Temporalis Function in Anthropoids and Strepsirrhines…rosslab.uchicago.edu/publications/Hylander...
22
Temporalis Function in Anthropoids and Strepsirrhines: An EMG Study William L. Hylander, 1,2 * Christine E. Wall, 1 Christopher J. Vinyard, 3 Callum Ross, 4 Mathew R. Ravosa, 5 Susan H. Williams, 6 and Kirk R. Johnson 1 1 Department of Biological Anthropology and Anatomy, Duke University Medical Center, Durham, North Carolina 27710 2 Duke University Primate Center, Durham, North Carolina 27705 3 Department of Anatomy, NEOUCOM, Rootstown, Ohio 44272 4 Department of Anatomical Sciences, Stony Brook University, Health Sciences Center, Stony Brook, New York 11794 5 Department of Cell and Molecular Biology, Northwestern University Feinberg School of Medicine, Chicago, Illinois 60611 6 Department of Biomedical Sciences, Ohio University College of Osteopathic Medicine, Athens, Ohio 45701 KEY WORDS temporalis; electromyography; biomechanics; jaw-adductor muscle force; mastication; primates ABSTRACT The major purpose of this study is to ana- lyze anterior and posterior temporalis muscle force recruit- ment and firing patterns in various anthropoid and strepsir- rhine primates. There are two specific goals for this project. First, we test the hypothesis that in addition to transversely directed muscle force, the evolution of symphyseal fusion in primates may also be linked to vertically directed balancing- side muscle force during chewing (Hylander et al. [2000] Am. J. Phys. Anthropol. 112:469 – 492). Second, we test the hypothesis of whether strepsirrhines retain the hypothe- sized primitive mammalian condition for the firing of the anterior temporalis, whereas anthropoids have the derived condition (Weijs [1994] Biomechanics of Feeding in Verte- brates; Berlin: Springer-Verlag, p. 282–320). Electromyo- graphic (EMG) activities of the left and right anterior and posterior temporalis muscles were recorded and analyzed in baboons, macaques, owl monkeys, thick-tailed galagos, and ring-tailed lemurs. In addition, as we used the working-side superficial masseter as a reference muscle, we also recorded and analyzed EMG activity of the left and right superficial masseter in these primates. The data for the anterior tem- poralis provided no support for the hypothesis that symph- yseal fusion in primates is linked to vertically directed jaw muscle forces during mastication. Thus, symphyseal fusion in primates is most likely mainly linked to the timing and recruitment of transversely directed forces from the balanc- ing-side deep masseter (Hylander et al. [2000] Am. J. Phys. Anthropol. 112:469 – 492). In addition, our data demonstrate that the firing patterns for the working- and balancing-side anterior temporalis muscles are near identical in both strep- sirrhines and anthropoids. Their working- and balancing- side anterior temporalis muscles fire asynchronously and reach peak activity during the power stroke. Similarly, their working- and balancing-side posterior temporalis muscles also fire asynchronously and reach peak activity during the power stroke. Compared to these strepsirrhines, however, the balancing-side posterior temporalis of anthropoids ap- pears to have a relatively delayed firing pattern. Moreover, based on their smaller W/B ratios, anthropoids demonstrate a relative increase in muscle-force recruitment of the balanc- ing-side posterior temporalis. This in turn suggests that anthropoids may emphasize the duration and magnitude of the power stroke during mastication. This hypothesis, how- ever, requires additional testing. Furthermore, during the latter portion of the power stroke, the late activity of the balancing-side posterior temporalis of anthropoids appar- ently assists the balancing-side deep masseter in driving the working-side molars through the terminal portion of occlu- sion. Am J Phys Anthropol 128:35–56, 2005. © 2005 Wiley-Liss, Inc. Craniofacial biomechanics continue to attract a sig- nificant amount of research interest. Furthermore, as the masticatory apparatus has a major influence on craniofacial form, considerable efforts have been di- rected toward studying the biomechanics of chewing and biting. These studies often require an estimate of the direction, magnitude, and/or timing of various jaw- closing muscle forces. For this reason, we have been actively involved in analyzing jaw-muscle electromyo- graphic (EMG) activity patterns among various spe- cies of mammals (e.g., Hylander et al., 2000, 2004; Vinyard et al., 2004a, b). This study is a continuation of these analyses, and here we focus on the recruit- ment and firing patterns of the anterior and posterior temporalis muscles in primates. Grant sponsor: NIH; Grant numbers: DE04531, DE05595, DE05663; Grant sponsor: NSF; Grant numbers: SBR-9420764, BCS- 0138565, BNS-91-00523. *Correspondence to: Professor William L. Hylander, Department of Biological Anthropology and Anatomy, Duke University Medical School, Box 3170, Durham, NC 27710. E-mail: [email protected] Received 14 October 2003; accepted 29 January 2004. DOI 10.1002/ajpa.20058 Published online 15 February 2005 in Wiley InterScience (www.interscience.wiley.com). AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 128:35–56 (2005) © 2005 WILEY-LISS, INC.
Transcript of Temporalis Function in Anthropoids and Strepsirrhines…rosslab.uchicago.edu/publications/Hylander...

Temporalis Function in Anthropoids and Strepsirrhines:An EMG StudyWilliam L. Hylander,1,2* Christine E. Wall,1 Christopher J. Vinyard,3 Callum Ross,4 Mathew R. Ravosa,5
Susan H. Williams,6 and Kirk R. Johnson1
1Department of Biological Anthropology and Anatomy, Duke University Medical Center, Durham, North Carolina 277102Duke University Primate Center, Durham, North Carolina 277053Department of Anatomy, NEOUCOM, Rootstown, Ohio 442724Department of Anatomical Sciences, Stony Brook University, Health Sciences Center, Stony Brook, New York 117945Department of Cell and Molecular Biology, Northwestern University Feinberg School of Medicine, Chicago, Illinois 606116Department of Biomedical Sciences, Ohio University College of Osteopathic Medicine, Athens, Ohio 45701
KEY WORDS temporalis; electromyography; biomechanics; jaw-adductor muscle force;mastication; primates
ABSTRACT The major purpose of this study is to ana-lyze anterior and posterior temporalis muscle force recruit-ment and firing patterns in various anthropoid and strepsir-rhine primates. There are two specific goals for this project.First, we test the hypothesis that in addition to transverselydirected muscle force, the evolution of symphyseal fusion inprimates may also be linked to vertically directed balancing-side muscle force during chewing (Hylander et al. [2000]Am. J. Phys. Anthropol. 112:469–492). Second, we test thehypothesis of whether strepsirrhines retain the hypothe-sized primitive mammalian condition for the firing of theanterior temporalis, whereas anthropoids have the derivedcondition (Weijs [1994] Biomechanics of Feeding in Verte-brates; Berlin: Springer-Verlag, p. 282–320). Electromyo-graphic (EMG) activities of the left and right anterior andposterior temporalis muscles were recorded and analyzed inbaboons, macaques, owl monkeys, thick-tailed galagos, andring-tailed lemurs. In addition, as we used the working-sidesuperficial masseter as a reference muscle, we also recordedand analyzed EMG activity of the left and right superficialmasseter in these primates. The data for the anterior tem-poralis provided no support for the hypothesis that symph-yseal fusion in primates is linked to vertically directed jawmuscle forces during mastication. Thus, symphyseal fusionin primates is most likely mainly linked to the timing and
recruitment of transversely directed forces from the balanc-ing-side deep masseter (Hylander et al. [2000] Am. J. Phys.Anthropol. 112:469–492). In addition, our data demonstratethat the firing patterns for the working- and balancing-sideanterior temporalis muscles are near identical in both strep-sirrhines and anthropoids. Their working- and balancing-side anterior temporalis muscles fire asynchronously andreach peak activity during the power stroke. Similarly, theirworking- and balancing-side posterior temporalis musclesalso fire asynchronously and reach peak activity during thepower stroke. Compared to these strepsirrhines, however,the balancing-side posterior temporalis of anthropoids ap-pears to have a relatively delayed firing pattern. Moreover,based on their smaller W/B ratios, anthropoids demonstratea relative increase in muscle-force recruitment of the balanc-ing-side posterior temporalis. This in turn suggests thatanthropoids may emphasize the duration and magnitude ofthe power stroke during mastication. This hypothesis, how-ever, requires additional testing. Furthermore, during thelatter portion of the power stroke, the late activity of thebalancing-side posterior temporalis of anthropoids appar-ently assists the balancing-side deep masseter in driving theworking-side molars through the terminal portion of occlu-sion. Am J Phys Anthropol 128:35–56, 2005.© 2005 Wiley-Liss, Inc.
Craniofacial biomechanics continue to attract a sig-nificant amount of research interest. Furthermore, asthe masticatory apparatus has a major influence oncraniofacial form, considerable efforts have been di-rected toward studying the biomechanics of chewingand biting. These studies often require an estimate ofthe direction, magnitude, and/or timing of various jaw-closing muscle forces. For this reason, we have beenactively involved in analyzing jaw-muscle electromyo-graphic (EMG) activity patterns among various spe-cies of mammals (e.g., Hylander et al., 2000, 2004;Vinyard et al., 2004a, b). This study is a continuationof these analyses, and here we focus on the recruit-ment and firing patterns of the anterior and posteriortemporalis muscles in primates.
Grant sponsor: NIH; Grant numbers: DE04531, DE05595,DE05663; Grant sponsor: NSF; Grant numbers: SBR-9420764, BCS-0138565, BNS-91-00523.
*Correspondence to: Professor William L. Hylander, Department ofBiological Anthropology and Anatomy, Duke University MedicalSchool, Box 3170, Durham, NC 27710.E-mail: [email protected]
Received 14 October 2003; accepted 29 January 2004.
DOI 10.1002/ajpa.20058Published online 15 February 2005 in Wiley InterScience
(www.interscience.wiley.com).
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 128:35–56 (2005)
© 2005 WILEY-LISS, INC.

HOW JAW-CLOSING MUSCLES WORKDURING CHEWING
Prior to outlining the details about what we pro-pose to do, it is important to provide some back-ground information about how mammalian jawmuscles function during mastication. For manymammals, the jaw-closing muscles are thought toact as two force couples during jaw closing (e.g.,Moller, 1966; Herring and Scapino, 1973; Luscheiand Goodwin, 1974; de Vree and Gans, 1976; Gor-niak, 1977; Weijs and Dantuma, 1981).1 These two“couples,” called “diagonals” or “diagonal pairs” byHerring (1976, 1985a, b, 1992, 1993) and “triplets”by Weijs and Dantuma (1981) and Weijs (1994), aremade up of a combination of various muscles. Eachdiagonal or triplet has one muscle that pulls themandible upward and backward, the temporalis,and two muscles that pull the mandible upward andforward, the opposite-side superficial masseter andmedial pterygoid muscles (Herring, 1992; Weijs,1994) (Fig. 1). Furthermore, these two musclegroups fire asymmetrically. The working-side (w-s)temporalis and the balancing-side (b-s) superficialmasseter and medial pterygoids fire first (diagonalor triplet I). This causes the lower jaw to be elevatedand its working side shifted laterally. The firing oftriplet or diagonal I is followed by the firing of tripletor diagonal II, which consists of the b-s temporalisand the w-s superficial masseter and medial ptery-goids. This causes the jaw to continue elevating, andshortly thereafter its working side is shifted medi-ally, back towards the midline.
As described above, for many mammals as well asall primates, the w-s and b-s posterior temporalis arethought to fire with triplets and diagonals I and II,respectively. On the other hand, for “prosimians”and many other “primitive” mammals, the left andright anterior temporalis muscles are said to fire asa third muscle group, the symmetric vertical closers(SVC) (Weijs, 1994; Langenbach and van Eijden,2001). Following the suggestion of Weijs (1994), theSVC of “primitive” mammals (including “prosim-ians”) are said to fire synchronously, prior to tripletI and only during fast closing (Fig. 2A). In contrast,the so-called “transverse” chewers (including an-thropoids) are said to fire their w-s and b-s anteriortemporalis with triplets (or diagonals) I and II, re-spectively (Fig. 2B) (Weijs, 1994).
PURPOSE OF STUDY
The purpose of this study is to analyze the EMGactivity of the anterior and posterior temporalis invarious anthropoid and strepsirrhine primates dur-ing mastication. There are two specific goals for thisproject. First, and most importantly, as part of ourcontinuing efforts to identify possible links betweenbiomechanical stress and symphyseal morphology,we propose to characterize the recruitment of rela-tive force of the anterior and posterior temporalismuscles in baboons, macaques, owl monkeys, thick-tailed galagos, and ring-tailed lemurs. We are par-ticularly interested in determining whether anthro-poids recruit relatively more force from their b-santerior temporalis than do strepsirrhines with mo-bile unfused symphyses. If so, this would provideevidence to support the hypothesis that the evolu-tion of symphyseal fusion in primates is also linkedto vertically directed force from their b-s jaw mus-cles during chewing (Beecher, 1977, 1979; Hylander,1975, 1977, 1979a, 1979b; Ravosa and Hylander,1994; Hylander et al., 2000).
Second, we want to determine firing patterns ofthe anterior and posterior temporalis. We are par-ticularly interested in whether strepsirrhines retainthe hypothesized primitive mammalian conditionfor the firing of the anterior temporalis, whereasanthropoids have a derived condition (Weijs, 1994).If so, among strepsirrhines, the w-s and b-s anteriortemporalis should fire only during fast closing asmembers of the SVC muscles, whereas for anthro-poids, the w-s and b-s anterior temporalis should firealong with members of triplets (or diagonals) I andII, respectively. Furthermore, we are interested inwhether there are important differences betweenstrepsirrhines and anthropoids in the firing patternsof the posterior temporalis.
MATERIALS AND METHODS
Subjects
The following primates served as subjects: 3 adultmale Papio anubis (baboons 2–4), 3 adult male Ma-caca fuscata (macaques 11, 13, and 14), 2 adultAotus trivirgatus, one male (owl monkey 1) and onefemale (owl monkey 2), 2 adult male Otolemur cras-sicaudatus and one adult male O. garnetti (galagos6–8), and 4 adult Lemur catta, 2 males and 2 fe-males (lemurs 1–4). With the exception of the gala-gos, all subjects were used for two or more recordingsessions. These different sessions are indicated inour tables by letters A–D.
EMG electrodes and electrodeplacement procedure
As many as eight fine-wire indwelling bipolar elec-trodes were placed within the left and right middlesuperficial masseter and the anterior and posteriortemporalis muscles (Fig. 3). For all subjects, the tipsof the electrodes were first placed into a fine-gauge
1The concept of the jaw-closing muscles acting as force couples is avery useful first approximation for understanding their mechanicaleffects during unilateral mastication. In a strict sense, however, thejaw-closing muscles do not act solely as force couples, because a coupleis characterized by two forces that are separated, parallel, equal inmagnitude, and opposite in direction (Panjabi and White, 2001). Al-though separated, the muscle forces within each diagonal or tripletare not parallel, equal, or opposite. Nevertheless, if we resolve eachmuscle force into its vertical and horizontal components, the horizon-tal components of these forces closely approximate a force couple (Fig.1B,C).
36 W.L. HYLANDER ET AL.

Fig. 1. Jaw-closing muscle firing patterns according to triplet(primitive mammalian condition) and diagonal hypotheses. Seetext for further discussion as to how triplets (A–C) differ fromdiagonals (B and C). A: Symmetric vertical closers (SVC). Simul-taneous activity of working- (w-s) and balancing-side (b-s) ante-rior temporalis (and deep masseters) initiate vertical jaw closing.Peak activity of SVC occurs well before initiation of power stroke.Fb-s svc is balancing-side force of anterior temporalis and deepmasseter muscles. Fw-s svc is working-side force for anterior tem-poralis and deep masseter muscles. B: Triplet I. Following SVC,triplet I, which is made up of b-s superficial masseter, b-s medialpterygoid, and w-s posterior temporalis muscles, fires more orless simultaneously. Diagonal I consists of above three muscles aswell as w-s anterior temporalis. These groups reach peak activityearly in power stroke. Vertical components of these muscles con-tinue to close jaws as their horizontal components shift w-s cor-pus and teeth laterally by rotating mandible about a vertical axislocated in vicinity of w-s mandibular condyle. Diagonal hypothe-sis does not recognize SVC as a separate muscle group. Instead,jaw closing is initiated by diagonal I. Fb-s sm � mpt is balancing-sideforce of superficial masseter and medial pterygoid. Fw-s pt is w-sforce of posterior temporalis for triplets and w-s force of entiretemporalis for diagonals. C: Triplet II. Following triplet I, tripletII, which is made up of w-s superficial masseter, w-s medialpterygoid, and b-s posterior temporalis, fires more or less simul-taneously.Diagonal II consists of above three muscles as well asb-s anterior temporalis muscles. These groups reach peak activitylate in power stroke. Vertical components of these muscles con-tinue to close jaws as their horizontal components shift w-s cor-pus and teeth back toward midline by rotating mandible about avertical axis located in vicinity of w-s mandibular condyle.Fw-s sm � mpt is the w-s force of superficial masseter and medialpterygoid. Fb-s pt is b-s force of posterior temporalis for tripletsand w-s force of entire temporalis for diagonals.
Fig. 2. Jaw muscle firing patterns. A: Predictions of firingpatterns of jaw closing muscles for “primitive” mammals accord-ing to triplet hypothesis (Weijs, 1994). Note that symmetric ver-tical closers fire first, followed by triplet I, and then triplet II. B:Predictions of firing patterns of jaw-closing muscle for anthro-poids according to triplet and diagonal hypotheses (Herring,1992; Weijs, 1994). C: Predictions of masseter firing pattern as-sociated with wishboning of symphysis. This hypothesis predictsoccurrence of this pattern in only those primates with a partiallyor completely fused mandibular symphysis, but not in thosestrepsirrhines with a highly mobile mandibular symphysis, suchas thick-tailed galagos and ring-tailed lemurs (Hylander andJohnson, 1994; Hylander et al., 2000, 2004). Note that working-side deep masseter peaks first and balancing-side deep masseterpeaks last. Furthermore, balancing-side superficial masseterpeaks after working-side deep and before working-side superficialmasseter. Vertical line indicates peak activity in reference mus-cle, i.e., working-side superficial masseter. Solid squares andcircles, working-side superficial and deep masseter, respectively;open squares and circles, balancing-side superficial and deepmasseter, respectively. See text for further description.

needle, and the needle was then inserted into themasseter or temporalis muscles until its point en-countered mandibular or cranial bone, respectively.The needle was then withdrawn, leaving the elec-trode tips embedded in muscle near the surface ofthe bone. The electrodes in the superficial masseterwere positioned midway between its anterior andposterior borders, between 5–10 mm above the lowerborder of the angle of the mandible, depending onthe size of the subject. Inserting these electrodesnear the ventral edge of the mandible avoids inad-vertent recording from the deep masseter. The elec-trode tips in the anterior temporalis were positioned5–10 mm behind the postorbital bar at the level ofthe center of the eyeball. The electrode tips in theposterior temporalis were positioned 5–10 mm abovethe external auditory meatus.
All electrodes were inserted with the subjects un-der light sedation. For baboons, macaques, owl mon-keys, and galagos, we used a combination ofacepromazine and ketamine (Connolly and Quimby,1978). For ring-tailed lemurs, we used a medetomi-dine-based sedation procedure (Williams et al.,2003). As the subjects were not killed, electrode po-sition was not verified by dissection, althoughproper electrode depth was easily confirmed whenremoving the electrodes at the end of the recordingsession.
Recording procedure
Prior to recovery from the sedative, subjects wererestrained as described elsewhere (Hylander et al.,2000). Once the animal fully recovered from seda-tion, it was fed relatively easy-to-chew foods, i.e.,pieces of fruit pulp with attached skin (apple orpear), fresh carrot, and moist raisins. We also fedthe subjects relatively difficult-to-chew brittle ortough (B/T) foods, i.e., unpopped popcorn kernels,cherry pits, dried gelatin candy (dried gummybears), dried apricots, and dried prunes. The EMGpotentials were simultaneously amplified and fil-tered (bandpass, 100–3,000 Hz) and then recorded
with a 14-channel FM tape recorder at 15 inches persecond. At this time, we identified whether the sub-ject chewed on the left or right side by proceduresoutlined elsewhere (Hylander et al., 2000). Details ofthe training and recording procedures were de-scribed previously (Hylander and Johnson, 1989,1994).
The entire data set is based on 42 separate elec-trode placement and recording sessions: 9 for ba-boons and 9 for macaques (3 for each subject), 8 forowl monkeys (4 for each subject), 3 for thick-tailedgalagos (one for each subject), and 9 for ring-tailedlemurs (1–3 for each subject). All EMG data wererecorded as subjects vigorously chewed various fooditems.
EMG quantification
All EMG recordings of each chewing sequencewere initially reproduced for visual examination byplaying the raw EMG data from the tape recorderinto two multiple-channel storage oscilloscopes. Asmacaques and baboons rarely shift chewing sidesduring a chewing sequence, we selected for analysisone long sequence of chewing on the left and one forchewing on the right for each food. In contrast, owlmonkeys, galagos, and lemurs usually shift chewingsides frequently during a sequence. Therefore, whenpossible and for each food, we selected for analysis atleast one sequence that started with chewing on theleft and one that started with chewing on the right.
For each chewing sequence analyzed, the rawEMG data were played from the FM tape recorderinto a 16-channel analog-to-digital converter (12-bitresolution), and the digitized values were written tothe hard disk of a microcomputer. Each channel wassampled and digitized at a rate of 10,000 Hz. Thedigitized values were then read back into the micro-computer for subsequent processing and analysis.
The digitized raw EMG values were quantified bycalculating the root-mean-square (rms) values in2-millisecond (ms) intervals from the raw digitizedvalues, using a 42-ms time constant (Fig. 4) (Hy-lander and Johnson, 1993). We then identified thelargest rms value based on peak amplitude for eachelectrode throughout a given experiment, regardlessas to whether it was a working- (w-s) or balancing-side (b-s) muscle. For each power stroke, the peakvalues for each electrode were then scaled by assign-ing a value of 1.0 to the largest value, and theremaining peak values were scaled in a linear fash-ion. Thus, for each power stroke and muscle, therewas a scaled value for peak EMG activity for valuesbetween 0–1.0.
Parenthetically, for purposes of this analysis, theEMG timing values of one muscle relative to anotherwere not necessarily dependent on employing whathas been called “the optimal time constant” (Hy-lander and Johnson, 1993). This is true so long asthe selected time constant is neither too short (toosmall) nor too long (too large). If too small (e.g.,about 20 ms or less), the rms EMG in our subjects
Fig. 3. Drawing of lateral view of a male macaque skull andsuperficial masseter and temporalis muscles. Solid black dotsindicate location of bipolar fine-wire indwelling electrodes in su-perficial masseter (sm) and anterior (at) and posterior (pt) tem-poralis.
38 W.L. HYLANDER ET AL.
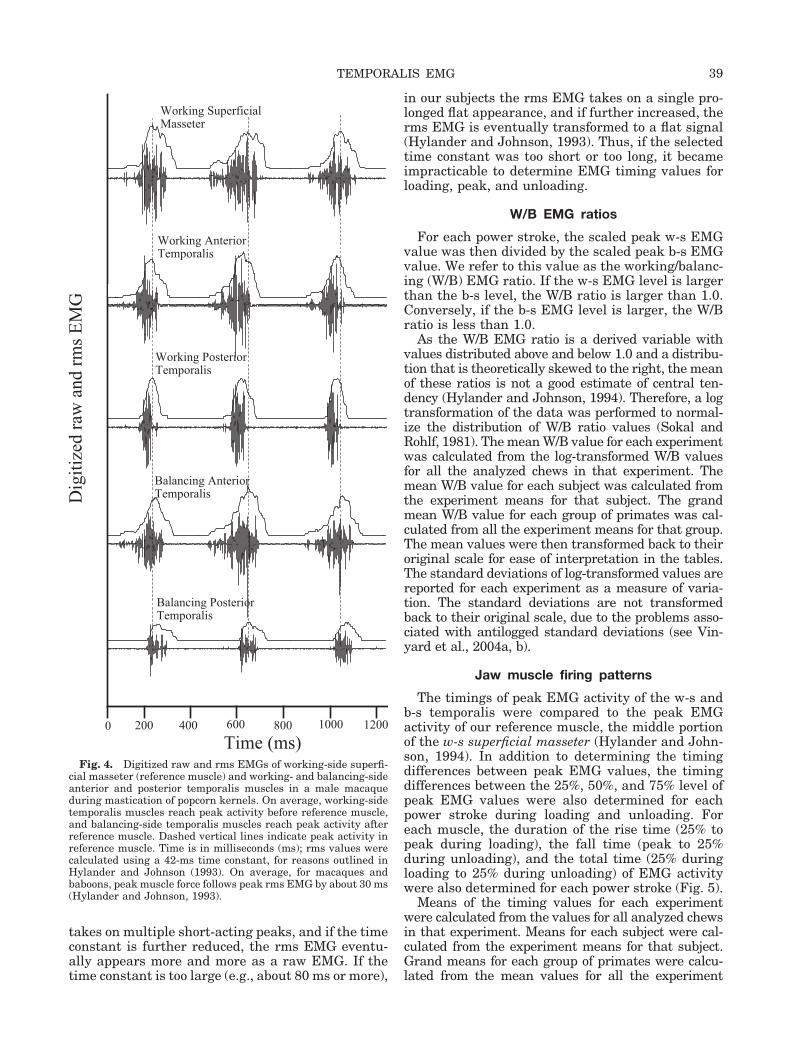
takes on multiple short-acting peaks, and if the timeconstant is further reduced, the rms EMG eventu-ally appears more and more as a raw EMG. If thetime constant is too large (e.g., about 80 ms or more),
in our subjects the rms EMG takes on a single pro-longed flat appearance, and if further increased, therms EMG is eventually transformed to a flat signal(Hylander and Johnson, 1993). Thus, if the selectedtime constant was too short or too long, it becameimpracticable to determine EMG timing values forloading, peak, and unloading.
W/B EMG ratios
For each power stroke, the scaled peak w-s EMGvalue was then divided by the scaled peak b-s EMGvalue. We refer to this value as the working/balanc-ing (W/B) EMG ratio. If the w-s EMG level is largerthan the b-s level, the W/B ratio is larger than 1.0.Conversely, if the b-s EMG level is larger, the W/Bratio is less than 1.0.
As the W/B EMG ratio is a derived variable withvalues distributed above and below 1.0 and a distribu-tion that is theoretically skewed to the right, the meanof these ratios is not a good estimate of central ten-dency (Hylander and Johnson, 1994). Therefore, a logtransformation of the data was performed to normal-ize the distribution of W/B ratio values (Sokal andRohlf, 1981). The mean W/B value for each experimentwas calculated from the log-transformed W/B valuesfor all the analyzed chews in that experiment. Themean W/B value for each subject was calculated fromthe experiment means for that subject. The grandmean W/B value for each group of primates was cal-culated from all the experiment means for that group.The mean values were then transformed back to theiroriginal scale for ease of interpretation in the tables.The standard deviations of log-transformed values arereported for each experiment as a measure of varia-tion. The standard deviations are not transformedback to their original scale, due to the problems asso-ciated with antilogged standard deviations (see Vin-yard et al., 2004a, b).
Jaw muscle firing patterns
The timings of peak EMG activity of the w-s andb-s temporalis were compared to the peak EMGactivity of our reference muscle, the middle portionof the w-s superficial masseter (Hylander and John-son, 1994). In addition to determining the timingdifferences between peak EMG values, the timingdifferences between the 25%, 50%, and 75% level ofpeak EMG values were also determined for eachpower stroke during loading and unloading. Foreach muscle, the duration of the rise time (25% topeak during loading), the fall time (peak to 25%during unloading), and the total time (25% duringloading to 25% during unloading) of EMG activitywere also determined for each power stroke (Fig. 5).
Means of the timing values for each experimentwere calculated from the values for all analyzed chewsin that experiment. Means for each subject were cal-culated from the experiment means for that subject.Grand means for each group of primates were calcu-lated from the mean values for all the experiment
Fig. 4. Digitized raw and rms EMGs of working-side superfi-cial masseter (reference muscle) and working- and balancing-sideanterior and posterior temporalis muscles in a male macaqueduring mastication of popcorn kernels. On average, working-sidetemporalis muscles reach peak activity before reference muscle,and balancing-side temporalis muscles reach peak activity afterreference muscle. Dashed vertical lines indicate peak activity inreference muscle. Time is in milliseconds (ms); rms values werecalculated using a 42-ms time constant, for reasons outlined inHylander and Johnson (1993). On average, for macaques andbaboons, peak muscle force follows peak rms EMG by about 30 ms(Hylander and Johnson, 1993).
TEMPORALIS EMG 39

means for that group. Unless specifically indicated, alltests of significance are on the grand means. As ourEMG timing data may not satisfy all of the require-ments for parametric statistical tests, the mean timingdifferences in peak EMGs between muscles weretested for significance at the 0.05 level using a non-parametric test, i.e., the Wilcoxon signed-ranks test forpaired comparisons. When peak EMG of a muscle waspredicted to either precede or follow peak EMG ofanother muscle, as when testing a diagonal or triplet Imuscle relative to a diagonal or triplet II muscle, aone-tailed test of significance was utilized. Conversely,when testing peak EMG of those muscles within eachtriplet or diagonal group, all tests of significance weretwo-tailed tests.
Data selection
As biomechanical adaptations in the craniofacialregion are likely to be in response to the more force-ful loading regimes, we focus on data recorded dur-ing the chewing of hard, brittle, or tough (B/T) foods.Earlier work established that among various pri-mates, W/B ratios are generally influenced by over-all masticatory force levels, and that the lowest W/Bratios are usually associated with the more forcefulpower strokes (Hylander, 1985; Hylander et al.,1992, 2000). This is also true for certain nonprimatemammals (Didelphis virginiana) (Crompton andHylander, 1986; Crompton, 1995). Therefore, for themore difficult-to-chew foods, we also analyze W/Bratios for 10 of the largest balancing-side peak EMGvalues so as to focus the analysis on the more force-ful power strokes. The identification of these powerstrokes is operationalized by selecting those powerstrokes with the five largest b-s EMG values forchewing on the left side and the five largest b-svalues for chewing on the right side for the anteriortemporalis muscle. This procedure arguably doesnot identify the 10 most forceful power strokes, but
it does allow us to focus our analysis on some of themore forceful power strokes.
At the very beginning of a chewing sequence, oursubjects occasionally engaged in two or three pro-longed and near-isometric bites interrupted by ex-tensive intraoral manipulation of the food item. Im-mediately following these behaviors, they engagedin rhythmic chewing behaviors. Our W/B ratio andtiming data are based entirely on these rhythmicchewing behaviors.
RESULTS
For baboons, macaques, owl monkeys, and ring-tailed lemurs, we recorded a substantial amount ofEMG data for many recording sessions. For galagos,however, we were only able to record data from theanterior temporalis during one recording session(galago 6), and from the posterior temporalis duringthree recording sessions (one session for each galagosubject). As we are unable to increase these samplesdue to the current unavailability of adult galagosubjects, we present the galago data with the under-standing that these data are preliminary.
Some subjects refused to eat certain foods. All,however, chewed at least one difficult-to-chew brit-tle or tough (B/T) food on both the left and rightsides. Therefore, EMG data were recorded for eachsubject during episodes of relatively forceful masti-cation. Unlike the anthropoids, the galagos alwaysand the lemurs usually refused to eat apple skin.
For baboons and macaques, the b-s posterior tem-poralis EMGs were often barely detectable duringthe chewing of less resistant foods. When this oc-curred, this caused the W/B ratio for this muscle tobecome very large. In order to prevent unusuallylarge ratio values from grossly distorting the overallmean W/B ratios, any ratios larger than 10.0 weresimply assigned a value of 10.0 (for further discus-sion of this issue, see Hylander et al., 2000).
W/B EMG ratios of anterior and posteriortemporalis muscles
Table 1 presents the descriptive statistics of W/Bratios for all five primate groups during chewing ofB/T foods. Table 1 also presents mean values of W/Bratios associated with the 10 more forceful powerstrokes (in parentheses). Table 2 consists of sum-mary data of the grand means of W/B ratios for allfive groups.
Anterior temporalis. The grand mean W/B ratiosduring the chewing of B/T foods for baboons, ma-caques, owl monkeys, and lemurs are 1.2, 1.2, 1.4,and 1.5, respectively. For the 10 more forceful powerstrokes, these values are reduced to 1.0, 1.0, 1.2, and1.2. For the single galago recording session, themean for all power strokes and the 10 more forcefulpower strokes are much larger (4.4 and 3.1, respec-tively).
During the chewing of less resistant foods, theW/B ratios are much larger for baboons and ma-
Fig. 5. Method for determining rise time and fall time ofjaw-muscle EMG values. Figure indicates rms EMG of one powerstroke for superficial masseter muscle of a female baboon duringmastication. Rise time and fall time are operationally defined astime between 25–100% of peak EMG during loading and unload-ing, respectively. Total time of activity is operationally defined asrise time plus fall time (from Hylander and Johnson, 1994).
40 W.L. HYLANDER ET AL.

caques (2.8 and 4.1), and only slightly larger for owlmonkeys (1.5).
Posterior temporalis. The W/B ratios during thechewing of B/T foods for baboons, macaques, owlmonkeys, galagos, and lemurs are 1.0, 1.2, 1.4, 2.4,and 2.0, respectively. For the 10 more forceful powerstrokes, these values are reduced to 0.9, 1.1, 1.3, 2.1,and 1.8.
During the chewing of less resistant foods, theW/B ratios are much larger for baboons and ma-caques (7.2 and 7.6), and nearly identical for owlmonkeys (1.3).
% EMG activity of the anterior and posteriortemporalis muscles
Figure 6 is a bar graph indicating average % EMGvalues for all five groups during the chewing of B/Tfoods.
Anterior temporalis. On average, all subjectsexhibit about the same percentage of peak EMGactivity of their w-s anterior temporalis (62–70%)during the chewing of B/T foods. With the exceptionof the single galago recording session, the b-s ante-rior temporalis exhibits 41–59% of peak activity.For galagos, the b-s anterior temporalis is less than20%.
During the chewing of less resistant foods (anthro-poids only), the w-s percentages are reduced to be-tween 50–54%. For baboons and macaques thesevalues fall to 17% and 19%, respectively, whereas forowl monkeys they fall to only 39%.
Posterior temporalis. Baboons and macaquesexhibit about the same percentage of peak EMGactivity for their w-s posterior temporalis during thechewing of B/T foods (42% and 39%, respectively),whereas owl monkeys, galagos, and lemurs exhibit a
TABLE 1. W/B EMG ratios for anterior and posterior temporalis muscles during masticationof brittle/tough foods for each of five groups
TABLE 1A. Baboons1
Subject N
Anterior temporalis Posterior temporalis
Mean (Mean) SDlog10 Mean (Mean) SDlog10
Baboon 3Exp. A 64 1.1 (0.9) 0.07 0.8 (0.8) 0.20Exp. B 52 1.1 (0.9) 0.08 1.0 (0.9) 0.10Exp. C 48 1.0 (0.9) 0.09 0.9 (0.8) 0.56
Subject means 1.1 (0.9) 0.08 0.9 (0.8) 0.29Baboon 4
Exp. A 66 1.0 (0.9) 0.07 0.8 (0.8) 0.09Exp. B 60 1.8 (1.4) 0.18 1.3 (1.0) 0.21Exp. C 81 1.1 (1.0) 0.07 0.8 (0.8) 0.17
Subject means 1.3 (1.1) 0.11 0.9 (0.9) 0.16Baboon 5
Exp. A 75 1.3 (1.1) 0.09 1.0 (0.9) 0.12Exp. B 70 1.0 (1.1) 0.07 1.0 (0.7) 0.30Exp. C 58 1.2 (1.0) 0.11 1.1 (1.2) 0.08
Subject means 1.2 (1.1) 0.09 1.0 (0.9) 0.17Grand means 1.2 (1.0) 0.09 1.0 (0.9) 0.20
1 Values in parentheses (Mean) indicate mean W/B ratio for 10 power strokes (5 from each side), with greatest balancing-sidecontribution based on BAT muscle. N, number of power strokes; SDlog10, standard deviation of log-transformed values.
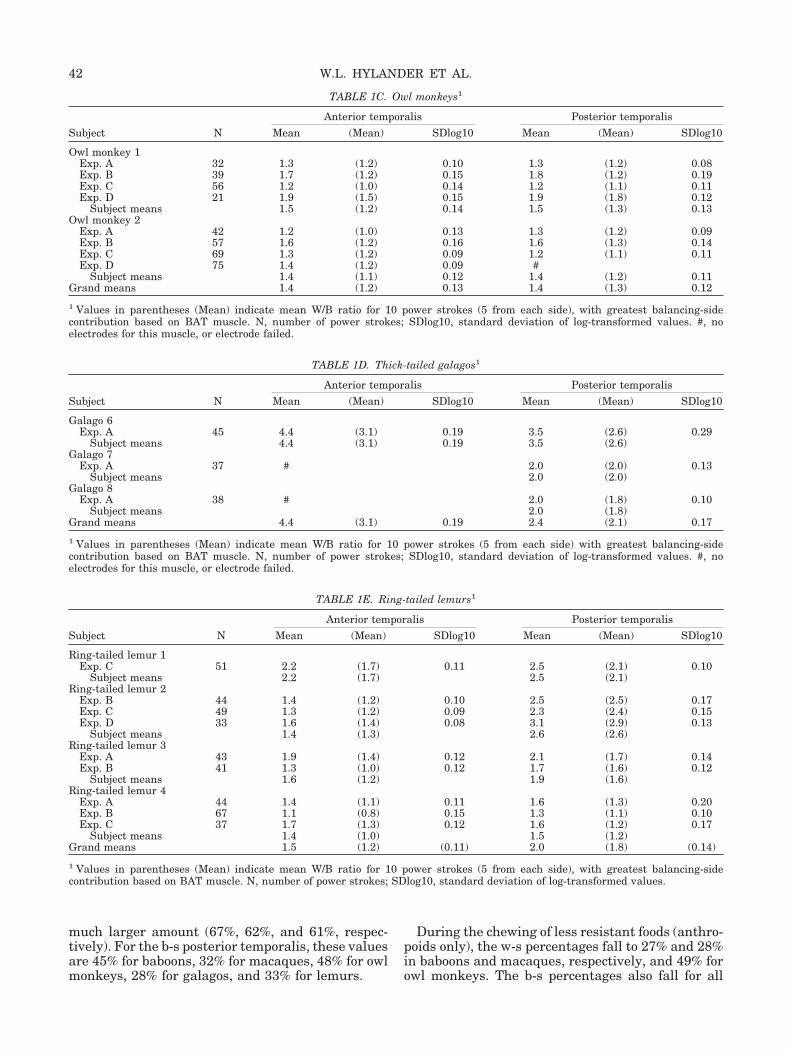
TABLE 1B. Macaques1
Subject N
Anterior temporalis Posterior temporalis
Mean (Mean) SDlog10 Mean (Mean) SDlog10
Macaque 11Exp. A 83 1.5 (1.0) 0.16 1.8 (1.4) 0.18Exp. B 48 1.3 (1.0) 0.09 1.2 (1.1) 0.13Exp. C 113 1.2 (0.9) 0.09 1.0 (1.0) 0.12
Subject means 1.3 (1.0) 0.11 1.3 (1.2) 0.14Macaque 13
Exp. A 68 1.5 (1.0) 0.15 #Exp. B 83 1.0 (0.9) 0.11 1.0 (1.0) 0.12Exp. C 85 1.1 (1.0) 0.07 1.2 (0.9) 0.20
Subject means 1.2 (1.0) 0.11 1.1 (0.9) 0.16Macaque 14
Exp. A 53 1.3 (1.1) 0.11 1.2 (1.1) 0.14Exp. B 71 1.2 (0.9) 0.12 1.4 (1.3) 0.16Exp. C 84 1.1 (0.9) 0.08 1.2 (1.2) 0.13
Subject means 1.2 (1.0) 0.10 1.3 (1.2) 0.14Grand means 1.2 (1.0) 0.11 1.2 (1.1) 0.15
1 Values in parentheses (Mean) indicate mean W/B ratio for 10 power strokes (5 from each side), with greatest balancing-sidecontribution based on BAT muscle. N, number of power strokes; SDlog10, standard deviation of log-transformed values. #, noelectrodes for this muscle, or electrode failed.
TEMPORALIS EMG 41
much larger amount (67%, 62%, and 61%, respec-tively). For the b-s posterior temporalis, these valuesare 45% for baboons, 32% for macaques, 48% for owlmonkeys, 28% for galagos, and 33% for lemurs.
During the chewing of less resistant foods (anthro-poids only), the w-s percentages fall to 27% and 28%in baboons and macaques, respectively, and 49% forowl monkeys. The b-s percentages also fall for all
TABLE 1C. Owl monkeys1
Subject N
Anterior temporalis Posterior temporalis
Mean (Mean) SDlog10 Mean (Mean) SDlog10
Owl monkey 1Exp. A 32 1.3 (1.2) 0.10 1.3 (1.2) 0.08Exp. B 39 1.7 (1.2) 0.15 1.8 (1.2) 0.19Exp. C 56 1.2 (1.0) 0.14 1.2 (1.1) 0.11Exp. D 21 1.9 (1.5) 0.15 1.9 (1.8) 0.12
Subject means 1.5 (1.2) 0.14 1.5 (1.3) 0.13Owl monkey 2
Exp. A 42 1.2 (1.0) 0.13 1.3 (1.2) 0.09Exp. B 57 1.6 (1.2) 0.16 1.6 (1.3) 0.14Exp. C 69 1.3 (1.2) 0.09 1.2 (1.1) 0.11Exp. D 75 1.4 (1.2) 0.09 #
Subject means 1.4 (1.1) 0.12 1.4 (1.2) 0.11Grand means 1.4 (1.2) 0.13 1.4 (1.3) 0.12
1 Values in parentheses (Mean) indicate mean W/B ratio for 10 power strokes (5 from each side), with greatest balancing-sidecontribution based on BAT muscle. N, number of power strokes; SDlog10, standard deviation of log-transformed values. #, noelectrodes for this muscle, or electrode failed.
TABLE 1D. Thick-tailed galagos1
Subject N
Anterior temporalis Posterior temporalis
Mean (Mean) SDlog10 Mean (Mean) SDlog10
Galago 6Exp. A 45 4.4 (3.1) 0.19 3.5 (2.6) 0.29
Subject means 4.4 (3.1) 0.19 3.5 (2.6)Galago 7
Exp. A 37 # 2.0 (2.0) 0.13Subject means 2.0 (2.0)
Galago 8Exp. A 38 # 2.0 (1.8) 0.10
Subject means 2.0 (1.8)Grand means 4.4 (3.1) 0.19 2.4 (2.1) 0.17
1 Values in parentheses (Mean) indicate mean W/B ratio for 10 power strokes (5 from each side) with greatest balancing-sidecontribution based on BAT muscle. N, number of power strokes; SDlog10, standard deviation of log-transformed values. #, noelectrodes for this muscle, or electrode failed.
TABLE 1E. Ring-tailed lemurs1
Subject N
Anterior temporalis Posterior temporalis
Mean (Mean) SDlog10 Mean (Mean) SDlog10
Ring-tailed lemur 1Exp. C 51 2.2 (1.7) 0.11 2.5 (2.1) 0.10
Subject means 2.2 (1.7) 2.5 (2.1)Ring-tailed lemur 2
Exp. B 44 1.4 (1.2) 0.10 2.5 (2.5) 0.17Exp. C 49 1.3 (1.2) 0.09 2.3 (2.4) 0.15Exp. D 33 1.6 (1.4) 0.08 3.1 (2.9) 0.13
Subject means 1.4 (1.3) 2.6 (2.6)Ring-tailed lemur 3
Exp. A 43 1.9 (1.4) 0.12 2.1 (1.7) 0.14Exp. B 41 1.3 (1.0) 0.12 1.7 (1.6) 0.12
Subject means 1.6 (1.2) 1.9 (1.6)Ring-tailed lemur 4
Exp. A 44 1.4 (1.1) 0.11 1.6 (1.3) 0.20Exp. B 67 1.1 (0.8) 0.15 1.3 (1.1) 0.10Exp. C 37 1.7 (1.3) 0.12 1.6 (1.2) 0.17
Subject means 1.4 (1.0) 1.5 (1.2)Grand means 1.5 (1.2) (0.11) 2.0 (1.8) (0.14)
1 Values in parentheses (Mean) indicate mean W/B ratio for 10 power strokes (5 from each side), with greatest balancing-sidecontribution based on BAT muscle. N, number of power strokes; SDlog10, standard deviation of log-transformed values.
42 W.L. HYLANDER ET AL.

three anthropoid species, but for baboons and ma-caques their values dramatically fall to 4% and 2%,respectively, whereas for owl monkeys the fall isonly to about 37%.
Firing patterns of anterior and posteriortemporalis muscles
Table 3 presents the descriptive statistics for thefiring patterns of the temporalis muscles for all fivegroups. Positive values indicate that peak EMG of agiven muscle precedes peak activity of the referencemuscle, the w-s superficial masseter (a triplet II anddiagonal II muscle), and negative values indicatethe reverse. Table 4 contains summary data of thegrand mean values for each group.
Figure 7 presents the mean timing of the refer-ence muscle, the w-s superficial masseter, and theanterior and posterior temporalis muscles through-out the power stroke for one subject of each of thefive groups. In addition to the timing of peak EMGvalues, these figures also indicate the relative tim-ing of 25%, 50%, and 75% of peak EMGs duringloading and unloading.
Baboons and macaques
Similar to our earlier study of the superficial anddeep masseter (Hylander et al., 2000), the overallfiring patterns of the anterior and posterior tempo-ralis for the two cercopithecine species (Fig. 7A,B)are remarkably similar, and therefore will be de-scribed together.
Working-side temporalis. Peak EMG activity ofthe w-s anterior temporalis (a triplet I and diagonalI muscle) precedes activity of the w-s superficialmasseter in baboons and macaques (grand means �16 ms and 8 ms, respectively). These values are allsignificantly larger than zero (P � 0.02). Peak EMGactivity of the w-s posterior temporalis (a triplet Iand diagonal I muscle) also precedes activity of thew-s superficial masseter in baboons and macaques(grand means � 20 msec and 7 msec, respectively).
Once again, these values are all significantly largerthan zero (P � 0.02).
Balancing-side temporalis. Unlike the w-s an-terior temporalis, peak EMG of the b-s anterior tem-poralis (a triplet II and diagonal II muscle) occursafter the w-s superficial masseter. The grand meansindicate that peak EMG in both species occurs about6 ms after the w-s superficial masseter, but thesevalues are not significantly different from zero (P �0.05). Peak EMG of the b-s posterior temporalis (atriplet II and diagonal II muscle) also occurs afterthe w-s superficial masseter (grand means � �19ms and �16 ms, respectively). These mean valuesare significantly different from zero (P � 0.01).
Anterior vs. posterior temporalis. None of thegrand mean values for the w-s anterior temporalisare significantly different from the correspondingw-s posterior temporalis, whereas the b-s values forthese two muscles are significantly different fromone another (baboons, P � 0.01; macaques, P �0.02). Overall, the b-s posterior temporalis peaksafter the b-s anterior temporalis.
Owl monkeysWorking-side temporalis. Similar to baboonsand macaques, peak EMG activity of the w-s ante-rior and posterior temporalis muscles (Fig. 7C) pre-cedes activity of the w-s superficial masseter (grandmeans � 13 ms for both the anterior and posteriortemporalis), and these values are significantlylarger than zero (P � 0.01).
Balancing-side temporalis. Also similar to ba-boons and macaques, the b-s anterior and posteriortemporalis muscles reach peak activity after the w-ssuperficial masseter (grand means � �14 ms and�15 ms for the anterior and posterior temporalis,respectively). Unlike baboons and macaques, how-ever, rather than just the grand mean for the poste-rior temporalis, grand mean values for both the an-terior and posterior temporalis are significantlydifferent from zero (P � 0.05). Moreover, note thatthe amount of asynchrony between the b-s posteriortemporalis and the reference muscle is quite pro-nounced for owl monkey 1. This is the same subjectthat exhibited the reversed timing of the two super-ficial masseters (Hylander et al., 2000).
Anterior vs. posterior temporalis. The timingvalues of the ipsilateral (same side) anterior andposterior temporalis muscles are virtually identical,and not significantly different.
Thick-tailed galagos
Our results for galagos (Fig. 7D) should be consid-ered preliminary because our sample of successfulrecording sessions is small. Furthermore, recall thatwhereas for anthropoids, the w-s and b-s anteriortemporalis muscles are said to be members of trip-lets I and II, respectively, for strepsirrhines, these
TABLE 2. Summary of mean W/B EMG ratios1
Anteriortemporalis
Posteriortemporalis
Mean (Mean) Mean (Mean)
Softer foodsBaboons 2.8 7.2Macaques 4.1 7.6Owl monkeys 1.5 1.3
Brittle/tough foodsBaboons 1.2 (1.0) 1.0 (0.9)Macaques 1.2 (1.0) 1.2 (1.1)Owl monkeys 1.4 (1.2) 1.4 (1.3)Thick-tailedgalagos
4.4 (3.1) 2.4 (2.1)
Ring-tailedlemurs
1.5 (1.2) 2.0 (1.8)
1 Values in parentheses (Mean) indicate mean W/B ratio for 10power strokes (5 from each side) with greatest balancing-sidecontribution.
TEMPORALIS EMG 43

muscles may be members of the symmetric verticalclosers (Weijs, 1994).
Working-side temporalis. In the one galago re-cording session, the w-s anterior temporalis pre-cedes the w-s superficial masseter (experimentmean � 13 ms). This value, which is based on theindividual chews from one experiment, is signifi-cantly larger than zero (P � 0.001). Similarly, peakEMG activity of the w-s posterior temporalis alsoprecedes activity of the w-s superficial masseter(grand mean � 14 ms). Our sample for the posteriortemporalis is restricted to only three experiments,and the grand mean value of 14 ms is not signifi-cantly larger than zero (P � 0.125). Furthermore, foreach individual recording session, each experimentmean value, which is based on individual chews, issignificantly larger than zero (P � 0.001).
Balancing-side temporalis. Similar to anthro-poids, the galago b-s anterior and posterior tempo-
ralis muscles reach peak activity after the w-s su-perficial masseter (mean � �12 ms and about �3ms, respectively). The mean value for the one record-ing session of the anterior temporalis is significantlydifferent from zero (P � 0.002). The grand meanvalue for the posterior temporalis is not significantlydifferent from zero. Furthermore, in one of the threerecording sessions, the b-s posterior temporalis ac-tually preceded the w-s superficial masseter.
Anterior vs. posterior temporalis. In the onesession where simultaneous EMG from both the an-terior and posterior temporalis were recorded, thedata suggest that peak EMG values for the ipsilat-eral anterior and posterior temporalis occur nearlysimultaneously.
Ring-tailed lemursWorking-side temporalis. Similar to the anthro-poids and galagos, peak EMG activity of the w-santerior and posterior temporalis muscles (Fig. 7E)
Fig. 6. Bar graphs of average percent EMG activity in (A) baboons, (B) macaques, (C) owl monkeys, (D) thick-tailed galagos, and(E) ring-tailed lemurs during chewing of B/T foods. WAT, working-side anterior temporalis; BAT, balancing-side anterior temporalis;WPT, working-side posterior temporalis; BPT, balancing-side posterior temporalis.
TABLE 3. Timing differences (in ms) between peak EMG activity of working- and balancing-side anteriorand posterior temporalis muscles relative to reference muscle, i.e., working-side superficial masseter,
during mastication of brittle/tough foods for each of five groupsTABLE 3A. Baboons1
Subject N
Triplet I and diagonal I, working Triplet II and diagonal II, balancing
Anteriortemporalis
Posteriortemporalis
Anteriortemporalis
Posteriortemporalis
Mean SD Mean SD Mean SD Mean SD
Baboon 3Exp. A 64 28 20.9 22 20.9 10 17.9 �16 19.0Exp. B 52 24 24.0 20 19.2 0 20.4 �18 19.9Exp. C 48 9 25.3 18 24.1 �15 26.9 �29 28.9
Subject means 20 23.4 20 21.4 �2 21.7 �21 22.6Baboon 4
Exp. A 68 18 18.5 20 19.5 �6 16.7 �21 20.6Exp. B 60 18 23.0 26 23.2 �15 21.6 �17 25.9Exp. C 81 20 20.2 24 19.7 �4 19.1 �25 23.8
Subject means 19 20.6 23 20.8 �8 19.1 �21 23.4Baboon 5
Exp. A 75 17 15.1 19 14.5 �6 14.8 �14 12.9Exp. B 70 6 16.8 15 17.0 �8 21.0 �17 16.8Exp. C 58 6 14.1 20 14.6 �12 12.8 �13 15.1
Subject means 10 15.3 18 15.4 �9 16.2 �15 14.9Grand means 16.2 19.8 20.4 19.2 �6.2 19.0 �18.9 20.3
1 Positive values indicate that balancing-side superficial masseter or temporalis muscles precede working-side superficial masseter. N,number of power strokes; SD, standard deviation.
44 W.L. HYLANDER ET AL.

precedes activity of the w-s superficial masseter(grand means � 23 ms and 17 ms, respectively).Each of these means is significantly larger than zero(P � 0.01).
Balancing-side temporalis. The b-s anteriortemporalis muscle reaches peak activity before thew-s superficial masseter (grand mean � 6 ms),whereas the b-s posterior temporalis reaches peakactivity along with the w-s superficial masseter(grand mean � �0.1 ms). Unlike the mean value forthe posterior temporalis, the anterior temporalis issignificantly different from zero (P � 0.01).
Anterior vs. posterior temporalis. On average,the ipsilateral anterior temporalis for both the work-ing and balancing sides tends to fire significantlyearlier than the ipsilateral posterior temporalis (P �0.01).
Rise, fall, and total time of activity ofsuperficial masseter and temporalis muscles
Table 5 presents the descriptive statistics for theduration of the rise, fall, difference between rise andfall, and total time of EMG activity of the superficialmasseter and temporalis muscles during the chew-ing of B/T foods. Positive values for the differencebetween rise and fall times indicate that the risetime is of longer duration. Negative values indicatethe reverse condition. Table 6 contains summarydata of the grand mean values for each group.
There is a considerable amount of variability inthe mean values for the five groups. Nevertheless,an interesting pattern emerged. For baboons andmacaques, the rise and fall times for all musclesanalyzed tended to be either of near equal duration,or the fall time tended to be longer than the rise
TABLE 3B. Macques1
Subject N
Triplet I and diagonal I, working Triplet II and diagonal II, balancing
Anteriortemporalis
Posteriortemporalis
Anteriortemporalis
Posteriortemporalis
Mean SD Mean SD Mean SD Mean SD
Macaque 11Exp. A 83 11 18.1 12 18.2 �9 12.5 �17 14.8Exp. B 48 4 16.5 2 20.1 �6 13.0 �23 12.9Exp. C 113 15 14.2 20 14.0 �2 11.1 �4 10.2
Subject means 10 16.3 11 17.4 �6 12.2 �15 12.6Macaque 13
Exp. A 68 15 13.1 # �6 9.2 #Exp. B 83 7 15.0 17 17.4 1 13.7 �16 12.8Exp. C 85 18 20.3 7 28.8 10 19.1 �6 20.2
Subject means 13 16.1 12 23.1 2 14.0 �11 16.5Macaque 14
Exp. A 53 0 7.2 3 13.0 �17 13.5 �22 13.3Exp. B 71 �4 15.8 �6 14.9 �14 12.1 �27 14.9Exp. C 84 5 21.1 2 22.8 �15 21.0 �15 20.1
Subject means 0 14.7 0 16.9 �15 15.5 �21 16.1Grand means 7.9 15.7 7.1 19.1 �6.4 13.9 �16.3 15.1
1 Positive values indicate that balancing-side superficial masseter or temporalis muscles precede working-side superficial masseter. N,number of power strokes; SD, standard deviation. #, no electrodes for this muscle.
TABLE 3C. Owl monkeys1
Subject N
Triplet I and diagonal I, working Triplet II and diagonal II, balancing
Anteriortemporalis
Posteriortemporalis
Anteriortemporalis
Posteriortemporalis
Mean SD Mean SD Mean SD Mean SD
Owl monkey 1Exp. A 32 22 22.4 21 19.2 �13 21.5 �12 18.6Exp. B 39 14 28.5 30 31.0 �21 26.0 �21 35.3Exp. C 56 4 36.1 6 34.0 �47 27.9 �47 29.5Exp. D 21 18 11.7 12 8.3 �19 13.9 �22 18.2
Subject means 15 24.7 17 23.1 �25 22.3 �26 25.4Owl monkey 2
Exp. A 42 11 8.4 9 9.5 0 6.6 �5 8.0Exp. B 57 18 11.6 14 12.0 1 10.4 �1 9.2Exp. C 69 3 7.7 6 5.8 �9 9.0 �11 6.6Exp. D 75 13 10.2 82 10.3 �2 9.6 �12 9.2
Subject means 11 9.5 10 9.4 �3 8.9 �6 8.3Grand means 12.9 17.1 13.3 16.3 �13.8 15.6 �15.0 16.8
1 Positive values indicate that balancing-side superficial masseter or temporalis muscles precede working-side superficial masseter. N,number of power strokes; SD, standard deviation.2 Times based on right posterior temporalis electrode only.
TEMPORALIS EMG 45

time. This contrasts with galagos and lemurs, wherefor the same muscles, the duration of the rise timetended to be longer than the fall time. Finally, owlmonkeys were intermediate in that the rise and falltimes of their w-s muscles were similar to those ofbaboons and macaques, whereas their b-s musclesbehaved more like those of galagos and lemurs.
DISCUSSION
This study has two main goals. First, in order toidentify possible links between biomechanical stressand the evolution of symphyseal fusion, we charac-terized the recruitment of relative force of the work-ing- (w-s) and balancing-side (b-s) anterior and pos-terior temporalis muscles for baboons, macaques,owl monkeys, thick-tailed galagos, and ring-tailedlemurs. We also characterized the rise (loading), fall(unloading), and total time of EMG activity for these
muscles. Second, to increase our understanding ofthe biomechanics of the power stroke, we analyzedthe firing patterns of these same muscles.
Jaw muscle recruitment and symphyseal fusion
It was suggested that symphyseal fusion strength-ens the symphysis so as to counter increased sym-physeal stress during chewing and biting (Beecher,1977, 1979; Hylander, 1975, 1977, 1979a, b; Ravosaand Hylander, 1994). Based primarily on an in vivoanalysis of strain along the mandibular symphysisof macaques (Hylander, 1984, 1985), we identifiedthree loading regimes that may be associated withincreased symphyseal stress and the evolution ofsymphyseal fusion in primates (Hylander et al.,1998). These regimes are: 1) lateral transverse bend-ing or “wishboning” due to increased transverselydirected b-s jaw-muscle force; 2) dorsoventral shear-
TABLE 3D. Thick-tailed galagos1
Subject N
Triplet I and diagonal I, working Triplet II and diagonal II, balancing
Anteriortemporalis2
Posteriortemporalis
Anteriortemporalis2
Posteriortemporalis
Mean SD Mean SD Mean SD Mean SD
Galago 6Exp. A 45 13 21.5 12 17.3 �12 19.4 �13 19.8
Subject means 13 21.5 12 17.3 �12 19.4 �13 19.8Galago 7
Exp. A 37 # 12 12.0 # �3 11.0Subject means 12 12.0 �3 11.0
Galago 8Exp. A 38 # 18 14.0 # 6 8.9
Subject means 18 14.0 6 8.9Grand means 13.0 21.5 14.0 14.4 �12.0 19.4 �3.3 13.2
1 Positive values indicate that balancing-side superficial masseter or temporalis muscles precede working-side superficial masseter. N,number of power strokes; SD, standard deviation. #, no electrodes for this muscle.2 Anterior temporalis is not considered to be a triplet I or II muscle for strepsirrhines (Weijs, 1994).
TABLE 3E. Ring-tailed lemurs1
Subject N
Triplet I and diagonal I, working Triplet II and diagonal II, balancing
Anteriortemporalis2
Posteriortemporalis
Anteriortemporalis2
Posteriortemporalis
Mean SD Mean SD Mean SD Mean SD
Ring-tailed lemur 1Exp. C 51 7 11.2 6 11.1 �4 13.1 �7 14.1
Subject means 7 11.2 6 11.1 �4 13.1 �7 14.1Ring-tailed lemur 2
Exp. B 44 18 18.3 15 20.1 5 11.7 �6 12.1Exp. C 49 27 16.9 25 11.0 9 12.2 3 11.0Exp. D 33 31 14.8 28 10.5 5 15.7 �1 8.3
Subject means 25 16.7 23 13.9 6 13.2 �1 10.5Ring-tailed lemur 3
Exp. A 43 31 13.8 25 11.9 6 12.7 1 7.1Exp. B 41 28 22.4 16 16.0 8 13.7 1 14.3
Subject means 30 18.1 21 14.0 7 13.2 1 10.7Ring-tailed lemur 4
Exp. A 44 22 15.1 5 11.0 6 18.9 �4 10.2Exp. B 67 22 15.3 19 13.8 11 15.0 4 13.1Exp. C 37 22 11.6 16 10.4 9 13.1 8 16.4
Subject means 22 14.0 13 11.7 9 15.7 3 13.2Grand means 23.1 15.5 17.2 12.9 6.1 14.0 �0.1 11.8
1 Positive values indicate that balancing-side superficial masseter or temporalis muscles precede working-side superficial masseter. N,number of power strokes; SD, standard deviation.2 Anterior temporalis is not considered to be a triplet I or II muscle for strepsirrhines (Weijs, 1994).
46 W.L. HYLANDER ET AL.

ing of the symphysis due to increased vertically di-rected b-s jaw-muscle force; and 3) frontal bending ofthe symphysis associated with twisting of the man-dibular corpus due to increased vertically directedb-s jaw-muscle force.
As one or more of these three loading regimes maybe importantly linked to symphyseal fusion in pri-mates, we formulated the following three hypothe-ses (Hylander et al., 1998, 2000). Hypothesis 1 statesthat the evolution of symphyseal fusion in primatesis linked to both increased vertically directed andtransversely directed b-s forces from the jaw closingmuscles. Hypothesis 2 states that the evolution ofsymphyseal fusion in primates is linked to only in-creased transversely directed b-s jaw muscle force.Hypothesis 3 states that the evolution of symphysealfusion in primates is linked to only increased verti-cally directed b-s jaw muscle force.
Following our EMG analysis of the superficial anddeep masseter in baboons, macaques, owl monkeys,galagos, and ring-tailed lemurs (Hylander et al.,2000, 2002, 2004), we concluded that the data pro-vide strong support for hypothesis 2, refute hypoth-esis 3, and arguably refute hypothesis 1. Moreover,data from Lieberman and Crompton (2000) demon-strated small W/B ratios for the vertically alignedjaw-closing muscles in goats and opossums, two spe-cies with unfused mobile symphyses. Therefore,these data also arguably refute hypotheses 1 and 3.We previously noted, however, that an EMG analy-sis of additional portions of the jaw adductor musclemass in primates is needed so as to provide a morethorough test of these hypotheses. The present anal-ysis of the temporalis muscle provides additionaldata to do so, particularly hypothesis 1, which statesthat symphyseal fusion is linked to both verticallyand transversely directed muscle forces.
For functional reasons, the temporalis of primatesis arbitrarily divided into anterior and posterior por-tions. The muscle fiber arrangements of these por-tions are such that the anterior temporalis pulls the
mandible primarily upward, whereas the posteriorportion pulls the mandible mainly upward and back-ward. If symphyseal fusion is an adaptation tostrengthen the symphysis so as to counter increasedmechanical stress associated with increased verticalforce, as stated by hypotheses 1 and 3, then duringforceful chewing, anthropoids should recruit moreb-s anterior temporalis force than do those strepsir-rhines with highly mobile symphyses.
Using our temporalis EMG data, one test of thesehypotheses is the following. If we focus exclusivelyon data recorded for the three anthropoids and ring-tailed lemurs, the grand mean values for the W/Bratios of the anterior temporalis for baboons, ma-caques, owl monkeys, and lemurs are 1.2, 1.2, 1.4,and 1.5, respectively. For the more forceful powerstrokes, these mean values are all reduced to 1.0,1.0, 1.2, and 1.2, respectively. It is apparent thatthese values are all quite similar to one another.Furthermore, in all four species there is consider-able overlap in mean values for the individual re-cording sessions. We conclude, therefore, that thereare no significant differences in the recruitment ofrelative force of the b-s anterior temporalis betweenanthropoids and ring-tailed lemurs. Therefore,these data refute or at least provide no evidence tosupport hypotheses 1 and 3.
In contrast, the data for the one recording sessionfor galagos indicate relatively large W/B ratios forthe anterior temporalis (overall mean � 4.4 andlarger power stroke mean � 3.1), and these dataarguably support hypothesis 1. Nevertheless, even ifa large sample of galago recordings were to demon-strate that galagos do indeed have large W/B ratiosfor the anterior temporalis, the data for ring-tailedlemurs lead us to conclude that increased recruit-ment of vertically directed b-s jaw muscle force is notlikely linked to the evolution of symphyseal fusion inprimates. Furthermore, and as already noted, pre-vious studies of the superficial and deep massetermuscles for baboons, macaques, owl monkeys, gala-
TABLE 4. Summary of mean timing differences1
Triplet I and diagonal I, working Triplet II and diagonal II, balancing
Anterior temporalis,2
meanPosterior temporalis,
meanAnterior temporalis,2
meanPosterior temporalis,
mean
Soft foodsBaboons 7.9*** 11.9*** �2.4 �20.4*Macaques 8.1*** 15.3*** �2.6 �14.2*Owl monkeys 3.0 2.0 �6.5 �9.5
Brittle/tough foodsBaboons 16.2*** 20.4*** �6.2* �18.9***Macaques 7.9** 7.1** �6.4* �16.3***Owl monkeys 12.9*** 13.3*** �13.8** �15.0***Thick-tailed galagos 13.0 14.0 �12.0 �3.3Ring-tailed lemurs 23.1*** 17.2*** 6.1*** �0.1
1 Positive values indicate that balancing-side superficial masseter or temporalis muscles precede working-side superficial masseter.SD, standard deviation.2 Anterior temporalis is not considered to be a triplet I or II muscle for strepsirrhines (Weijs, 1994).*P � .10**P � .05.***P � .01.
TEMPORALIS EMG 47

Fig. 7. Mean values for timing of average percent EMG activity of working-side superficial masseter and working- and balancing-side anterior and posterior temporalis muscles during mastication of popcorn kernels in (A) baboon 5 and (B) macaque 11, and driedgelatin candy in (C) owl monkey 1, (D) galago 6, and (E) ring-tailed lemur 2. Vertical line indicates peak activity in reference muscle,i.e., working-side superficial masseter. Positive timing values indicate that a muscle reaches peak activity prior to peak activity ofreference muscle, whereas negative values indicate peak activity after reference muscle. Abbreviations as in Figure 6.
48 W.L. HYLANDER ET AL.

gos, and ring-tailed lemurs led us to a similar con-clusion, and provided strong support for hypothesis2, i.e., that increased recruitment of transverselydirected b-s muscle force is linked to symphysealfusion in primates (Hylander et al., 2000, 2002,2004).
The large galago ratios do, however, suggest thatcertain aspects of symphyseal morphology and in-creased vertically directed forces are linked. That is,compared to ring-tailed lemurs, galagos have littlesymphyseal fibrous connective tissue binding theirtwo dentaries together (Beecher, 1977). Further-
more, compared to ring-tailed lemurs, galagos havelarger W/B ratios for all jaw muscles analyzed by us(superficial and deep masseter and anterior and pos-terior temporalis; Hylander et al., 2000, 2004). Thus,although increased vertical force in lemurs may belinked to the increased strength of their symphysis,this increased force is not sufficient to necessitatesymphyseal fusion.
Finally, we are impressed by the variability inW/B ratios for the posterior temporalis muscles.These data indicate that anthropoids on averagehave relatively greater recruitment of their b-s pos-
TABLE 5. Rise, fall and total time of EMG activity durations during mastication of brittle/tough foods for each of five groupsTABLE 5A. Baboons1
Subject N
Anterior temporalis Posterior temporalis
Working Balancing Working Balancing
Rise2 Fall3 Diff.4 Total5 Rise Fall Diff. Total Rise Fall Diff. Total Rise Fall Diff. Total
Baboon 3Exp. A 64 65 94 �29 159 64 106 �42 170 48 59 �11 107 53 70 �17 123Exp. B 52 59 86 �27 145 60 85 �25 145 45 52 �7 97 51 62 �11 113Exp. C 48 73 98 �25 171 71 88 �17 159 * * * *
Subject means 66 93 �27 158 65 93 �28 158 47 56 �9 102 52 66 �14 118Baboon 4
Exp. A 68 79 84 �5 163 83 96 �13 179 48 51 �3 99 56 77 �21 133Exp. B 60 62 72 �10 134 70 69 1 139 53 60 �7 113 38 60 �22 98Exp. C 81 69 82 �13 151 76 95 �19 171 36 49 �13 85 44 79 �35 123
Subject means 70 79 �9 149 76 87 �11 163 46 53 �7 99 46 72 �26 118Baboon 5
Exp. A 75 66 67 �1 133 63 68 �5 131 39 50 �11 89 42 62 �20 104Exp. B 70 77 65 12 142 68 64 4 132 43 46 �3 89 42 48 �6 90Exp. C 58 61 66 �5 127 63 54 9 117 51 67 �16 118 43 63 �20 106
Subject means 68 66 2 134 65 62 3 127 44 54 �10 99 42 58 �16 100Grand means 68 79 �11 147 69 81 �12 149 45 54 �9 100 46 65 �19 111
1 N, number of chews in each experiment. Mean values in milliseconds.2 Duration of time from 25% of peak EMG activity to 100% of peak EMG activity during loading.3 Duration of time from 100% of peak EMG activity to 25% of peak EMG activity during unloading.4 Difference equals rise time minus fall time.5 Duration of time from 25% of peak EMG activity during loading to 25% of peak EMG activity during unloading.* Very low values prevent determination of rise and fall times.
TABLE 5B. Macaques1
Subject N
Anterior temporalis Posterior temporalis
Working Balancing Working Balancing
Rise2 Fall3 Diff.4 Total5 Rise Fall Diff. Total Rise Fall Diff. Total Rise Fall Diff. Total
Macaque 11Exp. A 83 59 66 �7 125 35 43 �8 78 56 56 0 112 27 43 �16 70Exp. B 48 59 66 �7 125 45 51 �6 96 55 56 �1 111 31 41 �10 72Exp. C 113 45 41 4 86 35 42 �7 77 41 50 �9 91 32 40 �8 72
Subject means 54 58 �3 112 38 45 �7 84 51 54 �3 105 30 41 �11 71Macaque 13
Exp. A 68 47 58 �11 105 37 44 �7 81 54 56 �2 110 31 44 �13 75Exp. B 83 57 57 0 114 48 50 �2 98 58 58 0 116 41 48 �7 89Exp. C 85 69 72 �3 141 55 62 �7 117 61 66 �5 127 36 52 �16 88
Subject means 58 62 �4 120 47 52 �5 99 58 60 �2 118 36 48 �12 84Macaque 14
Exp. A 53 46 62 �16 108 38 42 �4 80 41 58 �17 99 30 39 �9 69Exp. B 71 50 64 �14 114 36 49 �13 85 45 59 �14 104 30 44 �14 74Exp. C 84 64 78 �14 142 50 57 �7 107 64 68 �4 132 42 58 �16 100
Subject means 53 68 �15 121 41 49 �8 91 50 62 �12 112 34 47 �13 81Grand means 55 63 �8 118 42 49 �7 91 53 59 �6 111 33 45 �12 79
1 N, number of chews in each experiment. Mean values in milliseconds.2 Duration of time from 25% of peak EMG activity to 100% of peak EMG activity during loading.3 Duration of time from 100% of peak EMG activity to 25% of peak EMG activity during unloading.4 Difference equals rise time minus fall time.5 Duration of time from 25% of peak EMG activity during loading to 25% of peak EMG activity during unloading.
TEMPORALIS EMG 49

terior temporalis muscles. That is, W/B ratios forbaboons, macaques, owl monkeys, galagos, and le-murs are 1.0, 1.2, 1.4, 2.4, and 2.0, respectively. Thissuggests that anthropoids may emphasize an in-crease in posteriorly directed b-s muscle force on themandible during mastication Similar to the lateralcomponent of force from the b-s deep masseter, per-haps posteriorly directed force on the mandible fromthe b-s posterior temporalis is also linked to symph-yseal fusion in primates. We do not see this as likely,however, simply because force from the b-s posteriortemporalis does not appear to cause forceful bendingor twisting of the macaque symphysis (Hylander,1984). On the other hand, as compared to strepsir-rhines, for anthropoids this increase in relative forcefrom the b-s posterior temporalis may result in anincrease in magnitude of the power stroke.
In summary, our anterior temporalis data for pri-mates provide no support for linking symphysealfusion and increased recruitment of vertically di-
rected b-s muscle force. We are, however, reluctantto dismiss completely hypothesis 1, because in-creased vertically directed force in crown anthro-poids may simply be due to an absolute increase inthe mass of the anterior temporalis (Cachel, 1979;Ross, 2000; Ross and Hylander, 2000), rather thanto the evolution of altered recruitment patterns, asrevealed by W/B ratios.2 Also, compared to strepsir-rhines, extant anthropoids are characterized by amore anteriorly repositioned and vertically alignedanterior temporalis muscle (Ross, 1995a, b, 2000;Ross and Hylander, 2000). This repositioning (andincreased size?) likely results in an increase in clos-ing moments associated with muscle force from the
2Perhaps increased vertically directed b-s force due to enlargedtemporalis muscles is largely responsible for symphyseal fusion invarious large-bodied carnivorans, such as certain ursids and felids(Scapino, 1981).
TABLE 5C. Owl monkeys1
Subject N
Anterior temporalis Posterior temporalis
Working Balancing Working Balancing
Rise2 Fall3 Diff.4 Total5 Rise Fall Diff. Total Rise Fall Diff. Total Rise Fall Diff. Total
Owl monkey 1Exp. A 32 53 63 �10 116 71 46 25 117 54 63 �9 117 68 56 12 124Exp. B 39 86 85 1 171 102 69 33 171 72 101 �29 173 98 70 28 168Exp. C 56 73 81 �8 154 84 59 25 143 81 80 1 161 98 60 38 158Exp. D 21 63 56 7 119 91 47 44 138 77 57 20 134 99 46 53 145
Subject means 69 71 �3 140 87 55 32 142 71 75 �4 146 91 58 33 149Owl monkey 2
Exp. A 42 37 42 �5 79 44 41 3 85 36 42 �6 78 43 41 2 84Exp. B 57 41 46 �5 87 50 45 5 95 39 43 �4 82 52 42 10 94Exp. C 69 40 46 �6 86 41 41 0 82 41 49 �8 90 46 42 4 88Exp. D 75 42 48 �6 90 48 43 5 91 38b 45 �7 83 456 43 2 88
Subject means 40 46 �6 86 46 43 3 88 39 45 �6 83 47 42 5 89Grand means 54 58 �4 113 66 49 18 115 55 60 �5 115 69 50 19 119
1 N, number of chews in each experiment. Mean values in milliseconds.2 Duration of time from 25% of peak EMG activity to 100% of peak EMG activity during loading.3 Duration of time from 100% of peak EMG activity to 25% of peak EMG activity during unloading.4 Difference equals rise time minus fall time.5 Duration of time from 25% of peak EMG activity during loading to 25% of peak EMG activity during unloading.6 Times based on right posterior temporalis electrode only.
TABLE 5D. Thick-tailed galagos1
Subject N
Anterior temporalis Posterior temporalis
Working Balancing Working Balancing
Rise2 Fall3 Diff.4 Total5 Rise Fall Diff. Total Rise Fall Diff. Total Rise Fall Diff. Total
Galago 6Exp. A 45 74 56 18 130 65 44 21 109 73 49 24 122 42 42 0 84
Subject means 74 56 18 130 65 44 21 109 73 49 24 122 42 42 0 84Galago 7
Exp. A 37 97 46 51 143 100 42 58 142Subject means 97 46 51 143 100 42 58 142
Galago 8Exp. A 38 62 49 13 111 62 47 15 109
Subject means 62 49 13 111 62 47 15 109Grand means 74 56 18 130 65 44 21 109 77 48 29 125 68 44 24 112
1 N, number of chews in each experiment. Mean values in milliseconds.2 Duration of time from 25% of peak EMG activity to 100% of peak EMG activity during loading.3 Duration of time from 100% of peak EMG activity to 25% of peak EMG activity during unloading.4 Difference equals rise time minus fall time.5 Duration of time from 25% of peak EMG activity during loading to 25% of peak EMG activity during unloading.
50 W.L. HYLANDER ET AL.

b-s anterior temporalis, which in turn may causeincreased levels of dorsoventral shear and frontalbending of the symphysis.
Rise time, fall time, and symphyseal fusion
We originally hypothesized that compared tostrepsirrhines, anthropoid jaw-closing muscles mayhave a relatively longer fall time during forcefulmastication. For example, a comparison of w-s man-dibular bone strain patterns of macaques, owl mon-keys, and galagos (Fig. 8A–C) demonstrates differ-ences between the duration of rise and fall times ofmasticatory forces during the power stroke. Fur-thermore, we also suspected that longer fall times inanthropoids are functionally linked to symphysealfusion. Data in Tables 5 and 6 partially allow us totest this hypothesis.
For the five primate groups analyzed, there isconsiderable amount of variability in the rise, fall,and total time duration for the anterior and poste-rior temporalis muscles. For example, for baboonsand macaques, either the rise and fall times tend tobe of near equal duration, or the fall time is longerthan the rise time. This contrasts with galagos andlemurs, where the duration of the rise time is usu-ally relatively longer than the fall time. For owlmonkeys, however, the rise and fall times of theirw-s muscles are similar to the pattern of baboonsand macaques, whereas their b-s muscles are moresimilar to those of galagos and lemurs.
We conclude from these data that there is no link-age between a prolonged fall time of individual mus-cles and symphyseal fusion. This is simply because ifprolonged fall times are linked to increased symph-
TABLE 5E. Ring-tailed lemurs1
Subject N
Anterior temporalis Posterior temporalis
Working Balancing Working Balancing
Rise2 Fall3 Diff.4 Total5 Rise Fall Diff. Total Rise Fall Diff. Total Rise Fall Diff. Total
Ring-tailed lemur 1Exp. C 51 56 45 11 101 58 44 14 102 55 43 12 98 62 43 19 105
Subject means 56 45 11 101 58 44 14 102 55 43 12 98 62 43 19 105Ring-tailed lemur 2
Exp. B 44 74 59 15 133 88 50 38 138 61 51 10 112 74 45 29 119Exp. C 49 74 58 16 132 91 50 41 141 58 51 7 109 74 49 25 123Exp. D 33 80 58 22 138 92 48 44 140 53 53 0 106 66 43 23 109
Subject means 76 58 18 134 90 49 41 140 57 52 6 109 71 46 26 117Ring-tailed lemur 3
Exp. A 43 78 60 18 138 94 45 49 139 83 52 31 135 86 42 44 128Exp. B 41 68 67 1 135 97 50 47 147 81 51 30 132 101 45 56 146
Subject means 73 64 10 137 96 48 48 143 82 52 31 134 94 44 50 137Ring-tailed lemur 4
Exp. A 44 71 60 11 131 89 50 39 139 76 46 30 122 82 42 40 124Exp. B 67 54 54 0 108 65 52 13 117 61 49 12 110 70 48 22 118Exp. C 37 54 52 2 106 72 48 24 120 55 46 9 101 61 50 11 111
Subject means 60 55 4 115 75 50 25 125 64 47 17 111 71 47 24 118Grand means 68 57 11 125 83 49 34 131 65 49 16 114 75 45 30 120
1 N, number of chews in each experiment. Mean values in milliseconds.2 Duration of time from 25% of peak EMG activity to 100% of peak EMG activity during loading.3 Duration of time from 100% of peak EMG activity to 25% of peak EMG activity during unloading.4 Difference equals rise time minus fall time.5 Duration of time from 25% of peak EMG activity during loading to 25% of peak EMG activity during unloading.
TABLE 6. Summary for rise, fall, and total time of EMG activity for brittle/tough foods
Brittle/Tough Foods
Anterior temporalis Posterior temporalis
Working Balancing Working Balancing
Rise1 Fall2 Diff.3 Total4 Rise Fall Diff. Total Rise Fall Diff. Total Rise Fall Diff. Total
Baboons 68 79 �11** 147 69 81 �12* 149 45 54 �9*** 100 46 65 �19*** 111Macaques 55 63 �8** 118 42 49 �7*** 91 53 59 �6** 111 33 45 �12*** 79Owl Monkeys 54 58 �4 113 66 49 18** 115 55 60 �5 115 69 50 19 119Thick-tailed galagos 74 56 18 130 65 44 21 109 77 48 29 125 68 44 24 112Ring-tailed lemurs 68 57 11*** 125 83 49 34*** 131 65 49 16*** 114 75 45 30*** 120
N � number of chews in each experiment. Mean values in milliseconds.1 Duration of time from 25% of peak EMG activity to 100% of peak EMG activity during loading.2 Duration of time from 100% of peak EMG activity to 25% of peak EMG activity during unloading.3 The difference equals rise time minus fall time.4 Duration of time from 25% of peak EMG activity during loading to 25% of peak EMG activity during unloading.*P � .10**P � .05***P � .01
TEMPORALIS EMG 51

yseal stress and a necessary strengthening of thesymphysis, we would expect that this would be truefor the balancing-side muscles. Instead, anthropoidsuniformly differ from strepsirrhines in only havingprolonged fall times of their working-side muscles.As noted above, unlike baboons and macaques, owlmonkeys do not have a prolonged fall time of theirb-s muscles. Finally, for individual muscles, the to-tal time of EMG from 25% of peak activity duringloading to 25% of peak activity during unloadingdoes not indicate significant differences in the dura-tion of the power stroke between anthropoids andstrepsirrhines.
On the other hand, as noted earlier, the delayedactivity of the b-s posterior temporalis and b-s deepmasseter in anthropoids leads us to suspect thatanthropoids may have a power stroke of longer du-ration than these strepsirrhines (Hiiemae and Kay,1973). Nevertheless, an EMG analysis of jaw closingmuscles may not the best method available for char-acterizing the duration of the power stroke. Instead,
perhaps the best method for characterizing its du-ration (and magnitude) is by utilizing w-s mandib-ular corpus bone strain data (Hylander, 1986; Hy-lander et al., 1987).
Differential firing and recruitment patterns ofjaw muscles and symphyseal fusion
Previous work on macaques, baboons, owl mon-keys, tamarins, and marmosets suggested that theseanthropoids share a common b-s deep masseter fir-ing pattern during mastication (Hylander and John-son, 1994; Hylander et al., 2000; Vinyard et al.,2001, 2004a). On the other hand, galagos, ring-tailed lemurs, treeshrews, and opossums do nothave this pattern 1980 (Hylander et al., 2000, 2002,2004; Vinyard et al., 2004b). For nonhuman anthro-poids, the last masseter muscle to reach peak activ-ity is the b-s deep (Figs. 2C, 9A). For galagos, ring-tailed lemurs, tree shrews, and opossums, the b-sdeep masseter reaches peak activity earlier, and thelast muscle to reach peak activity is the w-s super-ficial masseter (Fig. 9B–D). Thus, there is a distinctdifference in firing of the b-s deep masseter betweenthick-tailed galagos and ring-tailed lemurs on theone hand, and anthropoids on the other. We arguedelsewhere that the presence or absence of the late-firing b-s deep masseter determines whether or notthe primate mandible experiences wishboning dur-ing mastication (Hylander and Johnson, 1994). Fur-thermore, when wishboning does occur, it has thepotential to cause intense tensile stress along thelingual aspect of the symphysis. The most effectiveway to counter these loads is to fuse the symphysis,as cortical bone is 5–10 times stronger than liga-ment (Panjabi and White, 2001). The cost of fusion,however, includes loss of independent movement ofthe dentaries, which in turn may influence occlusalfunction (Hylander, 1979b; Lieberman and Cromp-ton, 2000).
In summary, the data on recruitment and firingpatterns of jaw-closing muscles support the hypoth-esis that in primates, symphyseal fusion and trans-versely directed muscle force during the powerstroke are functionally linked (hypothesis 2). Unlikeanthropoids, muscle recruitment and firing patterndata for the b-s deep masseter suggest that galagoand lemur symphyses experience little if any wish-boning. This is because of two major factors. First,for galagos and lemurs (also opossums and treeshrews), the overall bending moment that causeswishboning is relatively small because the force as-sociated with this moment, i.e., the force from theirbalancing-side deep masseter is relatively small.Second, and more importantly, whereas the b-s deepmasseter (and b-s posterior temporalis) of nonhu-man anthropoids reaches peak activity followingpeak activity of the other jaw-closing muscles, ingalagos and lemurs (also opossums and treeshrews), peak activity of the b-s deep masseter (andb-s posterior temporalis) occurs at a time when theother jaw-closers are highly active. Therefore, it is
Fig. 8. Working-side mandibular corpus shear strain duringmastication of (A) monkey chow in macaque 4, (B) hard prune inowl monkey 2, and (C) dried gelatin candy in galago 7 (data fromHylander et al., 1998).
52 W.L. HYLANDER ET AL.

quite likely that for galagos and lemurs, the mo-ments associated with the force from the medial(and lateral?) pterygoid muscles likely counter thewishboning moments of their deep (and superficial)masseter muscles. This is not the case for anthro-poids, simply because peak b-s deep masseter forcefollows well after most of the force occurs from theother jaw muscles, and therefore the moments asso-ciated with the remaining jaw muscles cannotcounter the wishboning moments associated withthe b-s deep masseter.
Stiffness, strength, and symphyseal fusion
Lieberman and Crompton (2000) suggested thatalthough increased transverse b-s muscle force isassociated with symphyseal fusion, and that fusionresults in increased symphyseal strength, the mostimportant characteristic associated with fusion is
that it provides increased stiffness (for a convincingcontrary view, see Hogue and Ravosa, 2001). Lieber-man and Crompton (2000) went on to suggest thatincreased symphyseal stiffness is particularly im-portant for those mammals that have a large trans-verse component to their power stroke, i.e., thosemammals that have relatively flat occlusal surfacesthat are nearly parallel to the horizontal plane. Thebasis for their argument as to the functional signif-icance of symphyseal fusion is that increased sym-physeal stiffness facilitates a more complete (moreefficient) transfer of transverse b-s muscle forceacross the symphysis.
In an attempt to deal with this issue, Liebermanand Crompton (2000) defined what they meant byefficiency. As Lieberman and Crompton (2000, p.519) clearly noted, “The term efficiency here is usedin this paper in the context of the timing and degree
Fig. 9. Mean values for timing of average percent EMG activity of deep and superficial masseters during chewing of B/T foods in(A) macaque 11, (B) galago 6, (C) opossum 1, and (D) treeshrew 1 (data from Hylander et al., 2000, unpublished data; Vinyard et al.,2004b). Vertical line indicates peak activity in reference muscle, i.e., working-side superficial masseter. WSM and BSM, w-s superficialmasseter and b-s superficial masseter; WDM and BDM, w-s deep masseter and b-s deep masseter. Remaining abbreviations as inFigure 6.
TEMPORALIS EMG 53

of force transfer. Stiffness increases the efficiency offorce transfer between two objects because no forceis stored elastically, or dissipated through move-ment or the generation of heat.”
There are a number of problems associated withtheir overall argument. For starters, when theseauthors claim that “no force is stored elastically, ordissipated through movement or the generation ofheat” in a stiff object such as a fused symphysis, theyare presumably referring to energy, rather than toforce. If so, their claim is incorrect, as all solid ob-jects (e.g., ligament, cartilage, and bone) are capableof storing strain energy. Moreover, and most impor-tantly, if energy storage, energy dissipation throughmovement, and energy dissipated as heat are impor-tant issues associated with the transfer of transverseb-s muscle force across an unfused symphysis, whydon’t these same issues apply to the transfer ofvertical b-s muscle force across an unfused symphy-sis? As Hogue and Ravosa (2001) noted, shouldn’tthe symphysis also fuse in response to increasedvertical forces so as to avoid these undesirable con-ditions? Finally, during force transfer across thesymphysis, mechanical energy can indeed be lost tothe system, and one way this can occur is when thetissues of the unfused symphysis experience therapid accumulation of microdamage. If so, increasedstrength due to partial or complete fusion helps pre-vent structural failure of the tissues of the symphy-sis.
Triplets, diagonals, and primates:predictions and data
Weijs (1994) noted that for “primitive” mammals(including “prosimians”),3 the w-s and b-s anteriortemporalis muscles fire simultaneously as membersof the SVC, whereas for anthropoids, these musclesfire as members of triplets I and II. Our secondmajor goal is to test two aspects of jaw-muscle func-tion. First, for anthropoids and strepsirrhines dur-ing rhythmic chewing, does the w-s posterior tempo-ralis fire as triplet I and diagonal I, and does the b-sposterior temporalis fire as triplet II and diagonal II(Weijs, 1994; Herring, 1992)? Second, for strepsir-rhines, do the two anterior temporalis muscles firesimultaneously, and do they fire prior to triplet I(Weijs, 1994)? Or, do their anterior temporalis mus-cles simply fire as triplets and diagonals I and II?
Triplets and diagonals I and II and posteriortemporalis. For all subjects, the w-s posteriortemporalis muscles reach peak activity prior to thew-s superficial masseter. Thus, these data indicatethat the firing pattern of the w-s posterior tempora-lis of primates conforms to the pattern predicted byboth the triplet and diagonal hypotheses. For ba-boons, macaques, and owl monkeys, the b-s posteriortemporalis reaches peak activity well after the w-s
superficial masseter, whereas for galagos and le-murs, it fires with the w-s superficial masseter.These data also indicate that the firing pattern forthe b-s posterior temporalis in primates conformsmore or less to the triplet and diagonal hypotheses.Furthermore, it appears that anthropoids may haveprolonged the duration of the power stroke by delay-ing the firing of the b-s posterior temporalis, as wellas the b-s deep masseter.
Symmetric vertical closers and anterior tempo-ralis. In all five groups of primates, the w-s ante-rior temporalis reaches peak activity prior to the w-ssuperficial masseter, and its timing is not signifi-cantly different from that of the w-s posterior tem-poralis. In baboons, macaques, owl monkeys, andgalagos, the b-s anterior temporalis reaches peakactivity slightly after the w-s superficial masseter,but only for owl monkeys are the mean values sig-nificantly different (P � 0.05). For lemurs, the b-santerior temporalis reaches peak activity before thew-s superficial masseter (P � 0.01), but well afterthe b-s superficial masseter muscle.
These data demonstrate that in strepsirrhine pri-mates, the w-s and b-s anterior temporalis musclesdo not fire symmetrically, nor do they only fire dur-ing fast closing, prior to triplet I. Instead, as inanthropoids, the w-s anterior temporalis alwaysleads the b-s anterior temporalis. Furthermore, thew-s anterior temporalis fires near simultaneouslywith the w-s posterior temporalis. On the balancingside, the anterior temporalis firing pattern is morevariable. For baboons and macaques, the b-s ante-rior temporalis fires slightly after the w-s superficialmasseter, but these differences are not statisticallysignificant from one another (P � 0.1). For owl mon-keys, the b-s anterior temporalis also fires after thew-s superficial masseter (P � 0.05). Finally, for le-murs, the b-s anterior temporalis fires before the w-ssuperficial masseter (P � 0.01), but still well aftertriplet I and diagonal I muscles.
In summary, it is clear that similar to anthro-poids, among strepsirrhines the w-s and b-s anteriortemporalis muscles do not fire simultaneously, norwell before triplet I and diagonal I. As a first approx-imation, the w-s and b-s temporalis muscles of pri-mates simply fire as members of triplets and diago-nals I and II, respectively. An arguable exception tothis pattern is that for anthropoids, the b-s posteriortemporalis firing peaks very late in the powerstroke. As in pigs (Herring, 1993), this late firing ofthe b-s posterior temporalis in anthropoids likelyassists the b-s deep masseter in driving the w-smolars through the terminal portion of the powerstroke (Hylander and Johnson, 1994).
CONCLUSIONS
The purpose of this study was to analyze the forcerecruitment and firing patterns of the anterior andposterior temporalis muscles in strepsirrhine andanthropoid primates. As indicated by the W/B ratios,3Weijs (1994) included treeshrews as prosimians.
54 W.L. HYLANDER ET AL.

the data for the anterior temporalis provide no sup-port for the hypothesis that symphyseal fusion inprimates is linked to vertically directed jaw muscleforces during forceful mastication. Thus, indirectly,these data provide support for the hypothesis thatsymphyseal fusion in primates is most importantlylinked to transversely directed forces.
Another purpose of this study was to confirmwhether primates conform to current notions as tohow mammalian jaw muscles work during mastica-tion. The firing patterns of the anterior and poste-rior temporalis of anthropoids conform well to thediagonal hypothesis of Herring (1993) and the trip-let hypothesis of Weijs (1994). This is only partiallytrue for strepsirrhines. Although the firing patternsof the posterior temporalis of our two groups ofstrepsirrhines conform to these two hypotheses,data for the anterior temporalis only conform to thediagonal hypothesis. Weijs (1994) hypothesized thatprosimians (plus an array of other “primitive” mam-mals) retain the primitive mammalian condition forthe firing pattern of their anterior temporalis. Thatis, that the working- and balancing-side anteriortemporalis muscles in these mammals fire synchro-nously, only during fast closing, and do not fire dur-ing the power stroke of mastication. Our data dem-onstrate that this is not the case for thick-tailedgalagos and ring-tailed lemurs. Instead, the firingpatterns for their anterior temporalis muscles arevirtually identical to those of anthropoids. That is,they fire asynchronously, reach peak activity duringthe power stroke, and are an important source ofmasticatory force during the power stroke of masti-cation. Moreover, both increased recruitment andlate firing patterns of the b-s posterior temporalissuggest that anthropoids may emphasize the dura-tion and magnitude of the power stroke. Finally,during the late portion of the power stroke, the b-sposterior temporalis assists the b-s deep masseter indriving the working-side molars through the termi-nal portion of occlusion.
ACKNOWLEDGMENTS
This study was supported by research grants fromthe NIH (Merit Award DE04531 to W.L.H.;DE05595 to M.J.R. and W.L.H.; and DE05663 toC.E.W. and W.L.H.) and NSF (SBR-9420764 toWLH and BCS-0138565 to W.L.H., C.E.W., andC.J.V.; and BNS-91-00523 to C.F.R. and W.L.H.).Thanks go to Dr. Don Schmechel for providing usaccess to the galagos in his care, and to the veteri-nary staff at the Duke University Primate Center.This is Duke University Primate Center publication#765.
LITERATURE CITED
Beecher RM. 1977. Function and fusion at the mandibular sym-physis. Am J Phys Anthropol 47:325–336.
Beecher RM. 1979. Functional significance of the mandibularsymphysis. J Morphol 159:117–130.
Cachel SM. 1979. A functional analysis of the primate mastica-tory system and the origin of the anthropoid post-orbital sep-tum. Am J Phys Anthropol 50:1–18
Connolly R, Quimby FW. 1978. Acepromazine-ketamine anesthe-sia in the rhesus monkey (Macaca mulatta). Lab Anim Sci28:72–74.
Crompton AW. 1995. Masticatory function in nonmammaliancynodonts and early mammals. In Thomason JJ, editor. Func-tional morphology in vertebrate paleontology. Cambridge:Cambridge University Press. p 55–75.
Crompton AW, Hylander WL. 1986. Changes in mandibular func-tion following the acquisition of the dentary-squamosal jawarticulation. In: Hotton N, MacLean PD, Roth JJ, editors. Theecology and biology of mammal-like reptiles. Washington, DC:Smithsonian Institution Press. p 263–282.
de Vree F, Gans C. 1976. Mastication in pygmy goats (Cuprahircus). Ann Soc R Zool Belg 105:255–306.
Gorniak GC. 1977. Feeding in golden hamsters, Mesocricetusauratus. J Morphol 154:427–458.
Herring SW. 1976. The dynamics of mastication in pigs. ArchOral Biol 21:473–480.
Herring SW. 1985a. Morphological correlates of masticatory pat-terns in peccaries and pigs. J Mammol 66:603–617.
Herring SW. 1985b. The ontogeny of mammalian mastication.Am Zool 25:339–349.
Herring SW. 1992. Muscles of mastication: Architecture and func-tional organization. In: Davidovitch Z, editor. Biological mech-anisms of tooth movement and craniofacial adaptation. Colum-bus, OH: Ohio State University College of Dentistry. p 541–548.
Herring SW. 1993. Epigenetic and functional influences on skullgrowth. In: Hanken J, Hall BK, editors. The skull, volume 1:development. Chicago: University of Chicago. p 153–206.
Herring SW, Scapino RP. 1973. Physiology of feeding in minia-ture pigs. J Morphol 141:427–460.
Hiiemae KM, Kay RF. 1973. Evolutionary trends in the dynamicsof primate mastication. In: Zingeser MR, editor. Craniofacialbiology of primates. Basel: Karger. p 28–62.
Hogue AS, Ravosa MJ. 2001. Transverse masticatory movements,occlusal orientation, and symphyseal fusion in selenodont ar-tiodactyls. J Morphol 249:221–241.
Hylander WL. 1975. The human mandible: lever or link? Am JPhys Anthropol 43:227–242.
Hylander WL. 1977. In vivo bone strain in the mandible of Galagocrassicaudatus. Am J Phys Anthropol 46:309–326.
Hylander WL. 1979a. Mandibular function in Galago crassicau-datus and Macaca fascicularis: an in vivo approach to stressanalysis of the mandible. J Morphol 159:253–296.
Hylander WL. 1979b. The functional significance of primate man-dibular form. J Morphol 160:223–240.
Hylander WL. 1984. Stress and strain in the mandibular sym-physis of primates: a test of competing hypotheses. Am J PhysAnthropol 64:1–46.
Hylander WL. 1985. Mandibular function and biomechanicalstress and scaling. Am Zool 25:315–330.
Hylander WL. 1986. In-vivo bone strain as an indicator of mas-ticatory bite force in Macaca fascicularis. Arch Oral Biol 31:149–157.
Hylander WL, Johnson KR. 1989. The relationship between mas-seter force and masseter electromyogram during mastication inthe monkey Macaca fascicularis. Arch Oral Biol 34:713–722.
Hylander WL, Johnson KR. 1993. Modelling relative masseterforce from surface electromyograms during mastication in non-human primates. Arch Oral Biol 38:233–240.
Hylander WL, Johnson KR. 1994. Jaw muscle function and wish-boning of the mandible during mastication in macaques andbaboons. Am J Phys Anthropol 94:523–547.
Hylander WL, Johnson KR, Crompton AW. 1987. Loading pat-terns and jaw movements during mastication in Macaca fas-cicularis: a bone-strain, electromyographic, and cineradio-graphic analysis. Am J Phys Anthropol 72:287–314.
Hylander WL, Johnson KR, Crompton AW. 1992. Muscle forcerecruitment and biomechanical modeling: an analysis of mas-
TEMPORALIS EMG 55

seter muscle function in Macaca fascicularis. Am J Phys An-thropol 88:365–387.
Hylander WL, Ravosa MJ, Ross CF, Johnson KR. 1998. Mandib-ular corpus strain in primates: further evidence for a functionallink between symphyseal fusion and jaw-adductor muscle force.Am J Phys Anthropol 107:257–271.
Hylander WL, Ravosa MJ, Ross CF, Wall CE, Johnson KR. 2000.Symphyseal fusion and jaw-adductor muscle force: an EMGstudy. Am J Phys Anthropol 112:469–492.
Hylander WL, Vinyard CJ, Wall CE, Williams SH, Johnson KR.2002. Recruitment and firing patterns of jaw muscles duringmastication in ring-tailed lemurs. Am J Phys. Anthropol[Suppl] 34:88.
Hylander WL, Wall CE, Vinyard C, Ross CF, Ravosa MJ, et al.2004. Jaw adductor force and symphyseal fusion. In: Anapol F,German R, Jablonski N, editors. Shaping primate evolution.Cambridge: Cambridge University Press. p 229–257.
Langenbach GEJ, van Eijden TMGJ. 2001. Mammalian feedingmotor patterns. Am Zool 41:1338–1351.
Lieberman DE, Crompton AW. 2000. Why fuse the mandibularsymphysis? A comparative analysis. Am J Phys Anthropol 112:517–540.
Luschei ES, Goodwin GM. 1974. Patterns of mandibular move-ment and jaw muscle activity during mastication in the mon-key. J Neurophysiol 37:954–966.
Moller E. 1966. The chewing apparatus: an electromyographic studyof the action of the muscles of mastication and its correlation tofacial morphology. Acta Physiol Scand [Suppl] 69:1–229.
Panjabi MM, White AA. 2001. Biomechanics in the musculoskel-etal system. New York: Churchhill Livingstone.
Ravosa MJ, Hylander WL. 1994. Function and fusion of themandibular symphysis in primates: stiffness or strength? In:Fleagle JG, Kay RF, editors. Anthropoid origins: the fossilevidence. New York: Plenum Press. p 447–468.
Ross CF. 1995a. Muscular and osseous anatomy of the primateanterior temporal fossa and the function of the postorbitalseptum. Am J Phys Anthropol 98:275–306.
Ross CF. 1995b. Allometric and functional influences on orbitorientation in primates and the origins of the Anthropoidea. JHum Evol 29:201–227.
Ross CF. 2000. Into the light: the origins of the Anthropoidea.Ann Rev Anthropol 29:147–194.
Ross CF, Hylander WL. 2000. Electromyography of the anteriortemporalis and masseter muscles of owl monkeys (Aotus trivir-gatus) and the function of the postorbital septum. Am J PhysAnthropol 112:455–468.
Scapino RP. 1981. Morphological investigations into functions ofthe jaw symphysis in carnivorans. J Morphol 167:339–375.
Sokal RR, Rohlf FJ. 1981. Biometry. New York: W.H. Freeman.Vinyard CJ, Williams SH, Wall CE, Johnson KR, Hylander WL.
2001. Deep masseter recruitment patterns during chewing incallitricids. Am J Phys Anthropol [Suppl] 32:156.
Vinyard CJ, Ravosa MJ, Wall CE, Williams SH, Johnson KR, etal. 2004a. Jaw-muscle function and the origins of primates. In:Ravosa MJ, Dagosto M, editors. Primate origins and adapta-tions: a multidisciplinary perspective. New York: Kluwer Press(in press).
Vinyard CJ, Williams SH, Wall CE, Johnson KR, Hylander WL.2004b. Jaw-muscle electromyography during chewing in Be-langer’s treeshrews (Tupaia belangeri). Am J Phys Anthropol(accepted for publication).
Weijs WA. 1994. Evolutionary approach of masticatory motorpatterns in mammals. In: Bels M, Chardon VL, Vandewalle P,editors. Biomechanics of feeding in vertebrates. Berlin: Spring-er-Verlag. p 282–320.
Weijs WA, Dantuma R. 1981. Functional anatomy of the masti-catory apparatus in the rabbit (Oryctolagus cuniculus L.). NethJ Zool 31:99–147.
Williams CV, Glenn KM, Levine JF, Horne WA. 2003. Compari-son of the efficacy and cardiorespiratory effects of medetomi-dine-based anesthetic protocols in ring-tailed lemurs (Lemurcatta). J Zoo Wildlife Med 34:163–170.
56 W.L. HYLANDER ET AL.