![TumorBehaviorinTransitionalCellCarcinomaoftheBladderinRela ...cancerres.aacrjournals.org/content/47/24_Part_1/6800.full.pdf · [CANCERRESEARCH47,6800-6805,December!5,1987] TumorBehaviorinTransitionalCellCarcinomaoftheBladderinRelationto](https://static.fdocuments.us/doc/165x107/5c085d3a09d3f23a458c00a0/tumorbehaviorintransitionalcellcarcinomaofthebladderinrela-cancerresearch476800-6805december51987.jpg)
TheIntegrina6ß4onTA3 ...cancerres.aacrjournals.org/content/53/15/3611.full.pdf ·...
8
[CANCER RESEARCH 53. 3611-3617. August I. 1993] The Integrin a6ß4on TA3/Ha Mammary Carcinoma Cells Is Involved in Adhesion to Hepatocytes1 Hans Kemperman, Yvonne Wijnands, Diana de Rijk, and Ed Roos2 Division of Cell Biology. The Netherlands Cancer Institute. Amsterdam, the Netherlands ABSTRACT lA.VIla mammary carcinoma cells form liver métastases upon in- traportal injection. We have studied the interaction between these cells and hepatocytes that is likely to be important for liver metastasis forma tion. We show that the integrin «,,/i4.which is highly expressed on TA3 cells, is involved in this interaction. Fab fragments, generated from a polyclonal serum against TA3 cells, inhibited TA3-hepatocyte adhesion. By affinity purification on purified •¿Â»,,)>.,. we isolated a6fJ4-specific Fab frag ments from this anti-TA3 serum. These antibodies were highly spécifiefor c*604, as demonstrated by Western blot analysis and immunoprecipitation, and inhibited TA3-hepatocyte adhesion. This shows that a6ß4, which thus far has only been implicated in cell-matrix adhesion, can also mediate interactions with cell surfaces. Our results suggest that the high levels of <*6ß4 often expressed by metastatic carcinoma cells contribute to liver metastasis formation. INTRODUCTION Carcinomas, i.e., tumors originating from epithelial cells, constitute 90% of human malignancies, and metastasis formation by these tu mors is the major cause of therapeutic failure ( 1). For many carcino mas the liver is a major site of metastasis (2). A key step in the metastatic process is the adhesion of circulating tumor cells to cells and extracellular matrix components in the target organ. Therefore it is likely that cellular adhesion molecules, which mediate such inter actions, are major determinants of metastasis (3), and recently direct evidence has been provided for this notion. Chan et al. (4) showed that nonmetastatic rhabdomyosarcoma cells transfected with the copy DNA encoding the integrin VLA-23 (a2ßi),exhibited increased ad hesion to collagen and laminin, and produced lung métastasesupon i.v. injection, whereas untransfected cells and control transfectants did not. Furthermore, we have shown that mutants of a T-cell hybridoma deficient in expression of the integrin LFA-1 (aLß2) exhibited greatly reduced metastatic potential (5). For carcinomas, however, there is thus far no direct evidence for involvement of particular cell adhesion molecules in colonization of metastasis target tissues. The liver is exceptional in that it contains no basement membrane around the sinusoids where we have observed invasion of tumor cells to occur (6). Therefore, the only relevant interactions are those with sinusoidal endothelial cells and hepatocytes. These interactions can be studied in vitro using cultures of isolated liver cells (7, 8). For the present study, we used TA3/Ha murine mammary carci noma cells, which readily form liver métastasesupon intraportal in jection (6). These cells grow in suspension and do not attach to extracellular matrix components like fibronectin, vitronectin, col lagen, or laminin and do not interact with freshly isolated hepatic Received 12/14/92; accepted 5/26/93. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 1 Supported by Grant NKI 88-18 from the Dutch Cancer Society. 2 To whom requests for reprints should be addressed. 3 The abbreviations used are: VLA, very late activation antigen; LFA-1, lymphocyte function activated antigen 1; mAb. monoclonal antibody; SDS, sodium dodecyl sulfate; PAGE, polyacrylamide gel electrophoresis; PBS, phosphate-buffered saline; DMEM. Dul- becco's modified Eagle's medium; BSA, bovine serum albumin: Hepes, 4-(2-hydroxy- ethyl)-1 -piperazineethanesulfonic acid. sinusoidal endothelial cells,4 but they do adhere to hepatocytes. This indicates that adhesion to hepatocytes is particularly important for the formation of liver métastasesby these carcinoma cells. To identify the adhesion molecules involved in TA3-hepatocyte adhesion we have attempted to generate rat and hamster monoclonal antibodies against TA3 cells that inhibit TA3-hepatocyte adhesion, thus far without success. However, Fab fragments made from a poly clonal rabbit serum against the TA3 cells did inhibit this adhesion. Presently, we show that one of the relevant antigens is the integrin a6ß4. This result indicates that c*6ß4, which thus far has only been implicated in cell-matrix adhesion, can also mediate interactions with cell surfaces. Furthermore, it suggests that the high levels of a6ß4 often expressed by metastatic carcinoma cells (9-12) contribute to liver metastasis formation. MATERIALS AND METHODS Tumor Cells Mouse TA3/Ha mammary carcinoma cells were maintained in ascites in syngeneic A mice. Metastasis formation in vivo and adhesion to hepatocyte cultures in vitro by these cells have been described (6). Antibodies Rabbit antiserum 81435 directed against TA3 cells was obtained by immu nizing rabbits with 3 mg of TA3 plasma membrane protein in Freund's com plete adjuvant, followed by monthly booster injections in Freund's incomplete adjuvant. Rabbit antiserum 7880 directed against ß4wasobtained by immu nizing rabbits with purified denatured ß4 (see "Purification of the M, 195,000 Protein"). Rat mAb GoH3 directed against mouse a6 was a generous gift from Dr. Sonnenberg (15), and rat mAb 346-11A specific for the ß4 subunit of the mouse a6ß4integrin was a generous gift from Dr. Kennel (16). The hamster mAb C28 that binds the a6 subunit of the mouse a6ß4 integrin is produced by a hybridoma generated by fusion of mouse Sp2/0 cells with spleen cells of Armenian hamsters immunized with TA3 plasma membranes, using standard procedures (17). The immunization schedule was as follows. Hamsters were given s.c. injections of 400 fig TA3 plasma membrane protein in Freund's complete adjuvant, followed by two booster injections in Freund's incomplete adjuvant after 2 and 4 weeks. A final booster injection was given i.v. without adjuvant after 8 weeks. Four days later the fusion was performed. Electrophoresis and Immunoblotting Proteins were solubilized in Laemmli sample buffer, separated by reduced SDS-PAGE according to the method of Laemmli, and subsequently electro- phoretically transferred to nitrocellulose using the Tris-glycine buffer system. To visualize protein transfer, blots were reversibly stained with 0.4% Ponceau S in 3% trichloroacetic acid. All subsequent incubations were done on a rotary platform at 20°C.Blots were treated for l h in block buffer (1% low fat milk powder, 0.1% Triton X-100 in PBS) and then incubated with first antibodies in block buffer for 1 h. Polyclonal sera were used at a dilution of 1:1000, and affinity-purified antibodies were used at a concentration of 1 fig/ml. Next, biotin-conjugated second antibodies were applied, followed by streptavidin- alkaline phosphatase at a dilution of 1:1000 in block buffer. After each incu bation blots were washed three times with block buffer and also once with PBS before the color reaction. The alkaline phosphatase was detected by incubating the blot in substrate buffer (100 HIMTris, pH 9.5; 100 mm NaCl; and 5 mm 4 E. Roos et ai. unpublished observations. 3611 Research. on October 21, 2018. © 1993 American Association for Cancer cancerres.aacrjournals.org Downloaded from
Transcript of TheIntegrina6ß4onTA3 ...cancerres.aacrjournals.org/content/53/15/3611.full.pdf ·...

[CANCER RESEARCH 53. 3611-3617. August I. 1993]
The Integrin a6ß4on TA3/Ha Mammary Carcinoma Cells Is Involved in Adhesionto Hepatocytes1
Hans Kemperman, Yvonne Wijnands, Diana de Rijk, and Ed Roos2
Division of Cell Biology. The Netherlands Cancer Institute. Amsterdam, the Netherlands
ABSTRACT
lA.VIla mammary carcinoma cells form liver métastases upon in-
traportal injection. We have studied the interaction between these cellsand hepatocytes that is likely to be important for liver metastasis formation. We show that the integrin «,,/i4.which is highly expressed on TA3cells, is involved in this interaction. Fab fragments, generated from apolyclonal serum against TA3 cells, inhibited TA3-hepatocyte adhesion. Byaffinity purification on purified •¿�»,,)>.,.we isolated a6fJ4-specific Fab fragments from this anti-TA3 serum. These antibodies were highly spécifiefor
c*604, as demonstrated by Western blot analysis and immunoprecipitation,and inhibited TA3-hepatocyte adhesion. This shows that a6ß4,which thusfar has only been implicated in cell-matrix adhesion, can also mediate
interactions with cell surfaces. Our results suggest that the high levels of<*6ß4often expressed by metastatic carcinoma cells contribute to livermetastasis formation.
INTRODUCTION
Carcinomas, i.e., tumors originating from epithelial cells, constitute90% of human malignancies, and metastasis formation by these tumors is the major cause of therapeutic failure ( 1). For many carcinomas the liver is a major site of metastasis (2). A key step in themetastatic process is the adhesion of circulating tumor cells to cellsand extracellular matrix components in the target organ. Therefore itis likely that cellular adhesion molecules, which mediate such interactions, are major determinants of metastasis (3), and recently directevidence has been provided for this notion. Chan et al. (4) showed thatnonmetastatic rhabdomyosarcoma cells transfected with the copyDNA encoding the integrin VLA-23 (a2ßi),exhibited increased ad
hesion to collagen and laminin, and produced lung métastasesuponi.v. injection, whereas untransfected cells and control transfectants didnot. Furthermore, we have shown that mutants of a T-cell hybridomadeficient in expression of the integrin LFA-1 (aLß2)exhibited greatly
reduced metastatic potential (5). For carcinomas, however, there isthus far no direct evidence for involvement of particular cell adhesionmolecules in colonization of metastasis target tissues. The liver isexceptional in that it contains no basement membrane around thesinusoids where we have observed invasion of tumor cells to occur(6). Therefore, the only relevant interactions are those with sinusoidalendothelial cells and hepatocytes. These interactions can be studied invitro using cultures of isolated liver cells (7, 8).
For the present study, we used TA3/Ha murine mammary carcinoma cells, which readily form liver métastasesupon intraportal injection (6). These cells grow in suspension and do not attach toextracellular matrix components like fibronectin, vitronectin, collagen, or laminin and do not interact with freshly isolated hepatic
Received 12/14/92; accepted 5/26/93.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1Supported by Grant NKI 88-18 from the Dutch Cancer Society.2 To whom requests for reprints should be addressed.3 The abbreviations used are: VLA, very late activation antigen; LFA-1, lymphocyte
function activated antigen 1; mAb. monoclonal antibody; SDS, sodium dodecyl sulfate;PAGE, polyacrylamide gel electrophoresis; PBS, phosphate-buffered saline; DMEM. Dul-becco's modified Eagle's medium; BSA, bovine serum albumin: Hepes, 4-(2-hydroxy-
ethyl)-1 -piperazineethanesulfonic acid.
sinusoidal endothelial cells,4 but they do adhere to hepatocytes. This
indicates that adhesion to hepatocytes is particularly important for theformation of liver métastasesby these carcinoma cells.
To identify the adhesion molecules involved in TA3-hepatocyte
adhesion we have attempted to generate rat and hamster monoclonalantibodies against TA3 cells that inhibit TA3-hepatocyte adhesion,
thus far without success. However, Fab fragments made from a polyclonal rabbit serum against the TA3 cells did inhibit this adhesion.Presently, we show that one of the relevant antigens is the integrina6ß4.This result indicates that c*6ß4,which thus far has only beenimplicated in cell-matrix adhesion, can also mediate interactions with
cell surfaces. Furthermore, it suggests that the high levels of a6ß4often expressed by metastatic carcinoma cells (9-12) contribute to
liver metastasis formation.
MATERIALS AND METHODS
Tumor Cells
Mouse TA3/Ha mammary carcinoma cells were maintained in ascites insyngeneic A mice. Metastasis formation in vivo and adhesion to hepatocytecultures in vitro by these cells have been described (6).
Antibodies
Rabbit antiserum 81435 directed against TA3 cells was obtained by immunizing rabbits with 3 mg of TA3 plasma membrane protein in Freund's complete adjuvant, followed by monthly booster injections in Freund's incomplete
adjuvant. Rabbit antiserum 7880 directed against ß4was obtained by immunizing rabbits with purified denatured ß4(see "Purification of the M, 195,000Protein"). Rat mAb GoH3 directed against mouse a6 was a generous gift from
Dr. Sonnenberg (15), and rat mAb 346-11A specific for the ß4subunit of the
mouse a6ß4integrin was a generous gift from Dr. Kennel (16). The hamstermAb C28 that binds the a6 subunit of the mouse a6ß4integrin is produced bya hybridoma generated by fusion of mouse Sp2/0 cells with spleen cells ofArmenian hamsters immunized with TA3 plasma membranes, using standardprocedures (17). The immunization schedule was as follows. Hamsters weregiven s.c. injections of 400 fig TA3 plasma membrane protein in Freund'scomplete adjuvant, followed by two booster injections in Freund's incomplete
adjuvant after 2 and 4 weeks. A final booster injection was given i.v. withoutadjuvant after 8 weeks. Four days later the fusion was performed.
Electrophoresis and Immunoblotting
Proteins were solubilized in Laemmli sample buffer, separated by reducedSDS-PAGE according to the method of Laemmli, and subsequently electro-phoretically transferred to nitrocellulose using the Tris-glycine buffer system.
To visualize protein transfer, blots were reversibly stained with 0.4% PonceauS in 3% trichloroacetic acid. All subsequent incubations were done on a rotaryplatform at 20°C.Blots were treated for l h in block buffer (1% low fat milk
powder, 0.1% Triton X-100 in PBS) and then incubated with first antibodies in
block buffer for 1 h. Polyclonal sera were used at a dilution of 1:1000, andaffinity-purified antibodies were used at a concentration of 1 fig/ml. Next,biotin-conjugated second antibodies were applied, followed by streptavidin-
alkaline phosphatase at a dilution of 1:1000 in block buffer. After each incubation blots were washed three times with block buffer and also once with PBSbefore the color reaction. The alkaline phosphatase was detected by incubatingthe blot in substrate buffer (100 HIMTris, pH 9.5; 100 mm NaCl; and 5 mm
4 E. Roos et ai. unpublished observations.
3611
Research. on October 21, 2018. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from

ii„H4IN CARCINOMA CELL ADHESION TO HF.PATOCYTES
MgCI2) containing nilrohlue tetra/olium and 5-bromo-4-chloro-3-indolylphos-
phate (both from Pierce Chemical Co., Rockford. IL).
Generation of Fab Fragments
Immobilized papain (Pierce Chemical Co.) was used to prepare Fab frag
ments. Undigested IgG and Fc fragments were separated from Fab with proteinA-Sepharose. and the purity was checked by SDS-PAGE. Fab fragments were
dialyzed against PBS before they were tested in an adhesion assay.
Hepatocyte Isolation and Culture
Rat hepatocytes were isolated as described previously (18). Cells wereseeded in 96-well microtiter plates (Primaria; Falcon Labware. Oxnard. CA) at4 X IO4 cells/well or in 24-well plates (Primaria) at 2.5 X 10s cells/well in
DMEM (Flow Laboratories. Irvine, Ayrshire, Scotland) supplemented with 5/xg/ml bovine insulin (Sigma Chemical Co.. St. Louis, MO). 2 ITIMglutamine(Sigma). 20 mm Hepes. and 20 fig/liter dexamethasone. Before seeding, wellshad been coated with 10 tig/ml bovine fibronectin (Gibco BRL. Grand Island.NY). Two h after seeding, the hepatocytes were washed twice and culturedovernight in the same medium.
In Vitro Adhesion Assay
After overnight culture, the hepatocytes were washed twice with DMEMcontaining 20 HIMHepes. Adhesion of tumor cells to hepatocytes was quanti-tated using TA3 cells labeled with 5lCr (Amersham International. Amersham.United Kingdom) as described (19). Briefly, 5 x IO4 5lCr-labeled TA3 cells
were preincubated in 15 fil PBS with or without Fab fragments for 30 min at37°C.Subsequently, this 15-(xl sample was added to a microtiter well containing hepatocytes in 50 jul DMEM-Hepes and incubated for l h at 37°C.The
nonadherent cells were washed away, cultures were lysed in 100 jxl l s NaOH,and radioactivity was counted and used as a measure for the adhesion. Allmeasurements were performed in triplicate with at least two controls per plate.
In addition, adhesion was quantitated by light microscopy as describedpreviously (20). Briefly. 3 x IO5TA3 cells were incubated in 100 /j.1PBS withor without Fab fragments for 30 min at 37°C.Subsequently this 100-/¿Isample
was added to a hepatocyte culture in a 16-mm well containing 300 /xl DMEM-Hepes. The cultures were incubated at 37°Cfor l h and nonadherent cells were
washed away with PBS. Cultures were fixed with 2.5% glutaraldehyde, dehydrated, and scraped from the dish. The culture fragments were pelleted andembedded in Epon. 1-fxm sections were stained with \Vi toluidine in 1% borax
and viewed with a light microscope. The ratio of TA3 cells to hepatocyte nucleiwas determined and used as a measure for adhesion. At least 150 hepatocytenuclei were counted in 4 different sections per sample.
Affinity Purification of Anti-TA3 Antibodies on Octylglucoside-
solubilized TA3 Plasma Membrane Proteins
Plasma membranes from TA3 cells were isolated as described (21). Plasmamembranes (900 /xg protein) were lysed in 1.5 ml lysis buffer containing 30mM octylglucoside, 5 rriMMgCI2 and 40 mvi Tris supplemented with a proteaseinhibitor mixture (2 mm phenylmethylsulfonyl fluoride, 1 fig/ml soybeantrypsin inhibitor, and 2 /Ltg/mlaprotinin) for l h at 4°C.The lysate was cleared
by spinning at 14.000 X g for 10 min. The proteins in the lysate were allowedto bind to nitrocellulose (6.25 cm2) for l h at 4°Con a rotary platform. Empty
sites were blocked with block buffer. The blots were incubated with 900 figIgG from the anti-TA3 serum (81435) in block buffer for l h at 4°C,followed
by 3 washes in PBS. Bound antibodies were eluted with 0.1 Mglycine (pH 2.5)and immediately neutrali/ed with I MTris (pH 8) and dialyzed against PBS.
Affinity Purification of Anti-TA3 Antibodies on the Western-blotted M,
195,000 Protein
TA3 plasma membranes (3 mg protein) were lysed in octylglucoside lysisbuffer containing the protease inhibitor mixture for l h at 4°Cand centrifuged
at 2700 x g for 10 min. The proteins in the pellet, containing the M, 195.000protein, were solubilized in Laemmli sample buffer, separated by preparativeSDS-PAGE, and blotted to nitrocellulose. Proteins were visualized by staining
with Ponceau S, and the M, 195.000 protein band was excised and cut intosmall pieces. These nitrocellulose pieces were treated with block buffer for I
h at 4°C.then incubated with 900 /xg IgG from the anti-TA3 serum (81435) in
block buffer, and washed three times for 15 min with PBS. Bound antibodieswere eluted with 0.1 Mglycine (pH 2.5), immediately neutralized with 1 MTris,
and dialyzed against PBS.
Purification of the Mr 195,000 Protein
TA3 plasma membranes (3 mg of protein) were lysed in octylglucoside lysisbuffer containing the protease inhibitor mixture for l h at 4°Cand centrifuged
at 2700 X £for 10 min. The proteins in the pellet, containing the M, 195,000
protein, were solubilized in Laemmli sample buffer and separated on a preparative SDS-PAGE column, containing an 1-cm 39r stacking gel and a 2.5-cm
5% running gel. The collected fractions were analyzed by SDS-PAGE, andthose containing only the M, 195,000 protein were pooled and freeze-dried.Purity was again checked by SDS-PAGE and the protein concentration was
estimated based on Coomassie Brilliant Blue staining. Rabbits were immunized with approximately 50 fig of denatured M, 195,000 protein in Freund'scomplete adjuvant, followed by monthly booster injections in Freund's incom
plete adjuvant. After each boost, the sera were tested by Western blot analysis
and immunoprecipitation. The serum from rabbit 7880 was used for furtherstudy.
Affinity Purification of ahß4-.specificAntibodies from the Anti-TA3
Serum
On «„ßjPurified with the Anti Mr 195,000 Serum (7880). IgG (35 mg)from the anti-A/r 195,000 serum (7880) was coupled to 5 ml CNBr-Sepharose4B (Pharmacia) and packed into a column. TA3 cells (10s1) were lysed in 1%
Triton X-100, 0.5% deoxycholate, 0.1% SDS. 150 mm NaCl, 25 HIMTris, 5 min
MgCl2. and 1% BSA supplemented with the protease inhibitor mixture. Thelysate was cleared by spinning at 2700 x g for 10 min and applied to the
column. The ahß4complex bound so strongly to the column that it was noteluted by 0.1 Mglycine (pH 2.5), 0.1 Mtriethanolamine (pH 11.5). 3 M MgCI2,or 20% ethylene glycol. This enabled us to use the column directly to affinitypurify ov,ß4-specificantibodies from the anti-TA3 serum (81435). Fab frag
ments (60 mg). prepared from this serum, were applied to the column andbound Fab fragments were eluted with glycine (pH 2.5), neutralized with 1 MTris (pH 8), and dialyzed against PBS. The purity of the eluted Fab wasanalyzed by SDS-PAGE and specificity for a6ß4was confirmed by immuno-
blotting and immunoprecipitation. The same Fab fragments were analyzed for
binding to intact TA3 cells by FACScan analysis and tested in an adhesionassay.
On a,,ß.4Purified with the avspecific mAb GoH3. Eleven mg of thea()-specific mAb GoH3 were coupled to 2.5 ml of CNBr-Sepharose 4B (Pharmacia) and packed into a column. TA3 cells (4 X 10") were lysed in 1% Triton
X-100, 0.5% deoxycholate, 0.1% SDS, 150 min NaCl, 25 imi Tris, 5 IHM
MgCli, and 1% BSA supplemented with the protease inhibitor mixture. Thelysate was cleared by spinning at 2700 X # for 10 min and applied to thecolumn. The ott,ßjwas not eluted by 0.1 Mglycine (pH 2.5). 0.1 Mtriethanolamine (pH 11.5). 3 M MgCl2. or 20% ethylene glycol. Therefore, the columnwas used directly to affinity purify a6ß4specific antibodies from the anti-TA3
serum (81435). similar to the method described above.
Immunoprecipitation
TA3 cells (2 x 10'Yprecipitation) were surface labeled with '-*! (AmershamInternational) using the lactoperoxidase method or metabolically with [<5S]-
methionine (Amersham International). Cells were lysed in lysis buffer (1%Triton X-100, 0.5% deoxycholate, 0.1% SDS, 150mM NaCl, 25 ITIMTris, 5 mm
MgCl2, and 1% BSA) supplemented with the protease inhibitor mixture for 1h at 4°C.Lysates were cleared by spinning at 14,000 X g for 10 min. Rabbit
antibodies were precipitated with Staphylttcoccus aureus. Rat and hamstermonoclonal antibodies were first bound to rabbit anti-rat IgG antibodies andanti-hamster IgG antibodies, respectively (both from Nordic, Tilburg, the Neth
erlands). Two fji\ from polyclonal sera, 0.5 tig from affinity purified antibodies,and 100 til supernatant or 2 /j.1 ascites in the case of mAb were used perprecipitation. The immunoprecipitates were boiled in Laemmli sample bufferand analyzed by SDS-PAGE.
3612
Research. on October 21, 2018. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from

i IN CARCINOMA CELL ADHESION TO HEPATOCYTES
FACScan Analysis
FACScan analysis was performed as described (22). For analysis of intra-
cellular antigens, cells were permeabilized by incubating the cells in 90%methanol for 15 min at 4°C.Polyclonal sera were used at a dilution of 1:50, and
affinity purified antibodies were used at a concentration of 20 fig/ml. Allmeasurements were performed using a FACScan (Becton-Dickinson, Sunny
vale, CA).
RESULTS
Adhesion of TA3 Cells to Hepatocytes Is Inhibited by PolyclonalAnti-TA3 Antibodies. TA3 mammary carcinoma cells adhere to cul
tures of hepatocytes, the levels varying between 10 and 30% of addedTA3 cells in individual experiments. This level of adhesion may seemlow, but it should be noted that hepatocyte cultures are far fromconfluent, so that many of the added cells sediment onto the areasbetween the hepatocyte islands. Fab fragments prepared from a poly-
clonal serum against TA3 cells (81435) inhibited this adhesion, theextent of maximal inhibition varying from 40 to 70% between experiments. The results from one of three experiments are shown in Fig.\A (81435). Similar inhibition was seen when TA3 cells were prein-
cubated with Fab fragments and washed before adding the TA3 cellsto the hepatocyte cultures (results not shown). Thus, the anti-TA3 Fab
affected the TA3 cells and not the hepatocytes. We never saw anyeffect of normal rabbit Fab fragments.
Identification of a Mr 195,000 Protein on TA3 Cells PotentiallyInvolved in TA3-Hepatocyte Adhesion. Our strategy to identify therelevant TA3 antigen was to affinity purify Fab fragments on separated and blotted membrane proteins and test eluted Fab for inhibitionof adhesion. As a first step, we used total membrane protein. TA3plasma membranes were lysed with octylglucoside, and the solubi-lized proteins were immobilized on nitrocellulose. Anti-TA3 Fab frag
ments were affinity purified on these blots and tested in an adhesion
assay but unexpectedly did not inhibit TA3-hepatocyte adhesion
(Fig. \A, 81435,sup). Western blot analysis showed that the elutedFab fragments reacted with the same TA3 plasma membrane proteinsas the original anti-TA3 serum with the exception of a A/r 195,000
protein that was not solubilized by octylglucoside (Fig. Iß).Thissuggested that the Mr 195,000 protein was the relevant antigen.
To demonstrate this, we purified Fab fragments specific for thisMr 195,000 protein. Hereto, TA3 plasma membrane proteins thatwere not solubilized by octylglucoside, were separated by SDS-
PAGE and blotted to nitrocellulose. The band containing the Mr195,000 protein was excised and Fab fragments were affinity purified on this protein. These Fab fragments were highly specific forthe Mr 195,000 protein as shown by Western blot analysis (Fig. Iß,81435,195 kD) and inhibited TA3-hepatocyte adhesion. Maximal in
hibition varied between 30 and 60%. The results from a representative experiment, of three, are shown in Fig. \A (81435,195 kD). As acontrol, the same procedure was applied to a major A/r 57,000 TA3plasma membrane protein, but the resulting Fab fragments, althoughhighly specific, did not inhibit TA3-hepatocyte adhesion (Fig. \A).
This strongly suggested that the Mr 195,000 protein was the relevantantigen.
Purification of the Mr 195,000 Protein and Production of aSpecific Antiserum. To obtain additional evidence for the involvement of the Mr 195,000 protein in TA3-hepatocyte adhesion, wepurified the Mr 195,000 protein on a preparative SDS-PAGE column.Purity was checked by SDS-PAGE, and rabbits were immunized with
the purified denatured protein. The obtained polyclonal rabbit serum(7880) specifically stained the Mr 195,000 protein on a Western blotof TA3 plasma membrane proteins (Fig. 2A). However, antibodiesisolated from this serum did not inhibit TA3-hepatocyte adhesion.
FACScan analysis showed that the antibodies did not react with intactcells (Fig. 3A), indicating that the serum was directed against intra-
% inhibition of adhesionB 81435 81435,sup 81435,195 kD
-210 kD
81435.57 kD
antibody
-111
- 68
- 45
P a1
P »2
Fig. I. A, effect of antibodies on the adhesion of TA3 cells to hepatocyte cultures. *' Cr-labeled TA3 cells were preincubated for 30 min with Fab fragments prepared from the anti-TA3serum (8Ì435:300 /¿g/ml).Fab that had been affinity-purified on octylglucoside-solubilized TA3 plasma membrane proteins (81435,sup; 300 /ig/ml). Fab affinity purified onWestern-blotted M, 195.000 protein (81435.195 kD; 10 /ig/ml), and Fab affinity purified on Western-blotted M, 57,000 protein (81435,57 kD; 10 ng/ml). Cells were then allowed to
adhere to hepatocyte cultures for 1 h, in the presence of the Fab fragments, and the nonadherent cells were washed away. Cultures were lysed and counted in a gamma counter. Eachcolumn is the average of three separate wells. Shown is the percentage of inhibition compared to control values {cells incubated with PBS only) of one experiment of three with similarresults. Bars. SD. ß.Western blot of TA3 plasma membrane proteins probed with the same antibodies as in A. Plasma membranes were lysed in octylglucoside buffer and centrifugedat 2700 X g for 10 min. Proteins in the pellet (p) and in the supernatant (s) were separated by reduced 10% SDS-PAGE, electrophoretically transferred to nitrocellulose and probedwith anti-TA3 antibodies (81435). antibodies affinity purified on octylglucoside-solubilized TA3 plasma membrane proteins (81435,sup), and antibodies affinity purified on Western
blotted M, 195.000 protein (81435,195 kD).
3613
Research. on October 21, 2018. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from

a„B4IN CARCINOMA CELL ADHESION TO HEPATOCYTES
A 7880 B
Fig. 2. Characterization of the anti-A/r 195,000serum (7880). A, Western blot of TA3 plasma membrane proteins separated by reduced 7% SDS-PAGE,electrophoretically transferred to nitrocellulose, andprobed with the anti-Air 195,000 serum (7880). B,antigens precipitated from lysates of I25l-surface-labeled TA3 cells by anti-TA3 serum (81435), ami-
Ai, 195,000 serum (7880). a mAb specific for ß4(346-11 A), or mAb specific for a6 (GoH3 and C28),
and normal rabbit serum (NRS). Precipitates wereanalyzed by reduced 7.5% SDS-PAGE. C, immuno-
precipitation using the same cells and antibodies asin B but after preclearance with the (»„-specificmAbGoH3.
-aos
-116
- 80
- 49
:-205 kD
-116
- 80
- 49
preclear GoH3
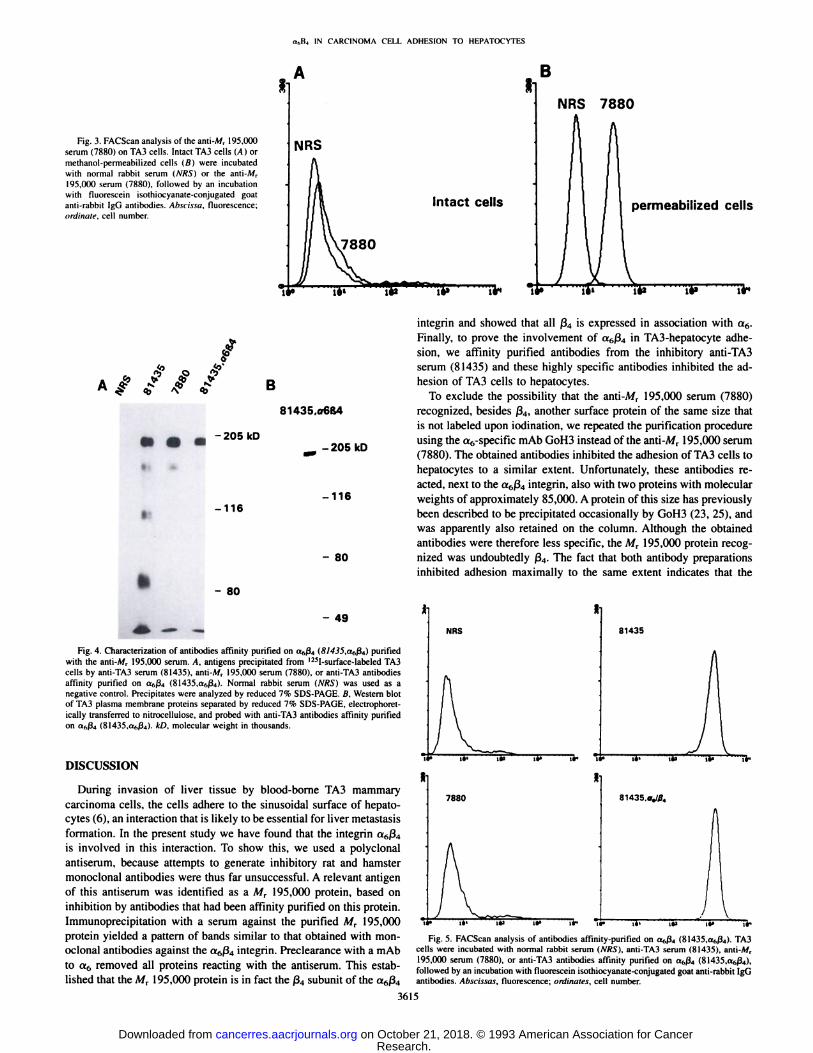
cellular epitopes. This was in fact the case since the antibodies didreact with permeabilized TA3 cells as shown by FACScan analysis(Fig. 3B).
Identification of the V/, 195,000 Protein as the Integrin ß4Subunit. Immunoprecipitation with the anti-A/r 195,000 serum(7880), using I25l-surface-labeled TA3 cells, yielded a pattern of
bands that was similar to that observed with antibodies against the a6or ß4subunit of the a6ß4integrin (Fig. IB): on reduced SDS-PAGE,
a major band with a molecular weight of 200,000 which is the matureß4subunit, two minor bands at Mr 180,000 and 150,000 which areproteolytic products of ß4,and a faint band with a molecular weightof 115,000 which is the a6 subunit (23). The band containing a6 isonly visible after longer exposure of the film due to the poor iodina-tion of a6 (23). On nonreduced SDS-PAGE the largest band ran at
almost the same position (results not shown), as has been described byothers for ß4(23). After preclearing of a TA3 cell lysate with thea6-specific mAb (GoH3), the anti-Mr 195,000 serum (7880) no longer
precipitated any protein (Fig. 2C), showing that the M, 195,000 protein is in fact the ß4subunit of the c*6ß4integrin. Even after very longexposure, no precipitated proteins were seen, showing that ß4is theonly iodinatable cell surface protein reacting with the anti-Mr 195,000
serum. Since the serum reacts only with intracellular epitopes (Fig. 3),it is apparently directed against the very long cytoplasmic domain ofß4(24).
Affinity Purification of <*6ß4-specificAntibodies from the Anti-TA3 Serum. To obtain inhibitory a6ß4-specificantibodies, we decided to purify a6ß4-specific antibodies from the anti-TA3 serum
(81435) on the a6ß4complex. Hereto, we attempted to affinity purifythe native a6ß4integrin on a Sepharose-coupled anti-Mr 195,000 IgG
(7880) column. However, a6ß4bound so strongly to the column thatit could not be eluted by low or high pH, high salt, or ethylene glycol.This enabled us to purify a6ß4-specificFab fragments prepared fromthe anti-TA3 serum directly on the a6ß4bound to the column. Anti-
TA3 Fab fragments (60 mg) were loaded on the column and 40 ¿xga6ß4-specificFab fragments were eluted. The eluted Fab fragments
immunoprecipitated the a6ß4integrin (Fig. 4A ) and were highly specific for ß4as demonstrated by Western blot analysis (Fig. 4ß).Theadditional faint band with a molecular weight of approximately150,000 is most likely a proteolytic product of ß4,since the intensity
of this band is enhanced after incubation of the lysate at 37°Cas
described by Hemler et al. (23). The Fab fragments also react withintact cells as shown by FACScan analysis (Fig. 5, 81435,a6ß4).
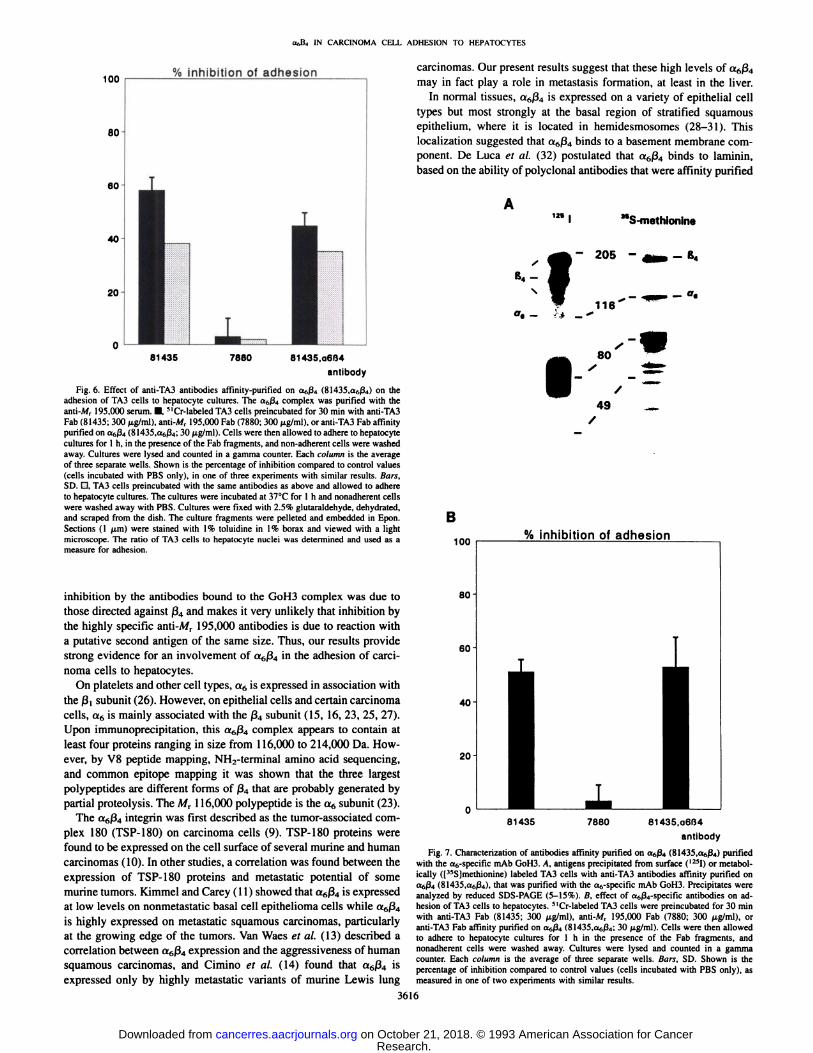
The same a6ß4-specificantibodies were tested in an adhesion assay. Adhesion, as quantitated with 51Cr-labeled TA3 cells, was inhib
ited maximally to an extent varying between 30 and 50%. The resultsfrom one of three experiments are shown in Fig. 6 (black bars). Thiswas confirmed by direct counting of adherent TA3 cells in sections ofembedded hepatocyte cultures. Again, maximal inhibition was approximately 40% (Fig. 6, stippled bars).
To exclude the possibility that the anti-Mr 195,000 serum also reacts
with another noniodinatable cell surface protein of exactly the samesize, we repeated the procedure using the a6-specific mAb GoH3.a6ß4could also not be eluted from the GoH3 column, so that «6ß4-specific Fab fragments from the anti-TA3 serum again could be purified directly on the column. Anti-TA3 Fab fragments (55 mg) wereloaded on the column and 200 ¿tga6ß4-specificFab fragments were
eluted. The eluted Fab fragments precipitated not only the a6ß4integrin but in addition also two proteins with molecular weight of approximately 85,000 from both surface and metabolically labeled TA3cell lysates (Fig. 7A). Also others described that the a6-specific mAb
GoH3 sometimes precipitates proteins with molecular weights of approximately 85,000 (23, 25). These proteins were quite likely retainedon the GoH3 column, so that anti-Mr 85,000 antibodies were alsoenriched from the anti-TA3 serum. The antibodies obtained were
therefore less specific, but the Mr 195,000 band recognized is undoubtedly ß4.
The eluted Fab fragments again inhibited TA3-hepatocyte adhesion
to a similar extent, approximately 50%. The results of one of twoexperiments is shown in Fig. IB. Taken together, the results with bothantibody preparations provide strong evidence for the involvement ofO6ß4in the adhesion of TA3 carcinoma cells to hepatocytes.
TA3-hepatocyte adhesion is not inhibited by the a6-specific mAbGoH3 and C28. Also mAb 346-11A directed against murine ß4did not
inhibit this interaction, indicating that their epitopes are not located inthe vicinity of the binding site for the hepatocyte ligand.5
5 H. Kemperman et al., unpublished observations.
3614
Research. on October 21, 2018. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from
t.,,IÌ4IN CARCINOMA CELL ADHESION TO HEPATOCYTES
Fig. 3. FACScan analysis of the anti-Mr 195.ÃœOO
serum (7880) on TA3 cells. Intact TA3 cells (A) ormethanol-permeabilized cells (ß)were incubatedwith normal rabbit serum (A'/f.S")or the anti-A/r
195.000 serum (7880). followed by an incubationwith fluorescein ¡sothiocyanate-conjugated goatanti-rabbit IgG antibodies. Abscissa, fluorescence;ordinate, cell number.
té»
NRS
7880
B
Intact cells
ir*
NRS 7880
permeabilized cells
le«
f* / f /
81435,
-205kD
-116
- 80
-205 kD
-116
- 80
- 49
Fig. 4. Characterization of antibodies affinity purified on a,,ß4(8/4J5,a6ß4) purifiedwith the anti-Mr 195,000 serum. A, antigens precipitated from I25l-surface-labeled TA3cells by anti-TA3 serum (81435). anti-Mr 195,000 serum (7880). or anti-TA3 antibodies
affinity purified on a,,ß4(81435.afiß4). Normal rabbit serum (NRS) was used as anegative control. Precipitates were analyzed by reduced 7% SDS-PAGE. B, Western blotof TA3 plasma membrane proteins separated by reduced 7% SDS-PAGE, electrophoret-ically transferred to nitrocellulose, and probed with anti-TA3 antibodies affinity purifiedon a^ßa(81435,a,,ß4). kD, molecular weight in thousands.
DISCUSSION
During invasion of liver tissue by blood-borne TA3 mammarycarcinoma cells, the cells adhere to the sinusoidal surface of hepato-
cytes (6), an interaction that is likely to be essential for liver metastasisformation. In the present study we have found that the integrin a6ß4is involved in this interaction. To show this, we used a polyclonalantiserum, because attempts to generate inhibitory rat and hamstermonoclonal antibodies were thus far unsuccessful. A relevant antigenof this antiserum was identified as a M, 195,000 protein, based oninhibition by antibodies that had been affinity purified on this protein.Immunoprecipitation with a serum against the purified Mr 195,000protein yielded a pattern of bands similar to that obtained with monoclonal antibodies against the a6ß4integrin. Preclearance with a mAbto a6 removed all proteins reacting with the antiserum. This established that the MT 195,000 protein is in fact the ß4subunit of the a6ß4
integrin and showed that all ß4is expressed in association with a6.Finally, to prove the involvement of a6ß4in TA3-hepatocyte adhesion, we affinity purified antibodies from the inhibitory anti-TA3
serum (81435) and these highly specific antibodies inhibited the adhesion of TA3 cells to hepatocytes.
To exclude the possibility that the anti-Mr 195,000 serum (7880)
recognized, besides ß4,another surface protein of the same size thatis not labeled upon iodination, we repeated the purification procedureusing the a6-specific mAb GoH3 instead of the anti-Mr 195,000 serum
(7880). The obtained antibodies inhibited the adhesion of TA3 cells tohepatocytes to a similar extent. Unfortunately, these antibodies reacted, next to the a6ß4integrin, also with two proteins with molecularweights of approximately 85,000. A protein of this size has previouslybeen described to be precipitated occasionally by GoH3 (23, 25), andwas apparently also retained on the column. Although the obtainedantibodies were therefore less specific, the Mr 195,000 protein recognized was undoubtedly ß4.The fact that both antibody preparationsinhibited adhesion maximally to the same extent indicates that the
I
NRS 81435
7880 81435. a,l04
Fig. 5. FACScan analysis of antibodies affinity-purified on aftß4(81435,a^ßj). TA3cells were incubated with normal rabbit serum (NRS). anti-TA3 serum (81435), anti-A/r195,000 serum (7880), or anti-TA3 antibodies affinity purified on a„ß4(81435,a,,ß4),followed by an incubation with fluorescein isothiocyanate-conjugated goat anti-rabbit IgGantibodies. Abscissas, fluorescence; ordinales, cell number.
3615
Research. on October 21, 2018. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from
IN CARCINOMA CELL ADHESION TO HEPATOCYTES
100
80
% inhibition of adhesion
81435 7880 81435,o6ß4antibody
Fig. 6. Effect of anti-TA3 antibodies affinity-purified on aftß4(8l435,af,ß4) on theadhesion of TA3 cells to hepatocyte cultures. The a6ß4complex was purified with theanti-M, 195,000 serum. •¿�''Cr-labeled TA3 cells preincubated for 30 min with anti-TA3Fab (81435; 300 ng/ml). anti-Mr 195,000 Fab (7880; 300 ng/ml), or anti-TA3 Fab affinitypurified on aftß4(81435,anß4;30 ng/ml). Cells were then allowed to adhere to hepatocytecultures for 1 h, in the presence of the Fab fragments, and non-adherent cells were washedaway. Cultures were lysed and counted in a gamma counter. Each column is the averageof three separate wells. Shown is the percentage of inhibition compared to control values(cells incubated with PBS only), in one of three experiments with similar results. Bars,SD. D, TA3 cells preincubated with the same antibodies as above and allowed to adhereto hepatocyte cultures. The cultures were incubated at 37°Cfor l h and nonadherent cells
were washed away with PBS. Cultures were fixed with 2.5% glutaraldehyde, dehydrated,and scraped from the dish. The culture fragments were pelleted and embedded in Epon.Sections (1 (¿m)were stained with 1% toluidine in 1% borax and viewed with a lightmicroscope. The ratio of TA3 cells to hepatocyte nuclei was determined and used as ameasure for adhesion.
inhibition by the antibodies bound to the GoH3 complex was due tothose directed against ß4and makes it very unlikely that inhibition bythe highly specific anti-A/r 195,000 antibodies is due to reaction with
a putative second antigen of the same size. Thus, our results providestrong evidence for an involvement of afiß4in the adhesion of carcinoma cells to hepatocytes.
On platelets and other cell types, a6 is expressed in association withthe ß,subunit (26). However, on epithelial cells and certain carcinomacells, a6 is mainly associated with the ß4subunit (15, 16, 23, 25, 27).Upon immunoprecipitation, this cv6ß4complex appears to contain atleast four proteins ranging in size from 116,000 to 214,000 Da. However, by V8 peptide mapping, NH2-terminal amino acid sequencing,
and common epitope mapping it was shown that the three largestpolypeptides are different forms of ß4that are probably generated bypartial proteolysis. The Mr 116,000 polypeptide is the a6 subunit (23).
The a6ß4integrin was first described as the tumor-associated complex 180 (TSP-180) on carcinoma cells (9). TSP-180 proteins were
found to be expressed on the cell surface of several murine and humancarcinomas (10). In other studies, a correlation was found between theexpression of TSP-180 proteins and metastatic potential of some
murine tumors. Kimmel and Carey (11) showed that a6ß4is expressedat low levels on nonmetastatic basal cell epithelioma cells while a6ß4is highly expressed on metastatic squamous carcinomas, particularlyat the growing edge of the tumors. Van Waes et al. (13) described acorrelation between a6ß4expression and the aggressiveness of humansquamous carcinomas, and Cimino et al. (14) found that a6ß4isexpressed only by highly metastatic variants of murine Lewis lung
carcinomas. Our present results suggest that these high levels of <*6ß4may in fact play a role in metastasis formation, at least in the liver.
In normal tissues, a6ß4is expressed on a variety of epithelial celltypes but most strongly at the basal region of stratified squamousepithelium, where it is located in hemidesmosomes (28-31). This
localization suggested that a6ß4binds to a basement membrane component. De Luca et al. (32) postulated that a6ß4binds to laminin,based on the ability of polyclonal antibodies that were affinity purified
1211 "S-methionine
j- 205 -__-B4
B
100
80
60-
40-
20
% inhibition of adhesion
81435 7880 81435,06(34antibody
Fig. 7. Characterization of antibodies affinity purified on a<,ß4(81435,a6ß4) purifiedwith the a6-specific mAb GoH3. A, antigens precipitated from surface (I25I) or metabol-ically ([15S]methionine) labeled TA3 cells with anti-TA3 antibodies affinity purified on<«(,/'i(81435,Of6ß4),that was purified with the a6-specific mAb GoH3. Precipitates wereanalyzed by reduced SDS-PAGE (5-15%). B, effect of a6ß4-specific antibodies on adhesion of TA3 cells to hepatocytes. 5lCr-labeled TA3 cells were preincubated for 30 minwith anti-TA3 Fab (81435; 300 fig/ml), anti-A/r 195,000 Fab (7880; 300 M-gAiil), oranti-TA3 Fab affinity purified on a6ß4(81435.a6ß4; 30 fig/ml). Cells were then allowedto adhere to hepatocyte cultures for l h in the presence of the Fab fragments, andnonadherent cells were washed away. Cultures were lysed and counted in a gammacounter. Each column is the average of three separate wells. Bars, SD. Shown ¡sthepercentage of inhibition compared to control values (cells incubated with PBS only), asmeasured in one of two experiments with similar results.
3616
Research. on October 21, 2018. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from

a«B4 IN CARCINOMA CELL ADHESION TO HEPATOCYTES
on a6ß4,to detach keratinocytes from Matrigel or purified laminin.Also Lotz et al. (33) suggested this, on the basis of the inhibition ofcarcinoma cell adhesion to laminin by an anti-a6 mAb. However, this
laminin receptor function for a6ß4is not widely accepted, becauseother groups were unable to inhibit adhesion to laminin of cellsexpressing a6ß4but not a6ßi,with monoclonal antibodies to a6 or ß4.Furthermore, attempts to purify a6ß4on a laminin column wereunsuccessful (31, 33-36). Recently, however, Lee et al. (37) showed
that cells expressing a6 only in association with ß4bind to the lamininE8 fragment and that this binding can be blocked by both a mAb to ß4(30%) and a mAb to a6 (60%). Moreover, these authors did succeedin purification of a6ß4on a laminin column, strongly suggesting thatlaminin is in fact a ligand of a6ß4.
It is unlikely, however, that a6ß4on TA3 cells binds to hepatocytesvia laminin, because TA3 cells do not adhere to purified laminincoated on plastic or glass and because we found that cultured hepatocytes produced little, if any, laminin.6 Moreover, the a6-specific
mAb GoH3, which completely blocks a6ß,binding to laminin (38)and partly the binding of a6ß4to laminin (37), did not inhibit TA3-
hepatocyte adhesion. This suggests that «6ß4binds to a differentligand on the hepatocyte surface via a distinct binding domain that isnot affected by GoH3. This notion is not without precedent, becausethe binding of the integrin VLA-4 (a4ß,)to its matrix ligand fibronec-tin and its cellular ligand VCAM-1 occurs via binding domains that
are blocked by distinct antibodies (39). A candidate ligand on hepatocytes is an antigen recognized by the OPAR mAb, directed againstrat hepatocytes, that inhibits adhesion of TA3 cells (19). Preliminaryresults indicate that this antigen is secreted by hepatocytes and istherefore a cell surface-associated rather than a transmembrane
protein.
ACKNOWLEDGMENTS
We are grateful to T. Hamers for performing immunizations and collectingsera from rabbits, to A. J. Schrauwers for biotechnical assistance, to E. Note-
boom for assistance with FACScan analysis, to N. Ong and J. Lomecky forpreparing photographs, and to C. van Niele-Pouw for secretarial assistance. We
further wish to thank Dr. A. Sonnenberg and Dr. S. J. Kennel for their generousgift of antibodies.
REFERENCES
1. Fidler, I. J., and Balch, C. M. The biology of cancer metastasis and implications fortherapy. Cum Probi. Surg., 24: 137-209, 1987.
2. Bross, I. D., Viadana, E., and Pickren, J. W. The metastatic spread and myeloma andleukemias in men. Virchows Arch., 365: 91-101, 1975.
3. Roos, E. Adhesion molecules in lymphoma metastasis. Cancer Metastasis, 10: 33^8,1991.
4. Chan, B. M. C., Matsuura, N., Takada, Y., Zetter, B. R., and Hemler, M. E. In vitroand in vivo consequences of VLA-2 expression on rhabdomyosarcoma cells. Science(Washington DC), 251: 1600-1602, 1991.
5. Roossien, F. F., de Rijk, D., Bikker, A., and Roos, E. Involvement of LFA-1 inlymphoma invasion and metastasis demonstrated with LFA-1-deficient mutants. J.Cell Biol., 108: 1979-1985, 1989.
6. Roos, E., Dingemans, K. P., van de Pavert, I. V., and van den Berg Weerman.Mammary carcinoma cells in the mouse liver: infiltration of liver tissue and interaction with Kupffer cells. Br. J. Cancer, 38: 88-99, 1978.
7. Roos, E., Middelkoop, O. P., and van de Pavert, I. V. Adhesion of tumor cells tohepatocytes: different mechanisms for mammary carcinoma compared with lympho-sarcoma cells. J. Nati. Cancer Inst., 73: 963-969, 1984.
8. Roos, E., Tulp, A., Middelkoop, O. P., and van de Pavert, I. V. Interactions betweenlymphoid tumor cells and isolated liver endothelial cells. J. Nati. Cancer Inst., 72:1173-1180, 1984.
9. Falcioni, R., Kennel, S. J., Oiacomini, P., Zupi, G., and Sacchi, A. Expression oftumor antigen correlated with metastatic potential of Lewis lung carcinoma and B16melanoma clones in mice. Cancer Res., 46: 5772-5778, 1986.
10. Falcioni, R., Sacchi, A., Resau, J., and Kennel, S. J. Monoclonal antibody to humancarcinoma-associated protein complex: quantitation in normal and tumor tissue. Cancer Res., 48: 816-821, 1988.
12.
13.
14.
16.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
s G. La Rivièreet ai, unpublished observations.
Kimmel, K. A., and Carey, T. E. Altered expression in squamous carcinoma cells ofan orientation restricted epithelial antigen detected by monoclonal antibody A9.Cancer Res., 46: 3614-3623. 1986.
Costantini, R. M., Falcioni, R., Battista, P., Zupi. G., Kennel, S. J., Colasante, A.,Venturo, I., Curcio, C. G., Sacchi, A. Integrin (aftß4)expression in human lung canceras monitored by specific monoclonal antibodies. Cancer Res., 50: 6107-6112. 1990.Van Waes, C., Kozarsky, K. F., Warren, A. B., Kidd, L., Paugh, D., Lieben, M., andCarey, T. E. The A9 antigen associated with aggressive human squamous carcinomais structurally and functionally similar to the newly defined integrin a^ß^.CancerRes., 5/: 2395-2402, 1991.
Cimino, L., Perrotti, D., Falcioni, R., Kennel, S. J., and Sacchi, A. ß4integrinexpression on in vitro and in vivo metastatic variants of Lewis lung carcinoma.Cytotechn., J: S45-S48, 1991.
Sonnenberg. A., Hogervorst, F., Osterop, A., and Veltman, F. E. M. Identification andcharacterization of a novel antigen complex on mouse mammary tumor cells using amonoclonal antibody against platelet glycoprotein Lc. J. Biol. Chem., 263: 14030-
14038, 1988.Kennel, S. J., Foote, L. J., Falcioni, R., Sonnenberg, A., Stringer, C. D., Crouse, C.,and Hemler, M. E. Analysis of the tumor-associated antigen TSP-180. Identity witha6(34 in the integrin superfamily. J. Biol. Chem., 264: 15515-15521, 1989.
Bright, S. W., Chen, T., Flebbe, L. M., Lei, M., and Morrison, D. C. Generation andcharacterization of hamster-mouse hybridomas secreting monoclonal antibodies withspecificity for lipopolysaccharide receptor. J. Immunol., 745: 1-7, 1990.Roos, E., and van de Pavert, I. V. Effect of tubulin-binding agents on infiltration oftumor cells into primary hepatocyte cultures. J. Cell Sci., 55: 223-245, 1982.Middelkoop, O. P., van Bavel, P., Calafat, J., and Roos, E. Hepatocyte surfacemolecule involved in the adhesion of TA3 mammary carcinoma cells to rat hepatocytecultures. Cancer Res., 45: 3825-3835, 1985.
Roos, E., van de Pavert, I. V., and Middelkoop, O. P. Infiltration of tumor cells intocultures of isolated hepatocytes. J. Cell Sci., 47: 385-397, 1981.Record, M., Bes, J-C., Chap. H., and Douste-Blazy, L. Isolation and characterization
of plasma membranes from Krebs II ascites cells using Percoli gradient. Biochim.Biophys. Acta, 688: 57-65, 1982.
La Rivière,G., Schipper, C. A., Collard, J. G., and Roos, E. Invasiveness in hepatocyte and fibroblast monolayers and metastatic potential of T-cell hybridomas in mice.Cancer Res., 48: 3405-3410, 1988.
Hemler, M. E., Crouse, C., and Sonnenberg, A. Association of the VLA Q6 subunitwith a novel protein. A possible alternative to the common VLA ß,subunit on certaincell lines. J. Biol. Chem., 264: 6529-6535, 1989.
Hogervorst, F., Kuikman, I., von dem Borne, A. E. G. Kr., and Sonnenberg, A.Cloning and sequence analysis of beta-4cDNA: an integrin subunit that contains aunique 118 kD cytoplasmic domain. EMBO J., 9: 765-770, 1990.Sonnenberg, A., Linders, C. J. T., Daams, J. H., and Kennel, S. K. The afiß,VLA-6)and a6ß4complexes: tissue distribution and biochemical properties. J. Cell Sci., 96:207-217, 1990.
Hemler, M. E., Crouse, C., Takada, Y., and Sonnenberg, A. Multiple very late antigen(VLA) heterodimers on platelets. J. Biol. Chem., 263: 7660-7665, 1988.Kajiji, S., Tamura, R. N., and Quaranta, V. A novel integrin (aEß4)from humanepithelial cells suggests a fourth family of integrin adhesion receptors. EMBO J., 8:673-680, 1989.Stepp, M. A., Spurr-Michaud, S., Tisdale, A., Elwell, J., and Gipson, I. K. a6ß4
integrin heterodimer is a component of hemidesmosomes. Proc. Nati. Acad. Sci. USA,87: 8970-8974, 1990.
Carter, W. G., Kaur, P., Gill, S. G., Gahr, P. J., and Wayner, E. A. Distinct functionsfor integrins in focal adhesions and Qöß^ullouspemphigoid antigen in a new stableanchoring contact (SAC) of keratinocytes: relation to hemidesmosomes. J. Cell Biol.,;;/: 3141-3154, 1991.
Jones. J. C. R., Kurpakus, M. A., Cooper, H. M., and Quaranta, V. A function for theintegrin a6ß„in the hemidesmosome. Cell Regul., 2: 427-438, 1991.Sonnenberg, A., Calafat, J., Janssen, H., Daams, H., van der Raaij-Hemler, L. M. H.,Falcioni, R., Kennel, S. J., Aplin, J. D., Baker, J., Loizidou, M., and Garrod, D.Integrin o6ß4complex is located in hemidesmosomes, suggesting a major role inepidermal cell-basement membrane adhesion. J. Cell Biol.. 113: 907-917, 1991.De Luca, M., Tamura, R. N., Kajiji, S., Bondanza, S., Rossino, P.. Cancedda, R.,Marchisio, P. C., and Quaranta, V. Polarized integrin mediates human keratinocyteadhesion to basal lamina. Proc. Nati. Acad. Sci. USA, 87: 6888-6892. 1990.Lotz, M. M., Korzelius, C. A., and Mercurio, A. M. Human colon carcinoma cells usemultiple receptors to adhere to laminin: involvement of a6ß4and a2ß,integrins. CellRegul., /: 249-257, 1990.
Sonnenberg, A., Linders, C. J. T., Modderman, P. W., Damsky, C. H., Aumailley, M.,and Timpl., R. Integrin recognition of different cell-binding fragments of laminin (PI,E8) and evidence that a6ßibut not aöß4functions as a major receptor for fragmentE8. J. Cell Biol., 110: 2145-2156, 1990.
Quaranta, V., and Jones, J. C. R. The internal affairs of an integrin. Trends Cell Biol.,/: 2-4, 1991.
Carter, W. G., Ryan, M. C., and Gahr, P. J. Epiligrin, a new cell adhesion ligand forintegrin a3ß,in epithelial basement membranes. Cell, 65: 599-610, 1991.Lee, E. C., Lotz, M. M., Steele, G. D., Jr., and Mercurio, A. M. The integrin a6ß4¡sa laminin receptor. J. Cell Biol., 117: 671-678, 1992.Sonnenberg, A., Modderman, P. W., and Hogervorst, F. Laminin receptor on plateletsis the integrin VLA-6. Nature (Lond.), 336: 487^189, 1988.Elices, M. J., Osborn, L., Takada, Y, Crouse, C., Luhowskyj, S., Hemler, M. E., andLobb, R. R. VCAM-1 on activated endothelium interacts with the leukocyte integrinVLA-4 at a site distinct from the VLA-4/fibronectin binding site. Cell, 60: 577-584,
1990.
3617
Research. on October 21, 2018. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from

1993;53:3611-3617. Cancer Res Hans Kemperman, Yvonne Wijnands, Diana de Rijk, et al. Involved in Adhesion to Hepatocytes
on TA3/Ha Mammary Carcinoma Cells Is4β6αThe Integrin
Updated version
http://cancerres.aacrjournals.org/content/53/15/3611
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/53/15/3611To request permission to re-use all or part of this article, use this link
Research. on October 21, 2018. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from









![StatisticalAnalysisofHormonalEffectsontheSteroid ...cancerres.aacrjournals.org/content/30/1/221.full.pdf[CANCERRESEARCH30,221-227,January1970]. StatisticalAnalysisofHormonalEffectsontheSteroid](https://static.fdocuments.us/doc/165x107/5ab039c17f8b9a25088e7528/statisticalanalysisofhormonaleffectsonthesteroid-cancerresearch30221-227january1970.jpg)

![InVitroHematopoiesisfollowingInductionChemotherapyforAcute ...cancerres.aacrjournals.org/content/45/11_Part_2/5921.full.pdf[CANCERRESEARCH45,5921-5925,November1985] InVitroHematopoiesisfollowingInductionChemotherapyforAcuteLeukemia1](https://static.fdocuments.us/doc/165x107/5b0a316b7f8b9a45518be441/invitrohematopoiesisfollowinginductionchemotherapyforacute-cancerresearch455921-5925november1985.jpg)


![ClinicalValueofSerumGlycoproteinGalactosyltransferaseLevel ...cancerres.aacrjournals.org/content/43/9/4491.full.pdf · [CANCERRESEARCH43,4491-4496,September1983] ClinicalValueofSerumGlycoproteinGalactosyltransferaseLevelsin](https://static.fdocuments.us/doc/165x107/5ac139b17f8b9ac6688d1490/clinicalvalueofserumglycoproteingalactosyltransferaselevel-cancerresearch434491-4496september1983.jpg)

![HeterotransplantationofHumanLymphoidNeoplasmsUsingaNudeMou ...cancerres.aacrjournals.org/content/50/10/3078.full.pdf · (CANCERRESEARCH50,3078-3086.May15,1990] HeterotransplantationofHumanLymphoidNeoplasmsUsingaNudeMouse](https://static.fdocuments.us/doc/165x107/5e83f48eaae3144d7c04ca6b/heterotransplantationofhumanlymphoidneoplasmsusinganudemou-cancerresearch503078-3086may151990.jpg)

