
The SH-3 and SH-4 Antibodies Recognize Distinct Epitopes on CD73 from Human Mesenchymal Stem Cells
6
The SH-3 and SH-4 Antibodies Recognize Distinct Epitopes on CD73 from Human Mesenchymal Stem Cells Frank Barry, 1 Raymond Boynton, Mary Murphy, and Joseph Zaia Osiris Therapeutics, Inc., Baltimore, Maryland 21231 Received October 18, 2001 Mesenchymal stem cells (MSCs) are pluripotent cells in the bone marrow that have the capacity to differ- entiate along a number of connective tissue lineages, including cartilage, bone, adipose tissue, and stroma. The SH-3 and SH-4 monoclonal antibodies recognize epitopes present on the surface of human MSCs. This study describes the isolation and characterization of the antigen that is recognized by these antibodies. A protein of molecular weight approximately 67 kDa was immunoprecipitated from a solubilized mem- brane preparation of human MSCs using the SH-3 an- tibody. Analysis of peptides derived from this protein by mass spectrometry and sequencing identified it as CD73 (ecto-5*-nucleotidase). The SH-4 antibody was also shown to react with purified bovine CD73 by im- munoblotting, but the SH-3 antibody failed to react with the bovine protein. These results indicate that both SH-3 and SH-4 epitopes are present on CD73, but they are distinct. CD73, present in lymphoid tissue, plays a role in the activation of B-lymphocytes and in signal transduction in the hematopoietic compart- ment of bone marrow. The role that CD73 may play in bone marrow stromal interactions and in the differen- tiation of MSCs is discussed. © 2001 Elsevier Science Mesenchymal stem cells (MSCs) have the capacity to differentiate along a number of lineages including the chondrogenic (1, 2), osteogenic (3), and adipogenic path- ways (4). This population of cells is maintained through- out adulthood and provide a source of progenitor cells during tissue turnover and repair. MSCs may be easily isolated from bone marrow aspirates (5) and expanded in culture through many generations without loss of the capacity to differentiate. Therefore they may potentially be used in therapeutic applications and a number of approaches have been described for the use of MSCs in cell therapy and tissue engineering (6 –10). Several monoclonal antibodies have been raised in an effort to provide reagents for the characterization and isolation of cells found in the human marrow stro- mal compartment. STRO-1 was identified as an anti- body that reacted with nonhematopoietic progenitor bone marrow stromal cells (11). The SB-10 antibody (12) was shown to be reactive with an antigen present on undifferentiated MSCs which disappeared once the cells embarked upon an osteogenic pathway and began to express cell surface alkaline phosphatase. The spe- cific SB-10 antigen was identified as CD166 (activated leukocyte-cell adhesion molecule, ALCAM (13)), which may play a role in the progression of osteogenic differ- entiation, although the precise mechanism of this pro- cess remains to be elucidated. The SH-2 antibody (14), also raised against human MSCs, reacts with an epitope present on CD105 (15). The SH-3 and SH-4 antibodies also recognize epitope(s) present on MSCs (14). As was reported with SH-2, these antibodies do not react with hematopoietic cells or with osteocytes (14). This study describes the immunoprecipitation of a protein of molecular weight approximately 67 kDa from a solubilized human MSC membrane fraction. Peptides derived from the purified protein were char- acterized by matrix-assisted laser desorption ioniza- tion time-of-flight (MALDI-TOF) mass spectrometry. Analysis of these peptide mass values indicated that the epitope recognized by the SH-3 antibody was present on CD73 (ecto-59-nucleotidase). This was con- firmed by amino acid sequence analysis. The SH-4 antibody was then found to react with purified bovine CD73 by immunoblotting, indicating that this epitope was present on the same protein. This finding provides useful information on the application of these antibod- ies in the isolation and characterization of MSCs and the possible role that the antigen may play in the regulation of differentiation. MATERIALS AND METHODS Cell culture and processing. MSCs were isolated from bone mar- row aspirates taken from normal adult human donors and culture- expanded using established procedures (4, 5). Cells from 3 donors 1 To whom correspondence and reprint requests should be ad- dressed at Osiris Therapeutics Inc., 2001 Aliceanna Street, Balti- more, MD 21231. Fax: 410-522-6999. E-mail: [email protected]. Biochemical and Biophysical Research Communications 289, 519 –524 (2001) doi:10.1006/bbrc.2001.6013, available online at http://www.idealibrary.com on 519 0006-291X/01 $35.00 © 2001 Elsevier Science All rights reserved.
-
Upload
frank-barry -
Category
Documents
-
view
213 -
download
0
Transcript of The SH-3 and SH-4 Antibodies Recognize Distinct Epitopes on CD73 from Human Mesenchymal Stem Cells

Biochemical and Biophysical Research Communications 289, 519–524 (2001)
doi:10.1006/bbrc.2001.6013, available online at http://www.idealibrary.com on
The SH-3 and SH-4 Antibodies Recognize DistinctEpitopes on CD73 from Human Mesenchymal Stem Cells
Frank Barry,1 Raymond Boynton, Mary Murphy, and Joseph ZaiaOsiris Therapeutics, Inc., Baltimore, Maryland 21231
Received October 18, 2001
Several monoclonal antibodies have been raised in
Mesenchymal stem cells (MSCs) are pluripotent cellsin the bone marrow that have the capacity to differ-entiate along a number of connective tissue lineages,including cartilage, bone, adipose tissue, and stroma.The SH-3 and SH-4 monoclonal antibodies recognizeepitopes present on the surface of human MSCs. Thisstudy describes the isolation and characterization ofthe antigen that is recognized by these antibodies. Aprotein of molecular weight approximately 67 kDawas immunoprecipitated from a solubilized mem-brane preparation of human MSCs using the SH-3 an-tibody. Analysis of peptides derived from this proteinby mass spectrometry and sequencing identified it asCD73 (ecto-5*-nucleotidase). The SH-4 antibody wasalso shown to react with purified bovine CD73 by im-munoblotting, but the SH-3 antibody failed to reactwith the bovine protein. These results indicate thatboth SH-3 and SH-4 epitopes are present on CD73, butthey are distinct. CD73, present in lymphoid tissue,plays a role in the activation of B-lymphocytes and insignal transduction in the hematopoietic compart-ment of bone marrow. The role that CD73 may play inbone marrow stromal interactions and in the differen-tiation of MSCs is discussed. © 2001 Elsevier Science
Mesenchymal stem cells (MSCs) have the capacity todifferentiate along a number of lineages including thechondrogenic (1, 2), osteogenic (3), and adipogenic path-ways (4). This population of cells is maintained through-out adulthood and provide a source of progenitor cellsduring tissue turnover and repair. MSCs may be easilyisolated from bone marrow aspirates (5) and expanded inculture through many generations without loss of thecapacity to differentiate. Therefore they may potentiallybe used in therapeutic applications and a number ofapproaches have been described for the use of MSCs incell therapy and tissue engineering (6–10).
1 To whom correspondence and reprint requests should be ad-dressed at Osiris Therapeutics Inc., 2001 Aliceanna Street, Balti-more, MD 21231. Fax: 410-522-6999. E-mail: [email protected].
519
an effort to provide reagents for the characterizationand isolation of cells found in the human marrow stro-mal compartment. STRO-1 was identified as an anti-body that reacted with nonhematopoietic progenitorbone marrow stromal cells (11). The SB-10 antibody(12) was shown to be reactive with an antigen presenton undifferentiated MSCs which disappeared once thecells embarked upon an osteogenic pathway and beganto express cell surface alkaline phosphatase. The spe-cific SB-10 antigen was identified as CD166 (activatedleukocyte-cell adhesion molecule, ALCAM (13)), whichmay play a role in the progression of osteogenic differ-entiation, although the precise mechanism of this pro-cess remains to be elucidated. The SH-2 antibody (14),also raised against human MSCs, reacts with anepitope present on CD105 (15). The SH-3 and SH-4antibodies also recognize epitope(s) present on MSCs(14). As was reported with SH-2, these antibodies donot react with hematopoietic cells or with osteocytes(14). This study describes the immunoprecipitation of aprotein of molecular weight approximately 67 kDafrom a solubilized human MSC membrane fraction.Peptides derived from the purified protein were char-acterized by matrix-assisted laser desorption ioniza-tion time-of-flight (MALDI-TOF) mass spectrometry.Analysis of these peptide mass values indicated thatthe epitope recognized by the SH-3 antibody waspresent on CD73 (ecto-59-nucleotidase). This was con-firmed by amino acid sequence analysis. The SH-4antibody was then found to react with purified bovineCD73 by immunoblotting, indicating that this epitopewas present on the same protein. This finding providesuseful information on the application of these antibod-ies in the isolation and characterization of MSCs andthe possible role that the antigen may play in theregulation of differentiation.
MATERIALS AND METHODS
Cell culture and processing. MSCs were isolated from bone mar-row aspirates taken from normal adult human donors and culture-expanded using established procedures (4, 5). Cells from 3 donors
0006-291X/01 $35.00© 2001 Elsevier ScienceAll rights reserved.
were excised and placed into 1.5 ml microcentrifuge tubes and
Vol. 289, No. 2, 2001 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
were used in these experiments and for each donor a total of 100T-185 cell culture flasks (Costar) were seeded and the cells grown toconfluence under standard culture conditions. Upon reaching conflu-ence the culture medium was removed and 20 ml Tyrode’s saltsolution was added to each flask. The cells were washed three timesfor 10–15 min each with gentle rocking. After washing, 3 ml ice coldlysis buffer (50 mM Tris–Cl, pH 7.4, 20 mM CaCl2, 0.15 M NaCl, and1 mM phenylmethylsulfonyl fluoride) was added to each flask andthe cells were removed by scraping. The cells were disrupted bysonication at 4°C in 3 bursts of 20-s duration at setting 4 on a SonicDismembrator 550 (Fisher Scientific). The cell homogenate was cen-trifuged at 4°C for 30 min at 2000g, the supernatant was collectedand then recentrifuged at 50,000g at 4°C for 60 min. The precipitatedmaterial was placed on ice and ice cold solubilization buffer (20 mMTris–Cl, pH. 8.0, 0.14 M NaCl, 0.5% (v/v) Chaps, 1 mM phenylmeth-ylsulfonyl fluoride) was added. The pooled solubilized materialwas stored at 280°C until sufficient material was accumulated forcharacterization.
Immunoprecipitation. The pooled solubilized membrane fractionderived from approximately 1.5 3 109 cells had a final volume of15–20 ml. The SH-3 antibody was added at a concentration of 10mg/ml and mixed end-over-end at 4°C for 6 h. 550 ml of a 50% slurryof protein G-agarose (Boehringer Mannheim) was added. The proteinG-agarose was pre-equilibrated in solubilization buffer prior to ad-dition. The immunoprecipitation mixture was incubated at 4°C over-night with mixing end-over-end. The protein G-agarose was removedby centrifugation and the supernatant was collected and stored at280°C. The protein G-agarose pellet was washed twice with 25 mlsolubilization buffer.
Elution of the bound antigen and SDS–PAGE purification. SDS–PAGE analysis was carried out using a Novex Electrophoresis sys-tem with 12% separating gel and 4% stacking gel. The washedprotein G-agarose pellet was boiled for 3 min in 250 ml of SDS–PAGEreducing sample buffer to elute the bound antigen, centrifugedbriefly and the supernatant was electrophoresed at a constant volt-age of 100 V until the dye front migrated through the stacking gel atwhich time the voltage was increased to 200 V. The gel was stainedusing a reversible zinc staining procedure (16). Individual bands
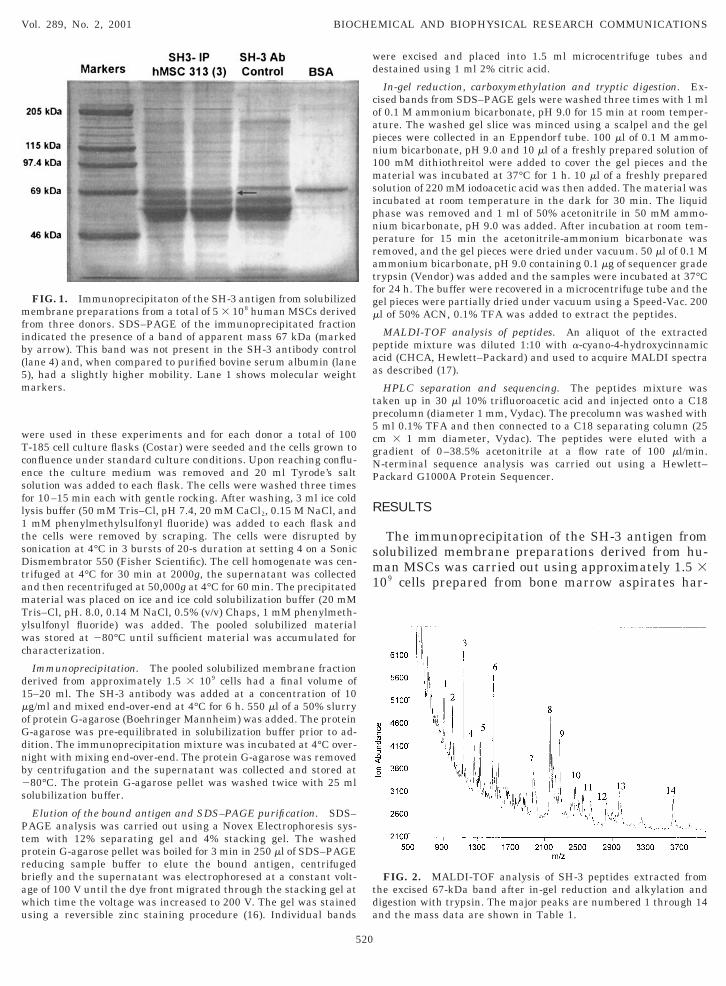
FIG. 1. Immunoprecipitaton of the SH-3 antigen from solubilizedmembrane preparations from a total of 5 3 108 human MSCs derivedfrom three donors. SDS–PAGE of the immunoprecipitated fractionindicated the presence of a band of apparent mass 67 kDa (markedby arrow). This band was not present in the SH-3 antibody control(lane 4) and, when compared to purified bovine serum albumin (lane5), had a slightly higher mobility. Lane 1 shows molecular weightmarkers.
520
destained using 1 ml 2% citric acid.
In-gel reduction, carboxymethylation and tryptic digestion. Ex-cised bands from SDS–PAGE gels were washed three times with 1 mlof 0.1 M ammonium bicarbonate, pH 9.0 for 15 min at room temper-ature. The washed gel slice was minced using a scalpel and the gelpieces were collected in an Eppendorf tube. 100 ml of 0.1 M ammo-nium bicarbonate, pH 9.0 and 10 ml of a freshly prepared solution of100 mM dithiothreitol were added to cover the gel pieces and thematerial was incubated at 37°C for 1 h. 10 ml of a freshly preparedsolution of 220 mM iodoacetic acid was then added. The material wasincubated at room temperature in the dark for 30 min. The liquidphase was removed and 1 ml of 50% acetonitrile in 50 mM ammo-nium bicarbonate, pH 9.0 was added. After incubation at room tem-perature for 15 min the acetonitrile-ammonium bicarbonate wasremoved, and the gel pieces were dried under vacuum. 50 ml of 0.1 Mammonium bicarbonate, pH 9.0 containing 0.1 mg of sequencer gradetrypsin (Vendor) was added and the samples were incubated at 37°Cfor 24 h. The buffer were recovered in a microcentrifuge tube and thegel pieces were partially dried under vacuum using a Speed-Vac. 200ml of 50% ACN, 0.1% TFA was added to extract the peptides.
MALDI-TOF analysis of peptides. An aliquot of the extractedpeptide mixture was diluted 1:10 with a-cyano-4-hydroxycinnamicacid (CHCA, Hewlett–Packard) and used to acquire MALDI spectraas described (17).
HPLC separation and sequencing. The peptides mixture wastaken up in 30 ml 10% trifluoroacetic acid and injected onto a C18precolumn (diameter 1 mm, Vydac). The precolumn was washed with5 ml 0.1% TFA and then connected to a C18 separating column (25cm 3 1 mm diameter, Vydac). The peptides were eluted with agradient of 0–38.5% acetonitrile at a flow rate of 100 ml/min.N-terminal sequence analysis was carried out using a Hewlett–Packard G1000A Protein Sequencer.
RESULTS
The immunoprecipitation of the SH-3 antigen fromsolubilized membrane preparations derived from hu-man MSCs was carried out using approximately 1.5 3109 cells prepared from bone marrow aspirates har-
FIG. 2. MALDI-TOF analysis of SH-3 peptides extracted fromthe excised 67-kDa band after in-gel reduction and alkylation anddigestion with trypsin. The major peaks are numbered 1 through 14and the mass data are shown in Table 1.
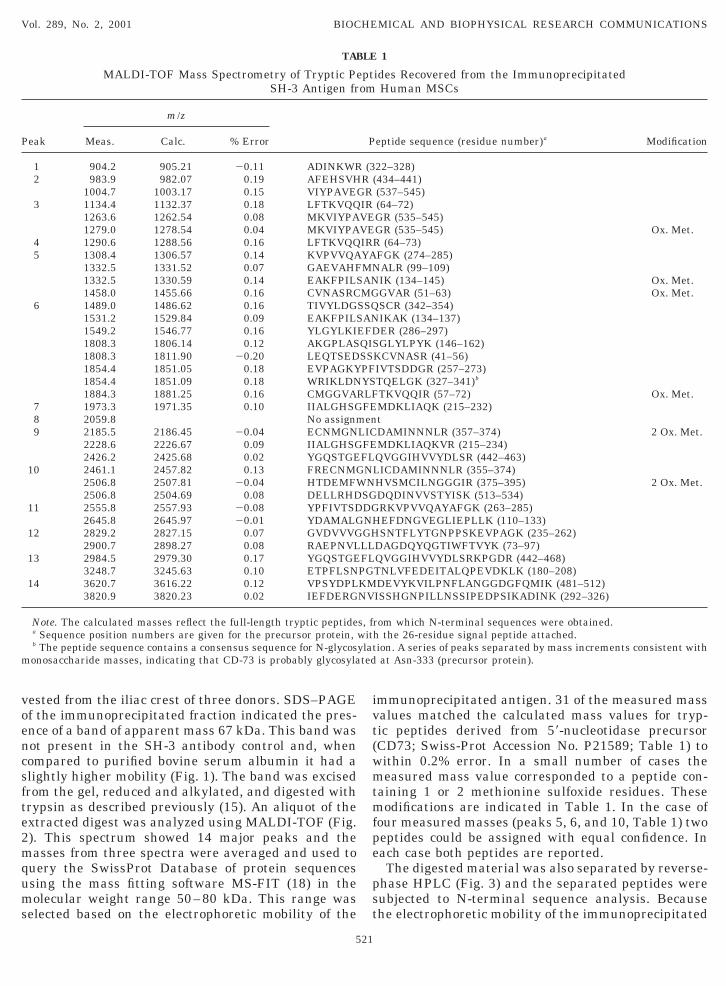
TABLE 1
Vol. 289, No. 2, 2001 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
vested from the iliac crest of three donors. SDS–PAGEof the immunoprecipitated fraction indicated the pres-ence of a band of apparent mass 67 kDa. This band wasnot present in the SH-3 antibody control and, whencompared to purified bovine serum albumin it had aslightly higher mobility (Fig. 1). The band was excisedfrom the gel, reduced and alkylated, and digested withtrypsin as described previously (15). An aliquot of theextracted digest was analyzed using MALDI-TOF (Fig.2). This spectrum showed 14 major peaks and themasses from three spectra were averaged and used toquery the SwissProt Database of protein sequencesusing the mass fitting software MS-FIT (18) in themolecular weight range 50–80 kDa. This range wasselected based on the electrophoretic mobility of the
MALDI-TOF Mass Spectrometry of Tryptic PSH-3 Antigen fr
Peak
m/z
Meas. Calc. % Error
1 904.2 905.21 20.11 ADINKWR2 983.9 982.07 0.19 AFEHSVH
1004.7 1003.17 0.15 VIYPAVE3 1134.4 1132.37 0.18 LFTKVQQ
1263.6 1262.54 0.08 MKVIYPA1279.0 1278.54 0.04 MKVIYPA
4 1290.6 1288.56 0.16 LFTKVQQ5 1308.4 1306.57 0.14 KVPVVQA
1332.5 1331.52 0.07 GAEVAHF1332.5 1330.59 0.14 EAKFPIL1458.0 1455.66 0.16 CVNASRC
6 1489.0 1486.62 0.16 TIVYLDG1531.2 1529.84 0.09 EAKFPIL1549.2 1546.77 0.16 YLGYLKI1808.3 1806.14 0.12 AKGPLAS1808.3 1811.90 20.20 LEQTSED1854.4 1851.05 0.18 EVPAGKY1854.4 1851.09 0.18 WRIKLDN1884.3 1881.25 0.16 CMGGVA
7 1973.3 1971.35 0.10 IIALGHSG8 2059.8 No assignm9 2185.5 2186.45 20.04 ECNMGN
2228.6 2226.67 0.09 IIALGHSG2426.2 2425.68 0.02 YGQSTGE
10 2461.1 2457.82 0.13 FRECNMG2506.8 2507.81 20.04 HTDEMFW2506.8 2504.69 0.08 DELLRHD
11 2555.8 2557.93 20.08 YPFIVTSD2645.8 2645.97 20.01 YDAMALG
12 2829.2 2827.15 0.07 GVDVVVG2900.7 2898.27 0.08 RAEPNVL
13 2984.5 2979.30 0.17 YGQSTGE3248.7 3245.63 0.10 ETPFLSN
14 3620.7 3616.22 0.12 VPSYDPL3820.9 3820.23 0.02 IEFDERG
Note. The calculated masses reflect the full-length tryptic peptidea Sequence position numbers are given for the precursor protein,b The peptide sequence contains a consensus sequence for N-glycos
monosaccharide masses, indicating that CD-73 is probably glycosyla
521
immunoprecipitated antigen. 31 of the measured massvalues matched the calculated mass values for tryp-tic peptides derived from 59-nucleotidase precursor(CD73; Swiss-Prot Accession No. P21589; Table 1) towithin 0.2% error. In a small number of cases themeasured mass value corresponded to a peptide con-taining 1 or 2 methionine sulfoxide residues. Thesemodifications are indicated in Table 1. In the case offour measured masses (peaks 5, 6, and 10, Table 1) twopeptides could be assigned with equal confidence. Ineach case both peptides are reported.
The digested material was also separated by reverse-phase HPLC (Fig. 3) and the separated peptides weresubjected to N-terminal sequence analysis. Becausethe electrophoretic mobility of the immunoprecipitated
ides Recovered from the ImmunoprecipitatedHuman MSCs
eptide sequence (residue number)a Modification
22–328)(434–441)(537–545)(64–72)GR (535–545)GR (535–545) Ox. Met.R (64–73)FGK (274–285)ALR (99–109)IK (134–145) Ox. Met.GVAR (51–63) Ox. Met.SCR (342–354)IKAK (134–137)ER (286–297)
SGLYLPYK (146–162)KCVNASR (41–56)IVTSDDGR (257–273)TQELGK (327–341)b
TKVQQIR (57–72) Ox. Met.MDKLIAQK (215–232)tDAMINNNLR (357–374) 2 Ox. Met.MDKLIAQKVR (215–234)QVGGIHVVYDLSR (442–463)LICDAMINNNLR (355–374)HVSMCILNGGGIR (375–395) 2 Ox. Met.DQDINVVSTYISK (513–534)RKVPVVQAYAFGK (263–285)
HEFDNGVEGLIEPLLK (110–133)SNTFLYTGNPPSKEVPAGK (235–262)
DAGDQYQGTIWFTVYK (73–97)QVGGIHVVYDLSRKPGDR (442–468)TNLVFEDEITALQPEVDKLK (180–208)DEVYKVILPNFLANGGDGFQMIK (481–512)ISSHGNPILLNSSIPEDPSIKADINK (292–326)
rom which N-terminal sequences were obtained.h the 26-residue signal peptide attached.tion. A series of peaks separated by mass increments consistent with
at Asn-333 (precursor protein).
eptom
P
(3R
GRIRVEVEIRYAMN
SANMG
SSQSANEFDQISSPFYS
RLFFEen
LICFEFLNN
SGDGNGHLLFLPGKMNV
s, fwitylated

TABLE 2
Vol. 289, No. 2, 2001 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
material was close to that of the antibody heavy chain,the gel band corresponding to the latter was also re-duced, alkylated digested with trypsin and subjected toHPLC separation. This chromatogram is also shown(Fig. 3). Seven fractions were collected, labeled A-G,which were present in the immunoprecipitate digestand not in the antibody heavy chain digest. From thesefractions 9 peptide sequences were obtained (Table 2).In every the sequence matched 59-nucleotidase.
Several experiments had suggested that the SH-3and SH-4 antigens shared some similarities in terms ofmolecular mass. In one experiment (data not shown) a
FIG. 3. Reverse-phase capillary HPLC chromatograms of a tryp-tic digest of the excised 67-kDa immunoprecipitated band (top) andof the excised band corresponding to the SH-3 antibody heavy chain(bottom). The eluate peaks labeled A–G were collected and subjectedto N-terminal sequence analysis (Table 2).
522
35S-labeled protein was immunoprecipitated with theSH-4 antibody. The mass of this protein, based onSDS–PAGE was 67 kDa, identical to that of the SH-3antigen. Further studies based in flow cytometry, sug-gested that the two antibodies competed for binding tothe same surface antigen, or else bound to neighboringantigens (data not shown). To determine whether theepitope recognized by the SH-4 antibody might exist onthe same antigen both antibodies were tested for reac-tivity with authentic purified 59-nucleotidase by immu-noblotting. The result of this experiment (Fig. 4) indi-cated that the SH-4 antibody did indeed react withpurified bovine 59-nucleotidase but the SH-3 antibodydid not. When a membrane preparation of humanMSCs was probed with both SH-3 and SH-4 antibodiestwo bands corresponding to proteins of molecularweight approximately 67 and 72 kDa were detected.The SH-3 antibody bound more strongly to the smallerof these and the SH-4 antibody to the larger (Fig. 4).
N-Terminal Sequence Analysis of Peptides Derived fromthe SH-3 Antigen Immunoprecipitated from Human MSCs
PeptideHPLC retention
time (min) Sequence
A 29.67 ADINK (222)B 33.01 IKLDXYSTQE (329)
TXVYL (342)C 33.35 TIVYLDGSSX (342)D 33.98 LFTKVQQIR (64)E 38.8 HDSLDQDINVV (518)
VVKLDVLCTK (469)F 43.80 WR (327)G 45.23 IIALGHSGFEMDKLI (215)
FIG. 4. Reactivity of the SH3 (A) and SH4 (B) antibodies with themembrane extract supernatant (lanes 1 and 4) and precipitate (lane2 and 5) of human MSCs and with purified bovine CD73 (lanes 3and 6). Lanes 4–6 contain a fivefold higher loading compared tolanes 1–3.

DISCUSSION is adenosine it may be involved in the activation of
Vol. 289, No. 2, 2001 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
The results of this study indicate that the SH-3 an-tibody recognizes an epitope present on CD73 or ecto-59-nucleotidase. This is based on analysis of the proteinimmunoprecipitated by the antibody in a solubilizedmembrane fraction derived from human MSCs. Bothmass spectrometric analysis and amino acid sequenc-ing confirmed the identity of the antigen as CD73.
Based on a number of observations the SH-4 anti-body recognizes an epitope present on the same mole-cule. This is based on the following: [1] the SH-3 andSH-4 antigens have identical molecular mass; [2] bind-ing of the two antibodies to the surface of hMSCsapparently occurs in a competitive manner; and [3] theSH-4 antibody binds to purified bovine CD-73 (Fig. 4).The epitopes recognized by both antibodies are likely tobe distinct, however, even though they exist on thesame molecule. The result of the immunoblotting ex-periment indicated that the SH-3 epitope is not con-served between human and bovine CD-73 while theSH-4 epitope is conserved. Bovine and human CD73are 88.9% homologous (19, 20) so it is possible that theantibody binding region on the human molecule is al-tered in the bovine protein. It is also possible that theSH-3 antibody does not bind to a denatured epitope,while the SH-4 epitope is not sensitive to denaturation.Finally it may be possible that the SH-3 antibody maybind to a region in the molecule that may be differen-tially glycosylated in the two species.
Two forms of the molecule were identified by immu-noblotting (Fig. 4) and had molecular mass of 67 and 72kDa respectively. These may represent the soluble andmembrane-bound species previously described (21)which carry several N-linked oligosaccharides (19). Inaddition the mature form of the protein lacks theC-terminal peptide which is replaced by the glyco-sylphosphatidylinositol (GPI) anchor.
Earlier studies using the SH-3 and SH-4 antibodiesindicated that they were specific for bone marrow mes-enchymal stem cells and that they did not react withother marrow components, including hematopoieticcells (14). Furthermore the antibodies were reported tobe unreactive with osteoblasts, and were specific to theprimitive stem cell (14). However, CD73 is widely ex-pressed on many cells. It is distributed in murine lym-phoid tissues and the level of expression increases withlymphocyte maturation (22) As already mentioned it isa glycosylphosphatidylinositol-anchored protein withboth ecto-59-nucleotidase activity and signal transduc-tion capability. It catalyzes the dephosphorylation ofpurine and pyrimidine ribo- and deoxyribonucleosidemonophosphates to the corresponding nucleosides. Inthis capacity it plays a role in the salvage of purinesand pyrimidines, presumably in all cell types where itis expressed. CD73 has the potential to act in cellsignaling and since the product of the enzyme reaction
523
adenosine receptors (23, 24). Thus CD73 may indeedreact with the bone marrow microenvironment tomediate cell-cell interactions and it may representa common element between stromal and lymphocytedevelopment.
REFERENCES
1. Mackay, A. M., Beck, S. C., Murphy, J. M., Barry, F. P., Chich-ester, C. O., and Pittenger, M. F. (1998) Chondrogenic differen-tiation of cultured human mesenchymal stem cells from marrow.Tissue Eng. 4, 415–428.
2. Johnstone, B., Hering, T. M., Caplan, A. I., Goldberg, V. M., andYoo, J. U. (1998) In vitro chondrogenesis of bone marrow-derivedmesenchymal progenitor cells. Exp. Cell Res. 238, 265–272.
3. Jaiswal, N., Haynesworth, S. E., Caplan, A. I., and Bruder, S. P.(1997) Osteogenic differentiation of purified, culture-expandedhuman mesenchymal stem cellsin vitro. J. Cell Biochem. 64,295–312.
4. Pittenger, M. F., Mackay, A. M., Beck, S. C., Jaiswal, R. K.,Douglas, R., Mosca, J. D., Moorman, M. A., Simonetti, D. W.,Craig, S., and Marshak, D. R. (1999) Multilineage potential ofadult human mesenchymal stem cells. Science 284, 143–147.
5. Haynesworth, S. E., Goshima, J., Goldberg, V. M., and Caplan,A. I. (1992) Characterization of cells with osteogenic potentialfrom human marrow. Bone 13, 81–88.
6. Wakitani, S., Goto, T., Pineda, S. J., Young, R. G., Mansour,J. M., Caplan, A. I., and Goldberg, V. M. (1994) Mesenchymalcell-based repair of large, full-thickness defects of articular car-tilage. J. Bone Joint Surg. Am. 76, 579–592.
7. Bruder, S. P., Kurth, A. A., Shea, M., Hayes, W. C., Jaiswal, N.,and Kadiyala, S. (1998) Bone regeneration by implantation ofpurified, culture-expanded human mesenchymal stem cells.J. Orthop. Res. 16, 155–162.
8. Lazarus, H. M., Haynesworth, S. E., Gerson, S. L., Rosenthal,N. S., and Caplan, A. I. (1995) Ex vivo expansion and subsequentinfusion of human bone marrow-derived stromal progenitor cells(mesenchymal progenitor cells): Implications for therapeutic use.Bone Marrow Transplant 16, 557–564.
9. Horwitz, E. M., Prockop, D. J., Fitzpatrick, L. A., Koo, W. W.,Gordon, P. L., Neel, M., Sussman, M., Orchard, P., Marx, J. C.,Pyeritz, R. E., Brenner, M. K. (1999) Transplantability and ther-apeutic effects of bone marrow-derived mesenchymal cells inchildren with osteogenesis imperfecta. Nat. Med. 5, 309–313.
10. Young, R. G., Butler, D. L., Weber, W., Caplan, A. I., Gordon,S. L., and Fink, D. J. (1998) Use of mesenchymal stem cells in acollagen matrix for Achilles tendon repair. J. Orthop. Res. 16,406–413.
11. Simmons, P. J., and Torok-Storb, B. (1991) Identification ofstromal cell precursors in human bone marrow by a novel mono-clonal antibody, STRO-1. Blood 78, 55–62.
12. Bruder, S. P., Horowitz, M. C., Mosca, J. D., Haynesworth, S. E.(1997) Monoclonal antibodies reactive with human osteogeniccell surface antigens. Bone 21, 225–235.
13. Bruder, S. P., Ricalton, N. S., Boynton, R. E., Connolly, T. J.,Jaiswal, N., Zaia, J., and Barry, F. P. (1998) Mesenchymal stemcell surface antigen SB-10 corresponds to activated leukocytecell adhesion molecule and is involved in osteogenic differentia-tion. J. Bone Miner. Res. 13, 655–663.
14. Haynesworth, S. E., Baber, M. A., and Caplan, A. I. (1992) Cellsurface antigens on human marrow-derived mesenchymal cellsare detected by monoclonal antibodies. Bone 13, 69–80.
15. Barry, F. P., Boynton, R. E., Haynesworth, S., Murphy, J. M.,

and Zaia, J. (1999) The monoclonal antibody SH-2, raised Taguchi, R., Nakamura, S., Suzuki, Y., and Ikezawa, H. (1993)
Vol. 289, No. 2, 2001 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
against human mesenchymal stem cells, recognizes an epitopeon endoglin (CD105). Biochem. Biophys. Res. Commun. 265,134–139.
16. Castellanos-Serra, L., Proenza, W., Huerta, V., Moritz, R. L., andSimpson, R. J. (1999) Proteome analysis of polyacrylamide gel-separated proteins visualized by reversible negative stainingusing imidazole-zinc salts. Electrophoresis 20, 732–737.
17. Zaia, J., Liu, B., Boynton, R., and Barry, F. (2000) Structuralanalysis of cartilage proteoglycans and glycoproteins usingmatrix-assisted laser desorption/ionization time-of-flight massspectrometry. Anal. Biochem. 277, 94–103.
18. Baker, P. R., and Clauser, K. R. http://prospector.ucsf.com.19. Misumi, Y., Ogata, S., Ohkubo, K., Hirose, S., Ikehara, Y. (1990)
Primary structure of human placental 59-nucleotidase and iden-tification of the glycolipid anchor in the mature form. Eur. J. Bio-chem. 191, 563–569.
20. Suzuki, K., Furukawa, Y., Tamura, H., Ejiri, N., Suematsu, H.,
524
Purification and cDNA cloning of bovine liver 59-nucleotidase, aGPI-anchored protein, and its expression in COS cells. J. Bio-chem. (Tokyo) 113, 607–613.
21. Klemens, M. R., Sherman, W. R., Holmberg, N. J., Ruedi, J. M.,Low, M. G., and Thompson, L. F. (1990) Characterization ofsoluble vs membrane-bound human placental 59-nucleotidase.Biochem. Biophys. Res. Commun. 172, 1371–1377.
22. Yamashita, Y., Hooker, S. W., Jiang, H., Laurent, A. B., Resta,R., Khare, K., Coe, A., Kincade, P. W., and Thompson, L. F.(1998) Eur. J. Immunol. 28, 2981–2990.
23. Resta, R., Yamashita, Y., and Thompson, L. F. (1998) Ecto-enzyme and signaling functions of lymphocyte CD73. Immunol.Rev. 161, 95–109.
24. Resta, R., Jiang, H., Hooker, S. W., Laurent, A. B., and Thomp-son, L. F. (1998) Insights into adenosine deaminase deficiencyprovided by murine fetal thymic organ culture with 29-deoxy-coformycin. Adv. Exp. Med. Biol. 431, 451–454.





![Phosphorylation-dependentepitopes antibodies Alzheimertau · ment antibodies SMI31, SMI34, SMI35, or SMI310 (with phosphorylated epitopes) and SM133 [unphosphorylated epitopes (3)].](https://static.fdocuments.us/doc/165x107/5e62d2f4d3d32f22a55ed9e3/phosphorylation-dependentepitopes-antibodies-alzheimertau-ment-antibodies-smi31.jpg)












