
Harpactorini Phylogeny
14
Molecular phylogeny of Harpactorini (Insecta: Reduviidae): correlation of novel predation strategy with accelerated evolution of predatory leg morphology Guanyang Zhang a, * ,† and Christiane Weirauch a a Department of Entomology, University of California, Riverside, CA, 92521, USA; † Present address: School of Life Sciences, Arizona State University, Tempe, AZ, 85281, USA Accepted 5 July 2013 Abstract Much research and discussion have focused on the effects of key innovations on lineage diversification, whereas little has been done to investigate their role in morphological evolution using phylogenetic approaches. Here we present the first comprehensive molecular phylogeny of the Harpactorini (Insecta: Reduviidae), the largest assassin bug tribe, sampling 229 terminal taxa and using five gene segments (28S D2, D3–D5, 16S, COI, and Deformed). Employing comparative phylogenetic methods, we demon- strate the correlation of a putative key innovation, the sticky trap predation strategy, with accelerated rates of morphological evolution of the predatory fore leg in assassin bugs. We show that bugs exhibiting sticky trap predation have evolved more slen- der and longer fore femora than non-sticky bugs. Using phylogenetically independent contrast analyses, we document correlated evolution between femoral thickness and length. We argue that the novel sticky trap predation strategy may allow sticky bugs to alleviate functional constraints on the fore femur and thus to attain a higher rate of evolution than other Harpactorini or Redu- viidae. We discuss the possibility that sticky bugs represent a case of adaptive radiation. We also test historical supra-generic groups within the Harpactorini, and show that most of them are not monophyletic. We confirm the paraphyly of Harpactorini with respect to Rhaphidosomini. © The Willi Hennig Society 2013. The term “key innovation” refers to a character that is considered to be critically linked to the origin of a higher taxon or niche, lineage diversification, or adap- tive radiation (reviewed by Hunter, 1998; Yoder et al., 2010). The connection of key innovations to these phe- nomena of biological diversifications has been, and continues to be, the subject of extensive theoretical dis- cussion and numerous empirical or comparative phylo- genetic studies (Cracraft, 1990; Slowinski and Guyer, 1993; Bond and Opell, 1998; Ree, 2005; Alfaro et al., 2009a; Moore and Donoghue, 2009; Drummond et al., 2012; Erkens et al., 2012). However, relatively little attention has been paid to the role of key innovations in phenotypic or morphological diversification. Fur- thermore, as discussed by Wainwright (2007) and Price et al. (2010), most studies and discussions on this topic have not utilized explicit phylogenetic approaches, nor have they measured phenotypic or morphological diversity directly (Thomas et al., 2006; Dornburg et al., 2011). Besides, a taxonomic bias towards verte- brates is evident in the literature. In contrast to key innovations, a number of recent comparative phyloge- netic studies have focused on the correlation of extrin- sic factors, such as “ecological opportunity” and “habitat use”, with rates of morphological diversifica- tion (Harmon et al., 2008; Collar et al., 2009, 2010, 2011; Moen and Wiens, 2009; Hulsey et al., 2010; Mahler et al., 2010; Derryberry et al., 2011; Martin and Wainwright, 2011; Price et al., 2011). Taking an explicit phylogenetic approach, the current study explores the relationship between a potential key *Corresponding author: E-mail address: [email protected] Cladistics Cladistics 30 (2014) 339–351 10.1111/cla.12049 © The Willi Hennig Society 2013
-
Upload
darius-trujillo -
Category
Documents
-
view
36 -
download
10
description
Phylogenetic tree of tribe Harpactorini (Reduviidae)
Transcript of Harpactorini Phylogeny

Molecular phylogeny of Harpactorini (Insecta: Reduviidae):correlation of novel predation strategy with accelerated evolution
of predatory leg morphology
Guanyang Zhanga,*,† and Christiane Weiraucha
aDepartment of Entomology, University of California, Riverside, CA, 92521, USA; †Present address: School of Life Sciences, Arizona State
University, Tempe, AZ, 85281, USA
Accepted 5 July 2013
Abstract
Much research and discussion have focused on the effects of key innovations on lineage diversification, whereas little has beendone to investigate their role in morphological evolution using phylogenetic approaches. Here we present the first comprehensivemolecular phylogeny of the Harpactorini (Insecta: Reduviidae), the largest assassin bug tribe, sampling 229 terminal taxa andusing five gene segments (28S D2, D3–D5, 16S, COI, and Deformed). Employing comparative phylogenetic methods, we demon-strate the correlation of a putative key innovation, the sticky trap predation strategy, with accelerated rates of morphologicalevolution of the predatory fore leg in assassin bugs. We show that bugs exhibiting sticky trap predation have evolved more slen-der and longer fore femora than non-sticky bugs. Using phylogenetically independent contrast analyses, we document correlatedevolution between femoral thickness and length. We argue that the novel sticky trap predation strategy may allow sticky bugs toalleviate functional constraints on the fore femur and thus to attain a higher rate of evolution than other Harpactorini or Redu-viidae. We discuss the possibility that sticky bugs represent a case of adaptive radiation. We also test historical supra-genericgroups within the Harpactorini, and show that most of them are not monophyletic. We confirm the paraphyly of Harpactoriniwith respect to Rhaphidosomini.© The Willi Hennig Society 2013.
The term “key innovation” refers to a character thatis considered to be critically linked to the origin of ahigher taxon or niche, lineage diversification, or adap-tive radiation (reviewed by Hunter, 1998; Yoder et al.,2010). The connection of key innovations to these phe-nomena of biological diversifications has been, andcontinues to be, the subject of extensive theoretical dis-cussion and numerous empirical or comparative phylo-genetic studies (Cracraft, 1990; Slowinski and Guyer,1993; Bond and Opell, 1998; Ree, 2005; Alfaro et al.,2009a; Moore and Donoghue, 2009; Drummond et al.,2012; Erkens et al., 2012). However, relatively littleattention has been paid to the role of key innovationsin phenotypic or morphological diversification. Fur-
thermore, as discussed by Wainwright (2007) and Priceet al. (2010), most studies and discussions on this topichave not utilized explicit phylogenetic approaches, norhave they measured phenotypic or morphologicaldiversity directly (Thomas et al., 2006; Dornburget al., 2011). Besides, a taxonomic bias towards verte-brates is evident in the literature. In contrast to keyinnovations, a number of recent comparative phyloge-netic studies have focused on the correlation of extrin-sic factors, such as “ecological opportunity” and“habitat use”, with rates of morphological diversifica-tion (Harmon et al., 2008; Collar et al., 2009, 2010,2011; Moen and Wiens, 2009; Hulsey et al., 2010;Mahler et al., 2010; Derryberry et al., 2011; Martinand Wainwright, 2011; Price et al., 2011). Taking anexplicit phylogenetic approach, the current studyexplores the relationship between a potential key
*Corresponding author:E-mail address: [email protected]
CladisticsCladistics 30 (2014) 339–351
10.1111/cla.12049
© The Willi Hennig Society 2013

innovation, the novel sticky trap predation strategy,and rates of evolution of the predatory fore leg inassassin bugs (Insecta: Hemiptera: Reduviidae).One of the mechanisms by which a key innovation
provides the potential for morphological diversificationis by breaking existing constraints and therefore facili-tating increased evolutionary change (Galis, 2001; Priceet al., 2010). For instance, powered flight as a novellocomotory system in birds has eased the constraint ofmaintaining running proficiency on the hind limbs,resulting in greater morphological disparities comparedwith those of non-avian bipedal theropods (Gatesy andMiddleton, 1997). Price et al. (2010) demonstratedeight times faster rates of jaw diversification in parrotfishes that possess two “design breakthroughs”: the in-tramandibular joint modifications and the pharyngealjaw. They argued that these innovations reduce con-straints on the structural elements of the jaw byincreasing the degree of freedom of the jaw morpho-space. Evidence for the opposite phenomenon—limitedmorphological diversification due to the conservationof functional constraints—also exists. For example,Cooper and Steppan (2010) showed that in marsupialmammals, functional requirements of the natal climblimit the evolution of forelimb shape diversity.Based on comparative morphological studies, a
novel predation strategy called “sticky trap predation”has been previously documented for 12 New Worldgenera in the assassin bug tribe Harpactorini (Hemip-tera: Heteroptera: Reduviidae: Harpactorinae; Zhangand Weirauch, 2013). Members of these genera, called“sticky bugs”, are characterized by the presence ofunique dermal glands—“sticky glands”—on the legs,which secretes a sticky substance (Barth, 1952;Ambrose, 1999; Weirauch, 2006). The sticky glandscan be broadly categorized into two types, one withelongate or pear-shaped saccules and another withrounded saccules, as exemplified by the conditions inZelus Fabricius and Heza Amyot and Serville (Zhangand Weirauch, 2013), respectively. The former hasbeen found in the majority of sticky bugs and the lat-ter only from species of Heza. Sticky bugs also possessone of two kinds of modified setae on the legs: “sun-dew setae” that have projections resembling trichomeson the leaves of sundews (Drosera Linnaeus), or“lamellate setae” that have undulate lamellae alongthe setal body (Barth, 1952; Wolf and Reid, 2001;Weirauch, 2006; Zhang and Weirauch, 2013). Similarto the two types of sticky glands, sundew setae havebeen observed in most of the sticky-bug genera, and
lamellate setae only from species of Heza. The sundewsetae are usually covered with sticky secretions andtheir presence probably increases the surface and reachof the sticky fore leg. However, examination of lamel-late setae did not reveal a conspicuous presence ofsticky material on the legs. Zhang and Weirauch(2013) speculated that the morphological conditions ofsticky glands and modified setae in Heza may be aresult of convergent evolution. In the present study,we investigate the origin(s) of sticky glands and themodified setae.We propose that the sticky trap predation strategy
may allow sticky bugs to break functional constraintson the fore femur and therefore may promote a rapidrate of morphological evolution. The fore femur con-tains extensor and flexor muscles that move the foretibia, with the flexor muscle generating the graspingforce used to hold a prey item (Chapman, 1998). Thefore femur in raptorial assassin bugs is commonlyenlarged compared with the femora of the mid andhind legs, and contains a large flexor muscle. We fur-ther argue that there is probably a trade-off betweenthe thickness and length of the fore femur: to retainmobility, a thick femur may have to be relatively shortand a long femur relatively slender. Therefore wehypothesize that the thickness and the length of thefore femur in Reduviidae evolve in a correlated man-ner. We suggest that sticky bugs may rely to a lesserdegree on mechanical power to capture prey, poten-tially resulting in the reduced size of the flexor muscleand reduced femur thickness, and providing increasedpotential for the fore femur to become diversified.Thus we predict that the rate of morphological evolu-tion of the fore femur is higher in sticky bugs thanthat in non-sticky assassin bugs, and we hypothesizethat their fore femur is longer and more slender.We conduct phylogenetically informed comparative
analyses to test these hypotheses. We reconstruct thefirst comprehensive molecular phylogeny of the Har-pactorini, sampling 195 taxa of Harpactorini and 34other reduviid taxa and utilizing five gene segments.Harpactorini consist of > 2000 species (ca. 30% of thedescribed species diversity of assassin bugs) in ca. 290genera, are the largest tribe in the Reduviidae, haveworldwide distribution, and contain many charismaticspecies, such as the wheel bugs in the genus ArilusHahn, and the wasp or bee mimicking bugs (e.g. Har-pagocoris St�al, Hiranetis Spinola, Notocyrtus Burmei-ster, Xystonyttus Kirkaldy; Fig. 1) and natural enemies(Maldonado, 1990; Ambrose, 1999).
Fig. 1. Phylogeny of Harpactorini reconstructed using maximum likelihood (ML) and habitus images of representatives of Harpactorini andRhaphidosomini. Numbers after taxon names are RCW voucher numbers. Taxa in blue are New World, taxa in green, Old World. S, sticky bugclade; H, Heza clade (including Isocondylus). Three other clades are identified by circled numbers and referred to in the results. Asterisks (*) indi-cate the evolution of the mesopleural tubercle (plica). Numbers adjacent to nodes are bootstrap support values from the ML analysis and jack-knife support values from the parsimony analysis (shown only if > 50).
340 G. Zhang & C. Weirauch / Cladistics 30 (2014) 339–351
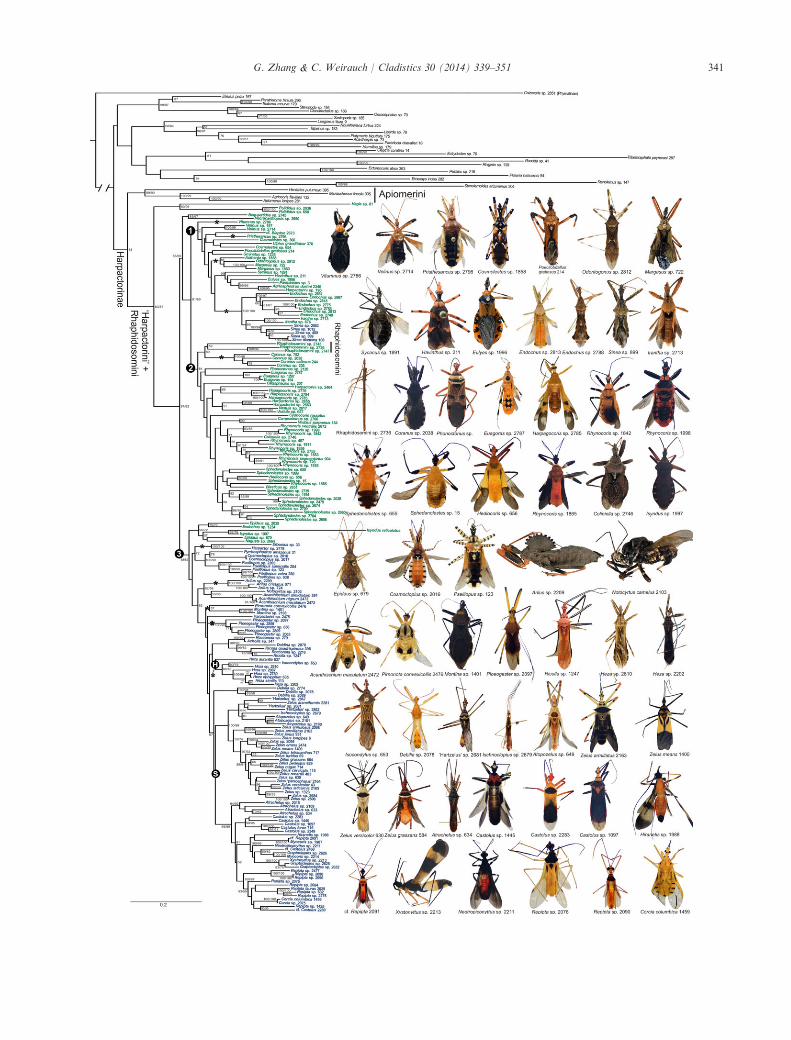
G. Zhang & C. Weirauch / Cladistics 30 (2014) 339–351 341

In addition to reconstructing the evolution of stickytrap predation within Harpactorini, the phylogeny isused to test the monophyly of Harpactorini and previ-ous hypotheses of supra-generic relationships and clas-sifications within the Harpactorini (Amyot andServille, 1843; Distant, 1904; Villiers, 1982; reviewedby Forero, 2011). These groups have a tendency to bedefined by superficially similar characters. Forinstance, St�al (1859) and Villiers (1982, 1983) used thecharacter plica, a tubercle on the mesopleuron, to deli-mit groups within the Harpactorini. We are interestedin testing whether this character is homoplasious orhomologous.Furthermore, to test the hypothesis of correlated
evolution of fore femur thickness and length, we usethe phylogenetically independent contrasts method(Felsenstein, 1985; Garland et al., 1992). We quantifythe thickness and length of the fore femur and employa maximum likelihood (ML) method utilizing aBrownian motion model to test the rates of morpho-logical evolution of those characters (O’Meara, 2006).
Materials and methods
Taxon sampling and molecular methods
The current study comprises 229 taxa. These include72 genera and 195 species of Harpactorini, represent-ing almost 30% of the generic diversity of the tribeand covering all major zoogeographical regions (Afro-tropical, Palearctic, Oriental, Australasian, Nearctic,and Neotropical). We also included seven samples oftwo other tribes in the subfamily Harpactorinae (Api-omerini and Rhaphidosomini) and 27 representativesof nine non-harpactorine reduviid subfamilies, generat-ing a dataset of 229 taxa. We here refer to Harpacto-rini and Rhaphidosomini as the ingroup as, accordingto our and previous analyses, Rhaphidosomini arenested within Harpactorini; we treat the remainingtaxa as outgroup. For identification and information-association purposes, each specimen was given amolecular voucher number (RCW_XXX) and a uniquespecimen identifier number (e.g. UCR_ENT 00004115or AMNH_PBI 00218711) following the methoddescribed by Schuh (2012), the latter used to log speci-men information into the Plant Bug Planetary Biodi-versity Inventory project database (https://research.amnh.org/pbi/locality). DNA extraction, PCR amplifi-cation, and sequencing reaction protocols includingprimer information follow those described by Weir-auch and Munro (2009) with modifications (Support-ing information, Table S1). The five gene segmentsused include three ribosomal DNAs (16S, 302–562 bp;28S D2 region, 332–694 bp; 28S D3–D5 region, 293–632 bp), the mitochondrial protein-coding cytochrome
c oxidase subunit I (COI) gene (331–833 bp), and the5′ end of a nuclear protein-coding Homeobox gene,Deformed (5′ Dfd, 223–590 bp; Tian et al., 2011). Atotal of 783 DNA sequences were generated in the cur-rent study, and a small number of sequences (134sequences, 15% of the entire data set) were obtainedfrom previous studies (Liu et al., 2009; Weirauch andMunro, 2009). The entire data set is 80% complete(917 sequences out of 1145 possible). For specimenvoucher information and Genbank accession numbers,see Table S2.
Alignments and phylogenetic reconstructions
Ribosomal DNAs (16S, 28S D2, 28S D3–D5) werealigned with MAFFT (Katoh et al., 2005) using the E-INS-i algorithm and default settings. Protein codinggenes, COI and 5′ Dfd, were aligned with MUSCLE(Edgar, 2004) via the European Bioinformatics Insti-tute portal (http://www.ebi.ac.uk/Tools/msa/muscle).The five gene data sets were concatenated with theprogram “Sequence Matrix” (Vaidya et al., 2011) togenerate a matrix of 229 taxa and 3820 bp. Phyloge-netic reconstruction using ML was conducted withRAxML 7.3.0 (Stamatakis, 2006; Stamatakis et al.,2008) via the Cyberinfrastructure for PhylogeneticResearch (CIPRES) portal (RAxML-HPC2 onXSEDE; default parameters; http://www.phylo.org).Analyses were performed with and without partitions.Five hundred rapid bootstrap replicates were per-formed to assess branch supports (random seed value12345). The best likelihood phylogeny from this analy-sis was reported and the bootstrap values indicated onthe phylogeny. Parsimony phylogenetic reconstructionwas performed using TNT ver. 1.1 (downloaded Feb-ruary 2013; Goloboff et al., 2008). Analyses were car-ried out with gaps read as missing data. We used adriven search with sectorial searches, ratcheting, drift-ing, and tree fusing (Goloboff, 1999; Nixon, 1999).For searching the most parsimonious tree(s), the fol-lowing parameters were specified: new technologysearch, level 50, initial addseqs = 60, find minimumtree length 40 times, random seed = 2000. The strictconsensus of the most parsimonious trees wasreported. Nodal support was assessed by jackkniferesampling (Farris et al., 1996; 500 replicates with36% character deletion) using the new technologysearch with sectorial searches, ratcheting, drifting, andtree fusing (search parameters: level 50, initial add-seqs = 9, find minimum tree length 5 times, randomseed = 2000). Jackknifing support values above 50were indicated on the strict consensus tree obtained inthe previous analysis. All phylogenies (parsimony andlikelihood) were rooted with a species of Phymatinae,a taxon that has been consistently recovered as part ofthe sister group to all other Reduviidae included in
342 G. Zhang & C. Weirauch / Cladistics 30 (2014) 339–351

this study (Weirauch, 2008; Weirauch and Munro,2009; Hwang and Weirauch, 2012).
Morphological observations and fore femurmeasurements
Information on sticky glands and modified setaewas extracted from Zhang and Weirauch (2013) for36 species. Fifty-four additional taxa, mostly genericrepresentatives (Table S3), were sampled across majorclades included in the phylogeny and examined forabsence/presence of sticky glands and modified setaefollowing protocols used in Zhang and Weirauch(2013). Despite the differences in sticky gland mor-phology between Heza spp. and the remaining sticky-bug genera that may indicate non-homology (Zhangand Weirauch, 2013), we here coded sticky glands asone absent/present character to test the possibilitythat the glands are homologous across the stickybugs.To quantify the thickness of the fore femur, we used
the ratio of length to width of the femur as a thicknessindex (TI; the higher the value of TI, the more slenderthe femur). The width of the fore femur at the mid-point in lateral view was measured using an ocularmicrometer mounted on a Nikon SMZ1500 dissectingmicroscope. For specimens with unequal widths alongthe femur, measurements were taken at three points(distally, medially, and proximally) and the average ofthe three values was used. This is a more conservativevalue than that taken at the thickest point. To quan-tify the relative length of the fore femur, a relativelength index (RLI) was determined as the ratiobetween the femur and body length measured from theanterior margin of the clypeus to the apex of the abdo-men (the larger the RLI, the longer the fore femur rel-ative to the body length). Data of morphologicalobservations and fore femur measurements are pro-vided in Table S3.
Ancestral state reconstruction, phylogenetic correlation,and rates of morphological evolution
Three character states were coded for the charactersticky glands: 0, absent; 1, with elongate or pear-shaped saccules; 2, with rounded saccules. Likewise,three states were coded for the condition of the forefemoral setae: 0, simple, without modifications; 1,modified to sundew setae; 2, modified to lamellatesetae. Ancestral state reconstruction of these two char-acters was performed based on both of the ML bestphylogeny and strict consensus tree from the parsi-mony phylogeny reconstruction. Mesquite v. 2.75(Maddison and Maddison, 2011) was used to performthe ancestral state reconstruction, and the parsimonymethod was employed. When the ML phylogeny was
used as the basis for ancestral state reconstruction, theoptimization of the character “sticky glands” wasunambiguous. However, multiple equally parsimoniousreconstructions existed for the evolution of setae.Instead of using ACCTRAN or DELTRAN to choosea favourable reconstruction (Agnarsson and Miller,2008), we used the most parsimonious reconstructions(MPRs) mode in Mesquite, which is able to show allthe reconstructions. When the parsimony strict consen-sus was used as the basis for ancestral state reconstruc-tion, the optimizations of both characters wereunequivocal. The evolutionary history of the characterplica was also reconstructed on the ML phylogenyusing parsimony in Mesquite.Fore femoral thickness and length (as thickness and
relative length indices) were mapped on the ML phy-logeny as continuous characters using two parsimony-based methods: weighted squared-change parsimony(Maddison, 1991) and linear parsimony (Farris, 1970;Swofford and Maddison, 1987), as implemented inMesquite. The former method minimizes the sum ofthe square of change on each branch of the phylogeny,causing changes to spread out over the tree. The linearparsimony minimizes the absolute sum of changesacross the phylogeny. It considers only the three clos-est nodes in order to calculate the ancestral value of anode. This method tends to produce ambiguities forthe reconstructions and in Mesquite a “min” and a“max” value are reported from a set of equally parsi-monious values.In the subsequent comparative analyses, the unparti-
tioned ML best phylogeny was used. This is primarilybecause of the requirement for branch length in theanalyses. To test for the correlation between the twocontinuous characters (fore femur thickness andlength), the phylogenetically independent contrastsmethod (Felsenstein, 1985) was used as implementedin PDAP (Garland et al., 1992). The TI was Ln-trans-formed and the RLI was square rooted to create nor-malized distributions. Branch length transformation ofNee’s method (Purvis, 1995) was used, and diagnostictests were performed to ensure statistical assumptionswere met (Garland et al., 1992). Linear regressionanalysis of the contrasts was performed using the soft-ware QI Macros 2011 (www.qimacros.com) embeddedin Microsoft Excel 2010.We used the program Brownie (O’Meara et al.,
2006) to test if rates of morphological evolution havechanged across Reduviidae and Harpactorini, in par-ticular between non-sticky and sticky assassin bugs.Three analyses were performed that contrasted therates in the following groups with each other: (i) stickybugs vs. non-sticky Harpactorini; (ii) sticky bugs vs.all other reduviids; and (iii) Harpactorini (includingRhaphidosomini) vs. other tribes and subfamilies. Ourphylogenetic analyses suggest that sticky bugs, as
G. Zhang & C. Weirauch / Cladistics 30 (2014) 339–351 343

defined by the unique morphological characters, arenot monophyletic and form two clades (H and S inFig. 1). For the analyses of rate evolution, we definedsticky bugs as those forming the clade S, which exclu-sively consists of 13 sticky-bug genera (versus one inthe clade H), represented by 60 species. We consideredthe clade H, containing Heza and Isocondylus Amyotand Serville, as part of the non-sticky bugs. We madethis treatment because the specialized glandular andcuticular morphology in Heza spp. may not representsticky trap predation, as suggested by the phylogeneticresults and comparative morphological evidence (seeDiscussion).In Brownie we used the censored rate test method
and both the hypothesis-testing and model-selectionapproaches (O’Meara et al., 2006). In the hypothesis-testing approach, 1000 parametric bootstrapping repli-cates were performed. For the likelihood ratio test ofthis approach, a P value < 0.05 was considered as sig-nificant for rejection of the null hypothesis (i.e. rate ofevolution is constant throughout the tree). In themodel-selection approach, two models were comparedfor their fit to the data, one with a single rate andanother with two rates. A smaller value of the second-order Akaike information criterion (AICc) indicates abetter-fitting model. An absolute difference in theAICc of the two models (ΔAICc) ≥ 8 suggests that thebetter fitting model is strongly favoured, and a valueof ΔAICc ≥ 4 indicates the better-fitting model is mod-erately supported. We used the ML phylogram shownin Fig. 1 that has branch lengths proportional to thenumber of substitutions, which is appropriate for therate tests implemented in Brownie (Litsios and Salam-in, 2012; B.C. O’Meara, pers. commun.). The Ln- orsquare root-transformed thickness and relative lengthindices were used for the tests.
Results
Sticky gland observations and fore femoralmeasurements
Three previously unstudied genera of Harpactorini,Ischnoclopius St�al, Neotropiconyttus Kirkaldy, andXystonyttus are here shown to possess sticky glandsand sundew setae, increasing the known number ofsticky-bug genera to 15 (Table S3). Sticky glands andsundew setae are present in all the sampled species ofthe 13 sticky-bug genera included in the phylogeny.The genus Heza includes species with both stickyglands and lamellate setae, and species that lack oneor both structures. The thickest fore femora werefound in non-harpactorine taxa—Racelda sp. (Ectri-chodiinae; TI = 2.57), Cnizocoris sp. (Phymatinae;2.62), and Peirates sp. (Peiratinae; 3.10). The most
slender fore femora were found in Ischnoclopius (Har-pactorini; TI = 49.50), Rhaphidosomini sp.RCW_2736 (29.02) and “Hartzelus” (manuscriptname) RCW_2862 (Harpactorini; 28.64). Two of these,Ischnoclopius and “Hartzelus”, are sticky bugs. Thethree relatively shortest fore femora were also found inthe outgroup taxa: Ploiaria hirticornis (Banks) (Emesi-nae; RLI = 0.17), Rhiginia sp. (Ectrichodiinae; 0.19),and Platymeris biguttatus (Linnaeus) (Reduviinae;0.19). The three relatively longest fore femora werefound in the sticky bugs: Ischnoclopius (0.75) and twospecies of Zelus (0.66 and 0.58). The sticky bug Isch-noclopius has both the most slender and the relativelylongest fore femur.
Phylogenetic relationships with the Harpactorini
The best tree from the unpartitioned ML analysis isshown in Fig. 1. The phylogeny from the partitionedML analysis is provided in the supporting information(Fig. S1) and the strict consensus of 85 equally mostparsimonious trees in Fig. S2. These phylogenetichypotheses are highly congruent. The ML phylogenetictree with bootstrap support values shown in Fig. 1 willbe the primary basis for describing and discussing phy-logenetic relationships and evolutionary scenariosbelow. Harpactorinae, represented in the present studyby Apiomerini, Harpactorini, and Rhaphidosomini,was recovered as monophyletic. Apiomerini is mono-phyletic and the sister group to the remaining Harpac-torinae included in this analysis. The tribeHarpactorini is paraphyletic with respect to the Rhap-hidosomini, which is the sister group to the Harpacto-rini genus Coranus Curtis. Fourteen genera of stickybugs were represented in the phylogeny, and fall intotwo clades. The genus Heza and the non-sticky Isoc-ondylus form a clade (clade H; Fig. 1), with Hezabeing paraphyletic. All the remaining 13 sticky-buggenera form another monophyletic clade (clade S;Fig. 1). In the unpartitioned ML analysis (Fig. S1)and the parsimony consensus (Fig. S2), the paraphylyof Harpactorini, the three clades defined in the phylog-eny shown in Fig. 1, and the monophyly of the stickybug clade S were also recovered. The parsimony strictconsensus tree failed to resolve the monophyly of Har-pactorinae and did not recover clade H.Of the 34 genera represented by at least two species
(Table 1), 18 were recovered as monophyletic, includ-ing some relatively speciose genera such as Coranus,Euagoras Burmeister, Ploeogaster Amyot and Serville,and Sinea Amyot and Serville. Genera recovered asparaphyletic are Cosmolestes St�al, “Hartzelus”, Heza,and Ricolla St�al. The genera Endochus St�al, EpidausSt�al, Graptocleptes St�al, Myocoris Burmeister, Rhynoc-oris Hahn, Rocconota St�al, Sphedanolestes St�al, andZelus are polyphyletic based on our analyses. The
344 G. Zhang & C. Weirauch / Cladistics 30 (2014) 339–351
monophyly of Repipta St�al and Castolus St�al was notdetermined because they include specimens whose gen-eric identification is uncertain. Nearly all historicallyproposed supra-generic groups within the Harpactoriniare paraphyletic or polyphyletic in the current phylog-eny (Table 1).To describe aspects of the overall phylogeny of Har-
pactorini and to place sticky bugs in that context, wehighlight three well supported clades within the Har-
pactorini + Rhaphidosomini group (bootstrap value> 85; indicated by numbers in black circles; Fig. 1):clade 1 contains some of the largest harpactorineassassin bugs (e.g. Eulyes Amyot & Serville, SycanusAmyot & Serville, Pantoleistes St�al; body length 27–29 mm), with species mostly found in the Old Worldtropics. Interestingly, the New World genus Sinea isnested within this clade and recovered as sister to theOld World genus Irantha St�al. Members of these two
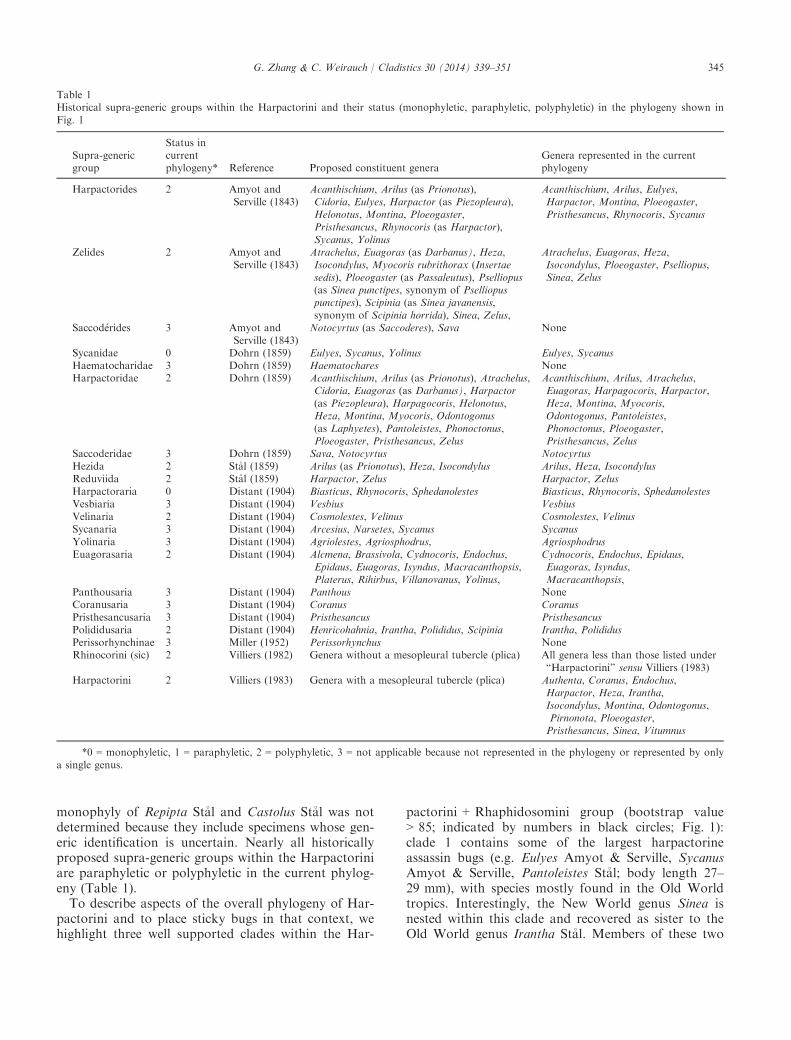
Table 1Historical supra-generic groups within the Harpactorini and their status (monophyletic, paraphyletic, polyphyletic) in the phylogeny shown inFig. 1
Supra-genericgroup
Status incurrentphylogeny* Reference Proposed constituent genera
Genera represented in the currentphylogeny
Harpactorides 2 Amyot andServille (1843)
Acanthischium, Arilus (as Prionotus),Cidoria, Eulyes, Harpactor (as Piezopleura),Helonotus, Montina, Ploeogaster,Pristhesancus, Rhynocoris (as Harpactor),Sycanus, Yolinus
Acanthischium, Arilus, Eulyes,Harpactor, Montina, Ploeogaster,Pristhesancus, Rhynocoris, Sycanus
Zelides 2 Amyot andServille (1843)
Atrachelus, Euagoras (as Darbanus), Heza,Isocondylus, Myocoris rubrithorax (Insertaesedis), Ploeogaster (as Passaleutus), Pselliopus(as Sinea punctipes, synonym of Pselliopuspunctipes), Scipinia (as Sinea javanensis,synonym of Scipinia horrida), Sinea, Zelus,
Atrachelus, Euagoras, Heza,Isocondylus, Ploeogaster, Pselliopus,Sinea, Zelus
Saccod�erides 3 Amyot andServille (1843)
Notocyrtus (as Saccoderes), Sava None
Sycanidae 0 Dohrn (1859) Eulyes, Sycanus, Yolinus Eulyes, SycanusHaematocharidae 3 Dohrn (1859) Haematochares NoneHarpactoridae 2 Dohrn (1859) Acanthischium, Arilus (as Prionotus), Atrachelus,
Cidoria, Euagoras (as Darbanus), Harpactor(as Piezopleura), Harpagocoris, Helonotus,Heza, Montina, Myocoris, Odontogonus(as Laphyetes), Pantoleistes, Phonoctonus,Ploeogaster, Pristhesancus, Zelus
Acanthischium, Arilus, Atrachelus,Euagoras, Harpagocoris, Harpactor,Heza, Montina, Myocoris,Odontogonus, Pantoleistes,Phonoctonus, Ploeogaster,Pristhesancus, Zelus
Saccoderidae 3 Dohrn (1859) Sava, Notocyrtus NotocyrtusHezida 2 St�al (1859) Arilus (as Prionotus), Heza, Isocondylus Arilus, Heza, IsocondylusReduviida 2 St�al (1859) Harpactor, Zelus Harpactor, ZelusHarpactoraria 0 Distant (1904) Biasticus, Rhynocoris, Sphedanolestes Biasticus, Rhynocoris, SphedanolestesVesbiaria 3 Distant (1904) Vesbius VesbiusVelinaria 2 Distant (1904) Cosmolestes, Velinus Cosmolestes, VelinusSycanaria 3 Distant (1904) Arcesius, Narsetes, Sycanus SycanusYolinaria 3 Distant (1904) Agriolestes, Agriosphodrus, AgriosphodrusEuagorasaria 2 Distant (1904) Alcmena, Brassivola, Cydnocoris, Endochus,
Epidaus, Euagoras, Isyndus, Macracanthopsis,Platerus, Rihirbus, Villanovanus, Yolinus,
Cydnocoris, Endochus, Epidaus,Euagoras, Isyndus,Macracanthopsis,
Panthousaria 3 Distant (1904) Panthous NoneCoranusaria 3 Distant (1904) Coranus CoranusPristhesancusaria 3 Distant (1904) Pristhesancus PristhesancusPolididusaria 2 Distant (1904) Henricohahnia, Irantha, Polididus, Scipinia Irantha, PolididusPerissorhynchinae 3 Miller (1952) Perissorhynchus NoneRhinocorini (sic) 2 Villiers (1982) Genera without a mesopleural tubercle (plica) All genera less than those listed under
“Harpactorini” sensu Villiers (1983)Harpactorini 2 Villiers (1983) Genera with a mesopleural tubercle (plica) Authenta, Coranus, Endochus,
Harpactor, Heza, Irantha,Isocondylus, Montina, Odontogonus,Pirnonota, Ploeogaster,Pristhesancus, Sinea, Vitumnus
*0 = monophyletic, 1 = paraphyletic, 2 = polyphyletic, 3 = not applicable because not represented in the phylogeny or represented by onlya single genus.
G. Zhang & C. Weirauch / Cladistics 30 (2014) 339–351 345

genera are heavily tuberculate and possess acute spineson the fore leg (Fig. 1), features that we now proposeto be homologous in the two taxa and potentially syn-apomorphic for the Irantha + Sinea clade. Clade 2 hasan exclusive Old World distribution and containsRhaphidosomini and some small to mid-sized Har-pactorini. The Afrotropical wasp-mimicking Harpagoc-oris and two of the most speciose genera ofHarpactorinae and Reduviidae, Rhynocoris and Sphe-danolestes, belong to this clade. Clade 3 contains allthe New World taxa sampled except for Sinea, butalso a small Old World group that is recovered as thesister group to the Neotropical taxa. The sticky-buggenera are nested within this Neotropical clade.Eighty-five equally most parsimonious trees were
produced from the parsimony analysis, each with alength of 23 798. Clades 1–3 and clade S were consis-tently recovered, but members of Heza and Isocondylus(clade H) were not recovered as monophyletic. This isdue to the placement of Heza aurantia as the sister toa non-Heza species. The ML tree (Fig. 1) is not one ofthe most parsimonious trees and has a length of23 966, which is 168 steps longer than the parsimonytrees.
Character evolution and correlation, and rates ofmorphological evolution
Results of ancestral state reconstruction of the twodiscrete characters of special interest (gland and setaltypes) are largely congruent between the two analysesusing either the ML phylogeny or the parsimony strictconsensus tree. Both show a single origin of the stickyglands with elongate saccules and two origins of thekind with rounded saccules (Fig. 2 and Fig. S3). Sun-dew setae evolved once at the base of the clade S andare retained in all the taxa examined within this clade.The evolution of sticky glands and sundew setae co-occur at the base of clade S. We hereafter refer toclade S as the “sticky bugs”. According the analysisbased on the ML phylogeny, three MPRs exist for theevolution of lamellate setae: one gain and three losses;three gains and one loss; and four gains (the first sce-nario is shown in Fig. 2 and all three are shownFig. S4). In contrast, the analysis results based on theparsimony strict consensus tree unambiguously showthree origins and loss of the lamellate setae (Fig. 3).This corresponds to one of the MPRs when using theML phylogeny. For the character plica, the MPRshows 10 independent origins (see Fig. 1 for branchesmarked with asterisks).Weighted squared-change parsimony reconstruction
assigned a thick and short fore femur to the ancestor ofassassin bugs (TI = 4.94, RLI = 0.25). The majority ofthe outgroup taxa possess a thick and short forefemur (Table S3). The ancestor of Harpactorinae was
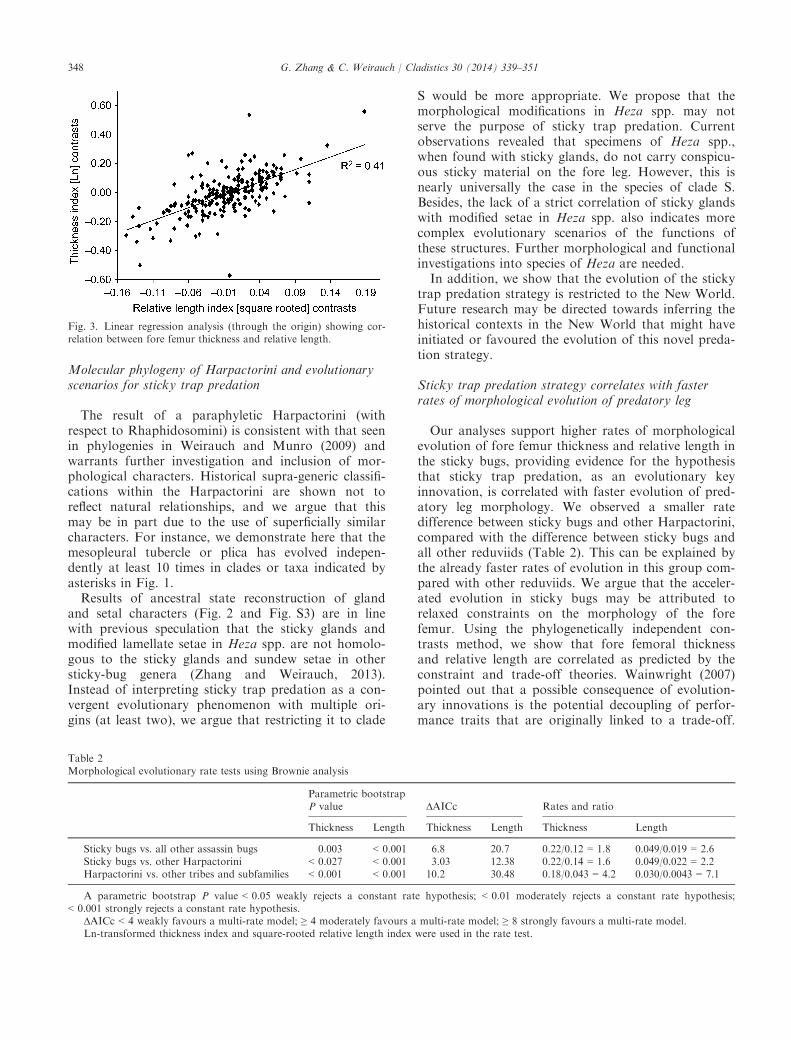
reconstructed to have values of TI = 7.64 andRLI = 0.32. Those values are even higher in sticky bugs(clade S; TI = 12.9, RLI = 0.41), indicating that theancestor of this clade had a more slender and longerfore femur. The reconstructions based on the linearparsimony method demonstrated a similar pattern(ancestor of Reduviidae—TI: 2.62–7.78 [min–max],RLI: 0.21–0.24; ancestor of Harpactorinae—TI: 5.18–8.44, RLI: 0.32–0.35; ancestor of sticky bugs—TI:10.83–12.0, RLI: 0.37–0.39). The phylogenetically inde-pendent contrasts analysis shows that fore femur thick-ness and relative length are strongly correlated(R2 = 0.41, d.f. = 227, P < 0.001; Fig. 3). This correla-tion is also evident in the lateral habitus images andline drawings of the fore femur of selected Reduviidae(Fig. 2 and Fig. S5), and can be visualized in the mir-rored trees showing the evolution of the two characters.When sticky bugs were compared with all other
assassin bugs, the hypothesis of a constant rate ofevolution throughout the tree was rejected for bothfore femur thickness (P = 0.003) and relative length(P < 0.001) using the parametric bootstrapping likeli-hood ratio test (Table 2). The model-selectionapproach also found moderate or strong support,respectively, for a two-rate model for the evolution offore femur thickness (ΔAICc = 6.8) and relative length(ΔAICc = 20.7). In clade S, the fore femur TI (Ln-transformed) exhibited a Brownian motion rate 1.8times faster than the rate in the remaining Reduvii-dae; the rate for relative femur length index (squarerooted) was 2.6 times higher than the rate in otherReduviidae. For the analyses comparing sticky bugsto other Harpactorini, and Harpactorini to otherreduviids, a two-rate model was also favoured(Table 2). The rate differences were smaller betweensticky bugs and other Harpactorini than betweensticky bugs and all other reduviids. Within the stickybugs (clade S), multiple taxa show exceptionally slen-der and long fore femora (e.g. Ischnoclopius sp.RCW_2679, “Hartzelus” sp. RCW_2681, Zelus sp.RCW_2095; Fig. 1).
Discussion
The current study investigates the evolutionaryhistory of a novel predation strategy—sticky trappredation—and tests for its correlation with the mor-phological diversification of the predatory fore leg inassassin bugs. We show that the evolution of stickytrap predation is correlated with the evolution of a forefemur that is more slender and longer than that of atypical raptorial fore leg as seen in most other assassinbugs. We argue that sticky strap predation may havereduced constraints on the fore femur and acceleratedthe evolution of more slender and longer fore legs.
346 G. Zhang & C. Weirauch / Cladistics 30 (2014) 339–351
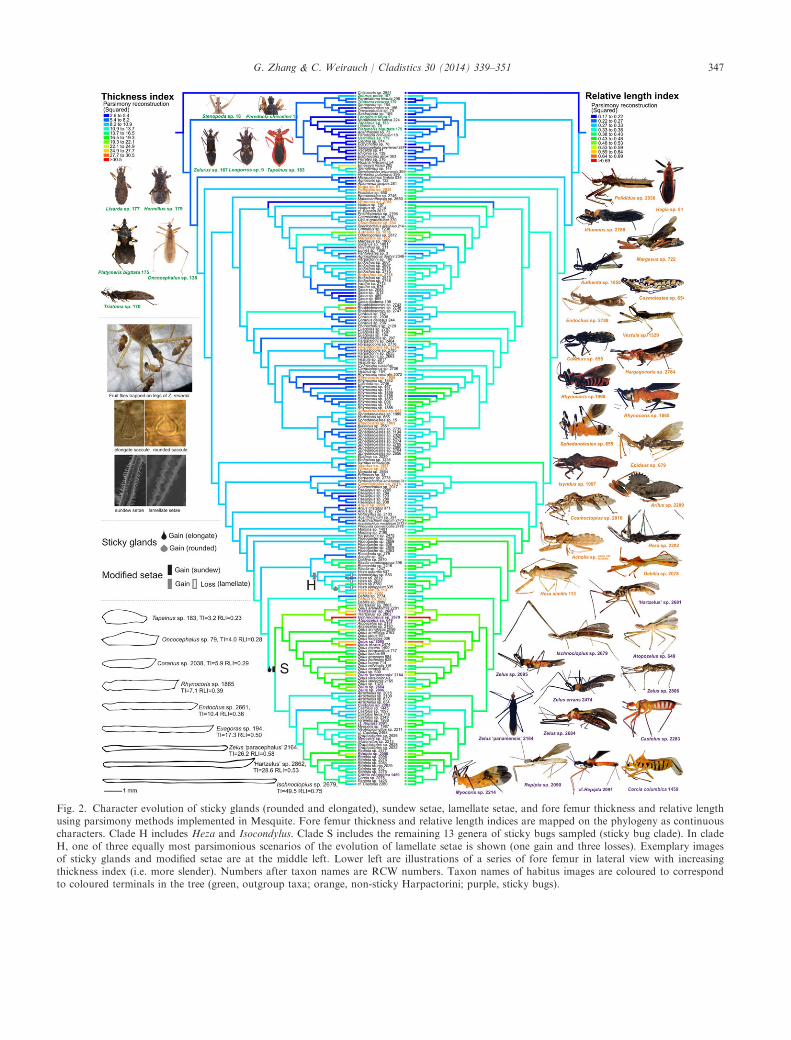
Fig. 2. Character evolution of sticky glands (rounded and elongated), sundew setae, lamellate setae, and fore femur thickness and relative lengthusing parsimony methods implemented in Mesquite. Fore femur thickness and relative length indices are mapped on the phylogeny as continuouscharacters. Clade H includes Heza and Isocondylus. Clade S includes the remaining 13 genera of sticky bugs sampled (sticky bug clade). In cladeH, one of three equally most parsimonious scenarios of the evolution of lamellate setae is shown (one gain and three losses). Exemplary imagesof sticky glands and modified setae are at the middle left. Lower left are illustrations of a series of fore femur in lateral view with increasingthickness index (i.e. more slender). Numbers after taxon names are RCW numbers. Taxon names of habitus images are coloured to correspondto coloured terminals in the tree (green, outgroup taxa; orange, non-sticky Harpactorini; purple, sticky bugs).
G. Zhang & C. Weirauch / Cladistics 30 (2014) 339–351 347
Molecular phylogeny of Harpactorini and evolutionaryscenarios for sticky trap predation
The result of a paraphyletic Harpactorini (withrespect to Rhaphidosomini) is consistent with that seenin phylogenies in Weirauch and Munro (2009) andwarrants further investigation and inclusion of mor-phological characters. Historical supra-generic classifi-cations within the Harpactorini are shown not toreflect natural relationships, and we argue that thismay be in part due to the use of superficially similarcharacters. For instance, we demonstrate here that themesopleural tubercle or plica has evolved indepen-dently at least 10 times in clades or taxa indicated byasterisks in Fig. 1.Results of ancestral state reconstruction of gland
and setal characters (Fig. 2 and Fig. S3) are in linewith previous speculation that the sticky glands andmodified lamellate setae in Heza spp. are not homolo-gous to the sticky glands and sundew setae in othersticky-bug genera (Zhang and Weirauch, 2013).Instead of interpreting sticky trap predation as a con-vergent evolutionary phenomenon with multiple ori-gins (at least two), we argue that restricting it to clade
S would be more appropriate. We propose that themorphological modifications in Heza spp. may notserve the purpose of sticky trap predation. Currentobservations revealed that specimens of Heza spp.,when found with sticky glands, do not carry conspicu-ous sticky material on the fore leg. However, this isnearly universally the case in the species of clade S.Besides, the lack of a strict correlation of sticky glandswith modified setae in Heza spp. also indicates morecomplex evolutionary scenarios of the functions ofthese structures. Further morphological and functionalinvestigations into species of Heza are needed.In addition, we show that the evolution of the sticky
trap predation strategy is restricted to the New World.Future research may be directed towards inferring thehistorical contexts in the New World that might haveinitiated or favoured the evolution of this novel preda-tion strategy.
Sticky trap predation strategy correlates with fasterrates of morphological evolution of predatory leg
Our analyses support higher rates of morphologicalevolution of fore femur thickness and relative length inthe sticky bugs, providing evidence for the hypothesisthat sticky trap predation, as an evolutionary keyinnovation, is correlated with faster evolution of pred-atory leg morphology. We observed a smaller ratedifference between sticky bugs and other Harpactorini,compared with the difference between sticky bugs andall other reduviids (Table 2). This can be explained bythe already faster rates of evolution in this group com-pared with other reduviids. We argue that the acceler-ated evolution in sticky bugs may be attributed torelaxed constraints on the morphology of the forefemur. Using the phylogenetically independent con-trasts method, we show that fore femoral thicknessand relative length are correlated as predicted by theconstraint and trade-off theories. Wainwright (2007)pointed out that a possible consequence of evolution-ary innovations is the potential decoupling of perfor-mance traits that are originally linked to a trade-off.
Fig. 3. Linear regression analysis (through the origin) showing cor-relation between fore femur thickness and relative length.
Table 2Morphological evolutionary rate tests using Brownie analysis
Parametric bootstrapP value ΔAICc Rates and ratio
Thickness Length Thickness Length Thickness Length
Sticky bugs vs. all other assassin bugs 0.003 < 0.001 6.8 20.7 0.22/0.12 = 1.8 0.049/0.019 = 2.6Sticky bugs vs. other Harpactorini < 0.027 < 0.001 3.03 12.38 0.22/0.14 = 1.6 0.049/0.022 = 2.2Harpactorini vs. other tribes and subfamilies < 0.001 < 0.001 10.2 30.48 0.18/0.043 = 4.2 0.030/0.0043 = 7.1
A parametric bootstrap P value < 0.05 weakly rejects a constant rate hypothesis; < 0.01 moderately rejects a constant rate hypothesis;< 0.001 strongly rejects a constant rate hypothesis.
ΔAICc < 4 weakly favours a multi-rate model; ≥ 4 moderately favours a multi-rate model; ≥ 8 strongly favours a multi-rate model.Ln-transformed thickness index and square-rooted relative length index were used in the rate test.
348 G. Zhang & C. Weirauch / Cladistics 30 (2014) 339–351

In the context of a raptorial fore leg, these two perfor-mance traits could be the reach (determined by thelength of fore femur) and the speed and strength withwhich a prey organism can be captured and held(determined by the thickness of the fore femur as aproxy for size and power of the femoral flexor muscle).A thick femur containing a larger flexor can generatea larger force, but would be compromised in its lengthor how far it can reach due to the difficulty of movinga long and heavy femur. On the other hand, a longerfemur enables the bug to reach further, but would bemore slender, thus compromising mechanical strengthor efficiency of prey capture. Sticky trap predationmay break these constraints by exchanging mechanicalstrength for a sticky trap, and therefore provides anopportunity for faster evolution of the fore femur,which becomes both longer and more slender withinthe sticky bug group. However, the realization of thisopportunity varies across sticky bugs, with some taxaretaining legs that are comparable with those of non-sticky harpactorines (e.g. Castolus sp. RCW_1097).Furthermore, it could be argued that this predationstrategy does not necessarily replace the raptorial func-tion of the fore leg, but supplements it. We suspectthat faster morphological evolution may be realizedonly under certain ecological circumstances. In fact,the degree of femoral elongation and slenderness variesamong the sticky bugs, with some showing morphol-ogy similar to that in the non-sticky bugs.
Sticky trap predation and adaptive radiation
Simpson (1953) argued that the acquisition of anovel trait that influences the ability to exploit unusedresources may be critical for the initiation of an adap-tive radiation. Under this theoretical framework, thesticky trap predation strategy may have initiated anadaptive radiation of sticky bugs (clade S). Accordingto Schluter (2000) and Glor (2010), three criteria needto be met to qualify as adaptive radiation—commonancestry, rapid speciation and extraordinary diversity,and trait utility. In the present study, we have shownthat the sticky bugs (clade S, Fig. 1) share a commonancestor. Second, the extraordinary diversification andrapid speciation of the sticky bugs will need to bedemonstrated. Sticky bugs currently have 134described species. They are an order of magnitudemore speciose than their sister group (Debilia; ninespp.), and comprise a large proportion (ca. 43%) of alldescribed New World species of Harpactorini (Maldo-nado, 1990). Additional studies should investigate ifsticky bugs also show elevated speciation rates, using atime-calibrated phylogeny and speciation rate esti-mates (Alfaro et al., 2009b). Finally, the trait utility oradaptive function of the sticky trap predation strategywill need to be demonstrated. This will require tests of
prey-capture efficiency of sticky and non-sticky bugswith various prey items. We speculate that sticky trapprobably increases the efficiency of capturing small,fast-flying prey and therefore opens a previously rela-tively inaccessible ecological opportunity. Sticky trappredation has been shown to be effective for trappingsmall flying insects such Drosophila melanogaster Mei-gen (Edwards, 1966; Law and Sediqi, 2010; Zhang andWeirauch, 2013). In contrast, other assassin bugs,including non-sticky Harpactorini, usually grasp theirprey items between the fore femur and tibia (Ambrose,1999). In summary, we propose that sticky bugs mayindeed represent an adaptive radiation, but the above-mentioned studies are required to further test thishypothesis.
Acknowledgements
Financial support is provided primarily by the NSFgrant “Partnership in Enhancing Expertise in Taxon-omy” (PEET) #0933853 awarded to C. Weirauch.Additional financial support comes from the Robertvan den Bosch Scholarship (Center for Biological Con-trol, University of California, Berkeley), the Disserta-tion Year Fellowship and the Dissertation ResearchGrant (University of California, Riverside, UCR)awarded to G. Zhang, and the Beneficial ArthropodExploration fund (UCR) awarded to C. Weirauch.The Willi Hennig Society is acknowledged for makingTNT available for public use. We thank James Bald-win, Michael Forthman, John Heraty (UCR), WeiSong Hwang (National University of Singapore,NUS), Sunghoon Jung (Seoul National University),and Dimitri Forero (Pontificia Universidad Javeriana)for critical reading of earlier versions of the manu-script. We are grateful to the following colleagues whohave assisted with acquisition or loans of specimens:Caroline Chaboo (University of Kansas), Jason Cryan(New York State Museum), Torsten Dikow (NationalMuseum of Natural History, USNM), Jos�e A. Fernan-des (Universidade Federal do Par�a), Dimitri Forero,Dave Hawks (UCR), Thomas Henry (USNM), JeremyHuff (American Museum of Natural History,AMNH), Hui Liu (Kyushu University), DmitriMaslov (UCR), Rudolf Meier (NUS), Jim Miller(AMNH), Jason Mottern (UCR), Norman Penny(California Academy of Sciences), Michael Sharkey(University of Kentucky), and Steffen Roth (Univer-sity of Bergen). We would like to thank VincentCareau (UCR), David Collar (University of Califor-nia, Santa Cruz), Dimitri Forero, Wei Song Hwang,Brian O’Meara (University of Tennessee, Knoxville),and Qiang Xie (Nankai University) for technical dis-cussion and advice on molecular methods, use of pro-grams, or identification of specimens. We also thank
G. Zhang & C. Weirauch / Cladistics 30 (2014) 339–351 349

two anonymous reviewers for reviewing the manu-script.
References
Agnarsson, I., Miller, J.A., 2008. Is ACCTRAN better thanDELTRAN? Cladistics 24, 1032–1038.
Alfaro, M.E., Brock, C.D., Banbury, B., Wainwright, P.C., 2009a.Does evolutionary innovation in pharyngeal jaws lead to rapidlineage diversification in labrid fish? BMC Evol. Biol. 9, 255.
Alfaro, M.E., Santini, F., Brock, C., Alamillo, H., Dornburg, A.,Rabosky, D.L., Carnevale, G., Harmon, L.J., 2009b. Nineexceptional radiations plus high turnover explain species diversityin jawed vertebrates. Proc. Natl Acad. Sci. USA 106, 13410–13414.
Ambrose, D.P., 1999. Assassin Bugs. Science Publishers, Enfield,NH.
Amyot, C.J.B., Serville, A., 1843. Histoire Naturelle des Insectes.H�emipt�eres. Librairie encyclop�edique de Roret, Fain et Thunot,Paris.
Barth, R., 1952. Die Fangdr€usen an den Beinen von Zelus(Diplocodus) leucogrammus (Perty). Zool. Jb. 73, 323–336.
Bond, J.E., Opell, B.D., 1998. Testing adaptive radiation and keyinnovation hypotheses in spiders. Evolution 52, 403–414.
Chapman, R.F., 1998. The Insects: Structure and Function.Cambridge University Press, Cambridge, UK.
Collar, D.C., O’Meara, B.C., Wainwright, P.C., Near, T.J., 2009.Piscivory limits morphological diversification in centrarchidfishes. Evolution 63, 1557–1573.
Collar, D.C., Schulte, J.A. II, O’Meara, B.C., Losos, J.B., 2010.Habitat use affects morphological diversification in dragonlizards (Agamidae). J. Evol. Biol. 23, 1033–1049.
Collar, D.C., Schulte, J.A. II, Losos, J.B., 2011. Evolution ofextreme body size disparity in monitor lizards (Varanus).Evolution 65, 2664–2680.
Cooper, D.W., Steppan, S.J., 2010. Developmental constraint on theevolution of marsupial forelimb morphology. Aust. J. Zool. 58,1–15.
Cracraft, J., 1990. The origin of evolutionary novelties: pattern andprocess at different hierarchical levels. In: Nitecki, M. (Ed.),Evolutionary Innovations. University of Chicago Press, Chicago,IL, pp. 21–44.
Derryberry, E.P., Claramunt, S., Derryberry, G., Chesser, R.T.,Cracraft, J., Aleixo, A., P�erez-Em�an, J., Remsen, J.V. Jr,Brumfield, R.T., 2011. Lineage diversification and morphologicalevolution in a large-scale continental radiation: the Neotropicalovenbirds and woodcreepers (aves: Furnariidae). Evolution 65,2973–2986.
Distant, W.L., 1904. The Fauna of British India, Including Ceylonand Burma. Rhynchota, Vol. 2. (Heteroptera). Taylor & Francis,London.
Dohrn, A., 1859b. Catalogus hemipterorum. Entomologischer Vereinin Stettin, Buchdruckerei von Herrcke & Lebeling, Stettin.
Dornburg, A., Sidlauskas, B., Santini, F., Sorenson, L., Near, T.J.,Alfaro, M.E., 2011. The influence of an innovative locomotorstrategy on the phenotypic diversification of triggerfish (Family:Balistidae). Evolution 65, 1912–1926.
Drummond, C.S., Eastwood, R.J., Miotto, S.T., Hughes, C.E., 2012.Multiple continental radiations and correlates of diversification inLupinus (Leguminosae): testing for key innovation withincomplete taxon sampling. Syst. Biol. 61, 443–460.
Edgar, R.C., 2004. MUSCLE: multiple sequence alignment with highaccuracy and high throughput. Nucleic Acids Res. 32, 1792–1797.
Edwards, J.S., 1966. Observations on the life history and predatorybehaviour of Zelus exsanguis (St�al) (Heteroptera: Reduviidae).Proc. R. Entomol. Soc. Lond. A, Gen. Entomol. 41, 21–24.
Erkens, R.H.J., Chatrou, L.W., Couvreur, T.L.P., 2012. Radiationsand key innovations in a basal angiosperm lineage (Annonaceae;Magnoliales). Bot. J. Linn. Soc. 169, 117–134.
Farris, J.S., 1970. Methods of computing Wagner trees. Syst. Zool.19, 83–92.
Farris, J.S., Albert, V.A., K€allersj€o, M., Lipscomb, D., Kluge, A.G.,1996. Parsimony jackknifing outperforms neighbor-joining.Cladistics 12, 99–124.
Felsenstein, J., 1985. Phylogenies and the comparative method. Am.Nat. 125, 1–15.
Forero, D., 2011. Classification of Harpactorinae assassin bugs(Hemiptera: Heteroptera: Reduviidae). Bol. Mus. Entomol.Francisco Luis. Gallego 3, 9–24.
Galis, F., 2001. Key innovations and radiations. In: Wagner, G.P.(Ed.), The Character Concept in Evolutionary Biology. AcademicPress, San Diego, CA, pp. 583–607.
Garland, T. Jr, Harvey, P.H., Ives, A.R., 1992. Procedures for theanalysis of comparative data using phylogenetically independentcontrasts. Syst. Biol. 41, 18–32.
Gatesy, S.M., Middleton, K.M., 1997. Bipedalism, flight, and theevolution of theropod locomotor diversity. J. Vertebr. Paleontol.17, 308–329.
Glor, R.E., 2010. Phylogenetic insights on adaptive radiation. Annu.Rev. Ecol. Evol. Syst. 41, 251–270.
Goloboff, P.A., 1999. Analyzing large data sets in reasonable times:solutions for composite optima. Cladistics 15, 415–428.
Goloboff, P.A., Farris, J.S., Nixon, K.C., 2008. TNT, a freeprogram for phylogenetic analysis. Cladistics 24, 774–786.
Harmon, L.J., Melville, J., Larson, A., Losos, J.B., 2008. The role ofgeography and ecological opportunity in the diversification ofday geckos (Phelsuma). Syst. Biol. 57, 562–573.
Hulsey, C.D., Mims, M.C., Parnell, N.F., Streelman, J.T., 2010.Comparative rates of lower jaw diversification in cichlid adaptiveradiations. J. Evol. Biol. 23, 1456–1467.
Hunter, J.P., 1998. Key innovations and the ecology ofmacroevolution. Trends Ecol. Evol. 13, 31–36.
Hwang, W.S., Weirauch, C., 2012. Evolutionary history of assassinbugs (Insecta: Hemiptera: Reduviidae): insights from divergencedating and ancestral state reconstruction. PLoS One 7, e45523.
Katoh, K., Kuma, K., Toh, H., Miyata, T., 2005. MAFFT version5: improvement in accuracy of multiple sequence alignment.Nucleic Acids Res. 33, 511–518.
Law, Y.-H., Sediqi, A., 2010. Sticky substance on eggs improvespredation success and substrate adhesion in newly hatched Zelusrenardii (Hemiptera: Reduviidae) instars. Ann. Entomol. Soc.Am. 103, 771–774.
Litsios, G., Salamin, N., 2012. Effects of phylogenetic signal onancestral state reconstruction. Syst. Biol. 61, 533–538.
Liu, H., Zhao, P., Li, S., Cai, W., 2009. Taxonomic status ofVelinoides Matsumura (Hemiptera: Reduviidae: Harpatorinae)inferred from mitochondrial and nuclear genes. Zootaxa 2080,55–68.
Maddison, W.P., 1991. Squared-change parsimony reconstructionsof ancestral states for continuous-valued characters on aphylogenetic tree. Syst. Zool. 40, 304–314.
Maddison, W.P., Maddison, D.R., 2011. Mesquite: a modularsystem for evolutionary analysis. Version 2.75. Available at:http://mesquiteproject.org.
Mahler, D.L., Revell, L.J., Glor, R.E., Losos, J.B., 2010. Ecologicalopportunity and the rate of morphological evolution in thediversification of Greater Antillean anoles. Evolution 64, 2731–2745.
Maldonado, J., 1990. Systematic catalogue of the Reduviidae of theworld (Insecta: Heteroptera). Carib. J. Sci., Spec. Edn No. 1.Mayaguez, Puerto Rico.
Martin, C.H., Wainwright, P.C., 2011. Trophic novelty is linked toexceptional rates of morphological diversification in twoadaptive radiations of Cyprinodon pupfish. Evolution 65, 2197–2212.
O’Meara, B.C., Ane, C., Sanderson, M.J., Wainwright, P.C., 2006.Testing for different rates of continuous trait evolution usinglikelihood. Evolution 60, 922–933.
Miller, N.C.E., 1952. Three new subfamilies of Reduviidae. EOS,Revista Espa~nola de Entomolog�ıa 28, 85–90.
350 G. Zhang & C. Weirauch / Cladistics 30 (2014) 339–351

Moen, D.S., Wiens, J.J., 2009. Phylogenetic evidence forcompetitively-driven divergence: body-size evolution in Caribbeantreefrogs (Hylidae: Osteopilus). Evolution 63, 195–214.
Moore, B.R., Donoghue, M.J., 2009. A Bayesian approach forevaluating the impact of historical events on rates ofdiversification. Proc. Natl Acad. Sci. USA 106, 4307–4312.
Nixon, K.C., 1999. The parsimony ratchet, a new method for rapidparsimony analysis. Cladistics 15, 407–414.
Price, S.A., Wainwright, P.C., Bellwood, D.R., Kazancioglu, E.,Collar, D.C., Near, T.J., 2010. Functional innovations andmorphological diversification in parrotfishes. Evolution 64, 3057–3068.
Price, S., Holzman, A.R., Near, T.J., Wainwright, P.C., 2011. Coralreefs promote the evolution of morphological diversity andecological novelty in labrid fishes. Ecol. Lett. 14, 462–469.
Purvis, A., 1995. A composite estimate of primate phylogeny. Philos.Trans. R. Soc. Lond. B, Biol. Sci. 348, 405–421.
Ree, R.H., 2005. Detecting the historical signature of keyinnovations using stochastic models of character evolution andcladogenesis. Evolution 59, 257–265.
Schluter, D., 2000. The Ecology of Adaptive Radiation. OxfordUniversity Press, Oxford.
Schuh, R., 2012. Integrating specimen databases and revisionarysystematics. ZooKeys 209, 255–267.
Simpson, G.G., 1953. The Major Features of Evolution. ColumbiaUniversity Press, New York.
Slowinski, J.B., Guyer, C., 1993. Testing whether certain traits havecaused amplified diversification–an improved method based on amodel of random speciation and extinction. Am. Nat. 142, 1019–1024.
St�al, C., 1859. Till k€annedomen om Reduvini. Ofv. K. Vet.-Ak.F€orhandl. 16, 175–204.
Stamatakis, A., 2006. RAxML-VI-HPC: maximum likelihood-basedphylogenetic analyses with thousands of taxa and mixed models.Bioinformatics 22, 2688–2690.
Stamatakis, A., Hoover, P., Rougemont, J., 2008. A fastbootstrapping algorithm for the RAxML web-servers. Syst. Biol.57, 758–771.
Swofford, D.L., Maddison, W.P., 1987. Reconstructing ancestralcharacter states under Wagner parsimony. Math. Biosci. 87, 199–229.
Thomas, G.H., Freckleton, R.P., Szekely, T., 2006. Comparativeanalysis of the influence of developmental mode on phenotypicdiversification rates in shorebirds. Proc. R. Soc. Lond. B, Biol.Sci. 273, 1619–1624.
Tian, X., Xie, Q., Li, M., Gao, C., Cui, Y., Xi, L., Bu, W., 2011.Phylogeny of pentatomomorphan bugs (Hemiptera–Heteroptera:Pentatomomorpha) based on six Hox gene fragments. Zootaxa2888, 57–68.
Vaidya, G., Lohman, D.J., Meier, R., 2011. SequenceMatrix:concatenation software for the fast assembly of multi-genedatasets with character set and codon information. Cladistics 27,171–180.
Villiers, A., 1983. H�emipt�eres Reduviidae africains. Localisations etdescriptions. 4. Harpactorinae, Harpactorini. Revue Francaised’Entomologie (Nouvelle Serie) 5, 10–14.
Villiers, A., 1982. H�emipt�eres Reduviidae africains. Localisations etdescriptions. 3. Harpactorinae, Rhinocorini. Rev. Franc.Entomol. (Nouvelle Serie) 4, 126–136.
Wainwright, P.C., 2007. Functional versus morphological diversityin macroevolution. Annu. Rev. Ecol. Evol. Syst. 38, 381–401.
Weirauch, C., 2006. Observations on the sticky trap predator Zelusluridus St�al (Heteroptera: Reduviidae: Harpactorinae), with thedescription of a novel gland associated with the female genitalia.Denisia 50, 1169–1180.
Weirauch, C., 2008. Cladistic analysis of Reduviidae (Heteroptera:Cimicomorpha) based on morphological characters. Syst.Entomol. 33, 229–274.
Weirauch, C., Munro, J.B., 2009. Molecular phylogeny of theassassin bugs (Hemiptera: Reduviidae), based on mitochondrialand nuclear ribosomal genes. Mol. Phylogenet. Evol. 53, 287–299.
Wolf, K.W., Reid, W., 2001. Surface morphology of legs in theassassin bug Zelus longipes (Hemiptera: Reduviidae): a scanningelectron microscopy study with an emphasis on hairs and pores.Ann. Entomol. Soc. Am. 94, 457–461.
Yoder, J.B., Des Roches, S., Eastman, J.M., Gentry, L., Godsoe,W.K.W., Hagey, T., Jochimsen, D., Oswald, B.P., Robertson, J.,Sarver, B.A.J., Schenk, J.J., Spear, S.F., Harmon, L.J., 2010.Ecological opportunity and the origin of adaptive radiations. J.Evol. Biol. 23, 1581–1596.
Zhang, G., Weirauch, C., 2013. Sticky predators: a comparativestudy of sticky glands in harpactorine assassin bugs (Insecta:Hemiptera: Reduviidae). Acta Zool. 94, 1–10.
Supporting Information
Additional Supporting Information may be found inthe online version of this article:Table S1. Molecular protocols and primers.Table S2. Specimen voucher information and Gen-
Bank accession numbers.Table S3. Fore femoral measurements and observa-
tions of glands and setae.Figure S1. Best likelihood phylogeny from a parti-
tioned analysis using RAxML.Figure S2. Strict consensus of 85 equally most parsi-
monious trees from a TNT analysis.Figure S3. Ancestral state reconstructions of the
gland and setal characters based on the parsimonystrict consensus.Figure S4. Three most parsimonious reconstructions
of the evolution of lamellate setae based on the maxi-mum likelihood phylogeny.Figure S5. Character evolution of fore femoral
thickness and relative length mapped on the maximumlikelihood phylogeny using the linear parsimony modelas implemented in Mesquite.
G. Zhang & C. Weirauch / Cladistics 30 (2014) 339–351 351

Copyright of Cladistics is the property of Wiley-Blackwell and its content may not be copiedor emailed to multiple sites or posted to a listserv without the copyright holder's expresswritten permission. However, users may print, download, or email articles for individual use.


















