
The cartilaginous skeleton of an elasmobranch fish does ...
8
Matrix Biology 23 (2004) 15–22 0945-053X/04/$30.00 2004 Elsevier B.V. yInternational Society of Matrix Biology. All rights reserved. doi:10.1016/j.matbio.2004.02.001 The cartilaginous skeleton of an elasmobranch fish does not heal Doreen E. Ashhurst* Department of Anatomy, St. George’s Hospital Medical School, Tooting, London SW17 0RE, UK Received 11 August 2003; received in revised form 2 February 2004; accepted 2 February 2004 Abstract The inability of articular cartilage to heal satisfactorily is becoming, with ageing populations, an important medical problem. One question that has not been raised is whether a mechanism for the repair of cartilage evolved in animals with cartilaginous skeletons. Fin rays of dogfish were cut and the fish maintained for up to 6 months. The initial inflammatory reaction around the cut rays lasts for 2 weeks. By 4 weeks the cut ends are covered by fibrous tissue. At 12 weeks some areas of cartilage-like tissue are developing. Development of these areas continues and at 26 weeks large chondrocyte-like cells are surrounded by matrix. This tissue is in regions of poor vascularity. It does not have the typical appearance of hyaline cartilage, nor is it integrated with the cartilage of the fin rays. No changes in the cut surfaces of the fin rays are observed at any time. It is concluded that no mechanism has evolved in the elasmobranch fishes for the repair of their cartilaginous skeleton. This is discussed in relation to previous investigations of the reactions of cartilage to injury in embryonic, neonatal and adult tissues of higher vertebrates. 2004 Elsevier B.V.yInternational Society of Matrix Biology. All rights reserved. Keywords: Elasmobranch fish; Cartilage; Cartilage repair; Fracture 1. Introduction An important problem in medicine at the present time is the inability of articular cartilage to heal satisfactorily (Frenkel and Di Cesare, 1999; Temenoff and Mikos, 2000). The prognosis after damage to an articular cartilage, or the onset of osteoarthritis, is poor. In contrast, bone heals very well. The ability of bone to heal so well is attributable to its continuous remodelling, which is brought about by two cells, osteoclasts and osteoblasts; new bone matrix is laid down on bone surfaces. In cartilage, turnover is intrinsic; the chondro- cytes produce enzymes that breakdown the matrix, while at the same time, they secrete more matrix molecules. There is no cell in cartilage that is equivalent to the osteoclast, nor is there a mechanism whereby new matrix is added by apposition to pre-existing matrix. These facts have implications for healing. Many experiments have been performed to induce the repair of damaged articular cartilage. New tissue is formed, but it is usually fibrocartilaginous. It does not integrate with the pre-existing matrix and is eventually *Tel.: q44-20-8725-5224; fax: q44-20-8725-3326. E-mail address: [email protected] (D.E. Ashhurst). sloughed off. Previous studies of mature amphibian and avian tissues indicate that there is no mechanism for cartilage repair in these groups (Subba Rao, 1954; Goss, 1977; Hall and Hanken, 1985). An early investigation of chicken embryos showed that healing occurs only when the cartilage in the limb rudiments is very imma- ture, that is, when the cells are actively dividing and there is little matrix (Niven, 1931). A question that has never been asked is whether during evolution a mechanism for the repair of cartilage developed. It appeared that an answer to this question might be found in the elasmobranch fishes, which have a cartilaginous skeleton. Lesions were, therefore, made in the fin rays of the dogfish and examined at periods up to 6 months later. The fin rays were chosen as the site for study because of their easy operative accessibil- ity and the minimal effect of the trauma on the fish. No evidence of true healing was observed. 2. Results 2.1. Normal fin rays The fin rays are cylindrical rods approximately 0.5– 0.75 mm in diameter (Fig. 1). They are composed of
Transcript of The cartilaginous skeleton of an elasmobranch fish does ...

Matrix Biology 23 (2004) 15–22
0945-053X/04/$30.00� 2004 Elsevier B.V.yInternational Society of Matrix Biology. All rights reserved.doi:10.1016/j.matbio.2004.02.001
The cartilaginous skeleton of an elasmobranch fish does not heal
Doreen E. Ashhurst*
Department of Anatomy, St. George’s Hospital Medical School, Tooting, London SW17 0RE, UK
Received 11 August 2003; received in revised form 2 February 2004; accepted 2 February 2004
Abstract
The inability of articular cartilage to heal satisfactorily is becoming, with ageing populations, an important medical problem.One question that has not been raised is whether a mechanism for the repair of cartilage evolved in animals with cartilaginousskeletons. Fin rays of dogfish were cut and the fish maintained for up to 6 months. The initial inflammatory reaction around thecut rays lasts for 2 weeks. By 4 weeks the cut ends are covered by fibrous tissue. At 12 weeks some areas of cartilage-like tissueare developing. Development of these areas continues and at 26 weeks large chondrocyte-like cells are surrounded by matrix.This tissue is in regions of poor vascularity. It does not have the typical appearance of hyaline cartilage, nor is it integrated withthe cartilage of the fin rays. No changes in the cut surfaces of the fin rays are observed at any time. It is concluded that nomechanism has evolved in the elasmobranch fishes for the repair of their cartilaginous skeleton. This is discussed in relation toprevious investigations of the reactions of cartilage to injury in embryonic, neonatal and adult tissues of higher vertebrates.� 2004 Elsevier B.V.yInternational Society of Matrix Biology. All rights reserved.
Keywords: Elasmobranch fish; Cartilage; Cartilage repair; Fracture
1. Introduction
An important problem in medicine at the present timeis the inability of articular cartilage to heal satisfactorily(Frenkel and Di Cesare, 1999; Temenoff and Mikos,2000). The prognosis after damage to an articularcartilage, or the onset of osteoarthritis, is poor. Incontrast, bone heals very well. The ability of bone toheal so well is attributable to its continuous remodelling,which is brought about by two cells, osteoclasts andosteoblasts; new bone matrix is laid down on bonesurfaces. In cartilage, turnover is intrinsic; the chondro-cytes produce enzymes that breakdown the matrix, whileat the same time, they secrete more matrix molecules.There is no cell in cartilage that is equivalent to theosteoclast, nor is there a mechanism whereby new matrixis added by apposition to pre-existing matrix. Thesefacts have implications for healing.Many experiments have been performed to induce the
repair of damaged articular cartilage. New tissue isformed, but it is usually fibrocartilaginous. It does notintegrate with the pre-existing matrix and is eventually
*Tel.: q44-20-8725-5224; fax:q44-20-8725-3326.E-mail address: [email protected](D.E. Ashhurst).
sloughed off. Previous studies of mature amphibian andavian tissues indicate that there is no mechanism forcartilage repair in these groups(Subba Rao, 1954; Goss,1977; Hall and Hanken, 1985). An early investigationof chicken embryos showed that healing occurs onlywhen the cartilage in the limb rudiments is very imma-ture, that is, when the cells are actively dividing andthere is little matrix(Niven, 1931).A question that has never been asked is whether
during evolution a mechanism for the repair of cartilagedeveloped. It appeared that an answer to this questionmight be found in the elasmobranch fishes, which havea cartilaginous skeleton. Lesions were, therefore, madein the fin rays of the dogfish and examined at periodsup to 6 months later. The fin rays were chosen as thesite for study because of their easy operative accessibil-ity and the minimal effect of the trauma on the fish. Noevidence of true healing was observed.
2. Results
2.1. Normal fin rays
The fin rays are cylindrical rods approximately 0.5–0.75 mm in diameter(Fig. 1). They are composed of
16 D.E. Ashhurst / Matrix Biology 23 (2004) 15–22
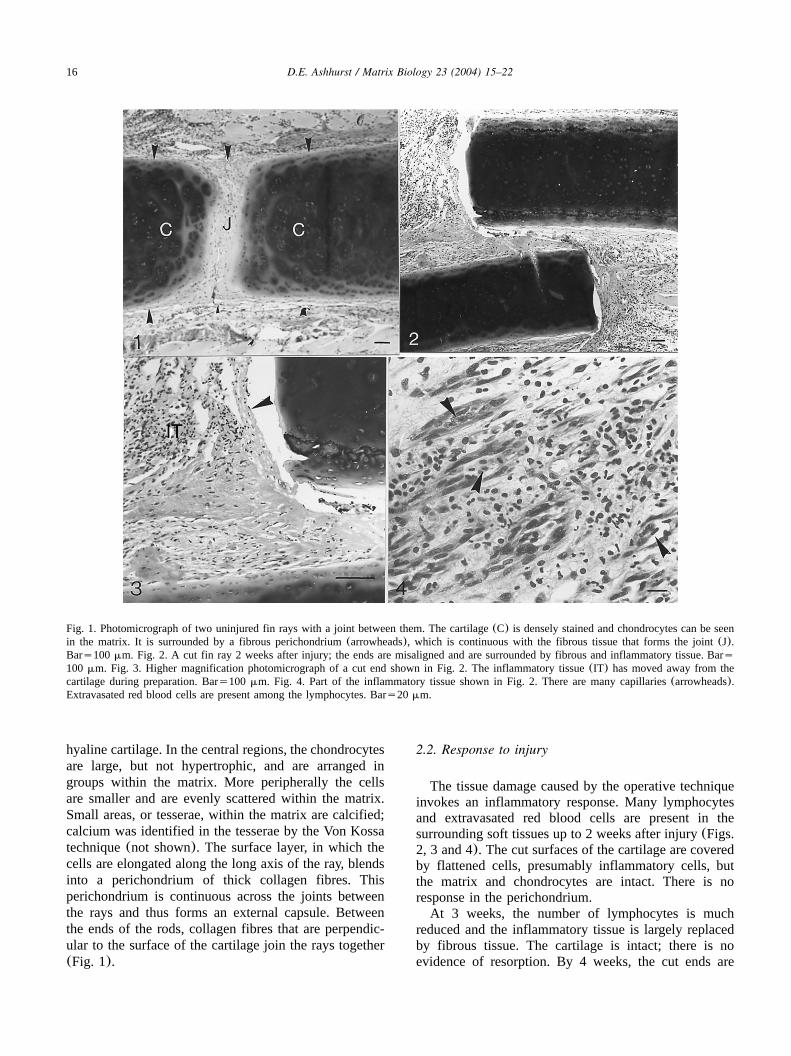
Fig. 1. Photomicrograph of two uninjured fin rays with a joint between them. The cartilage(C) is densely stained and chondrocytes can be seenin the matrix. It is surrounded by a fibrous perichondrium(arrowheads), which is continuous with the fibrous tissue that forms the joint(J).Bars100mm. Fig. 2. A cut fin ray 2 weeks after injury; the ends are misaligned and are surrounded by fibrous and inflammatory tissue. Bars100 mm. Fig. 3. Higher magnification photomicrograph of a cut end shown in Fig. 2. The inflammatory tissue(IT) has moved away from thecartilage during preparation. Bars100 mm. Fig. 4. Part of the inflammatory tissue shown in Fig. 2. There are many capillaries(arrowheads).Extravasated red blood cells are present among the lymphocytes. Bars20 mm.
hyaline cartilage. In the central regions, the chondrocytesare large, but not hypertrophic, and are arranged ingroups within the matrix. More peripherally the cellsare smaller and are evenly scattered within the matrix.Small areas, or tesserae, within the matrix are calcified;calcium was identified in the tesserae by the Von Kossatechnique(not shown). The surface layer, in which thecells are elongated along the long axis of the ray, blendsinto a perichondrium of thick collagen fibres. Thisperichondrium is continuous across the joints betweenthe rays and thus forms an external capsule. Betweenthe ends of the rods, collagen fibres that are perpendic-ular to the surface of the cartilage join the rays together(Fig. 1).
2.2. Response to injury
The tissue damage caused by the operative techniqueinvokes an inflammatory response. Many lymphocytesand extravasated red blood cells are present in thesurrounding soft tissues up to 2 weeks after injury(Figs.2, 3 and 4). The cut surfaces of the cartilage are coveredby flattened cells, presumably inflammatory cells, butthe matrix and chondrocytes are intact. There is noresponse in the perichondrium.At 3 weeks, the number of lymphocytes is much
reduced and the inflammatory tissue is largely replacedby fibrous tissue. The cartilage is intact; there is noevidence of resorption. By 4 weeks, the cut ends are

17D.E. Ashhurst / Matrix Biology 23 (2004) 15–22
Fig. 5. The cut ends of a fin ray 4 weeks after injury. They are surrounded by fibrous tissue(FT). The cut surfaces of the fin rays are unchanged(arrows). Bars100mm. Fig. 6. A cut end shown in Fig. 5 at higher magnification to show that it has not been eroded(arrow) Bars100mm.Fig. 7. An injury site at 12 weeks. The fragments of fin ray are surrounded by fibrous tissue. The cut surfaces are indicated by arrows. The gapbetween two surfaces is filled by cells. Bars100 mm. Fig. 8. At higher magnification the cells in the gap on Fig. 7 appear chondrocyte-like.Bars20 mm.
covered by fibrous tissue(Figs. 5 and 6) and there isno change at 8 weeks. A few lymphocytes may still bepresent.By 12 weeks, there may be areas of the fibrous tissue
in which the cells appear round and similar to chondro-cytes(Figs. 7 and 8). Where there is a cut that has notpenetrated right through the ray, or the cut surfacesremain parallel and close together, the gap may be filledby round chondrocyte-like cells(Fig. 8). No bloodvessels penetrate the gaps.The tissues at 26 weeks remain similar. The cut ends
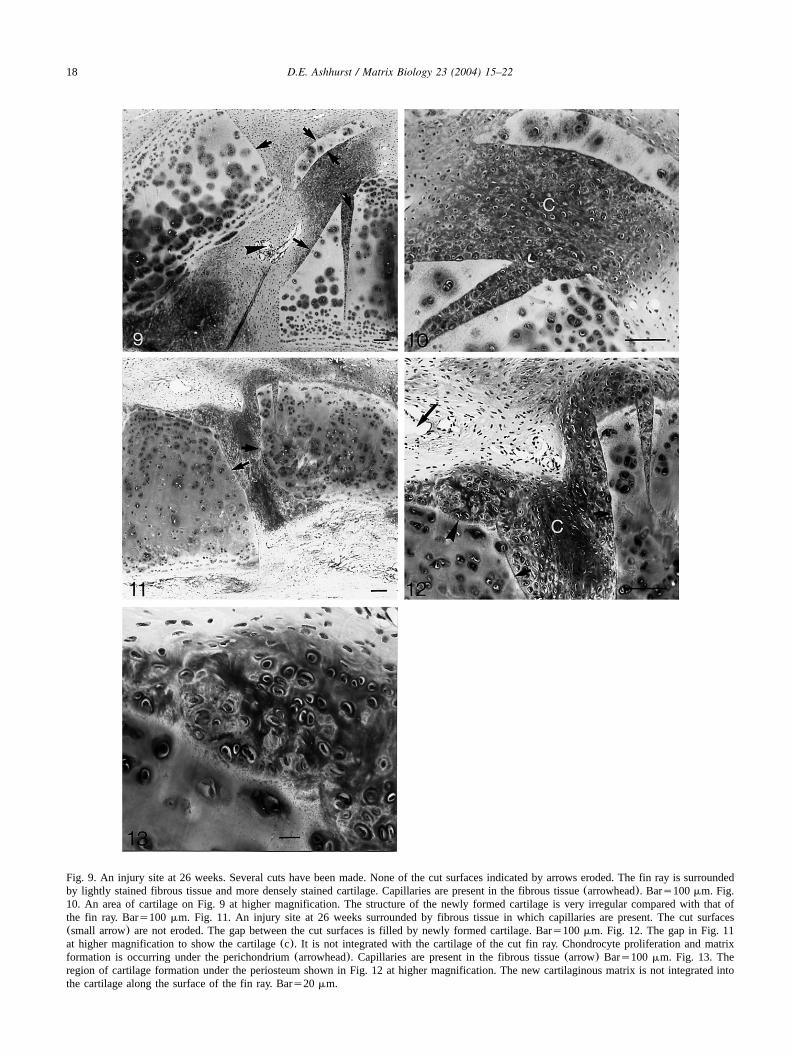
and any small pieces of cartilage are held within a densefibrous matrix(Figs. 9–13). Blood vessels are confinedto the fibrous tissue, which is not found in the gapsbetween the cut ends of the fin rays(Figs. 9 and 11).In the gaps chondrocyte-like cells are surrounded by a
matrix that binds Alcian Blue(Figs. 10, 12 and 13).The cells appear very similar to those within the fin ray(Fig. 13). Examination of these regions with polarizedlight does not reveal collagen fibres crossing the junctionbetween the old and new matrices. Thus, the newmatrices are not integrated with those beyond the cutsurface of the fin ray. Near the cuts there may beevidence of cell proliferation and matrix formation underthe perichondrium, but again there is no integration ofthe original and new matrices(Fig. 13).
3. Discussion
The aim of these experiments was to determinewhether the cartilaginous skeleton of an elasmobranchfish will heal. Repair processes take longer in cold-
18 D.E. Ashhurst / Matrix Biology 23 (2004) 15–22
Fig. 9. An injury site at 26 weeks. Several cuts have been made. None of the cut surfaces indicated by arrows eroded. The fin ray is surroundedby lightly stained fibrous tissue and more densely stained cartilage. Capillaries are present in the fibrous tissue(arrowhead). Bars100mm. Fig.10. An area of cartilage on Fig. 9 at higher magnification. The structure of the newly formed cartilage is very irregular compared with that ofthe fin ray. Bars100 mm. Fig. 11. An injury site at 26 weeks surrounded by fibrous tissue in which capillaries are present. The cut surfaces(small arrow) are not eroded. The gap between the cut surfaces is filled by newly formed cartilage. Bars100mm. Fig. 12. The gap in Fig. 11at higher magnification to show the cartilage(c). It is not integrated with the cartilage of the cut fin ray. Chondrocyte proliferation and matrixformation is occurring under the perichondrium(arrowhead). Capillaries are present in the fibrous tissue(arrow) Bars100 mm. Fig. 13. Theregion of cartilage formation under the periosteum shown in Fig. 12 at higher magnification. The new cartilaginous matrix is not integrated intothe cartilage along the surface of the fin ray. Bars20 mm.

19D.E. Ashhurst / Matrix Biology 23 (2004) 15–22
blooded animals. For this reason the experiments wereextended to 6 months. The initial response to thefractured fin ray is an inflammatory reaction. Manylymphocytes and extravasated red cells mingle withfibroblasts. This takes 3 to 4 weeks to resolve at whichtime the area around the cut ends is filled by fibroustissue. It is a further 8 weeks(i.e. at 12 weeks post-operation) before cells resembling chondrocytes arepresent in regions of poor blood supply, but there islittle cartilaginous matrix. Later, at 26 weeks cartilage-like tissue is present, but it bears little similarity to thehyaline cartilage of the rays. An important point is thatthere has been no change in the cut surfaces of the finrays. These are covered by cells from an early stage,but there is no evidence of resorption or other modifi-cation. New tissue produced adjacent to the cut surfaceis not integrated with it. In contrast, new bone laid downon a fractured surface is subsequently integrated withthe old bone as a result of remodelling. In all tissuerepair processes, the anchorage of the repair tissue tothe existing healthy tissue is essential.The absence of any modification of the cut surfaces
of the fin rays may be caused by the lack of cellscompetent to perform this function. In mammals onlycalcified cartilage is resorbed and resorption is per-formed by osteoclasts, which are derived from mono-cytes. No fishes possess bone marrow. Haematopoiesistakes place at other sites including the liver and spleen(Hyder et al., 1983; Tavassoli, 1986; Torroba et al.,1995). Both elasmobranch and teleost fishes possesserythrocytes, several types of lymphocyte and monocyte-macrophages(Hyder et al., 1983; Ellis, 1986). The roleof lymphocytes in inflammation is similar to that inmammals and the macrophages are phagocytic. A sig-nificant difference between the cartilaginous elasmo-branchs and the bony teleosts is that osteoclasts arefound only in the bony fish(Lopez et al., 1980;Huysseune and Sire 1992; Hughes et al., 1994). Typicalmultinucleated osteoclasts are described and they wereshown to resorb both bone and Meckel’s cartilage. Theabsence of osteoclasts, or similar cells, in elasmobranchscould account for the absence of resorption of thedamaged surfaces of the fin rays.Elasmobranchs cannot, therefore, repair their skeleton,
that is, no repair process for hyaline cartilage hasevolved in these fish. It may be argued that the likeli-hood of a fractured bone in a fish is small and, becausethey are supported by water, it is not imperative fortheir survival that all damaged skeletal elements arerepaired. There is evidence in teleosts that when skeletalstructures, which are essential for survival are damaged,for example, jaw bones, barbels, or large area of fin,healing or regeneration occurs. Thus, callus containingboth bone and cartilage is formed after fracture ofteleost jaw bones(Moss, 1962). The long projectionsfrom the head, or barbels, of catfish are covered by taste
buds and are essential for food selection. After ampu-tation, the cut surface is covered by a blastema and thenskin. Cells from the perichondrium migrate over the cutend of the cartilaginous supporting rod, differentiate intochondrocytes and produce new matrix until the barbelachieves its original length(Goss, 1954; Saxena andAggarwal, 1971). The knifefish (Eigenmannia vires-cens) is able to regenerate its tail(Kirschbaum andMeunier, 1981). If up to 20 vertebrae are removed, ablastema develops and cartilage grows into it from thenotochord and later undergoes endochondral ossification.Teleost fins also regenerate when a piece is removed,but bone and cartilage are not involved and the resultsare frequently imperfect(Wagner and Misof, 1992;Becerra et al., 1996). Regeneration, as in the barbelsand tail, differs from healing, or repair, in that theinteraction of mesenchymal and epithelial tissues and anerve supply are required(Wagner and Misof, 1992).Skeletal healing is independent of epithelial influence.Terrestrial vertebrates depend upon their skeleton for
support and locomotion. They have evolved a mecha-nism for the repair of bones that is remarkably similarin amphibians, reptiles and mammals(Pritchard andRuzicka, 1950; Robertson, 1969; Hall and Hanken,1985; Ashhurst, 1986). After the initial blastema isremoved, newly formed bone and cartilage create acallus around the fractured ends. This cartilage is laterreplaced by bone endochondrally. In contrast, whencartilaginous structures are injured, or fractured, they donot heal. From an evolutionary viewpoint this is notsurprising because by birth or hatching most of theskeleton is ossified. Damage to the remaining cartilagi-nous regions, such as, articular cartilage and sternum, isnot life threatening so repair is of little, or no, selectiveadvantage.A number of experiments have, however, been per-
formed over the years to investigate the healing potentialof cartilage. In early experiments Niven(1931) isolatedchicken embryo tibiae of different developmental ages,cut the diaphysis and maintained the tibiae in culturefor various periods. In those tibiae isolated and fracturedon day 5, the chondrocytes around the cut proliferatedand a perfect repair was seen within 24 h in vitro. Thesame experiment was repeated on 5.5- and 7-day-oldchicken embryo tibiae and they were cultured for 6days. There was no union of cartilage or proliferationof chondrocytes. Instead the chondrocytes adjacent tothe cut became necrotic. In the 7-day embryo, bone isdeveloping around the diaphysis and this achieves amechanical repair by holding the two parts of the tibiatogether. In older embryos, bone formation is responsiblefor the repair. There is, therefore, a critical time atwhich diaphyseal chondrocytes lose their ability todivide and repair an injury. This appears to coincidewith chondrocyte maturity and matrix formation, priorto hypertrophy and bone formation.

20 D.E. Ashhurst / Matrix Biology 23 (2004) 15–22
Niven (1931) extended her studies to the epiphysesof tibiae from 6-day embryos. Vertical cuts into theputative articular regions where the chondrocytes aresmall and round heal perfectly, but those in regions offlattened cells at the disphyseal–epiphyseal junction donot. More recently, similar experiments were performedon femoral condyles of sheep fetuses of 75 days gesta-tion (terms145 days) at which time the putative artic-ular cartilage contains randomly arranged chondrocytesseparated by matrix(Namba et al., 1998). Cuts 100mmdeep were made in the femoral condyles. Dead chondro-cytes are seen in the incision at 3 days, but at 7 daysthe incision has healed although the region is hypocel-lular. At 28 days, the area is said to be normal.The experiments above help to explain the results of
later experiments with developing avian bones. Chickenembryo radii and quail embryo femora and tibiaebetween 6.5 and 8 days old were cut and either main-tained in ovo, or isolated and maintained on allantoicmembranes(Simmons and Kahn, 1979; McCullagh etal., 1990). In both experiments, the cartilage does notheal, nor is new cartilage formed; union is achieved bybone formation as ossification proceeds. In anotherexperiment on embryonic chicken cartilage a trephinewound was made in sterna isolated at HH stage 42(sapprox. 15 days) and subsequently maintained in vitrofor up to 10 days, but although both cellular and matrixchanges were seen, there was no evidence of repair(Walker et al., 2000). These results can be predictedfrom Niven’s (1931) work because the chondrocytesare too mature at the beginning of the experiments.Comparable experiments on mammalian fetuses(Risand Wray, 1972; Longaker et al., 1992) were performedafter ossification had started and are, therefore, notrelevant here.Other sites used to investigate cartilage healing
include Meckel’s cartilage, nasal septum, pinna of themammalian ear and the xiphisternum. Meckel’s cartilageof adult salamanders, xiphisternum of adult rats andnasal septum of neonatal rats do not heal although thereis some production of fibrous tissue(Subba Rao, 1954;Goss, 1977; Hall and Hanken, 1985). An area ofimmature cartilage develops, however, around the cutsurfaces twenty days after removal of a piece of elasticcartilage from the rabbit ear(Joseph et al., 1961); theappearance in the figures is very similar to that of Fig.12 in this paper. The gap in the cartilage is not, however,bridged by elastic cartilage. More recent experimentsinvolving removal of tissue from newborn and adultmouse pinnae resulted in healing of the neonatal, butnot of the adult, elastic cartilage(Wagner et al., 2001).Cell proliferation is detected in the 4-day neonatal elasticcartilage, which from the figures appears immature andhighly cellular. It bears a close resemblance to thecartilage of the 5-day chicken embryo(see above;
Niven, 1931) and the healing processes appear to becomparable.These experiments taken together indicate that only
developing cartilage can heal and that the injury mustoccur while the chondrocytes are still dividing rapidlyand there is little intervening matrix. The potential ofchondrocytes to divide after injury decreases with agein both articular and other permanent cartilages(Niven,1931; Tew et al., 2001). As pointed out by Nodder andMartin (1997) embryonic tissue repair re-uses the devel-opmental mechanisms of morphogenesis and tissue dif-ferentiation. Thus, it may be argued that this is not truehealing, but simply the response to a disturbance of arapid, ongoing, developmental process. Once the tissueis mature any repair tissue originates outside the carti-lage. Thus, in Meckel’s cartilage and embryonic chickenlong bones, not only is no new cartilage formationstimulated, but no cell division in the perichondrium isseen. Similarly, there is no cell proliferation in theperichondrium of the fin rays until 26 weeks and thenit is very localized. Some cartilage is produced in thedamaged dogfish fin and mammalian pinna, but it is nothyaline and is not integrated into the original cartilage.What are the factors that control the development of
cartilage in tissues? One is the blood supply, or ratherits absence in areas of chondrocytic differentiation. Indeveloping embryos, the blood vessels regress from thecentral core of the limb buds before the mesenchymalcells form condensations in which cartilage develops(Caplan and Koutroupas, 1973; Feinberg et al., 1986;Latker et al., 1986; Wilson, 1986; Hallman et al., 1987).Recently, Yin and Pacifici(2001) confirmed that theabsence of blood vessels is necessary for cartilagedevelopment when they showed directly that enhancedvascularity induced by vascular endothelial growth fac-tor inhibits digit development and cartilage formation indeveloping chicken limb buds. A comparable conclusionwas drawn from the callus of rabbit tibial fractureshealing under unstable mechanical conditions(Ashhurst,1986, 2000). A large callus develops and cartilageformation occurs only in the region over the fracturesite, which is avascular. Similarly, it is noted in thispaper that the regions in which cartilage develops nearthe fin rays are avascular. These observations indicatethat skeletal progenitor cells differentiate into chondro-cytes only in regions devoid of blood vessels.It is not pertinent to review here the many experi-
mental studies of articular cartilage repair, but theexperiments discussed above do serve, however, toemphasize the problems. These include the low prolif-erative rate of mature chondrocytes and the inability toreproduce the original structure of the matrix and tointegrate it. Thus, the conclusion is that a mechanismfor the repair of damaged cartilage has not evolved,possibly because from an evolutionary viewpoint itsrepair would be of little selective advantage. Present

21D.E. Ashhurst / Matrix Biology 23 (2004) 15–22
interest in the repair of cartilage arises, however, fromthe longer human life span and from the excessiveloading and strains put on articular cartilage by modernsporting activities.
4. Experimental procedures
Dogfish, Scyliorhinus spp., were obtained from theMarine Biological Association, Plymouth and kept intanks of artificial sea water(Tropic Marine, TropicalMarine Centre, Rickmansworth, Herts, UK) circulatingat a constant temperature of 158C. Under anaesthesia,a small incision was made in the skin and underlyingtissues of the upper surface of the pectoral fin to exposethe fin rays. Three rays were cut through with a scalpel.The skin was sutured. The fish were returned to thetank and recovered from the anaesthetic within 5 min.Postoperatively the fish used the fin normally. Theseexperiments were conducted under the Animals(Scien-tific Procedures) Act 1986.Three fish were killed at each of the following times-
1, 2, 3, 4, 8 and 12 and 26 weeks after operation. Thetissue containing the damaged fin rays was removedand fixed in either Bouin’s fluid or 4% paraformalde-hyde in 0.1 M phosphate buffer, pH 7.4, for approxi-mately 18 h. After washing, the tissues were dehydratedin graded ethanols, cleared in methyl salicylate andembedded in paraffin wax. Sections were cut at 7mmand stained with Alcian Blue, pH 2.5, followed byhaematoxylin and eosin. Calcium deposits were identi-fied by the Von Kossa method.
Acknowledgments
I wish to thank Yvette Bland, Joanna Bryson andKrystala Michael for their expert assistance with thehistology.
References
Ashhurst, D.E., 1986. The influence of mechanical conditions on thehealing of experimental fractures in the rabbit: a microscopicalstudy. Philos. Trans. R. Soc. Lond. B 313, 271–302.
Ashhurst, D.E., 2000. Bone healing and the differentiation of osteo-progenitor cells in mammals. In: Malhotra, S.K.(Ed.), Advancesin Structural Biology, Vol. 6. JAI Press, Stamford, Connecticut,pp. 83–116.
Becerra, J., Junqueira, L.C.U., Bechara, I.J., Montes, G.S., 1996.Regeneration of fin rays in teleosts: a histochemical, radioauto-graphic, and ultrastructural study. Arch. Histol. Cytol. 59, 15–35.
Caplan, A.I., Koutroupas, S., 1973. The control of muscle andcartilage development in the chick limb: the role of differentialvascularization. J. Embryol. Exp. Morphol. 29, 571–583.
Ellis, A.E., 1986. The function of teleost fish lymphocytes in relationto inflammation. Int. J. Tissue React. 8, 263–270.
Feinberg, R.N., Latker, C.H., Beebe, D.C., 1986. Localized vascularregression during limb morphogenesis in the chicken embryo. I.Spatial and temporal changes in the vascular pattern. Anat. Rec.214, 405–409.
Frenkel, S.R., Di Cesare, P.E., 1999. Degradation and repair ofarticular cartilage. Front. Biosci. 4, D671–D685.
Goss, A.N., 1977. Intra-uterine healing of fetal rat oral mucosal, skinand cartilage wounds. J. Oral Pathol. 6, 35–43.
Goss, R.J., 1954. The role of the central cartilaginous rod in theregeneration of the catfish taste barbel. J. Exp. Zool. 127, 181–199.
Hall, B.K., Hanken, J., 1985. Repair of fractured lower jaws in thespotted salamander: do amphibians form secondary cartilage? J.Exp. Zool. 233, 359–368.
Hallman, R., Feinberg, R.N., Latker, C.H., Sasse, J., Risau, W., 1987.Regression of blood vessels precedes cartilage differentiation dur-ing chick limb development. Differentiation 34, 98–105.
Hughes, D.R., Bassett, J.R., Moffat, L.A., 1994. Structure and originof the tooth pedicel(the so-called bone of attachment) and dental-ridge bone in the mandibles of the sea breamsAcanthopagrusaustralis, Pagrus auratus and Rhabdosargus sarba (Sparidae,Perciformes, Telesotei). Anat. Embryol. 189, 51–69.
Huysseune, A., Sire, J.-Y., 1992. Bone and cartilage resorption inrelation to tooth development in the anterior part of the mandiblein cichlid fish: a light and TEM study. Anat. Rec. 234, 1–14.
Hyder, S.L., Cayer, M.L., Pettey, C.L., 1983. Cell types in peripheralblood of the nurse shark: an approach to structure and function.Tissue Cell 15, 437–455.
Joseph, J., Thomas, G.A., Tynen, J., 1961. The reaction of the earcartilage of the rabbit and guinea-pig to trauma. J. Anat. 95,564–568.
Kirschbaum, F., Meunier, F.J., 1981. Experimental regeneration of thecaudal skeleton of the glass knifefish,Eigenmannia virescens(Rhamphichthydae, Gymnotoidei). J. Morphol. 168, 121–135.
Latker, C.H., Feinberg, R.N., Beebe, D.C., 1986. Localized vascularregression during limb morphogenesis in the chicken embryo. II.Morphological changes in the vasculature. Anat. Rec. 214,410–417.
Longaker, M.T., Moelleken, B.R., Cheng, J.C., Jennings, R.W.,Adzick, N.S., Mintorovich, J., et al., 1992. Fetal fracture healingin a lamb model. Plastic Reconst. Surg. 90, 161–171.
Lopez, E., Mac Intyre, I., Martelly, E., Lallier, F., Vidal, B., 1980.Paradoxical effect of 1,25 dihydroxy-cholecalciferol on osteoblasticand osteoclastic activity in the skeleton of the eelAnguilla anguillaL. Calcif. Tissue Int. 32, 83–87.
McCullagh, J.J., Gill, P., Wilson, D.J., 1990. Repair of cartilaginousfractures during chick limb development. J. Orthop. Res. 8,127–131.
Moss, M.L., 1962. Studies of the acellular bone of teleost fish. II.Response to fracture under normal and acalcemic conditions. ActaAnat. 48, 46–60.
Namba, R.S., Meuli, M., Sullivan, K.M., Le, A.X., Adzick, N.S.,1998. Spontaneous repair of superficial defects in articular cartilagein a fetal lamb model. J. Bone Joint Surg. 80A, 4–10.
Niven, J.S.F., 1931. The repair in vitro of embryonic skeletal rudi-ments after experimental injury. J. Pathol. Bacteriol. 34, 307–324.
Nodder, S., Martin, P., 1997. Wound healing in embryos: a review.Anat. Embryol. 195, 215–228.
Pritchard, J.J., Ruzicka, A.J., 1950. Comparison of fracture repair inthe frog, lizard and rat. J. Anat. 84, 236–261.
Ris, P.M., Wray, J.B., 1972. A histological study of fracture healingwithin the uterus of the rabbit. Clin. Orthop. Relat. Res. 87,318–321.
Robertson, D.R., 1969. The ultimobranchial body ofRana pipiens.X. Effect of glandular extirpation on fracture healing. J. Exp. Zool.172, 425–442.
Saxena, P.K., Aggarwal, S., 1971. Structure and regeneration ofbarbels inHeteropneustis fossilis (Bloch.). Anat. Anzeiger 128,354–364.
Simmons, D.J., Kahn, A.J., 1979. Cell lineage in fracture healing inchimeric bone grafts. Calcif. Tissue Int. 27, 247–253.

22 D.E. Ashhurst / Matrix Biology 23 (2004) 15–22
Subba Rao, K.V., 1954. An experimental study of regeneration incartilage. J. Pathol. Bacteriol. 67, 455–459.
Tavassoli, M., 1986. Bone marrow in boneless fish: lessons ofevolution. Med. Hypotheses 20, 9–15.
Temenoff, J.S., Mikos, A.G., 2000. Tissue engineering for regenera-tion of articular cartilage. Biomaterials 21, 431–440.
Tew, S., Redman, S., Kwan, A., Walker, E., Khan, I., Dowthwaite,G., et al., 2001. Differences in repair responses between immatureand mature cartilage. Clin. Orthop. Relat. Res. 391S, S142–S152.
Torroba, M., Chiba, A., Vicente, A., Varas, A., Sacedon, R., Jimenez,E., et al., 1995. Macrophage-lymphocyte cell clusters in thehypothalamic ventricle of some elasmobranch fish: ultrastructuralanalysis and possible functional significance. Anat. Rec. 242,400–410.
Wagner, G.P., Misof, B.Y., 1992. Evolutionary modification of regen-erative capability in vertebrates: a comparative study on teleostpectoral fin regeneration. J. Exp. Zool. 261, 62–78.
Wagner, W., Reichl, J., Wehrmann, M., Zenner, H.-P., 2001. Neonatalrat cartilage has the capacity for tissue regeneration. Wound RepairRegen. 9, 531–536.
Walker, E.A., Verner, A., Flannery, C.R., Archer, C.W., 2000. Cellularresponses of embryonic hyaline cartilage to experimental woundingin vitro. J. Orthop. Res. 18, 25–34.
Wilson, D.J., 1986. Development of avascularity during cartilagedifferentiation in the embryonic limb. Differentiation 30, 183–187.
Yin, M., Pacifici, M., 2001. Vascular regression is required formesenchymal condensation and chondrogenesis in the developinglimb. Dev. Dyn. 222, 522–533.


















