
AutocrineTransformingGrowthFactoraIsDispensibleforv-rasHa...
12
[CANCER RESEARCH 55. 1883-1893, May 1, 1995] Autocrine Transforming Growth Factor a Is Dispensible for v-rasHa-induced Epidermal Neoplasia: Potential Involvement of Alternate Epidermal Growth Factor Receptor Ligands1 Andrzej A. Dlugosz,2 Christina Cheng, Erin K. Williams, Nadine Darwiche, Peter J. Dempsey, Bruce Mann, Ashley R. Dunn, Robert J. Coffey, Jr., and Stuart H. Yuspa Laboratory of Cellular Carcinogenesis and Tumor Promotion, Division of Cancer Etiology, National Cancer Institute, Bethesda, Maryland 20892 [A. A. D., C. C„E. K. W., N. D., S. H. YJ; Departments of Medicine and Cell Biology, Vanderbill University School of Medicine, Nashville, Tennessee 37232 ¡P.J. D., R. J. C.¡;and Ludwig Institute for Cancer Research, Royal Melbourne Hospital, Victoria 3050, Australia [B. M., A. R. DJ ABSTRACT Autocrine epidermal growth factor receptor activation by transforming growth factor a (TGFa) has been implicated in growth stimulation during epithelial neoplasia. Using keratinocytes isolated from mice with genetic defects in TGFa expression, we tested whether TGFa is required for transformation by the v-ras"" oncogene. Introduction of v-ras"" into primary epidermal cultures using a retroviral vector stimulated growth of both control (TGFa +/+, BALB/c) and TGFa-deficient (TGFa -/-, ira-I i keratinocytes. Moreover, v-ra.v"1' elicited characteristic changes in marker expression (keratin 1 was suppressed; keratin 8 was induced), previously shown to be associated with epidermal growth factor (EGF) receptor activation, in both TGFa +/+ and TGFa -/- keratinocytes. v-ra.v"" markedly increased secreted o 10-fold) and cell-associated (2-3- fold) TGFa levels in keratinocytes from TGFa +/+ and BALB/c mice, but not TGFa —/— or wa-l mice. Based on Northern blot analysis, \-ras"" induced striking up-regulation of transcripts encoding the additional EGF family members amphiregulin, heparin-binding EGF-like growth factor, and betacellulin in cultured keratinocytes from all four mouse strains. Interestingly, in addition to the normal 4.5-kilobase TGFa transcript, wa-l keratinocytes expressed two additional TGFa transcripts, 4.7 and 5.2 kilobases long. All three transcripts were up-regulated in response to \-ras"", as well as exogenous TGFa or keratinocyte growth factor treat ment, and were also detected in RNA isolated from wa-l brain and skin. In vivo, v-rasH" keratinocytes from control as well as TGFa-deficient mice produced squamous tumors when grafted onto nude mice, and these lesions expressed high levels of amphiregulin, heparin-binding EGF-like growth factor, and betacellulin mRNA, regardless of their TGFa status. These findings indicate that TGFa is not essential for epidermal neoplasia induced by the v-ra.v"1'oncogene and suggest that another EGF family member(s) may contribute to autocrine growth stimulation of ra.v-trans- formed keratinocytes. INTRODUCTION The majority of malignancies affecting humans arise in the epithe lial component of organs including the skin, breast, lung, and gastro intestinal and genitourinary tracts. Both clinical data and results of studies using animal models indicate that epithelial neoplasia is a stepwise process driven by the accumulation of multiple genetic and epigenetic changes (reviewed in Ref. l). Chemical carcinogenesis in mouse skin has provided a powerful experimental model for studying both early changes involved in the development of premalignant lesions, as well as later events important in neoplastic progression (2). Application of a subtumorigenic dose of the chemical carcinogen Received 12/1/94; accepted 2/24/95. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance wilh 18 U.S.C. Section 1734 solely to indicate this fact. 1This work was supported in part by a Thomas B. Fitzpatrick-Kao Corporation Research Award (A. A. D.), and by Grant MCICA46413 and by the Veterans Adminis tration (R. J. C.). R. J. C. is a Veterans Administration Clinical Investigator. 2 To whom requests for reprints should be addressed, at LCCTP/NCI, Bldg 37/Rm 3B08, 37 Convent DR MSC 4255, Bethesda, MD 20892-4255. DMBA3 to mouse skin results in mutation of the c-ra.vHa gene (3). Repeated treatment with the phorbol ester 12-O-tetradecanoylphor- bol-13-acetate promotes the clonal expansion of cells expressing the mutant ras gene to form a benign tumor capable of autonomous growth. The central role of ras in the pathogenesis of benign papil- lomas was corroborated in studies where introduction of oncogenic ras into wounded skin could replace chemical initiation by DMBA (4), as well as nude mouse experiments in which primary keratino cytes transduced with v-rasHa in vitro produced tumors when grafted as a reconstituted skin (5). In addition to its involvement in early stages of cutaneous neoplasia, raÃ-has also been implicated in malig nant progression of skin tumors (reviewed in Ref. l). Since mutation of cellular raÃ-genes has been detected in a variety of human cancers (6, 7), understanding how activated Ras influences keratinocyte biology is likely to provide insights relevant to human disease. Numerous studies have documented alterations in growth factor signaling pathways during epithelial neoplasia, suggesting that these changes may be important in the development and/or maintenance of the neoplastic phenotype. TGFa and its receptor, EGFR, appear to play a particularly prominent role in epithelial neoplasia (reviewed in Refs. 8 and 9). Expression of v-raiHa in cultured mouse keratinocytes induces a large increase in TGFa mRNA and protein expression (10, 11). In addition, high levels of TGFa have been detected in papillo- mas produced either by grafting v-raiHa keratinocytes onto nude mice (10), or generated using a two-stage chemical carcinogenesis protocol (12-14). Treatment of normal keratinocytes in vitro with exogenous TGFa elicits many of the same phenotypic alterations as v-raÃ-Ha(11), suggesting that oncogenic raÃ-may alter keratinocyte behavior, at least in part, via activation of the TGFa/EGFR signaling pathway. Con sistent with this concept, transgenic mice in which TGFa is targeted to the epidermis using keratin 14 (K14) or keratin 1 (Kl) promoters develop squamous papillomas following tumor promotion by wound ing or 12-O-tetradecanoylphorbol-13-acetate treatment (15, 16), sug gesting that overexpression of TGFa can substitute for raÃ-Hagene activation as an initiator in two-stage cutaneous carcinogenesis. It has also been suggested that TGFa can function as a tumor-promoting stimulus, based on the results of two-stage chemical carcinogenesis studies (13) and grafting experiments using papilloma cell lines in an environment where TGFa is overexpressed (17). The development of cutaneous papillomas and carcinomas following DMBA treatment of a TGFa transgenic mouse strain (18) supports the concept that TGFa can function as a tumor promoter. In addition to TGFa and EGF, several other structurally related proteins bind and activate the EGFR, including amphiregulin (19), HB-EGF (20), and betacellulin (21). The role of these "alternate" EGFR ligands in the physiology of normal or neoplastic keratinocytes has not yet been defined. 3 The abbreviations used are: DMBA, 7,12-dimethylbenz[a]anthracene; EGFR, epi dermal growth factor receptor; TGFa, transforming growth factor a; HB-EGF, heparin- binding epidermal growth factor-like growth factor; KGF, keratinocyte growth factor; CM, conditioned medium; TBS, Tris-buffered saline (20 mM Tris, pH 7.4-500 IHMNaCI); poly(A)+ RNA, polyadenylated RNA; PL-7, ribosomal protein L7. 1883 on July 18, 2018. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Transcript of AutocrineTransformingGrowthFactoraIsDispensibleforv-rasHa...

[CANCER RESEARCH 55. 1883-1893, May 1, 1995]
Autocrine Transforming Growth Factor a Is Dispensible for v-rasHa-induced
Epidermal Neoplasia: Potential Involvement of AlternateEpidermal Growth Factor Receptor Ligands1
Andrzej A. Dlugosz,2 Christina Cheng, Erin K. Williams, Nadine Darwiche, Peter J. Dempsey, Bruce Mann,
Ashley R. Dunn, Robert J. Coffey, Jr., and Stuart H. Yuspa
Laboratory of Cellular Carcinogenesis and Tumor Promotion, Division of Cancer Etiology, National Cancer Institute, Bethesda, Maryland 20892 [A. A. D., C. C„E. K. W.,N. D., S. H. YJ; Departments of Medicine and Cell Biology, Vanderbill University School of Medicine, Nashville, Tennessee 37232 ¡P.J. D., R. J. C.¡;and Ludwig Institute forCancer Research, Royal Melbourne Hospital, Victoria 3050, Australia [B. M., A. R. DJ
ABSTRACT
Autocrine epidermal growth factor receptor activation by transforminggrowth factor a (TGFa) has been implicated in growth stimulation duringepithelial neoplasia. Using keratinocytes isolated from mice with geneticdefects in TGFa expression, we tested whether TGFa is required fortransformation by the v-ras"" oncogene. Introduction of v-ras"" into
primary epidermal cultures using a retroviral vector stimulated growth ofboth control (TGFa +/+, BALB/c) and TGFa-deficient (TGFa -/-,ira-I i keratinocytes. Moreover, v-ra.v"1' elicited characteristic changes in
marker expression (keratin 1 was suppressed; keratin 8 was induced),previously shown to be associated with epidermal growth factor (EGF)receptor activation, in both TGFa +/+ and TGFa -/- keratinocytes.v-ra.v"" markedly increased secreted o 10-fold) and cell-associated (2-3-
fold) TGFa levels in keratinocytes from TGFa +/+ and BALB/c mice,but not TGFa —/—or wa-l mice. Based on Northern blot analysis, \-ras""
induced striking up-regulation of transcripts encoding the additional EGFfamily members amphiregulin, heparin-binding EGF-like growth factor,
and betacellulin in cultured keratinocytes from all four mouse strains.Interestingly, in addition to the normal 4.5-kilobase TGFa transcript,wa-l keratinocytes expressed two additional TGFa transcripts, 4.7 and 5.2kilobases long. All three transcripts were up-regulated in response to\-ras"", as well as exogenous TGFa or keratinocyte growth factor treat
ment, and were also detected in RNA isolated from wa-l brain and skin.In vivo, v-rasH" keratinocytes from control as well as TGFa-deficient mice
produced squamous tumors when grafted onto nude mice, and theselesions expressed high levels of amphiregulin, heparin-binding EGF-like
growth factor, and betacellulin mRNA, regardless of their TGFa status.These findings indicate that TGFa is not essential for epidermal neoplasiainduced by the v-ra.v"1'oncogene and suggest that another EGF family
member(s) may contribute to autocrine growth stimulation of ra.v-trans-
formed keratinocytes.
INTRODUCTION
The majority of malignancies affecting humans arise in the epithelial component of organs including the skin, breast, lung, and gastrointestinal and genitourinary tracts. Both clinical data and results ofstudies using animal models indicate that epithelial neoplasia is astepwise process driven by the accumulation of multiple genetic andepigenetic changes (reviewed in Ref. l). Chemical carcinogenesis inmouse skin has provided a powerful experimental model for studyingboth early changes involved in the development of premalignantlesions, as well as later events important in neoplastic progression (2).Application of a subtumorigenic dose of the chemical carcinogen
Received 12/1/94; accepted 2/24/95.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance wilh18 U.S.C. Section 1734 solely to indicate this fact.
1This work was supported in part by a Thomas B. Fitzpatrick-Kao Corporation
Research Award (A. A. D.), and by Grant MCICA46413 and by the Veterans Administration (R. J. C.). R. J. C. is a Veterans Administration Clinical Investigator.
2 To whom requests for reprints should be addressed, at LCCTP/NCI, Bldg 37/Rm
3B08, 37 Convent DR MSC 4255, Bethesda, MD 20892-4255.
DMBA3 to mouse skin results in mutation of the c-ra.vHa gene (3).
Repeated treatment with the phorbol ester 12-O-tetradecanoylphor-bol-13-acetate promotes the clonal expansion of cells expressing the
mutant ras gene to form a benign tumor capable of autonomousgrowth. The central role of ras in the pathogenesis of benign papil-
lomas was corroborated in studies where introduction of oncogenicras into wounded skin could replace chemical initiation by DMBA(4), as well as nude mouse experiments in which primary keratinocytes transduced with v-rasHa in vitro produced tumors when grafted
as a reconstituted skin (5). In addition to its involvement in earlystages of cutaneous neoplasia, raÃhas also been implicated in malignant progression of skin tumors (reviewed in Ref. l). Since mutationof cellular raÃgenes has been detected in a variety of human cancers(6, 7), understanding how activated Ras influences keratinocytebiology is likely to provide insights relevant to human disease.
Numerous studies have documented alterations in growth factorsignaling pathways during epithelial neoplasia, suggesting that thesechanges may be important in the development and/or maintenance ofthe neoplastic phenotype. TGFa and its receptor, EGFR, appear toplay a particularly prominent role in epithelial neoplasia (reviewed inRefs. 8 and 9). Expression of v-raiHa in cultured mouse keratinocytes
induces a large increase in TGFa mRNA and protein expression (10,11). In addition, high levels of TGFa have been detected in papillo-mas produced either by grafting v-raiHa keratinocytes onto nude mice
(10), or generated using a two-stage chemical carcinogenesis protocol(12-14). Treatment of normal keratinocytes in vitro with exogenousTGFa elicits many of the same phenotypic alterations as v-raÃHa(11),
suggesting that oncogenic raÃmay alter keratinocyte behavior, at leastin part, via activation of the TGFa/EGFR signaling pathway. Consistent with this concept, transgenic mice in which TGFa is targetedto the epidermis using keratin 14 (K14) or keratin 1 (Kl) promotersdevelop squamous papillomas following tumor promotion by wounding or 12-O-tetradecanoylphorbol-13-acetate treatment (15, 16), suggesting that overexpression of TGFa can substitute for raÃHagene
activation as an initiator in two-stage cutaneous carcinogenesis. It hasalso been suggested that TGFa can function as a tumor-promotingstimulus, based on the results of two-stage chemical carcinogenesis
studies (13) and grafting experiments using papilloma cell lines in anenvironment where TGFa is overexpressed (17). The development ofcutaneous papillomas and carcinomas following DMBA treatment ofa TGFa transgenic mouse strain (18) supports the concept that TGFacan function as a tumor promoter. In addition to TGFa and EGF,several other structurally related proteins bind and activate the EGFR,including amphiregulin (19), HB-EGF (20), and betacellulin (21). Therole of these "alternate" EGFR ligands in the physiology of normal or
neoplastic keratinocytes has not yet been defined.
3 The abbreviations used are: DMBA, 7,12-dimethylbenz[a]anthracene; EGFR, epi
dermal growth factor receptor; TGFa, transforming growth factor a; HB-EGF, heparin-binding epidermal growth factor-like growth factor; KGF, keratinocyte growth factor;CM, conditioned medium; TBS, Tris-buffered saline (20 mM Tris, pH 7.4-500 IHMNaCI);poly(A)+ RNA, polyadenylated RNA; PL-7, ribosomal protein L7.
1883
on July 18, 2018. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

EOFR LÕGANOS AND EPIDERMAL NEOPLASIA
To test whether TGFa expression is obligatory in ras-mediatedepidermal neoplasia, we have studied the effects of v-rasHa on kera-
tinocytes from two mouse strains deficient in TGFa expression, thewa-1 spontaneous mutant (22) and the TGFa-null mouse, generated
by homologous recombination (23). Our findings argue against anobligatory role for autocrine TGFa in epidermal neoplasia and raisethe possibility that another EGFR ligand(s) may be capable of substituting for TGFa to stimulate the growth of v-rasHa-transformed
keratinocytes.
MATERIALS AND METHODS
Reagents. Recombinant human TGFa, KGF, and PCS were purchasedfrom UBI (Lake Placid, NY).
Mouse Strains and Cell Culture. Primary epidermal keratinocytes wereobtained from newborn mouse skin following overnight flotation on trypsin, asdescribed previously (24). In some experiments, keratinocytes were isolatedfrom adult mouse skin using a slightly modified protocol (25). Cells wereobtained from the following mouse strains: ABP/Le (The Jackson Laboratory,Bar Harbor, ME), a linkage strain homozygous for the wa-l mutation, which
arose spontaneously in a mouse colony of unknown genetic background (22);BALB/c (or BALB/cByJ,The Jackson Laboratory), a representative "normal"
strain; TGFa —/-, homozygous TGFa-null mice produced by homologous
recombination (23); and TGFa +/+, the TGFa-null control strain. TGFa +/+and TGFa —/—mice were generated on a C57BL6 background (23). In
addition to the wa-\ alÃele,ABP/Le mice are homozygous for the followingrecessive mutations: a, non-agouti; b, brown; bt, belted; p, pink eye; and se,
short ear.Medium was prepared with Eagle's MEM (without Ca2+ and Mg2+) sup
plemented with 8% PCS [Ca2+-depleted (24)] and 0.25% penicillin-streptomycin solution (GIBCO, Grand Island, NY). The concentration of Ca2* in the
medium was adjusted by addition of 0.3 M CaCl2 stock. Cells were cultured inmedium with 0.05 mM Ca2 * to maintain a proliferating population of basal
cells. Primary keratinocytes were transduced with a defective retrovirus harboring the v-ra.cHa oncogene as described previously (5). Growth factor treat
ments were initiated 3 days after plating and continued until cells wereharvested for RNA isolation, which was performed 8-10 days after plating.For preparation of keratinocyte CM, cells grown in 60-mm culture dishes were
washed with PBS and 2 ml of fresh medium containing 1% PCS were addedto dishes. Medium was removed after approximately 24 h, centrifuged at1000 x g to remove any floating cells and culture debris, and stored at —20°C
until RIA. For the analysis of cell-associated TGFa, cultures were washed with
PBS, and the attached cells remaining in the dish were frozen on dry ice andstored at -20°C until processed for RIA. Where indicated, keratinocytes wereinduced to terminally differentiate by raising the Ca2+ concentration from 0.05
to 0.12 mM for 48 h (26).Immortalized cell lines were established from v-ra£H"-transduced TGFa
+/+ and TGFa —/—keratinocytes as follows. Primary \-rasHa keratinocyteswere trypsinized, pelleted, resuspended in medium containing 1.4 mM Ca2+
medium, and plated into 6-well culture dishes that had been precoated with asolution of fibronectin-collagen (25). As an alternative approach, floating cellsfrom v-ra.v"a-transduced primary cultures were collected, resuspended in medium with 1.4 mM Ca2+, and plated in precoated 6-well dishes. One to 2 daysafter plating, cells were washed with Car* and Mg2+-free PBS and replenishedwith medium containing 0.05 mM Ca24 medium. Keratinocytes were propagated continuously in 0.05 mM Ca2 * medium. A similar protocol was used toestablish an immortalized cell line, designated wa-lrp, from v-rasH"-trans-
duced H'a-1 primary keratinocytes. The SP-1 cell line was established from
SENCAR mouse papillomas (27). The C57r cell line was generated usingprimary v-ra.cHa keratinocytes obtained from newborn C57BL mice, and was
generously provided by Dr. Wendy Weinberg (Laboratory of Cellular Carci-
nogenesis and Tumor Promotion/National Cancer Institute).Growth Curves. Cells were plated in 6-well tissue culture plates (Costar,
Cambridge, MA) at a density of 3 X 10s cells/well in 2 ml of medium. The
following day, cultures were washed with PBS and fresh medium was added.Additional medium changes were performed on alternate days, and cells weretransduced on day 5 in culture using a retrovirus containing \-ras"* (5). To
assess keratinocyte growth, cells from replicate wells were trypsinized on
alternate days and counted using a Coulter Counter (model ZB1; CoulterElectronics, Hialeah, FL).
Skin Grafting. Tumorigenicity was tested by grafting a slurry of keratinocytes (2.5-6 x IO6 cells) and SENCAR mouse fibroblasts (5-8 X IO6 cells)
onto prepared graft sites on the backs of nude mice, as described previously (5,28). This procedure results in the development of a benign squamous papil-loma at the graft site when v-ra.?Ha-transduced keratinocytes are tested,
whereas normal skin is produced when control keratinocytes are grafted (5). Atthe completion of experiments, animals were sacrificed and tumors wereprocessed for routine histológica! analysis. For in situ hybridization, tissue wasalso embedded in OCT (Miles, Elkhart, IN), frozen by flotation in an ethanol/dry ice bath, and stored at -70°C.
Tissue for RNA isolation was used either fresh or frozen in liquid nitrogenand stored at —70°Cuntil processing (see below).
Radioimmunoassay and Antibodies. A RIA specific for murine TGFa(29) was used to measure immunoreactive TGFa in CM from mouse keratinocyte cultures and cell lysates. A polyclonal sheep antibody was raised to ratTGFa (East Acres Biologicals, Southbridge, MA) which was also used asstandard and 125I-labeled tracer. Dose-response curves of immunoreactiveTGFa detected in CM from normal and v-rasHa-transformed mouse keratino
cyte cultures were parallel to rat TGFa standards in the TGFa RIA. Levels ofmouse TGFa are therefore expressed in equivalent units of rat TGFa standard.For determining the level of cell-associated TGFa, cells in 60-mm dishes were
harvested in 1 ml of buffer containing 25 mM Tris (pH 8.0), 50 mM NaCl, 0.5%NP40, 0.5% sodium deoxycholate, 5 pig/ml leupeptin, 5 fig/ml aprotinin, and2 mM phenylmethylsulfonyl fluoride. Lysates were vortexed, extracted on icefor 45 min, vortexed again, and centrifuged at 16,000 X g for 10 min.Supernatants were used for determination of cell-associated TGFa by RIA.
The generation and specificity of rabbit anti-peptide antibodies recognizing
keratins Kl and K14 were described in a previous report (30). For immunoblotanalysis, anti-Kl was used at a dilution of 1:1000 and anti-K14 was diluted1:15,000. A rat monoclonal antibody, TROMA-1 (31), was used at a dilution
of 1:500 to detect keratin K8. This antibody was a gift from Dr. Rolf Kemler(Max Planck Institute, Freiburg, Germany), generously provided by Dr. IvanDamjanov (Thomas Jefferson University, Philadelphia, PA).
Polyacrylamide Gel Electrophoresis and Immunoblotting. Cultures werewashed with PBS and whole cell extracts were prepared by harvesting cells in lysisbuffer [0.25 M Tris (pH 6.8), 5% SDS, and 20% 0-mercaptoethanol] and heatingthe lysates at 100°Cfor 5 min. Proteins were separated in 8.5% polyacrylamide
gels for determination of keratin expression and electrophoretically transferred toreinforced nitrocellulose (BA-S 83) (Schleicher & Schuell, Keene, NH) in a buffer
containing 25 mM Tris (pH 8.3), 192 mM glycine, and 20% methanol for l h at 100V. Nitrocellulose blots were stained with Ponceau S (Sigma, St. Louis, MO) todetermine uniformity of protein loading and transfer (32). Following blocking for30 min with 5% milk in TBS, blots were exposed to primary antibodies for l h atroom temperature. Filters were washed for 30 min using three changes of TBSwith 0.2% Tween 20 (Bio-Rad Laboratories, Richmond, CA) and incubated withhorseradish peroxidase-conjugated secondary antibody, diluted 1:5000 in 2% milk
in TBS, for 30 min. Filters were washed as before, and immunoreactive proteinsdetected using enhanced chemiluminescence reagents purchased from Amersham(Arlington Heights, IL).
RNA Isolation and Northern Blot Analysis. Poly(A)+ RNA was isolated
from cells growing in 150-mm tissue culture dishes as described previously
(33). Total RNA from epidermal tumors or normal mouse tissues was preparedby homogenization in a guanidine isothiocyanate-phenol solution (TRIzol;
GIBCO, Gaithersburg, MD) based on a previously published protocol (34). Insome cases, total RNA served as a source for poIy(A)+ RNA isolation using
a modification of a protocol described earlier (33). RNA species were separated by electrophoresis through a 1% agarose gel containing 0.66 M formaldehyde and 0.66 fig/ml ethidium bromide and transferred to reinforced nitrocellulose (BA-S NC; Schleicher and Schuell, Keene, NH) (35). Transcript
lengths were determined using rRNA as markers. The following probes wereused, labeled with 32P by nick-translation, for Northern blot analysis: rat
TGFa, full-length cDNA (36); glyceraldehyde phosphate dehydrogenase, full-length cDNA (37); keratin 1 (Kl), 400-base pair cDNA (26); 400-base pair ratamphiregulin, 1.5-kilobase rat HB-EGF, 650-base pair mouse betacellulin, and784-base pair mouse EOF cDNA fragments (38). A 380-base pair cDNAfragment recognizing mRNA encoding PL-7 (39) was labeled by random
priming.
1884
on July 18, 2018. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
EGFR LÕGANOS AND EPIDERMAL NEOPLASIA
BALB/c TGFCC+/+ TGFa-/-
Control
M-ras Ha
B107
LU
EI 1°6CO
HiO
105
-I- control
1 3 5 7 9 11 13 15 17
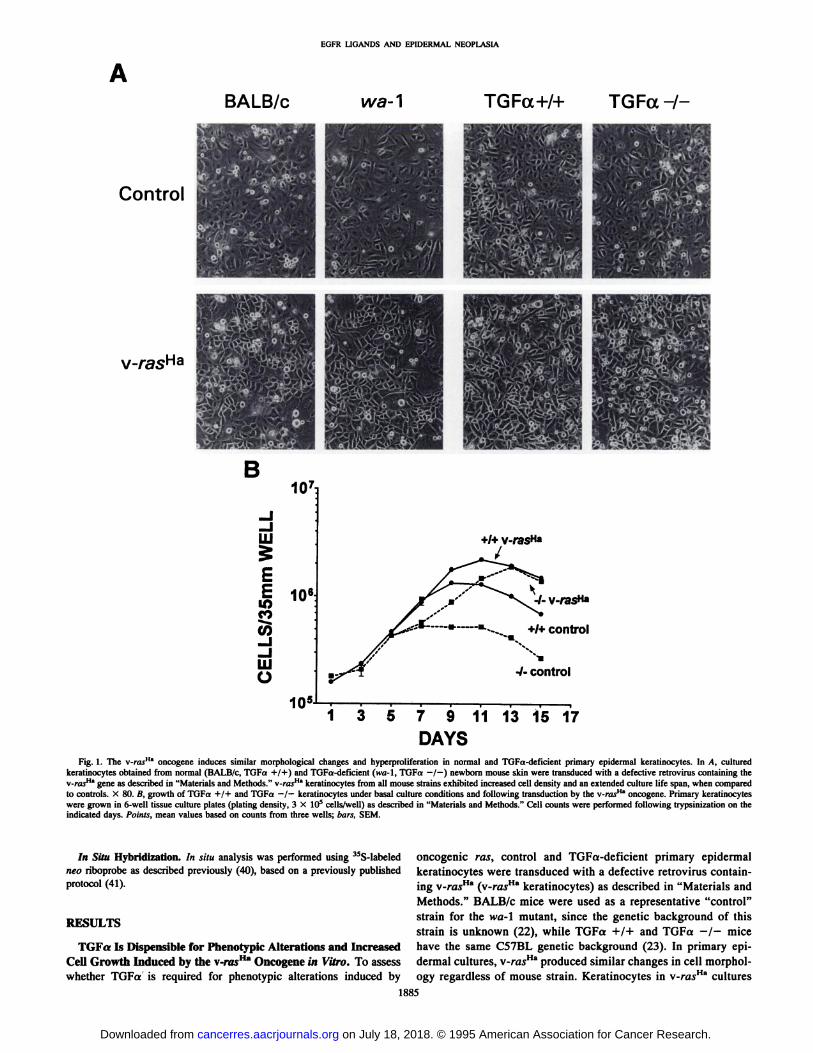
DAYSFig. 1. The v-rasHa oncogene induces similar morphological changes and hyperproliferation in normal and TGFa-deficienI primary epidermal keratinocytes. In A, cultured
keratinocytes obtained from normal (BALB/c, TGFa +/+) and TGFa-deficient (wa-1, TGFa —/—)newborn mouse skin were transduced with a defective retrovirus containing thev-rasH" gene as described in "Materials and Methods." v-rasH" keratinocytes from all mouse strains exhibited increased cell density and an extended culture life span, when comparedto controls. X 80. B, growth of TGFa +/+ and TGFa —/—keratinocytes under basal culture conditions and following transduction by the v-rasHa oncogene. Primary keratinocyteswere grown in 6-well tissue culture plates (plating density, 3 X IO5 cells/well) as described in "Materials and Methods." Cell counts were performed following trypsinizalion on the
indicated days. Points, mean values based on counts from three wells; bars, SEM.
In Situ Hybridization. In situ analysis was performed using 35S-labeled
neo riboprobe as described previously (40), based on a previously publishedprotocol (41).
RESULTS
TGFa Is Dispensible for Phenotypic Alterations and IncreasedCell Growth Induced by the v-r<wHaOncogene in Vitro. To assess
whether TGFa is required for phenotypic alterations induced by
oncogenic ras, control and TGFa-deficient primary epidermalkeratinocytes were transduced with a defective retrovirus containing v-ras"" (v-rasHd keratinocytes) as described in "Materials andMethods." BALB/c mice were used as a representative "control"
strain for the wa-l mutant, since the genetic background of thisstrain is unknown (22), while TGFa +/+ and TGFa -/- mice
have the same C57BL genetic background (23). In primary epidermal cultures, v-rasHil produced similar changes in cell morphology regardless of mouse strain. Keratinocytes in \-rasH" cultures
1885
on July 18, 2018. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

EOFR LÕGANOS AND EPIDERMAL NEOPLASIA
Control v-rasHa
r I I
l.05 .12 .05 .12
I I I.05 .12 .05 .12
K1
K8
K14
Fig. 2. v-raiHa induces similar changes in keratin expression in TGFa null (-/-) andcontrol ( + /+) keratinocytes. Primary control and v-ra.vHakeratinocytes from the indicatedmouse strains were grown in medium with 0.12 mM Ca2+ for 48 h to induce terminal
differentiation. Western blot analysis was performed using total lysates as described in"Materials and Methods." v-ra.cHa induced similar alterations in keratin expression in both
normal and TGFa-null keratinocytes. The constitutively expressed keratin K14 was usedas a control for loading.
were generally more crowded than in corresponding controlcultures and exhibited a more phase-dense cytoplasm with prominent, phase-lucent intercellular spaces (Fig. IA). The growth ofcontrol and v-rasHa-transduced keratinocytes was assessed by per
forming cell counts over a 2-week period. From day 7 to 15 inculture, the number of cells in control TGFa —/—cultures was
significantly lower than in control TGFa +/+ cultures (Fig. 15),suggesting that endogenous TGFa plays a role in maintainingkeratinocyte growth under basal conditions in vitro. Transduction
with v-rasHa stimulated growth of keratinocytes of both genotypes;
interestingly, despite their slower growth under control conditions,TGFa —/—v-rasHa keratinocytes achieved the same final celldensity as TGFa +/+ v-rasH" keratinocytes (Fig. IB).
We have previously reported that v-rasHa keratinocytes induced to
terminally differentiate fail to express the spinous cell keratins Kl andK10 but aberrantly express keratins K8 and K18, and these changes inkeratin expression appear to be mediated by autocrine stimulation ofthe EGFR (11). Immunoblot analysis presented in Fig. 2 indicates thatv-rasHa induces comparable alterations in the keratin expression pat
tern in both TGFa +/+ and TGFa —/—keratinocytes. In controlkeratinocytes, raising extracellular Ca2+ from 0.05 to 0.12 mM in
duces expression of the differentiation-specific keratin Kl; in contrast,v-rasHa-transduced keratinocytes of either strain fail to express Kl but
synthesize K8 instead (Fig. 2). K14, which is constitutively expressedin cultured keratinocytes, is expressed at similar levels in all treatmentgroups.
TGFa Is Not Detected in Normal or v-ras"a-transformed Cul
tured Keratinocytes from TGFa-null Mice and Is Markedly Reduced in v-rasHa-transformed wa-1 Keratinocytes. Since v-rasHa
keratinocytes from all mouse strains examined exhibited phenotypicchanges previously attributed to activation of the EGFR signalingpathway, the levels of secreted and cell-associated TGFa were determined in control (BALB/c, TGFa +/+) and TGFa-deficient (wa-1,TGFa -/-) cultures (Fig. 3). Secreted TGFa in CM from either
BALB/c or wa-l control cultures was below the level of detection,
which was 39 pg/ml for this assay (Fig. 3A). TGFa secretion waselevated to 490 ±20 (SEM) pg/ml in CM from v-rasHa-transformed
BALB/c keratinocytes; in contrast, the level of TGFa in CM fromwa-l v-rasHa keratinocyte cultures was marginally increased, to87 ±2.5 pg/ml (Fig. 3A). v-ra.vHa also induced dramatic changes in
the level of cell-associated TGFa in BALB/c, but not wa-l, kerati-
B525-
£ "50J0 375-
9t300
1 8 225K 150-
j BALB/c
ras contr
TREATMENT
^2zmTGFa */+••TGFa •/-
ras contr
TREATMENT
ras contr
TREATMENT
contr
TREATMENT
Fig. 3. Induction of secreted and cell-associated TGFa by the v-rasH" oncogene is blocked in wa-l and TGFa null keratinocytes. A, wa-l keratinocytes. The level of TGFa wasdetermined by RIA in CM (2 ml/60-mm dish) collected following a 24-h exposure to control or v-rasHa-transduced primary keratinocytes. Cell-associated TGFa was measured in cellextracts prepared from keratinocytes attached to the substratum, as described in "Materials and Methods." Results are presented as mean values based on determinations performedusing three dishes; bars, SEM; B, TGFa-null primary keratinocytes; C, immortalized v-raiHa-transformed keratinocyte cell lines. Keratinocyte cell lines were established followingv-ru.!Ha transduction of epidermal cultures isolated from TGFa -/- mice (lines a-1, a-2, and a-6) or the parental mouse strain TGFa +/+ (line a + 2), as described in "Materialsand Methods." TGFa levels were assayed in CM and cell lysates for each cell line as described above. , RIA "background," determined using medium not previously exposed to
cells (for CM) or lysis buffer (for cell lysates).
1886
on July 18, 2018. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

EOFR LÕGANOS AND EPIDERMAL NEOPLASIA
BALB/c iva-1
TGFa
GAPDH
u.o O CD <O I- s¿ >
HB-EGF
GAPDH ...*
GAPDH
Fig. 4. Expression of transcripts encoding multiple EGFR ligands is coordinatelyregulated in cultured BALB/c and wa-l keratinocytes. Primary cultures were either grownunder basal conditions (Control), transduced with the v-r«.vll;loncogene, or treated with 10
ng/ml TGFa or KGF for 3-4 days. Northern blot analysis was performed using 1 fig ofpoly(A) ' RNA/lane, as described in "Materials and Methods." Expression of transcripts
for EGFR ligands was assessed by reprobing filters multiple times, and uniformity ofmRNA loading was evaluated by probing for glyceraldehyde phosphate dehydrogenase(GAPDH) transcripts. The three TGFa transcripts detected in stimulated wa-l keratino
cytes have approximate lengths of 4.5, 4.7, and 5.2 kilobases. Based on densitometricanalysis and normalization to glyceraldehyde phosphate dehydrogenase, the maximumcombined expression level of the three wa-l TGFa mRNAs (v-rai"" lane) was 6-foldlower than the level of the TGFa transcript in corresponding BALB/c v-r«i"akeratinocyte
cultures. AR, amphiregulin; BC, betacellulin.
nocytes. In control cultures, TGFa measured in cell lysates was at orslightly above the assay background (31 ±4 pg/60-mm dish) in bothBALB/c and wa-l keratinocytes. In contrast, v-raiHa increased the
abundance of cell-associated TGFa in BALB/c keratinocytes to176 ±50 pg/60-mm dish, while the TGFa level in wa-l v-rasHa
keratinocyte lysates was only slightly higher than in control wa-l
keratinocytes (47 ±1.2 versus 34 ±2.1 pg/60 mm dish, respectively)(Fig. 3A).
A similar analysis was performed using v-ras"" primary keratino
cytes from TGFa +/+ and TGFa -/- mice. Like BALB/c keratinocytes, TGFa +/+ primary keratinocytes transduced with v-rasHa
expressed high levels of both secreted (799 ±81 pg/ml) and cell-associated (118 ±29 pg/60-mm dish) TGFa; in contrast, in cultures
of TGFa —/—keratinocytes, TGFa was not increased by \-ras"*
above control levels (Fig. 3ß).CM and cell lysates were also preparedusing established cell lines derived from v-ra.vlla-transduced primarykeratinocytes, as described in "Materials and Methods." The raj-
transformed control cell line a + 2 produced 980.6 ±111 pg/ml ofsecreted TGFa and 420.2 ±116 pg/60-mm dish of cell-associated
TGFa. In contrast, there was no detectable increase in either secretedor cell-associated TGFa in three ras-transformed cell lines derivedfrom TGFa -/- epidermal cultures (Fig. 3C). Together, these dataconfirm that cultured TGFa -/- and wa-l keratinocytes exhibit aprofound defect in v-ra.vHa-induced TGFa expression, supporting
results reported previously in which TGFa expression was measuredin tissues from these mouse strains (23, 42). Combined with the datapresented in Figs. 1 and 2, these results indicate that phenotypicchanges induced by v-rasH* in cultured epidermal keratinocytes,
involving growth regulation as well as marker expression, are notdependent on autocrine stimulation by TGFa.
Expression of Transcripts Encoding Multiple EGFR Ligands IsRegulated Coordinately in Normal and TGFa-deficient Keratinocytes. To address the possibility that another growth factor may becompensating for TGFa in wa-l and TGFa —/—keratinocytes, ex
pression of transcripts encoding other mammalian EGFR ligands wasdetermined by Northern blot analysis of poly(A)+ RNA from controland TGFa-deficient keratinocytes. v-raiHa increased the expression
of TGFa transcripts in both wa-l and BALB/c cultures, although the
induction in BALB/c keratinocytes was substantially higher than inwa-l cultures (Fig. 4). In contrast to TGFa mRNA in normal keratinocytes, which is detected as a single species —4.5kilobases long,three TGFa transcripts were present in RNA from wa-l keratinocytes,with estimated sizes of —4.5,4.7, and 5.2 kilobases (Fig. 4). Thesethree bands were also detected in RNA from wa-l brain and skin (Fig.
nu wa-1
.E e -E5 15 2
CQ C/) Cu
TGFa
PL-7
Fig. 5. Multiple TGFa transcripts are expressed in wa-l tissues in vivo. Northern hlotanalysis was performed using 15 jig of poly(A)+-enriched RNA isolated from the
indicated wa-l or nude (nu) mouse tissues. The filter was reprobed for PL-7 mRNA as acontrol for loading. Based on densitometric scanning and normalization to PL-7, thecombined abundance of the three TGFa transcripts in RNA from wa-1 brain was 42% ofthe level detected in nu mouse brain RNA.
1887
on July 18, 2018. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

EGFR LÕGANOS AND EPIDERMAL NEOPLASIA
i _ II __
Ìf> if»C R> C (Oo v o >u > u >
•-TGF-a
AR
HB-EGF
BTCL
GAPDH
Fig. 6. v-raslia coordinately induces transcripts encoding multiple EGFR ligands in
TGFa-null and control keratinocytes in vitro. Primary epidermal keratinocytes fromTGFa null (-/-) and control (+/+) mice were grown under basal conditions ortransduced with \-ras"'. Poly(A)+ RNA from the indicated cultures (3 /j.g/lane) was
analyzed by Northern blotting. The targeting strategy used for generating TGFa-null miceresults in the production of a 6.2-kilobase TGFa transcript in —/—keratinocytes. GAPDH,
glyceraldehyde phosphate dehydrogenase; AR, amphiregulin; BTCL, betacellulin.
5), indicating that the expression of multiple TGFa transcripts is notrestricted to stimulated wa-\ keratinocytes in vitro. In agreement witha previous report (42), the expression of TGFa mRNA in wa-\ brainwas lower than in control brain; however, only the 4.5-kilobase TGFatranscript was detected in the earlier study (see "Discussion").
Stimuli which induce TGFa gene expression also increase the levelof amphiregulin, HB-EGF, and betacellulin transcripts in culturedkeratinocytes (Fig. 4). In addition to v-rasHa, treatment with TGFa or
KGF up-regulated expression of multiple EGFR ligands in BALB/cand wa-\ cultures (Fig. 4), supporting our recent findings suggesting
that KGF influences keratinocyte biology via activation of the EGFRsignaling pathway (43). Up-regulation of multiple EGFR ligands in
stimulated keratinocytes does not involve all members of this growthfactor family, since transcripts encoding EOF were not detected inepidermal cultures under any of the conditions tested (data notshown).
The effects of v-ras"" on EGFR ligand gene expression were alsoexamined in keratinocytes from TGFa +/+ and TGFa —/- mice.
The usual single 4.5-kilobase TGFa mRNA was up-regulated inv-rasHa-transduced TGFa +/+ keratinocytes, whereas a 6.2-kilobase
TGFa transcript, resulting from the introduction of a 1.7-kilobasePGK-neo cassette in the targeted TGFa gene (23), was induced inTGFa null v-rasHa keratinocytes. Furthermore, as observed with wa-land BALB/c keratinocytes, v-rasHa coordinately induced expression
of amphiregulin, HB-EGF, and betacellulin mRNAs in TGFa-null
and control keratinocytes (Fig. 6). Together, these data indicate thattranscripts encoding multiple EGFR ligands are coordinately regulated in normal as well as TGFa-deficient cultured keratinocytes,
providing a potential mechanism whereby loss, or deficiency, of oneEGFR ligand may be supplanted by another.
Control and TGFa-deficient v-ras"" Keratinocytes Produce
Tumors When Grafted onto Athymic Mice. Tumorigenicity wasassessed by grafting v-rasHa keratinocytes as a reconstituted skin onto
nude mouse hosts (5). Control (BALB/c and TGFa +/+) as well asTGFa-deficient (wa-l and TGFa -/-) v-rasHa keratinocytes pro
duced epidermal tumors in skin grafts (Table 1; Fig. 7). In severalexperiments, the appearance of tumors produced by ras-transformedwa-l and TGFa -/- keratinocytes was delayed when compared
with control tumors. Interestingly, tumors produced by primary wa-lv-raiHa keratinocytes exhibited a greater propensity for malignant
conversion than control BALB/c cells (Table 1). In addition, animmortalized wa-l v-rasHa-transformed cell line (wa-lrp; see"Materials and Methods") also exhibited an increased tendency for
malignant conversion, when compared to two control cell lines, SP-1and C57rp, expressing activated ras11*(Table 1; Fig. 7). Since a region
Table 1 Tumorigenicity of TGFa-deficient keratinocytes expressing oncogenic ras " in
nude mouse skin grafis
v-ras a-transduced primary keratinocytes
Latency" Tumor
Experiment Strain (wk) incidence Histology'
- Graft ageat sacrifice
(wk)
wa-l 3-7 4/5 Carcinoma/spindle 13cell tumor (l/4)d
Papilloma/spindle celltumor (1/4)
Skin (2/4)f
BALB/c 3 5/5 Papilloma (3/4)Skin (1/4)
wa-l 2-4 10/10 Papilloma/carcinoma (1/8) 10
Dysplastic papilloma (2/8)Papilloma (4/8)Skin (1/8)
34BALB/cTGFa-/-TGFa
+/+TGFa
-/-TGFa
+/+2-32-32MVNA3/54/42/23/44/5Skin
(2/2)Papilloma
(3/4)Skin(1/4)Papilloma(2/2)Papilloma
(3/4)Skin(1/4)Papilloma(3/4)Skin
(1/4)55Celllines*56wa-lrpSP-1wa-lrp2-52-54-89/1010/109/10Spindle
cell tumor(1/4)Carcinoma(3/4)Papilloma/carcinoma(1/1)Carcinoma
(1/3)1210Carcinoma/spindle cell tumor (2/3)
7 wa-lrp 2-4 9/9 Papilloma/carcinoma (2/8) 5Carcinoma (1/8)Carcinoma/spindle cell tumor (4/8)Carcinoma/spindle cell
tumor/papilloma (1/8)C57rp 4 5/5 PapUloma (5/5)
4 a-1 NA 5/5 Papilloma (4/5) 5Dysplastic papilloma (1/5)
8 a-2 NA 3/4 Spindle cell tumor (3/4) 7Skin (1/4)
wa-lrp NA 3/4 Papilloma/carcinoma (2/4)Spindle cell tumor (1/4)Skin (1/4)SP-1 NA 2/2 Papilloma (2/2)
a Time interval after grafting when tumor(s) first reported.h Maximum number of tumor-bearing animals/total animals.' Certain tumors contained different regions with distinct histopathologies.''Tumor-bearing animals/total animals remaining at completion of experiment.
Significant animal mortality occurred in some experiments.e Presence of relatively normal-appearing skin in graft site. May result from sponta
neous regression of certain benign lesions or poor graft "take" at the outset of the
experiment.^NA, data not available.* wa-lrp, v-rajHa-transformed iva-1 keratinocyte cell line; SP-1, SENCAR papilloma-
derived cell line containing mutated c-ras a; C57rp, v-ras a-transformed C57BL keratinocyte cell line; a-1, a-2, v-rasHa-transformed TGFa -/- keratinocyte cell lines. See"Materials and Methods" for additional details.
1888
on July 18, 2018. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

EOFR LÕGANOS AND EPIDERMAL NEOPLASIA
Fig. 7. Control and TGFa-deficient keratino-cyles expressing the v-ra.(Ha oncogene produce tu
mors when grafted onto nude mice. A, B, grossappearance of tumors generated following graftingof control (TGFa +/+) (A) or TGFa-null (-/-)(B) v-ra.sllu keratinocytes onto nude mice, as described in "Materials and Methods." Animals were
photographed 5 weeks after grafting. C-F, histopa-thology. Tumor sections were stained with H & E.Photomicrographs of representative lesions reveal apattern typical for benign squamous papillomas.Major changes were not detected when comparingtumors arising from v-rajH"-transduced TGFa+/+ (C, £)and TGFa -/- (D. F) keratinocytes.G, 7-week-old tumor produced by grafted wa-lrp
cell line. Carcinoma in lower portion of photomicrograph is contiguous to overlying squamouspapilloma. C. D. X 15; E, F, G, X 45.
of papilloma was detected in several wa-lrp malignant tumors In similar experiments performed using TGFa-null keratinocytes,harvested either 5 or 7 weeks after grafting (Table 1), it appears the gross appearance and histology of 5-week-old tumors produced bythat these cells underwent rapid neoplastic progression in vivo and grafting either TGFa +/+ or TGFa -/- v-rasHa keratinocytes waswere not malignant at the time of grafting. typical for benign squamous papillomas (Fig. 7). Detection of neo'
1889
on July 18, 2018. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

EOFR LÕGANOS AND EPIDERMAL NEOPLASIA
TG Fa^- IG Foc+/+
Fig. 8. TGFa -/- v-rajH" papillomas are comprised of grafted TGFa-null keratinocytes, and not host keratinocytes recruited into the graft site. Dark-field in situ detection of neo'transcripts in papillomas produced by TGFa -/— v-rajHa keratinocyles. Tumor sections were hybridized to an antisense riboprobe specific for neoT mRNA. A positive signal is detectedin the epithelial component of tumors produced by grafting TGFa -/- keratinocytes but not TGFa control (+/+) keratinocytes. Corresponding bright-field images in lower panelsdemonstrate neoplastic epithelium in both +/+ and -/— grafts. X 160.
transcripts by in situ hybridization confirmed that TGFa —/—v-rasHa papillomas were comprised of grafted TGFa-null kerati
nocytes, and not host keratinocytes recruited into the graft site(Fig. 8). Interestingly, a TGFa-null v-raiHa-transformed cell line,
a-2, produced malignant tumors with a spindle cell histology 7
weeks after grafting (Table 1). Together, these results indicate thatautocrine TGFa is dispensible for the growth of benign as well asmalignant epidermal tumors.
Squamous Papillomas Express Transcripts Encoding MultipleEGFR Ligands. In light of the in vitro findings presented in Figs. 4and 6, Northern blot analysis was performed to determine the expression pattern of EGFR ligand mRNAs in papillomas produced bygrafting v-raiHa-transformed control or TGFa-null keratinocytes.
Consistent with earlier reports using other mouse strains (10, 13),TGFa mRNA was readily detected in total RNA samples preparedfrom TGFa +/+ papillomas; in contrast, the recombinant 6.2-kilo-
base TGFa transcript was expressed in tumors produced by TGFa-/- v-rasHa keratinocytes (Fig. 9). The paucity of native TGFatranscripts in TGFa -/- tumors validates the use of this model
system for studying in vivo effects of autocrine TGFa deficiency onneoplastic keratinocytes. Further analysis of tumor RNA revealed thatamphiregulin, HB-EGF, and betacellulin transcripts are similarly up-regulated in both control and TGFa -/- v-ras"" tumors (Fig. 9).
Intriguingly, the only EGFR ligand transcript detected in poly(A)"1"
RNA isolated from newborn mouse epidermis was that encodingbetacellulin (Fig. 9). Collectively, these data indicate that autocrineTGFa is not required for the development of v-rasHa-induced squa-
mous papillomas and are in keeping with in vitro results demonstrating coordinate up-regulation of multiple EGFR ligands in stimulated
keratinocytes.
DISCUSSION
Utilizing two mouse strains with distinct genetic defects in TGFaexpression, we have demonstrated that formation of tumors comprisedof epidermal keratinocytes expressing the v-rasHa oncogene is not
dependent on autocrine TGFa. TGFa-deficient and control tumorsexpress transcripts encoding amphiregulin, HB-EGF, and betacellulin,and mRNAs encoding these EGFR ligands are coordinately up-regulated in cultured keratinocytes in response to v-rasHa as well as
exogenous TGFa or KGF. These findings raise the possibility thatanother EGFR ligand, or ligands, may contribute to autocrine growthregulation of ras-transformed keratinocytes.
Increased TGFa expression has been demonstrated in a variety ofhuman epithelial tumors, cultured primary and immortalized tumorcells, and normal epithelial cells transformed in vitro (reviewed in
1890
on July 18, 2018. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

EGFR LÕGANOSAND EPIDERMAL NEOPLASIA
,Epi.
TGF-a
HB-EGF
GAPDH
BC
GAPDH
keratin 1
Fig. 9. Transcripts encoding multiple EGFR ligands are coordinately up-regulated insquamous papillotnas. Tolal RNA was isolated from 8 tumors generated by graftingcontrol ( + /+) and TGFot-null (~/~) v-ra.sH" keratinocytes onto nude mice, as describedin "Materials and Methods." Lanes 1-4 and 5-8, RNA isolations from tumors generated
in two independent experiments. Each of the paired lanes contains RNA from a separatetumor produced by grafting v-rajHa keratinocytes of the indicated genotype onto nudemice, as described in "Materials and Methods." Thirty /Ag of tumor RNA were loaded/lane, with 1.5 ftg of poly(A) + RNA from newborn mouse epidermis (Epi.). Despite
the extremely low level of glyceraldehyde phosphate dehydrogenase (GAPDH) transcriptsin epidermal mRNA, keratin 1 mRNA is easily detected. AR, amphiregulin; BC,betacellulin.
Refs. 8 and 9). The observation that EGFR levels are frequentlyup-regulated in human carcinomas (reviewed in Refs. 44 and 45)
strengthens the hypothesis that autocrine activation of this signalingpathway is important for tumor growth. Furthermore, TGFa trans-
genie mouse strains are predisposed to the development of neoplasiainvolving the skin (15, 16), breast (46, 47), pancreas (48), and liver(49). Involvement of an autocrine TGFa loop in tumor cell growth hasbeen substantiated using neutralizing antibodies or antisense DNA insome neoplastic cell types in vitro (50-53) and in xenografts of an
ovarian carcinoma cell line (54). However, neutralizing EGFR antibodies are substantially more effective at blocking tumor growth thanligand-specific antibodies (reviewed in Ref. 55), suggesting that up-
regulation of multiple EGF family members, as reported here, mayoccur in other tumor models. In addition, the ability of TGFa toinduce multiple other EGFR ligands (Fig. 4; Ref. 38) raises thepossibility that one of these other growth factors may play a criticalrole in the development of epithelial tumors in TGFa transgenic mice.
The delayed appearance of tumors produced by TGFa-deficient
keratinocytes suggests that although this growth factor is not obligatory for rai-mediated epidermal neoplasia, it may be an important
regulator of tumor growth during certain stages of this process. A
similar conclusion was reached in a study where human TGFa wasoverexpressed in skin grafts of the papilloma-producing SP-1 cell line
(17). On the other hand, a relatively high incidence of carcinomadevelopment was observed in ra.ï-transformed wa-l keratinocyteswhen compared to ras-transformed control cells. This was the case forprimary wa-l keratinocytes as well as an immortalized cell line,wa-lrp (Table 1). Although it is conceivable that the aggressive nature
of these tumors is related to TGFa deficiency, it may also reflectstrain differences in the behavior of keratinocytes from wa-l versus
other mouse strains. Alternatively, since the ABP/Le mouse strainused for wa-l studies is also homozygous for 5 additional recessivemutations (non-agouti, brown, belted, pink eye, and short ear), one ofthese other alÃelesmay be influencing the behavior of ras-transformed
keratinocytes. Strain differences can influence premalignant progression,as demonstrated by the high incidence of chemically induced epidermalcarcinomas developing in FVB/N mice when compared to other strains(56). However, the development of spindle cell tumors in TGFa-null a-2
grafts is consistent with the notion that the absence of autocrine TGFapredisposes to neoplastic progression. Additional grafting studies, chemical carcinogenesis experiments using TGFa-null and control mice, andbreeding the wa-l mutation onto a defined genetic background will be
required to critically address this issue.We have detected several intriguing abnormalities in the expression
of TGFa in wa-l keratinocytes and tissues: (a) the induction of TGFamRNA by v-rajHa is markedly lower in wa-l keratinocyte cultures
than control cultures, consistent with previous reports suggesting thatthe wa-l mutation influences the regulation of TGFa gene expression
(23, 42); (b) in contrast to normal mouse TGFa, which is detected asa single 4.5-kilobase transcript, we have identified two additional
TGFa species, 4.7 and 5.2 kilobases long, in RNA isolated fromcultured wa-l keratinocytes and tissues (Figs. 4 and 5); (c) in addition
to the quantitative changes in expression of TGFa mRNA, the level ofimmunoreactive TGFa produced by wa-l v-raiHa keratinocytes ap
pears to be disproportionately low relative to the amount of mRNApresent, suggesting an abnormality at the protein level as well. Ourfindings differ from those reported in a previous study, in which onlya single 4.5-kilobase TGFa transcript was detected in RNA isolatedfrom wa-l brain (42). However, since the probe used in the earlier
report was a partial TGFa cDNA encoding 2.0 kilobases of exon 6, itis conceivable that we are detecting transcript variants hybridizing tofull-length cDNA that are missing this region of the normal TGFatranscript. Studies aimed at identifying the nature of the wa-l muta
tion should provide an explanation for the complex alteration in TGFaexpression observed in these cells.
Our findings suggest that another EGFR ligand may be capable ofsubstituting for TGFa in autocrine growth stimulation of ras-trans-
formed keratinocytes. Consistent with this hypothesis, amphiregulinhas been shown to act as an autocrine growth regulator in raj-
transformed human mammary epithelial cells (57) and a colon carcinoma cell line (58) in vitro. However, the potential autocrine functionof HB-EGF or betacellulin, both of which are up-regulated by v-rasHa
in vitro (Figs. 4 and 6) and in vivo (Fig. 9), has not been previouslyaddressed in neoplastic keratinocytes. We have recently reported thatCM from v-rasHa keratinocyte cultures induces phenotypic changes in
normal cells mimicking those observed in ras-transformed cells.
Moreover, these alterations are blocked using a neutralizing TGFaantibody (11), indicating that TGFa is the principal secreted factorresponsible for v-raiHa effects mediated through the EGFR. The
results presented in this report suggest that in cultures of TGFa —/—v-ra.ïHakeratinocytes, other EGFR ligands may be capable of induc
ing responses previously attributed solely to TGFa. We are currentlyassessing the overall importance of the EGFR signaling pathway in
1891
on July 18, 2018. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

EOFR LÕGANOS AND EPIDERMAL NEOPLASIA
normal keratinocyte biology and neoplasia by disrupting EGFR function using antisense cDNA and a dominant-negative mutant (59, 60).
Simultaneous induction of mRNA encoding four EGFR ligands inepidermal keratinocytes raises questions regarding the role of thesegrowth factors, both individually and collectively, in epidermal ho-
meostasis and disease. Despite certain similarities, these growth factors exhibit distinct biochemical differences that may influence theirfunction and localization both in vitro and in vivo; for example,amphiregulin, HB-EGF, and betacellulin bind heparin, whereas ma
ture TO Fa does not (21, 61). Furthermore, although heparin blocksthe mitogenic response of cultured keratinocytes to amphiregulin (62),it enhances HB-EGF-induced proliferation (63) and migration (64)
but appears to have no effect on cell growth stimulated by betacellulin(65). Interaction with heparan sulfate proteoglycans in vivo may havesimilar functional consequences and could also provide a means ofrestricting activity of individual growth factors to specific sites withina tissue, e.g., during wound-healing.
Several studies have explored the expression of EGFR ligands inepidermis and/or cultured keratinocytes. TGFo (66), amphiregulin(62), and HB-EGF (67) are expressed by cultured human epidermal
keratinocytes. In addition, TGFa is present in human epidermis atrelatively low levels (66, 68), and HB-EGF mRNA has been detectedby RT-PCR in RNA isolated from skin (69). Up-regulation of TGFa
and amphiregulin mRNA, as well as TGFa protein, has been described in the hyperproliferative skin disease psoriasis (68, 70, 71),and elevated expression of TGFa protein has been detected in severaltypes of human epidermal tumors (68). Moreover, HB-EGF was
identified in wound fluid obtained from a porcine skin experimentalmodel (63). Expression of the most recently described EOF familymember, betacellulin, has not been reported previously in skin orcultured keratinocytes; however, the preponderance of betacellulinmRNA in newborn mouse epidermis (Fig. 9) suggests that it is a majorEGFR ligand produced by this tissue. Additional studies to determinethe abundance and distribution of EGFR ligands, in both newborn andadult mouse skin, may provide clues into the normal role of thesegrowth factors in cutaneous biology, and help explain the selectivealteration of hair follicle function observed in both wa-\ and TGFa
null mice.
ACKNOWLEDGMENTS
The authors are grateful to Dr. Rolf Kemler for K8 antibody, Dr. WendyWeinberg for providing the C57rp cell line, Drs. Kay Gray Nelson andCaroline Stanwell for PL-7 cDNA, and Dr. Glen Merlino for critical reading of
the manuscript. The technical assistance of David Morgan during skin graftingstudies was greatly appreciated, as was animal breeding performed by MarthaShaver, Karen Baxley, Susan Kirchhoff, and Linda Sly (Biocon, Inc.,Rockville, MD).
REFERENCES
1. Yuspa, S. H. The pathogènes»of squamous cell cancer: lessons learned from studiesof skin carcinogenesis—Thirty-third G. H. A. Clowes Memorial Award lecture.Cancer Res., 54: 1178-1189, 1994.
2. Yuspa, S. H., and Dlugosz, A. A. Cutaneous carcinogenesis: natural and experimental. In: L. Goldsmilh (ed.). Physiology, Biochemistry, and Molecular Biology of theSkin. pp. 1365-1402. New York: Oxford University Press, 1991.
3. Balmain. A., and Brown, K. Oncogene activation and chemical carcinogenesis. Adv.Cancer Res., 5/: 147-182. 1988.
4. Brown. K., Quintanilla, M., Ramsden. M., Kerr, 1. B., Young, S., and Balmain, A.v-ruj genes from harvey and BALB murine sarcoma viruses can act as initiators oftwo-stage mouse skin carcinogenesis. Cell, 46: 447-456, 1986.
5. Roop, D. R., Lowy, D. R.. Tambourin, P. E., Strickland, ¡., Harper, J. R.,Balaschak, M.. Spangler, E. F.. and Yuspa, S. H. An activated Harvey rasoncogene produces benign tumours on mouse epidermal tissue. Nature (Lond.),323: 822-824, 1986.
6. Conti, C. J. Mutations of genes of the ras family in human and experimental tumors.Prog. Clin. Biol. Res., 376: 357-378, 1992.
7.
8.
9.
10.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
Barbacid, M. ras oncogenes: their role in neoplasia. Eur. J. Clin. Invest., 20: 225-235,1990.Derynck, R. The physiology of transforming growth fac!or-a. Adv. Cancer Res., 58:27-52, 1992.Lee, D. C, Luetteke, N. C, and Petch, L. A. Transforming growth factor-« and itsrole in neoplaslic progression. Cancer Treat. Res., 6.?: 233-254, 1992.Click, A. B., Sporn, M. B., and Yuspa, S. H. Altered regulation of TGF-ßl andTGF-a in primary keratinocytes and papillomas expressing v-Ha-ras. Mol. Carcinog.,4: 210-219, 1991.
Cheng, C., Tennenbaum, T., Dempsey, P. J., Coffey, R. J., Yuspa, S. H., and Dlugosz.A. A. Epidermal growth factor receptor ligands regulate keratin 8 expression inkeratinocytes, and transforming growth factor or mediates the induction of keratin 8by the v-ra.v-Ha oncogene. Cell Growth & Differ., 4: 317-327, 1993.
Hashimoto, Y., Tajima, O., Hashiba, H., Nose, K., and Kuroki, T. Elevated expressionof secondary, but not early, responding genes to phorbol ester tumor promoters inpapillomas and carcinomas of mouse skin. Mol. Carcinog., 3: 302-308, 1990.
Imamoto, A., Beltran, L. M., and DiGiovanni, J. Evidence for autocrine/paracrinegrowth stimulation by transforming growth factor-a during the process of skin tumorpromotion. Mol. Carcinog., 4: 52-60, 1991.Rho, O., Beltran, L. M., Giminez-Conti, I. B., and DiGiovanni, J. Alteredexpression of the epidermal growth factor receptor and transforming growthfactor-a during multistage skin carcinogenesis in SENCAR mice. Mol. Carcinog.,//.- 19-28, 1994.
Vassar, R.. Hutton, M. E., and Fuchs, E. Transgenic overexpression of transforminggrowth factor a bypasses the need for c-Ha-rai mutations in mouse skin tumorigen-esis. Mol. Cell. Biol., 12: 4643-4653, 1992.
Wang, X.. Greenhalgh, D. A., Eckhardt, J. N., Rothnagel, J. A., and Roop, D. R.Epidermal expression of transforming growth factor-a in Iransgenic mice: inductionof spontaneous and 12-O-tetradecanoyl-phorbol-13-acetate-induced papillomas via amechanism independent of Ha-ras activation of overexpression. Mol. Carcinog., 10:15-22, 1994.
Pinzi, E., Kilkenny, A., Strickland, J. E., Balaschak, M., Bringman, T., Derynck, R.,Aaronson, S., and Yuspa, S. H. TGF a stimulates growth of skin papillomas byautocrine and paracrine mechanisms but does not cause neoplastic progression. Mol.Carcinog., /: 7-12, 1988.
Jhappan, C., Takayama. H., Dickson, R. B., and Merlino, G. Transgenic mice providegenetic evidence that transforming growth factor a promotes skin tumorigenesis viaH-raj-dependent and H-ra.v-independent pathways. Cell Growth & Differ., 5:385-394, 1994.
Shoyab, M., Plowman, G. D., McDonald, V. L., Bradley, J. G., and Todaro, G. J.Structure and function of human amphiregulin: a member of the epidermal growthfactor family. Science (Washington DC). 243: 1074-1076, 1989.Higashiyama. S., Abraham, J. A., Miller, J., Fiddes, J. C., and Klagsbrun, M. Aheparin-binding growth factor secreted by macrophage-like cells that is related toEOF. Science (Washington DC), 251: 936-939, 1991.
Shing, Y., Christofori, G., Hanahan, D., Ono, Y., Sasada, R., Igarashi. K., andFolkman. J. Betacellulin: a mitogen from pancreatic ßcell tumors. Science(Washington DC), 259. 1604-1607, 1993.
Crew, F. A. Waved: An autosomal recessive coat form in the mouse. J. Genet., 27:95-96, 1933.Mann, G. B., Fowler, K. J., Gabriel, A., Nice, E. C., Williams, R. L., and Dunn, A. R.Mice with a null mutation of the TGF a gene have abnormal skin architecture, wavyhair, and curly whiskers and often develop corneal inflammation. Cell, 73: 249-261,
1993.Hennings, H., Michael, D., Cheng, C., Steinert, P., Holbrook, K. A., and Yuspa, S. H.Calcium regulation of growth and differentiation of mouse epidermal cells in culture.Cell, 19: 245-254, 1980.Kawamura, H., Strickland, J. E., and Yuspa, S. H. Association of resistance toterminal differentiation with initiation of carcinogenesis in adult mouse epidermalcells. Cancer Res., 45: 2748-2752, 1985.
Yuspa, S. H.. Kilkenny, A. E., Steinert, P. M., and Roop, D. R. Expression of murineepidermal differentiation markers is tightly regulated by restricted extracellular calcium concentrations in vitro. J. Cell Biol., 109: 1207-1217, 1989.Strickland, J. E.. Greenhalgh, D. A., Koceva-Chyla, A., Hennings, H., Restrepo, C.,Balaschak, M., and Yuspa, S. H. Development of murine epidermal cell lines whichcontain an activated rasHa oncogene and form papillomas in skin grafts on athymicnude mouse hosts. Cancer Res., 48: 165-169, 1988.Strickland, J. E.. Dlugosz, A. A., Hennings, H., and Yuspa, S. H. Inhibition of tumorformation from grafted murine papilloma cells by treatment of grafts with stauro-sporine, an inducer of squamous differentiation. Carcinogenesis (Lond.), 14:205-209, 1993.
Russell, W. E., Dempsey, P. J., Sitaric, S., Peck, A. J., and Coffey, R. J., Jr.Transforming growth factor-a (TGFa) concentrations increase in regenerating ratliver: evidence for a delayed accumulation of mature TGFa. Endocrinology, 133:1731-1738, 1993.
Roop, D. R., Cheng, C. K., Titterington, L., Meyers, C. A., Stanley, J. R., Steinert,P. M., and Yuspa, S. H. Synthetic pcptides corresponding to keratin subunits elicithighly specific antibodies. J. Biol. Chem., 259: 8037-8040, 1984.
Brulet. P., Babinet, C., Kemler, R., and Jacob, F. Monoclonal antibodies againsttrophectoderm-specific markers during mouse blastocyst formation. Proc. Nati. Acad.Sci. USA, 77: 4113-4117, 1980.Salinovich, O.. and Montelaro, R. C. Reversible staining and peptide mapping ofproteins transferred to nitrocellulose after separation by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Anal. Biochem., ¡56:341-347, 1986.
Badley, J. E.. Bishop, G. A., St. John, T., and Frelinger, J. A. A simple, rapid methodfor the purification of poly A+ RNA. Biotechnology, 6: 114-116, 1988.
1892
on July 18, 2018. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

EOFR LÕGANOS AND EPIDERMAL NEOPLASIA
34. Chomczynski, P., and Sacchi, N. Single-step method of RNA isolation by acidguanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem., 762:156-159, 1987.
35. Davis, L. G., Dibner, M. D., and Battey, J. F. Basic Methods in Molecular Biology.New York: Elsevier Science Publishing Co., Inc., 1986.
36. Lee, D. C, Rose, T. M., Webb, N. R., and Todaro, G. J. Cloning and sequenceanalysis of a cDNA for rat transforming growth factor-a. Nature (Lond.), 313:489-491, 1985.
37. Fort, P., Marty, L., Piechaczyk, M., el Sabrouty, S., Dani, C., Jeanteur, P., andBlanchard, J. M. Various rat adult tissues express only one major mRNA species fromthe glyceraldehyde-3-phosphate-dehydrogenase multigenic family. Nucleic AcidsRes., 13: 1431-1442, 1985.
38. Barnard, J. A., Graves-Deal, R., Pittelkow, M. R., DuBois, R., Cook, P., Ramsey,G. W., Bishop, P. R., Damstrup, L., and Coffey, R. J. Auto- and cross-inductionwithin the mammalian epidermal growth factor-related peptide family. J. Biol. ( 'hem..
269: 22817-22822, 1994.
39. Meyuhas, O., and Klein, A. The mouse ribosomal protein L7 gene. J. Biol. Chem.,205.- 11465-11473, 1990.
40. Click, A. B., Lee, M. M., Darwiche, N., Kulkarni, A. B., Karlsson, S., and Yuspa,S. H. Targeted deletion of the TGF-ß1 gene causes rapid progression to squamouscell carcinoma. Genes Dev., 8: 2429-2440, 1994.
41. Young, W. S. I. Analysis of neuronal microcircuits and synaptic interactions. In: A.Bjorklund, T. Horkfelt, F. G. Wouterlood, and A. A. Van den Pol (eds.), Handbookof Chemical Neuroanatomy, Vol. 8, pp. 481-512. Amsterdam: Elsevier Science
Publishers, BV, 1990.42. Luetteke, N. C., Qiu, T. H., Peiffer, R. L., Oliver, P., Smithies, 0., and Lee, D. C.
TGF a deficiency results in hair follicle and eye abnormalities in targeted andwaved-1 mice. Cell, 73: 263-278, 1993.
43. Dlugosz, A. A., Cheng, C., Denning, M. F., Dempsey, P. J., Coffey, R. J., Jr., andYuspa, S. H. Keratinocyte growth factor-receptor ligands induce TGFa expressionand activate the epidermal growth factor-receptor signaling pathway in culturedepidermal keratinocytes. Cell Growth & Differ., J: 1283-1292, 1994.
44. Gullick, W. J. Prevalence of aberrant expression of the epidermal growth factorreceptor in human cancers. Br. Med. Bull., 47: 87-98, 1991.
45. Modjtahedi, H., and Dean, C. The receptor for EGF and its tigands: expression,prognostic value and target for therapy in cancer (review). Int. J. Oncol., 4: 277-296,
1994.46. Matsui, Y., Halter, S. A., Holt, J. T., Hogan, B. L., and Coffey, R. J. Development of
mammary hyperplasia and neoplasia in MMTV-TGF a transgenic mice. Cell, 67:1147-1155, 1990.
47. Sandgren, E. P., Luetteke, N. C, Palmiter, R. D., Brinster, R. L., and Lee, D. C.Overexpression of TGF-or in transgenic mice: induction of epithelial hyperplasia, pancreatic metaplasia, and carcinoma of the breast. Cell, 61: 1121-1135,
1990.48. Sandgren, E. P., Luetteke, N. C., Qiu, T. H., Palmiter, R. D., Brinster, R. L., and Lee,
D. C. Transforming growth factor a dramatically enhances oncogene-induced carci-nogenesis in transgenic mouse pancreas and liver. Mol. Cell. Biol., 13: 320-330,1993.
49. Jhappan, C., Stahle, C., Harkins, R. N., Fausto, N., Smith, G. H., and Merlino, G. T.TGFa Overexpression in transgenic mice induces liver neoplasia and abnormaldevelopment of the mammary gland and pancreas. Cell, 61: 1137-1146, 1990.
50. Sizeland, A. M., and Burgess, A. W. Anti-sense transforming growth factor a
oligonucleotides inhibit autocrine stimulated proliferation of a colon carcinoma cellline. Mol. Biol. Cell, 3: 1235-1243, 1992.
51. Ziober, B. L., Willson, J. K., Hymphrey, L. E., Childress-Fields, K., and Brattain,M. G. Autocrine transforming growth factor-a is associated with progression oftransformed properties in human colon cancer cells. J. Biol. Chem., 268: 691-698,
1993.52. Morishige, K., Kurachi, H., Amemiya, K., Fujita, Y., Yamamoto, T., Miyaké,A., and
Tanizawa, O. Evidence for the involvement of transforming growth factor a andepidermal growth factor receptor autocrine growth mechanism in primary humanovarian cancers in viiro. Cancer Res., 51: 5322-5328, 1991.
53. Morishige, K., Kurachi, H., Amemiya, K., Adachi, H., Inoue, M., Miyaké,A.,Tanizawa, O., and Sakoyama, Y. Involvement of transforming growth factor a/epidermal growth factor receptor autocrine growth mechanism in an ovarian cancer cellline in vitro. Cancer Res., 51: 5951-5955, 1991.
54. Kurachi, H., Morishige, K., Amemiya, K., Adachi, H., Hirota, K., Miyaké,A., andTanizawa, O. Importance of transforming growth factor a/epidermal growth factorreceptor autocrine growth mechanism in an ovarian cancer cell line in vivo. CancerRes., 51: 5956-5959, 1991.
55. Mendelsohn, J. Epidermal growth factor receptor as a target for therapy with antireceptor monoclonal antibodies. Nati. Cancer Inst. Monogr., 125-131, 1992.
56. Hennings, H., Glick, A. B., Lowry, D. T., Krsmanovic, L. S., Sly, L. M., and Yuspa,S. H. FVB/N mice: an inbred strain sensitive to the chemical induction of squamouscell carcinomas in the skin. Carcinogenesis (Lond.), 14: 2353-2358, 1993.
57. Normanno, N., Selvam, M. P., Oi, C. F., Saeki, T., Johnson, G., Kim, N., Ciardiello,F., Shoyab, M., Plowman, G., and Brandt, R. Amphiregulin as an autocrine growthfactor for c-Ha-ros- and c-erfcS-2-transformed human mammary epithelial cells. Proc.Nati. Acad. Sci. USA, 91: 2790-2794, 1994.
58. Johnson, G. R., Saeki, T., Gordon, A. W., Shoyab, M., Salomon, D. S., andStromberg, K. Autocrine action of amphiregulin in a colon carcinoma cell lineand immunocytochemical localization of amphiregulin in human colon. J. Cell Biol.,118: 741-751, 1992.
59. Moroni, M. C., Willingham, M. C., and Beguinot, L. EGF-R antisense RNA blocksexpression of the epidermal growth factor receptor and suppresses the transformingphenotype of a human carcinoma cell line. J. Biol. Chem., 267: 2714-2722, 1992.
60. Wu, J. X., and Adamson, E. D. Inhibition of differentiation in P19 embryonalcarcinoma cells by the expression of vectors encoding truncated or antisense EGFreceptor. Dev. Biol., 159: 208-222, 1993.
61. Prigent, S. A., and Lemoine, N. R. The type 1 (EGFR-related) family of growth factorreceptors and their ligands. Prog. Growth Factor Res., 4: 1—24,1992.
62. Cook, P. W., Mattox, P. A., Keeble, W. W., Pittelkow, M. R., Plowman, G. D.,Shoyab, M., Adelman, J. P., and Shipley, G. D. A heparin sulfate-regulated humankeratinocyte autocrine factor is similar or identical to amphiregulin. Mol. Cell. Biol.,//: 2547-2557, 1991.
63. Marikovsky, M., Breuing, K., Liu, P. Y., Eriksson, E., Higashiyama, S., Farber, P.,Abraham, J., and Klagsbrun, M. Appearance of heparin-binding EGF-like growth
factor in wound fluid as a response to injury. Proc. Nati. Acad. Sci. USA, 90:3889-3893, 1993.
64. Higashiyama, S., Abraham, J. A., and Klagsbrun, M. Heparin-binding EGF-likegrowth factor stimulation of smooth muscle cell migration: dependence on interactions with cell surface heparan sulfate. J. Cell Biol., 722: 933-940, 1993.
65. Watanabe, T., Shintani, A., Nakata, M., Shing, Y., Folkman, J., Igarashi, K., andSasada, R. Recombinant human betacellulin. Molecular structure, biological activities, and receptor interaction. J. Biol. Chem., 269: 9966-9973, 1994.
66. Coffey, R. J., Jr., Derynck, R., Wilcox, J. N., Bringman, T. S., Goustin, A. S., Moses,H. L., and Pittelkow, M. R. Production and auto-induction of transforming growthfactor-a in human keratinocytes. Nature (Lond.), 328: 817-820, 1987.
67. Hashimoto, K., Higashiyama, S., Asada, H., Hashimura, E., Kobayashi, T., Sudo, K.,Nakagawa, T., Damm, D., Yoshikawa, K., and Taniguchi. N. Heparin-binding epidermal growth factor-like growth factor is an autocrine growth factor for humankeratinocytes. J. Biol. Chem., 269: 20060-20066, 1994.
68. Gottlieb, A. B., Chang, C. K., Posnett, D. N., Fanelli, B., and Tarn, J. P. Detection oftransforming growth factor a in normal, malignant and hyperproliferative humankeratinocytes. J. Exp. Med., 167: 670-675, 1988.
69. Vaughan, T. J., Pascal!, J. C., and Brown, K. D. Tissue distribution of mRNA forheparin-binding epidermal growth factor. Biochem. J., 287: 681-684, 1992.
70. Elder, J. T., Fisher, G. J., Lindquist, P. B., Bennett, G. L., Pittelkow, M. R., Coffey,R. J., Jr., Ellingsworth, L., Derynck, R., and Voorhees, J. J. Overexpression oftransforming growth factor a in psoriatic epidermis. Science (Washington DC), 243:811-814, 1989.
71. Cook, P. W., Pittelkow, M. R., Keeble, W. W., Graves-Deal, R., Coffey, R. J., Jr., andShipley, G. D. Amphiregulin messenger RNA is elevated in psoriatic epidermis andgastrointestinal carcinomas. Cancer Res., 52: 3224-3227, 1992.
1893
on July 18, 2018. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1995;55:1883-1893. Cancer Res Andrzej A. Dlugosz, Christina Cheng, Erin K. Williams, et al. Alternate Epidermal Growth Factor Receptor Ligands
-induced Epidermal Neoplasia: Potential Involvement ofHaras Is Dispensible for v-αAutocrine Transforming Growth Factor
Updated version
http://cancerres.aacrjournals.org/content/55/9/1883
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/55/9/1883To request permission to re-use all or part of this article, use this link
on July 18, 2018. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

![RASHA Advisory And Consultancy Services Pvt. Ltd ... · RASHA Advisory And Consultancy Services Pvt. Ltd. Franchise Proposal [Business Advisory Services] Contact Us: - +91-8826983883](https://static.fdocuments.us/doc/165x107/5cc3cefe88c99343558d9968/rasha-advisory-and-consultancy-services-pvt-ltd-rasha-advisory-and-consultancy.jpg)






![5 VIG Alison & Stan [Kompatibilitätsmodus] · Ms Rasha ElMs Rasha El-Khadem AU ... 12 commercial cabbage lots (1 cv): ... 5 VIG Alison & Stan [Kompatibilitätsmodus] Author:](https://static.fdocuments.us/doc/165x107/5b4fc92c7f8b9a256e8cf9de/5-vig-alison-stan-kompatibilitaetsmodus-ms-rasha-elms-rasha-el-khadem-au.jpg)








