Snail Races Snail Races Using the Scientific Method and the Metric System.

THE JOURNAL OF BIOLOGICAL CHEMIBTRY Vol. 245, No. 24, Issue of December 25, pp. 6634-6641, 1970
Printed in U.S.A.
Carbamyl Phosphate Synthesis in a Land Snail, Strophocheilus oblongus*
(Received for publication, August 17, 1970)
PAUL R. TRAMELLS AND JAMES W. CAMPBELLS
From the Department of Biology, William Marsh Rice University, Houston, Texas 77001
SUMMARY
A carbamyl phosphate synthetase has been detected and partially characterized in hepatopancreas tissue of the land snail Strophocheitus oblongus. The enzyme is localized in mitochondria, utilizes L-glutamine, and shows an absolute requirement for N-acetyl-L-glutamate. This unique combi- nation of properties distinguishes the snail enzyme from either of the previously described animal carbamyl phosphate synthetases, carbamyl phosphate synthetase I and carbamyl phosphate synthetase Il. The snail enzyme was measured as the fixation of 14C-bicarbonate into citrulline in the pres- ence of excess L-ornithine and ornithine transcarbamylase. ATP and Mg2+ are also required in the reaction; optimal activity is obtained when the Mg2+:ATP ratio is 2. When this ratio is maintained, saturation with ATP occurs at ap- proximately 5 111~. The K, of the enzyme for L-glutamine is estimated to be 2.5 mu and for N-acetylglutamate, 0.3 mu.
The major portion of carbamyl phosphate synthetase ac- tivity is localized in the mitochondrial fraction of the snail tissue and, since this is also the fraction iu which ornithine transcarbamylase occurs, the enzyme is presumed to function in arginine biosynthesis. Aspartate transcarbamylase oc- curs in the cytosol of the tissue but the amount of carbamyl phosphate synthetase that can be measured in this fraction is negligible. The snail carbamyl phosphate synthetase may thus also function in pyrimidine biosynthesis.
Carbamyl phosphate, the initial compound in both arginine and pyrimidine biosynthesis, is formed in vertebrate animals by two distinct enzymes, carbamyl phosphate synthetase I and carbamyl phosphate synthetase II. Carbamyl phosphate syn- thetase I (EC 2.7.2.5) utilizes NH,, COZ, and ATP for carbamyl phosphate synthesis and requires N-acetyl+glutamate as a cofactor. Carbamyl phosphate synthetase II utilizes L-gluta-
* This invest.igation was supported by United States Public Health Service Grants AI-05006 and 5-TI-GM-884 and National Science Foundation Grant GB-8172.
$ Present address, Department of Pharmacology, Stanford University School of Medicine, Stanford, California 94305.
$ To whom inquiries should be sent. Recipient of United States Public Health Service Career Development Award 5-K3- GM-6780.
mine, COZ, and ATP, and does not require a cofactor. Enzyme I occurs in mitochondria and Enzyme II, in the cytosol or soluble cellular fraction. The mitochondrial localization of Enzyme I along with ornithine transcarbamylase (EC 2.1.3.3) indicates that its metabolic function is to provide carbamyl phosphate for arginine and urea biosynthesis, although carbamyl phosphate formed intramitochondrially may also be accessible to extramito- chondrial enzyme systems (1). The extramitochondrial local- ization of Enzyme II along with aspartate transcarbamylase (EC 2.1.3.2) indicates that the glutamine enzyme functions in tivo to provide carbamyl phosphate for pyrimidine biosyn- thesis (2). Enzyme II, but not Enzyme I, occurs in tissues capable of pyrimidine, but not arginine synthesis de novo (3-6).
With the exception of a carbamyl phosphate synthetase I-like enzyme that has been reported in the soluble cellular fraction of earthworm gut tissue (7)) the mechanism of carbamyl phosphate synthesis in invertebrate animals is unknown, although tracer studies in vivo with several species indicate that carbamyl phos- phate synthesis does occur (8). In certain land snails, for example, 14C-bicarbonate is incorporated into the ureido-C of citrulline and the guanidino-C of arginine (9, 10) and into the pyrimidine nucleotides UMP and CMP (11). Attempts to demonstrate either a carbamyl phosphate synthetase I- or carbamyl phosphate synthetase II-like enzyme in these species have previously yielded negative or inconclusive results (8). The work reported here was therefore directed toward the detection and characterization of the enzyme or enzymes re- sponsible for carbamyl phosphate synthesis in the land snail Strophocheilus oblongus.
MATERIALS AND METHODS
Animals-Specimens of S. oblongus were obtained from Porto Alegre, Brazil, through the courtesy of Dr. C. P. Jaeger of the Universidade do Rio Grande do Sul. They were kept under humid conditions and fed lettuce ad lititum.
ChemicatsL-Glutamine was obtained from Sigma and sodium l*C-bicarbonate from New England Nuclear. Sodium bicar- bonate was added to the 14C-compound to adjust the specific radioactivity to 1 PCi per pmole. N-Acyl amino acids and other substrates were also obtained from Sigma with the exception of N-acetyl+glutamine, N-acetyl+cysteine, and n-biotin which were obtained from Mann Research. The sources of the gluta- mine analogues were as follows : azaserine, S. W. Cancer Chemo- therapy Study Group Headquarters, M. D. Anderson Hospital, Houston, Texas; 0-carbamyl-L-serine, Cycle Chemicals, Los
6634
by guest on March 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Issue of December 25, 1970 P. R. Tramell and J. W. Campbell
FIG. 1. Electron micrograph of Strophocheilus hepatopancreas mitochondrial fraction prepared as described under “Materials and Methods.” The KCI-washed pellet was fixed in Millonig’s solution (13) and embedded in Epon 812. X 45,000.
Angeles, California, azotomycin, Cancer Therapy Evaluation Branch, National Institutes of Health, Bethesda, Maryland. Avidin was obtained from Boehringer Xannheim, and cetyltri- methylammonium bromide, from Eastman Organic Chemicals, Rochester, New York. Other inorganic and organic reagents were Fisher certified reagents.
EnxymesPyruvic kinase was obtained from Calbiochem. The ornithine transcarbamylase used as a supplementary enzyme was either purified from rat liver mitochondria through the second ammonium sulfate step as described by Caravaca and Grisolia (12) or was a preparation from Sfrepfmoccus D furnished by Dr. S. H. Bishop, Department of Biochemistry, Baylor College of Medicine, Houston, Texas. The specific enzyme activity of the rat enzyme was 20 units per mg of protein and, of the bacterial enzyme, 1200 units per mg. A unit of enzyme activity corresponds to 1 pmole of product per min at 30”. In experiments in R-hich the effect of NH4f on enzyme activity was tested, endogenous NH4+ in the supplementary enzyme prepara- tions was removed by dialysis against 50 mM Tris-chloride, pH 7.4, or by chromatography on Sephadex G-25 equilibrated with this buffer.
Preparation of Enzyme Extract-Hepatopancreas tissue was homogenized in 9 volumes of 0.25 BI sucrose by hand using a TenBroeck homogenizer kept cold in crushed ice. The homog- enate was centrifuged at 600 x g for 10 min. This and subse- quent centrifugations were at O-4”. The supernatant fluid was collected and centrifuged at 6,600 x g for 15 min. The sediment, which contained the mitochondria, was resuspended in 0.154 tf
KCl. This supension was then centrifuged at 20,000 x g for 20 min. The composition of the mitochondrial fraction pre- pared in this manlier is shown in Fig. 1. To solubilize the enzyme, the KCl-washed mitochondrial fraction was suspended in a solution containing 1 y0 (w/v) cetyltrimethylammonium bromide, 20y0 (v/v) glycerol, and 1 mM dithiothreitol. The volume of this solution was such that 1 ml of the suspension contained the mitochondrial fraction from 1 g of tissue. The suspended mitochondria were subjected to two 30-set pulses in a Bronwill 13iosonik II sonic disrupter operating at maximum output intensity. During sonic disruption, the suspension was kept cold in an ice-water mixture. After sonic disruption, the solution was centrifuged at 48,000 x g for 20 min. The resulting supernatant fluid was used as the enzyme source. The solu- bilized enzyme retained 100% of its activity when kept at - 15” for 11 days; 40yc of the original activity was lost during the same time period at 2-5”. Both glycerol and dithiothreitol were necessary to ret.ain enzyme activity (5, 14-16).
Measurement of Enzyme Activity-Clarbamyl phosphate syn- t.hesis was measured as the formation of i%Lcitrulline in a coupled assay system containing n-ornithine and ornithine transcarbam- ylase. The reaction was performed in thick walled, 12-ml centrifuge tubes maintained at 30” in a water bath. The reac- tion mixture for the basic assay system contained, in micromoles per 1 ml: ATP (sodium salt, pH 7), 5; l\lgSO+ 10; n-glutamine, 10; sodium l*C-bicarbonate (1 PC1 per pmole), 10; N-acetyl-n- glutamate (sodium salt, pH 7.5), 5; n-ornithine (n-ornithine hydrochloride adjusted to pH 7.4 with NaOH), 10; Tris-chloride,
by guest on March 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

6636 Carbamyl Phosphate Synthesis Vol. 245, No. 24
TABLE I General requirements for carbamyl phosphate synfhetase activity
The basic assay system was as described in the text with the indicated modifications. Incubation was for 20 min using 0.1 ml of the mitochondrial extract.
Assay system modifications Enzyme activity
dgm incorporatea into citr?l&ze
None......................................... Minus L-glutamine............................ Minus L-glutamine; plus 12.5 mM NH&l Minus N-acetyl-L-glutamate Minus L-glutamine and N-acetyl-L-glutamate;
20,000 108
2,570 1%
plus 12.5 mu NH&l 2,140 Minus ATP................................... 14 Minus L-ornithine............................. 450 Minus enzyme. 56 Zero time incubation.. 63
pH 7.4, 50; sodium phosphoenolpyruvate, 2.5; pyruvate kinase, 6.6 units; ornithine t&ntiearbamylase, 2 to 3 units; and 0.1 ml of Strophocheilus enzyme preparation. The reaction mixture components were mixed and kept at 0” until used. After they had equilibrated at 30”, the reaction was started by the addition of the mitochondrial extract. The reaction was stopped after 20 min by the addition of 0.25 ml of 30% (w/v) trichloracetic acid. After centrifuging to remove the protein precipitate, the supernatant fluid was gassed with CO2 to remove unreacted 14C- bicarbonate. Fifty microliters of the solution were then spotted on Whatman No. 3MM paper. ‘SC-Citrulline was separated from the other compounds by ascending chromatography using n-propyl alcohol-NHS-water (70:30, v/v). After drying, the citrulline area on the chromatogram was located by reference to a citrulline standard detected with p-dimethylaminobenzaldehyde (Ehrlich’s reagent). The area containing the 14C-citrulline was cut out and placed in a standard scintillation mixture (0.5% 2,5- diphenyloxazole and 0.05% 1,4-bis[2-(4-methyl-5-phenyloxa- zolyl)Jbenzene in toluene) for counting. The efficiency of count- ing was determined by the channels-ratio method (17). Enzyme activity is expressed as disintegrations per min incorporated into citrulline; 2.2 x lo3 dpm therefore corresponds approxi- mately to the formation of 1 nmole of carbamyl phosphate.
With this assay system, there was a linear incorporation of 14C into citrulline that was both time- and enzyme concentration-de- pendent under the experimental conditions used.
From 96 to 100% of the 14C measured in the isolated citrulline could be recovered as 14C02 following arsenolysis (18). For these measurements, the 14C-citrulline was eluted from the chromato- gram with water and incubated in 2 ml of a mixture containing 100 pmoles of sodium arsenate, pH 5.8, 2 pmoles of L-citrulline (added as carrier), and 200 units of rat liver ornithine transcar- bamylase. This incubation was in a closed flask and, at the end of 60 min, 1 ml of 5 N H&O4 was injected into the flask to liberate the 14C02 formed. The liberated 14C02 was trapped in 0.2 ml of methanolic p-(diisobutylcresoxyethoxyethyl)dimethylbenzylam- monium hydroxide which was contained in a polyethylene center well. The center well and its contents were transferred to the standard scintillation mixture for counting. The disappearance of citrulline from the reaction mixture, as measured colorimetri- tally (19), paralleled the liberation of 14C02. In preliminary ex- periments in which the 14C-citrulline produced by the enzyme
reaction was isolated by ion exchange chromatography (3), arsen- olysis yielded only about 80% recovery of the incorporated 14C. The non-citrulline component was identified as 14C-aspartic acid by paper and automated ion exchange chromatography and was found to contain 16% of the total incorporated 14C.
Measurements of Other EnzymesOrnithine transcarbamylase was measured in a reaction mixture containing, in l-ml volume: 10 pmoles of L-ornithine (the hydrochloride adjusted to pH 8.5 with NaOH); 50 pmoles of sodium glycylglycinate, pH 8.5; 10 pmoles of dilithium carbamyl phosphate; and the enzyme source as described. The same reaction mixture was used for aspartate transcarbamylase except that L-aspartate was substituted for L-ornithine. The reaction was started with carbamyl phosphate and stopped by adding 0.5 M perchloric acid. The citrulline or carbamyl aspartate formed was measured calorimetrically (19, 20).
Glutamine synthetase activity was estimated in a reaction mix- ture containing, in l-ml volume: 10 pmoles of ATP (sodium salt, pH 7) ; 10 pmoles of MgC12; 100 pmoles of Tris-chloride, pH 7.2; 25 pmoles of NH&l; 2 pmoles of 3, 4-14C-nL-glutamic acid (4.3 x lo5 dpm per pmole) ; and 0.1 ml of the same mitochondrial extract used for the assay of carbamyl phosphate synthetase activity. The reaction was stopped by adding 0.5 ml of 30% (w/v) tri- chloracetic acid. After removing the protein precipitate by centrifuging, 50 ~1 of the deproteinized solution were spotted on Whatman No. 3MM paper and developed with 75% (w/w) phenol in an ammoniacal atmosphere. The glutamine area on the chromatogram was located by reference to a glutamine stand- ard detected with ninhydrin and was cut out and counted as de- scribed above for citrulline. Incorporation of 14C into glutamine was linear with both time of incubation and enzyme concentration under the conditions used and was dependent upon the addition of both NH4+ and ATP to the reaction mixture.
Protein Measurement-Protein was estimated by the method of Lowry et al. (21) using dry bovine serum albumin as the standard.
RESULTS
The general requirements for carbamyl phosphate synthetase activity in Strophocheilus hepatopancreas tissue are shown in Table I. Maximal activity is dependent upon the addition of L-glutamine, N-acetyl+glutamate, ATP, and L-ornithine as an accessory substrate for the conversion of carbamyl phosphate to citrulline. The extent of dependence upon these compounds varies somewhat from one enzyme preparation to the other. This is also true of the activity found when NH,+ is used as the amino donor (see below).
The level of carbamyl phosphate synthetase activity in Stropho- cheilus hepatopancreas tissue ranges from around 150 to 500 nmoles of carbamyl phosphate formed per g per hour at 30”. This is higher than most extrahepatic mammalian tissue but falls within the range reported for several tumor tissues (6).
L-Glutamine Requirement-Evidence that L-glutamine is the substrate for carbamyl phosphate synthesis includes the inhibi- tion of the reaction by L-glutamine analogues (Table II). In concentration ratios with L-glutamine of 50: 1, azaserine and 0-carbamyl-L-serine inhibit the Strophocheilus enzyme 20% and 80%, respectively. Somewhat similar results have been obtained using both of these inhibitors with the Ehrlich ascites carcinoma enzyme (3).
The effect of L-glutamine concentration on Strophocheilus car- bamyl phosphate synthetase is shown in Fig. 2. Saturation oc-
by guest on March 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Issue of December 25, 1970 P. R. Tramell and J. W. Campbell 6637
TABLE II Inhibition of carbamvl phosphate synthesis by glutamine analogues
The basic assay procedure was modified as follows: the n-glu- tamine concentration was decreased to 0.5 rnr,r and the reaction was started by the addition of L-glutamine and the inhibitor. Incubation was for 20 min using 0.1 ml of the enzyme preparation.
Assay system modification
Experiment 1 None......................... Minus N-acetyl+glutamate Plus azaserine.. . . . . . Plus azaserine . . . . . . . . . Plus azaserine.. . . . . . Plus azaserine.. . . . . . . Plus 0-carbamyl-L-serine Plus 0-carbamyl-n-serine Plus 0-carbamyl-L-serine . . Plus 0-carbamyl-n-serine . .
Experiment 2 None......................... Minus N-acetyl-L-glutamate Plus azotomycin.............. Plus azotomycin.............. Plus azotomycin.............. Plus azotomycin..............
-
c I
-_
-
Final inhib- itor
oncen- :ration
?nM
0.5 1.0
10.0 25.0
0.5 1.0
10.0 25.0
0.5 1.0
10.0 25.0
-
: t
.-
-
i-Gh- amine to in- dbitor ratio
EIlZ.yIlX activity
14,700 950
15,200 15.300 13,200
8,300 15,200 13,800 11,000
4,670
35,300 450
35,900 35,800 35,400 27,620
%
0 0
10.1 43.7
0 6.2
25.5 68.3
0 0 0
21.8
Glutomih (mM)
I-L I I 3 4 5 6 7 8 9 IO
L-Glutamine (mM)
FIG. 2. Effect of n-glutamine concentration on the carbamyl phosphate synthetase reaction. The basic assay system in which the L-glutamine concentration was varied as indicated was used. Incubation at each substrate concentration was for 20 min with 0.1 enzyme preparation.
curs around 5 mu glutamine. A Lineweaver-Burk plot of the data indicates substrate inhibition at higher concentration. The K,,, for L-glutamine, estimated by extrapolation from such a plot (---, Fig. 2), is around 2.5 InM, which is considerably higher than that of carbamyl phosphate synthetase II. The reported K, values of carbamyl phosphate synthetase II for L-glutamine are around 0.01 mM (3, 6, 22). The higher K, of the snail en- zyme would not, however, render it nonfunctional in wivo since r,-glutamine is a major free amino acid in snail tissues, occurring in concentrations of from 1 to 2 pmoles per g (23).
6.4 7.2 8.0 8.0
PH
FIG. 3. Effect of pH on carbamyl phosphate synthesis from L-glutamine ( l ; 5 maa) and NH&l (0 ; 25 m@. The basic assay system was used except that L-glutamine was omitted when NH&l was added. Incubation was for 20 min with 0.1 ml of the enzyme preparation.
Because the snails are herbivorous and therefore ingest con- siderable L-asparagine in their diets, this compound was also tested as a substrate for the snail enzyme. This amide would not, however, substitute for L-glutamine as an amino donor: less than 2% of the glutamine activity was obtained with 25 mM as- paragine.
Ammonia also does not appear to be a substrate for Xtropho- cheilus carbamyl phosphate synthetase, although the nature of the reaction responsible for the small amount of 14C-bicarbonate fixation in the presence of NH,+ (Table I) is not known. The enzyme preparation does have glutamine synthetase activity but the measured level of this enzyme (less than 0.2 nmoles per 0.1 ml per 20 min) is not sufficient to account for the amount of fixation observed in the presence of NH4+, assuming there were endogenous glutamate present in the preparation. The residual activity with NH4+ is also not generally dependent upon added acetylglutamate. When NH4+-NH3 is used as substrate for the snail enzyme at 5 times the saturating glutamine concentration in the pH range 6.5 to 9.0 there is no optimum shown between pH 7 and 8 (Fig. 3). This is in contrast to the results obtained with carbamyl phosphate synthetase II which does utilize NH3 as a substrate (3,6). The pH optimum for the snail enzyme with glutamine as the substrate is around 7.8 as shown in Fig. 3.
N-A@ Amino Acid Requirement-As shown in Table III, of the derivatives tested, N-acetyl-L-glutamate is the most effective cofactor for the snail enzyme. Carbamylglutamate, N,-acetyl- glutamine, and acetylaspartate are also utilized, but to a lesser extent than acetylglutamate. With acetylglutamate, the en- zyme shows typical saturation kinetics when the concentration of this cofactor is varied between 0.125 and 10 mM (Fig. 4). The K, of the enzyme for acetylglutamate, estimated from these data, is 2.5 X 10m4 M.
The requirement for acetylglutamate is also shown by the en- zyme present in “intact” mitochondria. The requirement of carbamyl phosphate synthetase I for acetylglutamate can be satisfied in freshly prepared mammalian mitochondria by an oxidizable substrate (24), presumably because of endogenous syn- thesis of this cofactor. The physiological integrity of the Strophocheilus mitochondria was not determined but, as shown in
by guest on March 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from
6638 Carbamyl Phosphate Synthesis Vol. 245, No. 24
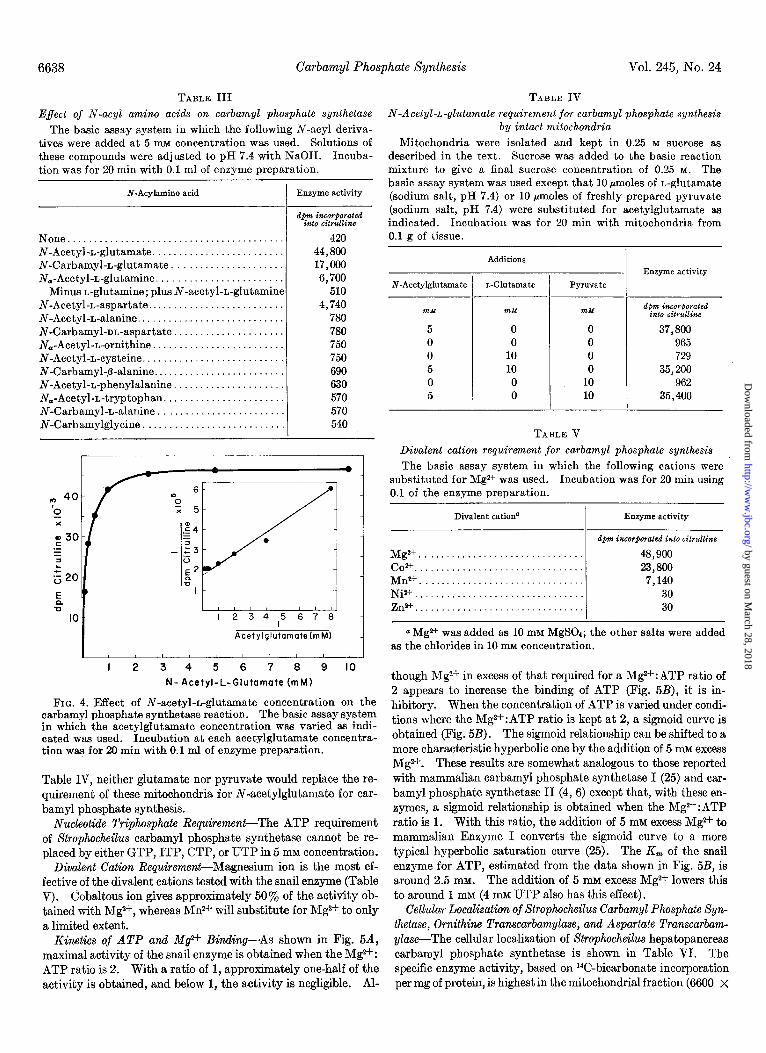
TABLE III Effect of N-acyl amino acids on carbamyl phosphate synthetase
The basic assay system in which the following N-acyl deriva- tives were added at 5 mM concentration was used. Solutions of these compounds were adjusted to pH 7.4 with NaOH. Incuba- tion was for 20 min with 0.1 ml of enzyme preparation.
N-Acylamino acid Enzyme activity
apm incorgoratea i&O &rullilze
None ......................................... 420 N-Acetyl-L-glutamate ......................... 44,800 N-Carbamyl-L-glutamate ..................... 17,000 N,-Acetyl-L-glutamine. ....................... 6,700
Minus L-glutamine; plus N-acetyl+glutamine 510 N-Acetyl-L-aspartate .......................... 4,740 N-Acetyl-L-alanine ............................ 780 N-Carbamyl-m-aspartate ..................... 780 N,-Acetyl-L-ornithine ......................... 750 N-Acetyl-L-cysteine. .......................... 750 N-Carbamyl-fi-alanine ......................... 690 N-Acetyl-L-phenylalanine ..................... 630 N,-Acetyl-L-tryptophan. ...................... 570 N-Carbamyl-L-alanine ........................ 570 N-Carbamylglycine ........................... 540
r I
Acetylglutomate(mM)
I I I I I
I 2 3 4 5 6 7 8 9 IO N- Acetyl-L-Glutamate (mM)
FIG. 4. Effect of N-acetyl-L-glutamate concentration on the carbamyl phosphate synthetase reaction. The basic assay system in which the acetylglutamate concentration was varied as indi- cated was used. Incubation at each acetylglutamate concentra- tion was for 20 min with 0.1 ml of enzyme preparation.
Table IV, neither glutamate nor pyruvate would replace the re- quirement of these mitochondria for N-acetylglutamate for car- bamyl phosphate synthesis.
Nucleotide Triphosphate Requirement-The ATP requirement of Strophocheilus carbamyl phosphate synthetase cannot be re- placed by either GTP, ITP, CTP, or UTP in 5 mm concentration.
Divalent Cation Requirement-Magnesium ion is the most ef- fective of the divalent cations tested with the snail enzyme (Table V). Cobaltous ion gives approximately 50% of the activity ob- tained with Mg”+, whereas Mn2+ will substitute for Mg2+ to only a limited extent.
Kinetics of ATP and Mg2+ Binding-As shown in Fig. 5A, maximal activity of the snail enzyme is obtained when the Mg2f: ATP ratio is 2. With a ratio of 1, approximately one-half of the activity is obtained, and below 1, the activity is negligible. Al-
TABLE IV N-Acetyl-L-glutamate requirement for carbamyl phosphate synthesis
by intact mitochondria
Mitochondria were isolated and kept in 0.25 M sucrose as described in the text. Sucrose was added to the basic reaction mixture to give a final sucrose concentration of 0.25 M. The basic assay system was used except that 10 pmoles of L-glutamate (sodium salt, pH 7.4) or 10 pmoles of freshly prepared pyruvate (sodium salt, pH 7.4) were substituted for acetylglutamate as indicated. Incubation was for 20 min with mitochondria from 0.1 g of tissue.
Additions Enzyme activity
N-Acetylglutamate L-Glutamate Pyruvate
n&M m&l %zM d~?n i?mrporated into citrulline
5 0 0 37,800 0 0 0 965 0 10 0 729 5 10 0 35,200 0 0 10 962 5 0 10 35,400
TABLE V Divalent cation requirement for carbamyl phosphate synthesis The basic assay system in which the following cations were
substituted for Mgz+ was used. Incubation was for 20 min using 0.1 of the enzyme preparation.
Divalent cationa Enzyme activity -
dpm incor@rated into citrullinc
Mgz+ ............................ 48,900 Co” ............................. 23,800 MrP+ ............................ 7,140 Ni* ............................. 30 Zn2+. ............................ 30
aMg* was added as 10 mM MgSOd; the other salts were added as the chlorides in 10 mM concentration.
though Mg2+ in excess of that required for a Mg2+:ATP ratio of 2 appears to increase the binding of ATP (Fig. 5B), it is in- hibitory. When the concentration of ATP is varied under condi- tions where the Mg 2+:ATP ratio is kept at 2, a sigmoid curve is obtained (Fig. 5B). The sigmoid relationship can be shifted to a more characteristic hyperbolic one by the addition of 5 mM excess Mg”. These results are somewhat analogous to those reported with mammalian carbamyl phosphate synthetase I (25) and car- bamyl phosphate synthetase II (4, 6) except that, with these en- zymes, a sigmoid relationship is obtained when the Mg2+:ATP ratio is 1. With this ratio, the addition of 5 mM excess Mg” to mammalian Enzyme I converts the sigmoid curve to a more typical hyperbolic saturation curve (25). The K, of the snail enzyme for ATP, estimated from the data shown in Fig. 5B, is around 2.5 MM. The addition of 5 mu excess Mg2+ lowers this to around 1 InM (4 mM UTP also has this effect).
Cellular Localization of Strophocheilus Carbamyl Phosphate Syn- thetase, Ornithine Transcarbamylase, and Aspartate Transcarbam- yZa.se--The cellular localization of Strophocheilus hepatopancreas carbamyl phosphate synthetase is shown in Table VI. The specific enzyme activity, based on 14C-bicarbonate incorporation per mg of protein, is highest in the mitochondrial fraction (6600 x
by guest on March 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Issue of December 25, 1970 P. R. I’ramell and J. W. Campbell 6639
n 40 ‘0 x Q) 30 .E =
E 20 .-
E
g IO
‘: 40 0 ;;
g 30 .- = 2
u 20 E a TI
IO
n 5mM ATP
LL 5 10 15 20 25 3*
c
A
Mg2+(mM)
-A
0
ATP(mM)
FIG. 5. Effect of Mg2+ and ATP concentrations on carbamyl phosphate synthetaae. Incubation was for 20 min with 0.1 ml of enzyme preparation in the basic assay system in which the concentrations of ATP and Mgz+ (as MgSOI) were varied as indi- cated. A, the ATP concentration was 5 mM and the Mg” con- centration was varied &s indicated; B, for O--O, the Mg*:ATP ratio was 2; and for O---O; 5 mM Mgz+ was added in excess of that required for a Mg 2+:ATP ratio of 2 at all ATP concentrations.
g residue). While some fixation is obtained in the soluble frac- tion, its dependence upon either added acetylglutamate or NH4+ is negligible. For the experiment shown in Table VI, 14C-citrul- line formed in the reaction was isolated by ion exchange chroma- tography. The high nonspecific fixation obtained may thus be due to fixation into aspartate. Additional evidence for the mito- chondrial localization of the snail enzyme is shown in Fig. 6. During sedimentation in a sucrose density gradient, the carbamyl phosphate synthetase activity coincides with that of cytochrome oxidase, an enzyme commonly used as a mitochondrial marker.
No more than 5% of the total ornithine transcarbamylase ac- tivity is localized in the soluble cellular fraction of Xtrophocheilus hepatopancreas tissue (Table VII). Although some activity is present in the residue obtained by low speed centrifugation (nuclei, cell debris, etc.) a major portion, around 60% of the total, occurs in the mitochondrial fraction. This is in contrast to the localization of the major portion of aspartate transcarbamylase in the tissue; approximately 70% of this latter activity is found in the soluble fraction obtained by high speed centrifugation, whereas only around 18% of the total aspartate transcarbamylase activity is present in the mitochondrial fraction. Qualitatively,
TABLE VI
Cellular localization of carbamyl phosphate synthetase activity
Hepatopancreaa tissue (5.7 g) was homogenized by hand in a TenBroeck homogenizer kept cold in ice in 9 volumes of 0.25 M sucrose. Centrifugations at 600 and 6,600 X g were for 30 min at o-4” in a Servall RCO-B centrifuge with a fixed-angle rotor (SS-34); centrifugations at 110,000 X g were the same except that a Beck- man Spinco model L centrifuge, also with a fixed-angle rotor (type 40)) was used. The residues were resuspended in a volume of 0.25 M sucrose equivalent to 0.1 the original homogenate volume; 0.2 to 0.4 ml of each fraction was assayed in the basic assay system, the modifications of which are indicated. When NH,+ was added, it was as 25 mM NH&l. W-Citrulline was isolated from the re- act.ion mixture by ion exchange chromatography (3).
Fraction
Homogenate (15.0)a. 600 X g
Residue (28.5) Supernatant fluid (11.3).
6,600 x g Residue (11.3). . Supernatant fluid (9.0).
110,000 x g Residue (5.6). Supernatant fluid (7.5).
Assay modifications in enzyme activity
bf%m~ Minus
M;;~“s Minus mine 0*p1-
gluta- and N- ‘iti? None acetyl- mine; acetyl- orni-
L-gluta- plus L-gluta- thine mate NHt+ mate; trans-
plus carbm- NHd+ YlW.
dpm incorporated into citrulline/mg protein
970 93 203 150 264
235 58 67 58 58 1480 129 216 131 433
8030 155
120 209
1240 1780 107 152
118 187 121 195
1470 148
184 239
489 37
13 55
a The protein content of each fraction as milligrams per ml is given in parenthesis.
o 161 ‘0 ! x I aa 12 1 .E =
i
1 0.08
t 15 13 II 9 7 5 3 f MENISCUS
Fraction No. BOTTOM
FIG. 6. Localization of carbamyl phosphate synthetase (0) and cytochrome oxidase (0) activities in Strophocheilus mito- chondria by density gradient centrifugation. Of the mitochon- drial fraction, 1 ml was layered onto a 49-ml gradient prepared with 2 M sucrose in the mixing chamber and 1 M sucrose in the reservoir (26). Centrifugation was for 45 min at 7000 X g in a Servall RCP-B centrifuge equipped with a swinging bucket rotor (HB-874). Fractions of approximately 2 ml were collected and assayed for carbamyl phosphate synthetase and cytochrome oxidase (27) activities.
by guest on March 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

T.YBLE VII
Cellular localization of transcarbamylase activity
Fr:lctionatjion of 4.5 g of hepatopancrcas tissue was as described in Table VI. Assays for transcarbamylnse activity were per- formed as described in the text using a heat-inactivated control (100” for 3 to 4 min) to correct, for spontarrrolls transcnrbamyla- tion and interfering substances.
Fraction
Homogetlnte. 600 x g:
Hesidue Supernatallt fluid.,
G,600 x g: Residue. Sllpernatant fluid..
110,000 x g: I:esidlle SI1perllatant fluid.,
I-
I
Enzyme activity
Ornithine : transcsrbamylase
Aspartate i transcarbamylase
Total % of Total unit9 total units
rmoles/hr
1080
308 059
c,22 (Xi. 4
17.6 53 0
3G.8 0 61.0 1.72
57. (i G 2
1.6 4.9
pnaoles/hr
1.92
0.34 1.M
0.22 1.34
97”
100
0 89.G
17.7 85.4
11.5 69.8
Tangos: VIII E$ecf oj pwine rind pyrimidine n/rcleo/itles on carbamyl
phosphate synthesis
The basic assay system was modified hy redtlcing the ATP con- cclltration t,o 2.5 m&r. The rrucleotides were added in 10 mu COW centrat,ion. Incubation was for 20 min with 0.1 ml of ellzyrne preparation.
Nucleotide added I Enzyme activity
None UTP AIll’ CTP AMP I’D GTP
llpnr incorpovated iuo citruzline
34,800 40,800 31,100 26,500 11,200 3,040
350
%
100 117 90 76 32 13
1
the distribution of the two transcarbamylascs is much the same as in mammalian tissue (28, 29).
Inhibition by Nucleotides-Since, if the Sfrophocheilus carbamyl phosphate synthetase were functioning in pyrimidine biosyn- thesis, it might be subject to feedback inhibition by pyrimidine nucleotides (30), the effect of t’hese compounds on its activity was tested. As shown in Table VIII, neither UTP nor CTP have a pronounced effect on the enzyme, although UTP gives a slight stimulation and CTP, a slight inhibition when these nucleotides are present in 10 rnlll concentration. UTP may possibly be an allosteric activator of the enzyme. In experiments identical to bhat described in Fig. 5B, the addition of 4 m>r UTP in place of 5 111x excess Mg2+ had t,he same effect as excess 1Ig*+ in convert- ing the sigmoid activity-concentration curve to a hyperbolic one, decreasing the apparent K, for ATP from 2.5 to 1 mM.
In contrast to the effect of the pyrimidine nucleotides, the purine nucleotides ARIP, ITP, and especially GTP markedly in- hibit the enzyme. Since ADP acts as an end product inhibitor
of carbamyl phosphat’e synthetase I (2.5, 31), thr inlril)it~iotl OF the snail enzyme by X;\IP 17-a’ of some interest. However, xttcmpts
to obtain an enzyme preparation free of interfering cnzpmc :LC- tivities with which the stoichiometry of ATP hydrolysis during the reaction could be measured have been unsurccq+ful. Since carbnmyl phosphate is not known to be inr-olvctl in purinc bio- synthesis, no phgyiological significance can 1~~ attributed t,o t,lre inhibitions given by CTP and ITI’ and they nlay, ill fact,, result from Xg2f binding. The effect of both ITl’ and G’I’P is conccn- tration dependent; in an experiment, identical to that, described in Table VIII, low concentrations of both IT1 and G’l’l’ gave progressive stimulation ul) to around 3 mv. Increasing the nucleotide concentrations above 3 mM resulted in progressive in- hibition of the enzyme. This concentration dependence ma> explain the various result,s obtained when these nuc*lcotides have been tested on other carbamyl phosphate sgnthetnses (5, 32).
Jliscellaneous Chmpoulzds-Phosphate, which at~imulatcs the Ehrlich ascites cell enzyme (3), is without effect on the snail ear- bamyl phosphate synthekise in 10 mM concent’ration. Sodium fluoride in 5 m&f concentration inhibits the enzyme around 4OC;. Fluoride (3 mM KF) completely inhibits the pea seedling enzyme (33)) although it stabilizes ngaricu,s carbamyl phosphate sy~l- thetase II (22). High concentrations of L-xrgininc (10 IIIM) in- hibit the snail enzyme around 27%). This inhibition by arginine is, however, unlikely to be of physiological aignificanc~c because of the generally low concent,rations of free argininc in the tissues of land snails (23). -%vidin and biotin, alolie and iii various combinations, have no effect on the snail cuzyme.
DISCUSSION
Previous :Lttempts to demonstrate either a carbamyl phosphate synthetase I- or carbamyl phosphate synthetase II-like enzyme in land snails were unsuccessful, although studies with intact or- ganisms and tissues indicated that carbamyl 1)hosphate was formed in vivo (8). The unique properties of the (Barbamy phos- phate synthetase in Strophocheilus hepatopancreas tissue may explain the negative results obtained using assay systcmstlesigned for either of the vertebrate enzymes. Although the siiail eii- zyme shares certain properties with carbamyl 1)hosphatc syn- thetase I and carbamgl phosphate synthetase II, its unique corn- bination of properties distinguishes it from either of these enzymes. The snail enzyme shares the proljerties of requiring 5-acety-L-glutamate and being a mitochondrial enzyme with vertebrate Enzyme I, the enzyme that provides carbamyl phos- phate for arginine and urea biosynthesis. However, unlike En- zyme I, the snail enzyme utilizes L-glutamine as a substrate and apparently does not utilize ammonia. Its utilization of gluta- mine is a property shared with carbamyl phosphate synthetaxe II from microorganisms (22, 30, 34, 35), nvian liver (4), and scvcral mammalian tissues and tumor cells (4-6). In vertebrates, the latter enzyme is localized in the cytosol and provides carbamyl phosphate for pyrimidine biosynthesis.
The properties the snail enzyme shares with carbamyl phos- phate synthetases I and II are qualitative and not necessarily quantitative ones. The k’, of the snail enzyme for r,-glutaminc is, for example, different by three orders of magnitude from that of carbamyl phosphate synthetase II, being 2.5 m&f versus 0.01 m&I for the latter (3, 6, 22). lJnlike Enzyme II, the snail car- bamyl phosphate synthetase does not ut’ilize ammonia in addit,ion to glutamine, although it is possible that the affinity for ammonia was lost during solubilization of the enzyme. The affinity for glutamine of the enzyme from Escherichia coli is much more labile
by guest on March 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

than that for ammonia and may be lost during purification of the enzyme (36). Uthough Enzyme II utilizes both glutamine and ammonia, the lower I<, for glutamine led to the conclusion that glutamine and not ammonia is the physiological substrate (e.g. 3, 6, 36). Sincte activity of the snail enzyme was only obtained with glutamine, it is not possible to evaluate the physiological significance of its high li, for ghltamine in terms of what is the normal substrate of the enzyme. Its inhibition by glutamine :malogucs, the failure of asparagine to function as a substrate, rind the occurrence of relatively high concentrations of free gluta- mine in the tissues of land snails are indirect evidence that glutaminc is the 1)hysiological substrate of the enzyme. The K, of the snail enzyme for acetylglutamate is also higher than that of vertebrate cnarbamyl phosphate synthetaxe I, being 0.3 mlr versus 0.03 to 0.06 mar for the latter (25, 31). The anomalous behavior of the Jnail enzyme toward .\TP and l\lg’f is similar to that shown by Enzymes I and II although, again, there are quantitative differences. Maximal activity of the snail enzyme is obtained when the ratio of Mg2+ to ATP is 2. With this ratio, the enzyme apl)ears to be saturated around .S mM ATP, which is about the same as that required by mammalian carbamyl l,hos- phatc synthetasc I (a;), but much less than that required by carbamyl phosphate synthetase II (4, 6). Varying the ATP concentration under conditions where the >Ig2+:ATP ratio is kel)t at 2 results in a sigmoid activity-concentration curve with the snail enzyme similar to that obtained with both Enzyme I and Enzyme II when the XIg2+:9TI’ ratio is 1. The addition of excess R4g2+ to both Enzyme I and the snail enzyme in the pres- ence of their respective Rig 2+:ATP ratios gives a more typical hyperbolic activit?--concelltration relation.+ip even though en- zyme activity is inhibited (25).
The mitochondrial localization of Strophockeilus carbamyl phosphate synthctase also serves to distinguish it from Enzyme II. In vertebrates, Enzyme II is localized along with aspartate transcarbamylase in the soluble cellular fraction or cytosol and is therefore felt to function in pyrimidine biosynthesis (2). Since the snail enzyme is localized along with ornit,hine transcarbamyl- ase in the mitoc+ondrial fraction, it presumably functions to pro- vide c~arbamyl phosphate for arginine and urea biosynthesis. The metabolism of some land snails is unique because they are the only lnuinotelic animals known that have retained the ability to s\-nthcsize arginine (8). In these snails, urea is formed from argininc even though it is subsequently hydrolyzed by a unique animal iirease rather than accumulat,ing for excretion as in ureo- telic species (37, 38). ‘G&carbonate is incorporated into nucleic acid 1)yrimidine nucleotides by the snails (11) and their possession of aspartate transcarbamylase indicates that their syn- thesis of pyrimidines is the same as in other species. The mito- chondrial carbamyl phosphate synthetase described here could thus also function in pyrimidine biosynthesis since intramito- chondrial c~rbamyl phosphate has been shown to be available t,o estramitocholldrial enzyme systems in vitro (1) and the glutamine enzymes from at least two bacteria are known to have dual func- tions (see Ilcl’erence 30).
REFERENCES
1. N.\T\ ~b:, I’. J., .YSD TRXMIIL.\Y, G. C., Wiochem. Biophys. Res. -. _- - ..-.,.
2.
3.
4.
5.
6.
7. 8.
9.
10.
11.
12.
13 14
15.
16 17.
18 19 20.
21
22 23
24.
25.
26. 27.
28.
29.
30.
31.
32.
33. 34.
35.
36.
H.IGER, S. E., ISD JOYES, M. 15.. J. Iliol. (‘/I(:uI., 242, 566: (1967).
YIP, M. C. M., .ca KNOX, W. E., J. Bid. Chc~r., 246, 2199 (1970).
BISHOP, S. H., .IP~D CAMPUELL, J. W., Scicnw, 142, 1583 (1963). C,IMPBELL, J. W., in B. SCHMIDYNIIX,SI~:S (I’:clil or), ~~UXI tint/
the kidney, Excerpta Medica Foundaiiolt. AIIIS~CI.~:LI~, 1970, D. 48.
CI\MPBELL, J. w., .1ND SPEEG, K. 5’., Jli., (:o?t/p. l~ioche,//. Physiol., 26, 3 (1968).
POI~I*:MBSK.I, Z., GORZKOWSIU, B., .\~n JI.~I.\\ SIC \, AI., .tctu Biochim. Pal., 13, 107 (1966).
C.IRAV.~C~, J., .IXD GRISOLII, S., J. Iliol. (I/w/r., 235, Ii84 (1960).
MILLONIG, G., J. Appl. Physics, 32, 1637 (19611. N~KANISHI, S., ITO, K., AND TATIBINI, &I., I~io~~he~~,. I~~o~IT~J,s.
Res. Commun., 33, 774 (1968). GUTII~HRLEIN, G., AND J~NAP~, J., Eur. J. IIio&ott., 7, 119
(1968). O’NEAL, II., .~ND N.~YLOR, A. W., 13iochenl. J., 113,271 (1969). BRUNO, G. A., AND CIIRISTIAN, J. E., iI r&. Cherrr., 33, G50
(1961). REICHARD, P., Acta Chem. Scantl., 11, 523 (1057). ARCHIBALD,~~~. M.,‘J. Biol. Chew!.. 166, 121 (1034). GERHART, J. C., AND PARDEE, A. B., J. Biol. Chew., 237, 891
(1962). LOWRY, 0. H., ROSEBROUGH, N. J., F.~rclt, A. T,., .INI) I:.IxT~.~LL,
R. J., J. Biol. Chem., 193, 265 (1951). LEVENBERG, B., J. Biol. Chem., 237, 2590 (1962) CAMPBELL, J. W., AND SPEEG, K. V., JJC., (YOIQ. Biochevr.
Physiol., 26, 3 (1968). CHARLES, R., TRAGOR, J. M., AND SLITI~, 15. C., Hiochitn.
Biophgs. Acta, 131, 29 (19671. KERS~N; L. A., 'AND-APP~L, s. II., J. Niol. Cltcn~., 243, 4270
(1968). BRITTEN, R. J., .IND Rom~n~s, R. I~., %icnc~, 131, 32 (1960). SMITH, L., in S. P. COLO~ICK .\XD N. 0. K.\PL\N (IjGlitors),
Methods in enzymology, Vol. 2, Academic I’I.CSS, No\v York, 1955, p. 732.
GRISOLIA, S., AND COHEN, P. P., J. /Go{. Ucorr., 198, 561 (1952).
LOWEN&EIN, J. M., AND COHEN, I'. P., J. Iliol. Clrc~l., 220, 57 (1956).
ISSALY, I. M., ISSALY, A. s., AND ~~ISFIC., -7. I,., I~io/~hiv~. Biophvs. Acta. 198, 482 (1970).
JON&, k E., in’S. P; COL~WIC~ .\XD N. 0. T<.IPL.IN (IlXtors), Methods in enzymology, Vol. 5, Acadrmic Ihss, New York, 1962, p. 903.
O’NUL, D., .IND NAYLOR, A. W., Uiodmt. fhph,p. 1i’c.q. Commun., 31, 322 (1968).
KLECZKOWSKI, K., Acta Biochim. Pal., 12, 243 (1965). PIER~RD, A., .4ND WIAME, J. &I., Niwhc~v~. J~ioph~~s. 1:r.c.
Commun., 16, 76 (1964). ~~~~~~~~~~~ F., PIERARD, A., GRENOS, RI., .INI) WI.\MI:, J. JI.,
J. Gen. iWicrobioZ., 40, 127 (1965). KALMAN, S. M., DUFFIELD, P. H., .\SD I~~tzozo\~~s~c~, T., 1.
BioZ. Chem., 241, 1871 (196G). 37. SPEEG, K. V., JR., AND C.IMPIEI.L, J. W., .I )u~I’. .I. ~lr!/,~iol.,
216, 1003 (1969). 38. MCDONALD, J. 8., BND C.\MPHI:I.L, J. Rv., Fctl. I'm., 29, !IO~
(19711~ Comm/Ln., 37, 512 (1YUY). \--.“,.
by guest on March 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Paul R. Tramell and James W. CampbellStrophocheilus oblongusCarbamyl Phosphate Synthesis in a Land Snail,
1970, 245:6634-6641.J. Biol. Chem.
http://www.jbc.org/content/245/24/6634Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/245/24/6634.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on March 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from