






Languages
Pages
Legal


A Mouse Model of Acinetobacter baumannii-Associated PneumoniaUsing a Clinically Isolated Hypervirulent Strain
Greg Harris,a Rhonda Kuo Lee,a Christopher K. Lam,b Gregory Kanzaki,b Girishchandra B. Patel,a H. Howard Xu,b Wangxue Chena,c
Vaccine Program, Human Health Therapeutics, National Research Council Canada, Ottawa, Ontario, Canadaa; Department of Biological Sciences, California StateUniversity, Los Angeles, Los Angeles, California, USAb; Department of Biology, Brock University, St. Catharines, Ontario, Canadac
Acinetobacter baumannii is an important emerging pathogen in health care-acquired infections and is responsible for severenosocomial and community-acquired pneumonia. Currently available mouse models of A. baumannii pneumonia show poorcolonization with little to no extrapulmonary dissemination. Here, we describe a mouse model of A. baumannii pneumonia us-ing a clinical isolate (LAC-4 strain) that reliably reproduces the most relevant features of human pulmonary A. baumannii infec-tion and pathology. Using this model, we have shown that LAC-4 infection induced rapid bacterial replication in the lungs, sig-nificant extrapulmonary dissemination, and severe bacteremia by 24 h postintranasal inoculation. Infected mice showed severebronchopneumonia and dilatation and inflammatory cell infiltration in the perivascular space. More significantly, 100% ofC57BL/6 and BALB/c mice succumbed to 108 CFU of LAC-4 inoculation within 48 h. When this model was used to assess the effi-cacy of antimicrobials, all mice treated with imipenem and tigecycline survived a lethal intranasal challenge, with minimal clini-cal signs and body weight loss. Moreover, intranasal immunization of mice with formalin-fixed LAC-4 protected 40% of micefrom a lethal (100� 100% lethal dose) intraperitoneal challenge. Thus, this model offers a reproducible acute course of A. bau-mannii pneumonia without requiring additional manipulation of host immune status, which will facilitate the development oftherapeutic agents and vaccines against A. baumannii pneumonia in humans.
Acinetobacter baumannii infection has recently emerged as amajor cause of health care-associated (hospital- and commu-
nity-acquired) infections worldwide (1). The overall 30-day mor-tality of Acinetobacter infection can be as high as 49%, with therespiratory tract being an important portal of entry (2). Indeed,the United States National Nosocomial Infections Surveillancedata indicate that Acinetobacter infections were responsible for 7%of intensive care unit (ICU) nosocomial cases of pneumonia in2003 (3). Moreover, A. baumannii infections have become in-creasingly difficult to treat because of the rapid development ofresistance to multiple antibiotics by the pathogen (4). There-fore, there is an urgent need for the development of novel ther-apeutics and other intervention strategies to combat this im-portant pathogen.
Animal models are crucial to the development of new thera-peutics and vaccines and play critical roles in the assessment of theefficacy and safety of the new products before they enter humanclinical trials. Over the years, a number of animal models of A.baumannii pneumonia have been developed, with the mouse be-ing the most widely used model (5). However, most laboratorystrains and clinical isolates of A. baumannii do not infect immu-nocompetent mice well and induce only a self-limiting pneumo-nia with no or very limited local bacterial replication and systemicdissemination, even when a large inoculum is used (6–10). Toovercome these shortcomings, several laboratories use immuno-compromised (such as neutropenic) mice or treat mice with mu-cin or agar to increase host susceptibility to A. baumannii andbacterial virulence. Despite their limitations, these models havebeen used for many years and have been instrumental in studies ofdisease pathogenesis and product development (7, 9, 11–21).However, these models fail to mimic the natural course of humaninfection and generally are not suitable for studying the host re-sponse to the infection or evaluating antimicrobials that involvethe host immune system.
The aim of this study was to evaluate the ability of severalclinical isolates and laboratory strains of A. baumannii to repro-ducibly infect commonly used laboratory mouse strains (BALB/cand C57BL/6) and to cause disease that would more closely resem-ble the spectrum of the human infections. We identified one clin-ical isolate (LAC-4) that caused 100% mortality in conventional,immunocompetent BALB/c and C57BL/6 mice when intranasally(i.n.) infected at a dose of 108 CFU. To test the utility of our model,we also assessed the efficacy of imipenem and tigecycline against alethal intranasal challenge of A. baumannii.
MATERIALS AND METHODSMice. Eight- to 12-week-old, specific-pathogen-free, male or femaleC57BL/6 and BALB/c mice were purchased from Charles River Laborato-ries (St. Constant, Quebec, Canada). The animals were maintained andused in accordance with the recommendations of the Canadian Councilon Animal Care Guide to the Care and Use of Experimental Animals, andexperimental procedures were approved by the institutional animal carecommittee.
A. baumannii isolates and species confirmation. The clinical isolatesand ATCC strains used in the study are listed in Table 1. The species of allLos Angeles County (LAC) isolates were further confirmed by DNA se-quencing analysis of the intergenic spacer (ITS) between 16S and 23SrRNA genes based on the methods of Chang et al. (22). Sequences wereanalyzed through comparison of the ITS sequence to those of multiplespecies of the Acinetobacter genus considered closest to the A. calcoaceti-cus-A. baumannii complex, including the A. baumannii strains AYE,
Received 3 May 2013 Accepted 12 May 2013
Published ahead of print 20 May 2013
Address correspondence to Wangxue Chen, [email protected].
Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/AAC.00944-13
August 2013 Volume 57 Number 8 Antimicrobial Agents and Chemotherapy p. 3601–3613 aac.asm.org 3601
ATCC 17978, and ATCC 19606 (strain BCRC 10591), the A. calcoaceticusstrain LMG 1046, the genomic species 3 strain LMG 1035, and thegenomic species 13TU strain BCRC 15417.
In vitro antimicrobial susceptibility testing. An antimicrobial sus-ceptibility profile of select clinical isolates and laboratory strains against apanel of 17 antimicrobials was determined using the broth microdilutionprotocols of the Clinical and Laboratory Standards Institute as describedpreviously (23).
Intranasal A. baumannii inoculation and sample collections. Forintranasal (i.n.) inoculation of mice, fresh inocula were prepared for eachexperiment from frozen stocks of A. baumannii isolates as previously de-scribed (7). Mice were anesthetized by intraperitoneal (i.p.) injection ofxylazine and ketamine and then inoculated intranasally with appropriatenumbers of various A. baumannii isolates in 50 �l of saline. Actual inoculain each experiment were determined by plating 10-fold serial dilutions onbrain heart infusion agar plates. The clinical appearance of the mice wasmonitored and scored as described previously (7). Groups of three to sixinfected mice were sacrificed 0, 4, and 24 h postinoculation (hpi). Therelevant organs (such as lungs, spleens, and lymph nodes) were asepticallyremoved and used for quantitative bacteriology or histopathology. Insome experiments, blood samples were collected for serum separation andlungs were lavaged, as described below, to collect bronchoalveolar lavage(BAL) fluid.
Quantitative bacteriology and histopathology. Lungs and spleenswere homogenized in sterile saline using aerosol-proof homogenizers.Lymph nodes were pressed through a 70-�m cell strainer (BD Falcon,Mississauga, Ontario) in sterile saline. Aliquots (100 �l) of 10-fold serialdilutions of the homogenates were cultured on brain heart infusion agarplates to quantify the number of viable A. baumannii organisms in therespective organs (7). In some experiments, blood samples were similarlycultured. For histopathology, lungs, spleens, livers, hearts, and kidneyswere fixed immediately in 10% neutral buffered formalin and processedby standard paraffin embedding methods (7). Sections were cut 4 �mthick, stained with hematoxylin-eosin (HE) (Department of LaboratoryMedicine, University of Ottawa, Ottawa, Canada), and examined by lightmicroscope.
BAL fluid. Lungs were lavaged five times with 1.0 ml saline supple-mented with 3 mM EDTA and 1% fetal bovine serum as previously de-
scribed (24). The total number of cells in BAL fluid was determined witha hemacytometer, and differential cell counts were determined by exam-ining 200 cells on cytospin slides (Cytospin 3; Shandon, Pittsburgh, PA)stained with Hema-3 (Fisher Scientific, Kalamazoo, MI). The lavage fluidwas centrifuged at 3,200 � g for 7 min, and the supernatant were collected,supplemented with protease inhibitors (Complete protease inhibitorcocktail tablets; Roche Applied Sciences, Laval, Quebec, Canada), andstored at �80°C.
Clinical blood chemistry. In some experiments, blood samples werecollected by incision of the posterior vena cava from mice sacrificed 24 hafter infection. The sera were separated and assayed for the levels of totalprotein, albumin, and globulin, the ratio of albumin to globulin, andlevels of �-glutamyl transpeptidase, aspartate aminotransferase (AST),alanine aminotransferase (ALT), bilirubin, creatinine, urea, and alkalinephosphatase (ALP) using the Roche Hitachi 917 analyzer (Vita-Tech,Markham, Canada) (25).
Determination of cytokine and chemokine levels. The levels of cyto-kines and chemokines in the sera and lung homogenate supernatant weremeasured using the 21-plex Milliplex MAP mouse cytokine/chemokinekits (Millipore, Ltd., Billerica, MA) on a Luminex 100IS system (Luminex,Austin, TX) as specified by the manufacturer. Samples were assayed induplicate, and cytokine/chemokine concentrations were calculatedagainst the standards using Beadview software (version 1.03; Upstate) (7).
In vivo antibiotic treatment efficacy studies. Groups of 10 femaleC57BL/6 mice were intranasally inoculated with LAC-4 and treated withimipenem (100 mg/kg of body weight/day, twice a day [b.i.d.], i.p.), ami-kacin (15 mg/kg/d, b.i.d., i.p.), tigecycline (10 mg/kg/d, b.i.d., subcutane-ously) or diluent (placebo; i.p.) starting 3 h after the LAC-4 inoculation.The blood and tissue (lungs and spleen) bacterial burdens at 24 hpi weredetermined, and clinical signs and body weight changes were observed for7 days.
Vaccination and protection studies. Groups of 5 female BALB/c micewere intranasally immunized with either 1 � 108 CFU of formalin-fixedLAC-4 (ffLAC-4) in 50 �l saline or saline only (sham-immunized mice).ffLAC-4 was prepared by incubating freshly grown LAC-4 in buffered 4%formaldehyde solution (Formalde-Fresh; Fisher Scientific, Ottawa, On-tario, Canada) for 24 h. The sterility of the preparation was confirmed bybacterial culture. The immunizations were performed under isoflurane
TABLE 1 A. baumannii isolates and strains used in the present study and initial bacterial burden screens following their i.n. inoculation in C57BL/6mice
Strain/isolate Strain/isolate reference SourceMultidrugresistancea
Bacterial burdenb
(log10 CFU/organ)
Lung Spleen
LAC-1 23 Clinical outbreak � 6.5 � 0.9*** 3.1 � 1.1LAC-4 23 Clinical outbreak � 8.1 � 0.8*** 6.6 � 0.6***LAC-5 23 Clinical outbreak � 5.6 � 0.4** 1.5 � 0.2LAC-6 23 Clinical outbreak � 6.0 � 0.3*** 3.0 � 1.0LAC-7 23 Clinical outbreak � 4.7 � 0.1 1.9 � 0.5LAC-11 23 Clinical outbreak � 5.0 � 0.1 1.3 � 0.0LAC-15 23 Clinical outbreak � 6.1 � 1.0*** 3.1 � 1.8LAC-16 23 Clinical outbreak � 5.2 � 0.2 1.8 � 0.4AYE 36 Clinical outbreak � 3.9 � 0.4 2.0 � 0.3SDF 36 Human body lice � 3.7 � 0.4 1.5 � 0.3NYB (isolate B) 37 Clinical isolate � 4.9 � 0.1 1.4 � 0.2ATCC 19606 23 Clinical isolate; type strain � 2.9 � 0.3 1.4 � 0.2ATCC 17978 23 Clinical isolate; reference strain � 4.0 � 0.3 1.8 � 0.2ATCC 17961 38 Clinical isolate; reference strain Not determined 5.2 � 0.2 1.3 � 0.015839 (ATCC item 202080) 23 Clinical isolate � 5.3 � 0.3 1.3 � 0.0a Multidrug resistance of the strains against a panel of 17 known antibiotics was determined using the broth microdilution protocols of the Clinical and Laboratory StandardsInstitute as described previously (23). �, multidrug resistant (resistant to three or more different classes of antibiotics); �, not multidrug resistant.b Female C57BL/6 mice (n � 3) were intranasally inoculated with �2 � 107 CFU of various strains of A. baumannii. The mice were sacrificed 24 h later, and the bacterial burdensin the lungs and spleen were determined by quantitative bacteriology. Data are presented as means � SD. The detection limit for bacterial burdens was 1.3 log10 CFU/organ. P 0.01 (**) and P 0.001 (***) versus results for ATCC 17961.
Harris et al.
3602 aac.asm.org Antimicrobial Agents and Chemotherapy

inhalation anesthesia at 0, 14, and 21 days. At 28 days, fecal pellets as wellas vaginal wash and blood (submandibular bleed) samples were collectedfrom individual mice for enzyme-linked immunosorbent assays (ELISAs)to determine A. baumannii-specific antibody (IgA and IgG) responses. Allof the mice were intraperitoneally challenged with 108 CFU (approxi-mately 100� the 100% lethal dose [LD100]) of freshly grown LAC-4 onday 47, and the survival of the mice was monitored for 7 days.
A. baumannii whole-cell-specific IgA and IgG ELISAs. Levels of A.baumannii whole-cell-specific antibodies in serum and mucosal sampleswere measured by ELISA (26). Briefly, 96-well microplates were precoatedwith 106 ffLAC-4 cells/well in 100 �l of sodium bicarbonate buffer (pH9.6). Samples were prediluted before assays (1:20 for vaginal wash sam-ples, 1:100 for serum IgG and IgA). Pooled samples collected from micethat had been intranasally immunized with ffLAC-4 or from naive micewere used as positive or negative controls, respectively.
Serum bactericidal assay. The serum bactericidal assay was per-formed based on the method of Luke et al. (14). The assays were set up intriplicate in 5-ml polystyrene tubes. Briefly, 100 �l mid-log-phase A. bau-mannii culture (5 � 107 CFU) in tryptic soy broth was mixed with 900 �lof either pooled normal human serum (NHS) (Valley Biomedical, Win-chester, VA) or heat-inactivated serum (HIS; 56°C for 30 min). The mix-tures were incubated at 37°C with rotation, and aliquots of 100 �l wereremoved from the culture at the indicated time for the determination ofviable bacterial counts. Results were expressed as percent survival, with100% being the number of viable bacteria grown in HIS.
Statistical analysis. Data are presented as means � standard devia-tions (SD) for each group, unless otherwise specified. Differences in quan-titative measurements were assessed by Student’s t test or one- or two-wayanalysis of variance (ANOVA), followed by Bonferroni’s post hoc multi-ple-comparison tests when appropriate. Differences were considered sig-nificant when P 0.05.
RESULTSIn vivo selection of the LAC-4 isolate in C57BL/6 mice. To de-velop a reproducible mouse model of A. baumannii pneumoniathat is more representative of natural human infection, we firstevaluated the relative virulence of a panel of 11 clinical isolates and4 ATCC type strains of A. baumannii in our collection, using themouse model of i.n. infection (7). We found that LAC-4 and, to alesser degree, LAC-1, LAC-5, LAC-6, and LAC-15, demonstratedsignificantly higher bacterial burdens than did ATCC 17961 inboth the lungs and the spleen at 24 hpi (P value of 0.01 to0.001) (Table 1).
Although LAC-4 was initially determined to belong to A. bau-mannii species using an API 20NE kit and the temperature test
(23), we reconfirmed its species designation in this study using amore quantitative ITS sequencing assay (22). The ITS regions ofLAC-4 are 607 bp long (Fig. 1), identical to known A. baumanniistrains (22). The LAC-4 ITS shares 99% (606/607) and 100% (607/607) sequence identity with the sequence of AYE (another clinicalstrain) and ATCC 19606 strains, respectively. LAC-4 was multi-drug resistant (i.e., resistant to three or more different classes ofantibiotics) according to antibiotic MICs and breakpoint designa-tions. Its multidrug resistance lies between AYE, which shows re-sistance to 12 of the 17 antibiotics tested, and ATCC 17978, whichis susceptible to all 17 antibiotics tested (Table 2).
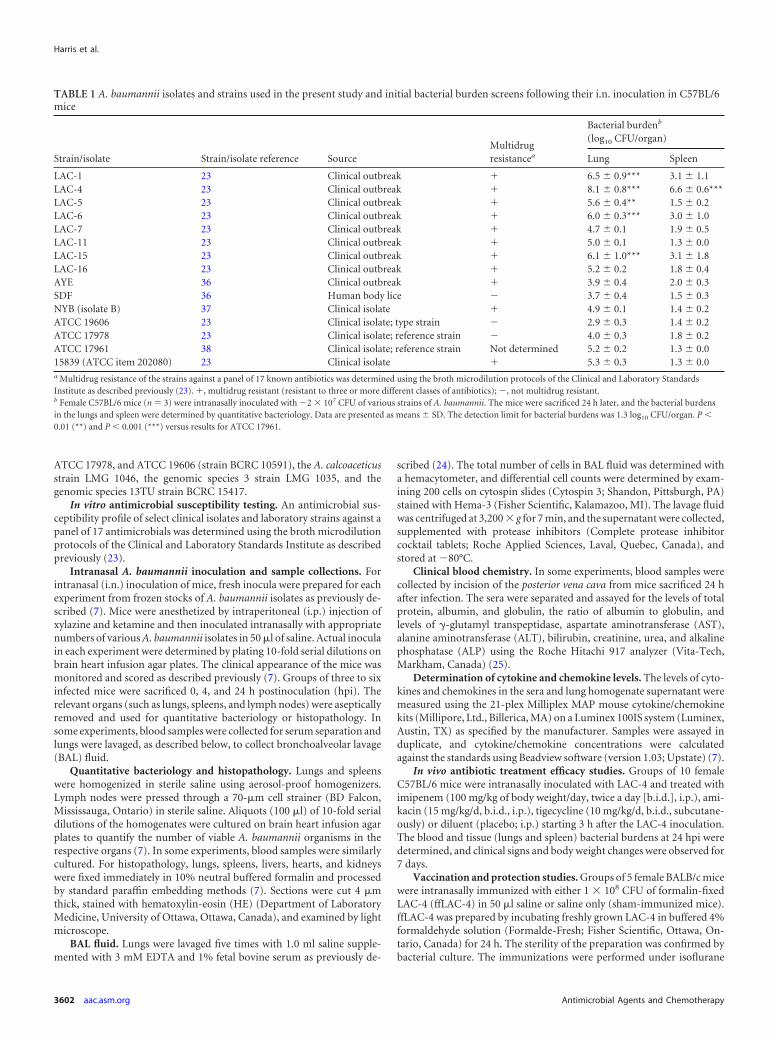
Susceptibility of C57BL/6 and BALB/c mice to intranasal in-oculation with LAC-4. To determine the relative susceptibilityof mice to i.n. infection with LAC-4, groups of male or femaleBALB/c and C57BL/6 mice were intranasally inoculated with105 to 108 CFU of freshly grown LAC-4. Clinical signs, bodyweight, and survival of the mice were monitored. FemaleBALB/c and C57BL/6 mice showed little or no clinical signsfollowing i.n. inoculation with 105 or 106 CFU of LAC-4,whereas their male counterparts showed transient but mildclinical signs at day 1 postinoculation (Fig. 2A). Mice inocu-lated with 107 CFU displayed various degrees of clinical signs,with the clinical scores of female mice being generally moresevere than those of male mice. All mice developed severe clin-ical signs when inoculated with 108 CFU of LAC-4 (Fig. 2A).Similarly, LAC-4-inoculated mice lost about 5 to 15% of theirinitial body weight at 24 h, depending on the size of the inoculareceived (Fig. 2B). The body weights of those mice that receivedthe lower inocula (105 or 106 CFU) generally recovered to theirpreinoculation level by day 3, and the mice continued to gainweight throughout the 6 to 7 days of the observation period(Fig. 2B). The body weights of the surviving mice from thegroup at 107 CFU were more variable, presumably reflectingthe various severities of the acute infection. Most significantly,i.n. inoculation with 108 CFU of LAC-4 resulted in 100% mor-tality by day 2 after inoculation, regardless of the mouse strainor sex (Fig. 2C). Up to 80% of the mice that received 107 CFU ofLAC-4 succumbed to the infection, whereas all mice that re-
FIG 1 PCR amplification of intergenic spacer (ITS) regions of four isolates:LAC-4 (lane 2), LAC-15 (lane 3), ATCC 17978 (lane 4), and AYE (lane 5). ADNA ladder is shown in lane 1. The ITS regions were amplified via PCR usingthe 1512F and 6R primers based on the method of Chang et al. (22). Due to theprimers used, the amplicons are approximately 800 bp in length. Removal of23S and 16S rRNA gene sequences from the sequencing reads will generate anITS of 607 to 638 bp in range, depending on the species (22).
TABLE 2 Antibiotic resistance profile of LAC-4, ATCC 17978, and AYEstrains
Antibiotic
LAC-4 ATCC 17978 AYE
MIC(�g/ml) Breakpointa
MIC(�g/ml) Breakpoint
MIC(�g/ml) Breakpoint
Amikacin 32 I �2 S 64 RGentamicin 16 R �2 S �256 RTobramycin 32 R �2 S 64 RImipenem �2 S �2 S �2 SMeropenem 4 S �2 S �2 SPiperacillin 256 R 16 S �256 RCefepime 16 I 4 S �256 RCefotaxime 256 R 8 S �256 RCeftazidime 128 R 8 S �256 RCeftriaxone �256 R 8 S �256 RCiprofloxacin 16 R 0.5 S 128 RGatifloxacin 2 S �2 S 8 RLevofloxacin �2 S �2 S 4 IDoxycycline 0.5 S �2 S 4 SMinocycline �0.25 S �0.25 S 0.5 STetracycline 8 I 4 S 128 RTigecycline �0.25 S �0.25 S 1 Sa R, resistant; I, intermediate; S, susceptible.
Mouse Model of A. baumannii-Associated Pneumonia
August 2013 Volume 57 Number 8 aac.asm.org 3603
ceived 105 or 106 CFU survived the infection (Fig. 2C). This isin contrast to previous studies with other A. baumannii strains,such as ATCC 17961 (7, 8), and other clinical isolates (9, 12),where no mice died after i.n. inoculation with 108 CFU. Theseresults suggest that the i.n. 50% lethal dose (LD50) for LAC-4infection in mice is about 107 CFU, and that, in both C57BL/6and BALB/c mice, LAC-4 is at least 100 times more lethal thanmost ATCC type strains and clinical isolates, including AYEand SFD (data not shown). Moreover, there were no significantdifferences in susceptibility due to mouse strain or sex. Conse-quently, we focused our subsequent characterization of thismodel primarily in female C57BL/6 mice, with the consider-ation that the majority of transgenic mice currently available tothe research community are on the C57BL/6 background.
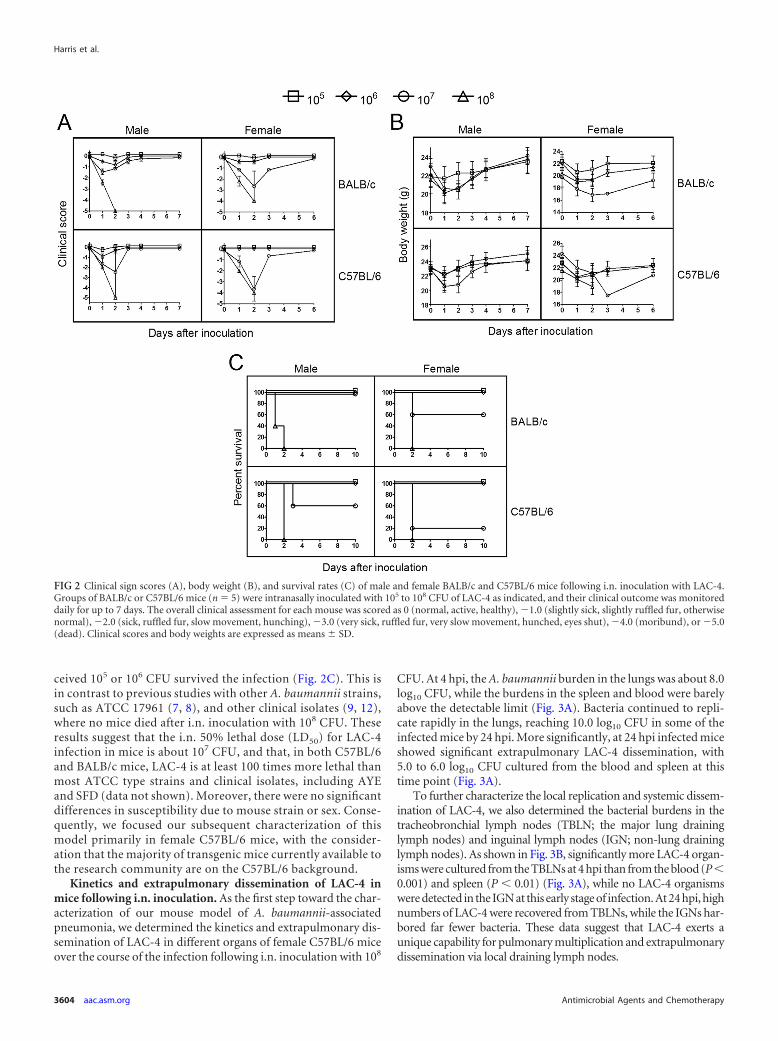
Kinetics and extrapulmonary dissemination of LAC-4 inmice following i.n. inoculation. As the first step toward the char-acterization of our mouse model of A. baumannii-associatedpneumonia, we determined the kinetics and extrapulmonary dis-semination of LAC-4 in different organs of female C57BL/6 miceover the course of the infection following i.n. inoculation with 108
CFU. At 4 hpi, the A. baumannii burden in the lungs was about 8.0log10 CFU, while the burdens in the spleen and blood were barelyabove the detectable limit (Fig. 3A). Bacteria continued to repli-cate rapidly in the lungs, reaching 10.0 log10 CFU in some of theinfected mice by 24 hpi. More significantly, at 24 hpi infected miceshowed significant extrapulmonary LAC-4 dissemination, with5.0 to 6.0 log10 CFU cultured from the blood and spleen at thistime point (Fig. 3A).
To further characterize the local replication and systemic dissem-ination of LAC-4, we also determined the bacterial burdens in thetracheobronchial lymph nodes (TBLN; the major lung draininglymph nodes) and inguinal lymph nodes (IGN; non-lung draininglymph nodes). As shown in Fig. 3B, significantly more LAC-4 organ-isms were cultured from the TBLNs at 4 hpi than from the blood (P0.001) and spleen (P 0.01) (Fig. 3A), while no LAC-4 organismswere detected in the IGN at this early stage of infection. At 24 hpi, highnumbers of LAC-4 were recovered from TBLNs, while the IGNs har-bored far fewer bacteria. These data suggest that LAC-4 exerts aunique capability for pulmonary multiplication and extrapulmonarydissemination via local draining lymph nodes.
FIG 2 Clinical sign scores (A), body weight (B), and survival rates (C) of male and female BALB/c and C57BL/6 mice following i.n. inoculation with LAC-4.Groups of BALB/c or C57BL/6 mice (n � 5) were intranasally inoculated with 105 to 108 CFU of LAC-4 as indicated, and their clinical outcome was monitoreddaily for up to 7 days. The overall clinical assessment for each mouse was scored as 0 (normal, active, healthy), �1.0 (slightly sick, slightly ruffled fur, otherwisenormal), �2.0 (sick, ruffled fur, slow movement, hunching), �3.0 (very sick, ruffled fur, very slow movement, hunched, eyes shut), �4.0 (moribund), or �5.0(dead). Clinical scores and body weights are expressed as means � SD.
Harris et al.
3604 aac.asm.org Antimicrobial Agents and Chemotherapy
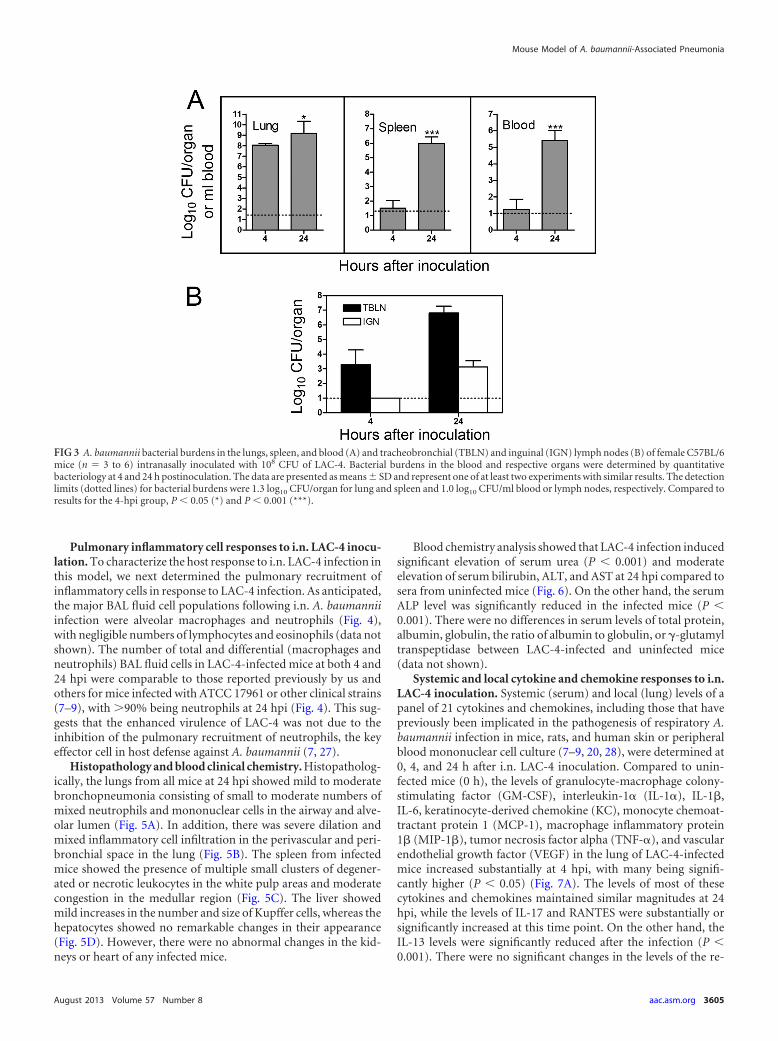
Pulmonary inflammatory cell responses to i.n. LAC-4 inocu-lation. To characterize the host response to i.n. LAC-4 infection inthis model, we next determined the pulmonary recruitment ofinflammatory cells in response to LAC-4 infection. As anticipated,the major BAL fluid cell populations following i.n. A. baumanniiinfection were alveolar macrophages and neutrophils (Fig. 4),with negligible numbers of lymphocytes and eosinophils (data notshown). The number of total and differential (macrophages andneutrophils) BAL fluid cells in LAC-4-infected mice at both 4 and24 hpi were comparable to those reported previously by us andothers for mice infected with ATCC 17961 or other clinical strains(7–9), with 90% being neutrophils at 24 hpi (Fig. 4). This sug-gests that the enhanced virulence of LAC-4 was not due to theinhibition of the pulmonary recruitment of neutrophils, the keyeffector cell in host defense against A. baumannii (7, 27).
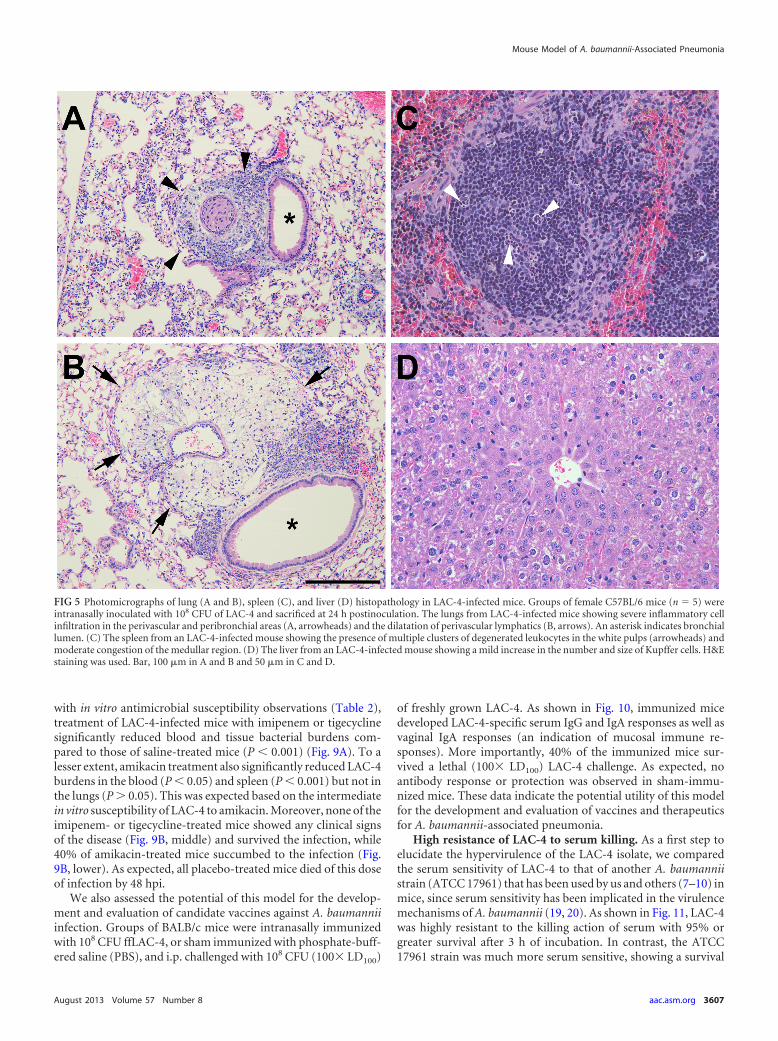
Histopathology and blood clinical chemistry. Histopatholog-ically, the lungs from all mice at 24 hpi showed mild to moderatebronchopneumonia consisting of small to moderate numbers ofmixed neutrophils and mononuclear cells in the airway and alve-olar lumen (Fig. 5A). In addition, there was severe dilation andmixed inflammatory cell infiltration in the perivascular and peri-bronchial space in the lung (Fig. 5B). The spleen from infectedmice showed the presence of multiple small clusters of degener-ated or necrotic leukocytes in the white pulp areas and moderatecongestion in the medullar region (Fig. 5C). The liver showedmild increases in the number and size of Kupffer cells, whereas thehepatocytes showed no remarkable changes in their appearance(Fig. 5D). However, there were no abnormal changes in the kid-neys or heart of any infected mice.
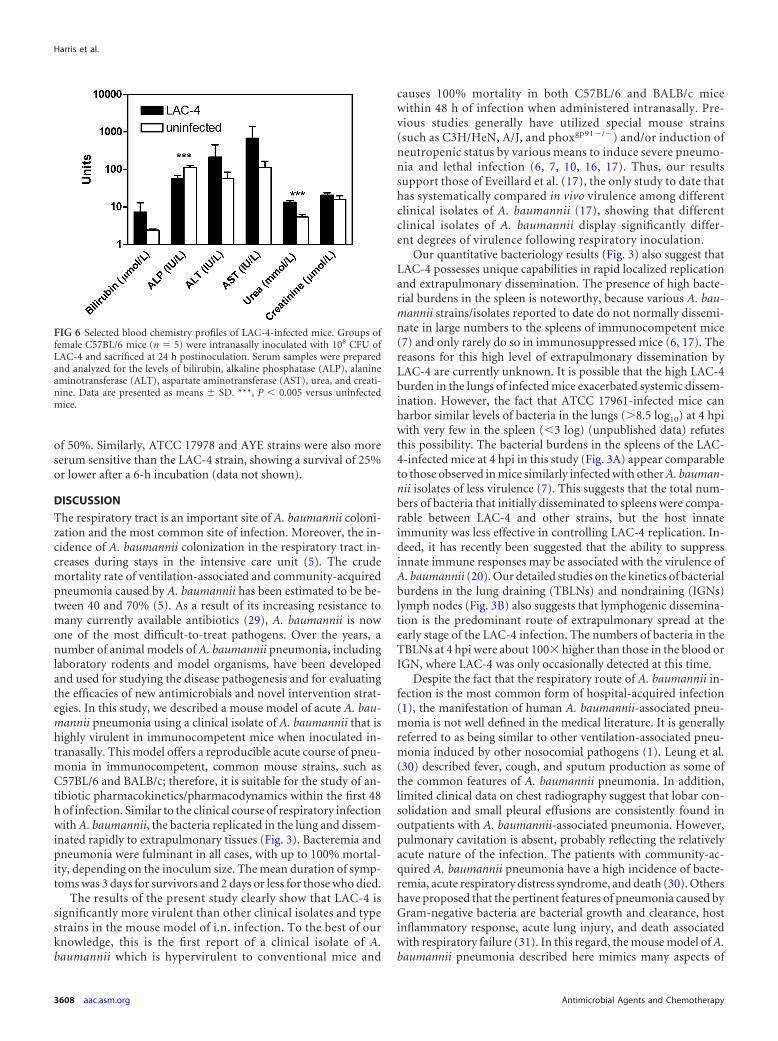
Blood chemistry analysis showed that LAC-4 infection inducedsignificant elevation of serum urea (P 0.001) and moderateelevation of serum bilirubin, ALT, and AST at 24 hpi compared tosera from uninfected mice (Fig. 6). On the other hand, the serumALP level was significantly reduced in the infected mice (P 0.001). There were no differences in serum levels of total protein,albumin, globulin, the ratio of albumin to globulin, or �-glutamyltranspeptidase between LAC-4-infected and uninfected mice(data not shown).
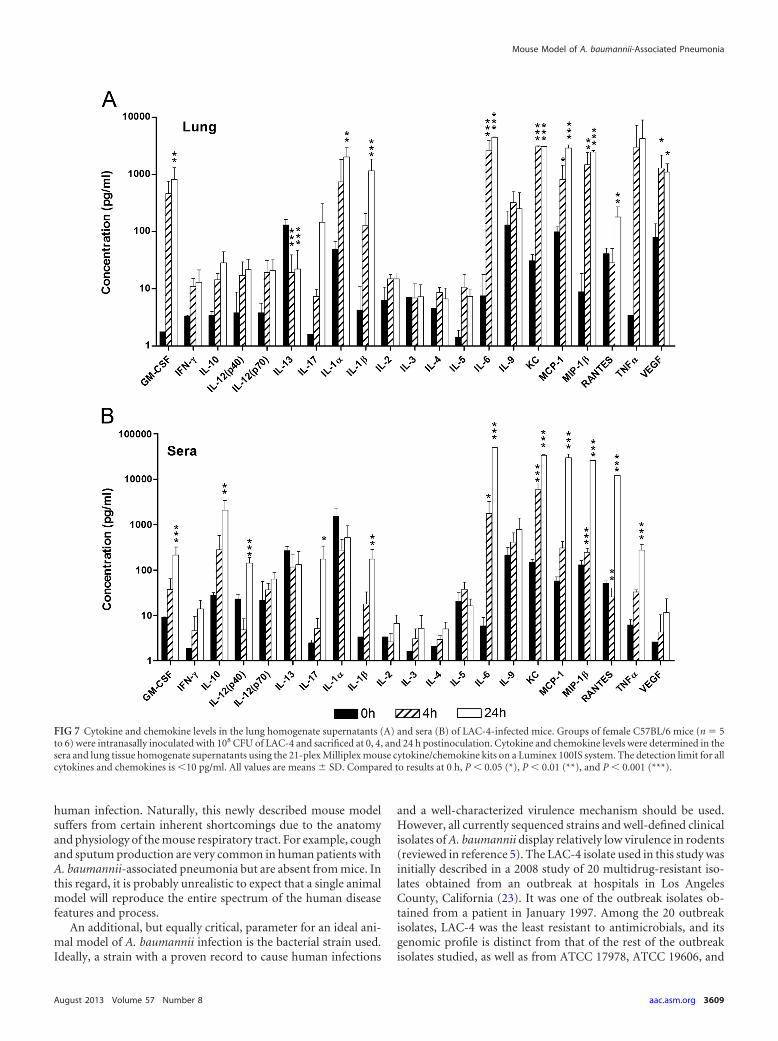
Systemic and local cytokine and chemokine responses to i.n.LAC-4 inoculation. Systemic (serum) and local (lung) levels of apanel of 21 cytokines and chemokines, including those that havepreviously been implicated in the pathogenesis of respiratory A.baumannii infection in mice, rats, and human skin or peripheralblood mononuclear cell culture (7–9, 20, 28), were determined at0, 4, and 24 h after i.n. LAC-4 inoculation. Compared to unin-fected mice (0 h), the levels of granulocyte-macrophage colony-stimulating factor (GM-CSF), interleukin-1� (IL-1�), IL-1�,IL-6, keratinocyte-derived chemokine (KC), monocyte chemoat-tractant protein 1 (MCP-1), macrophage inflammatory protein1� (MIP-1�), tumor necrosis factor alpha (TNF-�), and vascularendothelial growth factor (VEGF) in the lung of LAC-4-infectedmice increased substantially at 4 hpi, with many being signifi-cantly higher (P 0.05) (Fig. 7A). The levels of most of thesecytokines and chemokines maintained similar magnitudes at 24hpi, while the levels of IL-17 and RANTES were substantially orsignificantly increased at this time point. On the other hand, theIL-13 levels were significantly reduced after the infection (P 0.001). There were no significant changes in the levels of the re-
FIG 3 A. baumannii bacterial burdens in the lungs, spleen, and blood (A) and tracheobronchial (TBLN) and inguinal (IGN) lymph nodes (B) of female C57BL/6mice (n � 3 to 6) intranasally inoculated with 108 CFU of LAC-4. Bacterial burdens in the blood and respective organs were determined by quantitativebacteriology at 4 and 24 h postinoculation. The data are presented as means � SD and represent one of at least two experiments with similar results. The detectionlimits (dotted lines) for bacterial burdens were 1.3 log10 CFU/organ for lung and spleen and 1.0 log10 CFU/ml blood or lymph nodes, respectively. Compared toresults for the 4-hpi group, P 0.05 (*) and P 0.001 (***).
Mouse Model of A. baumannii-Associated Pneumonia
August 2013 Volume 57 Number 8 aac.asm.org 3605
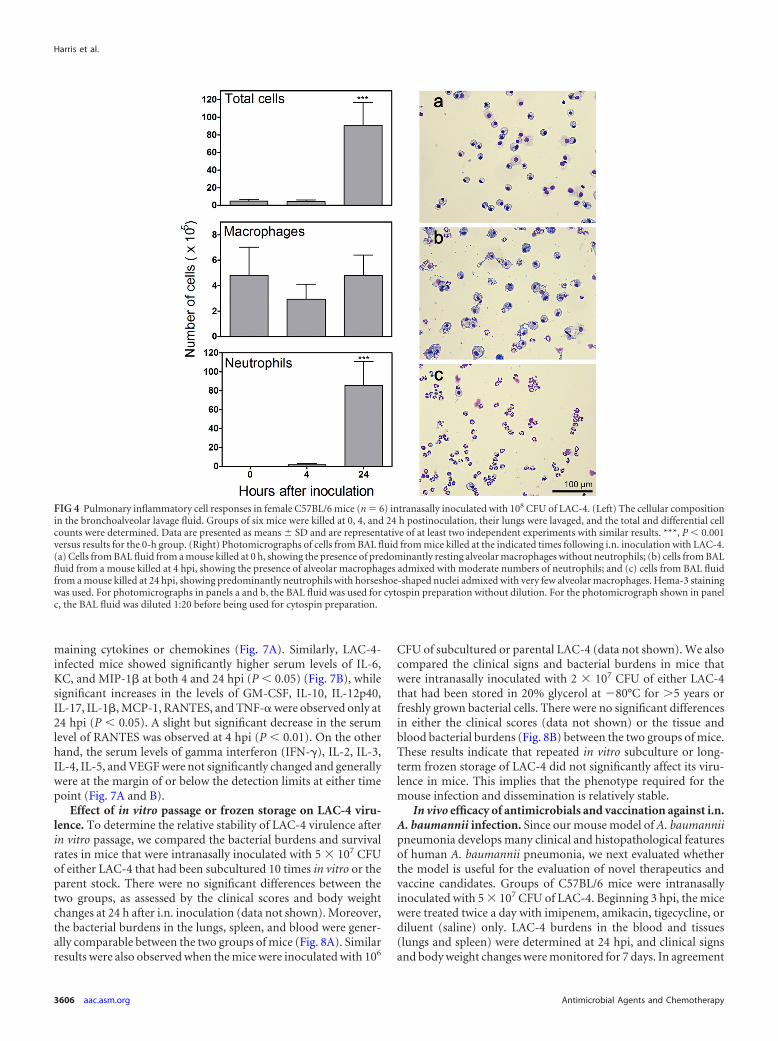
maining cytokines or chemokines (Fig. 7A). Similarly, LAC-4-infected mice showed significantly higher serum levels of IL-6,KC, and MIP-1� at both 4 and 24 hpi (P 0.05) (Fig. 7B), whilesignificant increases in the levels of GM-CSF, IL-10, IL-12p40,IL-17, IL-1�, MCP-1, RANTES, and TNF-� were observed only at24 hpi (P 0.05). A slight but significant decrease in the serumlevel of RANTES was observed at 4 hpi (P 0.01). On the otherhand, the serum levels of gamma interferon (IFN-�), IL-2, IL-3,IL-4, IL-5, and VEGF were not significantly changed and generallywere at the margin of or below the detection limits at either timepoint (Fig. 7A and B).
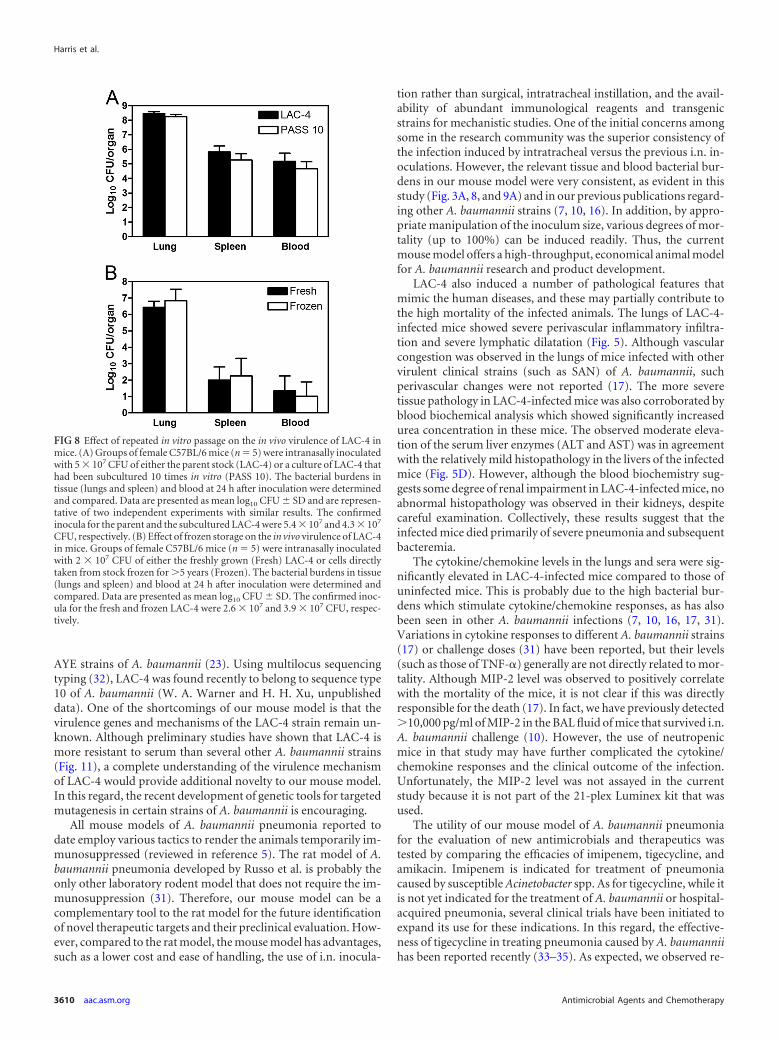
Effect of in vitro passage or frozen storage on LAC-4 viru-lence. To determine the relative stability of LAC-4 virulence afterin vitro passage, we compared the bacterial burdens and survivalrates in mice that were intranasally inoculated with 5 � 107 CFUof either LAC-4 that had been subcultured 10 times in vitro or theparent stock. There were no significant differences between thetwo groups, as assessed by the clinical scores and body weightchanges at 24 h after i.n. inoculation (data not shown). Moreover,the bacterial burdens in the lungs, spleen, and blood were gener-ally comparable between the two groups of mice (Fig. 8A). Similarresults were also observed when the mice were inoculated with 106
CFU of subcultured or parental LAC-4 (data not shown). We alsocompared the clinical signs and bacterial burdens in mice thatwere intranasally inoculated with 2 � 107 CFU of either LAC-4that had been stored in 20% glycerol at �80°C for 5 years orfreshly grown bacterial cells. There were no significant differencesin either the clinical scores (data not shown) or the tissue andblood bacterial burdens (Fig. 8B) between the two groups of mice.These results indicate that repeated in vitro subculture or long-term frozen storage of LAC-4 did not significantly affect its viru-lence in mice. This implies that the phenotype required for themouse infection and dissemination is relatively stable.
In vivo efficacy of antimicrobials and vaccination against i.n.A. baumannii infection. Since our mouse model of A. baumanniipneumonia develops many clinical and histopathological featuresof human A. baumannii pneumonia, we next evaluated whetherthe model is useful for the evaluation of novel therapeutics andvaccine candidates. Groups of C57BL/6 mice were intranasallyinoculated with 5 � 107 CFU of LAC-4. Beginning 3 hpi, the micewere treated twice a day with imipenem, amikacin, tigecycline, ordiluent (saline) only. LAC-4 burdens in the blood and tissues(lungs and spleen) were determined at 24 hpi, and clinical signsand body weight changes were monitored for 7 days. In agreement
FIG 4 Pulmonary inflammatory cell responses in female C57BL/6 mice (n � 6) intranasally inoculated with 108 CFU of LAC-4. (Left) The cellular compositionin the bronchoalveolar lavage fluid. Groups of six mice were killed at 0, 4, and 24 h postinoculation, their lungs were lavaged, and the total and differential cellcounts were determined. Data are presented as means � SD and are representative of at least two independent experiments with similar results. ***, P 0.001versus results for the 0-h group. (Right) Photomicrographs of cells from BAL fluid from mice killed at the indicated times following i.n. inoculation with LAC-4.(a) Cells from BAL fluid from a mouse killed at 0 h, showing the presence of predominantly resting alveolar macrophages without neutrophils; (b) cells from BALfluid from a mouse killed at 4 hpi, showing the presence of alveolar macrophages admixed with moderate numbers of neutrophils; and (c) cells from BAL fluidfrom a mouse killed at 24 hpi, showing predominantly neutrophils with horseshoe-shaped nuclei admixed with very few alveolar macrophages. Hema-3 stainingwas used. For photomicrographs in panels a and b, the BAL fluid was used for cytospin preparation without dilution. For the photomicrograph shown in panelc, the BAL fluid was diluted 1:20 before being used for cytospin preparation.
Harris et al.
3606 aac.asm.org Antimicrobial Agents and Chemotherapy
with in vitro antimicrobial susceptibility observations (Table 2),treatment of LAC-4-infected mice with imipenem or tigecyclinesignificantly reduced blood and tissue bacterial burdens com-pared to those of saline-treated mice (P 0.001) (Fig. 9A). To alesser extent, amikacin treatment also significantly reduced LAC-4burdens in the blood (P 0.05) and spleen (P 0.001) but not inthe lungs (P 0.05). This was expected based on the intermediatein vitro susceptibility of LAC-4 to amikacin. Moreover, none of theimipenem- or tigecycline-treated mice showed any clinical signsof the disease (Fig. 9B, middle) and survived the infection, while40% of amikacin-treated mice succumbed to the infection (Fig.9B, lower). As expected, all placebo-treated mice died of this doseof infection by 48 hpi.
We also assessed the potential of this model for the develop-ment and evaluation of candidate vaccines against A. baumanniiinfection. Groups of BALB/c mice were intranasally immunizedwith 108 CFU ffLAC-4, or sham immunized with phosphate-buff-ered saline (PBS), and i.p. challenged with 108 CFU (100� LD100)
of freshly grown LAC-4. As shown in Fig. 10, immunized micedeveloped LAC-4-specific serum IgG and IgA responses as well asvaginal IgA responses (an indication of mucosal immune re-sponses). More importantly, 40% of the immunized mice sur-vived a lethal (100� LD100) LAC-4 challenge. As expected, noantibody response or protection was observed in sham-immu-nized mice. These data indicate the potential utility of this modelfor the development and evaluation of vaccines and therapeuticsfor A. baumannii-associated pneumonia.
High resistance of LAC-4 to serum killing. As a first step toelucidate the hypervirulence of the LAC-4 isolate, we comparedthe serum sensitivity of LAC-4 to that of another A. baumanniistrain (ATCC 17961) that has been used by us and others (7–10) inmice, since serum sensitivity has been implicated in the virulencemechanisms of A. baumannii (19, 20). As shown in Fig. 11, LAC-4was highly resistant to the killing action of serum with 95% orgreater survival after 3 h of incubation. In contrast, the ATCC17961 strain was much more serum sensitive, showing a survival
FIG 5 Photomicrographs of lung (A and B), spleen (C), and liver (D) histopathology in LAC-4-infected mice. Groups of female C57BL/6 mice (n � 5) wereintranasally inoculated with 108 CFU of LAC-4 and sacrificed at 24 h postinoculation. The lungs from LAC-4-infected mice showing severe inflammatory cellinfiltration in the perivascular and peribronchial areas (A, arrowheads) and the dilatation of perivascular lymphatics (B, arrows). An asterisk indicates bronchiallumen. (C) The spleen from an LAC-4-infected mouse showing the presence of multiple clusters of degenerated leukocytes in the white pulps (arrowheads) andmoderate congestion of the medullar region. (D) The liver from an LAC-4-infected mouse showing a mild increase in the number and size of Kupffer cells. H&Estaining was used. Bar, 100 �m in A and B and 50 �m in C and D.
Mouse Model of A. baumannii-Associated Pneumonia
August 2013 Volume 57 Number 8 aac.asm.org 3607
of 50%. Similarly, ATCC 17978 and AYE strains were also moreserum sensitive than the LAC-4 strain, showing a survival of 25%or lower after a 6-h incubation (data not shown).
DISCUSSION
The respiratory tract is an important site of A. baumannii coloni-zation and the most common site of infection. Moreover, the in-cidence of A. baumannii colonization in the respiratory tract in-creases during stays in the intensive care unit (5). The crudemortality rate of ventilation-associated and community-acquiredpneumonia caused by A. baumannii has been estimated to be be-tween 40 and 70% (5). As a result of its increasing resistance tomany currently available antibiotics (29), A. baumannii is nowone of the most difficult-to-treat pathogens. Over the years, anumber of animal models of A. baumannii pneumonia, includinglaboratory rodents and model organisms, have been developedand used for studying the disease pathogenesis and for evaluatingthe efficacies of new antimicrobials and novel intervention strat-egies. In this study, we described a mouse model of acute A. bau-mannii pneumonia using a clinical isolate of A. baumannii that ishighly virulent in immunocompetent mice when inoculated in-tranasally. This model offers a reproducible acute course of pneu-monia in immunocompetent, common mouse strains, such asC57BL/6 and BALB/c; therefore, it is suitable for the study of an-tibiotic pharmacokinetics/pharmacodynamics within the first 48h of infection. Similar to the clinical course of respiratory infectionwith A. baumannii, the bacteria replicated in the lung and dissem-inated rapidly to extrapulmonary tissues (Fig. 3). Bacteremia andpneumonia were fulminant in all cases, with up to 100% mortal-ity, depending on the inoculum size. The mean duration of symp-toms was 3 days for survivors and 2 days or less for those who died.
The results of the present study clearly show that LAC-4 issignificantly more virulent than other clinical isolates and typestrains in the mouse model of i.n. infection. To the best of ourknowledge, this is the first report of a clinical isolate of A.baumannii which is hypervirulent to conventional mice and
causes 100% mortality in both C57BL/6 and BALB/c micewithin 48 h of infection when administered intranasally. Pre-vious studies generally have utilized special mouse strains(such as C3H/HeN, A/J, and phoxgp91�/�) and/or induction ofneutropenic status by various means to induce severe pneumo-nia and lethal infection (6, 7, 10, 16, 17). Thus, our resultssupport those of Eveillard et al. (17), the only study to date thathas systematically compared in vivo virulence among differentclinical isolates of A. baumannii (17), showing that differentclinical isolates of A. baumannii display significantly differ-ent degrees of virulence following respiratory inoculation.
Our quantitative bacteriology results (Fig. 3) also suggest thatLAC-4 possesses unique capabilities in rapid localized replicationand extrapulmonary dissemination. The presence of high bacte-rial burdens in the spleen is noteworthy, because various A. bau-mannii strains/isolates reported to date do not normally dissemi-nate in large numbers to the spleens of immunocompetent mice(7) and only rarely do so in immunosuppressed mice (6, 17). Thereasons for this high level of extrapulmonary dissemination byLAC-4 are currently unknown. It is possible that the high LAC-4burden in the lungs of infected mice exacerbated systemic dissem-ination. However, the fact that ATCC 17961-infected mice canharbor similar levels of bacteria in the lungs (8.5 log10) at 4 hpiwith very few in the spleen (3 log) (unpublished data) refutesthis possibility. The bacterial burdens in the spleens of the LAC-4-infected mice at 4 hpi in this study (Fig. 3A) appear comparableto those observed in mice similarly infected with other A. bauman-nii isolates of less virulence (7). This suggests that the total num-bers of bacteria that initially disseminated to spleens were compa-rable between LAC-4 and other strains, but the host innateimmunity was less effective in controlling LAC-4 replication. In-deed, it has recently been suggested that the ability to suppressinnate immune responses may be associated with the virulence ofA. baumannii (20). Our detailed studies on the kinetics of bacterialburdens in the lung draining (TBLNs) and nondraining (IGNs)lymph nodes (Fig. 3B) also suggests that lymphogenic dissemina-tion is the predominant route of extrapulmonary spread at theearly stage of the LAC-4 infection. The numbers of bacteria in theTBLNs at 4 hpi were about 100� higher than those in the blood orIGN, where LAC-4 was only occasionally detected at this time.
Despite the fact that the respiratory route of A. baumannii in-fection is the most common form of hospital-acquired infection(1), the manifestation of human A. baumannii-associated pneu-monia is not well defined in the medical literature. It is generallyreferred to as being similar to other ventilation-associated pneu-monia induced by other nosocomial pathogens (1). Leung et al.(30) described fever, cough, and sputum production as some ofthe common features of A. baumannii pneumonia. In addition,limited clinical data on chest radiography suggest that lobar con-solidation and small pleural effusions are consistently found inoutpatients with A. baumannii-associated pneumonia. However,pulmonary cavitation is absent, probably reflecting the relativelyacute nature of the infection. The patients with community-ac-quired A. baumannii pneumonia have a high incidence of bacte-remia, acute respiratory distress syndrome, and death (30). Othershave proposed that the pertinent features of pneumonia caused byGram-negative bacteria are bacterial growth and clearance, hostinflammatory response, acute lung injury, and death associatedwith respiratory failure (31). In this regard, the mouse model of A.baumannii pneumonia described here mimics many aspects of
FIG 6 Selected blood chemistry profiles of LAC-4-infected mice. Groups offemale C57BL/6 mice (n � 5) were intranasally inoculated with 108 CFU ofLAC-4 and sacrificed at 24 h postinoculation. Serum samples were preparedand analyzed for the levels of bilirubin, alkaline phosphatase (ALP), alanineaminotransferase (ALT), aspartate aminotransferase (AST), urea, and creati-nine. Data are presented as means � SD. ***, P 0.005 versus uninfectedmice.
Harris et al.
3608 aac.asm.org Antimicrobial Agents and Chemotherapy
human infection. Naturally, this newly described mouse modelsuffers from certain inherent shortcomings due to the anatomyand physiology of the mouse respiratory tract. For example, coughand sputum production are very common in human patients withA. baumannii-associated pneumonia but are absent from mice. Inthis regard, it is probably unrealistic to expect that a single animalmodel will reproduce the entire spectrum of the human diseasefeatures and process.
An additional, but equally critical, parameter for an ideal ani-mal model of A. baumannii infection is the bacterial strain used.Ideally, a strain with a proven record to cause human infections
and a well-characterized virulence mechanism should be used.However, all currently sequenced strains and well-defined clinicalisolates of A. baumannii display relatively low virulence in rodents(reviewed in reference 5). The LAC-4 isolate used in this study wasinitially described in a 2008 study of 20 multidrug-resistant iso-lates obtained from an outbreak at hospitals in Los AngelesCounty, California (23). It was one of the outbreak isolates ob-tained from a patient in January 1997. Among the 20 outbreakisolates, LAC-4 was the least resistant to antimicrobials, and itsgenomic profile is distinct from that of the rest of the outbreakisolates studied, as well as from ATCC 17978, ATCC 19606, and
FIG 7 Cytokine and chemokine levels in the lung homogenate supernatants (A) and sera (B) of LAC-4-infected mice. Groups of female C57BL/6 mice (n � 5to 6) were intranasally inoculated with 108 CFU of LAC-4 and sacrificed at 0, 4, and 24 h postinoculation. Cytokine and chemokine levels were determined in thesera and lung tissue homogenate supernatants using the 21-plex Milliplex mouse cytokine/chemokine kits on a Luminex 100IS system. The detection limit for allcytokines and chemokines is 10 pg/ml. All values are means � SD. Compared to results at 0 h, P 0.05 (*), P 0.01 (**), and P 0.001 (***).
Mouse Model of A. baumannii-Associated Pneumonia
August 2013 Volume 57 Number 8 aac.asm.org 3609
AYE strains of A. baumannii (23). Using multilocus sequencingtyping (32), LAC-4 was found recently to belong to sequence type10 of A. baumannii (W. A. Warner and H. H. Xu, unpublisheddata). One of the shortcomings of our mouse model is that thevirulence genes and mechanisms of the LAC-4 strain remain un-known. Although preliminary studies have shown that LAC-4 ismore resistant to serum than several other A. baumannii strains(Fig. 11), a complete understanding of the virulence mechanismof LAC-4 would provide additional novelty to our mouse model.In this regard, the recent development of genetic tools for targetedmutagenesis in certain strains of A. baumannii is encouraging.
All mouse models of A. baumannii pneumonia reported todate employ various tactics to render the animals temporarily im-munosuppressed (reviewed in reference 5). The rat model of A.baumannii pneumonia developed by Russo et al. is probably theonly other laboratory rodent model that does not require the im-munosuppression (31). Therefore, our mouse model can be acomplementary tool to the rat model for the future identificationof novel therapeutic targets and their preclinical evaluation. How-ever, compared to the rat model, the mouse model has advantages,such as a lower cost and ease of handling, the use of i.n. inocula-
tion rather than surgical, intratracheal instillation, and the avail-ability of abundant immunological reagents and transgenicstrains for mechanistic studies. One of the initial concerns amongsome in the research community was the superior consistency ofthe infection induced by intratracheal versus the previous i.n. in-oculations. However, the relevant tissue and blood bacterial bur-dens in our mouse model were very consistent, as evident in thisstudy (Fig. 3A, 8, and 9A) and in our previous publications regard-ing other A. baumannii strains (7, 10, 16). In addition, by appro-priate manipulation of the inoculum size, various degrees of mor-tality (up to 100%) can be induced readily. Thus, the currentmouse model offers a high-throughput, economical animal modelfor A. baumannii research and product development.
LAC-4 also induced a number of pathological features thatmimic the human diseases, and these may partially contribute tothe high mortality of the infected animals. The lungs of LAC-4-infected mice showed severe perivascular inflammatory infiltra-tion and severe lymphatic dilatation (Fig. 5). Although vascularcongestion was observed in the lungs of mice infected with othervirulent clinical strains (such as SAN) of A. baumannii, suchperivascular changes were not reported (17). The more severetissue pathology in LAC-4-infected mice was also corroborated byblood biochemical analysis which showed significantly increasedurea concentration in these mice. The observed moderate eleva-tion of the serum liver enzymes (ALT and AST) was in agreementwith the relatively mild histopathology in the livers of the infectedmice (Fig. 5D). However, although the blood biochemistry sug-gests some degree of renal impairment in LAC-4-infected mice, noabnormal histopathology was observed in their kidneys, despitecareful examination. Collectively, these results suggest that theinfected mice died primarily of severe pneumonia and subsequentbacteremia.
The cytokine/chemokine levels in the lungs and sera were sig-nificantly elevated in LAC-4-infected mice compared to those ofuninfected mice. This is probably due to the high bacterial bur-dens which stimulate cytokine/chemokine responses, as has alsobeen seen in other A. baumannii infections (7, 10, 16, 17, 31).Variations in cytokine responses to different A. baumannii strains(17) or challenge doses (31) have been reported, but their levels(such as those of TNF-�) generally are not directly related to mor-tality. Although MIP-2 level was observed to positively correlatewith the mortality of the mice, it is not clear if this was directlyresponsible for the death (17). In fact, we have previously detected10,000 pg/ml of MIP-2 in the BAL fluid of mice that survived i.n.A. baumannii challenge (10). However, the use of neutropenicmice in that study may have further complicated the cytokine/chemokine responses and the clinical outcome of the infection.Unfortunately, the MIP-2 level was not assayed in the currentstudy because it is not part of the 21-plex Luminex kit that wasused.
The utility of our mouse model of A. baumannii pneumoniafor the evaluation of new antimicrobials and therapeutics wastested by comparing the efficacies of imipenem, tigecycline, andamikacin. Imipenem is indicated for treatment of pneumoniacaused by susceptible Acinetobacter spp. As for tigecycline, while itis not yet indicated for the treatment of A. baumannii or hospital-acquired pneumonia, several clinical trials have been initiated toexpand its use for these indications. In this regard, the effective-ness of tigecycline in treating pneumonia caused by A. baumanniihas been reported recently (33–35). As expected, we observed re-
FIG 8 Effect of repeated in vitro passage on the in vivo virulence of LAC-4 inmice. (A) Groups of female C57BL/6 mice (n � 5) were intranasally inoculatedwith 5 � 107 CFU of either the parent stock (LAC-4) or a culture of LAC-4 thathad been subcultured 10 times in vitro (PASS 10). The bacterial burdens intissue (lungs and spleen) and blood at 24 h after inoculation were determinedand compared. Data are presented as mean log10 CFU � SD and are represen-tative of two independent experiments with similar results. The confirmedinocula for the parent and the subcultured LAC-4 were 5.4 � 107 and 4.3 � 107
CFU, respectively. (B) Effect of frozen storage on the in vivo virulence of LAC-4in mice. Groups of female C57BL/6 mice (n � 5) were intranasally inoculatedwith 2 � 107 CFU of either the freshly grown (Fresh) LAC-4 or cells directlytaken from stock frozen for 5 years (Frozen). The bacterial burdens in tissue(lungs and spleen) and blood at 24 h after inoculation were determined andcompared. Data are presented as mean log10 CFU � SD. The confirmed inoc-ula for the fresh and frozen LAC-4 were 2.6 � 107 and 3.9 � 107 CFU, respec-tively.
Harris et al.
3610 aac.asm.org Antimicrobial Agents and Chemotherapy
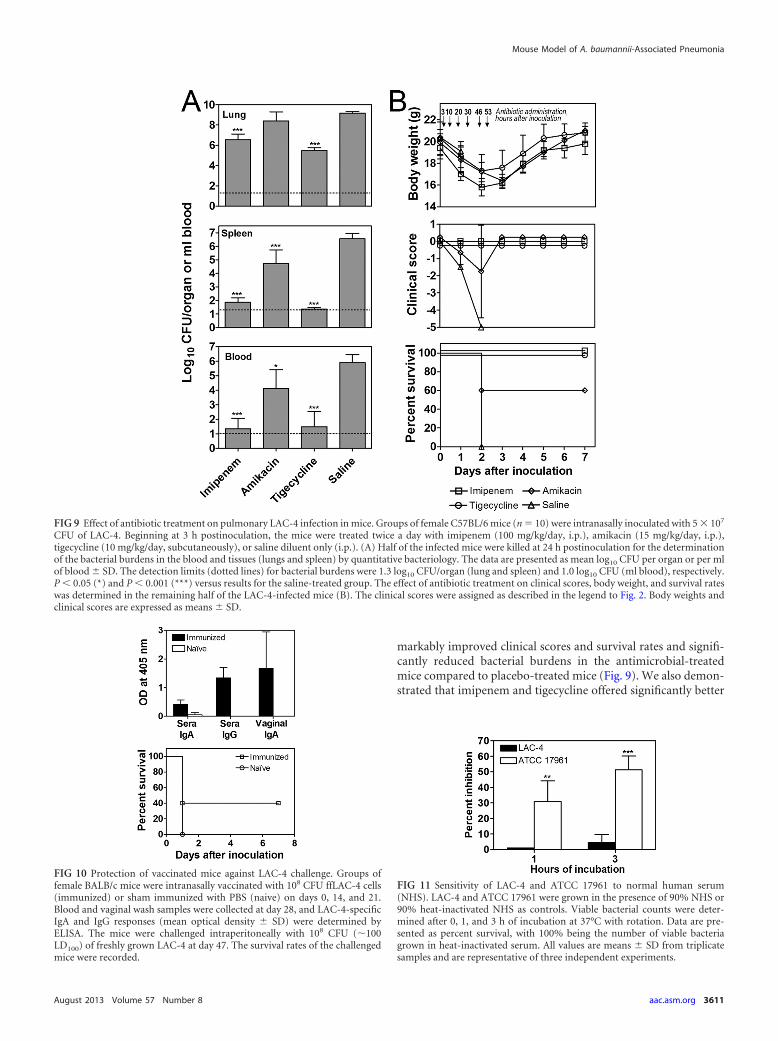
markably improved clinical scores and survival rates and signifi-cantly reduced bacterial burdens in the antimicrobial-treatedmice compared to placebo-treated mice (Fig. 9). We also demon-strated that imipenem and tigecycline offered significantly better
FIG 9 Effect of antibiotic treatment on pulmonary LAC-4 infection in mice. Groups of female C57BL/6 mice (n � 10) were intranasally inoculated with 5 � 107
CFU of LAC-4. Beginning at 3 h postinoculation, the mice were treated twice a day with imipenem (100 mg/kg/day, i.p.), amikacin (15 mg/kg/day, i.p.),tigecycline (10 mg/kg/day, subcutaneously), or saline diluent only (i.p.). (A) Half of the infected mice were killed at 24 h postinoculation for the determinationof the bacterial burdens in the blood and tissues (lungs and spleen) by quantitative bacteriology. The data are presented as mean log10 CFU per organ or per mlof blood � SD. The detection limits (dotted lines) for bacterial burdens were 1.3 log10 CFU/organ (lung and spleen) and 1.0 log10 CFU (ml blood), respectively.P 0.05 (*) and P 0.001 (***) versus results for the saline-treated group. The effect of antibiotic treatment on clinical scores, body weight, and survival rateswas determined in the remaining half of the LAC-4-infected mice (B). The clinical scores were assigned as described in the legend to Fig. 2. Body weights andclinical scores are expressed as means � SD.
FIG 10 Protection of vaccinated mice against LAC-4 challenge. Groups offemale BALB/c mice were intranasally vaccinated with 108 CFU ffLAC-4 cells(immunized) or sham immunized with PBS (naive) on days 0, 14, and 21.Blood and vaginal wash samples were collected at day 28, and LAC-4-specificIgA and IgG responses (mean optical density � SD) were determined byELISA. The mice were challenged intraperitoneally with 108 CFU (�100LD100) of freshly grown LAC-4 at day 47. The survival rates of the challengedmice were recorded.
FIG 11 Sensitivity of LAC-4 and ATCC 17961 to normal human serum(NHS). LAC-4 and ATCC 17961 were grown in the presence of 90% NHS or90% heat-inactivated NHS as controls. Viable bacterial counts were deter-mined after 0, 1, and 3 h of incubation at 37°C with rotation. Data are pre-sented as percent survival, with 100% being the number of viable bacteriagrown in heat-inactivated serum. All values are means � SD from triplicatesamples and are representative of three independent experiments.
Mouse Model of A. baumannii-Associated Pneumonia
August 2013 Volume 57 Number 8 aac.asm.org 3611

protection than amikacin, an observation that is consistent with invitro MIC values (Table 2). Our data also show that imipenem andtigecycline were significantly more efficient in controlling pulmo-nary bacterial multiplication than amikacin. Similarly, our pro-phylactic vaccination and challenge experiment showed that vac-cination of mice with inactivated LAC-4 cells protects againstlethal challenge with this pathogen.
In conclusion, we have described a mouse model of A. bau-mannii pneumonia which offers a reproducible, acute course ofpneumonia that, in many aspects (high tissue bacterial burdens,severe pulmonary pathology, and high mortality), mimics humandisease. One of the major limitations of our study is that the rela-tionship of LAC-4 to the severity of clinical infection in patientscould not be established due to the unavailability of clinical infor-mation from the hospital where LAC-4 was initially isolated. Be-cause of the difficulties in treating health care-acquired infectionscaused by A. baumannii, this model should be a useful tool forfuture identification and characterization of important virulencefactors, pathogenesis studies, and the evaluation of novel thera-peutics and vaccines for this emerging infectious agent.
ACKNOWLEDGMENTS
This work was partially funded by the intramural program (A-base) of theNational Research Council Canada (to W.C. and G.B.P.) and by theUnited States Department of Homeland Security (2009-ST-062-000018),the U.S. Army Research Laboratory, and the U.S. Army Research Office(W911NF-12-1-0059) (to H.H.X.).
We are grateful to Pfizer for providing tigecycline. We thank DidierRaoult (The University of the Mediterranean Aix-Marseille II, France),Carl Urban (New York Hospital Queens), and Sheena Chu (Los AngelesCounty Public Health Laboratory) for providing clinical isolates of A.baumannii used in this study. We thank Tom Devecseri for assistance inthe preparation of photomicrography, as well as Sonya Valentine, MelissaChong, Maria Gonzalez, and Wayne Warner for technical assistance.
REFERENCES1. Hartzell JD, Kim AS, Kortepeter MG, Moran KA. 2007. Acinetobacter
pneumonia: a review. Med. Gen. Med. 9:4.2. Kuo LC, Lai CC, Liao CH, Hsu CK, Chang YL, Chang CY, Hsueh PR.
2007. Multidrug-resistant Acinetobacter baumannii bacteraemia: clinicalfeatures, antimicrobial therapy and outcome. Clin. Microbiol. Infect. 13:196 –198.
3. Gaynes R, Edwards JR. 2005. Overview of nosocomial infections causedby gram-negative bacilli. Clin. Infect. Dis. 41:848 – 854.
4. Fournier PE, Richet H. 2006. The epidemiology and control of Acineto-bacter baumannii in health care facilities. Clin. Infect. Dis. 42:692– 699.
5. McConnell MJ, Actis L, Pachon J. 2012. Acinetobacter baumannii: hu-man infections, factors contributing to pathogenesis and animal models.FEMS Microbiol. Rev. [Epub ahead of print.] doi:10.1111/j.1574-6976.2012.00344.x.
6. Joly-Guillou ML, Wolff M, Pocidalo JJ, Walker F, Carbon C. 1997. Useof a new mouse model of Acinetobacter baumannii pneumonia to evalu-ate the postantibiotic effect of imipenem. Antimicrob. Agents Chemother.41:345–351.
7. van Faassen H, KuoLee R, Harris G, Zhao X, Conlan JW, Chen W.2007. Neutrophils play an important role in host resistance to respiratoryinfection with Acinetobacter baumannii in mice. Infect. Immun. 75:5597–5608.
8. Renckens R, Roelofs JJ, Knapp S, de Vos AF, Florquin S, van der PollT. 2006. The acute-phase response and serum amyloid A inhibit the in-flammatory response to Acinetobacter baumannii pneumonia. J. Infect.Dis. 193:187–195.
9. Knapp S, Wieland CW, Florquin S, Pantophlet R, Dijkshoorn L,Tshimbalanga N, Akira S, van der Poll T. 2006. Differential roles ofCD14 and toll-like receptors 4 and 2 in murine Acinetobacter pneumonia.Am. J. Respir. Crit. Care Med. 173:122–129.
10. Qiu H, Kuo Lee R, Harris G, Chen W. 2009. High susceptibility torespiratory Acinetobacter baumannii infection in A/J mice is associatedwith a delay in early pulmonary recruitment of neutrophils. MicrobesInfect. 11:946 –955.
11. Russo TA, MacDonald U, Beanan JM, Olson R, MacDonald IJ,Sauberan SL, Luke NR, Schultz LW, Umland TC. 2009. Penicillin-binding protein 7/8 contributes to the survival of Acinetobacter bauman-nii in vitro and in vivo. J. Infect. Dis. 199:513–521.
12. Jacobs AC, Hood I, Boyd KL, Olson PD, Morrison JM, Carson S,Sayood K, Iwen PC, Skaar EP, Dunman PM. 2010. Inactivation ofphospholipase D diminishes Acinetobacter baumannii pathogenesis. In-fect. Immun. 78:1952–1962.
13. Russo TA, Luke NR, Beanan JM, Olson R, Sauberan SL, MacDonald U,Schultz LW, Umland TC, Campagnari AA. 2010. The K1 capsular poly-saccharide of Acinetobacter baumannii strain 307-0294 is a major viru-lence factor. Infect. Immun. 78:3993– 4000.
14. Luke NR, Sauberan SL, Russo TA, Beanan JM, Olson R, Loehfelm TW,Cox AD, St Michael F, Vinogradov EV, Campagnari AA. 2010. Identi-fication and characterization of a glycosyltransferase involved in Acineto-bacter baumannii lipopolysaccharide core biosynthesis. Infect. Immun.78:2017–2023.
15. March C, Regueiro V, Llobet E, Moranta D, Morey P, Garmendia J,Bengoechea JA. 2010. Dissection of host cell signal transduction duringAcinetobacter baumannii-triggered inflammatory response. PLoS One5:e10033. doi:10.1371/journal.pone.0010033.
16. Qiu H, Kuolee R, Harris G, Chen W. 2009. Role of NADPH phagocyteoxidase in host defense against acute respiratory Acinetobacter baumanniiinfection in mice. Infect. Immun. 77:1015–1021.
17. Eveillard M, Soltner C, Kempf M, Saint-André Lemarié J-PC, Randri-anarivelo C, Seifert H, Wolff M, Joly-Guillou M-L. 2010. The virulencevariability of different Acinetobacter baumannii strains in experimentalpneumonia. J. Infect. 60:154 –161.
18. Kim SW, Choi CH, Moon DC, Jin JS, Lee JH, Shin JH, Kim JM, Lee YC,Seol SY, Cho DT, Lee JC. 2009. Serum resistance of Acinetobacter bau-mannii through the binding of factor H to outer membrane proteins.FEMS Microbiol. Lett. 301:224 –231.
19. King LB, Swiatlo E, Swiatlo A, McDaniel LS. 2009. Serum resistance andbiofilm formation in clinical isolates of Acinetobacter baumannii. FEMSImmunol. Med. Microbiol. 55:414 – 421.
20. de Breij A, Dijkshoorn L, Lagendijk E, van der Meer J, Koster A,Bloemberg G, Wolterbeek R, van den Broek P, Nibbering P. 2010. Dobiofilm formation and interactions with human cells explain the clinicalsuccess of Acinetobacter baumannii? PLoS One 5:e10732. doi:10.1371/journal.pone.0010732.
21. Bentancor LV, Routray A, Bozkurt-Guzel C, Camacho-Peiro A, PierGB, Maira-Litran T. 2012. Evaluation of the trimeric autotransporter Ataas a vaccine candidate against Acinetobacter baumannii infections. Infect.Immun. 80:3381–3388.
22. Chang HC, Wei YF, Dijkshoorn L, Vaneechoutte M, Tang CT, ChangTC. 2005. Species-level identification of isolates of the Acinetobacter cal-coaceticus-Acinetobacter baumannii complex by sequence analysis of the16S-23S rRNA gene spacer region. J. Clin. Microbiol. 43:1632–1639.
23. Valentine SC, Contreras D, Tan S, Real LJ, Chu S, Xu HH. 2008.Phenotypic and molecular characterization of Acinetobacter baumanniiclinical isolates from nosocomial outbreaks in Los Angeles County, Cali-fornia. J. Clin. Microbiol. 46:2499 –2507.
24. Chen W, Havell EA, Moldawer LL, McIntyre KW, Chizzonite RA,Harmsen AG. 1992. Interleukin 1: an important mediator of host resis-tance against Pneumocystis carinii. J. Exp. Med. 176:713–718.
25. Chen W, Kuo Lee R, Shen H, Conlan JW. 2004. Susceptibility ofimmunodeficient mice to aerosol and systemic infection with virulentstrains of Francisella tularensis. Microb. Pathog. 36:311–318.
26. Yan H, Kuo Lee R, Tram K, Qiu H, Zhang J, Patel GB, Chen W. 2009.3=,5=-Cyclic diguanylic acid elicits mucosal immunity against bacterialinfection. Biochem. Biophys. Res. Commun. 387:581–584.
27. Breslow JM, Meissler JJ, Jr, Hartzell RR, Spence PB, Truant A, GaughanJ, Eisenstein TK. 2011. Innate immune responses to systemic Acinetobac-ter baumannii infection in mice: neutrophils, but not interleukin-17, me-diate host resistance. Infect. Immun. 79:3317–3327.
28. de Breij A, Haisma EM, Rietveld M, El Ghalbzouri A, van den Broek PJ,Dijkshoorn L, Nibbering PH. 2012. Three-dimensional human skinequivalent as a tool to study Acinetobacter baumannii colonization. An-timicrob. Agents Chemother. 56:2459 –2464.
Harris et al.
3612 aac.asm.org Antimicrobial Agents and Chemotherapy

29. Boucher HW, Talbot GH, Bradley JS, Edwards JE, Gilbert D, Rice LB,Scheld M, Spellberg B, Bartlett J. 2009. Bad bugs, no drugs: no ESKAPE!An update from the Infectious Diseases Society of America Clin. Infect.Dis. 48:1–12.
30. Leung WS, Chu CM, Tsang KY, Lo FH, Lo KF, Ho PL. 2006. Fulminantcommunity-acquired Acinetobacter baumannii pneumonia as a distinctclinical syndrome. Chest 129:102–109.
31. Russo TA, Beanan JM, Olson R, MacDonald U, Luke NR, Gill SR,Campagnari AA. 2008. Rat pneumonia and soft-tissue infection modelsfor the study of Acinetobacter baumannii biology. Infect. Immun. 76:3577–3586.
32. Diancourt L, Passet V, Nemec A, Dijkshoorn L, Brisse S. 2010. Thepopulation structure of Acinetobacter baumannii: expanding multiresis-tant clones from an ancestral susceptible genetic pool. PLoS One 5:e10034.doi:10.1371/journal.pone.0010034.
33. Shin JA, Chang YS, Kim HJ, Kim SK, Chang J, Ahn CM, Byun MK.2012. Clinical outcomes of tigecycline in the treatment of multidrug-resistant Acinetobacter baumannii infection. Yonsei Med. J. 53:974 –984.
34. Moon SY, Peck KR, Chang HH, Kim SW, Heo ST, Son JS, Ryu SY,Moon C, Jung SI, Shin SY, Lee JA, Joung MK, Chung DR, Kang CI,
Song JH. 2012. Clinical experience of tigecycline treatment in infectionscaused by extensively drug-resistant Acinetobacter spp. Microb. Drug Re-sist. 18:562–566.
35. Tasbakan MS, Pullukcu H, Sipahi OR, Tasbakan MI, Aydemir S,Bacakoglu F. 2011. Is tigecyclin a good choice in the treatment of multi-drug-resistant Acinetobacter baumannii pneumonia? J. Chemother. 23:345–349.
36. Fournier PE, Vallenet D, Barbe V, Audic S, Ogata H, Poirel L, RichetH, Robert C, Mangenot S, Abergel C, Nordmann P, Weissenbach J,Raoult D, Claverie JM. 2006. Comparative genomics of multidrug resis-tance in Acinetobacter baumannii. PLoS Genet. 2:e7. doi:10.1371/journal.pgen.0020007.
37. Yoon J, Urban C, Terzian C, Mariano N, Rahal JJ. 2004. In vitro doubleand triple synergistic activities of polymyxin B, imipenem, and rifampinagainst multidrug-resistant Acinetobacter baumannii. Antimicrob.Agents Chemother. 48:753–757.
38. Baumann P, Doudoroff M, Stanier RY. 1968. A study of the Moraxellagroup. II. Oxidative-negative species (genus Acinetobacter). J. Bacteriol.95:1520 –1541.
Mouse Model of A. baumannii-Associated Pneumonia
August 2013 Volume 57 Number 8 aac.asm.org 3613
Top Related
