
The FGF-BMP Signaling Axis Regulates Outflow Tract Valve...
20
The FGF-BMP Signaling Axis Regulates Outflow Tract Valve Primordium Formation by Promoting Cushion Neural Crest Cell Differentiation Jue Zhang, Julia Y.F. Chang, Yanqing Huang, Xiang Lin, Yongde Luo, Robert J. Schwartz, James F. Martin, Fen Wang Rationale: Heart valves develop from precursor structures called cardiac cushions, an endothelial-lined cardiac jelly that resides in the inner side of the heart tube. The cushions are then invaded by cells from different sources, undergo a series of complicated and poorly understood remodeling processes, and give rise to valves. Disruption of the fibroblast growth factor (FGF) signaling axis impairs morphogenesis of the outflow tract (OFT). Yet, whether FGF signaling regulates OFT valve formation is unknown. Objective: To study how OFT valve formation is regulated and how aberrant cell signaling causes valve defects. Methods and Results: By using mouse genetic manipulation, cell lineage tracing, ex vivo heart culture, and molecular biology approaches, we demonstrated that FGF signaling in the OFT myocardium upregulated Bmp4 expression, which then enhanced smooth muscle differentiation of neural crest cells (NCCs) in the cushion. FGF signaling also promoted OFT myocardial cell invasion to the cushion. Disrupting FGF signaling interrupted cushion remodeling with reduced NCCs differentiation into smooth muscle and less cardiomyocyte invasion and resulted in malformed OFT valves. Conclusions: The results demonstrate a novel mechanism by which the FGF-BMP signaling axis regulates formation of OFT valve primordia by controlling smooth muscle differentiation of cushion NCCs. (Circ Res. 2010;107:00-00.) Key Words: FGF BMP heart development NCC differentiation cardiac valve defect C ongenital valve defects occur in 2% to 3% of the population. Cardiac valves developed from cardiac cush- ions through a series of complicated and poorly defined processes. At early stages, the cardiac cushions are com- posed of extracellular matrix lined by endothelial cells. The cushions are invaded by endothelial-derived mesen- chymal cells after completing the endothelial–mesenchy- mal transition (EMT) and neural crest cells (NCCs) from pharyngeal arches. 1 The cushions subsequently undergo a series of remodeling processes. A portion of the proximal cushion undergoes remodeling and forms valve primordia and their supporting apparatus. The valve primordia are then gradually transformed from primary bulbous swollen cushions into thinly tapered valve leaflets. 1 Although cushion formation has been extensively studied, how valve primordia are defined and how valve remodeling is regu- lated remain unclear. It has been proposed that proximal outflow tract (OFT) myocardial cells receive inductive signals and migrate into the cushion. This process, termed as myocardialization, 2 subsequently induces surrounding mesenchymal cells to undergo muscle differentiation. These newly differentiated myocardial cells further induce their adjacent mesenchymal cells to differentiate until the entire mesenchymal structure is muscularized. Although required for OFT septation, 2 whether this process is required for valve formation is unclear. The fibroblast growth factor (FGF) signaling axis has been shown to play important roles in OFT cushion formation. Yet, its role in OFT valve formation remains to be characterized. The FGF family consists of 18 receptor- binding members that regulate a broad spectrum of cellular activities, 3 and play important roles in heart development. 4 Among them, FGF4 has been shown to regulate valve precursor cell proliferation and differentiation, 5–7 and FGF8 contributes to OFT valve morphogenesis. 8,9 FGF elicits its regulatory activities via activating FGF receptor Original received January 9, 2010; resubmission received June 1, 2010; revised resubmission received September 2, 2010; accepted September 7, 2010. In August 2010, the average time from submission to first decision for all original research papers submitted to Circulation Research was 13.2 days. From the Center for Cancer and Stem Cell Biology (J.Z., J.Y.F.C., Y.H., X.L., Y.L., J.F.M., F.W.) and Center for Molecular Development and Disease (J.F.M.), Institute of Biosciences and Technology, Texas A&M Health Science Center, Houston; and Department of Biology and Biochemistry (R.J.S.), University of Houston, Tex. Correspondence to Fen Wang, Center for Cancer and Stem Cell Biology, Institute of Biosciences and Technology, Texas A&M Health Science Center, Houston, TX 77030-3303. E-mail [email protected] © 2010 American Heart Association, Inc. Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/CIRCRESAHA.110.225318 1 by guest on June 15, 2018 http://circres.ahajournals.org/ Downloaded from by guest on June 15, 2018 http://circres.ahajournals.org/ Downloaded from by guest on June 15, 2018 http://circres.ahajournals.org/ Downloaded from by guest on June 15, 2018 http://circres.ahajournals.org/ Downloaded from by guest on June 15, 2018 http://circres.ahajournals.org/ Downloaded from by guest on June 15, 2018 http://circres.ahajournals.org/ Downloaded from by guest on June 15, 2018 http://circres.ahajournals.org/ Downloaded from by guest on June 15, 2018 http://circres.ahajournals.org/ Downloaded from by guest on June 15, 2018 http://circres.ahajournals.org/ Downloaded from by guest on June 15, 2018 http://circres.ahajournals.org/ Downloaded from
-
Upload
duongthien -
Category
Documents
-
view
216 -
download
2
Transcript of The FGF-BMP Signaling Axis Regulates Outflow Tract Valve...

The FGF-BMP Signaling Axis Regulates Outflow TractValve Primordium Formation by Promoting Cushion Neural
Crest Cell DifferentiationJue Zhang, Julia Y.F. Chang, Yanqing Huang, Xiang Lin, Yongde Luo, Robert J. Schwartz,
James F. Martin, Fen Wang
Rationale: Heart valves develop from precursor structures called cardiac cushions, an endothelial-lined cardiacjelly that resides in the inner side of the heart tube. The cushions are then invaded by cells from different sources,undergo a series of complicated and poorly understood remodeling processes, and give rise to valves. Disruptionof the fibroblast growth factor (FGF) signaling axis impairs morphogenesis of the outflow tract (OFT). Yet,whether FGF signaling regulates OFT valve formation is unknown.
Objective: To study how OFT valve formation is regulated and how aberrant cell signaling causes valve defects.Methods and Results: By using mouse genetic manipulation, cell lineage tracing, ex vivo heart culture, and
molecular biology approaches, we demonstrated that FGF signaling in the OFT myocardium upregulated Bmp4expression, which then enhanced smooth muscle differentiation of neural crest cells (NCCs) in the cushion. FGFsignaling also promoted OFT myocardial cell invasion to the cushion. Disrupting FGF signaling interruptedcushion remodeling with reduced NCCs differentiation into smooth muscle and less cardiomyocyte invasion andresulted in malformed OFT valves.
Conclusions: The results demonstrate a novel mechanism by which the FGF-BMP signaling axis regulatesformation of OFT valve primordia by controlling smooth muscle differentiation of cushion NCCs. (Circ Res.2010;107:00-00.)
Key Words: FGF � BMP � heart development � NCC differentiation � cardiac valve defect
Congenital valve defects occur in 2% to 3% of thepopulation. Cardiac valves developed from cardiac cush-
ions through a series of complicated and poorly definedprocesses. At early stages, the cardiac cushions are com-posed of extracellular matrix lined by endothelial cells.The cushions are invaded by endothelial-derived mesen-chymal cells after completing the endothelial–mesenchy-mal transition (EMT) and neural crest cells (NCCs) frompharyngeal arches.1 The cushions subsequently undergo aseries of remodeling processes. A portion of the proximalcushion undergoes remodeling and forms valve primordiaand their supporting apparatus. The valve primordia arethen gradually transformed from primary bulbous swollencushions into thinly tapered valve leaflets.1 Althoughcushion formation has been extensively studied, how valveprimordia are defined and how valve remodeling is regu-lated remain unclear. It has been proposed that proximaloutflow tract (OFT) myocardial cells receive inductive
signals and migrate into the cushion. This process, termedas myocardialization,2 subsequently induces surroundingmesenchymal cells to undergo muscle differentiation.These newly differentiated myocardial cells further inducetheir adjacent mesenchymal cells to differentiate until theentire mesenchymal structure is muscularized. Althoughrequired for OFT septation,2 whether this process isrequired for valve formation is unclear.
The fibroblast growth factor (FGF) signaling axis hasbeen shown to play important roles in OFT cushionformation. Yet, its role in OFT valve formation remains tobe characterized. The FGF family consists of 18 receptor-binding members that regulate a broad spectrum of cellularactivities,3 and play important roles in heart development.4
Among them, FGF4 has been shown to regulate valveprecursor cell proliferation and differentiation,5–7 andFGF8 contributes to OFT valve morphogenesis.8,9 FGFelicits its regulatory activities via activating FGF receptor
Original received January 9, 2010; resubmission received June 1, 2010; revised resubmission received September 2, 2010; accepted September 7, 2010.In August 2010, the average time from submission to first decision for all original research papers submitted to Circulation Research was 13.2 days.
From the Center for Cancer and Stem Cell Biology (J.Z., J.Y.F.C., Y.H., X.L., Y.L., J.F.M., F.W.) and Center for Molecular Development and Disease(J.F.M.), Institute of Biosciences and Technology, Texas A&M Health Science Center, Houston; and Department of Biology and Biochemistry (R.J.S.),University of Houston, Tex.
Correspondence to Fen Wang, Center for Cancer and Stem Cell Biology, Institute of Biosciences and Technology, Texas A&M Health Science Center,Houston, TX 77030-3303. E-mail [email protected]
© 2010 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/CIRCRESAHA.110.225318
1
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from

(FGFR) tyrosine kinases encoded by 4 highly homologousgenes. FGFR substrate (FRS)2� is a broadly expressedadaptor protein that is required for the FGF to activatemitogen activated protein (MAP) and PI3K pathways, the2 major pathways in the FGF signaling cascade.10 –12
Frs2�-null embryos die between embryonic day (E)7.0 toE7.5.13 Ablation of Frs2�, or double ablation of Fgfr1/Fgfr2, in heart progenitor cells disrupts endocardial EMTand NCC deployment to the OFT, resulting in OFTalignment and septation defects.14 Ablation of Fgf8 alsoleads to OFT alignment and septation defects.9 Gain-of-function and dominant-negative mutations of SHP2 thatdirectly bind to FRS2� and mediate FGF signaling to theMAP kinase (MAPK) pathway causes enlarged valves inNoonan syndrome and LEOPARD syndrome patients,respectively.15,16
In this report, we demonstrated that ablation of Frs2�,or double ablation of Fgfr1/Fgfr2, in heart progenitor cellsled to enlarged OFT valves and bicuspid aortic valves(BAV). FGF regulated Bmp4 expression in the myocar-dium via activator protein (AP)1 transcription factor bind-ing sites located upstream of the Bmp4 coding sequence.Disrupting FGF signaling diminished Bmp4 expression inthe OFT myocardium and reduced smooth muscle (SM)differentiation of cushion NCCs, thus leaving excessiveundifferentiated NCC-derived mesenchymal cells withinvalve primordia. Treating in vitro cultured heart explantswith BMP4 partially rescued the defects. The resultsdemonstrate a novel role of NCC differentiation orches-trated by the FGF-BMP signaling axis during OFT valveformation.
MethodsAn expanded Methods section is available in the Online DataSupplement at http://circres.ahajournals.org.
All animals were housed in the Program for Animal Resources ofthe Institute of Biosciences and Technology and were handled inaccordance with the principles and procedures of the Guide for theCare and Use of Laboratory Animals. All experimental procedureswere approved by the Institutional Animal Care and Use Committee.
ResultsDisruption of FGF Signaling Leads to EnlargedOFT ValvesDisruption of FGF signaling in mouse second heart field(SHF) progenitors compromises OFT cushion formation andresults in OFT alignment and septation defects.9,14 BecauseOFT cushions also give rise to both pulmonary and aorticvalves, we then investigated whether ablating FGF signalingalso affected OFT valve development. Hematoxylin/eosin(H&E) staining of E14.5 embryo sections revealed that Fgfr1and Fgfr2 double conditional ablations in cardiac progenitorcells with Nkx2.5Cre (Fgfr1/r2cn/Nkx) led to enlarged OFTvalves (Figure 1A). Individual ablation of Fgfr1 or Fgfr2failed to cause such defects (data not shown), suggestingredundant activities of the 2 FGFRs in regulating OFT valvedevelopment. Because FRS2� is the major adaptor proteinlinking the FGFR to MAP kinase and PI3K/AKT pathways,we then ablated Frs2� alleles in the same domain withNkx2.5Cre (Frs2�cn/Nkx) or in the SHF with Mef2CCre
(Frs2�cn/Mef). Similar to Fgfr1/r2cn/Nkx, both Frs2�cn/Nkx andFrs2�cn/Mef embryos exhibited enlarged OFT valves (Figure1A). The phenotype remained obvious in neonatal hearts (OnlineFigure I). The average diameters of pulmonary and aortic valveswere 70�10 �m and 62�11 �m in E14.5 control embryos and103�26 �m and 99�29 �m in Frs2�cn/Nkx embryos, respec-tively (Figure 1B). Approximately 20% of Frs2�cn/Nkx embryoshad BAVs (Figure 1C). All valve defects were associated withOFT septation defects, indicating that OFT cushion remodelingwas affected. No apparent defects were found in mitral valvesand tricuspid valves (Figure 1D). Cell fate mapping experimentsshowed that atrioventricular valves were only composed ofendothelial lineage cells, whereas OFT valves had both endo-thelium- and NCC-derived cells (Online Figure II), suggestingthat the defects were likely associated with NCC-derived mes-enchymal cells.
Defects in OFT Cushion Cell Recruitment,Proliferation, and Apoptosis Are Not CausalFactors for Enlarged Valves in Frs2�cn OFTBoth Frs2�cn/Nkx and Frs2�cn/Mef embryos had enlarged OFTvalves (Figure 1A). Yet, only Frs2�cn/Nkx embryos showreduced endothelial and NCC contributions to OFT cushionsand have small OFT cushions.14 Thus, the valve defect likelydid not result from cushion insufficient recruitment of endo-thelium- and NCC-derived cells into OFT cushions. Becausethe enlarged Frs2�cn OFT valves had more mesenchymalcells at E14.5 day, bromodeoxyuridine labeling was used todetermine whether mutant cushions had increased cell prolif-eration activities. Both mesenchymal and myocardial cells inFrs2�cn/Nkx OFT cushions actually had reduced proliferation
Non-standard Abbreviations and Acronyms
AP activator protein
BAV bicuspid aortic valve
BMP bone morphogenetic protein
E embryonic day
EMT endothelial–mesenchymal transition
FGF fibroblast growth factor
FGFR fibroblast growth factor receptor
FHF first heart field
FRS2� fibroblast growth factor receptor substrate 2�
gal galactosidase
H&E hematoxylin/eosin
MAP mitogen-activated protein
MAPK mitogen-activated protein kinase
NCC neural crest cell
OFT outflow tract
SHF second heart field
SM smooth muscle
SMA smooth muscle actin
2 Circulation Research November 12, 2010
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from

at E10.5 and E12.5 (Figure 1E), which was consistent withour previous report.14 This suggests that enlarged valves arenot caused by proliferation defects.
To test whether the valve enlargement was caused byapoptosis defects, TUNEL assays were used to assess cushioncell apoptosis at E13.5. Compared with control Frs2�f/f OFTcushions, apoptotic cell numbers in Frs2�cn/Nkx OFT cush-ions were significantly reduced (Figure 2A and 2B).Consistently, Fgfr1/r2cn/Nkx OFT cushions also exhibitedreduced apoptosis at E13.5 (Figure 2C), suggesting that theFGF might be involved in regulating OFT cushion cellapoptosis. Surprisingly, most apoptotic cells were notlocated at the valve primordia. Instead, they were locatedbeneath the valve primordia, suggesting that enlargedvalves were not directly caused by reduced apoptosis.Moreover, Frs2�cn/Mef OFT that did not exhibit increasedapoptosis in the cushion (Figure 2D), nevertheless, stillhad enlarged OFT valves (Figure 1A), further indicating
that OFT valve hyperplasia was not caused by defectiveapoptosis. Consistent with previous findings in chickenmodels,17 cell lineage tracing revealed that only NCC-, butnot endocardium-, derived cushion mesenchymal cellsunderwent apoptosis (Online Figure III).
Ablation of Frs2� Causes Defects in CushionMyocardialization and Smooth MuscleDifferentiation, Leaving Excessive UndifferentiatedMesenchyme Within the OFT Valve PrimordiaUnlike wild-type cells that were round, Frs2�cn/Nkx OFTvalve cells were spindle-like (Figure 1A, insets), suggestingdifferentiation defects. We then investigated whether aberrantcushion cell differentiation caused the valve remodelingdefects. During this stage, proximal OFT myocardial cells,which express SM �-actin (SMA) and sarcomeric myosin,invade the cushion and induce adjacent mesenchymal cells to
Figure 1. Disruption of the FGF signaling axis leads to OFT valve hyperplasia. A, Transverse sections of E14.5 embryos were H&E-stained to demonstrate enlarged OFT valves. Pulmonary and aortic valves are highlighted with dotted lines. Insets are enlargedimages of the valve. B, Embryos were serially sectioned, and the valve dimensions were measured in every 5 sections. Averages of 3largest values were calculated from each embryo. Data represent an average dimension of at least 10 valves and are expressed asmeans�SD. C, Coronal sections of embryos were H&E-stained, demonstrating bicuspid aortic valves in E14.5 Frs2�cn/Nkx embryos. D,Transverse sections of E14.5 embryos were H&E-stained, demonstrating atrioventricular valves. E, Bromodeoxyuridine (BrdU) incorpo-ration demonstrating compromised cell proliferation in Frs2�cn/Nkx OFTs. Percentages of positive cells in the OFT myocardium andcushions from 4 embryos were calculated and expressed as means�SD. Fgfr1/r2f/f indicates Fgfr1 and Fgfr2 double floxed embryos;Fgfr1/r2cnNkx, Fgfr1/Fgfr2 double conditional ablation with Nkx2.5Cre; Frs2�flox, Frs2� floxed, Frs2�cn/Nkx, Frs2� conditional ablation withNkx2.5Cre; Frs2�cn/Mef, Frs2� conditional ablation with Mef2CCre. AV indicates aortic valve; C, cushion; M, myocardium; MV, mitralvalve; PV, pulmonary valve; TV, tricuspid valve.
Zhang et al FGF Signaling in Outflow Tract Valve Development 3
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from

undergo SM differentiation.18–20 Immunostaining with anti-SMA antibodies revealed that at E12.5, the SMA-positivemyocardial cells protruded into proximal OFT cushions ofwild-type embryos; some cells in the cushion also expressedlow level SMA (Figure 3A, a). In Frs2�cn/Nkx embryos,however, the myocardial cells failed to extend into thecushion, and SMA expressing cushion mesenchymal cellswas reduced (Figure 3A, b), indicating defects in bothmyocardial cell invasion and mesenchymal cell differentia-tion. At E13.5, increased SMA-positive cells appeared incontrol OFT cushions, which separated OFT cushions into 2parts and, thus, defined the valve primordia (Figure 3A, c).However, the SMA-positive cells in Frs2�cn/Nkx OFTcushions were reduced and failed to separate valve primor-dia from the rest of the cushion, leaving excessive mesen-chymal cells within the valve (Figure 3A, d). At E14.5,myocardialization and SM differentiation of the proximaloutlet septum were almost completed in control cushions;the valve was sculpted, and the aorta was separated fromthe right ventricle by muscular tissues (Figure 3A, e). Incontrast, the outlet septum in Frs2�cn/Nkx remained unmus-cularized, and the valve still remained bulbous and swollen(Figure 3A, f). Consistently, similar defects were found inFgfr1/r2cn/Nkx and Frs2�cn/Mef embryos (Figure 3B and3C), further confirming the role of FGF signaling in thisprocess.
Immunostaining with anti–sarcomeric myosin MF20antibody was carried out to further dissect whether theunmuscularized OFT cushions were caused by defectivemyocardialization or SM differentiation. In control OFT,the myocardial cells next to cushions were elongated andhad lamellipodia extending into the cushion mesenchyme(Figure 3D, a). In contrast, Frs2�cn/Nkx OFT myocardialcells were round and almost had no lamellipodia extendinginto the cushion (Figure 3D, b), suggesting compromisedinward migration of OFT myocardial cells. To confirmthis, cells from E13.5 proximal OFTs were isolated forTranswell migration assays, and the myocardial cells thatmigrated through the membrane were identified by sarco-
meric myosin expression. It was apparent that Frs2�cn/Nkx
OFTs had fewer cells migrating through the membranethan controls (Figure 3E), indicating that FRS2�-mediatedsignals promoted myocardial cell migration in a cellautonomous manner. Focal adhesion kinase (FAK) andVangl2 mediated noncanonical Wnt signaling pathwaysare involved in regulating OFT myocardium migration.18,21
However, expressions of FAK, Rock1, Vangl2, and Wnt11were not changed in E12.5 Frs2�cn/Nkx OFT (Figure 3F),suggesting that the FGF regulated OFT myocardium mi-gration through other pathways.
FRS2�-Mediated Signals Are Required forRegulating SM Differentiation of CushionMesenchymal Cells in the OFTThe OFT cushion mesenchyme mainly consists of bothendocardium- and NCC-derived cells. To determine howFRS2�-mediated signals regulated cushion cell differenti-ation and the origin of cushion muscular cells, cell fatemapping was carried out with ROSA26 reporter mice22 andTie2cre driver that was expressed in the endocardium23 orAp2�cre that was expressed in NCCs.24 Immunostainingrevealed that most mesenchymal cells in proximal OFTcushions were labeled by Ap2�cre, but not Tie2cre, sugges-tive NCC-origin of the cells. These NCC-derived cellswere SMA-positive and MF20-negative (Figure 4A, a andb), whereas endothelial-derived cells were both SMA andMF20 negative (Figure 4A, c and d), indicating that NCC-but not endothelial-derived cells in the OFT cushionsunderwent SM differentiation. All MF-20 –positive cellslocated at the junction of the OFT myocardium andcushion were �-galactosidase (�-gal) negative in bothTie2cre-ROSA26 and Ap2�cre-ROSA26 mice, suggestingthat these cells were neither from the endothelium nor fromNCCs, but from the myocardium (Figure 4A, b and d).Double staining of �-gal and SMA of E12.5 Nkx2.5Cre-
Figure 2. Disruption of FGF signalingreduces OFT cushion cell apoptosisduring remodeling. A and B, Apoptosisin E13.5 OFT cushions. Representativesections from anterior to posterior sidesare shown in A. Averages of total apo-ptotic cell numbers in all sections from 3individuals were calculated and areshown in B as means�SD. C, Apoptoticcells in Fgfr1/r2Nkx, Frs2�cn/Mef, and con-trol embryos at E13.5.
4 Circulation Research November 12, 2010
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from

Frs2�flox/W-ROSA26 embryos revealed that all SMA-positive OFT cushion mesenchymal cells were �-galnegative (Figure 4B, a). Because Nkx2.5Cre is expressed inboth OFT myocardial and endocardial cells, the resultsconfirmed that these SMA-positive mesenchymal cellswere neither myocardial nor endocardial derived. Further-more, because most proximal OFT cushion mesenchymalcells were NCC origin, these �-gal–negative and SMA-positive cells were most likely derived from NCCs. Thispopulation of cells were severely reduced in Nkx2.5Cre-Frs2�f/f-ROSA26 OFT cushions (Figure 4B, b), indicatingthat NCC differentiation was disrupted in mutant cushions.Furthermore, both �-gal and SMA negative cells in OFTvalves were increased from 17.1%�5 in Nkx2.5Cre-Frs2�f/w-ROSA26 embryos to 26.7%�10 in Nkx2.5Cre-Frs2�f/f-ROSA26 embryos at E14.5 (Figure 4C), suggesting thatmore undifferentiated NCCs were left in mutant than in
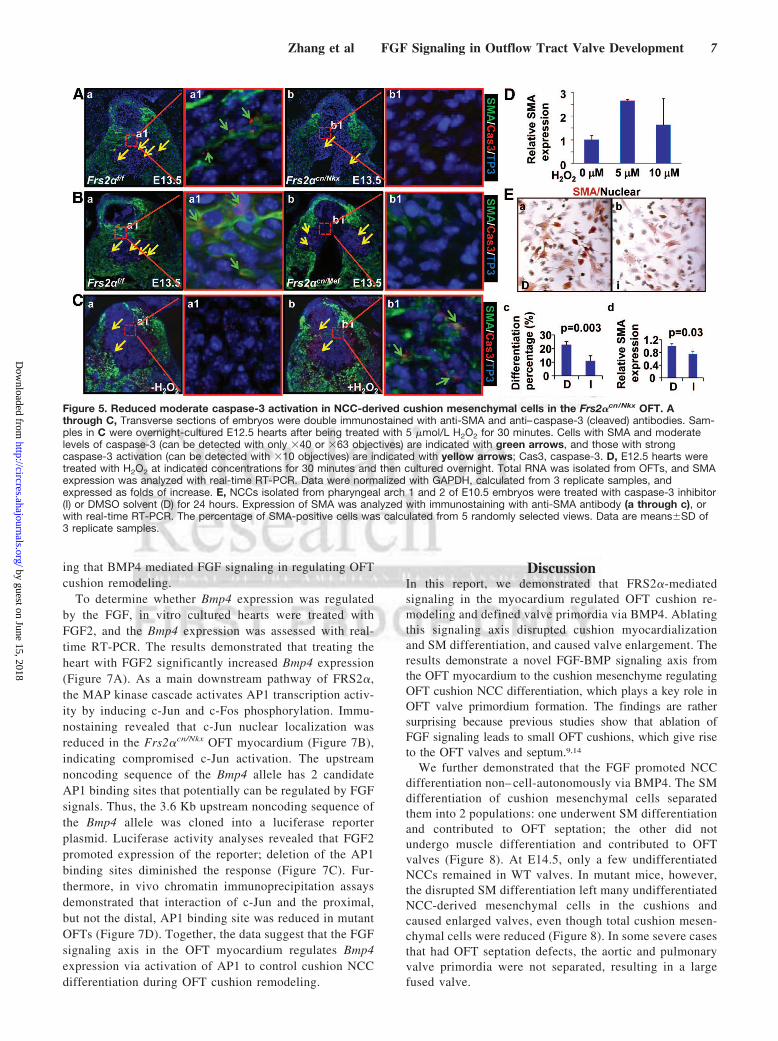
control valve primordia. Because moderate caspase-3 ac-tivity is required for stem cell differentiation,25–27 wesubsequently investigated whether caspase-3 activationlevels underlie cushion mesenchymal differentiation andapoptosis. Double immunostaining revealed that wild-typeOFT cushions had many mesenchymal cells exhibitingmoderate cleaved caspase-3 and SMA staining at E13.5(Figure 5A, a, green arrows). Frs2�cn/Nkx OFT cushionshad reduced caspase-3 positive (strongly and moderately)and SMA-positive cells (Figure 5A); Frs2�cn/Mef OFTcushions only had reduced moderate caspase-3 and SMAdouble positive cells (Figure 5B), suggesting that apoptoticand differentiated OFT cushion cells are different sub-populations. To confirm that caspase-3 activity played akey role in cushion cell differentiation, ex vivo culturedE12.5 embryonic hearts were treated with H2O2 to inducecaspase-3 activation. As expected, H2O2 treatment induced
Figure 3. Ablation of the FGF signalingaxis leads to defective myocardializa-tion and SM differentiation in OFTcushions. A through C, Transverse sec-tions of embryos were immunostainedwith anti-SMA antibody, demonstratingdefects in myocardialization and SM dif-ferentiation. The specifically bound anti-bodies were visualized either with perox-idase activities (A) or fluorescent dyes (Band C). Arrows indicate migrating myo-cardial cells (My), and arrowheads indi-cate cushion mesenchymal cells (Cu)undergoing SM differentiation. Dottedlines outline the valve primordia.To-Pro3 (TP3) was used for nuclearcounterstaining. D, Immunostaining withMF20 antibody revealed that the myo-cardial cells in control OFT were elon-gated and had long lamellipodia extend-ing into the cushion mesenchyme asindicated by arrows (a); reduced lamelli-podia were observed in Frs2�cn/Nkx OFTmyocardial cells (b). E, Myocardial cellsisolated from E13.5 OFT migratingthrough the membranes were identifiedby immunostaining with MF20 antibody(c and d). The ratios of migratedMF-20–positive cells over total MF-20–positive cells were calculated from 3individual samples and were expressedas means�SD (e). F, Real-time RT-PCRanalyses of E12.5 OFTs. Data are nor-malized to GAPDH and expressed asfolds of changes over wild-type samples.
Zhang et al FGF Signaling in Outflow Tract Valve Development 5
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from

caspase-3 activation at both high and moderate levels andSMA expression (Figure 5C). More importantly, increasedSMA expression was found in cells exhibiting moderatecaspase-3 activities. Real-time RT-PCR analyses also re-vealed that SMA expression was increased by H2O2
treatments (Figure 5D). Consistently, inhibition of caspase-3significantly suppressed expression of SMA in NCCs isolatedfrom pharyngeal arches 1 and 2 of E10.5 embryos, asrevealed by both immunostaining and real-time RT-PCRanalyses (Figure 5E). The results further demonstrated thatappearance of moderate caspase-3 activity induced OFTcushion cells to differentiate.
The BMP4 Pathway Mediates FGF Signals inRegulating OFT Cushion Cell DifferentiationBecause both Nkx2.5cre and Mef2ccre were not expressed inNCCs, FRS2�-mediated signals in regulating NCC-derived cushion mesenchymal differentiation must be non–cell-autonomous and mediated by other signaling path-ways. Furthermore, ablation of Frs2� in endothelial cellswith Tie2cre did not cause apparent defects,14 therefore, themediating signals likely were derived from the OFT
myocardium. BMP4 has been shown to control NCCapoptosis28; ablation of Bmp4 also leads to valve hyper-plasia.29 Because Bmp4 expression in the SHF is regu-lated by FRS2�-mediated signaling pathways,14 we thentested whether BMP4 was the downstream pathway thatmediated FGF signals in regulating OFT cushion remod-eling. Whole-mount in situ hybridization demonstratedthat Bmp4 expression was reduced in the Frs2�cn/Nkx OFTmyocardium at E12.5 (Figure 6A). Consistently, phosphor-ylation of Smad1/5/8, downstream targets of the BMPreceptor, was dramatically reduced in Frs2�cn/Nkx cushionmesenchymal cells (Figure 6A). Ablation of Bmp4 withMef2ccre also reduced apoptosis, expression of SMA, andactivation of caspase-3 in OFT cushion mesenchymal cells(Figure 6B). Together, it is suggested that BMP4 is thedownstream mediator of FGF for regulating OFT cushionremodeling. To test this possibility, E12.5 hearts werecultured in the presence or absence of BMP4 overnight.Both immunostaining and real-time RT-PCR analysesrevealed that BMP4 increased SMA expression in Frs2�cn/Nkx
OFT (Figure 6C and 6D) suggesting that BMP4 treatmentrescued the defects. Ablation of one Bmp4 allele inFrs2�cn/Mef embryos increased the penetrance of valvehyperplasia from 38% to 58% (Figure 6E), further suggest-
Figure 4. Ablation of Frs2� inheart progenitor cells com-promises NCC-derived cush-ion cells undergoing SM dif-ferentiation. A, Transversesections of embryos were im-munostained with anti–�-galand anti-SMA or MF20 anti-bodies as specified. B and C,Double immunostaining oftransverse embryo sectionsrevealed that SMA expressionin �-gal–negative NCCs wasreduced in Frs2�cn/Nkx OFTcushions (B) and valves (C).Percentage of �-gal–negativecells was scored from serialsections of Frs2�cn/Nkx andcontrol OFT valves and wasexpressed as means�SD.
6 Circulation Research November 12, 2010
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from
ing that BMP4 mediated FGF signaling in regulating OFTcushion remodeling.
To determine whether Bmp4 expression was regulatedby the FGF, in vitro cultured hearts were treated withFGF2, and the Bmp4 expression was assessed with real-time RT-PCR. The results demonstrated that treating theheart with FGF2 significantly increased Bmp4 expression(Figure 7A). As a main downstream pathway of FRS2�,the MAP kinase cascade activates AP1 transcription activ-ity by inducing c-Jun and c-Fos phosphorylation. Immu-nostaining revealed that c-Jun nuclear localization wasreduced in the Frs2�cn/Nkx OFT myocardium (Figure 7B),indicating compromised c-Jun activation. The upstreamnoncoding sequence of the Bmp4 allele has 2 candidateAP1 binding sites that potentially can be regulated by FGFsignals. Thus, the 3.6 Kb upstream noncoding sequence ofthe Bmp4 allele was cloned into a luciferase reporterplasmid. Luciferase activity analyses revealed that FGF2promoted expression of the reporter; deletion of the AP1binding sites diminished the response (Figure 7C). Fur-thermore, in vivo chromatin immunoprecipitation assaysdemonstrated that interaction of c-Jun and the proximal,but not the distal, AP1 binding site was reduced in mutantOFTs (Figure 7D). Together, the data suggest that the FGFsignaling axis in the OFT myocardium regulates Bmp4expression via activation of AP1 to control cushion NCCdifferentiation during OFT cushion remodeling.
DiscussionIn this report, we demonstrated that FRS2�-mediatedsignaling in the myocardium regulated OFT cushion re-modeling and defined valve primordia via BMP4. Ablatingthis signaling axis disrupted cushion myocardializationand SM differentiation, and caused valve enlargement. Theresults demonstrate a novel FGF-BMP signaling axis fromthe OFT myocardium to the cushion mesenchyme regulatingOFT cushion NCC differentiation, which plays a key role inOFT valve primordium formation. The findings are rathersurprising because previous studies show that ablation ofFGF signaling leads to small OFT cushions, which give riseto the OFT valves and septum.9,14
We further demonstrated that the FGF promoted NCCdifferentiation non– cell-autonomously via BMP4. The SMdifferentiation of cushion mesenchymal cells separatedthem into 2 populations: one underwent SM differentiationand contributed to OFT septation; the other did notundergo muscle differentiation and contributed to OFTvalves (Figure 8). At E14.5, only a few undifferentiatedNCCs remained in WT valves. In mutant mice, however,the disrupted SM differentiation left many undifferentiatedNCC-derived mesenchymal cells in the cushions andcaused enlarged valves, even though total cushion mesen-chymal cells were reduced (Figure 8). In some severe casesthat had OFT septation defects, the aortic and pulmonaryvalve primordia were not separated, resulting in a largefused valve.
Figure 5. Reduced moderate caspase-3 activation in NCC-derived cushion mesenchymal cells in the Frs2�cn/Nkx OFT. Athrough C, Transverse sections of embryos were double immunostained with anti-SMA and anti–caspase-3 (cleaved) antibodies. Sam-ples in C were overnight-cultured E12.5 hearts after being treated with 5 �mol/L H2O2 for 30 minutes. Cells with SMA and moderatelevels of caspase-3 (can be detected with only �40 or �63 objectives) are indicated with green arrows, and those with strongcaspase-3 activation (can be detected with �10 objectives) are indicated with yellow arrows; Cas3, caspase-3. D, E12.5 hearts weretreated with H2O2 at indicated concentrations for 30 minutes and then cultured overnight. Total RNA was isolated from OFTs, and SMAexpression was analyzed with real-time RT-PCR. Data were normalized with GAPDH, calculated from 3 replicate samples, andexpressed as folds of increase. E, NCCs isolated from pharyngeal arch 1 and 2 of E10.5 embryos were treated with caspase-3 inhibitor(I) or DMSO solvent (D) for 24 hours. Expression of SMA was analyzed with immunostaining with anti-SMA antibody (a through c), orwith real-time RT-PCR. The percentage of SMA-positive cells was calculated from 5 randomly selected views. Data are means�SD of3 replicate samples.
Zhang et al FGF Signaling in Outflow Tract Valve Development 7
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from

Although it has been reported that mature OFT valvecells are largely derived from the endocardium (de Langeet al30 and Jiang et al, 2000) the part of OFT cushions thatgave rise to valves and the underlying region were mainlycomprised of NCC-derived mesenchymal cells at E13.5(Online Figure II). These NCC lineage cells then eitherunderwent apoptosis or SM differentiation (Figures 4 and5), leaving mainly endothelial lineage cells in OFT valves.The data are consistent with previous findings that noendocardial lineage cells in OFT valves undergo apoptosisand that the endocardial lineage replaces NCCs during latestages of valve formation.30 Frs2�cn/Nkx OFT valves hadsignificantly increased non-Nkx2.5 lineage cells (Figure4C), suggesting that more NCC-lineage contributed tomutant OFT valves. Unlike OFT cushions, AV cushionsonly had the endocardium lineage mesenchymal cells(Online Figure II). This likely explains why AV valveswere not affected by ablation of FGF signaling.
Mutations in SM �-actin are associated with familialBAV defects,31 implying that defective SM differentiationof NCCs may contribute to BAV. However, although NCCdifferentiation was disrupted in both Frs2�cn/Nkx andFrs2�cn/Mef OFT cushions, only Frs2�cn/Nkx embryos ex-
hibited BAV defects, indicating that BAV defects in Frs2�mutants were not caused by disrupted NCC differentiation.Because cushion mesenchymal apoptosis was disrupted inFrs2�cn/Nkx, but not Frs2�cn/Mef, OFT cushions, it ispossible that reduced apoptosis contributed to failure inseparating the valve leaflets from each other.
FRS2� has 4 GRB2 and 2 SHP2 binding sites. SHP2 is aubiquitously expressed protein tyrosine phosphatase thatplays a central role in the MAP kinase and other signalingpathways.32,33 Gain of function of SHP2 enhances MAPKactivation and promotes the EMT and mesenchymal cellproliferation; dominant negative SHP2 suppresses MAPKactivation and fails to arrest the proliferation of mesenchymalcells at later stages.34 Thus, a narrow range of MAPK activityis required for embryonic development. Ablation of Frs2� inheart progenitor cells reduces MAPK activation in the myo-cardium. Here, we demonstrated that deletion of Frs2� led toreduced AP1 transcription activity, resulting in reduced Bmp4expression in the myocardium, inhibited cushion NCC dif-ferentiation, and caused valve hyperplasia. The results reveala novel mechanism that FGF signals in the OFT myocardiumnegatively regulate the valve size through promotion of NCCdifferentiation.
Figure 6. FRS2�-mediated signals regulate OFT cushion remodeling via BMP4. A, Whole-mount in situ hybridization with E12.5embryos demonstrates reduced Bmp4 expression in Frs2�cn/Nkx OFT (a and b). Red arrows indicate Bmp4 expression. c and d,Transverse sections of E12.5 embryos were immunostained with antibody against phosphorylated Smad1/5/8. B, Apoptotic cells intransverse sections of E13.5 embryos were detected with TUNEL assays (a and b). c and d, Double immunostaining of transverse sec-tions of E13.5 embryos with anti-SMA and anti–caspase-3 antibodies. Green arrows indicate cells with moderate caspase-3 activationand SMA expressions, and yellow arrow indicates strong caspase-3 activation. C, Embryonic hearts collected at E12.5 were culturedovernight in the presence or absence of 10 ng/mL BMP4. Paraffin sections of the hearts were then immunostained with anti-SMA anti-bodies. D, Real-time RT-PCR analyses of SMA expression in ex vivo– cultured OFTs. Data were normalized to GAPDH andexpressed as folds of changes over untreated samples. E, Ablation of one Bmp4 allele increases the penetrance of OFT valvedefects in Frs2�cn/Mef embryos.
8 Circulation Research November 12, 2010
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from

The regulatory mechanism underlying SM differentia-tion of OFT cushion NCCs remains poorly understood. Ithas been shown that transient and moderate caspase-3activities promote stem cell differentiation.25 Consistently,
only those cells with moderate caspase-3 activity in OFTcushions undergo SM differentiation. Ablation of Frs2�compromised BMP4 signaling in OFT cushion cells andreduced the population of cells with SMA expression and
Figure 7. The FGF signaling axis regu-lates Bmp4 expression via activatingthe AP1 transcription activity. A, E12.5heart explants were cultured in the pres-ence or absence of 10 ng/mL FGF2overnight, and Bmp4 expression wasanalyzed with real-time RT-PCR. Datawere normalized to GAPDH andexpressed as folds of increase overuntreated samples. B, Transverse sec-tions of E12.5 embryos were immuno-stained with anti–c-JUN antibody, show-ing reduced c-JUN nuclear localizationin the OFT myocardium of Frs2�cn/Nkx
embryos. Insets are high-magnificationviews of the boxed areas. OFT cushionsare highlighted with dotted lines. C,AP1-binding sites in the Bmp4 allelewere required for FGF2 stimulation. a,Luciferase reporter constructs. b, Lucif-erase reporter–transfected C2C12 cellswere incubated overnight in the pres-ence or absence of 10 ng/mL FGF2, andthe luciferase activity was analyzed. Dataare means and SD from 3 replicatewells. �AP indicates AP1 site–deletedmutant. D, E12.5 hearts were subjectedto chromatin immunoprecipitation assayanalyses with anti–C-Jun antibody.Pulled-down DNA fragments were ana-lyzed with real-time PCR and expressedas folds of changes over untreatedsamples.
Figure 8. Working model of FGF signaling inOFT valve formation. Defective cushion NCC dif-ferentiation fails to define valve primordia andleaves excessive cells within valve primordia,which results in a large valve. Valve primordia areoutlined with red lines.
Zhang et al FGF Signaling in Outflow Tract Valve Development 9
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from

moderate caspase-3 activity. Therefore, the results suggestthat BMP4 is an upstream regulator for caspase-3 activa-tion in OFT cushion NCCs. It remains unanswered whythese cushion mesenchymal cells have different caspase-3activities. One possibility is that differentiated NCCs areinsensitive to BMP4 stimulation, and undifferentiatedNCCs are BMP sensitive and will undergo apoptosis whenthey receive sustained BMP4 stimulation. However, fur-ther experimentation is needed to test this hypothesis.
In conclusion, we demonstrated a novel FGF-BMP signal-ing axis between the OFT myocardium and cushion mesen-chyme that regulated OFT cushion myocardialization and SMdifferentiation during cushion remodeling and that the 2processes play an important role in defining OFT valveprimordia. The results shed new light on understanding howperturbation of cell signaling and NCC differentiation lead tocongenital heart valve defects.
AcknowledgmentsWe thank Drs Juha Partanen and David Ornitz for generously sharingthe Fgfr1-floxed and Fgfr2-floxed mice and Mary Cole for criticalreading of the manuscript.
Sources of FundingThis work was supported by National Cancer Institute NIH grantCA96824 (to F.W.), NIH grants 2R01DE12324 and R01HL093484(to J.F.M.), and American Heart Association Grants AHA0655077Y(to F.W.) and 09PRE2010130 (to J.Z.).
DisclosuresNone.
References1. Armstrong EJ, Bischoff J. Heart valve development: endothelial cell
signaling and differentiation. Circ Res. 2004;95:459–470.2. van den Hoff MJ, Kruithof BP, Moorman AF. Making more heart muscle.
Bioessays. 2004;26:248–261.3. McKeehan WL, Wang F, Luo Y. The fibroblast growth factor (FGF)
signaling complex. In: Bradshaw RA, Dennis EA, eds. Handbook of CellSignaling. Vol I. 2nd ed. New York: Academic/Elsevier Press; 2009:253–259.
4. Chien KR, Domian IJ, Parker KK. Cardiogenesis and the complexbiology of regenerative cardiovascular medicine. Science. 2008;322:1494–1497.
5. Lincoln J, Alfieri CM, Yutzey KE. BMP and FGF regulatory pathwayscontrol cell lineage diversification of heart valve precursor cells. DevBiol. 2006;292:292–302.
6. Sugi Y, Ito N, Szebenyi G, Myers K, Fallon JF, Mikawa T, MarkwaldRR. Fibroblast growth factor (FGF)-4 can induce proliferation of cardiaccushion mesenchymal cells during early valve leaflet formation. Dev Biol.2003;258:252–263.
7. Zhao B, Etter L, Hinton RB Jr, Benson DW. BMP and FGF regulatorypathways in semilunar valve precursor cells. Dev Dyn. 2007;236:971–980.
8. Ilagan R, Abu-Issa R, Brown D, Yang YP, Jiao K, Schwartz RJ, Klin-gensmith J, Meyers EN. Fgf8 is required for anterior heart field devel-opment. Development. 2006;133:2435–2445.
9. Park EJ, Watanabe Y, Smyth G, Miyagawa-Tomita S, Meyers E, Klin-gensmith J, Camenisch T, Buckingham M, Moon AM. An FGF autocrineloop initiated in second heart field mesoderm regulates morphogenesis atthe arterial pole of the heart. Development. 2008;135:3599–3610.
10. Kouhara H, Hadari YR, Spivak-Kroizman T, Schilling J, Bar-Sagi D, LaxI, Schlessinger J. A lipid-anchored Grb2-binding protein that links FGF-receptor activation to the Ras/MAPK signaling pathway. Cell. 1997;89:693–702.
11. Xu H, Lee KW, Goldfarb M. Novel recognition motif on fibroblastgrowth factor receptor mediates direct association and activation of SNTadapter proteins. J Biol Chem. 1998;273:17987–17990.
12. Ong SH, Lim YP, Low BC, Guy GR. SHP2 associates directly withtyrosine phosphorylated p90 (SNT) protein in FGF-stimulated cells.Biochem Biophys Res Commun. 1997;238:261–266.
13. Hadari YR, Gotoh N, Kouhara H, Lax I, Schlessinger J. Critical role forthe docking-protein FRS2 alpha in FGF receptor-mediated signal trans-duction pathways. Proc Natl Acad Sci U S A. 2001;98:8578–8583.
14. Zhang J, Lin Y, Zhang Y, Lan Y, Lin C, Moon AM, Schwartz RJ, MartinJF, Wang F. Frs2alpha-deficiency in cardiac progenitors disrupts a subsetof FGF signals required for outflow tract morphogenesis. Development.2008;135:3611–3622.
15. Chen B, Bronson RT, Klaman LD, Hampton TG, Wang JF, Green PJ,Magnuson T, Douglas PS, Morgan JP, Neel BG. Mice mutant for Egfrand Shp2 have defective cardiac semilunar valvulogenesis. Nat Genet.2000;24:296–299.
16. Krenz M, Yutzey KE, Robbins J. Noonan syndrome mutation Q79R inShp2 increases proliferation of valve primordia mesenchymal cells viaextracellular signal-regulated kinase 1/2 signaling. Circ Res. 2005;97:813–820.
17. Poelmann RE, Mikawa T, Gittenberger-de Groot AC. Neural crest cells inoutflow tract septation of the embryonic chicken heart: differentiation andapoptosis. Dev Dyn. 1998;212:373–384.
18. Phillips HM, Murdoch JN, Chaudhry B, Copp AJ, Henderson DJ.Vangl2 acts via RhoA signaling to regulate polarized cell movementsduring development of the proximal outflow tract. Circ Res. 2005;96:292–299.
19. van den Hoff MJ, Moorman AF, Ruijter JM, Lamers WH, BenningtonRW, Markwald RR, Wessels A. Myocardialization of the cardiac outflowtract. Dev Biol. 1999;212:477–490.
20. van den Hoff MJ, Kruithof BP, Moorman AF, Markwald RR, Wessels A.Formation of myocardium after the initial development of the linear hearttube. Dev Biol. 2001;240:61–76.
21. Hakim ZS, DiMichele LA, Doherty JT, Homeister JW, Beggs HE,Reichardt LF, Schwartz RJ, Brackhan J, Smithies O, Mack CP, TaylorJM. Conditional deletion of focal adhesion kinase leads to defects inventricular septation and outflow tract alignment. Mol Cell Biol. 2007;27:5352–5364.
22. Soriano P. Generalized lacZ expression with the ROSA26 Cre reporterstrain. Nat Genet. 1999;21:70–71.
23. Kisanuki YY, Hammer RE, Miyazaki J, Williams SC, Richardson JA,Yanagisawa M. Tie2-Cre transgenic mice: a new model for endothelialcell-lineage analysis in vivo. Dev Biol. 2001;230:230–242.
24. Macatee TL, Hammond BP, Arenkiel BR, Francis L, Frank DU, MoonAM. Ablation of specific expression domains reveals discrete functions ofectoderm- and endoderm-derived FGF8 during cardiovascular and pha-ryngeal development. Development. 2003;130:6361–6374.
25. Abdul-Ghani M, Megeney LA. Rehabilitation of a contract killer:caspase-3 directs stem cell differentiation. Cell Stem Cell. 2008;2:515–516.
26. Janzen V, Fleming HE, Riedt T, Karlsson G, Riese MJ, Lo Celso C,Reynolds G, Milne CD, Paige CJ, Karlsson S, Woo M, Scadden DT.Hematopoietic stem cell responsiveness to exogenous signals is limitedby caspase-3. Cell Stem Cell. 2008;2:584–594.
27. Fujita J, Crane AM, Souza MK, Dejosez M, Kyba M, Flavell RA,Thomson JA, Zwaka TP. Caspase activity mediates the differentiation ofembryonic stem cells. Cell Stem Cell. 2008;2:595–601.
28. Graham A, Francis-West P, Brickell P, Lumsden A. The signallingmolecule BMP4 mediates apoptosis in the rhombencephalic neural crest.Nature. 1994;372:684–686.
29. McCulley DJ, Kang JO, Martin JF, Black BL. BMP4 is required in theanterior heart field and its derivatives for endocardial cushion remodeling,outflow tract septation, and semilunar valve development. Dev Dyn.2008;237:3200–3209.
30. de Lange FJ, Moorman AF, Anderson RH, Manner J, Soufan AT, deGier-de Vries C, Schneider MD, Webb S, van den Hoff MJ, ChristoffelsVM. Lineage and morphogenetic analysis of the cardiac valves. Circ Res.2004;95:645–654.
31. Guo DC, Pannu H, Tran-Fadulu V, Papke CL, Yu RK, Avidan N,Bourgeois S, Estrera AL, Safi HJ, Sparks E, Amor D, Ades L, McConnell
10 Circulation Research November 12, 2010
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from

V, Willoughby CE, Abuelo D, Willing M, Lewis RA, Kim DH, SchererS, Tung PP, Ahn C, Buja LM, Raman CS, Shete SS, Milewicz DM.Mutations in smooth muscle alpha-actin (ACTA2) lead to thoracic aorticaneurysms and dissections. Nat Genet. 2007;39:1488–1493.
32. Chan G, Kalaitzidis D, Neel BG. The tyrosine phosphatase Shp2(PTPN11) in cancer. Cancer Metastasis Rev. 2008;27:179–192.
33. Feng GS. Shp2-mediated molecular signaling in control of embryonicstem cell self-renewal and differentiation. Cell Res. 2007;17:37–41.
34. Edouard T, Montagner A, Dance M, Conte F, Yart A, Parfait B, TauberM, Salles JP, Raynal P. How do Shp2 mutations that oppositely influenceits biochemical activity result in syndromes with overlapping symptoms?Cell Mol Life Sci. 2007;64:1585–1590.
Novelty and Significance
What Is Known?
● Cardiac valves develop from cardiac cushions located inside the hearttube, and cardiac cushion formation and remodeling are the 2major processes of cardiac valve formation.
● Outflow tract (OFT) cushion mesenchymal cells are derived fromendothelial and neural crest cells (NCCs).
● Fibroblast growth factor (FGF) signaling regulates contributions ofendothelial cells and NCCs to the OFT cushion.
What New Information Does This Article Contribute?
● During cushion remodeling, the cushion mesenchymal myocardial-ization and smooth muscle differentiation defines the size of valveprimordium
● The FGF-BMP4 signaling axis regulates migration of cardiomyocytesinto OFT cushions and smooth muscle differentiation of cushionNCCs.
● Disruption of the FGF-BMP4 axis leads to OFT valve hyperplasia.
Congenital cardiac valve defect is a major birth defect. Howcardiac valves develop from cardiac cushions and how the valveprimordium is defined are poorly understood. Here, we demon-strated that myocardialization and smooth muscle differentiationof cushion mesenchyme defines the valve primordium. Wediscovered a novel FGF-BMP4 signaling axis from the OFTmyocardium to the cushion mesenchyme, which regulates NCCdifferentiation in the OFT. We further revealed that FGF signalingpromotes BMP4 expression through activation of the AP1transcription factor. Ablation of this signaling axis disruptedmyocardialization and smooth muscle differentiation of thecushion, which left too many undifferentiated NCCs within thevalves and, thus, caused OFT valve hyperplasia. We also foundthat ablation of FGF signaling in the myocardium compromisedcushion mesenchymal apoptosis and caused bicuspid aorticvalve defect. Collectively, our work provides evidence of how theOFT valve primordium is set apart from the rest of the cushionsand provides new insight into valve diseases at the molecularand cellular level.
Zhang et al FGF Signaling in Outflow Tract Valve Development 11
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from

James F. Martin and Fen WangJue Zhang, Julia Y.F. Chang, Yanqing Huang, Xiang Lin, Yongde Luo, Robert J. Schwartz,
Promoting Cushion Neural Crest Cell DifferentiationThe FGF-BMP Signaling Axis Regulates Outflow Tract Valve Primordium Formation by
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2010 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
published online September 16, 2010;Circ Res.
http://circres.ahajournals.org/content/early/2010/09/16/CIRCRESAHA.110.225318.citationWorld Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org/content/suppl/2010/09/16/CIRCRESAHA.110.225318.DC1Data Supplement (unedited) at:
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on June 15, 2018http://circres.ahajournals.org/
Dow
nloaded from

225318-R2
1
Online Supplemental Information
The FGF-BMP signaling axis regulates outflow tract valve primordium formation by promoting cushion neural crest cell differentiation
Jue Zhang, Julia Y. F. Chang, Yanqing Huang, Xiang Lin, Yongde Luo, Robert J. Schwartz, James F. Martin, and Fen Wang
From Center for Cancer and Stem Cell Biology (J.Z., J.Y-F.C., Y.H., X.L., Y.L., J.F.M., F.W.), Center for Molecular Development and Disease (J.F.M.), Institute of Biosciences and Technology, Texas A&M Health Science Center, Houston, TX; and Biology and Biochemistry Department (R.J.S.), University of Houston, Houston, TX.
Correspondence to Fen Wang, Center for Cancer and Stem Cell Biology, Institute of Biosciences and Technology, Texas A&M Health Science Center, Houston, TX 77030-3303. E-mail: [email protected]

225318-R2
2
Online Extended Methods
Animal models
All animals were housed in the Program for Animal Resources of the Institute of
Biosciences and Technology, and were handled in accordance with the principles and
procedures of the Guide for the Care and Use of Laboratory Animals. All experimental
procedures were approved by the Institutional Animal Care and Use Committee.
Short-term organ culture of mouse embryonic hearts
Mouse embryonic hearts were dissected in ice-cold PBS and cultured at 37 oC in 12-
well plates containing 1 ml DMEM medium with 10% FBS and penicillin and
streptomycin (100 U/ml each). FGF2 or BMP4 was added to the media to a final
concentration of 10 ng/ml.
Transwell migration assay
The OFTs dissected from E13.5 embryos were minced and rinsed extensively in the
MSS buffer. After being digested with 0.25% trypsin in PBS for 20 minutes at 37 °C, the
samples were triturated to obtain single-cell suspensions, followed by filtering with 8 μm
pore size filters. The cells were pelleted by centrifugation at 3000 rpm for 2 minutes
and resuspended in DMEM with 15% FBS. The cells were then placed on transwell
filters (Corning Costar, Corning, NY) and cultured at 37 °C for 24 hours. The medium
was replaced with serum-free DMEM in transwell inserts and serum-containing medium
in the chamber. Twenty-four hours later, the cells were immunostained with MF20. The

225318-R2
3
non-migrated cells on the upper surface of the filter were removed with a cotton swab;
the migrated MF20 positive cells were counted in eight randomly assigned fields per
sample.
Immunostaining and lacZ staining
Immunostaining was performed on 5 μm sections mounted on Superfrost/Plus slides
(Fisher Scientific, Pittsburgh, PA). The antigens were retrieved by incubating in the
citrate buffer (10 mM) for 20 minutes at 100 °C or as suggested by the manufacturers.
The sources and concentrations of primary antibodies are: anti-cleaved caspase-3
(1:1000 dilution) and anti-phosphorylated Smad1,5,8 (1:1000) from Cell Signaling
(Danvers, MA); anti-SM actin (1:1) from Sigma (St. Louis, MO); anti-c-Jun (1:1000)
from Santa Cruz (Santa Cruz, CA); anti-β-galactosidase (1:10,000) from MP
Biomedicals (Solon, OH), and anti-sarcomere myosin MF20 (1:50) from Developmental
Studies Hybridoma Bank (Iowa City, IA). The specifically-bound antibodies were
detected with HRP-conjugated secondary antibody (Bio-Rad Co., Hercules, CA) and
visualized using TSATM Plus Fluorescence Systems from PerkinElmer (Boston, MA) on
a Zeiss LSM 510 Confocal Microscope. For TUNEL assays, tissues were fixed and
sectioned, and the apoptotic cells were detected with the DeadEnd Fluorometric TUNEL
System from Promega, Inc (Madison, MI). For lacZ staining, the embryos were first
lightly fixed with 0.2% glutaraldehyde for 1-2 hours and then incubated overnight with
1mg/ml X-Gal at room temperature. The tissues were then dehydrated and paraffin
embedded for the subsequent analyses as described 1.

225318-R2
4
BrdU (5’-bromo-2’-deoxyuridine, Sigma, St. Louis, MO) was intraperitoneally injected
into pregnant female mice (0.5 mg per 10 g body weight) 2-3 hours prior to being
sacrificed. The embryos were collected and fixed in 4% PFA for 4 hours and paraffin-
embedded; incorporated BrdU was detected by immunostaining with anti-BrdU antibody
(1:1000 dilution) from Sigma (St. Louis, MO).
Gene expression analyses
Total RNA was extracted with the MaxRecovery™ BiooPure™ RNA Isolation Reagent
(Bioo Scientific, Austin, TX). Reverse transcriptions were carried out with SuperScript II
(GIBCO/BRL, Life Technologies, Grand Island, NY) and random primers. Real-time
RT-PCR analyses were carried out with the SYBR Green JumpStart Taq ReadyMix
(Sigma, St. Louis, MO). The primer sequences are: BMP4-F
(AGGAGGAGGAGGAAGAGCAG), BMP4-R (TGTGATGAGGTGTCCAGGAA), SMA-F
(TGACGCTGAAGTATCCGATAG), and SMA-R (GCCAAGTCCAGACGCATGA).
Relative abundance of the mRNA was calculated using the comparative threshold (CT)
cycle method, normalized with GADPH, and expressed as fold of differences between
Frs2αcn and control hearts. Whole-mount in situ hybridization was performed as
previously described 1.
Construction and analyses of the Bmp4-luciferase reporters
A 3.6 KB genomic DNA fragment that includes two AP1 binding sites upstream of the
Bmp4 transcription initiation site was PCR amplified and cloned into a pGL3-basic
vector (Promega, Madison, WI). PCR-mediated mutagenesis was used to generate
mutants with deletions of the two AP1 binding sites. For luciferase expression assays,

225318-R2
5
C2C12 cells (1X105 cells in 12-well plates) in serum free media were transfected with
0.3 μg plasmid premixed with 2 μl Lipofectamine (Invitrogen, Carlsbad, CA). The cells
were incubated with 10% FBS for 24 hours and then serum-free medium containing 10
μg/ml heparin at 37 °C for 16 hours. FGF2 was added to the medium at a final
concentration of 10 ng/ml. The cells were lysed 16 hours later for luciferase analyses
as previously described 1.
Chromatin immunoprecipitation (ChIP) assays
For the ChIP experiments, extracts were prepared from ten E12.5 hearts. The tissues
were lightly fixed in 2% formaldehyde-DMEM with 10% FBS for 30 min at room
temperature. ChIP assays were carried out with the EZ-ChIP Kit (Millipore, Billerica,
MA) according to the manufacturer’s protocols. The following two sets of real-time PCR
primers specifically for the AP1 containing regions were used: BMP4-F5
(TCAACGTTTGGGAATCCTTT) and BMP4-R5 (TGGGCTTCCCTGAGTTTAGA),
BMP4-F6 (TGCTATAGACCTATTTCTGGACG) and BMP4-R6
(AACCAGACAGTGATTGCAGG); BMP4-F7 (GCTGCTGGCCTCACCCCTTC) and
BMP4-R7 (TGGGAGGTGGAATGTGGCCG) were used as the control for a non-AP1
region in the Bmp4 promoter.

225318-R2
6
Supplemental figures
Online Figure I. Persistent phenotype of OFT valve enlargement in Frs2αcn/Nkx mice. H&E staining of neonatal hearts shows enlarged OFT valves. Arrows indicate the OFT valves. PV, pulmonary valve; AV, aortic valve.

225318-R2
7
Online Figure II. The atrioventricular valve cushion mesenchyme consists of endothelial, but not NCC, derived cells. Transverse sections of Tie2cre-ROSA26 and Ap2αcre-ROSA26 bigenic embryos at E13.5 were double immunostained with anti-β galactosidase and MF20 antibodies. To-Pro3 was used for nuclear counterstaining. MV, mitral valves; TV, tricuspid valve.
Online Figure III. NCC-, but not endothelium-, derived OFT cushion mesenchymal cells undergo apoptosis during valve formation. Tie2cre-ROSA26 and Ap2αcre-ROSA26 bigenic embryos collected at E13.5 were stained with X-Gal to trace the endothelium and NCC lineages, respectively. The embryos were then sectioned transversely for the TUNEL analyses. Only cushion cells in Ap2αcre, but not in Tie2cre, embryos exhibited double labeling.

225318-R2
8
REFERENCES
1. Zhang J, Lin Y, Zhang Y, Lan Y, Lin C, Moon AM, Schwartz RJ, Martin JF, Wang F. Frs2alpha-deficiency in cardiac progenitors disrupts a subset of FGF signals required for outflow tract morphogenesis. Development. 2008; 135:3611-3622.


















