
UvA-DARE (Digital Academic Repository) Identification and … · CHAPTERR 4 Interferenceewithth ......
15
UvA-DARE is a service provided by the library of the University of Amsterdam (http://dare.uva.nl) UvA-DARE (Digital Academic Repository) Identification and characterization of human Polycomb-group proteins. Satijn, D.P.E. Link to publication Citation for published version (APA): Satijn, D. P. E. (2000). Identification and characterization of human Polycomb-group proteins. Amsterdam General rights It is not permitted to download or to forward/distribute the text or part of it without the consent of the author(s) and/or copyright holder(s), other than for strictly personal, individual use, unless the work is under an open content license (like Creative Commons). Disclaimer/Complaints regulations If you believe that digital publication of certain material infringes any of your rights or (privacy) interests, please let the Library know, stating your reasons. In case of a legitimate complaint, the Library will make the material inaccessible and/or remove it from the website. Please Ask the Library: http://uba.uva.nl/en/contact, or a letter to: Library of the University of Amsterdam, Secretariat, Singel 425, 1012 WP Amsterdam, The Netherlands. You will be contacted as soon as possible. Download date: 25 Jun 2018
Transcript of UvA-DARE (Digital Academic Repository) Identification and … · CHAPTERR 4 Interferenceewithth ......

UvA-DARE is a service provided by the library of the University of Amsterdam (http://dare.uva.nl)
UvA-DARE (Digital Academic Repository)
Identification and characterization of human Polycomb-group proteins.
Satijn, D.P.E.
Link to publication
Citation for published version (APA):Satijn, D. P. E. (2000). Identification and characterization of human Polycomb-group proteins. Amsterdam
General rightsIt is not permitted to download or to forward/distribute the text or part of it without the consent of the author(s) and/or copyright holder(s),other than for strictly personal, individual use, unless the work is under an open content license (like Creative Commons).
Disclaimer/Complaints regulationsIf you believe that digital publication of certain material infringes any of your rights or (privacy) interests, please let the Library know, statingyour reasons. In case of a legitimate complaint, the Library will make the material inaccessible and/or remove it from the website. Please Askthe Library: http://uba.uva.nl/en/contact, or a letter to: Library of the University of Amsterdam, Secretariat, Singel 425, 1012 WP Amsterdam,The Netherlands. You will be contacted as soon as possible.
Download date: 25 Jun 2018

CHAPTERR 4
Interferencee with the expression of a novel human Polycombb protein, HPC2, results in cellular
transformationn and apoptosis. Satijn,, D.P.E., D. J. Olson, J. Van der Vlag, K.M. Hamer, C. Lambrechts, H. Masselink, M.J.
Gunster,, R.G.A.B. Sewalt, R. Van Driel and A.P.Otte.
Mol.. Cell. Biol., 1997,17 (10), 6076-6086.
49 9

50 0

4.. Human Poly comb protein and cellular transformation
Interferencee with the Expression of a Novel Human Polycomb Protein,, hPc2, Results in Cellular Transformation
andd Apoptosis DAVI DD P. E. SATIJN,1 DANIEL J. OLSON,2 JOHAN VAN DER VLAG,1 KARIEN M. HAMER,1
CAROO LAMBRECHTS,3 HANS MASSELINK,3 MARCO J. GUNSTER,1 RICHARD G. A. B. SEW ALT,1
ROELL VAN DRIEL,1 AND ARIE P. OTTE1* E.E. C. Slater Instituut, University of Amsterdam, 1018 TV Amsterdam,1 and Divisions of Experimental Therapy and
MolecularMolecular Carcinogenesis of the Netherlands Cancer Institute, 1066 CX Amsterdam,3 The Netherlands, and DepartmentDepartment of Oral Molecular Biology, School of Dentistry, Cancer Center, Hormonal and Reproductive Cancer
Division,Division, Oregon Health Sciences University, Portland, Oregon 972012
Receivedd 19 March 1997/Relurned for modification 3 May 1997/Accepted 1 July 1997
Polycombb (Pc) is involved in the stable and heritable repression of homeotic gene activity during Drosophila development.. Here, we report the identification of a novel human Pc homolog, hPc2. This gene is more closely relatedd to a Xenopus Pc homolog, XPc, than to a previously described human Pc homolog, CBX2 (hPcl). However,, the hPc2 and CBX2/hPcl proteins colocalize in interphase nuclei of human U-2 OS osteosarcoma cells,, suggesting that the proteins are part of a common protein complex. To study the functions of the novel humann Pc homolog, we generated a mutant protein, AhPc2, which lacks an evolutionarily conserved C-tenninal domain.. This C-terminal domain is important for hPc2 function, since the AhPc2 mutant protein which lacks thee C-terminal domain is unable to repress gene activity. Expression of the AhPc2 protein, but not of the wild-typee hPc2 protein, results in cellular transformation of mammalian cell lines as judged by phenotypic changes,, altered marker gene expression, and anchorage-independent growth. Specifically in AhPc2-trans-formedd cells, the expression of the c-myc proto-oncogene is strongly enhanced and serum deprivation results inn apoptosis. In contrast, overexpression of the wild-type hPc2 protein results in decreased c-myc expression. Ourr data suggest that hPc2 is a repressor of proto-oncogene activity and that interference with hPc2 function cann lead to derepression of proto-oncogene transcription and subsequently to cellular transformation.
Thee Drosophila Polycomb (Pc) gene is a member of the PolycombPolycomb group (PcG) gene family. These genes are part of a cellularr memory system that is responsible for the inheritance off gene activity by progeny cells (7, 19, 21, 28, 38). Stable and heritablee transmission of gene activity is crucial for the main-tenancee of the differentiated identity of cells over many cell generations.. It has been proposed that PcG proteins repress homeoticc gene expression via the formation of multimeric complexes.. This model is based on the observation that differ-entt PcG proteins, including Pc, bind in overlapping patterns on polytenee chromosomes in Drosophila salivary gland cells (32, 48).. In Drosophila, the PcG protein Polyhomeotic (Ph), but no otherr known protein, coimmunoprecipitates or cofractionates withh Pc (12). Recently, we found that two human counterparts off Ph, HPH1 and HPH2, coimmunoprecipitate, cofractionate, andd colocalize in nuclear domains with both the vertebrate PcGG protein BMI1 and a human Pc homolog, indicating that thesee proteins associate in vivo (16). A similar in vivo interac-tionn between Bmil and a Ph homolog has been found in mice (1).. These findings indicate the existence of a vertebrate PcG multimericc protein complex.
Thee Pc protein binds to about 100 loci on polytene chromo-somess in Drosophila salivary gland cells (32, 48). These loci includee the homeotic gene loci and other PcG gene loci (48), indicatingg that homeotic genes and PcG genes are target genes off Pc. Direct evidence for this idea stems from the observation
** Corresponding author. Mailing address: E. C. Slater Instituut, Universityy of Amsterdam, Plantage Muidergracht 12, 1018 TV Am-sterdam,, The Netherlands. Phone: 31-20-5255115. Fax: 31-20-5255124. E-mail:: [email protected].
thatt Pc is associated with chromatin of the silent part of the homeoticc bithorax (Ubx) complex (27). Physical association of thee Pc protein with chromatin of other target loci has not been reportedd so far.
Severall vertebrate homologs of Pc have been identified (15, 30,33),, suggesting that repression of gene activity, mediated by Pc,, is evolutionarily conserved. This idea is supported by the findingg that a mouse Pc homolog, M33, is able to rescue the DrosophilaDrosophila Pc phenotype (25). Recently, it has become evident thatt M33 displays functions in mice similar to those of Pc in Drosophila.Drosophila. M33-deficient mice show homeotic transforma-tionss of the axial skeleton, along with sternal and limb malfor-mationss (6).
Manyy aspects of the molecular mechanism underlying the rolee of PcG proteins in the stable transmission of gene activity aree enigmatic. An important clue about the molecular mech-anismm underlying Pc action is the observation that the Pc pro-teinn shares a homologous domain with the Drosophila hetero-chromatin-bindingg protein HP1 (29, 39). The shared motif betweenn Pc and HP1 has been termed the chromodomain (chromatinn organization modifier) (29). This discovery pro-videss an important, direct link between the regulation of gene activityy and chromatin structure. It suggests that Pc and HP1 operatee through common mechanisms, which may involve the formationn of heterochromatin-like structures. The chromodo-mainn is essential for binding of the Pc protein to chromatin. Whenn the chromodomain is either mutated or deleted, it no longerr binds to chromatin (24). Also, a conserved domain locatedd in the C terminus of the Pc protein (30) is crucial for Pcc function. In Drosophila, several naturally occurring Pc mu-tantss either are mutated in the C-terminal domain or lack this
51 1

4.. Human Polycomb protein and cellular transformation
domainn entirely (13). A mutant Pc gene lacking the C-terminal domainn is unable to repress gene activity (4, 24).
Inn the present study, we identified a novel human Pc ho-molog,, hPc2. We showed that the hPc2 protein and a previ-ouslyy characterized human Pc protein, CBX2 (15), colocalize inn nuclear domains of human U-2 OS osteosarcoma cells. To studyy the functions of hPc2, we designed an hPc2 mutant proteinn that lacks the C-terminal domain, which is crucial for thee ability of hPc2 to repress gene activity. Expression of this hPc22 mutant in mammalian cell lines results in (i) cellular transformation,, (ii) enhanced expression of the c-myc proto-oncogene,, and (iii ) apoptosis upon serum deprivation. Our dataa suggest that hPc2 is a repressor of proto-oncogene activity andd that interference with hPc2 function can lead to derepres-sionn of proto-oncogene transcription and subsequently to cel-lularr transformation.
MATERIAL SS AND METHOD S
Isolationn of the hPc2 gene. For screening a \gi1U human fetal brain cDNA libraryy (Clontech, Palo Alto, Calif.), a probe encompassing the entire coding regionn oiXPc (33) except for the chromodomain (ranging from amino acids [aa] 600 to 558) was generated by PCR. The filters were hybridized overnight at 50°C inn 0.5x SSC ( l x SSC is 0.15 M NaCl plus 0.015 M sodium citrate)-10x Den-hardt'ss solution-10% dextran sulfate-0.1% sodium dodecyl sulfate (SDS)-100 figg of denatured herring sperm DNA per ml-[a-32P]ATP-!abelled probe (5 x 105
cpm/ml).. After being washed twice for 60 min at 55°C in 0.5 x SSC-0.1% SDS, thee filters were exposed to X-ray films with intensifying screens for 2 days at -70°C.. Positive plaques were isolated and purified. The complete nucleotide sequencee of the longest cDNA was determined by sequence analysis.
Analysiss of hPc2 transcripts. Multitissue Northern blots containing approxi-matelyy 2 p.% of poly(A) * RNA per lane from different human tissues or human celll lines were obtained commercially (Clontech). The U-2 OS cell line was not presentt on the commercial Northern blot. Poly( A) * RNA of U-2 OS was isolated andd blotted, and the expression pattern of HPc2 was analyzed. To allow a com-parisonn with the commercial Northern blot, we blotted poly(A)+ RNA of SW480 cells,, which are represented on the commercial blot and in which all three genes aree strongly expressed. The blots were hybridized with [a-32P]dATP-labelled DNAA probes and autoradiographed with X-ray films and intensifying screens at -70°C.. Northern blot analyses were performed with an hPc2 probe that excludes thee conserved chromodomain and C-terminal domain (coding for aa 60 to 530).
Productionn of the M33 and hPc2 polyclonal antibodies. A cDNA fragment encodingg aa 157 to 432 of M33 was cloned into the expression vector pET-23b (Novagen,, Madison, Wis.). This fragment does not encompass the conserved chromodomainn or the conserved C-terminal domain. A cDNA fragment encod-ingg aa 60 to 558 of hPc2 was cloned in pET-23c (Novagen). This fragment does nott encompass the conserved chromodomain. Fusion proteins were produced in EscherichiaEscherichia colt BL21 (DE), and the purified fusion proteins were used to immunizee rabbits and chicken. Sera were affinity purified over an antigen-cou-pledd cyanogen bromide-Sepharose column (Pharmacia, Uppsala, Sweden). Fu-sionn proteins or extracts of U-2 OS cells were separated by SDS-polyacrylamide gell electrophoresis (PAGE) and transferred to nitrocellulose. The blots were probedd with a 1:10,000 dilution of T7 antibody (Novagen) or a 1:1,000 to 1:5,000 dilutionn of affinity-purified anti-hPc2 or anti-M33 antibodies. The secondary alkalinee phosphatase-goat anti-rabbit or anti-chicken IgG (H+L) antibodies (Jacksonn Immunoresearch Laboratories) were diluted 1:10,000, and nitrobtue tetrazolium-5-bromo-4-chloro-3-indolylphosphatee toluidinium (NBT/BCIP) (Boehringer)) was used as the substrate for detection.
Immunofluorescencee staining of tissue culture cells. U-2 OS ceils were cul-turedd and labeled as described recently (16). Labeling has been analyzed by confocall laser scanning microscopy, and single optical sections are shown (see Fig.. 5). The first two pictures represent the two different detection channels of thee dual image, and the third picture represents the false-color overlay. For labelling,, donkey anti-rabbit immunoglobulin G coupled to Cy3 (Jackson Immu-noresearchh Laboratories) and donkey anti-chicken IgG coupled to fluorescein isothiocyanatee (FITC) (Jackson) were used.
LexAA fusion reporter gene targeted repression assay. The LexA fusion re-porterr gene targeted repression assay was performed as described previously (4, 35).. The chloramphenicol acetyltransferase (CAT) reporter gene has been re-placedd by the luciferase gene (LUC). U-2 OS cells were cultured in a 25-cnr' flask andd cotransfected with the heat shock factor (HSF)-inducible LUC reporter plasmid,, 4 u,g of the LexA fusion constructs, and 2 \Lg of the pSV/B-Gal construct (Promega)) by the calcium phosphate precipitation method. The HSF-inducible LUCC reporter plasmid was activated by exposing the cells at 43°C for 1 h, followedd by a 6-h recovery at 37°C, as described previously (4,35). LUC activity wass normalized to B-galactosidase activity. The absolute values of LUC activity variedd between independent experiments. The LUC activity in cells transfected withh the LUC reporter plasmid only was therefore set at 100%, and LUC
activitiess in celts which were cotransfected with indicated plasmids were ex-pressedd as a percentage of this control value. The degree of repression is ex-pressedd as the mean the standard error of the mean (SEM).
Establishmentt and characterization of stable C57MG and U-2 OS cell lines. Indicatedd cDNAs were cloned into the pcDNA3 vector (Invitrogen, San Diego, Calif.),, which carries the neomycin resistance gene and in which the indicated cDNAss are under control of the enhancer from the immediate-early gene of humann cytomegalovirus (CMV). C57MG, U-2 OS, or Ratla cells were trans-fectedd with Lipofectin (Gibco) as specified by the manufacturer, and stably transfectedd lines were selected by culturing the cells for 2 weeks in medium containingg 800 u,g of Geneticin (G418) (Gibco) per ml. The surviving cells were clonallyy expanded for 2 to 4 weeks in medium containing 250 u-g of G418 per ml. Individuall C57MG and U-2 OS cell clones were selected and cultured in indi-viduall dishes. After five passages, the cell lines were characterized for [3H]thy-midinee incorporation and gene expression. [3H]thymidine incorporation was measuredd as described previously (26). The numbers were normalized to the proteinn content. From C57MG cells, expressing low levels of c-myc, poly(A)+
RNAA was isolated. From U-2 OS cells, expressing high levels of c-myc, total RNA wass isolated. Isolation of RNA and Northern analysis were performed by stan-dardd procedures. The blots were hybridized with |_a-32P]dATP-labelled DNA probes,, and the blots were autoradiographed with intensifying screens at -70°C withh preflashed X-ray films.
Westernn blot analysis of hPc2 and c-myc proteins. The expression levels of the hPc22 and c-myc proteins were analyzed by using cell lysates of the stably trans-fectedd C57MG and U-2 OS clones (35). For hPc2 detection, the blots were incubatedd with a 1:5,000 dilution of affinity-purified rabbit anti-hPc2 antibodies. Thee monoclonal 9E10 antibody was used for the detection of c-myc (8). Equal amountss of proteins were loaded, as measured by the bicinchoninic acid method (41)) and visualized by Coomassie blue staining of a gel.
Softt agar growth assays. Cell lines were analyzed for anchorage-independent growthh as described previously (34, 40, 44). Ratla cells were transfected by the calciumm phosphate transfection procedure with full-length hPc2 (hPc2WT), the C-terminall deletion mutant (AhPc2), and a mutant lacking the chromodomain (Achromo),, which were all cloned in the pcDNA3 vector. As a positive control, thee c-myc cDNA which was cloned in the pRc-CMV vector was transfected. The cellss were subjected to selection with 500 u.g of G418 per ml. The cells were then culturedd for 14 days, at which time the G418 resistant clones were counted. The clonesclones were trypsinized, and cells were counted. A total of 5 X 104 cells in 5 ml off 10% Dulbecco's modified Eagle's medium containing 0.4% (wt/vol) agarose weree seeded in 5-cm petri dishes containing 1% (wt/vol) agarose. The plates were inspectedd 14 to 21 days after seeding of the cells, and the colonies were counted. Thee entire procedure, including the transfection of the cDNAs, was performed inn triplicate.
Apoptosiss assays. Exponentially growing cells were washed with phosphate-bufferedd saline, and culture medium containing 10% serum was exchanged for culturee medium containing 0.1% serum.. After 3 or 6 h, the cells were washed withh cold phosphate-buffered saline and incubated with FITC-conjugated an-nexinn V (Nexins Research, Hoeven, The Netherlands) for 15 min at 4°C. The bindingg conditions were as recommended by the manufacturer. After being washed,, the cells were fixed in 2% paraformaldehyde for 15 min. The cells were stilll viable, since they excluded propidium iodine (36). The assay was repeated fivee times with similar results. Beside annexin V staining, C57MG/A/V cells showedd hallmarks of apoptosis, such as membrane blebbing, cell shrinkage, and chromatinn condensation after 24 to 48 h. As an independent assay to detect apoptosis,, we monitored apoptosis-induced DNA strand breaks by the TUNEL (TdT-mediatedd dUTP nick end labeling) assay (Boehringer), in which the ter-minall deoxynucleotidyltransferase (TdT) enzyme activity is measured. Also using thiss assay, we observed apoptosis, specifically in serum-deprived 4hPc2-trans-fectedd cells (36).
Nucleotidee sequence accession number. The complete hPc2 sequence has been depositedd with GenBank under accession no. AF013956.
RESULTS S
Isolationn and characterization of a novel human Pc ho-molog,, hPc2. To isolate human Pc homologs, we screened a humann fetal brain cDNA library with a probe that encom-passess the coding region of a Xenopus Pc homolog, XPc, except forr the chromodomain (33). We isolated an 1,867-bp cDNA clonee (Fig. 1). This clone contains a 1,674-bp open reading framee (Fig. 2A). The predicted 558-aa protein possesses a conservedd region, the chromodomain, which is 96% identical too the XPc chromodomain, 77% identical to the chromodo-mainn of the mouse Pc homolog, M33, and 55% identical to the DrosophilaDrosophila Pc chromodomain (Fig. 2A) (29, 30, 33). A con-servedd C-terminal domain is 100% identical to XPc at the proteinn level, 73% identical to M33, and 67% identical to Pc (Fig.. 2). Overall, the human protein is 48% identical to XPc.
52 2

g g g c g a g c g c g a c g c g g g a g c c g g g g c g g c g c g c a g c c g g g g c , ,
c c g g c c g g g c t g t g c g g g g c g a g c g g c g g c g c g g c g g g g g c c t t c g g g c g g g g c g g g c g c c g g c g g g a g c c a g c a g c g t c t g c a g c c g c g c c c g g c c a g c c c c c t c c g g c t o o
,GCTGCCAGCTGTTGGCGAGCACGTCTTCGCGGTGGAGAGCATCGAGAAGAAGCGGATCCGCAAGGGCAGAGTGGAGTATCTGGTGAAATGGAGAGGCTGGTCGCCCAAATAT^ ^
OO I B V r n V l J I I * * * !! 8 KK * K Q PP K T
ACGTGGGAACCGGAGGAGAACATCCTGGAGCGCAGGCTGCTGATCGCCTTCGAGAACAGGGAACGGCAGGAGCAGCTGATGGGATATCGGAAGAGAGGGCCGAAGCCCAAACCGCTAGTG G
T H « P « t » I t » » « l ' I ' T * * 0 N R E R Q E Q L M G YY E — K — K — S — E — K E — R P L v
«ITGCAGGTGCrTAGCTTTGCrCGTCGTTCCAATGTCCTGACCGGCCTCCAGGACTCCTCCACTGACAACCGTGCCAAGCTGGATTTGGGCGCGCAGGGGAAGGGCCAGGGGCATCAGTAr r
' v " a ' v P T F A R R S N V I . T G I . Q D S S T D N R A l t L D L G A Q G K G O G H Q Y Y
,..^CTCAACAGCAAGAAGCACCACCAGTACCAGCCGCACAGCAAGGAGGGGAAGCCCCGGCCGCCGGGCAAGAGCGGCAAGTACTACTACCAGCTCAACAGCAAGAAGCACCACCCCTAC C
" E LL „ S K K H H Q Y ( l P H S K I S G K P R P P G K S G K Y Y Y Q I . N S l t K H H P Y
rAGCGCGACCrrAAAATGTACGACCTGGAGTACCAGGGCGGCCACAAGGAGGCGGGCAGCCGCACCTGCCCGGACCTGGGGGCCAAGAGCCACCCGCCCGACAAGTGGGGGCAAGGTGCG G
0 P D P K H Y D L 0 Y Q G G H K E A P S P T C P D I . G » 1 ! S H P P D K U A Q G A A
-GCGCGAAAGGGTArCTGGGGGrCGTGAAGCCCTTGGCCGGTGCGGCGGGTGCTCCAGGCAAAGGCTCCGAGAAGGGCCCCCCCAACGGAATGATGCCGGCCCCCAAAGAGGCTGTGACG G
G A K G Y L G A V K P L A G A A G A P G K G S E K G P P M G M H P A P K E A V T T
-GCAA(-.^GA^TG, ;G-UCAAGATGAAGATAGTCAAGAACAAGAACAAGAACGGACGCATCGTGATCGTGATGAGCAAATACATGGAGAACGGCATGCAGGCGGTGAAGATCAAGTrCGGC C
oo H es I 1 1 I I I I T 1 l ' I I I n i l » i n i « ' " " 1 » i I > > «
GAGGTGGCAGAGGGGGAGGCTCGCTCCCCCAGCCACAAGAAGCGGGCAGCCGACGAGCGCCACCCTCCTGCCGACAGGACTTTTAAAAAGGCGGCGGGCGCAGAGGAGAAGAAGGTGGAG G
B VV A E G E A R S P S H K E R A A D E R H P P A D R T P K K A A G A E E K K V E
C C A P P K R R E E E V S G V S D P Q P 0 D A G S I 1 K L S P T E E A T G E Q P L ( ! L L
A^ rACCAAGCCCGACCTCCTTGCCTGGGRCcCGGCCCGGAACACGCACCCGCCCTCACACCACCCGCACCCGCACCCCCATCACCACCACCACCACCACCACCACCACCACCACGCCGTC C
T T K P D L L A W D P A R K T H P P S K H P M P H P H H H H H H H H H M K K A V V
.GrcTGAATCTCTCCCACGTGCGCAAGCGCTGCCTCTCCGAGACCCACGGCGAGCGCGAGCCCTGCAAGAAGCGGCTGACTGCGCGCAGCATCAGCACCCCCACCTGCCTGGGGGGCAGC C
" 0 L » l S H V » l [ » C t « I I H O I » I » C « K » L T » « « I » I F T C L O I > a a
.. .A,;,.:..;rT,;AGCG,-rcGGCCGACCTGGCACCAGCCGCCGCCCTCCGGCAGCCCGAGGTCATCCTGCTAGACTCAGACCTGGATGAACCCATAGACTTGCGCTCGGTCAAGAGCCGCAGC
P A A E R P A D L P P A A A L R O P E V I I . L D S D L D E P I D L R S V K S R S S
;AII ;<:T.-JG( IGAGC' ( ;< C ' E A G E P P S S L O V K P E T P A S A A V A V A A A A A P T T T A E E P P A E A A
VVTGF-AGAGTCGCTGAGCGAGTTCAAGCCCTTCTTTOGGAATATAATTATCACCGACGTCACCGCGAACTGCCTCACCGTTACTTTCAAGGAGTACGTGACGGTGTAG<:C-G o " D , E P A E S I . S E E E P P P G l I I I I I I ) » I * » C I , I T i r K « Y V T V . .
1440 0 480 0
55f l l
1692 2
FIGG 1 Nucleotide sequence of hPc2 and its predicted amino acid sequence. The chromodomain in the N terminus of the protein and the conserved C-terminal domainn of the protein are shaded. A putative nuclear localization signal is underlined. The stop codon of the hPc2 gene is indicated by an asterisk.
Whenn conservative changes are taken into account, the human proteinn is 80% similar to the XPc protein. Also, at the nucle-otidee level the human cDNA has a striking 52% identity to the XPcXPc cDNA in the entire coding region. We conclude, there-fore,, that we have isolated the human homolog of the XPc protein. .
Inn contrast, the overall homology between the human Pc proteinn and the mouse Pc homolog M33 is a mere 24% iden-tity.. At the nucleotide level, the novel human Pc homolog has 29%% identity to the M33 cDNA. This degree of homology is considerablyy lower than that between the human Pc homolog andd the XPc cDNA. However, when conservative changes are takenn into account, the novel human Pc homolog is 68% sim-ilarr to M33 at the protein level. This probably implies that the overalll three-dimensional shapes of the two proteins are very similar.. This may in turn indicate that the two proteins are functionallyy equivalent.
AA partially characterized human homolog of M33, CBX2 (15),, has 86% identity and 100% similarity to M33 at the proteinn level (Fig. 2B). Significant homology between CBX2 andd our novel human Pc protein is limited to the conserved C-terminall domain (Fig. 2). The CBX2 protein has a mere 25%% overall identity but a significantly higher 80% similarity to thee novel human Pc homolog (Fig. 2B).
Wee conclude that there are at least two human Pc homologs. Theyy encode proteins that are homologous only in the con-servedd C-terminal domain and presumably also in the chromo-domain.. The hPc gene that we have isolated is highly homol-ogouss to the XPc homolog, whereas the other human Pc homolog,, CBX2, is more homologous to the murine Pc ho-molog,, M33. For convenience, we named the CBX2 gene hPcl andd the gene that we isolated hPc2.
Distributio nn of hPc2 transcripts in human tissues and can-cerr cell lines. The hPc2 cDNA clone that we have identified is
1,8677 bp in length, of which 1,674 bp (encoding aa 1 to 558) encompassess the open reading frame. To study the distribution off hPc2 mRNA, we selected a probe which has littl e homology too the other vertebrate Pc genes, that is, between the conserved chromodomainn and C-terminal domain ranging from aa 60 to 530.. On a Northern blot, a single transcript of approximately 2.88 kb was detected in all the tissues and cell lines tested (Fig. 3).. In normal human tissues, the highest level of hPc2 expres-sionn is found in the thymus (Fig. 3A, lane 2) and in peripheral bloodd leukocytes (lane 8). Expression levels are still pro-nouncedd in the spleen (lane 1), prostate (lane 3), testis (lane 4), ovaryy (lane 5) and small intestine (lane 6) but are very low in thee colon (lane 7). This expression pattern of hPc2 is very similarr to the expression pattern of the vertebrate PcG genes BMI1BMI1 and HPH2. In human tissues, all three PcG genes are expressedd at a high level, except in the colon (16). The expres-sionn levels of hPc2 in several human cancer cell lines vary more thann in normal human tissues (Fig. 3B). The hPc2 transcripts aree hardly detectable in the Burkitt's lymphoma Raji (Fig. 3B, lanee 5) and lung carcinoma (lane 7) human cell lines. In con-trast,, the expression level of hPc2 is high in all other human celll lines.
Inn summary, the hPc2 gene is expressed in normal human tissuess with almost equal abundance. The hPc2 mRNA levels aree more variable in the different human cell lines.
Characterizationn of antibodies against hPc2 and hPcl/M33. Too further strengthen our conclusion that we have isolated a novell human Pc homolog that is different from the previously isolatedd human M33 homolog, CBX2 (hPcl) (15), we raised polyclonall antibodies against hPc2 and M33. We raised anti-bodiess against M33 since the full-length cDNA of CBX2 has nott been reported. The published CBX2 sequence is virtually identicall to M33 at the protein level, and we reasoned that polyclonall antibodies raised against M33 will recognize the
53 3

4.. Human Polycomb protein and cellular transformation
M33 3
CHROMODOMAI * * Humann P c2 HELPAfcGEHVFA^SIEHWIRKGRVEYLVKMWÏWSPKYrtTWEPEENIIiJPRLLIAF^ ^ 100
KELP ** TOEHVPAVESIEKKRIRKGRVEtLVKWRGHS-KYMTWEPEEKILDPRLL-AFC (RERQEQ-MGYRJUtGPKP K V- -P-FARRSKVL-GL-DS S V - A - E - I — K R - - K G - V E Y - V K W - G W — Y N T W E P E - N I L D - R - L I ™ ™ KRG- KK P S
- E L — p G E - V F A - E - II —KR-RKG~EYLVKWRGWS-K-N-WEPEENILOPRLL-AFd— E RKRG—P- P R L-
Humann P c2 XPc c
Humann P c2 XPc c
Humann P c2 XPc c
Humann P c2 XPc c
TDHRAJU.DI^A^KGQGHQYEIJlSlUUa^YePHSKEGKPRPPGKSGKYYYQLNSIU ^^ 2 00 - -NR-XLDL GG HQY-LNSKXHHQYQP-- K KYYYaLNSKXHH-YQPDPK-Y--Q Y KE T—D D Q—
T_00 KKH H H K s c m R s Q__ EE — S P E — P - P —K K K K - A — p R D A
GAKGYI^AVKPLAGAAGAPGKGSEKGPPNGMKPAP«AVTGNGIGCW«IVKHIW ^^ 300 GG p p p G-GGIUUCIVKNKNKNGRIVIVMSKYMENGMQ-VKIKS- E
vv G _ _ G - _ _ K _ s _ _ G p p K A E E
L- AA A - - A - - K - S - - GP P G—G S A K —A
HPPADRTFTUtAAGAEEKKVEAPPKRREEEVSGVSDPQPQDAGSRKLSPTKBAFGEQPLQLTTKPDLLAWDPARNTHPPSHHPHPHPHHHHHHHHHHHHA VV 4 00 TT K - V E K _ _E s R _ _ s E QPLQLTT K P—N
_pp K p p s L _ T R HHHHHHHHHH- -DD C _ E A _ _K G g GS — L - P T - E LQ AR P
GLWLSHVRKRCLSETHGEREPCKKRLTARSISTPTCLGCSPAAERPADLPPAAALRQPEVILLDSDLDEPIDLRSVKSRSEAGEPPSSLQVKPETPASA A A - LL RKRCLSE—G—E-CKK-LT -RS-S - P P L—P—ILLDSDLDEPIDLR-VKS R QV P - S— S00
H33 3 CBX22 ( h P c l )
p—KB-T—s_ _ - - G — A - EE P A -S E DSD-D L - S AGE—SS P GE—SSS P
XPc c
M33 3 CBX22 ( h P c l )
COOE-BOX X VAVAAAAAPTTTAEKPPAEAQDEPAESLSEFKPFFGH II ijlTDVTANCLTVTFK E fVTV
-S-FKPFFCNI-ITDVTANCLTVTFK EE f-T V ITDVT-N—TVT— E E - T D V T A N — T V T - K I — T ---TDVTAN--TVT-K EE — T -p p
B B h P c22 KSRSEACEPPSSLQVKPETPASAAVAVAAAAAPTTTAEKFPAEAQDEPAESLSErKPFFGN I I
CBX22 ( h P c l ) LPEMSAGEESSSSDSDPDSASPPSTGg.NP5VSVCTSQDWKPTRS
M333 LSELSTCEENSSSDSDPDSTSLPSAAQNLSVAIQTSQOWKPTRS
COOB-BOX X ITDVTANCLTVTFKjkVTV V
LIEHVP/TDVTANLITVTVKEE SPTSVCFFNLRHY
LIEHVP/TDVTANLITVTVKEE SPTSVCFFNLRHY
FIG.. 2. Comparison of the hPc2 protein with other Pc homologs. (A) The sequence is aligned to Xenopus Pc, Pc, M33, and the CBX2 (hPcl) protein sequences. Identicall amino acids are shown, and nonidentica! amino acids are indicated by dashes. The conserved chromodomain and C-terminal domain (COOH box) are boxed. (B)) Alignment of the identified amino acids of CBX2 (hPcl) with corresponding regions of the hPc2 and M33 proteins. Double dots indicate identical amino acids. andd single dots indicate conservative changes.
closelyy related human CBX2 protein as well. The predicted molecularr masses of the 558-aa hPc2 and the 519-aa M33 are 611 and 55 kDa, respectively. The fusion proteins include a 3-kDaa T7 tag. A mouse monoclonal antibody against T7 rec-ognizedd an 85-kDa T7-hPc2 fusion protein and a 79-kDa T7-M333 fusion protein in extracts of the E. coli strain in which the fusionn proteins were produced (Fig. 4, lanes 1 and 2, respec-tively).. Both proteins have an aberrant mobility on SDS-PAGE.. The predicted size difference of 6 kDa between hPc2 andd M33, however, remains visible on SDS-PAGE.
Wee raised polyclonal chicken antibodies against T7-tagged hPc22 and polyclonal rabbit antibodies against T7-tagged M33. Thee chicken anti-hPc2 antibody recognizes the T7-hPc2 fusion proteinn but not the T7-M33 fusion protein (Fig. 4, lanes 3 and 4,, respectively). The rabbit anti-M33 antibody recognizes the T7-M333 fusion protein but not the T7-hPc2 fusion protein (lanee 6 and 5, respectively). In extracts of U-2 OS human osteosarcomaa cells, the hPc2 antibody detected an 82-kDa proteinn (lane 7) and the M33 antibody detected a 76-kDa proteinn (lane 8). The detected molecular masses in extracts of humann cells are consistent with the molecular masses of the hPc22 and M33 fusion proteins. We will refer to the human proteinn that is detected by the anti-M33 antibody as hPcl/M33. Althoughh M33 is a murine protein, the human CBX2 (hPcl) proteinn is virtually identical to M33 (Fig. 2B), and it is there-foree likely that our anti-M33 antibody detects the hPcl protein inn extracts of human U-2 OS cells. In support of this notion, we
findfind that the hPc2 and M33 antibodies recognize proteins with identicall molecular masses of, respectively, 82 and 76 kDa in extractss of the murine P19 embryo carcinoma cell line (36) and thee murine mammary epithelial cell line C57MG (see also Fig. 7).. We conclude that the antibodies specifically recognize hPc2 andd hPcl/M33 in extracts of human and murine cells.
hPc22 and hPcl/M33 colocalize in nuclei of U-2 OS cells. We nextt analyzed the subcellular localization of the hPc2 protein inn relation to the hPcl/M33 protein by performing immuno-fluorescencefluorescence labelling experiments. The use of chicken anti-hPc22 and rabbit anti-M33 allowed double-labelling experi-ments.. We used human U-2 OS osteosarcoma cells in which severall human PcG genes are expressed at a high level (Fig. 3B,, lane 10) (1, 16). Both the hPc2 and hPcl/M33 proteins weree detected in the nucleus of U-2 OS cells, throughout the nucleoplasm.. They are detected in a fine granular pattern as welll as in large, brightly labelled domains (Fig. 5). hPc2 and hPcl/M333 colocalize in these large domains. Using these hPc2-andd M33-directed antibodies, we have shown previously that hPc22 coimmunoprecipitates and colocalizes with the vertebrate PcGG proteins BMI1 and HPH1 (35). Also, M33 protein coim-munoprecipitatess and colocalizes with BMI1 (1). We therefore concludee that hPc2 and hPcl/M33 colocalize in interphase nucleii of U-2 OS human osteosarcoma cells.
Ann hPc2 mutant lacking the conserved (..'-terminal region is nott able to repress gene activity. The PcG complex proteins aree known to be involved in repressing homeotic gene activity
54 4

ahPc22 aM33
Kb b
4.4--
^^ |É| M Ü M «i »» ^ ^ » ^ ^
11 2 3 4 5 6 7
v v VSSJ? VSSJ?
Kb b
4,4--
^__ ^^ ^~ ^~ ^_ ÉiKMft
1 2 3 4 5 6 7 88 9 10
FIG.. 3. Expression patterns of hPc2 in human tissues and human cancer cell lines.. (A) Expression levels in the spleen, thymus, prostate, testis, ovary, small intestine,, colon, and peripheral blood leukocytes. As a probe, we used the entire hPc22 clone, except for the conserved chromodomain and the C-terminal domain. Thee filter was rehybridized with a probe for glyceraldehyde-3-phosphate dehy-drogenasee (GAPDH) to verify the loading of RNA in each lane. (B) Expression levelss in promyelocyte leukemia HL-60, HeLa cell S3, chronic myelogenous leukemiaa K-562, lymphoblastic leukemia MOLT-4, Burkitt's lymphoma Raji, colorectall adenocarcinoma SW480, lung carcinoma A549, and melanoma G361 cells.. Lane 1 to 8 represent a commercially obtained Northern blot. We also isolatedd and blotted poly(A)4 RNA from U-2 OS. To allow comparison with the commerciall multiple-tissue Northern blot, we isolated and blotted poly(A)* RNAA from SW48Ü cells (lane 9).
inn Drosophila (21, 28). So far, no PcG protein has been found too bind directly to DNA (28). To investigate the ability of PcG proteinss to repress gene activity, they have been targeted to reporterr genes as LexA or Gal4 fusion proteins (4, 35). Previ-ously,, we have analyzed the ability of LexA-XPc and LexA-hPc22 fusion proteins to repress gene activity in different cell liness by using CAT reporter constructs (35). We observed that LexA-XPcc and LexA-hPc2 repress HSF-induced CAT expres-sionn to approximately 20% (35). In Drosophila, several natu-rallyy occurring Pc mutants either are mutated in the C-terminal domainn or lack this domain entirely (13). We tested the ability off an hPc2 deletion mutant protein, AhPc2, that lacks the last 300 aa of the C terminus (Fig. 2) to repress gene activity. U-2 OSS cells were transfected with a construct containing a tandem off four LexA operators, binding sites for the HSF transcrip-tionall activator, and the hsp70 TATA promoter immediately upstreamm of the LUC reporter gene. The endogenous HSF was usedd as a transcriptional activator. In the absence of HSF, no LUCC activity was observed (36). Maximum LUC activity in the presencee of HSF was set at 100% (Fig. 6, Control). Cotrans-fectionn of LexA alone had no significant influence on HSF-
•?? *F v v x> <r
1 22 3 4 5 6
FIG.. 4. Antibodies directed against the hPc2 and hPcl/M33 proteins. Full-lengthh cDNA clones of hPc2 and M33 were cloned into pET23 vectors to produce T7-taggedd fusion proteins. The T7-hPc2 (lane 1) and T7-M33 (lane 2) fusion proteinss were detected with a mouse monoclonal antibody against the T7 tag (Novagen).. A chicken antibody raised against the T7-hPc2 fusion protein detectedd the T7-hPc2 protein (lane 3) but not the T7-M33 protein (lane 4). A rabbit antibodyy raised against the T7-M33 fusion protein recognized the T7-M33 proteinn (lane 6) but not the T7-hPc2 protein (lane 5). In extracts of U-2 OS osteosarcomaa cells, the chicken anti-hPc2 and rabbit anti-M33 detected endogenouss hPc2 (lane 7) and hPcl/M33 (lane 8) proteins, respectively. Analysis was performedd with a 7% polyacrylamide gel. Molecular masses of the proteins are indicatedd (in kilodaltons).
inducedd (Fig. 6, 97% 6% [mean SEM], n = 4) LUC activity.. We found that LexA-hPc2 repressed LUC activity to approximatelyy 20% (14% 4% [mean SEM], n = 4). When LexA-AhPc22 is targeted to the reporter gene, LUC activity is nott significantly repressed (90% 9% [mean SEM], n = 4). Thee results show that the C-terminal deletion mutant, unlike hPc2,, is not able to repress HSF-induced LUC activity. This is inn agreement with earlier data which also showed that a Dro-sophilasophila Pc protein which lacks the C-terminal domain is unable too repress gene activity (4). This indicates that the conserved C-terminall region of hPc2 is involved in mediating gene repression. .
Expressionn of AhPc2 results in cellular transformation. To studyy potential functions of hPc2, we expressed the protein in severall different mammalian cell lines. In Drosophila, several naturallyy occurring Pc mutants either are mutated in the conservedd C-terminal domain or lack this domain entirely (13). A mutantt hPc2 protein that lacks the C-terminal domain has lost itss ability to repress gene activity (Fig. 6). On the other hand, thee chromodomain is essential for binding of the Pc protein to chromatinn (23). Based on the different functions of these two conservedd domains, we reasoned that a mutant hPc2 protein thatt possesses a functional intact chromodomain but does not containn the C-terminal domain might still be able to bind to the chromatinn but be unable to repress gene activity. Competition off the mutant hPc2 protein with the endogenous wild-type hPc22 protein might interfere with wild-type hPc2 function.
Too test this idea, we expressed wild-type hPc2 (hPc2WT) andd the C-terminal deletion mutant (AhPc2) in the murine mammaryy epithelial C57MG (2, 3, 26) and NIH 3T3 cell lines andd in the human U-2 OS osteosarcoma cell line. To test whetherr the proteins are indeed properly overexpressed, we analyzedd the hPc2 protein levels of the different, individual cloness by Western blotting (Fig. 7). We found higher levels of thee hPc2 protein in hPc2WT- (Fig. 7, lane 2) and in AhPc2 (lanee 3)-transfected C57MG cells than in the untransfected C57MGG cells (lane 1). The 82-kDa protein which is detected in thee untransfected C57MG cells is presumably the murine ho-mologg of hPc2. Similarly, we found higher levels of hPc2 proteinn in hPc2WT- (lane 5) and AhPc2 (lane 6)-transfected U-2 OSS cells than in the untransfected U-2 OS cells (lane 4). These
55 5

4.. Human Poly comb protein and cellular transformation
FIG.. 5. Colocalization of the hPc2 and hPcl/M33 proteins in nuclei of U-2 OS cells. Rabbit anti-M33 (A) and chicken anti-hPc2 (B) labelling is shown in single confocall optical sections. (A and B) Two different detection channels of the dual image; (C) false-color overlay. hPc2 and hPcl/M33 are homogeneously distributed inn the nucleus but are also concentrated in large, brightly labelled domains. hPc2 and hPcl/M33 colocalize in these large domains (C) (yellow).
resultss show that the transfected constructs are expressed in thee selected cell clones.
Whenn cultured to confluence, AhPc2-transfected C57MG cellss showed striking changes in morphology compared to the parentall cell line. Whereas untransfected C57MG cells exhibit aa "cuboidal" morphology (Fig. 8), C57MG/AhPc2 cells exhibit aa "streaming" phenotype, characterized by highly elongated cellss (Fig. 8). This latter phenotype is commonly observed in transformedd cells and is very similar to the phenotype obtained whenn the wntl oncogene is transfected into C57MG cells (Fig. 8)) (2, 3, 26). No phenotypic changes were observed in cells transfectedd with hPc2WT (Fig. 8). To test whether the pheno-typicc changes induced by the AhPc2 mutant were specific for thee deletion of the C-terminal domain, we also transfected a constructt containing a mutant lacking the chromodomain (Achromo)) but retaining the C-terminal domain. This did not resultt in phenotypic changes (36). We also transfected the AhPc2-transformedd cells with additional hPc2WT. This re-sultedd in reversal of the transformed phenotype (36), suggest-ingg that the cellular transformation caused by the AhPc2 mu-tantt is due to a dominant-negative effect.
Similarr phenotypic changes, characterized by elongated cells, weree observed in NIH 3T3 and U-2 OS cells after transfection withh AhPc2 but not after transfection with hPc2WT. However, transformedd C57MG cells are well characterized in terms of morphologyy and changes in the expression of marker genes, andd we therefore further characterized the C57MG cells. Transformedd C57MG cells that are transfected with either the neuT,neuT, wntl, or wnt2 gene have decreased expression of the wnt4wnt4 gene (2,3,14, 26). This indicates that wnt4 expression can bee used as a marker for cellular transformation of C57MG cells resultingg from the overexpression of proto-oncogenes. Consid-eringg the striking similarities between AhPc2-induced and M7i<7-inducedd phenotypic changes, we tested whether APc-in-ducedd phenotypic changes are accompanied by changes in wnt4 expressionn level. We found that wnt4 is strongly down-regu-latedd in both C57MG/AhPc2 and C57MG/tvnr; cells whereas noo changes in wnt4 levels were observed in C57MG/hPc2WT cellss (Fig. 9A). In contrast, overexpression of wild-type hPc2 didd not have such an effect.
Itt has further been shown that after reaching confluence, C51MGIwntlC51MGIwntl cells continue to divide, as measured by [^thy-midinee incorporation (2, 3, 26). Similarly, we found that
C57MG/AhPc22 but not C57MG/hPc2WT cells continued to proliferatee at the same rate as C57MG/HT»7 cells (Fig. 9B).
Wee conclude that expression of the AhPc2 mutant protein resultss in a partial transformation of C57MG, NIH 3T3 and U-22 OS cell lines. A likely mechanism is that this transforma-tionn is due to interference of the AhPc2 mutant protein with thee PcG protein complex.
Enhanced,, deregulated expression of c-myc in AhPc2-trans-formedd cells. The PcG protein complex is involved in repres-sionn of gene activity, and our results suggest that interference withh hPc2 protein function results in derepression of at least onee oncogene. Deregulated expression of this oncogene will subsequentlyy lead to cellular transformation. To test this idea, wee probed a Northern blot containing poly(A)+ mRNA of C57MG,, CSlMGIwntl, C57MG/hPc2WT, and C57MG/AhPc2
II 1 I UxAA binding ti les
* * * *
A A TT T-
Controii UxA UxA- UxA-hPe22 AhPe2
FIG.. 6. AhPc2, lacking the C-terminal domain, is not able to repress HSF-inducedd LUC gene activity. LUC reporter expression is maximally induced by HSFF in the absence of any LexA fusion protein (Control). Cotransfection of LexAA alone has hardly any influence on HSF-induced LUC expression, which is stilll about 97% of the total (LexA). Cotransfection of LexA and hPc2 represses LUCC activity fivefold. Cotransfection of LexA and AhPc2 has no significant repressionn effect on LUC activity, which is still 90% of the total. The bars representt the average degree of repression by LexA-hPc2 or LexA-AhPc2 in four independentt experiments (mean SEM). The actual values are indicated in the text. .
56 6

cfcf #* # c/ #" #
FIG.. 7. Western analysis of transfected hPc2 proteins from cell extracts of C57MGG and U-2 OS cells. Equal amounts of C57MG and U-2 OS cell extracts weree Western blotted. The blots were incubated with a rabbit anti-hPc2 antibody. Endogenouss hPc2 levels were detected in the untransfected U-2 OS cells (lane 4).. Endogenous levels of a murine hPc2 homolog were detected in untransfected C57MGG cells (lane 1). Elevated levels of the hPc2 protein were detected in the hPc2WT-- (lanes 2 and 5) and in the ihPc2 (lanes 3 and 6)-transfccted cell lines. Analysiss was performed with a 10% polyacrylamide gel. Molecular masses of the proteinss are indicated (in kilodaltons). The C-terminal 30-aa deletion was too smalll to be detected on these Western blots.
cellss with cDNA fragments of several oncogenes. We found thatt in C57MG/AhPc2 cells the expression of the c-myc proto-oncogenee is strikingly enhanced, concomitant with a decrease inn the wnt4 expression level (Fig. 10). In C57MGAvm7 cells, expressionn of c-myc was also increased, but this increase was sixfoldd lower than that in C57MG/AhPc2 cells. No significant
changess in the expression levels of the bmil, bcl-2, c-fos, and c-junc-jun oncogenes were observed (36). This underlines the spec-ificityy of the effect of AhPc2 expression on c-myc expression in C57MG/AhPc22 cells.
Thee U-2 OS osteosarcoma cell line shows phenotypic char-acteristicss of cellular transformation similar to those seen in C57MGG after the transfection with AhPc2. We therefore also examinedd the RNA levels of c-myc in the different U-2 OS clones.. We found that in U-2 OS/AhPc2 cells the expression of c-mycc-myc is enhanced (Fig. 11 A, lane 3) compared to the control U-22 OS cells (lane 1). Surprisingly, we detected reduced c-myc RNAA levels in U-2 OS/hPc2WT cells (lane 2). This effect was specificc for c-myc expression, since no reduced c-fos and c-jun expressionn in U-2 OS/hPc2WT cells was observed (36). We alsoo analyzed the expression of the 67-kDa c-myc protein, usingg the 9E10 monoclonal antibody, which specifically detects thee human c-myc protein (8). We found that also the c-myc proteinn levels are elevated in U-2 OS/AhPc2 (Fig. 11B, lane 3) cellss in comparison with control U-2 OS cells (lane 1). As with c-mycc-myc RNA levels, a lower level of c-myc protein was found in U-22 OS/hPc2WT cells (lane 2).
Inn many cell lines, as is the case for C57MG, the expression levelss of c-myc are very low. Therefore, we needed to isolate poly(A)++ RNA to be able to detect c-myc RNA in C57MG cells.. On the other hand, c-myc levels in U-2 OS cells are knownn to be relatively high. In many osteosarcomas, the c-myc genee is amplified (17, 37). Therefore, we were able to detect
C57MG G
C57MG// hPc2WT
C57MG// wntl
C57MG// AhPc2
FIG.. 8. Overexpression of a AhPc2 deletion mutant induces partial transformation of C57MG cells. The C57MG cells were stably transformed with the wntl oncogenee (C57MG/wntl), the full-length hPc2 gene (C57MG/hPc2WT), and the AhPc2 deletion mutant, which lacks the conserved C-terminal domain (C57MG/ AhPc2).. The cells were cultured to confluence before the photographs were taken.
57 7

4.. Human Polycomb protein and cellular transformation
fff/f fff/f
MMm m
GAPDH H
&& # # 4 $
, ,
FIG.. 9. Changes in gene expression and growth characteristics in C57MG/ AhPc22 cells. (A) Poly(A)* mRNA of C57MG control cells (lane 1). C57MG cells transfectedd with only the pcDNA3 plasmid carrying the neomycin resistance genee (lane 2), C57MG/KW/ (lane 3), C57MG/hPc2WT (lane 4), and C57MG/ AhPc22 (lane 5) cells was Northern blotted and probed with a fragment of the wnt4wnt4 gene. To verify equal RNA loading, the filter was hybridized with a GAPDHH probe. (B) Untransfected C57MG cells and C57MG/NEO, C57MG/ wnll,wnll, C57MG/hPc2WT, and C57MG/AhPc2 cells were cultured to confluence, andd pHJthymidine was added to the culture medium. The cells were cultured for ann additional 2 days before [3H]thymidine incorporation was determined. The countss were normalized to the protein content. The pHlthymidine incorporation byy control, untransfected C57MG cells is set at 100%. Representative results of fourr independent experiments are shown.
c-mycc-myc RNA expression in total RNA in U-2 OS cells. The differencee in expression levels of c-myc in C57MG and U-2 OS cellss is also a likely explanation for our detection of a reduction inn c-myc expression levels in U-2 OS/hPc2WT cells but not in C57MG/hPc2WTT cells. In the last cell line, the c-myc levels are alreadyy low.
Inn summary, we find that c-myc expression is elevated after expressionn of the AhPc2 protein in two different cell lines. We concludee that interference with the expression of hPc2 results inn deregulated, enhanced c-myc transcription. At the same time,, a reduction of c-myc RNA and c-myc protein levels is detectedd in U-2 OS cells by the overexpression of hPc2WT, providingg extra evidence for a role of hPc2 in the regulation of c-mycc expression.
AhPc22 induces anchorage-independent growth. Rat la cells, aa fibroblast cell line, have been found to transform and display anchorage-independentt growth in soft agarose by overexpres-sionn of c-myc alone (40, 44). Previous studies have shown that Bmi-1,, a vertebrate PcG protein and oncoprotein, is able to inducee cellular transformation and anchorage-independent growthh in Rat la cells (5). Since overexpression of AhPc2 enhancesenhances c-myc transcription, it is possible that AhPc2, like
c-myc,, is able to induce anchorage-independent growth of Rat laa cells. We therefore used Rat la cells to test the transform-ingg effect of AhPc2 as an independent assay for cellular trans-formation. .
Ratt la cells were transfected with c-myc (Rat la-myc), hPc2WTT (Rat la-hPc2WT), AhPc2 (Rat la-AhPc2), and an hPc22 mutant lacking the chromodomain (Rat la-Achromo). Thee results demonstrate that AhPc2 (Rat la-AhPc2) as well as c-mycc (Ratla-myc) overexpression alone is sufficient to trans-formm cells (Table 1). The number of colonies induced by AhPc2 overexpressionn is comparable to the effect of c-myc overexpres-sionn alone. In contrast, overexpression of hPc2WT and Achromoo did not induce colonies of Ratla cells (Table 1). Thesee results demonstrate that AhPc2 is able to induce an-chorage-independentt growth when expressed in Rat la cells.
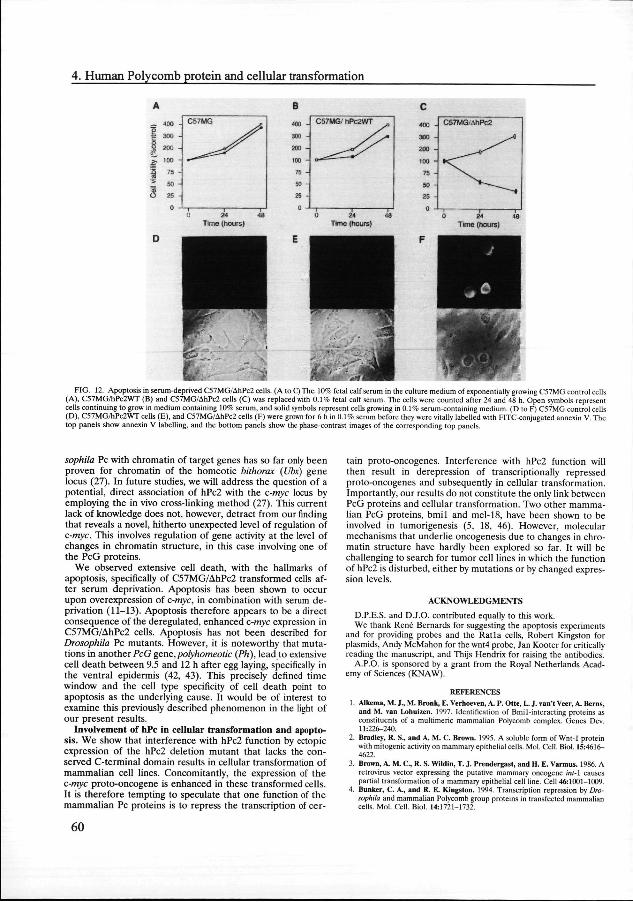
Inductionn of apoptosis, specifically in Ah I'c2-transformed cells.. We addressed the functional significance of the enhanced c-mycc expression in C57MG/AhPc2 and U-2 OS/AhPc2 cells. It hass been shown that overexpression of c-myc, in combination withh exposure to culture medium containing a low serum con-centration,, leads to apoptosis (9-11, 47). To test whether the enhancedd expression of c-myc in the two different cell lines also resultss in apoptosis, we cultured the cells in 0.1% serum. We observedd extensive cell death of C57MG/AhPc2 cells 24 to 48 h afterr serum deprivation (Fig. 12C). In contrast, the numbers of C57MGG cells and C57MG/hPc2WT cells had increased three-too fourfold 48 h after serum deprivation (Fig. 12A and B, respectively).. One of the earliest hallmarks of apoptosis is the redistributionn of phosphatidylserine from the inner face of the plasmaa membrane to the cell surface (20, 22). This redistribu-tionn can be detected with an FITC conjugate of annexin V, a proteinn that has a high affinity for phosphatidylserine (20, 22). Afterr 4 to 6 h of serum deprivation, we observed a substantial numberr of C57MG/AhPc2 cells and cells that could be labelled withh annexin V-FITC at the outside of the cell, 9% 3% in fivefive independent experiments (Fig. 12F). In contrast, no an-nexinn V-FITC-positive cells were observed in C57MG and C57MG/PcWTT cells 6 h after serum deprivation (Fig. 12D and E).. Similar results were detected with U-2 OS cells (36). We concludee that serum deprivation of C57MG/AhPc2 and U-2 OS/AhPc22 cells results in extensive cell death, which shows hallmarkss of apoptosis.
&/Z &/Z
WNT44 i 'S« 1 22 3 4
FIG.. 10. c-myc expression is enhanced in C57MG/AhPc2 cells, Poly(A)* mRNAA of C57MG control cells (lane 1) and C57MG/wntl (lane 2), C57MG/ hPc2WTT (lane 3), and C57MG/AhPc2 cells (lane 4) was Northern blotted and probedd with fragments of the c-myc and wnt4 genes. To verify equal RNA loading,, the filter was hybridized with a GAPDH probe.
58 8

>+& >+&
** / . r r
p p
82--
49--
33--
FIG.. 11. c-myc expression is enhanced in U-2 OS/AhPc2 cells. (A) Total RNAA of U-2 OS control (lane 1), U-2 OS/hPc2WT (lane 2), and U-2 OS/AhPc2 (lanee 3) cells was Northern blotted and probed with a fragment of the c-myc gene.. To verify equal RNA loading, the filter was hybridized with a GAPDH probe.. (B) Cell extracts of U-2 OS control (lane 1), U-2 OS/hPc2WT (lane 2), andd U-2 OS/AhPc2 (lane 3) cells were analyzed on a Western blot. The blots weree incubated with the mouse monoclonal antibody 9E10, which specifically recognizess the human c-myc protein. Molecular masses of the proteins are indicatedd (in kilodaltons).
DISCUSSION N
Twoo human Pc homologs exist. The Pc protein is involved in thee stable and heritable repression of gene activity during DrosophilaDrosophila development. To study the functions of vertebrate PcPc homologs, we have isolated and characterized a novel hu-mann Pc homolog. To isolate this gene, we used the Xenopus homologg of Pc, XPc, as a probe. Overall, the novel human Pc homologg has 48% identity and 80% similarity to the XPc protein.. Based on sequence homologies with another, previ-ouslyy characterized human Pc homolog, CBX2 or hPcl, and thee murine Pc homolog, M33, we conclude that we have iso-latedd a novel human Pc homolog, hPc2, which is distinct from hPcl/M33.hPcl/M33. Whereas the hPc2 protein is most closely related to XPc,, the hPcl protein is most closely related to M33.
Itt appears to be a common feature among vertebrate PcG proteinss that each of them exists as a pair of closely related proteins.. For instance, the vertebrate PcG proteins Bmil (33) andd mel-18 (45) have large identical regions (alignment shown inn reference 33). Furthermore, we recently identified two hu-mann proteins, HPH1 and HPH2, that both have extensive sequencee homology to the Drosophila PcG protein Polyho-meoticc (Ph) in two conserved homology domains (16). Strik-ingly,, homology between the HPH1 and HPH2 proteins them-selvess is restricted to these conserved homology domains; outsidee these domains, homologies are very limited. This is veryy similar to what we report here for hPcl/M33 and hPc2. Overalll homologies between hPc2 and hPcl/M33 do not ex-ceedd the homologies between these vertebrate Pc homologs andd Drosophila Pc. It is important, however, to note that when nott only identical amino acids but also conservative changes aree taken into consideration, the similarity between hPc2 and
hPcl/M333 is 68%. This is significantly higher than the 29% identity.. As already pointed out, this could indicate that the overalll three-dimensional structures of hPc2 and hPcl/M33 are veryy similar, which may imply that the two proteins are func-tionallyy equivalent. It is also possible, however, that the strong conservationn in the chromodomain and C-terminal domain is enoughh to provide functional equivalence to the proteins. Al-thoughh homology between M33 and Drosophila Pc is restricted too the chromodomain and the C-terminal domain, M33 is able too partly rescue the Drosophila Pc phenotype when overex-pressedd in the Pc mutant (25). This indicates that hPcl/M33 cann be considered a functional homolog of Pc, and it signifies thee importance of the conserved regions. The potential func-tionall relationship between the two human Pc homologs is furtherr underlined by our finding that hPcl/M33 and hPc2 colocalizee in nuclei of human U-2 OS cells, suggesting that hPcll and hPc2 are part of a human PcG protein complex. In thiss context, it is significant that the human PcG protein BMI1 andd the human Polyhomeotic-related HPH1 and HPH2 pro-teinss also colocalize with hPc2 and hPcl/M33 in the same nuclearr domains of several human cell lines (1, 16).
Wee conclude that there are at least two human Pc homologs. Itt is not clear why two closely related human Pc proteins exist. Sincee other vertebrate PcG-related proteins have been found too exist as functional pairs as well, this may have functional significance.. It is possible that small differences induce subtle changess in, for instance, their specificities for binding to target genes. .
c-mycc is a potential target gene of hPc2. In Drosophila, PcG proteinss have been identified as repressors of gene expression. Thee only identified target genes of PcG proteins are homeotic geness and gap genes (28, 31, 38, 48). These genes are all involvedd in developmental decisions. In this study we found thatt expression of a mutant hPc2 cDNA in two different mam-maliann cell lines, U-2 OS and C57MG, results in deregulated, enhancedd expression of a gene that controls a different process. Thiss gene, c-myc, is involved in cell cycle and differentiation events.. The mutant hPc2 protein lacks a conserved C-terminal domainn that is crucial for the ability of the hPc2 protein to represss gene activity. Further, overexpression of the wild-type hPc22 cDNA results in decreased expression of c-myc. It is thereforee likely that it is due to interference with hPc2 function thatt c-myc expression is deregulated and enhanced. From thesee data, however, it cannot be concluded whether the effect onn c-myc expression is a direct or indirect effect. It is tempting too speculate that hPc2 interacts directly with the c-myc locus. Unfortunately,, the fact that Pc proteins binds to chromatin and nott to naked DNA excludes the use of standard methods, such ass DNA footprinting, to assess whether the hPc2 protein is physicallyy associated with the c-myc locus. Association of Dro-
TABL EE 1. Colony formation by AhPc2-transfected Rat 1A cellss in soft agarose
No.. of colonies/5 x 104
C o n s t r u ctt transfected cells"
pRcCMVV c-myc 451 52 pcDNA33 0 pcDNA3-hPc2WTT 0 PcDNA3-AhPc22 337 27 pcDNA3-Achromoo 0
"" A total of 5 x 104 of each pool of transfected and geneticin-selected cells weree seeded into 0.4% top agarose, and colonies with diameters of >0.1 mm weree counted 14 to 21 days after seeding (34, 40, 44). The entire procedure, includingg the transfection of the cDNAs, was performed in triplicate, and the meann SEM is shown.
59 9
4.. Human Polycomb protein and cellular transformation
—— 400 -
|| 300 -
o o 11 200 -
ff 100 -
11 75~ 22 se-ess 26 -
C57M6 6
y* y* ^ = ^ ^
Timee (hours)
FIG.. 12. Apoptosis in serum-deprived C57MG/AhPc2 cells. (A to C) The 10% fetal calf serum in the culture medium of exponentially growing C57MG control cells (A),, C57MG/hPc2WT (B) and C57MG/AhPc2 cells (C) was replaced with 0.1% fetal calf serum. The cells were counted after 24 and 48 h. Open symbols represent cellss continuing to grow in medium containing 10% serum, and solid symbols represent cells growing in 0.1% serum-containing medium. (D to F) C57MG control cells (D),, C57MG/hPc2WT cells (E), and C57MG/AhPc2 cells (F) were grown for 6 h in 0.1% serum before they were vitally labelled with FITC-conjugated annexin V. The topp panels show annexin V labelling, and the bottom panels show the phase-contrast images of the corresponding top panels.
sophilasophila Pc with chromatin of target genes has so far only been provenn for chromatin of the homeotic bithorax (Ubx) gene locuss (27). In future studies, we wil l address the question of a potential,, direct association of hPc2 with the c-myc locus by employingg the in vivo cross-linking method (27). This current lackk of knowledge does not, however, detract from our finding thatt reveals a novel, hitherto unexpected level of regulation of c-myc.c-myc. This involves regulation of gene activity at the level of changess in chromatin structure, in this case involving one of thee PcG proteins.
Wee observed extensive cell death, with the hallmarks of apoptosis,, specifically of C57MG/AhPc2 transformed cells af-terr serum deprivation. Apoptosis has been shown to occur uponn overexpression of c-myc, in combination with serum de-privationn (11-13). Apoptosis therefore appears to be a direct consequencee of the deregulated, enhanced c-myc expression in C57MG/AhPc22 cells. Apoptosis has not been described for DrosophilaDrosophila Pc mutants. However, it is noteworthy that muta-tionss in another PcG gene,polyhomeotic (Ph), lead to extensive celll death between 9.5 and 12 h after egg laying, specifically in thee ventral epidermis (42, 43). This precisely defined time windoww and the cell type specificity of cell death point to apoptosiss as the underlying cause. It would be of interest to examinee this previously described phenomenon in the light of ourr present results.
Involvementt of hPc in cellular transformation and apopto-sis.. We show that interference with hPc2 function by ectopic expressionn of the hPc2 deletion mutant that lacks the con-servedd C-terminal domain results in cellular transformation of mammaliann cell lines. Concomitantly, the expression of the c-mycc proto-oncogene is enhanced in these transformed cells. I tt is therefore tempting to speculate that one function of the mammaliann Pc proteins is to repress the transcription of cer-
tainn proto-oncogenes. Interference with hPc2 function will thenn result in derepression of transcriptionally repressed proto-oncogeness and subsequently in cellular transformation. Importantly,, our results do not constitute the only link between PcGG proteins and cellular transformation. Two other mamma-liann PcG proteins, bmil and mel-18, have been shown to be involvedd in tumorigenesis (5, 18, 46). However, molecular mechanismss that underlie oncogenesis due to changes in chro-matinn structure have hardly been explored so far. It will be challengingg to search for tumor cell lines in which the function off hPc2 is disturbed, either by mutations or by changed expres-sionn levels.
ACKNOWLEDGMENTS S
D.P.E.S.. and D.J.O. contributed equally to this work. Wee thank René Bernards for suggesting the apoptosis experiments
andd for providing probes and the Ratla cells, Robert Kingston for plasmids,, Andy McMahon for the wnt4 probe, Jan Kooter for critically readingg the manuscript, and Thijs Hendrix for raising the antibodies.
A.P.O.. is sponsored by a grant from the Royal Netherlands Acad-emyy of Sciences (KNAW).
REFERENCES S 1.. Alkema, M. J., M. Bronk, E. Verhoeven, A. P. Otte, L. J. van't Veer, A. Berns,
andd M. van Lohuizen. 1997. Identification of Bmil-interacting proteins as constituentss of a multimeric mammalian Polycomb complex. Genes Dev. 11:226-240. .
2.. Bradley, R. S., and A. M. C. Brown. 1995. A soluble form of Wnt-1 protein withh mitogenic activity on mammary epithelial cells. Mol. Cell. Biol. 15:4616-4622. .
3.. Brown, A M. C, R. S. Wildin, T. J. Prendergast, and H. E. Varmus. 1986. A retroviruss vector expressing the putative mammary oncogene int-\ causes partiall transformation of a mammary epithelial cell line. Cell 46:1001-1009.
4.. Bunker, C. A, and R. E. Kingston. 1994. Transcription repression by Dro-sophilasophila and mammalian Polycomb group proteins in transfected mammalian cells.. Mol. Cell. Biol. 14:1721-1732.
60 0

5.. Cohen, K. J, J. S. Hanna, J. E. Prescott, and C. V. Dane- 1996. Transfor-mationn by the Bmi-1 oncoprotein correlates with its subnuclear localization butt not its transcriptional suppression activity. Mol. Cell. Biol. 16:5527-5535.
6.. Core, N., S. Bel, S. J. Gaunt, M. Aurrand-Lions , J. Pearce, A. Fischer, and M.. Djabali. 1997. Altered cellular proliferatio n and mesoderm patterning in porycomb-M33-denciemm mice. Development 124:721-729.
7.. Duncan, L, and E. B. Lewis. 1982. Genetic control of body segment differ-entiationn in Dwsophiia, p. 533-554. In G. Subtelny (ed.), Developmental order:: its origin and regulation. Alan R. Liss, Inc., New York, N.Y.
8.. Evan, G. I., G. K. Lewis, G. Ramsay, and M. Bishop. 1985. Isolation of monoclonall antibodies specific for human c-myc proto-oncogene product. Mol.. Cell. Biol. 5:3610-3616.
9.. Evan, G, E. Harrington , A. Fanadi, H. Land, B. Araati , and M. Bennett. 1994.. Integrated control of cell proliferatio n and cell death by the c-myc oncogene.. Philos. Trans. R. Soc. London Ser. B 345:269-275.
10.. Evan, G, A. Wyllie, C. Gilbert , T. Littlewood, H. Land, M. Brooks, C. Waters,, L. Penn, and D. Hancick. 1992. Induction of apoptosis in fibroblasts byy c-myc protein. Cell 69:119-125.
11.. FanidL A-, E. A. Harrington , and G. Evan. 1992. Cooperative interaction betweenn c-myc and bcl-2 proto-oncogenes. Nature (London) 359:554-556.
12.. Franke, A, M. DeCamillis, D. Zink , N. Cheng, H. W. Brock, and R. Paro. 1992.. Pofycomb and pofyhomeotic are constituents of a multimeri c protein complexx in chromatin of Dwsophiia melanogaster. EMBO J. 11:2941-2950.
13.. Franke, A, S„ Messmer, and R. Paro. 1995. Mapping functional domains of thee Polycotnb protein cif Dwsophiia melanogaster. Chromosome Res. 3:351— 360. .
14.. Gavin, B. J., and A. P. McManon. 1992. Differential regulation of the Wnl genee family during pregnancy and lactation suggests a role in postnatal developmentt of the mammary gland. Mol. Cell. Biol. 12:2418-2423.
15.. Gecz, J, S. J. Gaunt, E. Passage, R. D. Burton, C. Cudrey, J. J. Pearce, and M.. Pontes. 1995. Assignment of a Porycomb-like chromobox gene (CBX2) to humann chromosome 17q25. Genomics 26:130-133.
16.. Gunster, M. J, D. P. E. Satijn, R. M. Hamer, J. L. den Blaanwen, D. de Braijn ,, M. J. Alkema, M. van Lohuizen, R. van Driel, and A. P. Otte. 1997. Identificationn and characterization of interactions between the vertebrate Porycomb-groupp protein BMI 1 and human homologs of Poh/homeotic. Mol. Cell.. Biol. 17:2326-2335.
17.. Isfort , R. J, D. B. Cody, and C. J. Doersen. 1995. Analysis of oncogenes, tumorr suppressor genes, autocrine growth-factor production, and differen-tiationn state of human osteosarcoma cell lines. Mol. Carcinog. 14:170-178.
18.. Kanno, M , M. Hasegawa, A. Ishida, K. Isono, and M. Tanigucbi. 1995. mef-18, aa Pofycomb group-related mammalian gene, encodes a transcriptional negative regulatorr with tumor suppressive activity. EMBO J. 14^672-5678.
19.. Kingston, R. F_, C. A. Banker, and A. N. Imbalzano. 1996. Repression and activationn by multiprotei n complexes that alter chromatin structure. Genes Dev.. 10:905-920.
20.. Koopman, G, C. P. M. Reutelingsperger, G. A. M. Kuijten , R. M. J. Keeh-nen,, S. T, Pals, and M. H. J. van Oers. 1994. Annexin V for flow cytometric detectionn of phosphatidylserine expression on B cells undergoing apoptosis. Bloodd 84:1415-1420.
21.. Lewis, E. B. 1978. A gene complex controlling segmentation in Drosophila. Naturee (London) 276:565-570.
22.. Martin , S. J, C. P. M. Reutelingsperger, A. J. McGahon, J. A. Rader, R. C. vann Schle, D. M. LaFace, and D. R. Green. 1995. Early redistributio n of plasmaa membrane phosphatidylserine is a general feature of apoptosis le-gardlesss of the initiatin g stimulus: inhibitio n by overexpression of Bcl-2 and Abl.. J. Exp. Med. 182:1545-1556.
23.. Messmer, S., A. Franke, and R. Paro. 1992. Analysis of the functional role of thee Pofycomb chromo domain in Drosophila melanogaster. Genes Dev. 6: 1241-1254. .
24.. Muller , J. 1995. Transcriptional silencing by the Polycomb protein in Dro-sophilasophila embryos. EMBO J. 14:1209-1220.
25.. Muller , J., S. J. Gaunt, and P. A. Lawrence. 1995. Function of the Polycomb proteinn is conserved in mice and flies. Development 121:2847-2852.
26.. Olson, D. J-, and J. Papkoff. 1994. Regulated expression of Wnt family
memberss during proliferatio n of C57mg mammary cells. Cell Growth Differ. 5:197-206. .
27.. Orlando, V, and R. Paro. 1993. Mapping /•o/ycomfr-repressed domains in the BithoraxBithorax complex using in vivo formaldehyde cross-linked chromatin. Cell 75:1187-1198. .
28.. Paro, R. 1990. Imprinting a determined state into the chromatin of Drosoph-ila.ila. Trends Genet. 6:416-421.
29.. Paro, tL, and D. S. Hogness. 1991. The Pofycomb protein shares a homologouss domain with a heterochromatin-associatedd protein of Drosophila. Proc. Natl.. Acad. Sci. USA 88:263-267.
30.. Pearce, J. J. H, P. B. Singh, and S. J. Gaunt 1992. The mouse has a Pofycomb-\VttPofycomb-\Vtt chromobox gene. Development 114:921-929.
31.. Pelegri, F, and R. Lehmann. 1994. A role of Pofycomb group genes in the regulationn of Gap gene expression in Drosophila. Genetics 136:1341-1353.
32.. Rastelli, 1», C. S. Chan, and V. Pirotta. 1993. Related chromosome binding sitess for zeste, suppressors of zeste and Polycomb group proteins in Drosophik andd their dependence on Enhancer of zeste function. EMBO J. 12:1513-1522.
33.. Reijnen, M. J , K. M. Hamer, J. L den Blaauwen, C. Lambrechts, L Schoa-eveld,, R. van Driel, and A. P. Otte. 1995. Pofycomb and bmi-1 homologs are expressedd in overlapping patterns in Xenopus embryos and are able to interactt with each other. Mech. Dev. 53:35-46.
34.. Samuels, M. L, M. J. Weber, J. M. Bishop, and M. McMahon. 1993. Conditionall transformation of cells and rapid activation of the mitogen-activatedd protein kinase cascade by an estradiol-dependent human Raf-1 proteinn kinase. Mol. Cell. Biol. 13:6241-6252.
35.. Satijn, D. P. E, M. J. Gunster, J. van der Vlag, K. M. Hamer, W. Scnul, M. J. Alkenu,, A. J. Saurin, P. S. Freemont, R. van Driel, and A. P. Otte. 1997. RING11 is associated with the Poh/comb-group protein complex and acts as aa transcriptional repressor. Mol. Cell. Biol. 17:4105-4113.
36.. Satijn, D. P. E , and A. P. Otte. Unpublished data. 37.. Scbon, A, L. Mkhiels, M. Janowskt J. Merregaert, and V. Erfle. 1986.
Expressionn of proto-oncogenes in murine osteosarcomas. Int. J. Cancer 38:67-74. .
38.. Simon, J. 1995. Locking in stable states of gene expression: transcriptional controll during Drosophila development. Curr. Opin. Cell Biol. 7:376-385.
39.. Singh, P. B., J. K. Miller, J. Pearce, R. Kothaiy, R. D. Burton, R. Paro, T. C. James,, and S. Gaunt 1991. A sequence motif found in a Drosophila hetero-chromatinn protein is conserved in animals and plants. Nucleic Acids Res. 19:789-794. .
40.. Small, M. B, N. Hay, M. Schwab, and J. M. Bishop. 1987. Neoplastic transformationn by the human gene Nvnyc. Mol. Cell. Biol. 7:1638-1645.
41.. Smith, P. K-, R. I. Krobn, G. T. Hennanson, A. K. Mallia, F. H. Gartner, M.. D. Provenzano, E. K. Fujimoto, N. M. Goeke, B. J. Olson, and D. C. Klenk.. 1985. Measurement of protein using bicinchoninic acid. Anal. Bio-chem.. 150:76-85.
42.. Smoose, D., C. Goodman, A. Mahowald, and N. Perrimon. 1988. pofyho-meotic:meotic: a gene required for the embryonic development of axon pathways in thee central nervous system of Drosophila. Genes Dev. 2:830-842.
43.. Smouse, D , and N. Perrimon. 1990. Genetic dissection of a complex neurologicall mutant, pofyhomeotic, in Drosophila. Dev. Biol. 139:169-185.
44.. Stone, J-, T. de Lange, G. Ramsay, E. Jakobovits, J. M. Bishop, H. Varmus, andd W. Lee. 1987. Definition of regions in human c-myc that are involved in transformationn and nuclear localization. Mol. Cell. Biol. 7:1697-1709.
45.. Tagawa, M„ T. Sakamoto, K. Shigemoto, H. MaUnbara, Y. Tamara, T. Ito, I.. Nakamura, A. OUtsu, K. Imai, and M. Tanigndtf. 1990. Expression of novell DNA-binding protein with zinc finger structure in various tumor cells. J.. Biol. Chem. 265:20021-20026.
46.. Van Lohuizen, M„ S. Verbeek, B. Scneijen, E. Wienies, H. van der Gulden, andd A. Berns. 1991. Identification of cooperating oncogenes in E(mu)-myc transgenicc mice by provirus tagging. Cell 65:737-752.
47.. Wagner, A. J, J. M. Kokontis, and N. Hay. 1994. Myc-mediated apoptosis requiress wild-type p53 in a manner independent of cell cycle arrest and the abilityy of p53 to induce v2\w°f" cil". Genes Dev. 8:2817-2830.
48.. Zink, B., and R. Paro. 1989. In vivo binding pattern of a transregulator of homeoticc genes in Drosophila melanogaster. Nature (London) 337:468-471.
61 1

62 2




![UvA-DARE (Digital Academic Repository) Femtosecond …Chapterr 3 Solventt dependence of (sub)picosecond proton transfer inn photo-excited [2,2'-bipyridyI]-3,3'-diol Abstract t Weereporton(sub](https://static.fdocuments.us/doc/165x107/6008fb96ae77ee7b5d7f2532/uva-dare-digital-academic-repository-femtosecond-chapterr-3-solventt-dependence.jpg)




![UvA-DARE (Digital Academic Repository) Studies towards ... · CHAPTERR 2 STUDIESSTOWARDSlAZAADAMANTANE2CARBOXYLICACIDDERIVATIVES VIAVIAAZABICYCLO[3.3.1]NONENES 2.11 Introduction ...](https://static.fdocuments.us/doc/165x107/5e3087da1b14c3103013a01b/uva-dare-digital-academic-repository-studies-towards-chapterr-2-studiesstowardslazaadamantane2carboxylicacidderivatives.jpg)
![UvA-DARE (Digital Academic Repository) The internet and … · CHAPTERR SEVEN - I DEFINE MY OWN IDENTITY Introductionn Theeconcep to frac eitsel f[is ]inextricablywovenou to fth ehistor](https://static.fdocuments.us/doc/165x107/60c9d1cff7b86705ce6018ed/uva-dare-digital-academic-repository-the-internet-and-chapterr-seven-i-define.jpg)







