
The effect of visual similarity on short-term memory for spatial location: Implications for the...
22
Acta Psychologica 83 (1993) 203-224 North-Holland 203 The effect of visual similarity on short-term memory for spatial location: Implications for the capacity of visual short-term memory Peter Walker a, Graham J. Hitch b and Sallyann Duroe a ’ University of Central Lancashire, Preston, UK b Lancaster University, Lancaster, UK Accepted August 1992 Evidence from the recency effect suggests that visual short-term memory is limited to preserving information about a single pattern. Three experiments explored the capacity of visual short-term memory using a task which involved presenting a series of patterns in a random spatio-temporal sequence and probing memory for the spatial location of one of them. Experi- ment 1 used sequences of quasi-random block patterns which were either visually similar or dissimilar. Serial position curves showed a single-item recency effect. However, there was also a deleterious effect of visual similarity for the earlier patterns, indicating that visual short-term memory was supporting their retention. Experiment 2 showed that performance was unaffected by articulatory suppression, confirming that phonological coding is not an important factor in the localisation task. Experiment 3 switched to letter stimuli and, because of the known tendency for this type of stimulus to be phonologically recoded, required articulatory suppression. Letters were shown in a customised form in sequences which were either visually similar or visually dissimilar. The results replicated experiment 1 in showing restricted recency combined with an effect of visual similarity extending across earlier list items. When the same letters were shown in standard form, the recency effect remained but the similarity effect disappeared, ruling out possible interpretations in terms of semantic as well as phonological coding. It is concluded that the recency effect gives a misleading indication of the capacity of visual short-term memory. The visual similarity effect suggests that it is capable of supporting the retention of several patterns. The distinction between speech-based and visuo-spatial codes is now a standard feature of accounts of short-term memory. In the Working Memory model, for example, the slave systems controlled by a central executive comprise a phonological store, supported by articulation, Correspondence tot P. Walker, Faculty of Health, University of Central Lancashire, Preston PRl 2HE, UK. OOOl-6918/93/$06.00 0 1993 - Elsevier Science Publishers B.V. All rights reserved
-
Upload
peter-walker -
Category
Documents
-
view
216 -
download
1
Transcript of The effect of visual similarity on short-term memory for spatial location: Implications for the...

Acta Psychologica 83 (1993) 203-224
North-Holland
203
The effect of visual similarity on short-term memory for spatial location: Implications for the capacity of visual short-term memory
Peter Walker a, Graham J. Hitch b and Sallyann Duroe a ’ University of Central Lancashire, Preston, UK b Lancaster University, Lancaster, UK
Accepted August 1992
Evidence from the recency effect suggests that visual short-term memory is limited to
preserving information about a single pattern. Three experiments explored the capacity of visual
short-term memory using a task which involved presenting a series of patterns in a random
spatio-temporal sequence and probing memory for the spatial location of one of them. Experi-
ment 1 used sequences of quasi-random block patterns which were either visually similar or
dissimilar. Serial position curves showed a single-item recency effect. However, there was also a
deleterious effect of visual similarity for the earlier patterns, indicating that visual short-term
memory was supporting their retention. Experiment 2 showed that performance was unaffected
by articulatory suppression, confirming that phonological coding is not an important factor in the
localisation task. Experiment 3 switched to letter stimuli and, because of the known tendency for
this type of stimulus to be phonologically recoded, required articulatory suppression. Letters
were shown in a customised form in sequences which were either visually similar or visually
dissimilar. The results replicated experiment 1 in showing restricted recency combined with an
effect of visual similarity extending across earlier list items. When the same letters were shown in
standard form, the recency effect remained but the similarity effect disappeared, ruling out
possible interpretations in terms of semantic as well as phonological coding. It is concluded that
the recency effect gives a misleading indication of the capacity of visual short-term memory. The
visual similarity effect suggests that it is capable of supporting the retention of several patterns.
The distinction between speech-based and visuo-spatial codes is now a standard feature of accounts of short-term memory. In the Working Memory model, for example, the slave systems controlled by a central executive comprise a phonological store, supported by articulation,
Correspondence tot P. Walker, Faculty of Health, University of Central Lancashire, Preston PRl
2HE, UK.
OOOl-6918/93/$06.00 0 1993 - Elsevier Science Publishers B.V. All rights reserved

204 P. Walker et al. / Capacity of cisual short-term memory
and a visuo-spatial sketchpad (see Baddeley 1990). However, while considerable progress has been made in understanding phonological short-term memory, relatively little is known about its visuo-spatial counterpart. One reason for this disparity is that people prefer to rely heavily on speech-based processes, especially when undertaking the range of tasks devised by experimental psychologists. Because of this, researchers intent on investigating visual memory have had to devise methods to minimise the contribution of speech. One method involves presenting ‘non-nameable’ visual patterns as to-be-remembered stim- uli (e.g. Broadbent and Broadbent 1981; O’Connor and Hermelin 1978; Phillips and Christie 1977); another requires subjects to sup- press their articulation during the memory task (Broadbent and Broadbent 1981; Healy 1975; Kroll 1975; Stefurak and Boynton 1986). Once attempts have been made to minimise the contribution from speech, confirmation that visual memory has mediated performance can be sought in a number of ways. For example, it can be determined if performance is sensitive to the visual similarity of the to-be-remem- bered items (Broadbent and Broadbent 1981; Brown 1977; Conrad 1972; Frost 1972; Hayes and Schulze 1977; Hitch et al. 1988; 1989).
Although relatively little is known about visual short-term memory, several of its basic functional characteristics have been identified. For example, it seems to be more involved in preserving the spatial than the temporal features of stimuli (den Heyer and Barratt 1971; Healy 1975; Healy et al. 1992; Meudell 1972; Murray and Newman 1973). Thus, the sketchpad in working memory supports the spatial localisa- tion of items and is particularly susceptible to interference from secondary tasks that have a visuo-spatial, rather than a purely visual component (see Baddeley 1990). In addition, visual short-term mem- ory appears to be very vulnerable to the presentation of a subsequent stimulus, to an extent suggesting that its capacity may be limited to a single item (Phillips and Christie 1977). Thus, when subjects’ memory for short sequences of non-nameable matrix patterns is probed, recog- nition of the final pattern can be excellent while memory for all other patterns is extremely poor (Phillips and Christie 1977). Although Phillips and Christie (1977) argued for the involvement of a central processor in the retention of the final item, Logie et al. (1990) have demonstrated that memory for a single non-nameable pattern is much more sensitive to secondary tasks having specifically visuo-spatial components than to tasks that compete for central resources only.

P. Walker et al. / Capacity of visual short-term memory 205
Despite an apparently clear picture regarding the basic characteris- tics of visual short-term memory, doubts have remained on at least one fundamental point: It remains uncertain whether visual short-term memory is limited to preserving information about a single item. That it is not is suggested by the fact that Phillips and Christie (1977) observed levels of performance on pre-recency items that were above chance. Indeed, Phillips and Christie may have underestimated the potential of visual short-term memory by presenting their matrix patterns in the same spatial location. It is known, for instance, that the contribution of visual memory can be enhanced by presenting items in separate spatial locations (e.g. Broadbent and Broadbent 1981; Walker 19781.
On the other hand, Broadbent and Broadbent (1981) have pro- posed that visual short-term memory is limited to preserving informa- tion about a single item. They argue that a significant number of the matrix patterns used by Phillips and Christie will have contained familiar shapes, despite being generated on a random basis. They go on to suggest that memory for pre-recency items will reflect subjects’ use of phonological codes for patterns containing a familiar shape. In addition, it is possible that memory for pre-recency items is based on semantic codes. Avons and Phillips (1987) have recently demonstrated that the semantic categorisation of familiar shapes contained within random matrices can make a major contribution to memory for pre-recency items. It remains uncertain, therefore, whether visual memory is confined to a single item (typically the final item) in a series.
The three experiments reported here seek to clarify the contribu- tion of visual memory to the short-term retention of a series of items. A spatial localisation task is employed in which four shapes are presented in different spatial locations, and subjects are asked to recall the location of a probed shape. By using only four, clearly demarcated spatial locations, and by employing a shape probe to test subjects’ memory, the procedure focuses specifically on memory for a shape’s location and in doing so optimises the potential for a contribu- tion from visual memory. Furthermore, care is taken to dissociate the spatial and temporal aspects of the presentation sequence. Although in principle this is easily achieved with the probed location-recall procedure, it has seldom been a concern in previous investigations. For example, in several studies (Atkinson et al. 1964; Berth 1978;

206 P. Walker et al. / Capacity of c?sual short-term rnernoQ1
Bernbach 1967; Calfee et al. 1966; Hagen and Kingsley 1968; Hayes and Schulze 1977; Keely 1971; Siegel and Allik 1973) successive items have been placed in their locations systematically (typically in a left-right order), allowing subjects to infer an item’s spatial location on the basis of its temporal position in the sequence. In the task used here, the stimuli are always presented in a random spatio-temporal order, allowing memory for the spatial location of a shape to be assessed independently of memory for its temporal position.
In addition to encouraging a contribution from visual memory, steps are taken to preclude any influence from phonological coding. These steps include using quasi-random matrix patterns as to-be-re- membered stimuli (experiments 1 and 2) and having subjects suppress their articulation (experiments 2 and 3). The visual similarity of the to-be-remembered items is then manipulated to confirm the contribu- tion of visual memory to the localisation of pre-recency items (experi- ments 1 and 3). In addition, experiment 3 addresses the possibility that semantic coding could account for the results attributed to visual memory.
Experiment 1
In the first experiment, short sequences of non-nameable patterns were presented, in a random spatio-temporal order, with each pattern appearing at a distinct spatial location. After a brief interval subjects were asked to recall the location of a probed shape.
In common with previous observations of visual short-term memory, it was ex- pected that performance would be dominated by a recency effect focused on the final item. In order to explore whether visual memory can extend beyond the final item, sequences were made up of either visually similar or visually dissimilar patterns. If visual similarity disrupts memory for pre-recency patterns, this would be evidence that visual short-term memory is not restricted to a single pattern.
Response speed was monitored in addition to response accuracy. This permitted the issue of speed-accuracy trade-off to be addressed and also provided an alterna- tive, and potentially more sensitive measure of performance. Response speed, like accuracy, was expected to reveal a pronounced recency effect, with subjects respond- ing more quickly to tests of the final item (Phillips 1983).
Method
Subjects Eleven adults served as subjects. Across all of the experiments reported here, the
majority of subjects were drawn from the undergraduate population at the University

P. Walker et al. / Capacity of r:isual short-term memory 207
Fig. 1. The sixteen patterns used in experiment 1. The patterns in different subsets (rows) are
relatively distinct, while the four pattens within each subset are similar, varying on just two binary features.
of Central Lancashire. The remainder were drawn from friends and relatives of the authors.
Stimuli The stimuli were the sixteen quasi-random block patterns illustrated in fig. 1. They
comprised four visually similar subsets, the members of each subset being defined by two binary features, one in the top half of the patterns and one in the bottom half. For example, for the subset of patterns illustrated in the top row of fig. 1, the two features were the height of the to@ edge of the ‘hole’ in the centre of the pattern and the height of the stem at the base. The overall sizes of the patterns in the different subsets varied slightly, the maximum overall dimensions being 6 mm by 20 mm. The four locations at which the patterns were presented were marked out by a row of four outline rectangles, measuring 35 mm horizontally by 55 mm vertically, their adjacent edges being separated by 5 mm. An additional outline rectangle, within which the probe stimulus appeared, was located 15 mm directly below the centre of this row. These rectangles were continuously present on the screen and, like all the patterns, appeared as white figures on a black ground.
Design and procedure The experiment was conducted using a BBC (model B) microcomputer linked to a
colour monitor (Microvitec 452). Sequences of four visual stimuli were presented, with each stimulus appearing in a different spatial location on the monitor. The spatio- temporal order in which the stimuli appeared was randomly determined within

208 P. et al. Capacity of visual short-term memory
certain overall constraints. The stimuli were viewed from approximately 50 cm and were quasi-random block patterns, symmetrical about their vertical axis, presented in BBC teletext mode. Memory was tested after a short retention interval by presenting a probe stimulus in the neutral spatial location, below the horizontal array in which the list items appeared. The probe stimulus was identical to one of the list items and subjects had to indicate in which spatial location this list item had appeared. They responded to the probe via the computer keyboard, pressing keys S, F, J, and L to indicate successive spatial locations in a left-right directicn. They were encouraged to respond quickly and accurately, but to guess if necessary.
The lists of to-be-remembered items comprised four patterns which were equally likely to be taken either from the same subset or one each from the four different subsets. For a visually similar list one of the subsets of patterns was selected at random; for a visually distinct list one member from each subset was selected at random. When the patterns for a trial had been selected in this way, their temporal order of presentation was randomly determined. Each trial was initiated by the subject pressing the SPACE key. The trial commenced after a period of 2 s, with each pattern being visible for 1.0 s, a blank interstimulus interval (ISI) of 0.5 s separating successive patterns and a retention interval of 1.0 s following the final item in the sequence. The probe pattern then appeared and remained visible until the subject responded. For both levels of visual similarity, each serial position was tested once in association with each of the 24 (i.e. 4!) possible spatio-temporal sequences. In this way, all subjects completed a total of 192 trials.
Subjects were not provided with feedback regarding the speed or accuracy of their decisions. Their reaction times CRTS) were recorded to the nearest l/100 s and excessively slow correct RTs were identified as those in excess of two standard deviations above the subject’s mean correct RT. Prior to data summary and statistical analysis, the original values for these excessive RTs were replaced by the cut-off value. In the event, in no situation was the pattern of significant results changed by dealing with slow responses in this way.
Results
Accuracy and latency as a function of the temporal serial position of the probe are illustrated in fig. 2.
ANOVA of recall accuracy confirmed significant main effects of serial position, F(3, 30) = 12.77, MSe = 12.12, and similarity, F(1, 10) = 22.56, MSe = 12.73. The interaction between these two factors was also significant F(3, 30) = 4.34, MSe = 3.51. The serial position effect reflected pronounced recency, concentrated on the final item. The effect of visual similarity was examined at each serial position and, with the exception of serial position 3, performance was significantly worse for the visually similar lists, F(1, 40) = 25.4, 22.8, 3.78, and 5.28, respectively, for successive serial position, MSe = 5.81. The significant interaction was attributable to the greater effect of similarity at earlier serial positions. In interpreting the smaller effects of similarity for more recent items, and specifically with regard to the potentially distorting effects of performance at ceiling, it is worth noting that for only one subject was performance perfect at the final serial position, and this was in the visually dissimilar condition.

P. Walker et al. / Capacity of visual short-term memory 209
Gi i= K
1 2 3 4
serial position
L_Y
1.9 1.8
1.7
1.6 1.5
1 2 3 4
serial position
Fig. 2. Experiment 1: Serial position curves for the accuracy and latency of the probed
location-recall of visually similar (vis-sim) and visually dissimilar (vis-dissim) lists of patterns.
ANOVA of response latency confirmed the significance of the main effect of serial position, F(3, 30) = 17.48, MSe = 276.7. Neither the main effect of visual similarity, F(1, 10) < 1, MSe = 415.0, nor the interaction, F(3, 30) < 1, MSe = 206.9, was signifi- cant. The serial position effect reflected pronounced recency confined to the final item.
Error analysis
Because a response was required on every trial, a trivial interpretation of the recency effect has to be considered. It is possible that the relatively high hit rate for the final item reflects a strategy of selecting the location of this item whenever subjects were uncertain. To evaluate this possibility the distribution of error responses was examined for trials where the test item related to serial position 1, 2 or 3. If

210 P. Walker et al. / Capacity of csisual short-term memor);
subjects were guessing as described, these error responses should themselves yield a recency effect. However, a recency effect did not arise. The total numbers of errors mistakenly identified with serial positions 1, 2, 3, and 4 were 183, 210, 177, and 117, respectively. In the absence of a response bias the total number of responses directed at the final serial position should be 50% greater than the average number directed at each of the other positions. ’ Clearly, therefore, the recency effect observed for correct responses is not attributable to subjects selecting the most recently occupied location whenever they are uncertain. Indeed, it seems that subjects’ good memory for the last item led them not to select its location.
Replication
The results from this experiment were reproduced in an identical study in which the stimuli were slightly less discriminable (see fig. 3). In particular, stimulus similarity had an effect on accuracy but not speed, and this effect was more marked for pre-recency items. In more detail, for the eleven subjects involved in the replication, the average level of accuracy at each successive serial position was 62.5, 59.8, 61.4, and 84.1 percent, respectively, for visually dissimilar lists, and 39.4, 38.6, 46.6, and 71.6 percent, respectively, for visually similar lists. The main effects of serial position, F(3, 30) = 23.55, MSe = 9.62, and similarity, F(1, 10) = 26.98, MSc = 15.05, were both significant. Though the effect of similarity was greater on pre-recency items than on the final item, the overall interaction between serial position and similarity was not significant, F(3, 30) = 1.62, MSe = 5.02. The average RT at each successive serial position was 1.33, 1.36, 1.33, and 1.15 s, respectively, for visually dissimilar lists, and 1.49, 1.47, 1.35, and 1.20 s, respectively, for visually similar lists. The main effect of serial position was significant, F(3, 30) = 10.48, MSe = 296.6, while the main effect of similarity was not, F(1, 10) = 1.59, MSe = 944.7. There was no significant overall interaction between serial position and similarity, F(3, 30) = 0.83, MSe = 242.9.
Discussion
The serial position function that has previously characterised performance based on short-term visual memory was observed here. For both response accuracy and response speed this function was flat except for a recency effect concentrated on the final item. Moreover, it was confirmed that the effect with regard to accuracy did reflect superior memory for the final item, and was not attributable to a response bias whereby subjects selected the most recently occupied location whenever they were uncertain. Therefore, like item recognition and recall based on visual memory, the probed recall of spatial location appears vulnerable to the presentation of subsequent items.
It is difficult to determine what qualifies as the chance level of performance for pre-recency items. On one argument this should be 25% since there are four
’ While the spatial location of the fourth item is a possible error response to tests of any of the first three items, the location of any one of the first three items is a possible error response to
tests of just two of the first three items.

P. Walker et al. / Capacity of visual short-term memory 211
Fig. 3. The sixteen patterns used in the replication of experiment 1.
alternative responses. On another argument, however, if subjects are able to capitalise on their good memory for the final item when responding to a probe of an earlier item, as the pattern of the error responses suggests (see above), then something approaching 33% is more appropriate. Whatever argument is put forward, however, it is clear the level of performance on pre-recency items is above chance since visual similarity was able to bring performance down significantly. This key finding from the first experiment provides evidence that visual short-term memory is not confined to supporting the retention of a single item.
The detrimental effect of visual similarity mirrors the effect of phonological similarity on the temporally ordered recall of verbal items (Healy 1975; Watkins et al. 1974; Wickelgren 1965). In the same way that a phonological similarity effect confirms the contribution of phonological memory, so here the visual similarity effect confirms the involvement of visual memory in probed location-recall. The lack of any effect of visual similarity on response latency precludes an interpretation of the effect on accuracy in terms of speed-accuracy trade-off.
Predictability of the final item
When the number of to-be-remembered items in a list is fixed, superior recall of the final item may reflect features of the situation other than the item’s retention per se. For example, if subjects know that the last item is about to be presented, they may prepare themselves for it in a unique way. Although this is not consistent with subjects’ introspections concerning the passive way in which they approach the task, it has to be considered nevertheless. In a supplementary experiment this issue was addressed by examining location memory for non-nameable patterns in a situation where list length took on one of two values (5 or 6) and was varied unpredictably across trials. The results indicated quite clearly that the appearance of a final-item

212 P. Walker et al. / Capacity of visual short-term memory
recency effect in the probed location-recall task is not contingent on subjects being able to anticipate which item will be the final item. Not only did a recency effect appear in the context of the shorter lists, but its magnitude was no less than that observed for the longer lists.
Experiment 2
The patterns used in the first experiment were assumed to be meaningless, and hence non-nameable, with the effect that subjects would have been unable to use a phonological code to retain information about them or their spatial locations. In fact, however, resemblances can be seen between the patterns and a range of familiar objects (e.g. a perfume bottle) and on this basis it might be possible to assign verbal labels to them. The structure that the patterns possess by virtue of their vertical symmetry seems to guarantee a resemblance to some familiar object or other, and the repeated presentation of the small sample of patterns may have further facilitated the identification of such resemblances. It is conceivable, therefore, that the stimuli were amenable to phonological recoding, and that this contributed to performance. Fur- thermore, although an effect of visual similarity can be interpreted as confirming the contribution of visual memory, manipulations of similarity can confound an item’s visual features with its semantic and phonological features.
It is now accepted that articulation is required for the recoding and retention of visual information in phonological form. For example, the effects normally attributed to phonological recoding, such as those deriving from phonological similarity and word length, do not occur when subjects are prevented from articulating during the presentation of visual information (Baddeley et al. 1975; Estes 1973; Levy 1971; Murray 1968; Peterson and Johnson 1971; Richardson et al. 1980). Experiment 2 assessed the potential contribution of phonological coding to memory for the loca- tions of the quasi-random block patterns used in experiment 1 by sometimes requiring subjects to suppress their articulation.
Method
Subjects Eleven adults served as subjects.
Stimuli The experimental stimuli and all other aspects of the display were the same as in
experiment 1.
Design and procedure With the following two modifications, the basic procedure was identical to that
adopted experiment 1. First, there was no manipulation of visual similarity, only lists of dissimilar items were presented. Second, on half the trials, selected at random, a nonword appeared on the screen and cued subjects to suppress their articulation. For

P. Walker et al. / Capacity of visual short-term memory 213
0 .I 0.5 1 I 1 2 3 4 1 2 3 4
serial position serial position
Fig. 4. Experiment 2: Serial position curves for the accuracy and latency of the probed
location-recall of patterns in conditions requiring subjects to suppress their articulation (sup-
press) or not (control).
this purpose ten nonsense words were created whose pronunciation was considered to be quite demanding. These were PRIGGLE, WUMMLY, STIPPEL, KRIMMEL, SHUPPLE, PICKLY, LOBBEL, GRUDDLE, TRUMMET, and SHINNY. On the appropriate trials one of the nonsense words was chosen at random and presented at the top left corner of the screen. It remained visible for the full duration of the trial and subjects were instructed to begin repeating it to themselves, without pausing, from the moment they initiated the trial (2 s prior to the appearance of the first stimulus) to the point where they responded. They were asked to pronounce the nonsense word sufficiently loudly that it could just be heard by the experimenter when he/she sat next to them.
Every serial position was tested once for each of the 24 possible spatio-temporal sequences, making a total of 192 trials for each subject.
Accuracy and latency as a function of the temporal serial position of the probed item are illustrated in fig. 4.
The recall accuracy data were submitted to an ANOVA, with Serial Position and Suppression as within subjects factors. The main effect of serial position was signifi- cant, F(3, 30) = 17.73, MSe = 11.60, indicative of a marked final-item recency effect, but the main effect of suppression was not, F(1, 10) = 2.65, MSe = 7.57. The interac- tion between the two factors was insignificant, F(3, 30) = 1.72, MSe = 5.01. ANOVA of the response latency data confirmed the significance of the main effect of serial position, F(3, 30) = 8.91, MSe = 283.1, but failed to assign significance either to the main effect of suppression, F(1, 10) = 3.73, MSe = 264.9, or to the interaction be- tween the two factors, F(3, 30) < 1, MSe = 255.4. Although the main effect of suppression was almost significant, this reflects the fact that subjects responded more quickly in the suppression condition. The significance of serial position was again indicative of a final-item recency effect. Finally, the error responses for trials where the test item related to serial position 1, 2 or 3, did not yield a recency effect that

214 P. Wulker et al. / Capacity of visual short-term memov
would complicate the interpretation of the. pattern of correct responses. On the contrary, when subjects responded incorrectly on these trials they were less inclined to select the location of the final item than the location of an earlier item.
Discussion
The pattern of performance across serial position is essentially the same as that observed in experiment 1, being dominated by a final-item recency effect for both response accuracy and speed, suggesting that these are robust features of short-term memory for item location.
On the assumption that articulation is necessary for the translation of visual information into a phonological memory code (see Baddeley 1990), the absence of an effect of suppression indicates that such a code makes no contribution to the probed location-recall of quasi-random block patterns. The absence of a suppression effect here contrasts with the large and highly reliable effects of suppression on short-term memory for verbally recodable visual stimuli, such as letters and digits (e.g. Estes 1973; Levy 1971; Murray 1968).
The fact that a primacy effect was not evident in experiments 1 and 2 also argues against there having been a contribution from phonological memory. ’ That such a contribution would be expected to give rise to a primacy effect in probed location-re- call is suggested from two rather different sources. Bernbach (1967) presented 5-year-olds with short sequences of colour patches and then probed memory for the location of one of the colours. One group of subjects named the colours at presenta- tion while a second group did not. There was a pronounced recency effect for both groups, but primacy was present only in the group that named the stimuli. Similarly, Locke and Deck (1978) presented aphasic patients with short sequences of pictures and then probed memory for the location of one of them. For pictures that the patients could name, there was both reccncy and primacy, while for pictures the patients could not name there was recency but no primacy.
Experiment 3
Although suppressing articulation precludes the phonological recoding of visual information, it need not interfere with the semantic categorisation of familiar shapes and need not prevent such categorisation from supporting spatial localisation. For these reasons, articulatory suppression as a technique cannot address the potential confound between the visual similarity of items and their semantic features. Experi- ment 3 employs letters as the to-be-remembered stimuli in order to address these concerns about the potential contribution of semantic coding. The virtue of using letters for this purpose stems from the fact that their semantic features are relatively
’ The argument being put forward is that in the probed location-recall task subjects’ utilisation of speech-based processes can be expected to give rise to a primacy effect. It is not being implied
that in this or any other task a primacy effect can only arise from the involvement of speech.

P. Walker et al. / Capacity of Gual short-term memory 215
unambiguous and predetermined, and from the fact that their use allows visual similarity to be manipulated independently of semantic classification. Articulatory suppression was again used to preclude a contribution from phonological memory.
Method
Subjects Ten adults served as subjects in the main condition of the experiment in which
customised letters were used as stimuli. Eleven adults undertook the task in the control condition.
Stimuli
Experimental stimuli. The experimental stimuli comprised four relatively distinct subsets of visually similar letters (see fig. 5). The letters were created from the same block elements used to create the previous non-nameable patterns. They were designed so that the letters within a subset were endowed with a style that either exaggerated the visual features they already shared or provided them with features that were then the basis of their similarity. To give an indication of the sizes of the letters, the letter B measured 15 mm by 22 mm. More generally, the letters varied in width from 6 mm (F&L) to 17 mm (V, Q, &D, and in height from 20 mm (T, F, L, &J) to 24 mm (R, B, P, &S). Other aspects of the display, and in particular the demarca- tion of the four spatial locations at which the letters appeared, were exactly as in experiment 1.
Before accepting the letter designs for the experimental stimuli, and in an attempt to circumvent any problems associated with the differential encoding of visually
Y T
c I?
xv F 1
QG BP
x 1
0 s
Fig. 5. The sixteen, specially designed letters used as experimental stimuli in experiment 3. The
four letters within each subset (row) were created in a style that was the basis of their similar
appearance.

216 P. Walker et ul. / Capacity of Gsual short-term men-tory
Fig. 6. The sixteen letters used as control stimuli in experiment 3, reproduced in the standard
teletext font in which they appeared.
similar and dissimilar stimuli, a preliminary study required subjects to perform the probed location-recall task while at the same time attempting to name each letter as it appeared. Since main interest was in whether subjects would be able to name the letters, the requirement that each serial position was tested once for each of the 24 possible spatio-temporal sequences was relaxed. Instead, the spatio-temporal se- quence used on each of 64 trials was selected at random, though different serial positions were tested equally often. In the event, no subject made a single naming error, confirming that all the letters, whether part of a visually similar list or part of a visually dissimilar list, were encoded to the point where they could be discriminated from each other and named.
Control stimuli. Selection of the experimental letters and their inclusion in the different subsets could not be achieved on a random basis since it was necessary that

P. Walker et al. / Capacity of visual short-term memory 217
they could be made to appear visually similar. Other aspects of the letters, therefore, were confounded with their membership in the subsets. For example, one of the subsets comprised the letters Q, C, D, and G, and it could be argued that the phonological features shared by three of these letters would make remembering their locations difficult. The inclusion of a control condition assessed the contribution that any non-visual features might make to an apparent visual similarity effect in the experimental condition. For this purpose the same letters were presented in the standard BBC teletext font, thus removing the visual features that had been grafted on to them by virtue of their customised design. Although the change in the appearance of the letters is not guaranteed to remove all forms of visual similarity within the subsets, it does remove the most salient features on which the similarity manipulation is based. Fig. 6 is a reproduction of the letters, arranged in their subsets, as they appeared on the computer screen. All the letters measured 6 mm by 7 mm.
There is a further confound with the similarity manipulation that is addressed by the inclusion of this control condition. This confound relates the fact that successive items are more predictable in a similar list because the set from which a sequence of similar items is selected is smaller than the set from which a sequence of dissimilar items is selected. For example, the identity of the final item in a visually similar list is fully determined by the previous items, whereas the final item in a dissimilar sequence can be any one of the four items from the subset that has yet to be sampled. If this further confound is responsible for an apparent effect of similarity in the experimen- tal condition, then changing the font for the control condition should have no effect on the results.
Design and procedure
The procedure employed in experiment 1 was adopted, with two modifications. First, all subjects were required to suppress their articulation on every trial, in the same way as in experiment 2. Second, the rate of presentation of the stimuli was doubled in an attempt to ensure that subjects were unable to name or rehearse the stimuli covertly. Each stimulus appeared for 0.5 s, the ISI was 0.25 s, and the retention interval remained at 1.0 s.
Results
The two sets of results from this experiment are illustrated in figs. 7 and 8, for the experimental and control stimuli, respectively.
Experimental stimuli
ANOVA of recall accuracy, with Serial Position and Similarity as within subjects factors, confirmed the significance of the main effects of similarity, F(1, 9) = 19.38, MSe = 10.73, and serial position, F(3, 27) = 22.27, MSe = 17.27. The interaction between these two factors was also significant, F(3, 27) = 4.35, MSe = 3.44. The serial position effect reflected recency which, though concentrated on the final item, extended to the penultimate item. The effect of visual similarity was examined at each
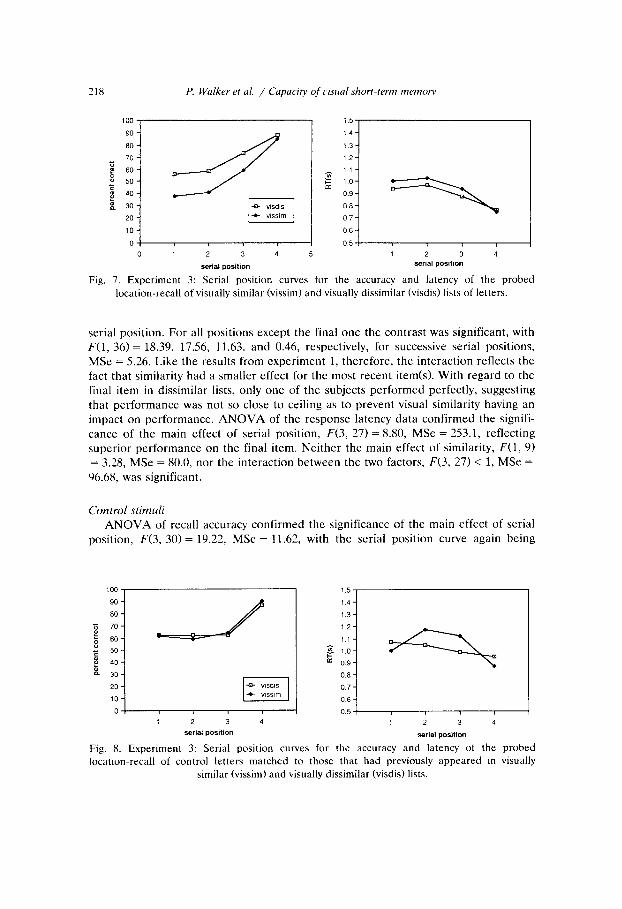
218 P. Walker et al. / Capacity of t,isual short-term memos
‘iI 0 1 2 3 4 5
serial position
Fig. 7. Experiment 3: Serial position curves for the accuracy and latency of the probed location-recall of visually similar (vissim) and visually dissimilar (visdis) lists of letters.
serial position. For all positions except the final one the contrast was significant, with F(1, 36) = 18.39, 17.56, 11.63, and 0.46, respectively, for successive serial positions, MSe = 5.26. Like the results from experiment 1, therefore, the interaction reflects the fact that similarity had a smaller effect for the most recent item(s). With regard to the final item in dissimilar lists, only one of the subjects performed perfectly, suggesting that performance was not so close to ceiling as to prevent visual similarity having an impact on performance. ANOVA of the response latency data confirmed the signifi- cance of the main effect of serial position, F(3, 27) = 8.80, MSe = 253.1, reflecting superior performance on the final item. Neither the main effect of similarity, F(1, 9) = 3.28, MSe = 80.0, nor the interaction between the two factors, F(3, 27) < 1, MSe = 96.68, was significant.
Control stimuli
ANOVA of recall accuracy confirmed the significance of the main effect of serial position, F(3, 30) = 19.22, MSe = 11.62, with the serial position curve again being
100 ,
0 I 1 2 3 4 1 2 3 4
serial position
Fig. 8. Experiment 3: Serial position curves for the accuracy and latency of the probed location-recall of control letters matched to those that had previously appeared in visually
similar (vissim) and visually dissimilar (visdis) lists.

P. Walker et al. / Capacity of Gsual short-term memory 219
dominated by a pronounced recency effect concentrated on the final item. Neither
the main effect of similarity, F(1, 10) < 1, MSe = 4.05, nor the interaction between these two factors F(3, 30) < 1, MSe = 3.76, was significant. ANOVA of the response latency data confirmed the significance of the main effect of serial position, F(3, 30) = 7.66, MSe = 326.0. Again, however, neither the effect of similarity, F(1, 10) = 2.41, MSe = 219.7, nor the interaction between the two factors, F(3, 30) = 2.79, MSe = 164.1, was significant. Finally, for experimental and control stimuli alike, error responses to tests of serial positions 1, 2 or 3, did not yield a recency effect to complicate the interpretation of the pattern of correct responses. Rather, when subjects responded incorrrectly to these test items they were less inclined to select the location of the final item than the location of an earlier item.
Discussion
This experiment addressed the possibility that familiar shapes are perceived in the quasi-random matrix patterns and that their semantic categorisation supports probed location-recall. Using letters as to-be-remembered stimuli allowed semantic coding to be eliminated from the equation because their semantic features are relatively unambiguous and predetermined, and because the visual similarity of different letters can be manipulated independently of their semantic classification.
The results for the customised letters closely resemble those obtained with non- nameable patterns (experiment 1). Not only are the effects of serial position again evident in measures of both accuracy and speed, being dominated by recency, but the effects of visual similarity are again confined to accuracy. Furthermore, the effect of visual similarity extends throughout the list, though less significantly for more recent items. Although the same serial position effects characterise performance with the control stimuli, there are no differences between the two types of list. It is clear, therefore, that the effects of similarity on the location-recall of the experimental stimuli did emanate from their visual features. These data argue strongly that short-term memory for location is mediated by a visual store which is not confined to the most recently presented item, as confirmed by the extension of the visual similarity effect to pre-recency items.
It is difficult to determine how close to floor subjects’ performance was for the first two items in visually similar lists. Certainly, if subjects can exploit their memory for the location of the final item when responding to a probe of an earlier one, then the level of performance is close to floor. To this extent the results imply that when articulation is suppressed only visual codes are available to support probed location- recall. The overlap in the results in experiments 1 and 3 (e.g. the fact that visual similarity influenced response accuracy but not response speed) confirms that visual short-term memory makes the same contribution to the spatial localisation of famil- iar/nameable stimuli as it does to meaningless stimuli when phonological recoding is precluded by articulatory suppression.
Spatial location and the distribution of error responses
Because of the nature of the displays in the present experiments, it is possible to examine the dependence of performance on spatial location independently of its

220 P. Walker et al. / Capacity of visual short-term memory
sensitivity to temporal position. The effect of spatial location and the spatial distribu- tion of errors relative to the correct (probed) location, were analysed after combining the results from all the experiments, including the replication of experiment 1. In total, therefore, 53 subjects contributed to the analysis.
Spatial location. For each subject the number of correct responses was determined (maximum = 48) in relation to each of the four spatial locations. Across all subjects and conditions the mean number of correct responses at successive (left-right) locations was 29.3, 30.6, 29.8, and 29.0, respectively. ANOVA of the number of correct responses, with Spatial Location as the only within-subjects factor, failed to reveal a significant effect of location, F(3, 156) = 1.47, MSe = 17.09.
Spatial distribution of errors. After determining the response matrix based on spatial location for each subject, the mean number of error responses at each level of separation from the correct location was calculated. Because spatial location itself did not influence performance, interpretation of the results is not complicated by the fact that spatial separation is confounded with spatial location. (For example, errors separated from the probed location by three positions must necessarily be a response to a probe of one of the extreme spatial locations.) The different levels of separation were 1, 2, and 3 positions from the probed location, and the mean number of errors associated with each level was 6.9, 5.3, and 5.2, respectively. When the three average values for each subject were submitted to ANOVA, with Spatial Separation as the only within subjects factor, the effect of separation was shown to be highly significant, F(2, 104) = 22.94, MSe = 2.10. From the mean values it is clear that subjects were more likely to select in error an immediately adjacent location than a more distant location. There was no difference in the probability with which subjects selected a location that was three rather than two positions from the probed location. In conclusion, the fact that errors did reflect something of the spatial location of the probed item indicates that the preservation of spatial location was not all-or-none but was coarse enough to confuse immediately adjacent locations.
General discussion
By using only four, clearly demarcated spatial locations and by employing a probe to test memory, the present task focused specifi- cally on the retention of a shape’s location rather than on memory for the shapes or locations themselves. In doing so, the task was assumed to facilitate a contribution from visual memory, care being taken to dissociate the spatial and temporal aspects of the presentation se- quences. To further guarantee the involvement of visual memory, non-nameable shapes were used as the to-be-remembered stimuli in the first two experiments.

P. Walker et al. / Capacity of cisual short-term memory 221
Several aspects of the present results converge on the view that these measures were successful. First, serial position had the same effect as in previous studies that have isolated the contribution of short-term visual memory to item recognition and reproduction (e.g. Phillips and Christie 1977; Phillips 1983). The absence of a primacy effects confirms the non-involvement of phonological memory since it is known that such an effect arises in situations where articulation supports the short-term retention of visual stimuli (e.g. Bernbach 1967; Locke and Deck 1978). Second, in experiment 2 articulatory suppression failed to influence memory for the location of matrix patterns. On the understanding that articulation is necessary for the phonological recoding of visual information, this major alternative to a visuo-spatial code is therefore eliminated. Finally, the fact that in- creasing the visual similarity of the to-be-remembered items had a detrimental effect on performance confirms the involvement of visual memory.
The results from supplementary studies, reported in the context of each experiment, confirm that the recency effect did emanate from enhanced memory for the final item and did not reflect a response bias whereby subjects selected the most recently occupied spatial location whenever they were uncertain. Equally, it was shown that the effect did not come about because subjects prepared themselves differently for what they knew would be the final item.
The recency effect observed here is unlikely to reflect the loss of perceptual information about all but the final item. The rate of presentation of the list items was not so fast as to give rise to perceptual masking. Indeed, when subjects were required to name successive list items in the context of experiment 3, they were able to do so without error. In addition, the visual short-term memory under observation in the present experiments is distinct from visual sensory storage (Phillips 19831, even with regard to the final item. Not only was the delay before the probe item was presented out of range (Phillips 19831, but the fact that the probe was presented in a different spatial location than the list items would preclude a contribution from sensory storage.
Broadbent and Broadbent (1981) interpreted the pronounced re- cency effect found in the visual short-term memory task as implying that the system has a very limited capacity, restricted to a single item. In particular, they suggested that memory for earlier items was medi-

222 P. Walker et al. / Capacity of visual short-term memory
ated by non-visual codes. However, aspects of the present results indicate that visual short-term memory is capable of supporting the retention of several items. First, despite any uncertainties about an appropriate baseline level of performance in the probed location-re- call task, pre-recency performance was almost certainly above such a level. This is particularly evident in the case of experiment 2 and in the case of the visually dissimilar conditions of experiments 1 and 3. Moreover, the level of pre-recency performance was higher than would be predicted by the argument that visual short-term memory is restricted to preserving a single item and that an earlier item can be retained only on those few occasions where the final item is not. This argument is most clearly refuted by the results from the visually dissimilar conditions of experiments 1 and 3. However, the clearest evidence that visual short-term memory can support the retention of several items comes with the observation that visual similarity was able to bring pre-recency performance down significantly. Experi- ments 2 and 3 were crucial in ruling out the possibility that this similarity effect was an artefact of the use of phonological or semantic coding. Finally, the fact that error responses, the bulk of which were made to early list items, were more likely to be directed at a spatial location immediately adjacent to the probed location, is consistent with the involvement of visual memory and with its capacity to support several items.
The visual short-term memory mediating probed location-recall appears to make the same contribution to the retention of meaning- ful/ nameable items and meaningless/ non-nameable items. The equivalence of this contribution is apparent in the finer points of performance, including the selective effect of visual similarity on response accuracy and not speed. Interestingly, the equivalence of the results for the different categories of stimulus indicates that articula- tory suppression is particularly useful as a technique because it can prevent a contribution from phonological memory without, at the same time, interfering with the normal operation of visual memory.
The present study did not aim to bear directly on the nature of the recency effect in visual short-term memory. However, since visual similarity had a greater detrimental effect on location recall for earlier list items, some support is provided for Broadbent and Broadbent’s (1981) overwriting hypothesis. According to this hypothesis visual memory for a stimulus suffers interference from the presentation of a

P. Walker et al. / Capacity of visual short-term memory 223
subsequent stimulus to the extent they share visual features. The interference is asymmetric insofar as it is retroactive, the later stimu- lus having a detrimental effect on retention of the preceding stimulus and not vice versa.
References
Atkinson, R.C., D.N. Hansen and H.A. Bernbach, 1964. Short-term memory with young children. Psychonomic Science 1, 255-256.
Avons, S.E. and W.A. Phillips, 1987. Representation of matrix patterns in long- and short-term
visual memory. Acta Psychologica 65, 227-246.
Baddeley, A.D., 1990. Human memory: Theory and practice. Hillsdale, NJ: Erlbaum.
Baddeley, A.D., N. Thomson and M. Buchanan, 1975. Word length and the structure of short-term memory. Journal of Verbal Learning and Verbal Behavior 14, 575-589.
Berth, D.R., 1978. The role of spatial cues in the probe-type serial memory task. Child Development 49, 749-754.
Bernbach, H.A., 1967. The effect of labels on short-term memory for colors with nursery school children. Psychonomic Science 7, 149-150.
Broadbent, D.E. and M.H.P. Broadbent, 1981. Recency effects in visual memory. Quarterly Journal of Experimental Psychology 33A, l-15.
Brown, R.M., 1977. An examination of visual and verbal coding processes in preschool children.
Child Development 48, 38-45.
Calfee, R.C., E.M. Hetherington and P. Waltzer, 1966. Short-term memory in children as a function of display size. Psychonomic Science 4, 153-154.
Conrad, R., 1972. Short-term memory in the deaf: A test for speech coding. British Journal of Psychology 63, 173-180.
den Heyer, K. and B. Barrett, 1971. Selective loss of visual and verbal information in STM by
means of visual and verbal interpolated tasks. Psychonomic Science 25, 100-102.
Estes, W.K., 1973. Phonemic coding and rehearsal in short-term memory for letter strings. Journal of Verbal Learning and Verbal Behavior 12, 360-372.
Frost, N., 1972. Encoding and retrieval in visual memory tasks. Journal of Experimental Psychology 95, 317-326.
Hagen, J.W. and P.R. Kingsley, 1968. Labelling effects in short-term memory. Child Develop- ment 39, 113-131.
Hayes, D.S. and S.A. Schulze, 1977. Visual encoding in preschoolers’ serial retention. Child Development 48, 1066-1070.
Healy, A.F., 1975. Coding of temporal-spatial patterns in short-term memory. Journal of Verbal Learning and Verbal Behavior 14, 481-495.
Healy, A.F., T.F. Cunningham, A.T. Gesi, R.E. Till and L.E. Bourne, 1992. ‘Comparing short-term recall of item, temporal, and spatial information in children and adults’. In: W.E.
Hockley and S. Lewandowsky (eds.), Relating theory and data: Essays on human memory in Honor of Bennet B. Murdock (pp. 127-154). Hillsdale, NJ: Erlbaum.
Hitch, G.J., MS. Halliday, A.M. Schaafstal and J.M.C. Schraagen, 1988. Visual working memory in young children. Memory and Cognition 16, 120-132.
Hitch, G.J., M.E. Woodin and S. Baker, 1989. Visual and phonological components of working memory in children. Memory and Cognition 17, 175-185.

224 P. Walker et al. / Capacity of visual short-term memory
Keely, K., 1971. Age and task effects in short-term memory of children. Perception & Psy-
chophysics 9, 480-482.
Kroll, N.E.A., 1975. ‘Visual short-term memory’. In: D. Deutsch and J.A. Deutsch (eds.),
Short-term memory (pp. 153-179). London: Academic Press.
Levy, B.A., 1971. The role of articulation in auditory and visual short-term memory. Journal of Verbal Learning and Verbal Behavior 10, 123-132.
Locke, J.L. and J.W. Deck, 1978. Retrieval failure, rehearsal deficiency, and short-term memory
loss in the aphasic adult. Brain and Language 5, 227-235.
Logic, R.H., G.M. Zucco and A.D. Baddeley, 1990. Interference with visual short-term memory.
Acta Psychologica 75, 55-74.
Meudell, P.R., 1972. Short-term visual memory: Comparative effects of two types of distraction
on the recall of visually presented verbal and nonverbal material. Journal of Experimental
Psychology 94, 244-247.
Murray, D.J., 1968. Articulation and acoustic confusability in short-term memory. Journal of
Experimental Psychology 78,679-684.
Murray, D.J. and F.M. Newman, 1973. Visual and verbal coding in short-term memory. Journal
of Experimental Psychology 100, 58-62.
O’Connor, N. and B. Hermelin, 1978. Seeing and hearing and space and time. London:
Academic Press.
Peterson, L.R. and ST. Johnson, 1971. Some effects of minimizing articulation on short-term
retention. Journal of Verbal Learning and Verbal Behavior 10, 346-354.
Phillips, W.A., 1983. Short-term visual memory. Philosophical Transactions of the Royal Society
of London B302, 295-309.
Phillips, W.A. and D.F.M. Christie, 1977. Components of visual memory. Quarterly Journal of
Experimental Psychology 29, 117-133. Richardson, J.T.E., D.E. Greaves, and M.M.C. Smith, 1980. Does articulatoty suppression
eliminate the phonemic similarity effect in short-term recall? Bulletin of the Psychonomic
Society 16, 417-420. Siegel, A.W. and J.P. Allik, 1973. A developmental study of visual and auditory short-term
memory. Journal of Verbal Learning and Verbal Behavior 12, 409-418.
Stefurak, D.L. and R.M. Boynton, 1986. Independence of memory for categorically different
colors and shapes. Perception and Psychophysics 39, 164-174. Walker, P., 1978. Short-term visual memory: The importance of the temporal and spatial
separation of successive stimuli. Quarterly Journal of Experimental Psychology 30, 665-679.
Watkins, M.J., O.C. Watkins and R.G. Crowder, 1974. The modality effect in free and serial
recall as a function of phonological similarity. Journal of Verbal Learning and Verbal
Behavior 13, 430-447. Wickelgren, W.A., 1965. Short-term memory for phonemically similar lists. American Journal of
Psychology 78, 567-574.


















