixers; iron precipita-tors; selenium oxidizers; oxidizers and reducersof manganese; facultative chemolithotrophs;acidophiles; alkalophiles; psychrophiles, thermo-philes and others (Slepecky, 1972; Norris et al.,1981; Claus and Berkeley, 1986) (see Table 2).Because of this vast diversity of physiologicaltypes, our knowledge of sporeformer ecologyis slight (Slepecky, 1972; Norris et al., 1981;Slepecky and Leadbetter, 1977, 1984). In themain, sporeformers as part of the zymogenousflora of the soil are viewed as opportunists. Uponaccess to the proper germinants and substratesfor subsequent outgrowth, they will actively con-tribute to and participate in the various micro-habitats which make up the soil’s heterogeneousenvironment. Aerial distribution of the dormantspores may explain the occurrence of Bacillusspecies in most habitats examined. This diversitywas apparent even with classical phenotypiccharacterizations based primarily on morphol-ogy (particularly size and position of theendospore within the vegetative cell), nutrition;growth characteristics; and various substrate uti-lization and physiological assessments.
At one time, 145 species made up the genus(Gordon, 1981). The understanding of the genushas been improved by augmenting the pheno-typic characterizations with measurements of theDNA base composition and DNA-DNA hybridi-gation. Currently, there are listed in Bergey’s
Fig. 1. B. megaterium sporeformingcells as seen in the phase contrast (A)or in the interference contrast (B)microscope. The refractile bodies inthe center are the spores. Bar
= 5
µm.
A B
Table 1. Origins of isolates of Bacillus species.
Based on Claus and Berkeley (1986).
Name of Bacillus species Habitats from which isolated
B. subtilis Soil, waterB. acidocaldarius Thermal acid water and soilB. alcalophilus pH 10 enrichment from soilB. alvei Soil, diseased bee larvaeB. amylolyticus SoilB. anthracis Anthrax-diseased animalsB. azotoformans SoilB. badius Feces, foods, marine sourcesB. brevis Soil, foodsB. cereus Soil, foodsB. circulans SoilB. coagulans Acid foodsB. fastidiosus Soil, poultry litterB. firmus Soil, salt marshesB. globisporus Soil, waterB. insolitus SoilB. larvae Diseased bee larvaeB. laterosporus Soil, waterB. lautus Soil, fecesB. lentimorbus Diseased honeybee larvaeB. lentus Soil, foodsB. licheniformis SoilB. macerans Plant materials, foodB. macquariensis Subantarctic soilB. marinus Marine sedimentB. megaterium SoilB. mycoides SoilB. pabuli Soil, fodderB. pantothenicus SoilB. pasteurii Soil, water, sewageB. popilliae Diseased scarabid beetlesB. psychrophilus Soil, waterB. pumilus SoilB. schlegelii Lake sedimentB. sphaericus Soil, water sediments, foodsB. stearothermophilus Soil, hot spring, foodsB. thermoglucosidasius SoilB. thuringiensis Soil, foodsB. validus Soil

532 R.A. Slepecky and H.E. Hemphill CHAPTER 1.2.16
Table 2. Media used for the isolation and cultivation of Bacillus species.
aNumerical amounts are grams unless specified.bMost other Bacillus cultures will grow on nutrient broth and nutrient agar.Based on Claus and Berkeley (1986).
B. acidocaldarius Part A: (NH4)2SO4, 0.4; MgSO4, 1.0; CaCl2·2H2O, 0.5; KH2PO4, 6.0; distilled H2O, 1 liter; pH adj.to 4.0
Part B: glucose, 2.0; yeast ext., 2.0 distilled H2O, 1 literCombine A and B after sterilization
B. alcalophilus Part A: glucose, 1.0; peptone, 5.0; yeast ext., 5.0; KH2PO4, 10.0 MgSO4·7H2O, 0.2, distilled H2O, 900ml.
Part B: Na2CO3·10H2O, 20; distilled H2O, 100ml.Combine A and B after sterilization (final pH
= 10.5)B. azotoformans Peptone, 10.0; Na2HPO4·12H2O, 3.6; MgSO4·7H2O, 0.03; MnSO4·H2O, 0.05; KH2PO4, 1.0; NH4Cl,
0.5; CaCl2·2H2O, 0.1; distilled H2O, 1 liter.B. brevis K2HPO4, 0.2; MgSO4·7H2O, 0.02; NaCl, 0.02; FeSO4·7H2O, 0.01; MnSO4·H2O, 0.01; betaine,
betaine·HCl or valine, 0.05M; agar, 16.0; distilled H2O, 1 liter.B. fastidiosus K2HPO4, 0.8; KH2PO4, 0.2; MgSO4·7H2O, 0.05; CaCl2·2H2O, 0.05; FeSO4·7H2O, 0.015;
MnSO4·H2O, 0.01; uric acid, 10.0; distilled H2O, 1 liter.B. lentus Peptone, 10.0; meat ext., 10.0; agar, 15.0; distilled H2O, 1 liter. adj. pH to 7.0–7.5; after sterilization,
add 100g urea, steam for 10min.B. licheniformis Peptone, 5.0; meat ext., 3.0; KNO3, 80.0; distilled H2O, 1 liter; adj. pH to 7.0; fill glass-stoppered
bottle to top for anacrobic conditions.B. marinus Peptone, 5.0; yeast ext., 1.0; FePO4·4H2O, 0.01; agar, 15.0; aged sea water, 750ml; distilled H2O,
250ml; adj. pH to 7.6.B. pantothenicus Nutrient broth + 4% (w/v) NaClB. pasteurii Nutrient broth + 2% (w/v) ureaB. schlegelii Na2HPO4·2H2O, 4.5; KH2PO4, 1.5; NH4Cl, 1.0; MgSO4·7H2O, 0.2; CaCl2·2H2O, 0.01; ferric
ammonium citrate, 0.005; NaHCO3, 0.5; trace element soln, 5ml (ZnSO4·7H2O, 0.1; MnCl2·4H2O, 0.03; H3BO3, 0.3; CoCl2·6H2O, 0.02; CuCl2·2H2O, 0.001; NiCl2·6H2O, 0.02; Na2MoO4·2H2O, 0.03; distilled H2O, 1 liter) Other: 65C, atmosphere of 0.05atm. O2 + 0.01atm. CO2 + 0.45atm. H2
B. stearothermophilus Nutrient agar; incubate cultures at 55°C
Table 3. Major distinguishing characteristics of some Bacillus species.
aSee Chapter 77.
Thermophile B. stearothermophilus Hydroxy aromatic B. gordonaeB. thermodenitrificans compounds degraderB. caldotenax Nitrate reduction to N2 B. azotoformans
Thermophilic acidophiles B. coagulans B. licheniformisB. acidocaldarius Growth restricted to uric
acid, allantoin or allantoicacid
B. fastidiosus
Psychrophiles B. psychrophilus Growth at high pH andNH4Cl
B. pasteurii
B. macquariensis Requires pantothenic acid B. pantothenicusB. globisporus Requires biotin B. pumilusB. insolitus Produces heat stable
glucosidaseB. thermoglucosidasius
B. psychrosaccharolyticus Insect pathogensa B. thuringiensisAlkalophile B. alcalophilus B. larvaeFacultative chemolithotroph B. schlegelii B. popilliaeNitrogen fixers B. polymyxa B. sphaericus
B. macerans B. lentimorbusB. azotofixans Human and animal
pathogenB. anthracis
Alginate degraders B. alginolyticus B. cereusB. chrondrotinus
Aromatic acid and phenoldegrader
B. benzoevorans

CHAPTER 1.2.16 The Genus Bacillus—Nonmedical 533
Manual of Systematic Bacteriology (BMSB) 40recognized species (Claus and Berkeley, 1986),Table 4 lists these with their GC content. Thereare several validly published new species shownto be genetically and phenotypically distinctfrom other Bacillus species that have not beendescribed in Bergey’s Manual. These include B.pulvifaciens (Nakamura, 1984); B. alginolyticusand B. chrondrotinus, two alginate-degradingspecies, (Nakamura, 1987); B. smithii (Nakamuraet al., 1988); B. thermoleovorans. an obligatelythermophilic hydrocarbon-utilizing organism(Zarilla and Perry, 1987); B. benzoevorans, an
aromatic acid and phenol degrader (Pichinoty etal., 1984); and B. gordonae, degrader of hydroxyaromatic compounds (Pichinoty et al., 1986).
There are more than 200 species of Bacillus inthe category “Species Incertae Sedis” (Claus andBerkeley, 1986). These have been inadequatelydescribed or the orginal isolates have been lost.Presumably, these can be revived for listing inBergey’s Manual after reisolation and moredetailed studies. For example, after extensivereconsideration of phenetic and molecular datait has been proposed that B. flexus, B. fusiformis,B. kaustophilus, B. psychrosaccharolyticus, B.
Table 4. DNA-base composition and sources of the type strains of Bacillus species.
aTm, GC content by thermal melting.bBD, GC content by buoyant density.cND, not determined.dATCC, American Type Culture Collection; DSM, Deutsche Sammlung von Mikroorganismen; NCIB, National Collectionof Industrial Bacteria; NCTC, National Collection of Type Cultures; NRRL, Northern Regional Research Laboratory.
Bacillus species
GC content (mol%) Culture collection number
Tma BDb ATCCd DSM NCIB NCTC NRRL
acidocaldarius 60.3 62.3 27009 446 11725 NRS1607alcalophilus 37.0 36.7 27647 485 10436 4553 B14309alvei 44.6 46.2 6344 29 9371 6352 B383amylolyticus NDc 53.0 3034 NRS290anthracis 33.2 ND 14578 9388 10340azotoformans ND 39.0 29788 1046 B14310badius 43.8 43.5 14574 123 9364 10333 NRS663brevis 47.3 47.4 8246 30 9372 2611 NRS604cereus 35.7 36.2 14579 31 9373 2599 B3711circulans 35.5 35.4 4513 11 9374 2610 B380coagulans 47.1 44.5 7050 1 9365 10334 NRS609fastidiosus 35.1 35.1 29604 91 11326firmus 41.4 40.7 14575 12 9366 10335 NRS613globisporus 39.8 39.7 23301 4 11434 NRS1533insolitus 35.9 36.1 23299 5 11433larvae ND 50.0 9545 B2605laterosporus 40.2 40.5 64 25 9367 6357 NRS314lautus ND 50–52 3035 NRS666lentimorbus 37.7 ND 14707 2049 11202 B2522lentus 36.3 36.4 10840 9 8773 4824 B396licheniformis 46.4 44.7 14580 13 9375 10341 NRS1264macerans 52.2 53.2 8244 24 9368 6355 B172macquariensis 39.3 41.6 23464 2 9934 10419 B14306marinus 37.6 38.0 29841 1297 B14321megaterium 37.3 37.6 14581 32 9376 10342 B14308mycoides 34.2 34.1 6462 2048 NRS273pabuli ND 48–50 3036 NRS924pantothenicus 36.9 36.8 14576 26 8775 8162 NRS1321pasteurii 38.5 38.4 11859 33 8841 4822 NRS673polymyxa 44.3 45.6 842 36 8158 10383 NRS1105popilliae 41.3 ND 14706 2047 B2309psychrophilus 39.7 40.5 23304 3 NRS1530pumilus 41.9 40.7 7061 27 9369 10337 NRS272schlegelii 64.6 66.3 43741 2000sphaericus 37.3 37.1 14577 28 9370 10338stearothermophilus 51.9 51.5 12980 22 8923 10339 B1172subtilis 42.9 43.1 6051 10 3610 3610 NRS744thermoglucosidasius 45–46 ND 43742 2542 B14516thuringiensis 33.8 34.3 10792 2046 9134 NRS996validus ND 53–54 3037 NRS1000

534 R.A. Slepecky and H.E. Hemphill CHAPTER 1.2.16
simplex (Priest et al., 1988), and B. thiaminolyti-cus (Nakumura, 1989) be recognized.
The literature contains many important exper-iments done with Bacillus isolates that have notyet been properly identified to species such as:Bacillus sp. strain SGI, a manganese-oxidizingand reducing organism (Johannes et al., 1986;Rosson and Nealson, 1982); Bacillus sp. strain C-59 and Bacillus sp. strain N-6, both alkalophilicorganisms with unusual bioenergetic properties(Kitada and Horikoshi, 1987; Kitada et al., 1989);Bacillus sp. strain Gx6638, a novel alkalineand heat stable serine protease-secreting strain(Durham et al., 1987); and Bacillus sp. strainMGA3, a thermophilic methanol-utilizing spe-cies, mutants of which are capable of producinglarge amounts of lysine (Guettler and Hanson,1988; Schendel et al., 1989).
An extensive list of phenetic characters ofmost members of the genus have been compliedand procedures for the isolation and identifica-tion of individual species have been presented(Gordon et al., 1973; Berkeley and Goodfellow,1981; Norris et al., 1981; Claus and Berkeley,1986). Summaries of one such rendition areshown in Tables 5 and 6 (Norris et al., 1981).
The GC content (32–69 mol%) of the knownBacillus species as well as DNA hybridizationexperiments have revealed the heterogeneity ofthe genus (Priest, 1981; Fahmy et al., 1985) (seeTable 4). Not only is there variation from speciesto species but there are differences in GC con-tent within strains of a species identified on otherbases. For example, the GC content of the B.megaterium group ranges from 36 to 45% (Hun-ger and Claus, 1981). It is thus understandable
Table 5. Simplified key for the tentative identification of typical strains of Bacillusspecies.
aNumbers on the right indicate the number (on the left) of the next test to be applieduntil the right-hand number is replaced by a species name.From Norris et al. (1981).
1. Catalase: positive ..................... 2negative .................... 17
2. Voges-Proskauer: positive ................ 3negative ............... 10
3. Growth in anaerobic agar: positive ............................ 4negative ........................... 9
4. Growth at 50°C: positive ............. 5negative ............ 6
5. Growth in 7% NaCl: positive ............................................. B. licheniformisnegative ............................................ B. coagulans
6. Acid and gas from glucose (inorganic N): positive.......... B. polymyxanegative......... 7
7. Reduction of NO3 to NO2: positive............................ 8negative .......................... B. alvei
8. Parasporal body in sporangium: positive .................. B. thuringiensisnegative ................. B. cereus
9. Hydrolysis of starch: positive ...................................... B. subtilisnegative ..................................... B. pumilus
10. Growth at 65°C: positive ............................................. B. stearothermophilusnegative ............................................ 11
11. Hydrolysis of starch: positive ...................................... 12negative ..................................... 15
12. Acid and gas from glucose (inorganic N): positive............... B. maceransnegative ............. 13
13. Width of rod 1.0µm or greater: positive ................... B. megateriumnegative................... 14
14. pH in V-P broth <6.0: positive .................................... B. circulansnegative ................................... B. firmus
15. Growth in anaerobic agar: positive ............................ B. laterosporusnegative ........................... 16
16. Acid from glucose (inorganic N): positive ................ B. brevisnegative ............... B. sphaericus
17. Growth at 65°C: positive ............................................. B. stearothermophilusnegative ............................................ 18
18. Decomposition of casein: positive.............................. B. larvaenegative............................. 19
19. Parasporal body in sporangium: positive .................. B. popilliaenegative ................. B. lentimorbus

CHAPTER 1.2.16 The Genus Bacillus—Nonmedical 535
that Priest et al. (1981, 1988), who have con-ducted extensive numerical analysis of many unitcharacters in addition to the DNA studies, haveproposed that the genus Bacillus be split intomultiple genera, since the intrageneric heteroge-neity is as great as exists in most bacterial fami-lies. Priest et al. (1988) assigned 80 organisms ofspecies rank to five or more cluster groups. Theirstudies reemphasized the heterogeneity of theB. brevis, B circulans, B. coagulans, B. sphaeri-cus, and B. stearothermophilus groups.
A variety of techniques have been employedto find either a simple approach to Bacillustaxonomy or a quick and painless identificationmethodology. Assessment of lipid analyses(reviewed by Minnikin and Goodfellow, 1981)indicated that Bacillus could not be separatedinto discrete groups. On the other hand, somespecies could be delineated from others. Forexample, B. acidocaldarius could be character-ized by its menaquinone (nine isoprenoid units,MK-9), cyclohexyl fatty acids, triterpenes, andcomplex lipids.
Using the API System (Analytab ProductsIncorporated) (a rapid identification systemwherein many standardized biochemical assess-ments can be made on test strips) and some
supplementary classical determinants, Loganand Berkeley (1981, 1984) have examined 1,075Bacillus strains. They were able to show that theAPI System tests were more reproducible thanthe classical tests.
Pyrolysis gas-liquid chromatography has beenapplied to the problems of Bacillus taxonomy(O’Donnell and Norris, 1981; O’Donnell et al.,1988). Although there are still some problemswith the technique, some promise for its use inclassification and identification has been shown.For example, as with DNA-DNA hybridizationstudies, a separation has been made between B.subtilis and B. amyloliquefaciens. When gas-liquid chromatography was applied to examinethe subgroups of B. megaterium, data wereobtained that confirmed the heterogeneity of thegroup even though there was some difficulty inresolving relationships within the group.
Shute et al. 1984 have used Curie-point pyrol-ysis mass spectrometry as a taxonomic tool. B.subtilis, B. pumilus, B. licheniformis, and B. amy-loliquefaciens could be separated using dataobtained from nonsporulating cultures (thosegrown on nutrient agar); however, such was notthe case with cultures sporulating on nutrientagar plus manganese.
Table 6. Summary of the characters used in the simplified key for Bacillus species.
+, Greater than 85% of strains examined by Gordon, Haynes, and Pang (1973) positive; !, greater than 85% of strainsnegative; V, variable character.aGrowth in 2% NaCl agar.
Cat
alas
e
V-P
rea
ctio
n
Gro
wth
in a
naer
obic
aga
r
Gro
wth
at
50°C
Gro
wth
in 7
% N
aCl
Aci
d an
d ga
s in
glu
cose
NO
3re
duce
d to
NO
2
Star
ch h
ydro
lyze
d
Gro
wth
at
65°C
Rod
s 1.
0µm
wid
e or
wid
er
pH in
V-P
med
ium
<6.
0
Aci
d fr
om g
luco
se
Hyd
roly
sis
of c
asei
n
Para
spor
al b
odie
s
B. megaterium + !!!+! V + !+ V + + !B. cereus + + + !+!++!++++ VB. thuringiensis + + + ! + ! + + ! + + + + +B. licheniformis + + + + + !++!! V + + !B. subtilis + + !++!++!! V + + !B. pumilus + + ! + + ! ! ! ! ! + + + !B. firmus + ! ! ! + ! + + ! ! ! + + !B. coagulans + + + + !! V + ! V + + V !B. polymyxa + + + !!+++!! V + + !B. macerans + ! + + ! + + + ! ! ! + ! !B. circulans + ! V + V ! V + !! V + V !B. stearothermophilus V ! ! +!! V + + V + + !B. alvei + + + !!!!+! V + + + !B. laterosporus + ! + + ! ! + ! ! ! ! + + +B. brevis + !!+!! V ! ! ! ! + + !B. larvae ! ! +!+ a ! V ! ! ! ! + + !B. popilliae ! ! +!+ a ! ! ! ! ! ! + ! +B. lentimorbus ! ! + ! ! ! ! ! ! ! ! + ! !B. sphaericus + !!! V ! ! ! ! V ! ! V +

536 R.A. Slepecky and H.E. Hemphill CHAPTER 1.2.16
Ribosomal RNA Sequencing
The most effective approach to Bacillus taxon-omy may be analysis of 16S rRNA molecules byoligonucleotide sequencing (Fox et al., 1977;Stackebrandt and Woese, 1979). That techniqueholds much promise for leading microbial taxon-omy into natural phylogenetic relationships.However, traditional taxonomists may be dis-mayed to find that Bacillus species show kinshipwith nonsporeforming species. Early studies withthis powerful tool showed a close relationshipamong Bacillus, Planococcus, Sporosarcina, Staph-ylococcus, and Thermoactinomycetes (Stacke-brandt et al., 1987; Stackebrandt and Woese,1981). In a recent study 16S rRNA catalogingshowed that B. subtilis and other ellipsoidal-sporeforming species, B. cereus, B. megaterium,and B. pumilus, formed a coherent cluster, whilethe round-sporeforming species, B. sphaericus,B. globisporus, and “B. aminovorans” did notcluster. Furthermore, the latter group werecloser phylogenetically to nonsporeformingorganisms as follows: B. sphaericus to Caryoph-anon latum; B. globisporus to Filibacter limicola;B. pasteuri to Sporosarcina urea and “B.aminovorans” to Planococcus citreus. Cell wallcomposition agreed except with the last case. B.stearothermophilus fell outside the main Bacilluscluster and showed some relationship to Ther-moactinomycete vulgaris (Stackebrandt et al.,1987).
In a more recent 16S rRNA sequencing survey,three major Bacillus taxonomic cluster groupswere defined (Jurtshuk et al., 1989). This wasaccomplished by determining complete or partialsequences of 16S RNA on 35 recognized neotypereference strains or type species by the techniqueof Lane et al. 1985. The partial sequences ana-lyzed typically exceeded 1,100 nucleotides. Phy-logenetic analyses were performed using three
different approaches (Sneath and Sokal, 1973;Fitch and Margoliash, 1967; Saitou and Nei,1987) which showed three major groupings ofBacillus spp., hereinafter referred to as clustersI, II, and III (see Table 7). The 16S rRNA Bacil-lus cluster groups were quite different fromthose previously noted by Stackebrandt et al.(1987). This is revealed by direct comparison tothe commonly used morphological groupings(see Table 7). Except for morphological group IIand the Unassigned Subgroup 2E, all strainssequenced fell into the B. subtilis cluster I group-ing. Bacillus strains of morphological group IIfell into all three 16S rRNA cluster groups andB. macquariensis, unlike other psychrophiles, fellinto the B. alvei cluster II group.
Comparative 16S rRNA analyses on thermo-philic and psychrophilic Bacillus strains (Wisotz-key et al., 1989) showed that the thermophiles,B. stearothermophilus, B. thermodenitrificansand B. caldotenax formed a subgroup within theB. subtilis cluster but separate from both the“thermotolerant” mesophilic, B. subtilis and B.licheniformis strains, and the moderate thermo-phile, B. coagulans. The psychrophilic strains, B.psychrophilus and B. insolitus, fell into cluster Iwhile B. macquariensis fell into cluster II.
Because several species were included in thecurrent study that had previously been examinedby 16S rRNA oligonucleotide cataloging, it ispossible to compare the two data sets directly.As a result, it is possible to augment the mem-bership of cluster I to include B. fastidiosus, B.firmus, B. badius and B. pasteurii (C. B. Woese,personal communication). In addition, it isextremely likely that at least two nonspore-forming strains, Planococcus citreus (Stacke-brandt and Woese, 1979) and Filibacter limicola(Clausen et al., 1985), as well as Sporosarcinaureae (Pechman et al., 1976), are properlyregarded as members of cluster I.
Table 7. Bacillus 16S rRNA cluster groups.
Table provided by Peter Jurtshuk.
Morphological group B. subtilis cluster I B. alvci cluster II B. brevis cluster III
I B. subtilis, B. cereus, B. licheniformis,B. pumilus, B. megaterium strainMohb, B. coagulans, B. smithil
II B. circulans, B. larvae, B. stearothermophilus B. alvei, B. polymyxa,B. macerans, B. azotofixans,B. pulvifaciens
B. brevis, B. laterosporus
III B. sphaericusSubgroup A “B. thiaminolyticus,” B. alcalophilusSubgroup B B. lentusSubgroup C B. freundenreichii, “B. aneurinolyticus”Subgroup D B. pantothenticusSubgroup E1 “B. psychrophilus,” B. insolitusSubgroup E2 B. macquariensis

CHAPTER 1.2.16 The Genus Bacillus—Nonmedical 537
Life Cycle; Sporulation and Germination as Models for Differentiation
IntroductionThe processes of resting cell formation and thechange back to the vegetative cell in a variety ofprokaryotes (Losick and Shapiro, 1984) presentexcellent models for studying differentiation,with the added attendant advantages of micro-bial systems: ease of handling, use of largenumbers of cells, fast growth, synchrony, andavailability of mutants. The endospore modelswere recognized early and, therefore, moreknowledge has been accumulated using themthan with other prokaryotic systems. As theattempt to categorize the many species of Bacil-lus has had a long history, so have the efforts tounravel the many aspects of the life cycle ofBacillus (for a historical treatment see Keynanand Sandler, 1983). Because of the enormousliterature in this area, the present treatment ofthe life cycle relies mainly on reviews of the sub-ject and covers mainly highlights of germinationand sporulation.
The cycle of germination, outgrowth,growth, and sporulation (shown schematicallyin Fig. 2) has been studied from many differ-ent aspects with many different species ofspore-formers but because of the genetic ver-satility of B. subtilis, most work has focused onthis species.
Germination and Outgrowth
Free spores usually must be activated for germi-nation. Activation is a reversible process whichconditions the spore for germination andincreases the number of spores undergoing ger-mination as well as the rate of germination.Spores can be activated by a variety of treat-ments, notably exposure to heat. During activa-tion there is a loss of some coat protein,dipicolinic acid (DPA), and Zn2+ along with anincrease in membrane fluidity. Germination, thebreaking of the spore’s highly dormant state,follows (recent reviews on germination includeSetlow, 1983, and Foster and Johnstone, 1989). Aseries of degradative reactions is triggered in anunknown manner by simple compounds such ascertain amino acids and ribosides or mixtures(no universal germinant has been described) orcertain nonnutrient conditions, and can be mon-itored by the loss of spore refractility as seen inthe phase-contrast microscope and by decreasein optical density. No metabolic activity can bedetected during the first 2 min of germination ofspores that require alaine or glucose for germi-nation. Generation of ATP or production ofknown metabolic products of these initiators hasnot been found. Mutants deficient in key glyco-lytic pathway enzymes can germinate, thus rulingout glycolysis in the case of spores requiringglucose for germination. The same spores canbe germinated by nonmetabolizable glucoseanalogs as well. However, metabolism may playa role in the germination of B. fastidiosus, whose
Fig. 2. Cycle of germination, out-growth, and sporulation of a typicalsporeforming bacterium. Also shownare some biochemical and physicalevents associated with various stages.(modification of a figure in Slepecky,R.A. (1978)).
GERMINATION OUTGROWTH
ACTIVATION
GERMINANTSSWELLING EMERGENCE ELONGATION
VEGETATIVECELLMICROCYCLE
SPORULATION
SYMMETRICALDIVISION
CONTINUED DIVISIONUNDER SOMECONDITIONS
ExoenzymesAntibiotics
Alanine Dehydrogenase
Alkaline PhosphataseGlucose DehydrogenaseAconitaseHeat-restant Catalase
CysteineIncorporationChemical & UVResistance
AlanineRacemaseHeat Resistance
RefractilityRibisidaseAdenosine DeaminaseDipicolinic acid
ASYMMETRICALDIVISION(STAGE II)
ENGULFMENT(STAGE III)
CORTEXFORMATION(STAGE IV)
SPORULATION(Stage II - Free Spore)
LYSIS OFLARGE CELL
FREE SPORE(REFRACTILE)
GERMINATEDSPORE(NON-REFRACTILE)
COATFORMATION(STAGE V)
MATURATION(STAGE VI)

538 R.A. Slepecky and H.E. Hemphill CHAPTER 1.2.16
spores can only be germinated with uric acid,which is the main carbon source for these uniqueorganisms (Aoki and Slepecky, 1973). Onehypothesis suggests that germinants act onreceptor proteins, possibly in the inner mem-brane, which then undergo conformationalchanges that alter permeability (Foster andJohnstone, 1989). This leads to an autocatalyticloss of heat resistance and to changes thatinitiate metabolism, leading to vegetativegrowth. Another view, based on the observationthat inhibition of the electron transport systemaffects germination, postulates that respirationand ATP create a proton motive force, lendingto the establishment of a proton gradient. Theproton motive force is used for transport of ionsfrom core to cortex to neutralize other ions(Gould, 1983).
Upon germination, the spores not only losetheir resistance to heat but also resistance toradiation and injurious chemicals; their stainabil-ity also increases. Concomitantly with “phase”darkening, the spores swell, break out of theircoats, and exude up to 30% of their dry weight;about one-half of the exudate consists of a cal-cium chelate of the spore-specific substance,DPA, and the remainder consists of peptidogly-can fragments (from the action of cortex lyticenzymes) and amino acids. The earliest measur-able events are the loss of calcium, DPA, andheat resistance. This is followed by metabolicevents using high-energy compounds producedearly in germination from energy reserves storedin the dormant spore.
RNA synthesis begins rapidly within 2 min ofgermination. The dormant spore lacks the abil-ity to produce amino acids and amino acid bio-synthesis is absent early in germination. Duringthe first minutes of germination, 20% of thespore’s protein is degraded, providing thesource of amino acids for biosynthesis of newprotein and small molecules (such as nucle-otides) during outgrowth (reviewed by Setlow,1988). The spore’s enzymes are not degraded.Rather, a group of small acid-soluble proteins(SASP) are the source of the amino acids. Theseunique proteins, located in the core and sensi-tive to proteolysis, conprise 8 to 20% of the pro-tein in the spore. Their molecular weight is low(5–11 kDa) and although they are not histones,they bind to the spore DNA. The proteins aredegraded by a unique protease that has an abso-lute specificity for these proteins. They are syn-thesized late in sporulation. Several of the fiveknown SASP genes (referred to as ssp) havebeen cloned and in addition to coding proteinssupplying amino acids in germination theirproducts may have other roles. One has beenshown to be involved in ultraviolet (UV)resistance.
Many germination mutants (abnormal germi-nation phenotypes) have been described(reviewed by Moir et al., 1986; Foster andJohnstone, 1989). The ger (germination) genesare considered a subclass of spo (sporulation)loci, and are made up of several classes: I,structural genes for germination mechanismcomponents; II, regulatory genes for class I; III,post-translation processing and assembly genes;and IV, genes for synthesizing spore structure(e.g., cortex) required for germination.
Outgrowth is the period during which thespore gradually becomes a vegetative cell andinitiates new macromolecular synthesis. Genesassociated with this period are out genes. DNAis replicated relatively late in outgrowth, justbefore division. The vegetative cell is then capa-ble of undergoing various morphological andbiochemical changes which lead either to a seriesof symmetric cell divisions if sufficient nutrientsare present, whereas in stressful times (particu-larly nutrient limitation), subsequent spore for-mation or the production of a spore withoutintermittent cell division can occur. The latter isknown as microcycle sporulation (Vinter andSlepecky, 1965).
SporulationElectron microscopical analyses of cells duringsporulation has revealed seven stages (reviewedby Fitz-James and Young, 1969). The variousstages are shown in Fig. 2. Vegetative cells areconsidered to be Stage 0. Upon induction inputprior to actual sporulation, the nuclear materialis in an axially disposed filament. This is stage I.However, since such a pattern does not appearto be unique to sporulating cells, current practicerefers to cells in stages prior to stage II as pre-septation cells. Segregation of the chromatinmaterial to the poles of the cell occurs concomi-tantly with the invagination of the plasma mem-brane in an asymmetrical position on the cellwhich fuses to complete the spore septum. Thisis stage II. The mode of formation of this septumis similar to the formation of the transverseseptum of symmetric vegetative cell division.Indeed, it has been proposed that sporulationbecause of this and some other similarities is amodified prokaryotic division (Hitchins andSlepecky, 1969) and models for that view havebeen presented (Freese and Heinze, 1983). Insporulation, the division of the cell is not equaland subsequent proliferation of the larger cell’splasma membrane leads to complete engulfmentof the “forespore” and liberation of the imma-ture spore, surrounded now by a double unitmembrane, into the cytoplasm of the larger cell.This completes stage III. This is a key step sincethis double membrane now has different trans-

CHAPTER 1.2.16 The Genus Bacillus—Nonmedical 539
port properties owing to the opposing polarity ofthe two membranes.
During sporulation the vegetative cell isdivided into two compartments each having adifferent fate and each displaying different pat-terns of gene expression (reviewed by Setlow,1989). At this time, the cell is “committed” tocomplete the process of sporulation. The smallcell eventually becomes the core of the sporewhile the large cell, the mother cell, goes on toproduce the outer protective layers of the sporeand then lyses to release the spore. Cortex mate-rial similar to vegetative cell peptidoglycan (butdiffering in the degree of cross-linking and otheraspects) is laid down between the unit mem-branes, and its deposition corresponds in time tothe accumulation in the core of DPA and cal-cium. Stage IV is now completed. Studies withcortex-less mutants show that the cortex isneeded for refractility of the spore (when thespores become refractile, they can be seen in thephase-contrast microscope) and for accumula-tion of DPA. The cortex plays a fundamental rolein the dehydration of the spore (reviewed byGould, 1983).
During Stage V, protein coats are synthesizedby the mother cell. There are about 10 major coatproteins in B. subtilis and they are encoded bycot genes, seven of which are known (Losick etal., 1986). The coat proteins are placed aroundthe outside of the forespore. In some species, anadditional protein layer called an exosporium issynthesized. Since the coat may play importantroles in protection of the spore and its subse-quent germination, it has been the subject ofmany investigations (reviewed by Aronson andFitz-James, 1976; Losick et al., 1986). Electronmicroscopy reveals that B. cereus contains anouter coat showing a cross-patched pattern, aninner pitted layer, and a thin layer, the under-coat, while other species show distinct dif-ferences. B. subtilis possesses a very thickmultilayered coat with an outer striped layer andB. thuringiensis has a coat deficient in the outercross-patched layer. Differences show up as wellwithin the major structural polypeptides andcoat-associated proteases. These differences maybe responsible for the variation found in germi-nation and certain resistant properties (otherthan heat and UV resistance) of various species.
During vegetative growth and subsequentsporulation, a variety of proteases are produced(reviewed by Priest, 1977). There are six extra-cellular proteases and at least three major intra-cellular proteases—ISP, esterase A, and esteraseB. They may be involved with turnover of intra-cellular proteins, the processing of protein pre-cursors for spore coats, or inactivation of latersporulation enzymes, as well as other functions.As with other aspects of B. subtilis physiology,
there are other considerations. Even though thegenes apr, npr, epr, and isp, which code for theproteases alkaline (subtilisin), neutral (metallo-)“new” serine, and major intracellular serine,respectively, can be deleted, there still is someprotease activity (Sloma et al., 1988). This findingsuggests that there are other unindentifiedproteases.
As the spore matures (stage VI), it becomesresistant to heat and to a variety of organic sol-vents. Final lytic enzymes lyse the sporangial ormother cell liberating the free spore (stage VII).Figure 3 shows a cross section of a mature andheat-resistant spore. If the free spore is placedamong the proper nutrients it will germinate,completing the cycle. The sporulation processtakes 6 to 8 h at 37°C in B. subtilis.
Many other biochemical and physiologicalevents occur during sporulation in addition tothose indicated in Fig. 2 and those that can besurmised to be linked with the morphologicallyidentified stages. Some vegetative enzyme activ-ities disappear, some remain, others are modi-fied, and new sporulation specific enzymes aresynthesized. The number of sporulation-associated events is uncertain, but the geneticevidence suggests as many as 200 genes in 40 to50 operons are involved (Losick et al., 1986;Mandelstam and Errington, 1987; Piggot, 1989;Youngman et al., 1989). Currently the geneticmap, which includes sporulation and germina-tion genes as well as all-known vegetative cellgenes, contains 700 loci, more than 300 of whichhave been cloned and 180 of those sequenced(reviewed by Piggott, 1989). The ordered appear-ance of cytological and biochemical changes
Fig. 3. Cross-section of Bacillus megaterium containing aspore and showing the sporangium (cell), cell protoplast(CP) and wall (CW), spore coat (SC), spore cortex (SCor),spore membrane (SM), and spore protoplast (SP). "120,000.(Norris et al., 1981.)
CW CP SC SCor SM SP

540 R.A. Slepecky and H.E. Hemphill CHAPTER 1.2.16
implies a sequential reading of the genome. Thegenetic data are consistent with there being asingle linear dependent sequence to stage IIIwith a more complex pattern of gene expressionbeyond that stage.
The genotype of mutants blocked at the vari-ous morphological stages in sporulation (asdetermined by electron microscopy) are desig-nates spoO, spoII, spoIII, etc. For example, aspoII mutant would be arrested in stage II.Within each genotype, the designation A, B, C,etc. identifies different genetic locations; forexample, spoIIA, spoIIB identify two separatemutations that cause arrest of developmentalstage II. Eight loci have been shown to be con-cerned with stage 0, the five principal spoOgenes being spo0A, B, E, F, and H. These spo0mutants have pleiotropic effects on the pheno-type, suggesting that their gene products regu-late the expression of several genes. They do notmake polar septa (the asymmetric division) orStage 0 associated products—proteases, antibi-otics and transformation competent cells. Thereare suppressors of spo0 mutations, i.e., suppres-sor mutations can restore sporulation in somespo0 mutants to the wild type level. For example,sof-I can suppress defects in spo0F, B, and E(Hoch et al., 1985). Others include arbA andarbB suppressors of spo0A mutations; rvtA, aspo0 repressor; ssa, a reliever of suppression ofalcohols; crs, an alleviator of glucose repressionof sporulation; and sapA and sapB. suppressorsinvolved with alkaline phosphate induction(Losick et al., 1986). Studies on these extragenicsuppressors have given insights into the role ofspo0 mutants.
Sporulation, normally repressed at highgrowth rates in the presence of excess nutrients,will ensue at the beginning of stationary phase(arbritrarily defined as t0) in batch culture uponlimitation of carbon, nitrogen, or phosphate(reviewed by Smith, 1989, and Sonenshein,1989). The initiating signal for sporulation is notknown. However, sporulation initiation is asso-ciated with a decrease in the intracellular GTPpool (reviewed by Freese and Heinze, 1983).Decoynine, a drug that artificially reduces GTPlevel, induces sporulation even in the presenceof excess nutrients. There may be a connectionbetween GTP levels and a proposed sensingmechanism as follows: spo0A, B, and F sharesignificant homology and A and F products showpartial homology with proteins such as OmpR,NtrC, and SfrA which are “sensing proteins”involved in the transduction of various environ-mental signals (Trach et al., 1985; Nixon et al.,1986). Thus, spo0A and spo0F may play a role insensing starvation and transferring a signal toother genes (see review by Smith, 1989, on theinitiation of sporulation). A gene for a protein
kinase that phosphorylates sporulation regula-tory proteins Spo0A and Spo0F has been char-acterized (Pergo et al., 1989). The gene, kinA,previously known as spo11J, has been shown tocode for a protein that is homologous to thetransmitter class of proteins (Stragier, 1989).Defects in the methylation of a membrane-associated 40-kDa protein in some B. subtilisspo0 mutants suggests that protein methylationalso may be a part of the nutrient-sensing system(Golden and Bernlohr, 1989).
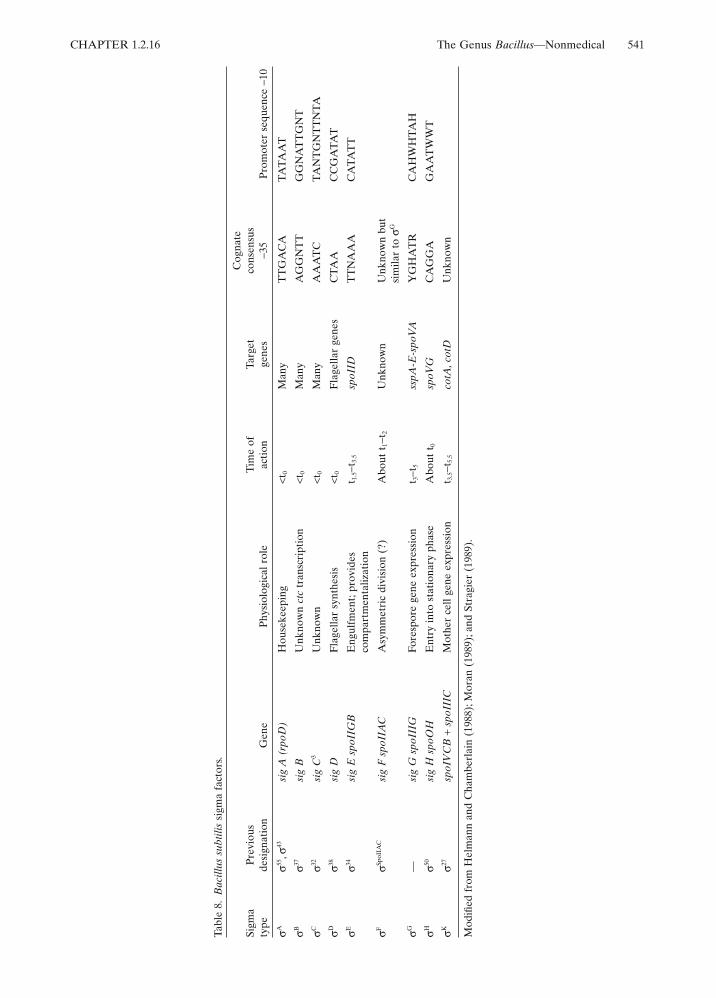
Multiple forms of RNA polymerase areof considerable importance in sporulation(reviewed by Doi and Wang, 1986; Losick et al.,1986; Losick and Kroos, 1989; Moran, 1989;Setlow, 1989; Stragier, 1989). The holoenzyme ismade up of an enzyme core comprised ofsubunits beta, beta# and alpha (the products ofgenes rpoB, rpoC, and rpoA, respectively) andattached sigma factors (the products of genes tobe designated), regulatory proteins that deter-mine the promoter specificity. Currently, thereare nine known sigma factors, four associatedwith vegetative cells and five involved in sporu-lation (see Table 8). Losick and Pero (1981) sug-gested there is a succession of different sigmafactors governing the transcription of the varioustemporal classes of sporulation genes.
The first acting sporulation sigma is ;gs;xHwhich is required for the initiation of sporulationand it is involved with the transcription of a latergene, spoVG.
A key point in sporulation is stage II, which isthe result of the asymmetric division event. stageII mutants and the associated gene products orfunctions are as follows: spoIIAA $E processing);IIAC ($E factor); IIE (pro $E); IIGA$E process-ing protease); IIGB (p31; $E); IIN (Fts homology)and IIJ (ntrB protein kinase) (Leighton cited inStragier (1989), and Stragier (1989)). That thespoIIN product has homology with E. coli Fts, aprotein involved in symmetrical cell division,may suggest that the level of Fts controls theasymmetric division. In turn, the functions of theother spoII mutations listed above implies arather complicated mechanism for the synthesisof $E. The sporulation septum (stage II), depen-dent on spoIIAC ($F) and spoIIE products, isrequired for spoIIGA processing activity, i.e.,conversion of pro-$E by proteolytic cleavage to$E. The important finding by Stragier (1989) ofthe coupling of gene expression to morphogene-sis may be a recurrent theme throughout otheraspects of sporulation.
After Stage II, compartmentalization of sigmafactors occurs. $E is active both in the mother celland forespore compartment. $G acts only in theforespore compartment (Setlow, 1989) while$K, the product of sig K, a composite genefrom IVCB plus IIC (due to a chromosome
CHAPTER 1.2.16 The Genus Bacillus—Nonmedical 541
Tabl
e 8.
Bac
illus
sub
tilis
sigm
a fa
ctor
s.
Mod
ified
fro
m H
elm
ann
and
Cha
mbe
rlai
n (1
988)
; Mor
an (
1989
); an
d St
ragi
er (
1989
).
Sigm
aty
pePr
evio
usde
sign
atio
nG
ene
Phys
iolo
gica
l rol
eTi
me
ofac
tion
Targ
etge
nes
Cog
nate
cons
ensu
s!3
5Pr
omot
er s
eque
nce
!10
$A$55
,$43
sig
A (
rpoD
)H
ouse
keep
ing
<t0
Man
yT
TG
AC
ATA
TAA
T
$B$37
sig
BU
nkno
wn
ctc
tran
scri
ptio
n<t
0M
any
AG
GN
TT
GG
NA
TT
GN
T
$C$32
sig
C3
Unk
now
n<t
0M
any
AA
AT
CTA
NT
GN
TT
NTA
$D$38
sig
DFl
agel
lar
synt
hesi
s<t
0Fl
agel
lar
gene
sC
TAA
CC
GA
TAT
$E$34
sig
E s
poII
GB
Eng
ulfm
ent;
prov
ides
com
part
men
taliz
atio
nt 1
.5!t
3.5
spoI
IDT
TN
AA
AC
ATA
TT
$F$Sp
oIIA
Csi
g F
spo
IIA
CA
sym
met
ric
divi
sion
(?)
Abo
ut t
1!t 2
Unk
now
nU
nkno
wn
but
sim
ilar
to $
G
$G—
sig
G s
poII
IGFo
resp
ore
gene
exp
ress
ion
t 3!t
5ss
pA-E
-spo
VA
YG
HA
TR
CA
HW
HTA
H
$H$50
sig
H s
poO
HE
ntry
into
sta
tiona
ry p
hase
Abo
ut t
0sp
oVG
CA
GG
AG
AA
TW
WT
$K$27
spoI
VC
B+
spoI
IIC
Mot
her
cell
gene
exp
ress
ion
t 3.5!t
5.5
cotA
, cot
DU
nkno
wn

542 R.A. Slepecky and H.E. Hemphill CHAPTER 1.2.16
rearrangement requiring SpoIIID and a recom-binase) acts only in the mother cell (Losick andKroos, 1989). $K directs the transcription of cotAand cotD, spore coat genes.
This brief overview of some of the emerginginformation on regulation of gene expressionduring sporulation reflects the current view of anetwork of dependent pathways in which activa-tion of developmental genes depends on theproducts of other developmental genes (Losickand Kroos, 1989).
Surface Structures of Bacillus
S-LayersCrystalline surface layers of protein or glycopro-tein subunits, called S-layers, are found in mem-bers of the genus Bacillus (reviewed by Sleytrand Messner, 1988). S-layers of individualstrains of Bacillus have been shown to differ inmolecular weight (40–200 kDa), the degree ofglycosylation of the subunits, and the geometryof the S-layer lattice. For example, B. stearother-mophilus contains one S-layer consisting of twotypes of glycan chains, one being a unique typeof protein-carbohydrate linkage. On the otherhand, B. brevis contains two S-layers, termedthe outer wall protein (OWP) and the middlewall protein (MWP), external to the peptidogly-can layer. These form a hexagonal array in thecell wall. The nucleotide sequence of the entireMWP-OWP gene operon is known (Tsuboi etal., 1988). The gene encoding an S-layer proteinof B. sphaericus has been cloned and sequenced(Bowditch et al., 1989). Not all Bacillus speciescontain S-layers and some strains within a spe-cies may lack such a layer. Furthermore, thetype of lattice may vary from species to speciesand within strains of a species. B. alvei, B.anthracis, and B. brevis show a hexagonal array;B. cereus, B. fastidiosus, “B. macroides,” B.megaterium, B. psychrophilus, and B. schlegeliipresent a square lattice, while strains of B.stearothermophilus can be obtained that individ-ually have one of the three types (Claus andBerkeley, 1986).
As with S-layers of other bacteria, their func-tion in Bacillus is unknown. However, since ithas been demonstrated that the S-layer can phys-ically mask the negatively charged peptidoglycansacculus in B. stearothermophilus and preventautoagglutination, it has been postulated that thelayer may play a key role in bacteria-metal inter-actions (Sleytr and Messner, 1988).
CapsulesThe capsule (a homopolypeptide of D-glutamicacid) of B. anthracis as a virulence factor has been
studied extensively (see The Genus Bacillus—Medical in this Volume). Other bacilli, such as B.subtilis, B. megaterium, and B. licheniformis, pos-sess capsules containing the homopolypeptide ofD- or L-glutamic acid as well (Makino et al.,1989). Some Bacillus species, e.g., B. circulans, B.mycoides, and B. pumilus, produce carbohydratecapsules. For example, B. circulans forms anextracellular polymer consisting of glucose andglucuronic acid (Claus and Berkeley, 1986). Inthe case of B. megaterium a heteropolysaccharidecomposed of D-glucose, D-xylose, D-galactose,and L-arabinose has been found in one strain(Cassity and Kolodziej, 1984).
FlagellaMost Bacillus species possess peritrichous fla-gella. Although some use has been made of H-antigens in setting up serotyping schemes, theyhave not been widely adopted (Claus and Ber-keley, 1986). Chemotaxis has been studied exten-sively in B. subtilis (Ordal and Nettleton, 1985).
Cell WallsAlmost all Bacillus species tested have vegeta-tive cell walls made up of peptidoglycan contain-ing meso-diaminopimelic acid (m-DAP). Theexceptions (B. sphaericus and the related species,B. pasteurii and B. globisporus) contain lysineinstead (Bartlett and White, 1985). But eventhose species, as all others, contain m-DAP in thepeptidoglycan of their spore cortex. Cell wallturnover in Gram-positive bacteria, particularlyBacillus species which have been useful models,has been reviewed by Doyle et al. (1988). Inaddition to peptidoglycan in the cell wall, allBacillus species contain large amounts of ananionic polymer, such as teichoic acid (a glycerolor ribitol-based polymer joined together byphosphodiester linkages to form a flexible linearstrand) or teichuronic acid (uronic acid-basedpolymer) which are bonded to muramic acid res-idues. The type of this anionic polymer presentdepends on the levels of phosphate and magne-sium in the growth medium. The glycerol teichoicacids vary a great deal between Bacillus speciesand within species. For example, B. subtilis cancontain either glucosyl % or & (1'2) glycerolor glucosyl % (1'6) galactosyl % (1'1 or 3)glycerol), while B. licheniformis contains galac-tosyl % (1'2) glycerol. However, they are joinedto the peptidoglycan through a common lin-kage disaccharide, acetylmannosaminyl(1'4)N-acetylglucosamine (Kaya et al., 1984).
As in other Gram-positive bacteria, lipote-ichoic acids are found associated with most of thecell membranes of Bacillus species. These com-pounds are involved in the synthesis of wall

CHAPTER 1.2.16 The Genus Bacillus—Nonmedical 543
teichoic acids as regulators of autholytic activityand as scavengers of bivalent ions. Those sotested in Bacillus fall into three groups based onthe presence or absence of N-acetylglucosaminebranches in the backbone chains of theirlipoteichoic acids—hydrophilic poly (glycerol-phosphate) chains and hydrophobic gentiobio-syldiacylglycerol anchors (Iwasaki et al., 1989).Group A is made up of strains of B. subtilis, B.licheniformis, and B. pumilus; group B, otherB. subtilis strains and B. cereus, and group C, B.polymyxa and B. circulans.
MacrofibersMacrofibers are multicellular and multistrandedstructures, hundreds of micrometers in length,produced by autocatalytic mutants of B. subtilis(Mendelson, 1978). These left- or right-handedhelical structures have been used to study cellwall structure and growth. They are thought toreveal cell-surface molecular organization andforce interactions in the cell wall not readily elu-cidated in the wild type, single-celled organism.The establishment and maintenance of macrofi-ber structure is influenced by both genetic andphysiological factors (Briehl and Mendelson,1987; Surana et al., 1988).
MembranesThe membranes of Bacillus species have beenstudied extensively because of their intrinsicinterest; as a model of membrane structure inGram-positive cells; with regard to their role insporulation and germination; with respect toexplanations for thermophily; and other reasons.For example, one explanation for the ability ofthermophilic microorganisms such as B. stearo-thermophilus to grow at high temperatures isthat the physical properties of the membrane arechanged due to changes in the lipid compositionin response to growth temperature (Gould,1983).
There is great diversity in the range and typeof lipids in Bacillus membranes (see reviewby Minnikin and Goodfellow, 1981) and widevariation in the fatty acids are found. The mainphospholipids present are phosphatidylglycerol,diphosphatidylglycerol, and phosphatidyletha-nolamine; however, others are found as well. Themajor isoprenoid quinones are menaquinone,and most species contain menaquinones withseven isoprenoid units (MK-7). B. acidocaldariusis the exception and possesses MK-9 (Minnikenand Goodfellow, 1981).
Two-dimensional polyacrylamide gel electro-phoresis (PAGE) has been used to attempt toresolve all B. subtilis membrane polypeptides(see Shohayer and Chapra [1985] for one such
study). Several membrane enzymes have beenisolated and characterized, such as the lactate,malate, glycerate-3-phosphate, NADH, and suc-cinate dehydrogenases.
Because of interest in synthesis and modifica-tion of the peptidoglycan layer, much attentionhas been paid to penicillin-binding proteins. Sixhave been found in B. subtilis, but one of themost penicillin-sensitive binding proteins, num-ber 4, has been found to be absent from B. sub-tilis 168 (Buchanan, 1987).
Genetic Studies
The discovery of transformation in B. subtilisstrain 168 by Spizizen (1958) was largely respon-sible for focusing attention on the genetics of thegenus Bacillus. Strain 168 is thought to be aderivative of the type strain B. subtilis Marburg(see Hemphill and Whiteley, 1975), and is one ofa relatively few bacilli in which competence forDNA uptake has been found to occur as a natu-ral part of the life cycle. As a consequence ofSpizizen’s discovery and the later isolation ofgeneralized transducing phages, our knowledgeof the chromosomal organization of B. subtilisis second only to that of the enteric bacteria.(About one-half as many genetic markers areknown in B. subtilis as in Escherichia coli.) Fur-thermore, the identification of numerous genesaffecting sporulation in B. subtilis is providing ameans for analyzing this complex developmentalprogram, which is largely unique to theGram-positives.
TransformationThe establishment of a competent state inbroth culture is most efficiently brought aboutin a minimal salts medium (Anagnostopoulosand Spizizen, 1961; Bott and Wilson, 1967). Asthe bacteria enter stationary phase in thismedium, a maximum of 20% become compe-tent, with a 1 to 2% transformation frequencyfor a given marker. The development of com-petence is associated with a reduction in mac-romolecular synthesis that is initiated wellbefore cells are transformable. Competentbacilli are relatively metabolically latent, andhave a lower buoyant density compared to non-competent bacteria (Dooley et al., 1971; Had-den and Nester, 1968). Although the period ofcompetence overlaps the time in which sporula-tion is initiated, the two forms of physiologicaldifferentiation are thought not to be connected.Different media are preferred for the two pro-cesses and commitment to competence isreversible. Several lines of research suggest that

544 R.A. Slepecky and H.E. Hemphill CHAPTER 1.2.16
the establishment of competence is coincidentalwith, and perhaps induces, a DNA-repair sys-tem analogous to the SOS regulon in E. coli(Love et al., 1985).
The development of the capacity of B. subtilisto take up DNA is associated with the appear-ance of several novel cellular proteins. Amongthese is a 75-kDa protein complex which hasbeen isolated from the membrane of competentcells (Smith et al., 1985). The complex consists oftwo subunits of 18 kDa (polypeptide A) and17 kDa (polypeptide B). Mutants lacking subunitA do not bind DNA to the cell surface whereasthose deficient in B are defective in DNA entry.The 75-kDa protein complex also has nucleaseactivity that is probably associated with polypep-tide B. A nuclease is expected, because it isknown that one strand of transforming DNA ishydrolyzed in the process of entering the cell,resulting in a single-stranded product on thecytoplasmic side of the membrane (Davidoff-Abelson and Dubnau, 1973a, 1973b). Compe-tence factors that interact with the membranehave also been reported in B. stearothermophilus(Streips and Welker, 1971). B. subtilis cells showlittle specificity with regard to DNA uptake andmay be transformed with homologous chromo-somal DNA, plasmid DNA, or transfected withbacteriophage DNA.
There is evidence that strands of transform-ing DNA enter the cell at sites on the mem-brane where the chromosomal DNA isattached (teRiele et al., 1984). The penetratingDNA, now reduced to a single-strand form, isthen brought in contact with the homologousregion of the recipient chromosome. This stepis probably mediated by the 45-kDa proteinproduct of the B. subtilis recE gene and resultsin a complex in which the transforming DNAbegins to displace one strand of the chromo-somal DNA while hydrogen-bonding to thecomplementary strand. A continuation of thedisplacement reaction allows pairing and inte-gration of several thousand bases of transform-ing DNA into the chromosome, while anequivalent amount of the homologous strand isremoved and degraded.
Considerable thought has been given to thequestion of whether transformation is associ-ated with genetic exchange in natural popula-tions of bacilli (see Stewart and Carlson, 1986,for a review). The complexity of the transfor-mation process with its requirement for uniquecompetence factors appearing only at station-ary phase suggests that the capacity to take-upexogenous DNA offers some selective advan-tage in the evolution of these bacteria. The factthat competence occurs only late in the growthcycle probably means the system is notdesigned to obtain DNA as a nutrient. Ephrati-Elizur 1968 found that B. subtilis cells excrete
high-molecular-weight DNA into liquid cultureas they grow. Under natural conditions thiscould be the source of donor DNA. Grahamand Istock (1978) demonstrated that geneticexchange, thought to be mediated by transfor-mation, occurs at high frequency betweengenetically labeled strains of B. subtilis in soil.Also, transformation frequencies in cultures inwhich the bacteria are allowed to attach tosand grains are much higher than in the stan-dard liquid culture procedure (Lorenz et al.,1988).
Generalized Transduction
Bacteriophage capable of mediating generalizedtransduction have been reported in manyspecies of Bacillus including B. subtilis, B. cereus,B. megaterium, B. thuringiensis, B. anthracis, andB. stearothermophilus. Thus, transduction offersan immediate advantage for genetic analysisover transformation in that it is applicable tomore strains of these bacteria. In addition, somephages transduce fragments of DNA muchlarger than can be transferred via transforma-tion, and this facilitates linking distant markers.PBS1, a bacteriophage that infects B. subtilis168, can incorporate 5 to 10% of the bacterialchromosome in a single virion particle, and wasinstrumental in constructing the complete circu-lar chromosomal map of B. subtilis (Lepesant-Kejzlarova et al., 1975). On the other hand,small DNA molecules such as those of someplasmids are not efficiently packaged in largephages, but can be transduced by a variety ofsmall bacteriophages (Canosi et al., 1982). Somegeneralized transducing phages have relativelybroad host ranges and can transfer plasmidsbetween different species of bacilli (Ruhfel etal., 1984).
Little is known about the mechanism by whichtransducing particles are formed. PBS1 appearsto package bacterial DNA randomly. There islittle evidence for packaging sites as are indi-cated in Salmonella phage P22 (Jackson et al.,1978; Schmieger, 1982, 1984). A pac site has beenlocated in the genome of the small generalizedtransducing phage SPP1. However, its relevance,if any, to packaging transducing DNA is not clear(Deichelbohrer et al., 1982, 1985).
The process by which transducing DNAbecomes incorporated into the recipient bacte-rial chromosome differs from that discussed ear-lier for transformation. In transduction, thedonor DNA is thought to enter the infected cellas double-stranded DNA which then synapseswith the homologous region of the recipientchromosome. Incorporation of the transducingfragment presumably results from a double

CHAPTER 1.2.16 The Genus Bacillus—Nonmedical 545
crossover between the two DNA molecules. Insupport of this model, mutants of B. subtilis defi-cient in recombination functions have a reducedcapacity to be transduced, while they can betransformed (Dodson and Hadden, 1980).
Other important Bacillus generalized trans-ducing phages include CP15, a phage originallyshown to transduce B. cereus (Thorne, 1968) butwhich can transfer plasmids between B. cereus,B. thuringiensis, and B. anthracis (Ruhfel et al.,1984). Bacteriophage MP13 has been importantin mapping the chromosome of B. megaterium(Vary et al., 1982), and TP-13 and TP18 havebeen similarly used in genetic studies of B. thur-ingiensis (Barsomian et al., 1984).
Specialized TransductionSpecialized transduction has been reported inseveral B. subtilis phages including (105 (Sha-piro et al., 1974), SP& (Zahler et al., 1977), and(3T (Odebralski and Zahler, 1982) and B.amyloliquefaciens phage H2 (Zahler et al.,1987). The most carefully examined of these isSP&, a temperate bacteriophage that is normallycarried as a prophage in B. subtilis 168 (Warneret al., 1977). SP& can transduce markers proxi-mal to its normal attachment site attB SP&between the markers ilvA (threonine dehy-dratase) and kauA (ketoacid uptake) locatednear the terminus of chromosomal replication.In addition, SP& prophage will insert at a varietyof aberrant positions when it lysogenizesmutants of B. subtilis lacking attB SP&. Induc-tion of these lysogens gives rise to specializedtransducing phages carrying genetic markersnear the novel sites of integration (see Zahler,1982, for a review). The gene order of SP&prophage is a circular permutation of thatpresent in the phage DNA (Spancake et al.,1984). Specialized transducing particles arethought to originate from errors in excisionwhen the prophage is induced. The crossing-over event that leads to reformation of a circu-lar phage DNA during induction may be dis-placed, resulting in removal of a portion of thebacterial DNA contiguous to the right or leftboundaries of the prophage. Encapsidation ofsuch hybrid DNA produces a specialized trans-ducing particle.
Two types of transductants are recognized inthe SP& system (Zahler, 1982). The bacterialportion of the transducing phage DNA mayundergo recombination with and replace thehomologous region of the genome of the recipi-ent. This so-called “replacement transduction” issimilar to generalized transduction, except thatonly a limited number of genetic markers areinvolved. On the other hand, the infectingphage-bacterial DNA may incorporate as aprophage to produce a bacterium diploid for the
B. subtilis genes associated with the prophage.Such “addition transductions” are most likely tooccur if the recipient bacterium is alreadylysogenic for SP&. The resulting merozygotemay be used in complementation studies if dif-ferent alleles are present on the prophage andbacterial chromosomes.
Conjugative PlasmidsFertility plasmids capable of bringing abouttheir own transfer from one bacterium toanother have been described in several speciesof Bacillus. The capacity to produce the insecti-cidal delta toxin crystal protein in B. thuring-iensis is encoded in large plasmids. Gonzalez etal. (1982) found that three strains of this bacte-rium transmitted the crystal-protein phenotypeto B. thuringiensis variants which had lost theplasmid. Moreover, these plasmids could alsobe transferred to B. cereus and yielded tran-scipients that produce crystal protein. Battistiet al. (1985) reported the transfer of plasmidspXO11 and pXO12 from B. thuringiensis to B.anthracis and B. cereus. The transcipients, inturn, became effective donors, and in the caseof those inheriting pXO12, also acquired theability to produce parasporal crystals. Strains ofB. anthracis that acquire plasmid pXO12 cansubsequently mobilize and transfer nonconjuga-tive plasmids present in the same cell. Usingthis system, the tetracycline resistance plasmidpBC16, the B. anthracis toxin plasmid pXO1,and the capsule plasmid pXO2 have beentransmitted to B. anthracis and B. cereus recipi-ents lacking these plasmids (Green et al., 1989).The small plasmid pBC16 is transferred at highfrequency without direct interaction withpXO12; such transfer of a nonconjunctive plas-mid is called donation. The large B. anthracisplasmids are apparently transferred by conduc-tion. The latter involves formation of cointegra-tive molecules in the donor, and resolution ofthe cointegrates into pXO12 and the respectiveB. anthracis plasmid in the recipient. Cell-to-cell contact is necessary for plasmid transferand is resistant to DNase, but little is knownabout the mechanisms or conjugative structuresthat may be involved.
A strain of B. subtilis (natto) has been foundwhich carries a 55-kb self-transmissible plasmid(pLS20), which can be transferred to closelyrelated strains and to restriction-deficient strainsof B. subtilis (Koehler and Thorne, 1987). Thisplasmid also promotes the transfer of the tetra-cycline-resistance plasmid pBC16 from B. subti-lis (natto) to a wide variety of Bacillus speciesincluding B. anthracis, B. megaterium, and B.subtilis. This is a much broader range of conju-gative transmission than has been observed withthe B. thuringiensis plamid. However, none of

546 R.A. Slepecky and H.E. Hemphill CHAPTER 1.2.16
the conjugative plasmids have been found tomobilize and transfer chromosomal markers asis observed with the F plasmid of E. coli.
In addition to the naturally occurring trans-missible plasmids of Bacillus, Christie et al.(1987) have identified a conjugative transposon(Tn925) which transfers from Streptococcusfaecalis to B. subtilis.
Bacteriophages
Bacteriophages that infect Bacillus are commonin soil. In addition, many strains of this genus arenaturally lysogenic for one or more prophages.The most extensively studied Bacillus phages arethose associated with B. subtilis, and these havebeen reviewed by Hemphill and Whiteley (1975),Rutberg (1982), and Zahler (1988).
With some exceptions, Bacillus phages haverelatively narrow host ranges, probably at leastin part because of restriction systems that makephage grown in one host incompatible withanother strain (Ando et al., 1982). With theexception of the phages of B. subtilis, no schemeof classification has been adopted to organize thephages of this genus. Therefore, the bacterioph-ages described here will be grouped according tolife cycle.
Temperate BacteriophagesMost strains of bacilli that have been carefullyexamined have been found to release phage par-ticles. These are of two types: defective phagesthat can kill but do not productively infect otherstrains (see “Defective Bacteriophages,” thischapter), and those that grow on and lysogenizenew host bacteria. B. subtilis 168 is lysogenic forphage SP& and also releases defective phagePBSX. As an extreme, B. thuringiensis subsp.aizawai is polylysogenic for five unique temper-ate phages (Reynolds et al., 1988). Temperatephages are easily obtained from nature. If sam-ples of soil are placed in broth and the mixtureheated 10 min at 80°C, most free phage and non-sporeforming bacteria are destroyed. When theculture is allowed to incubate several hours and
subsequently treated with mitomycin C, temper-ate phages are induced and released into themedium. (Some investigators inoculate the brothwith the Bacillus strain of interest to enrich forphage, and then add mitomycin C.) The phagesmay then be recovered by filtering the solutionand plating on an appropriate indicator.
Dean et al. 1978 have divided the temperatephages of B. subtilis and closely related speciesinto four groups based upon serology, immunity,and physical characteristics (Table 9); the defec-tive phages may be considered a fifth class. Sev-eral group III phages including B. subtilis phagesSP& and (3T and B. amyloliquefaciens phage H2can mediate specialized transduction. Group Iphage (105 also transduces genes close to itsprophage attachment site (Shapiro et al., 1974).In addition, (105, SP&, )11, and others have beenused as cloning vehicles, mostly for B. subtilisgenes (see Zahler, 1988, for a review).
Temperate bacteriophages often alter the bio-chemistry or phenotype of lysogenic bacteria andseveral examples of such prophage conversionhave been observed in B. subtilis. Strains of thisbacterium lysogenic for SP& release a bacterio-cin-like substance called betacin (Hemphill et al.,1980) which kills some Bacillus strains that donot harbor the SP& prophage. Most group IIIbacteriophages including (3T, )11 and Z (but notSP&) contain the structural gene for thymidylatesynthetase, and express this gene continuously inlysogens. Stains of B. subtilis lysogenic for SP02cannot be productively infected with virulentphage (1 (Rettenmier and Hemphill, 1974), andbacilli lysogenic for SP& are protected by asimilar interference system active against (1m(Rettenmeir et al., 1979).
Defective BacteriophagesMany species of Bacillus including B. subtilis,B. amyloliquefaciens, B. pumilis, and B. lat-erosporus release defective phages whose pres-ence is revealed by their bactericidal activityagainst other strains or species of this genus(Hemphill and Whiteley, 1975; Steensma et al.,1978; Zahler, 1988.) For example, B. subtilisstrain 168 releases a defective phage called
Table 9. Major groups of temperate B. subtilis phages.
Modified from Dean et al. (1978) and Zahler (1988).
Group Example DNA size (kb)
Virion dimensions (nm)
Other members of groupHead Tail
I *105 40 52 " 52 10 " 220 )14II SPO2 40 50 " 50 10 " 180 —III SP& 126 72 " 82 12 " 358 *3T, )11, Z, SPRIV SP6 53 61 " 61 12 " 192 —V PBSX 13 45 " 45 20 " 200 PBSZ

CHAPTER 1.2.16 The Genus Bacillus—Nonmedical 547
PBSX, which kills the cells of B. subtilis strainW23. Electron microscopic examination of theculture supernatant of strain 168 reveals typicalphage particles. However, the PBSX virions can-not replicate and produce plaques; rather, theyact much like a bacteriocin. B. subtilis strainW23, in turn, releases defective phage PBSZ,which has a bacteriocin activity against strain168.
Despite being defective, these phages actmuch like other temperature viruses. Althoughsmall numbers are typically present in the cul-ture supernatant, lysogens may be induced withUV light or mitomycin C and lyse and releaselarge numbers of particles. (The situation is oftenconfusing, however, because many bacilli includ-ing B. subtilis 168 are simultaneously lysogenicfor one or more nondefective temperate phages.)Virions of PBSX are unmistakably phage-like instructure. They have a rather small head (45 nmin diameter) and a cylindrical, contractile tail 18by 200 nm. The genes for structural proteins ofPBSX have been mapped to a chromosomalposition between the markers metA and metC(Thurm and Garro, 1975). PBSX and presum-ably other phages of this group are defective formore than one reason. First, the particles do notcontain an identifiable phage DNA. Instead,PBSX packages randomly selected 13-kb frag-ments of bacterial chromosomal DNA (Ander-son and Bott, 1985). Second, although PBSXvirions attach efficiently to susceptible strainsand the tail appears to contract, the DNA in thephage heads is not injected (Okamoto et al.,1968).
Lytic BacteriophagesLytic bacteriophages that infect Bacillus are alsocommon in soil and water. Again, the mostdetailed studies are of phages that infect B. sub-tilis or closely related species (see Hemphill andWhiteley, 1975, for a review.) Although many ofthese viruses are intrinsically interesting, twogroups of subtilis phages have been the focus ofstudies on the regulation of viral transcription(see Geiduschek and Ito, 1982). The first class,which includes SP01 and SP82, is distinguishedby the replacement of thymine by hydroxymeth-yluracil (hmU) in the viral DNA. The secondclass includes (29 and relatives, which are verysmall, linear double-stranded DNA phages.
SP01 and SP82 have very complex temporalprograms of transcription involving at least threephases of gene expression designated early, mid-dle, and late. Early gene expression starts within1 min of infection, and is thought to use the hostRNA polymerase. Middle genes are activatedabout 4 min after infection, and the expressionof late genes (which are turned on asynchro-
nously) is first detected 10 min into the latentperiod. Initiation of transcription of the middleand late genes involves structural modificationsof the B. subtilis RNA polymerase, such that theenzyme can recognize unique middle and latephage promoters that are not used by theunmodified polymerase (Tarkington and Pero,1979). The product of at least one SP01/SP82gene is thought to associate with the enzyme toactivate middle genes (Hyde et al., 1986), and atleast two other phage-encoded polypeptidesappear to modify the polymerase to synthesizelate mRNAs (Fox, 1976; Tjian and Pero, 1976).Other viral genes associated with late transcrip-tion have been identified, but these may berequired to initiate DNA replication, which isalso involved in transcriptional control. Even asnew mRNAs are made, transcription of at leastsome early and middle genes is repressed at char-acteristic times in the latent period of SP82 andSP01. Proteins that bind to phage DNA andblock RNA synthesis have been isolated frominfected cells, but the actual mechanism ofrepression is not clear. Although not directlyrelated, the study of gene expression in thesesubtilis phages has influenced the investigationof somewhat analogous changes in RNA poly-merase structure associated with sporulation.
Transcription of the (29 genome also involvesearly and late mRNAs, which in this case aretranscribed from different strands of the viralDNA. Early RNA is transcribed from the light(L) and late RNA from the heavy (H) strand. Invitro, B. subtilis RNA polymerase synthesizesearly mRNA from as many as eight promoters(Mellado et al., 1988). At least a portion of thehost RNA polymerase is also used for synthesisof late RNA, which is apparently initiated froma single promoter called A3. The latter has a !35region that differs somewhat from the consensus!35 region of B. subtilis promoters (Vlcek andPaces, 1986). Transcription from A3 requiresprotein P4 encoded by (29 gene 4 (Mellado etal., 1988).
Pseudotemperate PhagesPseudotemperate bacteriophages are lyticphages that establish a relatively long-term asso-ciation with bacteria that mimics true lysogeny(see Hemphill and Whiteley, 1975). Followingadsorption and penetration, further advance-ment of the viral development cycle is oftendelayed; some infected cells continue to grow,divide, and even sporulate. A portion of thedaughter cells may even be cured, and aboutone-half the spores produced from such culturesdo not contain viral DNA. Pseudotemperatebacteriophages characteristically produce turbidplaques, and “pseudolysogens” obtained from

548 R.A. Slepecky and H.E. Hemphill CHAPTER 1.2.16
these plaques may be subcultured. The pseu-dolysogens apparently consist of a balancebetween infected cells in a delayed latent period,bacteria containing maturing phage, and unin-fected bacilli.
Included among the pseudotemperate phagesare several of the best-known transducingphages of Bacillus such as B. subtilis phagesPBS1, SP10, and SP15 and B. thuringiensis phageTP-13. The genomes of these viruses oftencontain modified DNA. For instance, thechromosome of PBS1 has a complete re-placement of thymine by uracil (Takahashi andMarmur, 1963) and SP15 DNA substitutes 5-(4#-5#-dihydroxylpentyl)-uracil for thymine (Bran-don et al., 1972).
Phage PBS1 (also referred to as PBS2, aclear-plaque mutant) and phage PMB12 carryout their entire development cycle in the pres-ence of rifampin, an antibiotic that inhibitsRNA synthesis by interacting with the bacterialRNA polymerase (Rima and Takahashi, 1974;Bramucci et al., 1977). In the case of PBS1(PBS2), the resistance to rifampin is explainedin part by the synthesis of a new phage-inducedRNA polymerase that transcribes the viral latefunctions (Clark et al., 1974). Genetic evidencesuggests that early RNA synthesis in PBS1 maybe resistant to the drug because the virus, per-haps using a virion protein, induces a modifica-tion in the bacterial RNA polymerase thatconverts the latter to a resistant form (Osburneand Sonenshein, 1980).
One of the most intriguing phenomenaassociated with bacteriophage PMB12 is itscapacity to convert pseudolysogens of certainsporulation-negative (spo) mutants of B. subti-lis to a Spo+ phenotype (Kinney and Bramucci,1981). Conversion has been observed in sev-eral stage O mutants, as well as in B. subtilisvariants deficient in sporulation due to changes
in bacterial RNA polymerase or 30S ribo-somes. At least three PMB12 genes areinvolved in spore conversion. The products ofthese genes apparently interact with host-cellpathways at the earliest stages of sporulation.Conversion of spore-negative mutants has alsobeen observed with B. pumilis phage PMB1(Bramucci et al., 1977) and numerous otherphages which infect this species (Keggins etal., 1978). Bacteriophage TP-13 similarly con-verts oligosporogenic, acrystalliferous mutantsof B. thuringiensis to spore-positive, crystal-positive phenotype (Perlak et al., 1979). Thisphage also mediates generalized transductionof large DNA fragments.
PlasmidsPlasmids are of widespread occurrence in thegenus Bacillus and have been found in most spe-cies that have been screened. The vast majorityare cryptic plasmids, that is, their presence hasnot been correlated with any unique property ofthe bacterial host. Although efforts have beenmade to analyze the relationship between dif-ferent plasmids in a few species by comparingDNA-restriction fragment profiles, there is noaccepted scheme for systematically classifyingplasmids within the bacilli. Further confusing thestudy of plasmids in Bacillus is the fact that someof those used as vectors for gene cloning, such aspUB110, were actually derived from Staphylo-coccus aureus (Gryczan et al., 1978).
As shown in Table 10, some of the Bacillusplasmids do confer a recognizable phenotype tocells which carry them, such as antibiotic re-sistance, synthesis of toxins, unique metabolicactivities, and transconjugation. Because of themedical and economic importance of their hosts,the plasmids conferring virulence in B. anthracis
Table 10. Bacillus plasmids.
Bacterium PlasmidDNA
size (kb) Phenotype associated with plasmid Reference
B. anthracis pXO1 168 Exotoxin (lethal factor, edema factor,protective antigen)
Mikesell et al., 1983Tippetts and Robertson, 1988
pXO2 85.6 Capsule Green et al., 1985B. cereus pBC7 69 Bacteriocin Bernhand et al., 1978
pBC16 4.3 Tetracycline resistance Bernhand et al., 1978B. pumilus pBL10 6.8 Bacteriocin Lovett et al., 1976B. subtilis pIM13 2.2 Erythromycin resistance Mahler and Halvorson, 1980B. subtilis (natto) pLS19 5.4 Polyglutamate production Hara et al., 1982
pLS20 55 Self-transmissible plasmid, which alsopromotes transfer of other plasmids
Koehler and Thorne, 1987
B. thuringiensis pXO12 112.5 Production of insecticidal crystal protein, andis a self-transmissible plasmid, which canco-transfer unrelated plasmids
Green et al., 1989
Bacillus species(thermophilic)
pTB19 26 Kanomycin and tetracycline resistance Imanaka et al., 1981pTB20 4.3 Tetracycline resistance Imanaka et al., 1981

CHAPTER 1.2.16 The Genus Bacillus—Nonmedical 549
and B. thuringiensis have been of increasinginterest. Virulence in B. anthracis requires theproduction of a capsule composed of poly-D-glutamic acid and an exotoxin consisting of threecomponents: protective antigen, lethal factor,and edema factor. The capacity to produce thecapsule is associated with plasmid pXO2 (Greenet al., 1985), and the constituents of the toxin areencoded in plasmid pXO1 (Mikesell et al., 1983).Strains losing either plasmid are rendered aviru-lent. With the help of fertility plasmid pXO12derived from B. thuringiensis, both pXO1 andpXO2 have been transferred to bacilli lackingthese plasmids (Green et al., 1989). In the caseof pXO1, cured strains of B. anthracis have beenrestored to toxin production (Thorne, 1985). Theassociation of the toxin with plasmids is of his-torical interest. The original Pasteur vaccinestrains of B. anthracis were produced by subcul-turing this bacterium at high temperatures whichare now known to inhibit replication of pXO1.Indeed, two existing Pasteur strains lack thisplasmid (Mikesell and Vodkin, 1985).
The genes for delta-exotoxin crystal proteinsof B. thuringiensis strains are carried in largeplasmids (see The Genus Bacillus—InsectPathogens in this Volume). These plasmids aresomewhat divergent in size and the crystal pro-teins produced by B. thuringiensis strains differin serology and toxicity to various insects (Höfteand Whiteley, 1989). Moreover, several of theseplasmids, such as pXO12 of B. thuringiensissubsp. thuringiensis 4042, (Reddy et al., 1987;Green et al., 1989) have been shown to mediateconjugal transfer of themselves and other plas-mids (see “Genetic Studies” this chapter).
The production of the parasporal body con-taining the crystal protein toxin is coordinatedwith sporulation; crystal protein gene expressionbegins about stage II (see Whiteley and Schnepf,1986, for a review). Changes in transcription dur-ing sporulation are associated with modificationsof RNA polymerase which enable the enzymeto recognize sporulation-specific promoters (see“Sporulation”, this chapter). It has been sug-gested the activation of the crystal protein genesis also regulated by polymerase changes. DNAsequence analysis of the 133-kDa crystal proteingene from B. thuringiensis subsp. kurstaki HD-1-Dipel revealed two overlapping promoters, oneused early in sporulation (BtI) and the other(BtII) activated midway through development(Wong et al., 1983). Comparison of these pro-moters with those of the carefully studied B.subtilis system show little sequence consensuswith promoters that bind vegetative RNA poly-merase. Brown and Whiteley (1988) haveisolated an RNA polymerase from strain HD-1-Dipel that directs transcription in vitro from theBtI promoter and also transcribes crystal protein
genes from two other strains of B. thuringiensis.This polymerase contains a unique 35-kDa sigmasubunit that is different from the major sigmasubunit present in vegetative cells of B. thuring-iensis. In addition, the presence of two regionsof hyphenated dyad symmetry near the BtI andBtII promoters suggests that binding of otherregulatory proteins in this region could be asso-ciated with gene expression.
Transcriptional Regulation in Biosynthetic and Catabolic Pathways
Bacillus species demonstrate great catabolic andbiosynthetic versatility. It is only recently, how-ever, that significant progress has been made inunderstanding how synthesis of enzymes is reg-ulated. Again, most studies have concentrated onB. subtilis and closely related species.
Control of Biosynthetic Gene ExpressionB. subtilis can be grown in minimal salts mediawith glucose as carbon source. Thus, this bacte-rium must have the capacity to synthesize denovo all amino acids, nucleotides, etc. As in theenteric bacteria, the structural genes for enzymesof common biosynthetic pathways are clusteredon the B. subtilis chromosome (see Zalkin andEbbole, 1988, for a review.) Such operon-likeorganization has been observed for the genes ofthe tryptophan, arginine, and isoleucine-valine-leucine synthesis. In addition, the sequencesencoding the enzymes for purine (pur genes) andpyrimidine (pyr genes) occur in multicistronclusters in B. subtilis (Ebbole and Zalkin, 1987;Lerner et al., 1987), which is not the case in E.coli. Gene clustering is associated with coordi-nate control of gene expression at the level oftranscription by the products of the pathways.The trp, arg, and ilv operons are repressed,respectively, by tryptophan, arginine, and leu-cine. The pur genes are repressed by adenine andguanine nucleotides and pyr by pyrimidines.
It does not appear that regulation of the bio-synthetic operons is mediated primarily throughrepressors, although the arg pathway may bean exception (Smith et al., 1986). Rather, thefavored means of control appears to be termina-tion/antitermination (attenuation) mechanismswhich affect the elongation of nascent mRNAmolecules. In several operons studied, the 5#-endof the mRNA contains a leader of 100 to 300nucleotides which does not encode part of thefirst structural gene. Within this leader is a regionthat can fold into two mutually exclusive second-ary structures, one of which is a procaryotetranscription termination signal. The other con-

550 R.A. Slepecky and H.E. Hemphill CHAPTER 1.2.16
figuration is an antiterminator stem-and-loop(hairpin) which allows full-length message to bemade. Control then resides in changing the ratioof termination vs. antitermination secondarystructure.
In enteric bacteria, the leader sequence ofamino acid biosynthetic gene clusters includes anopen reading frame (ORF) for a short peptide,which is generally enriched in codons for theamino acid synthesized by that pathway. The for-mation of termination vs. antitermination hair-pins is modulated by the rate at which the leaderpolypeptide is synthesized. In the E. coli trpoperon, for example, the peptide is made slowlyif intracellular levels of tryptophan are low, andthe antitermination structure is generated. Theconverse is true if tryptophan levels are high. InB. subtilis those leader sequences that have beenanalyzed do not contain an ORF. Moreover,transcription of pur and pyr gene clusters is alsothought to be controlled by termination/anti-termination, and it is unlikely these operonscould be regulated by a mechanism preciselyanalogous to the E. coli system.
In the B. subtilis trp operon, attenuation isregulated in part by a trans-acting factor thoughtto be the product of the methyltryptophan resis-tance locus mtr (Shimotsu et al., 1986). Muta-tions in this gene result in constitutive expressionof the trp genes. It is likely that the mtr product,when activated by tryptophan, binds to theleader region of the nascent mRNA in such amanner as to prevent formation of the antitermi-nator structure, thus favoring termination. Inaddition, binding of tryptophan-activated mtrgene product to already completed mRNA mol-ecules is thought to inhibit the initiation of trans-lation, thereby further reducing the productionof trp enzymes (Kuroda et al., 1988).
Transcriptional regulation of the pur operon ofB. subtilis shows some similarities to that of thetrp system, but may be even more complicated.Synthesis of inosine monophosphate (IMP) isrepressed by both adenine and guanine nucle-otides, but apparently by different mechanisms.The latter are thought to promote prematurecessation of transcription at a rho-independenttermination site in the leader sequence, whileadenine nucleotides appear to repress transcrip-tion initiation (Zalkin and Ebbole, 1988). It isthought likely that when purine levels are high,a guanine nucleotide activates a regulatory mol-ecule, which in turn binds to the leader region ofthe nascent mRNA and blocks formation of theantiterminator secondary structure.
The products of polycistronic mRNAs aresometimes subunits in a common enzyme com-plex. In these instances, the polypeptides arerequired in 1 : 1 stoichiometry. Sequence analysisof the trp and pur operons shows that both are
characterized by many gene overlaps; that is,the 3#-end of one coding sequence overlaps withthe 5#-end of a contiguous downstream codingsequence (Henner et al., 1984; Ebbole andZalkin, 1987). The overlaps are thought to resultin “translational coupling” in which synthesis ofone polypeptide from a polycistronic mRNA isat least partially dependent on the translation ofthe contiguous upstream gene. In some instances,this coupled translation is thought to result in1 : 1 stoichiometric synthesis of the products ofthe overlapping genes.
Control of Catabolic Gene ExpressionMembers of the family Bacillaceae catabolize awide variety of simple and complex organic com-pounds, including mono-and disaccharides, andpolysaccharides such as starch which are par-tially digested by extracellular enzymes. As in theenteric bacteria, the structural genes for enzymesinvolved in degradation of these substrates areoften clustered on the Bacillus chromosome. Alloperons so far studied are transcribed from$xA promoters, and control of gene expressioninvolves repressors and termination/antitermina-tion systems (see Klier and Rapport, 1988, for areview).
The pathway involved in gluconate catabolismin B. subtilis is apparently regulated by negativecontrol mediated by a repressor. Two inducibleenzymes are required: the transport protein glu-conate permease and gluconate kinase whichphosphorylates gluconate. The genes for thesefunctions, gntP and gntK, respectively, are partof a four cistron operon gntR-gntK-gntP-gntZ.Mutations that inactivate gntR are constitutive.This gene encodes a 243 amino acid proteinthought to inhibit transcription by binding nearthe promoter located 40 bp upstream from gntR(Fujita and Fujita, 1987). Curiously, there is a fivebase overlap between the coding sequence ofgntR and gntK suggesting there may be a trans-lational coupling between these two proteins(Fujita et al., 1986). The genes required for thegrowth of B. subtilis on xylose polymers are alsoorganized in a five cistron regulon, in which oneof the genes, xylR, codes for a repressor thatregulates expression of the entire gene cluster(Hastrup, 1988). Synthesis of the enzymes isinduced by the presence of xylose in the medium.A repressor system has also been found to regu-late the induction of penicillinase in B. licheni-formis (Wittman and Wong, 1988).
Perhaps the most extensively studied catabolicsystem in the bacilli is that involved in sucrosecatabolism. B. subtilis produces two &,D-fructofuranosidases, sucrase and levansucrase,after induction by sucrose. The former is an intra-cellular enzyme while the latter (which also cat-

CHAPTER 1.2.16 The Genus Bacillus—Nonmedical 551
alyzes the formation of the fructose polymerlevan) is secreted (see Klier and Rapoport,1988). The structural genes for sucrase, sacA, andthat for levansucrase, sacB, are widely separatedon the B. subtilis chromosome. The sacA gene istightly linked to sacP, which is thought to be amembrane-associated protein that transportsand phosphorylates sucrose. The sacB gene andthe sacA-sacP cistrons are linked to regulatoryloci, sacR and sacT, respectively. Mutations ineither of the regulatory loci results in constitu-tive synthesis of the corresponding enzyme. Themost extensive studies of the mechanisms of con-trol have been made in sacR.
The sacR locus contains the sacB promoter.Between the transcription start site and the sacBcoding sequence is a 199-bp region that is thetarget of several regulatory effectors of expres-sion of the levansucrase gene (Aymerich et al.,1986). Part of this control is thought to involvea region of dyad symmetry that could form atranscription termination signal in the nascentmRNA. Deletion of this perspective terminatorresults in constitutive synthesis of levansucrase,and previously isolated constitutive sacRmutants map to this putative stem-and-loopstructure (Shimotsu and Henner, 1986; Stein-metz and Aymerich, 1986). Messenger RNA syn-thesis is thought to stop at this terminator unlessan antiterminator protein, presumably regulatedby sucrose, interacts with the nascent RNA toprevent formation or to allow bypass of the loop.One candidate for the antiterminator maps tosacS, a genetic locus that affects both the synthe-sis of levansucrase and sucrase. The sacS locusconsists of two cistrons, sacY and sacX (Aymer-ich and Steinmetz, 1987; Steinmetz et al., 1988;Zukowski et al., 1988). The sacY gene is thoughtto encode a sucrose-dependent antiterminatorprotein whose target probably is the region ofsecondary structure in sacR, where it presumablyacts to prevent formation of the stem-and-loop.Deletion of sacY abolishes levansucrase synthe-sis. Current data suggest the sacX gene directsthe production of a negative regulator of sacY;mutants in this cistron produce levansucraseconstitutively.
The products of two other regulatory genes,sacU and sacQ, have been implicated in regula-tion of levansucrase synthesis. The target of theproducts of these genes also appears to be thesacR region, and they may serve to modulatethe levels of gene expression (Shimotsu andHenner, 1986; Zukowski and Miller, 1986). sacQencodes a 46 amino acid polypeptide whichappears to stimulate the transcription of severalsecreted enzymes. Some mutations in sacQ leadto higher levels of expression of target loci, andgenes thought to be similar to sacQ have beencloned from B. licheniformis and B. amylolique-
faciens. It now appears that the sacQ polypeptideis one of many small polypeptides which some-how regulate the transcription of degradativeenzymes in the family Bacillaceae. Others mayinclude the products of the sacV, sin, and ptrRgenes (see Klier and Rapoport, 1988).
The enzymes for the histidase pathway respon-sible for the degradation of histidine are encodedin a cluster of four genes hutH-hutU-hutI-hutGin B. subtilis. Expression of this hut operon isinduced by histidine and is also subject to catab-olite repression. Between the $A promoter andthe first gene, hutH, is a regulatory locus, hutR.The latter has been cloned and sequenced andfound to contain two regions of possible regula-tory significance (Oda et al., 1988). One is anORF (ORF1) for a 151 amino acid protein thatacts trans in heterozygotes to regulate the hutoperon. A mutant having an amino-acid sub-stitution in this protein has been isolated andfound to be uninducible for the histidase path-way enzymes. This suggests that the ORF1 pro-tein is a positive regulator in expression of thehut operon, although the possibility that themutant is a superrepressor cannot be entirelyexcluded. The second regulatory region locatedbetween ORF1 and the first structural gene hutH(ORF2) is a region of dyad symmetry whichcould form a rho-independent terminator. It ispossible that the product of ORF1, presumablyactivated by histidine, acts as an antiterminatorto allow complete transcription of the hutoperon.
The picture emerging from these and otherstudies suggests that regulation of catabolic path-ways in B. subtilis involves a combination ofrepressors and positive controls through antiter-mination. It should be noted that in the attenu-ation systems observed in the biosyntheticpathways the leader sequences appear to be ableto form two mutually exclusive secondary struc-tures, one of which is the terminator. In theseveral leader sequences studied in catabolicpathways, only the terminator stem-and-loop isindicated. Thus, the putative antiterminator pro-teins in catabolic systems probably act by block-ing formation of or allowing bypass of theterminator, rather than acting to favor one sec-ondary structure over the other.
Catabolite Repression
Catabolic pathways in Bacillus are subject tocatabolite repression by high levels of glucoseand a variety of other rapidly metabolized sub-strates (Nihashi and Fujita, 1984). This phenom-enon is of special interest in bacteria of thisgenus because sporulation is also repressed byglucose. In the enteric bacteria, catabolite

552 R.A. Slepecky and H.E. Hemphill CHAPTER 1.2.16
repression is mediated in part by the levels ofcAMP and the interaction of this nucleotide withcatabolite activator protein (CAP). However,cAMP apparently is not present in Bacillus, andthus could not be involved in catabolite repres-sion in these organisms.
The promoters and associated control regionsof many catabolic operons from Bacillus havebeen cloned and sequenced (Laoide et al., 1989;Melin et al., 1987; Oda et al., 1988). In someinstances, these cloned regulatory systems, whichfor experimental purposes are sometimes fusedto an indicator gene such as E. coli lacZ, con-tinue to be subject to glucose suppression oftranscription. Among the most thoroughly stud-ied is the complex amyR1-amyE, the regulatoryand structural genes for alpha-amylase of B.subtilis, and amyR1-amyL, the correspondingregulatory region and structural gene for alpha-amylase of B. licheniformis. Amylase synthesis isactually under two forms of regulation. One is atemporal control manifested by the fact thatamylase genes are not fully induced until theonset of stationary phase. Temporal control maynot occur in all Bacillus strains (Rothstein et al.,1986). In addition, in both B. subtilis and B.licheniformis FDO2 production of alpha-amylase is repressed about 10-fold in glucose-containing cultures in early stationary phase,even when multiple copies of the complex arepresent on a plasmid (Laoide et al., 1989). Bothforms of regulation are associated with the amyRregions. A plasmid-borne construct of amyLmissing the promoter but including all tran-scribed sequences immediately adjacent anddownstream from the promoter has been pro-duced. When this fragment is attached at variousdistances from heterologous promoters, tran-scription continues to be subject to cataboliterepression. This suggests the target of repressionis not in the promoter itself or in regionsupstream from it. One possibility is that a regu-latory protein involved in mediating cataboliterepression binds to a cis-acting site very close tothe transcription start site, which in the case ofamyL is only 29 to 31 nucleotides from the trans-lation initiation codon (121 nucleotides foramyE). A search for consensus sequences down-stream from several promoters of B. subtilisgenes subject to catabolite repression has yieldeda candidate sequence: 5#-ATTGTNA-3# (Laoideet al., 1989). In the regulatory loci for amyL andamyE, this sequence is contained within aninverted repeat sequence that overlaps the tran-scription start site, and in the case of amyL, thetranslation initiation codon. A point mutation(gra-10) in this region of dyad symmetry relievescatabolite repression of amyE (Nicholson et al.,1987). In addition, Weickert and Chambliss(1989) have reported that deletion of DNA 3# to
the amyR1 promoter does not impair temporalactivation of chloramphenicol acetyltransferasein amyR1-cat86 transcriptional fusions, but abol-ishes catabolite repression.
Mutants of B. subtilis that are resistant to atleast some manifestations of catabolite repres-sion have been isolated. Among mutants origi-nally selected for resistance to glucose-mediatedrepression of sporulation (Takahashi, 1979) iscrysA43, which maps to rpoD, the structural genefor $A (Price and Doi, 1985). The alteration in $A
in this variant also relieves catabolite repressionin an amyRi=lacZ fusion cloned in B. subtilis(Laoide and McConnell, 1989). This may suggesta connection between the mechanism of glucoserepression affecting sporulation and cataboliterepression of at least those genes activated instationary phase. In addition, Sun and Takahashi(1984) localized another catabolite-resistantmutation, crysE1, to the rpo operon. This locusencodes the & and &# subunits of RNA poly-merase. Taken together the two types of muta-tions occurring in subunits of RNA polymerasemay suggest this enzyme is directly involved incatabolite repression or interacts with a protein,perhaps bound to the regulatory region, whichmediates this phenomenon.
Resistance of Spores
The resting forms of bacteria are usually moreresistant to various environmental stresses thantheir counterpart vegetative forms. The structureand composition of the resting (and dormant)form of endosporeforming bacteria, however,are quite different from other bacterial-restingforms (see Fig. 3). The core or protoplast con-taining the heat labile DNA, RNA, ribosomes,enzymes, and other proteins is surrounded by aprimitive “germ cell wall.” Moving toward thesurface, there is a layer called the cortex whichconsists of peptidoglycan of a similar nature tothat of the vegetative cell wall but with less crosslinking in the peptides among other differences(Warth, 1978). A second cell membrane sur-rounds the cortex. The protoplast and cortex andtheir membranes are enclosed by layers of pro-tein coat. A loose-fitting exosporium, append-ages, and internal protein crystals may be foundin some species.
Compared with vegetative cells, spores aremore resistant to heat by a factor of 105 or more,to UV and ionizing radiation by 100-fold ormore, and to desiccation, antibiotics, disinfec-tants, and other chemicals (reviewed by Robertsand Hitchins, 1969; Russell, 1982; Gould andDring, 1974; Gould, 1983). Since spore resistancehas been recognized for a long time and is im-portant in food processing and sterilization

CHAPTER 1.2.16 The Genus Bacillus—Nonmedical 553
considerations, it has been extensively studiedparticularly with the genus Bacillus. Spore resis-tance to physical and chemical agents other thanheat and irradiation indicated above is also note-worthy. Vegetative cells are killed at 88,000 p.s.i.for 14 h hydrostatic pressure while their sporeshave been shown to require 176,000 p.s.i. forthe same time period (Gould and Dring, 1974).Spores are about 10,000 times more resistantto hyperchlorites than are vegetative cells.Two hundred roentgens (r) " 103 of x-rays wererequired to kill 50% of treated B. megateriumspores, while 50% of treated E. coli cells werekilled at 5.6 " 103 r. In every process designed forkilling microorganisms, the spores are more re-sistant than vegetative cells and with few excep-tions are the most resistant biological entitiesknown. One exception is the nonsporeformerDeinococcus radiodurans, the most radiation-resistant organism yet discovered. Its resistanceis presumably due to the possession of efficientrepair mechanisms for radiation-inducedchanges in DNA.
The degree of heat resistance has been shownto depend not only on the species but also on thephysiological environment in which the sporeswere formed. B. stearothermophilus spores aremore heat resistant than B. subtilis spores whichare more resistant than spores of B. megaterium(Roberts and Hitchins, 1969; Khoury et al.,1987). In addition, the spore resistance of eachspecies depends on the temperature at which itwas grown. For example, B. subtilis grown andsporulated at 20, 30, and 45°C produced sporeshaving D90 values (the time required to kill 90%of the spores at 90°C) of 37, 78, and 99 min,respectively (Khoury et al., 1987). The sporeswere more temperature sensitive when formedin ethanol-supplemented media. Since tempera-ture and ethanol are known to perturb thedegree of order within membranes, this suggeststhat alteration of membrane function is an addi-tional factor in the multifactorial nature of heatresistance. Other factors to consider in explain-ing heat resistance include protection of essentialspore macromolecules (Murrell, 1981; Lindsay etal., 1985; Gerhardt and Marquis, 1989); specificeffects of calcium dipicolinate (Lindsay andMurrell, 1986); mineralization (Marquis, 1989),and possibly foremost, dehydration (Gerhardtand Marquis, 1989). Current hypotheses on theheat resistance of endospores center on thedehydration of the protoplast (core) andthe expandable cortex with its counterions(Gould, 1983). In the heat-resistant form, thespore coat is relatively impermeable to multiva-lent cations. The cortex (of high water content)contains an expanded electronegative pepti-doglycan and mobile counterions exerting highosmotic pressure. The protoplast, of low water
content, is osmotically dehydrated by the sur-rounding cortex and is, therefore, heat resistant.In the heat-sensitive form, there is a modifiedcoat leaky to multivalent cations. The neutralizedcortex, collapsed and free of counterions, exertslow osmotic pressure. The protoplast becomespartly hydrated and, therefore, heat sensitive.The theory of heat resistance, called the osmo-regulatory expanded cortex theory, fits all theknown facts but has yet to be proven or dis-proven. Germinated spores lose their heat resis-tance and yet, under special conditions, can bedehydrated to become both heat resistant anddormant once again (Gould, 1983). It is notewor-thy that the heat resistance of some nonspore-formers can be increased in dehydrated cells. Theexpansion or contraction of the cortex is thoughtto account for the dehydrated state of the sporeprotoplast.
This reduced water content may also play arole in radiation resistance. It is thought that con-formational differences between DNA in sporeand vegetative cells may be associated with dif-ferences in hydration levels. The greater resis-tance of spores to UV radiation is also related torepair processes. The photoproduct formed dur-ing sporulation, 5-thyminyl-5,6-dehydrothymine(TDHT), is different from the thymine dimersformed in vegetative cells, both types of photo-products being deemed as the cause of death.Dark repair mechanisms in germinating sporesconvert TDHT to thymine. The greater sporeresistance to UV radiation is attributed to themore efficient removal of the unusual photo-product (reviewed by Russell, 1982). However,as with the mechanism of spore heat resistance,the picture is complex and many factors areinvolved. For example, dipicolinic acid has beensuggested as a protectant of spore DNA(Lindsay and Murrell, 1986) and it has beenshown that the SASP may be involved (Setlow,1988).
Likewise, explanations for ionizing radiationresistance of spores are not yet complete (Rus-sell, 1982). Different spore DNA conformation(possibly due to dehydration), possession of coatlayer radioprotectant substances, dipicolinic acidprotection, and more efficient repair systemshave all been implicated.
Spore coat layers, both coat protein and cor-tex, act as permeability barriers to toxic agents.At the present, this is the accepted explanationfor the greater spore chemical resistance andresistance to lytic agents (Russell, 1982).
Spores as Biological Indicators
Endospores are often used as biological indica-tors. Known numbers of spores of various Bacil-

554 R.A. Slepecky and H.E. Hemphill CHAPTER 1.2.16
lus species of predictable death rate can beplaced on various solid substrates (usually stripsof filter paper) and placed with the items to besterilized. The strips are then checked for reten-tion of viable organisms by immersion into cul-ture medium (various strategies are employed)after the sterilization process is completed ifgrowth occurs then survival of the spores hasoccurred and hence the procedure for processingwas inadequate. A D-value (decimal reductiontime or time to kill 90% of the population at aparticular temperature or treatment) of 3.0 minfor B. subtilis spores is required for ethyleneoxide sterilization (600 mg ethylene oxide perliter at 50% relative humidity and 54°C). Formoist heat (121°C) B. stearothermophilus sporeswith a D-value of 1.5 min and for dry heat(170°C) spores of B. subtilis with a D-value of0.8 min are preferred. B. pumilus spores (D-value of 0.17 Mrad) are used when ionizing radi-ation is used for sterilization (Korczynski, 1981).
Production of Antibiotics
Members of the genus Bacillus are capable ofproducing antibiotics as secondary metabolitesin the late logarithmic or early stationary phase
of growth of batch cultures. As many as 169 ofthese secondary metabolites have been re-corded; for example, various strains of B. subtilishave been shown to produce 68 antibiotics whileB. brevis can produce 23 (Katz and Demain,1977). A partial updated listing modified fromthat of Katz and Demain (1977) is presented(Table 11) to illustrate that there are otherBacillus antibiotic producers. Most of the antibi-otics are active against Gram-positive organisms,although there are exceptions. The majority arepeptide antibiotics but some belong to otherchemical classes (e.g., butirosin is an aminogly-coside and protocin is a phosphorus-containingtriene). Also indicated in Table 11 are thoseantibiotics whose structural genes have beenmapped and the few whose genes have beencloned. The latter types of studies are importantin their own right and serve as models for thestudy of expression of the genes of otherantibiotics.
A controversy has existed for some timeconcerning the function of these antibiotics.Since they usually appear upon the onset ofsporulation, it has been proposed that they maybe important factors in the transition of vegeta-tive cells to spores (reviewed by Katz andDemain, 1977). Support for this hypothesis that
Table 11. Some Bacillus antibiotics.
aAnti-Gram-positive bacteria.bInhibitor of fibrin clotting.cBroad spectrum antibiotic.dAnti-fungal antibiotic.eAnti-Gram-negative bacteria.
Species Antibiotic Genes mapped Genes cloned
B. subtilis Subtilina + +Surfactinb + !Bacilysina + !Difficidinc ! !Oxydifficidinc ! !Bacillomycin Fd ! !Mycobacillind ! !
B. brevis Gramicidin Sa + +Lincar Gramicidina ! !Tyrocidina + +
B. licheniformis Bacitracina + +Proticin ! !
B. pumilus Pumilina
TetainRemaining antibiotics have neither
been mapped nor cloned.B. mesentericus Esperina
B. polymyxa Polymyxine
Colistine
B. thiaminolyticus Octopytina
Baciphelacina
B. circulans Circuline
Butirosina
B. laterosporus Laterospuraminea
Laterosporina
B. cereus Biocerina
Cerexina

CHAPTER 1.2.16 The Genus Bacillus—Nonmedical 555
antibiotic production and sporulation may beregulated by the same or similar control mech-anisms is now available. Transcription of the B.subtilis tycA gene (coding for tyrocidin synthe-sis) is dependent on the products of certainStage 0 sporulation regulatory genes (Marahielet al., 1987). Thus it does appear that the twophysiological events are partially coupled toregulatory events occurring at the onset ofsporulation.
In addition, it has recently been demonstratedthat gramicidin S functions as an inhibitor ofoutgrowth after germination (Daher et al., 1985).
Protein Secretion
Members of the genus Bacillus are able tosecrete a wide variety of enzymes into the culturemedium (reviewed by Priest, 1977; Mezes andLampen, 1985). Every Bacillus species which hasbeen checked produces at least one extracellularenzyme. These include many different carbohy-drates, several kinds of proteases, penicillinases,nucleases, phosphatases, lipase, phospholipase C,thiaminase, and bacteriolytic enzymes. A vastliterature exists on the use of Bacillus enzymemodels for studying secretion mechanisms, cellu-lar location, and regulation. For some time, therehas been considerable interest in producing largequantities of enzymes for industrial purposes—proteases for detergent supplementation, thebrewing industry, various uses in the food indus-try and in leather manufacturing; and differentamylases for brewery use, in bread making, andin the paper industry (see review by Debabov,1982). Since many prokaryotic and eukaryoticgenes can be fused to B. subtilis-derived regula-tory regions and signal peptide sequences, suchgenetically manipulated organisms can be usedfor expressing and secreting many different het-erologous proteins (for molecular cloning in B.subtilis, see Gryczan, 1982; Ganeson et al., 1982;Mezes and Lampen, 1985; Ganeson and Hoch,1988). Interferon (Palva et al., 1983; Schien etal., 1986); human growth hormone (Honjo et al.,1986); and human interleukin-1 (Motley andGraham, 1988) are three such examples. How-ever, yields of such proteins can be low, becauseextracellular neutral protease, subtilisin,esterases, and other proteases may degrade thesecreted proteins. One strategy to overcome theproblem and stabilize proteins has been to usestrong glucose-insensitive promoters, proteasedeficient mutants, and catabolite repression ofsporulation (Wong et al., 1986).
The widespread interest in protein secretionby B. subtilis has stimulated many studies onthe genetics of secretion. There appears to be ageneral mechanism for regulating the synthesis
of extracellular proteins. The activity of degra-dative enzymes can be increased by mutationsat a number of loci that are unlinked to thestructural genes for the affected enzymes. Onefamily of genes, senN, sacU, prtR and hpr,codes for small regulatory proteins. For exam-ple, mutations at the sacU and sacQ loci canincrease the expression of levan sucrase, alka-line protease, neutral protease, xylanase, beta-gluconase, alpha-amylase, and intracellularserine protease (Henner et al., 1988a). Thistype of stimulation of degradative enzymes isthought to be a global regulatory system turnedon by a requirement for other carbon ornitrogen sources. The SacU product shareshomology with other two-component sensor-regulator systems (Ronson et al., 1987; Henneret al., 1988; Kunst et al., 1988).
Other Considerations
Just as Bacillus species have been important asmodels for studying differentiation or secretion,they have been employed extensively to studyother important biological problems. Theseinclude DNA replication (Winston and Sueoka,1982) and repair (Yasbin, 1985); chemotaxis(Ordal and Nettleton, 1985); genetic transfor-mation (Dubnau, 1982); and translation appa-ratus (Smith, 1982; Hager and Rabinowitz,1985).
Literature Cited
Anagnostopoulos, C. J. Spizizen, 1961. Requirements fortransformation in Bacillus subtilis. J. Bacteriol. 81:741–746.
Anderson, L. M. K. F. Bott, 1985. DNA packaging by theBacillus subtilis defective bacteriophage PBSX. J. Virol.54:773–780.
Ando, T. E. Hayase, S. Ikawa, T. Shibata, 1982. Site-specificrestriction endodeoxyribonucleases in Bacilli. 66–70. D.Schlessinger (ed.) Microbiology-1982. American Societyof Microbiology. Washington, D.C.
Aoki, H. R. A. Slepecky, 1973. Inducement of a heat-shockrequirement for germination and production ofincreased heat resistance in Bacilla fastidiosus spores bymanganous ins. J. Bacterial 114:137–143.
Aronson, A. I. P. C. Fitz-James, 1976. Structure and morpho-genesis of the bacterial spore coat. Bacteriol. Rev.40:360–402.
Aymerich, S. G. Gonzy-Treboul, M. Steinmetz, 1986. 5#-Noncoding region sacR is the target of all identifiedregulation affecting the levansucrase gene in Bacillussubtilis. J. Bacteriol. 166:993–998.
Aymerich, S. M. Steinmetz, 1987. Cloning and preliminarycharacterization of the sacS locus from Bacillus subtilis,which controls the regulation of the exoenzyme levan-sucrase. Mol. Gen. Genet. 208:114–120.
Barsomian, G. D. N. J. Robillard, C. B. Thorne, 1984. Chro-mosomal mapping of Bacillus thuringiensis by transduc-tion. J. Bacteriol. 157:746–750.

556 R.A. Slepecky and H.E. Hemphill CHAPTER 1.2.16
Bartlett, A. T. M. P. J. White, 1985. Species of Bacillus thatmake a vegetative peptidoglycan containing lysine lackdiaminopimelate epimerase but have diaminopimelatedehydrogenase. J. Gen. Microbiol. 131:2145–2152.
Battisti, L. B. D. Green, C. B. Thorne, 1985. Mating systemfor transfer of plasmids among Bacillus anthracis, Bacil-lus cereus, and Bacillus thuringiensis. J. Bacteriol.162:543–550.
Berkeley, R. C. W. M. Goodfellow (ed.), 1981. The Aerobicendosporeforming bacteria: classification and identifica-tion. Academic Press. London,
Bernhard, K. H. Schrempf, W. Goebel, 1978. Bacteriocin andantibiotic resistance plasmids in Bacillus cereus andBacillus subtilis. J. Bacteriol. 133:897–903.
Bott, K. F. G. A. Wilson, 1967. Development of competencein the Bacillus subtilis transformation system. J. Bacte-riol. 94:562–570.
Bowditch, R. D. P. Baumann, A. A. Yousten, 1989. Cloningand sequencing of the gene encoding a 125-Kilodaltonsurface-layer protein from Bacillus sphaericus 2362 andof a related cryptic gene. J. Bacteriol. 171:4178–4188.
Bramucci, M. G. K. Keggins, P. S. Lovett, 1977. Bacterio-phage conversion of spore-negative mutants to spore-positive in Bacillus pumilis. J. Virol. 22:194–202.
Brandon, C. P. M. Gallop, J. Marmur, H. Hayashi, N. Naka-nishi, 1972. Structure of a new pyrimidine from Bacillussubtilis phage SP-15 nucleic acid. Nature New Biol239:70–71.
Briehl, M. N. H. Mendelson, 1987. Helix hand fidelity inBacillus subtilis macrofibers after spheroplast regenera-tion. J. Bacteriol. 169:5838–5840.
Brown, K. L. H. R. Whiteley, 1988. Isolation of a Bacillusthuringiensis RNA polymerase capable of transcribingcrystal protein genes. Proc. Natl. Acad. Sci. USA.85:4166–4170.
Buchanan, C. E. 1987. Absence of penicillin-binding protein4 from an apparently normal strain of Bacillus subtilis.J. Bacteriol. 169:5301–5303.
Canosi, V. G. Luber, T. A. Trautner, 1982. SPP1-mediatedplasmid tranduction. J. Virol. 44:431–436.
Cassity, R. R. B. J. Kolodziej, 1984. Isolation partial charac-terization and utilization of a polysaccharide fromBacillus megaterum ATCC 19213. J. Gen. Microbiol.130:535–539.
Christie, P. J. R. Z. Korman, S. A. Zahler, J. C. Adsit, G. M.Dunny, 1987. Two conjugation systems associated withStreptococcus faecalis plasmid pCF10: identification of aconjugative transposon that transfers between S. faecalisand B. subtilis. J. Bacteriol. 169:2529–2536.
Clark, S. R. Losick, J. Pero, 1974. New RNA polymerase fromBacillus subtilis infected with phage PBS2. Nature252:21–24.
Claus, D. R. C. W. Berkeley, 1986. The genus Bacillus. 1105–1139. P. H. A. Sneath (ed.) Bergey’s manual of system-atic bacteriology, vol. 2. Williams and Wilkins.Baltimore.
Clausen, V. J. G. Jones, E. Stackebrandt, 1985. 16S ribosomalRNA analysis of Filibacter limicola indicates a closerelationship to the genus Bacillus. J. Gen. Microbiol.131:2659–2663.
Cowan, S. T. K. J. Steel, 1974. Manual for the identificationof medical bacteria, 2nd ed. Cambridge UniversityPress. London.
Daher, E. E. Rosenberg, A. L. Demain, 1985. Germination-initiated spores of Bacillus brevis nagano retain theirresistance properties. J. Bacteriol. 161:47–50.
Davidoff-Abelson, R. D. Dubnau, 1973a. Conditions affect-ing the isolation from transformed cells of Bacillussubtilis of high-molecular-weight single-stranded deox-yribonucleic acid of donor origin. J. Bacteriol. 116:146–153.
Davidoff-Abelson, R. D. Dubnau, 1973b. Kinetic analysis ofthe products of donor deoxyribonucleate in transformedcells of Bacillus subtilis. J. Bacteriol. 116:154–162.
Dean, D. H. C. L. Fort, J. A. Hoch, 1978. Characterization oftemperate phages of Bacillus subtilis. Curr. Microbiol.1:213–217.
Debabov, V. G. 1982. The industrial use of Bacilli. 331–370.D. A. Dubnau (ed.) The molecular biology of the bacilli.Academic Press. New York,
Diechelbohrer, I. J. C. Alonso, G. Luder, T. Trautner, 1985.Plasmid transduction by Bacillus subtilis bacteriophageSPP1: Effects of DNA homology between plasmid andbacteriophage. J. Bacteriol. 162:1238–1243.
Diechelbohrer, I. W. Messer, T. A. Trautner, 1982. Genomeof Bacillus subtilis bacteriophage SPP1: structure andnucleotide sequence of pac, the origin of DNA packag-ing. J. Virol. 42:83–90.
Dodson, L. A. C. T. Hadden, 1980. Capacity for postreplica-tion repair correlated with transducibility in Recmutants of Bacillus subtilis. J. Bacteriol. 144:608–615.
Doetsch, R. N. 1981. Determinative methods of light micros-copy. 21–33. P. Gerhart (ed.) Manual of methods forgeneral microbiology.. American Society for Microbiol-ogy. Washington, D.C.
Doi, R. H. L. F. Wang, 1986. Multiple procaryotic ribonucleicacid polymerase sigma factors. Microbiol. Rev. 50:227–243.
Dooley, D. C. C. T. Hadden, E. W. Nester, 1971. Macro-molecular synthesis in Bacillus subtilis during develop-ment of the competent state. J. Bacteriol. 108:668–679.
Doyle, R. J. J. Chaloupka, V. Vinter, 1988. Turnover of cellwalls in microorganisms. Microbiol. Rev. 52:554–567.
Dubnau, D. A. 1982. Genetic transformation in Bacillus sub-tilis. 148–178. D. A. Dubnau (ed.) The molecular biologyof the bacilli, vol. 1. Bacillus subtilis Academic Press.New York.
Dubnau, D. A. (ed.) 1982. The molecular biology of thebacilli, vol. 1. Bacillus subtilis Academic Press. NewYork.
Dubnau, D. A. (ed.) 1985. The molecular biology of thebacilli, vol. 2. Academic Press. New York.
Durham, D. R. D. B. Stewart, E. J. Stellwag, 1987. Novelalkaline- and heat-stable serine proteases from alkalo-philic Bacillus sp. Strain GX 6638. J. Bacteriol. 169:2762–2768.
Ebbole, D. J. H. Zalkin, 1987. Cloning and characterizationof a 12-gene cluster from Bacillus subtilis encoding nineenzymes for de novo purine nucleotide synthesis. J. Biol.Chem. 262:8274–8287.
Ephrati-Elizur, E. 1968. Spontaneous transformation inBacillus subtilis. Genet. Res. 11:83–96.
Fahmy, F. J. Flossdorf, D. Claus, 1985. The DNA base com-position of the type strains of the genus Bacillus. Syst.and Appl. Microbiol. 6:60–65.
Fitch, W. M. E. Margoliash, 1967. Construction of phyloge-netic trees: A method based on mutational distances asestimated from cytochrome c sequences is of generalapplicability. Science 155:279–284.
Fitz-James, P. C. E. Young, 1969. Morphology of sporulation.39–72. G. W. Gould and A. Hurst (ed.) The bacterialspore. Academic Press. New York.

CHAPTER 1.2.16 The Genus Bacillus—Nonmedical 557
Foster, S. J. K. Johnstone, 1989. The trigger mechanism ofbacterial germination. 89–108. I. Smith, R. A. Slepecky,and P. Setlow (ed.) Regulation of procaryotic develop-ment, structural and functional analysis of bacterialsporulation and germination.. American Society forMicrobiology. Washington, D.C.
Fox, G. E. K. R. Pechan, C. R. Woese, 1977. Comparativecataloging of 16s ribosomal ribonucleic acid: molecularapproach to prokaryotic systematics. Int. J. Syst. Bacte-riol. 27:44–57.
Fox, T. D. 1976. Identification of phage SP01 proteins codedby regulatory genes 33 and 34. Nature 262:748–753.
Freese, E. J. Heinze, 1983. Metabolic and genetic control ofbacterial sporulation. 101–172. A. Hurst and G. W.Gould (ed.) The bacterial spore, vol. 2. Academic Press.New York.
Freese, E. J. Heinze, T. Mitani, E. B. Freese, 1978. Limitationof nucleotides induces sporulations. 277–285. G. Cham-bliss and J. C. Vary (ed.) Spores VII.. American Societyfor Microbiology. Washington, D.C.
Fujita, Y. T. Fujita, 1987. The gluconate operon gnt of Bacil-lus subtilis encodes its own transcriptional negative reg-ulator. Proc. Natl. Acad. Sci. USA. 84:4524–4528.
Fujita, Y. T. Fujita, Y. Miwa, J. Nihashi, Y. Aratani, 1986.Organization and transcription of the gluconate operon,gnt, of Bacillus subtilis. J. Biol. Chem. 261:13744–13753.
Ganesan, A. T. S. Chang, J. A. Hoch (ed.), 1982. Molecularcloning and gene regulation in bacilli. Academic Press.New York.
Ganesan, A. T. J. A. Hoch (ed.), 1988. Genetics and biotech-nology of bacilli, vol. 2. Academic Press, New York..
Geiduschek, E. P. J. Ito, 1982. Regulatory mechanisms in thedevelopment of lytic bacteriophages in Bacillus subtilis.203–245. D. A. Dubnau (ed.) The molecular biology ofthe bacilli, vol. 1. Academic Press. New York.
Gerhardt, P. R. E. Marquis, 1989. Spore thermo-resistancemechanisms. 43–64. I. Smith, R. A. Slepecky, and P.Setlow (ed.) Regulation of procaryotic developmentstructural and functional analysis of bacterial sporela-tion and generation. Am. Soc. Microbiol. Washington,D.C.
Golden, K. J. R. W. Bernlohr, 1989. Defects in the nutrient-dependent methylation of a membrane-associate pro-tein in spo mutants of Bacillus subtilis. Mol. and Gen.Genet 220:1–7.
Gonzalez, J. M., Jr. B. J. Brown, B. C. Carlton, 1982. Transferof Bacillus thuringiensis plasmids coding for delta-endotoxin among strains of B. thuringiensis and B.cereus. Proc. Natl. Acad. Sci. USA. 79:6951–6955.
Gordon, R. E. 1981. One hundred and seven years of thegenus Bacillus. R. C. Berkeley, and M. Goodfellow (ed.)The aerobic endosporeforming bacteria. AcademicPress. London,
Gordon, R. E. W. C. Haynes, C. H.-N. Pang, 1973. The genusBacillus. Handbook No. 427. U.S. Department of Agri-culture. Washington, D.C.
Gould, G. W. 1983. Mechanisms of resistance and dormancy.173–209. A. Hurst and G. W. Gould (ed.) The bacterialspore, vol. 2. Academic Press. New York.
Gould, G. W. G. J. Dring, 1974. Mechanisms of spore heatresistance. Adv. Microbiol. Physiol. 2:137–161.
Graham, J. B. C. A. Istock, 1978. Genetic exchange in Bacil-lus subtilis in soil. Mol. Gen. Genet. 166:287–290.
Green, B. D. L. Battisti, T. M. Koehler, C. B. Thorne, B. E.Ivins, 1985. Demonstration of a capsule plasmid in Bacil-lus anthracis. Infect. Immun. 49:291–297.
Green, B. D. L. Battisti, C. B. Thorne, 1989. Involvement ofTn 4430 in transfer of Bacillus anthracis plasmids medi-ated by Bacillus thuringiensis plasmid pXO12. J. Bacte-riol. 171:104–113.
Gryczan, T. J. 1982. Molecular cloning in Bacillus subtilis.307–330. D. A. Dubnau (ed.) The molecular biology ofthe bacilli. vol. 1. Academic Press. New York.
Gryczan, T. J. S. Contente, D. Dubnau, 1978. Characteriza-tion of Staphylococcus aureus plasmids introduced bytransformation into Bacillus subtilis. J. Bacteriol.134:318–329.
Guettler, M. R. S. Hanson, 1988. Characterization of a meth-anol oxidizing member of the genus Bacillus. Abs. Ann.Mtg. Am. Soc. Microbiol I-95:196.
Hadden, C. E. W. Nester, 1968. Purification of competentcells in the Bacillus subtilis transformation system.J. Bacteriol. 95:876–885.
Hager, P. W. J. C. Rabinowitz, 1985. Translation specificity inBacillus subtilis. 1–32. D. A. Dubnau (ed.) The molecu-lar biology of the bacilli, vol. 2. Academic Press. NewYork.
Hara, T. A. Aumayr, Y. Fujio, S. Veda, 1982. Elimination ofplasmid-linked polyglutamate production by Bacillussubtilis (natto) with acridine orange. Appl. Environ.Microbiol 44:1456–1458.
Hastrup, S. 1988. Analysis of the Bacillus subtilis xylose reg-ulon. 79–83. A. T. Ganesan and J. A. Hoch (ed.) Geneticsand biotechnology of bacilli, vol. 2. Academic Press.New York.
Helmann, J. D. M. J. Chamberlain, 1988. Structure and func-tion of bacterial sigma factors. Ann. Rev. Biochem.57:839–872.
Hemphill, H. E. I. Gage, S. A. Zahler, R. Korman, 1980.Prophage-mediated production of a bacteriocinlikesubstance by Sp& lysogens of Bacillus subtilis. Can. J.Microbiol. 23:45–51.
Hemphill, H. E. H. R. Whiteley, 1975. Bacteriophages ofBacillus subtilis. Bacteriol. Rev. 39:257–315.
Henner, D. J. L. Band, H. Shimotsu, 1984. Nucleotidesequence of the Bacillus subtilis tryptophon operon.Gene 34:169–177.
Henner, D. J. E. Ferrari, M. Perego, J. A. Hoch, 1988a. Loca-tion of the targets of the hpr-97, sacU32(hy) and sacQ36(Hy) mutations in upstream regions to the subtilispromoter. J. Bacteriol. 170:296–300.
Henner, D. J. M. Yang, E. Ferrari, 1988b. Localization of B.subtilis sacU (Hy) mutations to two linked genes withsimilarities to the conserved procaryotic family of two-component signalling systems. J. Bacteriol. 170:5102–5109.
Hitchins, A. D. R. A. Slepecky, 1969. Bacterial sporulation asa modified procaryotic cell division. Nature London,223:804–807.
Hoch, J. A. K. Trach, I. Kawamura, H. Saito, 1985. Identifi-cation of the transcriptional suppressor SOF-1 as analteration in the SpoOA protein. J. Bacteriol. 161:552–555.
Höfte, H. H. R. Whiteley, 1989. Insecticidal crystal pro-teins of Bacillus thuringiensis. Microbiol. Rev. 53:242–255.
Honjo, M. A. Akaoka, A. Nakayama, Y. Furutani, 1986.Secretion of human growth hormone in B. subtilis usingprepropeptide coding region of B. amyloliquefaciensneutral protease gene. J. Biotechnol. 4:63–71.
Hunger, W. D. Claus, 1981. Taxonomic studies on Bacillusmegaterium and on agarolytic Bacillus strains. 217–239.

558 R.A. Slepecky and H.E. Hemphill CHAPTER 1.2.16
R. C. Berkeley and M. Goodfellow (ed.) The aerobicendosporeforming bacteria. Academic Press. London.
Hyde, E. I. M. D. Hilton, H. R. Whiteley, 1986. Interactionsof Bacillus subtilis RNA polymerase with subunitsdetermining the specificity of initiation. J. Biol. Chem.261:16565–16570.
Imanaka, T. M. Fujii, S. Aiba, 1981. Isolation and character-ization of antibiotic resistance plamids from thermo-philic bacilli and construction of deletion plasmids.J. Bacteriol. 146:1091–1097.
Iwasaki, H. A. Shimada, K. Yokoyama, E. Iyo, 1989. Struc-ture and glycosylation of lipoteichoic acids in Bacillusstrains. J. Bacteriol. 171:424–429.
Jackson, E. N. D. A. Jackson, R. J. Deans, 1978. Eco R1analysis of bacteriophage P22 DNA packaging. J. Mol.Biol. 118:365–388.
Johannes, P. M. de V. F. G. Boogerd, E. W. deV-deJong, 1986.Manganese reduction by a marine Bacillus species.J. Bacteriol. 167:30–34.
Jurtshuk, R. J. C. Lin, P. Candela, J. D. Wisotzkey, P. Jurtshuk,Jr, G. E. Fox, 1989. 16S Ribosomal RNA sequencingstudies on organisms of the Bacillus species. Abst. Ann.Meet. Am. Soc. Microbiol. R-10:281.
Katz, E. A. L. Demain, 1977. The peptide antibiotics of Bacil-lus: chemistry, biogenesis and possible functions. Bacte-riol. Rev. 41:449–474.
Kaya, S. K. Yokoyama, Y. Araki, E. Ito, 1984. N-acetylman-nos-aminyl (14) N-acetylglucosamine, a linkage unitbetween glycerol teichoic acid and peptidoglycan in cellwalls of several Bacillus strains. J. Bacteriol. 158:990–996.
Keggins, K. M. R. K. Nauman, P. S. Lovett, 1978. Spor-ulation-converting bacteriophages for Bacillus pumilis.J. Virol. 27:819–822.
Keynan, A. N. Sandler, 1983. Spore research in historicalperspective. 1–48. A. Hurst and G. W. Gould (ed.) Thebacterial spore, vol. 2. Academic Press. New York.
Khoury, P. H. S. L. Lombardi, R. A. Slepecky, 1987. Theperturbation of the heat resistance of bacterial spores bysporulation temperatures and ethanol. Current Micro-biol. 15:15–19.
Kinney, D. M. M. G. Bramucci, 1981. Analysis of Bacillussubtilis sporulation with spore-converting bacteriophagePMB12. J. Bacteriol. 145:1281–1285.
Kitada, M. K. Horikoshi, 1987. Bioenergetic properties ofalkalophilic Bacillus sp. strain C-59 on an alkalinemedium containing K2CO3. J. Bacteriol 169:5761–5765.
Kitada, M. K. Onda, K. Horikoshi, 1989. The sodium/protonantiport system in a newly isolated alkalophilic Bacillussp. J. Bacteriol. 171:1879–1884.
Klier, A. F. G. Rapoport, 1988. Genetics and regulation ofcarbohydrate catabolism in Bacillus. Ann. Rev. Micro-biol. 42:65–95.
Koehler, T. M. C. B. Thorne, 1987. Bacillus subtilis (natto)plasmid pLS20 mediates interspecies plasmid transport.J. Bacteriol. 169:5271–5278.
Korczynski, M. 1981. Sterilization. 476–486. P. Gerhardt (ed.)Manual of methods for general microbiology.. AmericanSociety of Microbiology. Washington, D.C.
Kunst, F. M. Debarbouille, T. Msadek, M. Young, C. Mauel,D. Karomata, A. Klier, G. Rapoport, R. Dedonder,1988. Deduced polypeptides encoded by the Bacillussubtilis sac U locus share homology with two-componentsensor-regulator systems. J. Bacteriol. 170:5093–5101.
Kuroda, M. I. D. Henner, C. Yanofsky, 1988. cis-Acting sitesin the transcript of the Bacillus subtilis trp operon regu-
late expression of the operon. J. Bacteriol. 170:3080–3088.
Lane, D. J. B. Pace, G. J. Olsen, D. A. Stahl, M. L. Sogin,N. R. Pace, 1985. Rapid determination of 16S ribosomalRNA sequences for phylogenetic analysis. Proc. Natl.Acad. Sci. USA. 82:6955–6959.
Laoide, B. G. H. Chambliss, D. J. McConnell, 1989. Bacilluslicheniformis %-amylase gene, amyL, is subject to pro-moter-independent calabotite repression in Bacillus sub-tilis. J. Bacteriol. 171:2435–2442.
Laoide, B. M. D. J. McConnell, 1989. cis Sequences involvedin modulating expression of Bacillus licheniformis amyLin Bacillus subtilis: effect of sporulation and cataboliterepression resistance mutations on expression. J. Bacte-riol. 171:2443–2450.
Lepesant-Kejzlarova, J. J.-A. Lepeseant, J. Walle, A. Billaut,R. Dedonder, 1975. Revision of the linkage map ofBacillus subtilis 168: indications for circularity of thechromosome. J. Bacteriol. 121:823–834.
Lerner, C. G. B. T. Stephenson, R. L. Switzer, 1987. Structureof the Bacillus subtilis pyrimidine biosynthetic (Pyr)gene cluster. J. Bacteriol. 169:2202–2206.
Lindsay, J. A. W. G. Murrell, 1986. Solution spectroscopy ofdipicolinic acid interaction with nucleic acids: role inspore heat resistance. Curr. Microbiol. 13:255–259.
Lindsay, J. A. W. G. Murrell, A. D. Warth, 1985. Spore resis-tance and the basic mechanism of heat-resistance. 162–186. L. E. Harris and A. J. Skopek (ed.) Sterilization ofmedical products, Vol. 3. Johnson & Johnson Pty., Ltd.Botany NSW Australia.
Logan, N. A. R. C. W. Berkeley, 1981. Classification andidentification of members of the genus Bacillus usingAPI tests. 105–140. R. C. W. Berkeley and M. Goodfel-low (ed.) The aerobic endosporeforming bacteria: clas-sification and identification.” Academic Press. London,
Logan, N. A. R. C. W. Berkeley, 1984. Identification of Bacil-lus strains using the API system. J. Gen. Microbiol.130:1871–1882.
Lorenz, M. G. B. W. Aardema, W. Wackernagel, 1988. Highlyefficient genetic transformation of Bacillus subtilisattached to sand grains. J. Gen. Micro. 134:107–112.
Losick, R. L. Kroos, 1989. Dependence pathways for theexpression of genes involved in endospore formation inBacillus subtilis. 223–242. I. Smith, R. A. Slepecky, andP. Setlow (ed.) Regulation of procaryotic development,structural and functional analysis on bacterial sporula-tion and germination.. American Society for Microbiol-ogy. Washington, D.C.
Losick, R. J. Pero, 1981. Cascades of sigma factors. Cell25:582–584.
Losick, R. L. Shapiro (ed.), 1984. Microbial development.Cold Spring Harbor Laboratory. NY.
Losick, R. P. Youngman, P. J. Piggot, 1986. Genetics ofendospore formation in Bacillus subtilis. Annv. Rev.Genet. 20:625–669.
Love, P. E. M. V. Lyle, R. E. Yasbin, 1985. DNA-damageinducible (din) loci are transcriptionally activated incompetent Bacilus subtilis. Proc. Natl. Acad. Sciences82:6201–6205.
Lovett, P. S. E. J. Duvall, K. M. Keggins, 1976. Bacillus pumi-lis plasmed pPL10: Properties and insertion into Bacillussubtilis 168 by transformation. J. Bacteriol 127:817–828.
Mahler, I. H. O. Halvorson, 1980. Two erythromycin resis-tance plasmids of diverse origin and their effect onsporulation in Bacillus subtilis. J. Gen. Microbiol.120:259–263.

CHAPTER 1.2.16 The Genus Bacillus—Nonmedical 559
Makino, S-II. Uchida, N. Terakado, C. Sasakawa, M.Yoshikawa, 1989. Molecular characterization and pro-tein analysis of the cap region which is essential forencapsulation in Bacillus anthracis. J. Bacteriol. 171:722–730.
Mandelstam, J. J. Errington, 1987. Dependent sequences ofgene expression controlling spore formation in Bacillussubtilis. Microbiol. Sciences 4:238–244.
Marahiel, M. A. P. Zuber, G. Czekay, R. Losick, 1987. Iden-tification of the promoter for a peptide antibiotic bio-synthesis gene from Bacillus brevis and its regulation inBacillus subtilis. J. Bacteriol. 169:2215–2222.
Marquis, R. E. 1989. Minerals and bacterial spores. 147–161.T. J. Beveridge and R. J. Doyle (ed.) Bacterial interac-tions with metal ions. J. Wiley and Sons. New York.
Melin, L. K. Magnusson, L. Rutberg, 1987. Identification ofthe promoter of the Bacillus subtilis sdh operon.J. Bacteriol. 169:3232–3236.
Mellado, R. P. I. Barthelemy, M. Salas, 1988. Transcriptioninitiation and termination signals of the Bacillus subtilisphage (29 DNA. 215–219. A. T. Ganesan and J. A. Hoch(ed.) Genetics and biotechnology of bacilli, vol. 2. Aca-demic Press. New York.
Mendelson, N. H. 1978. Helical Bacillus subtilis macrofibers:morphogenesis of a bacterial multicellular macroorgan-ism. Proc. Natl. Acad. Sci. USA. 75:2478–2482.
Mezes, P. S. J. O. Lampen, 1985. Secretion of proteins byBacilli. 151–185. D. A. Dubnau (ed.) The molecular biol-ogy of the bacilli, vol. 2. Academic Press. New York.
Mikesell, P. B. E. Ivins, J. D. Ristroph, T. M. Dreier, 1983.Evidence for plasmid-mediated toxin production inBacillus anthracis. Infect. Immun. 39:371–376.
Mikesell, P. M. Vodkin, 1985. Plasmids of Bacillus anthracis.52–55. L. Leive (ed.) Microbiology-1985.. AmericanSociety for Microbiology. Washington, D.C.
Minnikin, D. E. M. Goodfellow, 1981. Lipids in the class-ification of Bacillus and related taxa. 59–103. Berkeley,R. C. and M. Goodfellow (ed.) The aerobic endospore-forming bacteria.. Academic Press. London,
Moir, A. I. M. Feavers, A. R. Zuberi, 1986. A spore germi-nation operon in Bacillus subtilis 168. 183–194. A. T.Ganesan and J. A. Hoch (ed.) Bacillus molecular genet-ics and biotechnology applications.. Academic Press.London,
Moran, C. P. 1989. Sigma factors and the regulation of tran-scription. 167–184. I. Smith, R. A. Slepecky and P. Set-low (ed.) Regulation of prokaryotic development,structural and functional analysis of bacterial sporula-tion and germination. American Society for Microbiol-ogy. Washington, D.C.
Motley, S. T. S. Graham, 1988. Expression and secretion ofhuman interleukin-1 in Bacillus subtilis. 371–376. A. T.Ganeson and J. A. Hoch (ed.) Genetics and biotechnol-ogy of bacilli, vol. 2. Academic Press. New York.
Murrell, W. G. 1981. Biophysical studies on the molecularmechanisms of spore heat resistance and dormancy. 64–77. H. S. Levinson, A. L. Sonenshein, and D. J. Tupper(ed.) Sporulation and germination.. American Societyfor Microbiology. Washington, D.C.
Nakamura, L. K. 1984. Bacillus pulvifaciens sp. nov., nom.rev. Int. J. Syst. Bacteriol. 34:410–413.
Nakamura, L. K. 1987. Bacillus alginolyticus sp. nov. andBacillus chondritinus sp. nov. Int. J. Syst. Bacteriol.37:284–286.
Nakamura, L. K. 1989. Bacillus thiaminolyticus sp. nov., nom.rev. Abst. Annu. Mtg. Am. Soc. Microbiol. R-11:282.
Nakamura, L. K. I. Blumenstock, D. Claus, 1988. Taxonomicstudy of Bacillus coagulans Hammer 1915 with a pro-posal for Bacillus smithii sp. nov. Int. J. Syst. Bacteriol.38:63–73.
Nicholson, W. L. Y.-K. Paris, T. M. Henkin, M. Won, M. J.Weickert, J. A. Gaskell, G. H. Chambliss, 1987. Catabo-lite repression-resistant mutations of the Bacillus subtilisalpha-amylase promoter affect transcription levels andare in an operator-like sequence. J. Mol. Biol. 198:609–618.
Nihashi, J-I. Y. Fujita, 1984. Catabolite repression of inositoldehydrogenase and gluconate kinase synthesis in Bacil-lus subtilis. Biochimica et Biophysica Acta 798:88–95.
Nixon, B. T. C. W. Ronson, F. M. Ausubel, 1986. Two compo-nent regulating systems responsive to environmentalstimuli share strongly conserved domains with the nitro-gen assimilation regulatory genes ntrB and ntrC. Proc.Natl. Acad. Sci. USA. 83:7850–7854.
Norris, J. R. R. C. W. Berkeley, N. A. Logan, A. G. O’Donnell,1981. The genera Bacillus and Sporalactobacillus. 1711–1742. M. P. Starr, A. Stolp, A. G. Truper, A. Balows, andH. G. Schlegel (ed.) The prokaryotes, vol. 2. Springer-Verlag. Berlin,
Oda, M. A. Sogishita, K. Furukawa, 1988. Cloning and nucle-otide sequences of histidase and regulatory genes in theBacillus subtilis hut operon and positive regulation ofthe operon. J. Bacteriol 170:3199–3205.
Odebralski, J. M. S. A. Zahler, 1982. Specialized transductionof the kauA and citK genes of Bacillus subtilis by bacte-riophage (3T. Abstr. Am. Soc. Microbial. 130.
O’Donnell, A. G. H. J. H. Macfie, J. R. Norris, 1988. Anassessment of taxononic congruence between DNA-DNA hybridization and pyrolysis gas-liquid chromato-graphic classifications. J. Gen. Microbiol. 134:743–749.
O’Donnell, A. G. J. R. Norris, 1981. Pyrolysis gas-liquid chro-matographic studies in the genus Bacillus. 141–179. R.C. W. Berkeley and M. Goodfellow (ed.) The aerobicendosporeforming bacteria, classification and identifica-tion.. Academic Press. New York.
Okamoto, K. J. A. Mudd, J. Mangan, W. M. Huang, T. V.Subbaiah, J. Marmur, 1968. Properties of the defectivephage of Bacillus subtilis. J. Mol. Biol. 34:413–428.
Ordal, G. W. D. O. Nettleton, 1985. Chemotaxis in Bacillussubtilis. 53–73. D. A. Dubnau (ed.) The molecular biol-ogy of the bacilli, vol. 2. Academic Press. New York.
Osburne, M. A. L. Sonenshein, 1980. Inhibition by Lipiarmy-cin of bacteriophage growth in Bacillus subtilis. J. Virol.33:945–953.
Palva, I. P. Lehtovaara, L. Kaariainen, M. Sibakov, L. Can-tell, C. H. Schein, K. Kashiwagi, C. Weismann, 1983.Secretion of interferon by Bacillus subtilis. Gene 22:229–235.
Pechman, K. J. B. J. Lewis, C. R. Woese, 1976. Phylogeneticstatus of Sporosarcina ureae. Int. J. Syst. Bacteriol.261:305–310.
Pergo, M. S. P. Cole, D. Burbulys, K. Trach, J. A. Hoch, 1989.Characterization of the gene for a protein kinase whichphosphorylates the sporulation-regulatory rooteinsSpoOA and SpoOF of Bacillus subtilis. J. Bacteriol.171:6187–6196.
Perlak, F. J. C. L. Mendelsohn, C. B. Thorne, 1979. Convert-ing bacteriophage for sporulation and crystal-formationin Bacillus thuringiensis. J. Bacteriol. 140:699–706.
Pichinoty, F. J. Asselineau, M. Mandel, 1984. Characterisa-tion biochimique de Bacillus benzoevorans sp. nov., unenouvelle espèce filamenteuse, engainée et mesophile,

560 R.A. Slepecky and H.E. Hemphill CHAPTER 1.2.16
dégradant divers acides aromatiques et phenols. Ann.Microbiol. 135B:209–217.
Pichinoty, F. J. B. Waterbury, M. Mandel, J. Asselineau, 1986.Bacillus gordonae sp. nov., Une nouvelle espèce appar-tenant au second groupe morphologique, dégradantdivers composes aromatiques. Ann. Inst. Pasteur137A:65–78.
Piggot, P. 1989. Revised genetic map of B. subtilis 168. 1–42.I. Smith, R. A. Slepecky, and P. Setlow (ed.) Regulationof procaryotic development, structural and functionalanalysis of bacterial sporulation and germination.American Society for Microbiology. Washington, D.C.
Potvin, B. W. R. J. Kelleher, Jr., H. Gooder, 1975. Pyrimidinebiosynthelic pathway of Bacillus subtilis. J. Bacteriol.123:604–615.
Price, C. W. R. H. Doi, 1985. Genetic mapping of rpoD impli-cates the major sigma factor of Bacillus subtilis RNApolymerase in sporulation intiation. Mol. Gen. Genet.201:88–95.
Priest, F. G. 1977. Extracellular enzyme synthesis in the genusBacillus. Bacterol. Rev. 41:711–753.
Priest, F. G. 1981. DNA homology in the genus Bacillus. 33–57. R. C. Berkeley and M. Goodfellow (ed.) The aerobicendosporeforming bacteria. Academic Press. London,
Priest, F. G. M. Goodfellow, C. Todd, 1981. The genus Bacil-lus: A numerical analysis. 91–103. R. C. Berkeley and M.Goodfellow (ed.) The aerobic endosporeforming bacte-ria.. Academic Press. London,
Priest, F. G. M. Goodfellow, C. Todd, 1988. A numericalclassification of the genus Bacillus. J. Gen. Microbiol.134:1847–1882.
Reddy, A. L. Battisti, C. B. Thorne, 1987. Identification ofself-transmissible plasmids in four Bacillus thuringiensissubspecies. J. Bacteriol. 169:5263–5270.
Rettenmier, C. W. H. E. Hemphill, 1974. Abortive infectionof lysogenic Bacillus subtilis 168(SPO2) by bacterio-phage (1. J. Virol. 13:870–880.
Rettenmeir, C. W. B. Gingell, H. E. Hemphill, 1979. The roleof temperate bacteriophage SP& in prophage-mediatedinterference in Bacillus subtilis. Can. J. Microbiol.25:1345–1351.
Reynolds, R. B. A. Reddy, C. B. Thorne, 1988. Five uniquetemperate phages from a polylysogenic stain of Bacillusthuringiensis subsp. aizawai. J. Gen. Microbiol.134:1577–1585.
Rima, B. K. I. Takahashi, 1974. The synthesis of nucleic acidsin Bacillus subtilis infected with phage PBSI. Can. J.Biochem. 51:1219–1224.
Roberts, T. A. A. D. Hichins, 1969. Resistance of spores. 611–670. G. W. Gould and A. Hurst (ed.) The bacterial spore.Academic Press. London.
Ronson, C. W. B. T. Nixon, F. Ausubel, 1987. Conserveddomains in bacterial regulatory proteins that respond toenvironmental stimuli. Cell 49:579–581.
Rosson, A. R. K. H. Nealson, 1982. Manganese binding andoxidation by spores of a marine Bacillus. J. Bacteriol.151:1027–1034.
Rothstein, D. M. P. E. Devlin, R. L. Cate, 1986. Expressionof %-amylase in Bacillus licheniformis. J. Bacteriol168:839–842.
Ruhfel, R. E. N. J. Robillard, C. B. Thorne, 1984. Interspeciestransduction of plasmids among Bacillus anthracis, B.cereus, and B. thuringiensis. J. Bacteriol. 157:708–711.
Russell, A. D. 1982. The bacterial spore. 1–24. A. D. Russell(ed.) The destruction of bacterial spores. AcademicPress. London.
Rutberg, L. 1982. Temperate bacteriophages of Bacillus sub-tilis. 247–268. D. A. Dubnau (ed.) The molecular biologyof the bacilli, vol. 1. Academic Press. New York.
Saitou, N. M. Nei, 1987. The neighbor-joining method: a newmethod for reconstructing phylogenetic trees. Molec.Biol. and Evol. 4:406–425.
Schendel, F. J. C. E. Bremmon, M. G. Flickinger, M. Guettler,R. S. Hanson, 1989. L-Lysine production from methanolat high cell densities of MGA3, a thermophilic Bacillus.Abst. Annu. Mtg. Am. Soc. Microbiol. 316.
Schien, C. H. K. Kashiwagi, A. Fujisawa, C. Weissmann,1986. Secretion of mature IFN-%2 and accumulation ofuncleared precursor by Bacillus subtilis transformedwith a hybrid %-amylase signal sequence-IFN-%2 geneBio/Technology. 4:719–725.
Schmieger, H. 1982. Packaging signals for phage P22 on thechromosome of Salmonella typhimureim. Mol. Gen.Genet. 187:516–518.
Schmieger, H. 1984. Pac sites are indispensible for in vivopackaging of DNA by P22. Mol. Gen. Genet. 195:252–255.
Setlow, P. 1983. Germination and outgrowth. 211–254. A.Hurst, and G. W. Gould (ed.) The bacterial spore, vol. 2.Academic Press. London.
Setlow, P. 1988. Small, acid-soluble spore proteins of Bacillusspecies: Structure, synthesis, genetics, function and deg-radation. Annv. Rev. Microbiol. 42:319–338.
Setlow, P. 1989. Forespore specific genes of Bacillus subti-lis: function and regulation of expression. 211–222. I.Smith, R. A. Slepecky, and P. Setlow (ed.) Regula-tion of procaryotic development, structural and func-tional analysis of bacterial sporulation andgermination. American Society for Microbiology.Washington, D.C.
Shapiro, J. M. D. H. Dean, H. O. Halvorson, 1974. Low-frequency specialized transduction with Bacillus subtilisbacteriophage (105. Virology 62:393–403.
Shimotsu, H. D. J. Henner, 1986. Modulation of Bacillussubtilis levansucrase gene expression by sucrose and reg-ulation of the steady state mRNA level by sac V and sacQ genes. J. Bacteriol. 168:380–388.
Shimotsu, H. M. I. Kuroda, C. Yanofsky, D. J. Henner, 1986.Novel form of transcription attenuation regulatesexpression of the Bacillus subtilis tryptophan operon.J. Bacteriol. 166:461–471.
Shohayer, M. I. Chapra, 1985. Composition of membranesfrom whole cells and minicells of Bacillus subtilis. J. Gen.Microbiol. 131:345–354.
Shute, L. A. C. S. Gutteridge, J. R. Norris, R. C. W. Berkeley,1984. Curepoint pyrolysis mass spectrometry applied tocharacterization and identification of selected Bacillusspecies. J. Gen. Microbiol. 130:343–355.
Slepecky, R. A. 1972. Ecology of bacterial sporeformers. 297–313. H. G. Halvorson, R. Hanson, and L. L. Campbell(ed.) Spores V. American Society for Microbiology.Washington, D.C.
Slepecky, R. A. 1978. Resistant forms. 14/1–14/31. J. R. Norrisand M. H. Richmond (ed.) Essays in microbiology. JohnWiley & Sons. New York.
Slepecky, R. A. E. R. Leadbetter, 1977. The diversity ofspore-forming bacteria: some ecological implications.869–877. A. N. Barker, J. Wolf, D. J. Ellar, G. J. Dring,and G. W. Gould (ed.) Spore Research. 1976. AcademicPress. London.
Slepecky, R. A. E. R. Leadbetter, 1984. On the prevalenceand roles of sporeforming bacteria and their spores in

CHAPTER 1.2.16 The Genus Bacillus—Nonmedical 561
nature. 79–99. A. Hurst and G. W. Gould (ed.) The bac-terial spore, vol. 2. Academic Press. London.
Sleytr, U. B. P. Messner, 1988. Crystalline surface layers inprocaryotes. J. Bacteriol. 170:2891–2897.
Sloma, A. A. Ally, D. Ally, J. Pero, 1988. Gene encoding aminor extracellular protease in Bacillus subtilis. J. Bac-teriol. 170:5557–5563.
Smith, H. K. Wiersma, G. Venema, S. Bron, 1985. Transfor-mation in Bacillus subtilis: Further characterization of a75,000-dalton protein complex involved in binding andentry of donor DNA. J. Bacteriol. 164:201–206.
Smith, I. 1982. The translational apparatus of Bacillus subti-lis. 111–147. D. A. Dubnau (ed.) The molecular biologyof the bacilli, vol. 1. Academic Press. New York.
Smith, I. 1989. The initiation of sporulation. 185–210. I.Smith, R. A. Slepecky and P. Setlow (ed.) Regulation ofprocaryotic development, structural and functional anal-ysis of bacterial sporulation and germination. AmericanSociety for Microbiology. Washington, D.C.
Smith, M. C. M. AMountain, S. Baumberg, 1986. Sequenceanalysis of the Bacillus subtilis argC promoter region.Gene 49:53–60.
Smith, N. R. R. E. Gordon, F. E. Clark, 1946. Aerobic meso-philic sporeforming bacteria. US Dept. Agri. Misc. Pub-lication 559. Washington, D.C.
Smith, N. R. R. E. Gordon, F. E. Clark, 1952. Aerobic spore-forming bacteria. US Dept. Agr. Washington, D.C.
Spancake, G. A. H. E. Hemphill, P. S. Fink, 1984. Genomeorganization of SP&c2 bacteriophage carrying the thy P3gene. J. Bacterial. 157:428–434.
Sneath, P. H. A. R. R. Sokal, 1973. Numerical Taxonomy.W. H. Freeman. San Francisco.
Sonenshein, A. L. 1989. Metabolic regulation of sporulationand other stationary-phase phenomena. 109–130. I.Smith, R. A. Slepecky, and P Setlow (ed.) Regulation ofprocaryotic development, structural and functional anal-ysis of bacerial sporulation and germination. AmericanSociety Microbiology. Washington, D.C.
Spizizen, J. 1958. Transformation of biochemically deficientstrains of Bacillus subtilis by deoxyribonucleate. Proc.Natl. Acad. Sci. USA. 44:1072–1078.
Stackebrandt, E. C. R. Woese, 1979. A phylogenetic dissec-tion of the family Micrococcaceae. Curr. Microbiol.2:317–322.
Stackebrandt, E. C. R. Woese, 1981. The evolution ofprokaryotes. 1–31. M. J. Carlile, J. F. Collins, and B. E.B. Moseley (ed.) Molecular and cellular aspects ofmicrobial evolution.. Cambridge University Press.Cambridge.
Stackebrandt, E. W. Ludwig, M. Weizenegger, S. Dorn, T. J.McGill, G. E. Fox, C. R. Woese, W. Schubert, K-H.Schleifer, 1987. Comparative 16S RNA oligonucleotideanalyses and murein types of round-sporeforming bacilliand nonsporeforming relatives. J. Gen. Microbiol.133:2523–2529.
Steensma, H. Y. L. A. Robertson, J. D. Van Elsas, 1978. Theoccurrence and taxonomic value of PBSX-like defectivephages in the genus Bacillus. Antonie van Leeuwenhock44:353–366.
Steinmetz, M. S. Aymerich, 1986. Analysis genetique desacR, regulateur en cis de la synthese de la levane-saccharose de Bacillus subtilis. Ann. Microbiol. Paris,137A:3–14.
Steinmetz, M. S. Aymerich, G. Goney-Treboul, D. LeCoq,1988. Levansucrase induction by sucrose in Bacillussubtilis involves an antiterminator. Homology with the
Escherichia coli bgl operon. 11–15. A. T. Ganesan and J.A. Hock (ed.) Genetics and biotechnology of bacilli, vol.2. Academic Press. New York.
Stewart, G. J. C. A. Carlson, 1986. The biology of naturaltransformation. Ann. Rev. Microbiol. 40:211–35.
Stragier, P. 1989. Temporal and spatial control of geneexpression during sporulation: from facts to specula-tions. 243–254. I. Smith, R. A. Slepecky, and P. Setlow(ed.) Regulation of procaryotic development, structuraland functional analysis of bacterial sporulation and ger-mination.. American Society for Microbiology. Wash-ington, D.C.
Streips, U. N. N. E. Welker, 1971. Competence-inducing fac-tor of Bacillus stearothermophilus. J. Bacteriol. 106:955–959.
Sun, D. I. Takahashi, 1984. A catabolite-resistant mutation islocalized in the rpo operon of Bacillus subtilis. Can. J.Microbiol. 30:423–429.
Surana, U. A. J. Wolfe, N. H. Mendelson, 1988. Regulationof Bacillus subtilis macrofibe twist development by D-alanine. J. Bacteriol. 170:2328–2335.
Takahashi, I. 1979. Catabolite repression-resistant mutants ofBacillus subtilis. Can. J. Microbiol. 25:1283–1287.
Takahashi, I. J. Marmur, 1963. Replacement of thymidylicacid by deoxyuridylic acid in the deoxyribonucleic acidof a transducing phage for Bacillus subtilis. Nature197:794–795.
Tarkington, C. J. Pero, 1979. Distinctive nucleotide sequencesof promoters recognized by RNA polymerase contain-ing a phage-coded “$-like” protein. Proc. Nat. Acad. Sci.USA. 76:5465–5469.
teRiele, H. P. J. G. Venema, 1984. Heterospecific transforma-tion in Bacillus subtilis: protein composition of amembrane DNA complex containing a heterologousdonor-recipient complex. Mol. Gen. Genetics. 197:478–485.
Thorne, C. B. 1968. Transducing bacteriophage for Bacilluscereus. J. Virol. 2:657–682.
Thorne, C. B. 1985. Genetics of Bacillus anthracis. 56–62.L. Leive, P. F. Bonventure, J. A. Morello, S.Schlesinger, S. D. Silver, and H. C. Wu (ed.) Microbiol-ogy-1985. American Society for Microbiology, Washing-ton, D.C.
Thurm, P. A. J. Garro, 1975. Isolation and characterization ofprophage mutants of the defective Bacillus subtilis bac-teriophage PBSX. J. Virol. 16:184–191.
Tippetts, M. T. D. L. Robertson, 1988. Molecular cloning andexpression of the Bacillus anthrasis edema factor toxingene: a calmodulin-dependent adenylcyclase. J. Bacte-riol. 170:2263–2266.
Tjian, R. J. Pero, 1976. Bacteriophage SPO1 regulatory pro-teins directing late gene transcription in vitro. Nature262:753–757.
Trach, K. A. J. W. Chapman, P. J. Piggot, J. A. Hoch, 1985.Deduced product of the stage O sporulation gene spoOFshares homology with the SpoOA, OmpR and SfrA pro-teins. Proc. Natl. Acad. Sci. USA. 82:7260–7264.
Tsuboi, A. R. Uchihi, T. Adachi, T. Sasaki, S. Hayakawa, H.Yamagata, N. Tsukagoshi, S. Udaka, 1988. Characteriza-tion of the genes for the hexagonally arranged surfacelayer proteins in protein-producing Bacillus brevis 47:Complete nucleotide sequence of the middle wall pro-tein gene. J. Bacteriol. 170:935–945.
Vary, P. S. J. C. Garbe, M. Franzen, E. W. Frampton, 1982.MP13, a generalized transducing bacteriophage forBacillus megaterium. J. Bacteriol. 149:112–119.

562 R.A. Slepecky and H.E. Hemphill CHAPTER 1.2.16
Vinter, V. R. A. Slepecky, 1965. Direct transition of outgrow-ing bacterial spores to new sporangia without interme-diate cell division. J. Bacteriol. 90:803–807.
Vlcek, C. V. Paces, 1986. Nucleotide sequence of the lateregion of Bacillus phage (29 complete the 19285-bpsequence of (29 genome. Comparison with the homolo-gous sequence of phage PZA. Gene 46:215–225.
Warner, F. D. G. A. Kitos, M. P. Romano, H. E. Hemphill,1977. Characterization of SP&: a temperate bacterioph-age from Bacillus subtilis 168M. Can. J. Microbiol.23:45–51.
Warth, A. D. 1978. Molecular structure of the bacterial spore.Adv. Microb. Physiol. 17:1–45.
Weickert, M. J. G. H. Chambliss, 1989. Genetic analysis ofthe promoter region of the Bacillus subtilis %-amylasegene. J. Bacteriol. 171:3656–3666.
Whiteley, H. R. H. E. Schnepf, 1986. The molecular biologyof parasporal crystal body formation in Bacillus thuring-iensis. Ann. Rev. Microbiol. 40:549–576.
Winston, S. N. Sueoka, 1982. DNA replication in Bacillussubtilis. 36–71. D. A. Dubnau (ed.) The molecular biol-ogy of the bacilli, vol. 1. Academic Press. New York.
Wisotzkey, J. D. P. Jurtshuk, Jr., G. E. Fox, 1989. Comparative16S rRNA analyses on thermophilic and psychrophilicbacillus species. Abst. Ann. Meet. Am. Soc. Microbiol.281.
Wittman, V. H. C. Wong, 1988. Regulation of the penicilli-nase genes of Bacillus licheniformis: Interaction of thepen repressor with its operators. J. Bacteriol. 170:3206–3212.
Wolf, J. A. N. Barker, 1968. The genus Bacillus: aids to theidentification of its species. 93–109. M. Gibbs and D. A.Shapton (ed.) Identification methods for microbiolo-gists. Part B. Academic Press. London,
Wong, H. C. H. E. Schnepf, H. R. Whiteley, 1983. Transcrip-tional and translational start sites for the Bacillus thur-ingiensis crystal protein gene. J. Biol. Chem. 258:1960–67.
Wong, S. F. Kawamura, R. H. Doi, 1986. Use of Bacillussubtilis subtilisin signal peptide for efficient secretion ofTEMB-lactamase during growth. J. Bacteriol. 168:1005–1009.
Yasbin, R. E. 1985. DNA repair in Bacillus subtilis. 33–52.Dubnau (ed.) The molecular biology of the bacilli, vol.2. Academic Press. New York.
Yasbin, R. E. P. I. Fields, B. J. Andersen, 1980. Properties ofBacillus subtilis 168 derivatives freed of their naturalprophages. Gene 121:155–159.
Yasbin, R. E. G. A. Wilson, F. E. Young, 1975. Transforma-tion and transfection in lysogenic strains of Bacillus sub-tilis: evidence for selective induction of prophage incompetent cells. J. Bacteriol. 121:296–304.
Youngman, P. H. Poth, B. Green, K. York, G. Olmedo, K.Smith, 1989. Methods for genetic manipulation, cloningand functional analysis of sporulation genes in Bacillussubtilis. 65–88. I. Smith. R. A. Slepecky, and P. Setlow(ed.) Regulation of procaryotic development, structuraland functional analysis of bacterial sporulation and ger-mination. American Society for Microbiology. Washing-ton, D.C.
Zahler, S. A. 1982. Specialized transduction in Bacillussubtilis. 269–305. D. A. Dubnau (ed.) The molecularbiology of the bacilli, vol. 1. Academic Press. NewYork.
Zahler, S. A. 1988. Temperate bacteriophages of Bacillussubtilis. 559–592. R. Calendar (ed.) The bacteriophages,vol. 1. Plenum Press. New York.
Zahler, S. A. R. Z. Korman, R. Rosenthal, H. E. Hemphill,1977. Bacillus subtilis bacteriophage SP&: localization ofthe prophage attachment site and specialized transduc-tion. J. Bacteriol 129:556–558.
Zahler, S. A. R. Z. Korman, C. Thomas, P. S. Fink, M. P.Weiner, J. M. Odebralski, 1987. H2, a temperate bacte-riophage isolated from Bacillus amyloliquefaciens strainH. J. Gen. Microbiol. 133:2937–2944.
Zalkin, H. D. J. Ebbole, 1988. Organization and regulationof gene encoding biosynthetic enzymes in Bacillus sub-tilis. J. Biol. Chem. 263:1595–1598.
Zarilla, K. A. J. J. Perry, 1987. Bacillus thermoleovorans,sp.nov., a species of obligately thermophilic hydrocarbonutilizing endosporeforming bacteria. System. App.Microbiol. 9:258–264.
Zukowski, M. M. L. Miller, 1986. Hyperproduction of anintracellular heterologous protein in a sac Uh mutant ofBacillus subtilis. Gene 46:247–55.
Zukowski, M. L. Miller, P. Cogswell, K. Chen, 1988. Induc-ible expression system based on sucrose metabolismgenes of Bacillus subtilis. 17–22. A. T. Ganesan and J. A.Hoch (ed.) Genetics and biotechology of bacilli, vol. 2.Academic Press. New York.


















