
Systematics and Biology of Silica Bodies in Monocotyledons
65
Systematics and Biology of Silica Bodies in Monocotyledons Author(s): Christina J. Prychid, Paula J. Rudall and Mary Gregory Reviewed work(s): Source: Botanical Review, Vol. 69, No. 4 (Oct. - Dec., 2003), pp. 377-440 Published by: Springer on behalf of New York Botanical Garden Press Stable URL: http://www.jstor.org/stable/4354467 . Accessed: 06/07/2012 14:10 Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp . JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. . New York Botanical Garden Press and Springer are collaborating with JSTOR to digitize, preserve and extend access to Botanical Review. http://www.jstor.org
-
Upload
yann-paranagua -
Category
Documents
-
view
47 -
download
1
Transcript of Systematics and Biology of Silica Bodies in Monocotyledons

Systematics and Biology of Silica Bodies in MonocotyledonsAuthor(s): Christina J. Prychid, Paula J. Rudall and Mary GregoryReviewed work(s):Source: Botanical Review, Vol. 69, No. 4 (Oct. - Dec., 2003), pp. 377-440Published by: Springer on behalf of New York Botanical Garden PressStable URL: http://www.jstor.org/stable/4354467 .Accessed: 06/07/2012 14:10
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
New York Botanical Garden Press and Springer are collaborating with JSTOR to digitize, preserve and extendaccess to Botanical Review.
http://www.jstor.org

The Botanical Review 69(4): 377-440
Systematics and Biology of Silica Bodies in Monocotyledons
CHRISTINA J. PRYCHID, PAULA J. RUDALL, AND MARY GREGORY
Jodrell Laboratory Royal Botanic Gardens
Kew, Richmond, Surrey TW9 3AB, England
I. Abstract . ................................................... 377 II. Introduction .................................................... 378
III. Historical Review ................................................. 378 IV. Composition of Silica in Plants . ........................................ 380 V. Techniques ................................................. 380
VI. Uptake and Deposition ................................................ 382 VII. Morphology and Location .............................................. 384
VIII. Ecology ................................................. 384 IX. Functions . ................................................... 385 X. Applications ................................................. 386
A. Soils and Archaeology .............................................. 386 B. Agricultural Crops and Their Evolution .......... ...................... 387 C. Medical Studies ................................................. 387 D. Animal Diets ................................................. 387 E. Other Applications ................................................. 388
XI. Systematic Distribution ................................................ 388 A. Orchidaceae ................................................... 390 B. Commelinids ................................................. 398
1. Arecaceae ................................................... 398 2. ZHC Clade (Zingiberales, Commelinales and Hanguana) ..... .......... 401 3. Poales . ................................................... 406 4. Dasypogonaceae ................................................. 425
XII. Literature Cited ................................................. 425
I. Abstract
Many plants take up soluble monosilicic acid from the soil. Some of these plants subse- quently deposit it as cell inclusions of characteristic structure. This article describes the distri- bution and diversity of opaline silica bodies in monocotyledons in a phylogenetic framework, together with a review of techniques used for their examination, and the ecology, function and economic applications of these cell inclusions. There are several different morphological forms of silica in monocot tissues, and the number of silica bodies per cell may also vary. The most common type is the "druse-like" spherical body, of which there is normally a single body per cell, more in some cases. Other forms include the conical type and an amorphous, fragmentary type (silica sand). Silica bodies are most commonly found either in the epidermis (e.g., in
Copies of this issue [69(4)] may be purchased from the NYBG Press, The New York Botanical Garden, Bronx, NY 10458-5128, U.S.A.; nybgpress(nybg.org. Please inquire as to prices.
Issued 07 April 2004 377 (D 2004 The New York Botanical Garden

378 THE BOTANICAL REVIEW
grasses, commelinas and sedges) or in the sheath cells of vascular bundles (e.g., in palms, bananas and orchids). Silica-bearing cells are most commonly associated either with subepi- dermal sclerenchyma or bundle-sheath sclerenchyma. Silica bodies are found only in orchids and commelinids, not in other lilioid or basal monocots. In orchids, silica bodies are entirely absent from subfamilies Vanilloideae and Orchidoideae and most Epidendroideae but present in some Cypripedioideae and in the putatively basal orchid subfamily Apostasioideae. Among commelinid monocots, silica bodies are present in all palms, Dasypogonaceae and Zingiberales but present or absent in different taxa of Poales and Commelinales, with at least four separate losses of silica bodies in Poales.
II. Introduction
Most plants have non-protoplasmic inclusions in some of their cells, such as calcium ox- alate crystals, starch grains, tannins and silica bodies. In some groups the presence of such cell inclusions may represent a potentially significant taxonomic character. For example, calcium oxalate styloids are a characteristic feature of the family Iridaceae (Goldblatt et al., 1984; Rudall, 1994); also crystal druses are largely restricted to some basal monocotyledons: Acorus and Alismatales (Prychid & Rudall, 1999, 2000; Keating, 2003). Silica bodies of various shapes and sizes occur in leaves of several groups of monocotyledons, always in well-defined tissues. This article reviews the presence, form and distribution of silica bodies in one large group of flowering plants, the monocotyledons, in the context of both a historical review and a phyloge- netic framework. Recent systematic analyses of monocotyledons that have considered this char- acter have recorded only presence or absence of silica bodies. However, their form and position, which are not greatly influenced by environmental factors but are clearly genetically controlled, may also have considerable systematic potential.
Silica in the form of bodies or particles deposited within or on cells, as distinct from silica incorporated in cell walls or completely filling hairs and other cells, -occurs in certain groups throughout the plant kingdom, including Selaginella (Bienfait et al., 1985; Le Coq et al., 1991), Equisetum (Laroche, 1968; Kaufman et al., 1971), some ferns (Rolleri et al., 1987) and to a small extent gymnosperm leaves and wood (Jiang & Zhou, 1989; Hodson et al., 1997; Sangster et al., 1997), as well as some angiosperm families. Pipemo (1988) listed pteridophytes, gymno- sperms and angiosperms that contain silica bodies, with notes on their abundance and position. Silica bodies may be found in all plant parts, although less commonly in roots. In wood they are sometimes termed "silica grains" and are characteristically situated in the ray or axial paren- chyma cells of certain genera in about 55 dicot families (see lists in Welle, 1976; Metcalfe & Chalk, 1983; Espinoza de Pemia, 1987). In reproductive parts they may be different in shape from those in vegetative parts, as Pipemo (1989) showed in many tropical angiosperms.
III. Historical Review
The early history of this subject was well covered by Netolitzky (1929), but since his book is not easily available a summary is given here. Davy (1814) was one of the first people to investigate the form of silica in plants, including the epidermis of Triticum, Avena, Arundo and Equisetum. Struve (1835) demonstrated for "tabaschir" in bamboo stems that silicified parts remain intact after ashing but dissolve in caustic potash solution. Criuger (1857) found silica bodies in Cauto bark (Moquilea or Hirtella species, Chrysobalanaceae) and Calamus paren- chyma; he used both Schulze's solution (nitric acid and potassium chloride) and a mixture of sulphuric and chromic acids to isolate the silica, which he thought was always deposited in dead cells. Von Mohl (1861) corroborated Cruiger's observations but disagreed with the

SILICA BODIES IN MONOCOTYLEDONS 379
hypothesis that silica deposition always occurs in dead cells. He found silica grains (reminis- cent of starch grains) and silica filling entire cells in various plants and used hydrofluoric acid to dissolve the silica and leave organic material. To obtain a silica skeleton he heated the mate- rial in Schulze's solution, then heated it in water and transferred it to alcohol before ashing in a platinum crucible. Kuister (1897) tested for silica with phenol, in which silica grains appear reddish or bluish.
Wiesner (1867) distinguished short "silica cells" from long epidermal cells in Zea and Sac- charum but thought they consisted of a wall thickening and silica incrustation; Wieler (1893, 1897) and Grob (1896) realized that the cell lumina are filled with silica. In Saccharum Wieler (1893, 1897) showed that silicification proceeds inward from a silicified wall and eventually fills the entire cell. The tiny cavities that are sometimes present in silica bodies of grasses were thought to be remains of cytoplasm or gas bubbles (Frohnmeyer, 1914; Molisch, 1918).
Mettenius (1864) coined the terms "Deckzellen" (cover cells) or "Stegmata" (from the Greek stegium, a roof or covering) for cells that contain inclusions and lie over sclerenchyma in ferns; these cells enclosed calcium oxalate crystals in Cyatheaceae, silica concretions in Dryopteris and true stegmata in most Trichomanes species. However, most subsequent authors have ap- plied the term "stegmata" only to silica-containing cells. Rosanoff (1871) termed these silica cells "Scheidenzellen" (sheath cells) because they often surround vascular bundles; he de- scribed them in many orchids, palms, bamboos and Marantaceae, where they contain one or sometimes two to three silica bodies per cell; the cells are often in axial files.
Pfitzer (1877) pointed out the positional homology between silica cells (stegmata) in or- chids and crystal-containing cells in similar locations accompanying the vascular bundles in many plants. Possession of either silica or calcium oxalate crystals in cells overlying vascular bundles occurs frequently in plants; e.g., in ferns (crystals and/or silica), palms (silica: Molisch, 1913), Pandanaceae (crystals: Kohl, 1889), Xanthorrhoea (crystals: Rudall & Chase, 1996) and Iridaceae (crystals: Goldblatt et al., 1984). Konstanty (1926) showed that files of crystal cells, or so-called chambered crystal fibers, develop from division of parenchyma cells, not fiber cells, and Netolitzky (1929) thought it likely that stegmata develop in a similar way. Rosanoff (1871, translated from Russian of 1867) studied the development of such cells in the root of Phoenix dactylifera and Syagrus botryophora; he observed that silica is laid down at a very early stage in these cells, while they are still thin walled and contain cytoplasm. He did not find any increase in number of the cells during development, although they are axially shorter than adjacent sclerenchyma and parenchyma cells; he therefore concluded that in these species the silica cells lose the capacity to grow and divide at an early stage.
Stegmata are characterized by a thickened wall adjacent to the underlying sclerenchyma cells, with progressively thinner lateral walls and thin outer walls. The wall next to scleren- chyma is often pitted, the pits corresponding to those of the sclerenchyma cell. Silica bodies in stegmata may be spherical, conical, hat shaped or of intermediate type and may resemble crys- tal druses (e.g., Ravenala, Strelitzia). The surface of the body is rarely completely smooth, often spinulose or nodular or with a crater-like cavity (e.g., Musa). The thin outer walls of stegmata often border large or small intercellular spaces.
The term "stegmata" is not normally applied to epidermal silica cells, even when they lie over sclerenchyma associated with vascular bundles and possess the thickened inner periclinal wall characteristic of stegmata, although Tomlinson (1969: 325) referred to "epidermal stegmata" in Phenakospermum (Strelitziaceae). Epidermal silica cells frequently lie over sclerenchyma fibers, either restricted to those accompanying vascular bundles as caps or girders or above independent fiber strands. However, silica also occurs in intercostal cells, where it is often of a different form from that in costal cells (e.g., many Cyperaceae and Poaceae).

380 THE BOTANICAL REVIEW
IV. Composition of Silica in Plants
Silica is an important component of many mineral soils and is the second most abundant ele- ment in the Earth's crust, after oxygen (Hodson & Evans, 1995). Soluble silica, the raw material of silica body formation, is released into the soil by weathering of silicate minerals such as quartz and feldspar (Piperno, 1988). For example, orthoclase feldspar, a mineral present in soil, is hydrolyzed by the hydrogen and hydroxyl ions of water into the mineral kaolinite, resulting in the release of potassium ions and monosilicic acid (Si(OH)4) into solution. Monosilicic acid is soluble in water giving a concentration of ca. 2mM at 250C, higher concentrations implying that polymeric forms of silica are present (Hodson & Evans, 1995). These latter two components are then taken up by plant roots and transported throughout the plant in xylem sap.
4KA1Si308 + 4H+ + 18H20 -e Si4Al 4O10(OH)8 + 4K+ + 8Si(OH)4 orthoclase feldspar kaolinite monosilicic acid
Minerals vary in their rates of weathering; for example, aluminosilicate clay minerals are more susceptible to weathering than is quartz (Brady, 1990). There are two proposed mechanisms of soluble silica uptake: active transport by metabolic processes or passive, nonselective flow in the transpiration stream (Piperno, 1988). In plant tissues silicic acid becomes highly polymerized, resulting in the deposition of solid, generally amorphous (non-crystalline) silicon dioxide (SiO2.nH2O), either within or external to cells, often referred to as "opal" or "opaline silica" (Piperno, 1988). This process can occur at a very early stage of plant development, in almost any plant tissue. Silica particles (ca. 10 nm in diameter) in mature hairs of the lemma of Phalaris canariensis may be laid down in lines, thereby resembling rods, may form sheet-like arrangements of discrete par- ticles or may be packed into disorganized arrays (Mann et al., 1983a, 1983b). Kaufman et al. (1970) and Lawton (1980), working on Avena sativa and Lolium temulentum respectively, sug- gested that the silica bodies in these species are made up of smaller silica rods that appear to grow from the sides of the cell toward the center. In Oryza sativa, individual silica bodies each consist of about 100,000 silica rods, each rod ca. 2.5 gm long and 0.4 ,um wide (Dayanandan, 1983). The silica particles in each rod have a diameter of 1-2 nm. Similarly, Jones et al. (1966) demonstrated that silica in various taxa is composed of spherical particles up to 100 nm in diameter. On the other hand, Hodson et al. (1984) cautioned that the spherical particles and the impression that the silica had aggregated into rods could all be artifacts due to the sectioning of the material.
The water content of the silica ranges from 4% to 9%. Crystalline silicon phases have been reported (Lanning, 1960; Sterling, 1967; Wilding & Drees, 1974), and it has been shown that the composition of amorphous silica changes into a crystalline form as it ages (Wilding et al., 1977). Silica bodies may also contain significant amounts of nitrogen and carbon, either within the body itself or on its surface, possibly arising from cytoplasmic material, cellulose and/or lignin. Other elements, such as aluminum, chlorine, copper, iron, manganese, phosphorus and titanium, may also be present. Plant silica is optically isotropic, ranging in refractive index from 1.41 to 1.47, has a specific gravity from 1.5 to 2.3 and, with a light microscope, ranges in appearance from colorless or light brown to opaque (Jones & Beavers, 1963). Carbon pigmen- tation may cause the darker forms of silica bodies. Frohnmeyer (1914) noticed that, in young material of Saccharum officinarum, silica bodies are only weakly refractive.
V. Techniques
Several different techniques have been utilized to examine silica in plants, although light microscopy is the primary method, often using standard anatomical methods of wax

SILICA BODIES IN MONOCOTYLEDONS 381
embedding and sectioning or epidermal peels (e.g., Frohnmeyer, 1914; Parry & Smithson, 1958; Tomlinson, 1966; Blackman, 1969; Hodson & Sangster, 1988). Silica may be observed in situ or extracted via various procedures, such as dry ashing under high temperatures (spodogram technique: Parry & Smithson, 1958; Lanning et al., 1980). The ash may be assessed for silicon dioxide content by difference of weights before and after treatment with hydrofluoric acid (Lanning & Eleuterius, 1989). Wet ashing involves extraction by liquids, such as sulphuric acid (Dayanandan, 1983), or digestion of organic material with nitric and perchloric acids (Hayward & Parry, 1980). For example, Blackman (1971) made observations on silica mor- phology in 26 species of grasses using either the spodogram technique or treatment with hydro- gen peroxide and/or chromic acid. Parr et al. (2001) investigated a microwave digestion method for the extraction of phytoliths from herbarium and/or fresh plant material without the need for wet or dry ashing. They found that phytolith assemblages comparable to those of conventional dry ashing were obtained quickly and without cross-contamination.
Localization of unstained and essentially transparent silica bodies using a light microscope relies on differences in refractive indices between the mounting medium and silica (Parry & Smithson, 1958; Dayanandan, 1983). However, there are a number of staining techniques. For example, silica deposits (actually the silanol [SiOH] groups on the surfaces of silica particles), may be localized by staining with silver-amine chromate, methyl red and crystal violet lactone (Dayanandan, 1983). Sections may also be stained with phenol (Davis et al., 1973), any silica present giving a magenta color reaction, and compared before and after treatment with hydro- fluoric acid, which dissolves the silica (Gattuso et al., 1998). Anionic dyes may also be used to visualize silica (Allingham et al., 1958). Total silica content may be determined colorimetri- cally by a chemical process that results in the formation of a blue silica-molybdate complex (Blackman, 1968; Yoshida et al., 1976; Gattuso et al., 1998).
Enzyme localization techniques were used by Blackman (1969) on developing silica cells in the leaf sheath of wheat Triticum aestivum to look for metabolic peculiarities of potential and developing silica cells. In addition to the greater level of succinic dehydrogenase activity found in potential silica cells, the cells may contain substances with a specific activity related to silica accumulation and precipitation or may even be manufacturing enzymes that will eventually degrade the cell contents during silica body formation.
The scanning electron microscope (SEM) and image analysis systems have also been used to evaluate and quantify variations in silica body morphological parameters and to investigate distribution of silica deposits within tissues (Sangster, 1968; Hayward & Parry, 1980; Hodson & Sangster, 1988; Rovner & Russ, 1992; Ball et al., 1993; Whang et al., 1998). Sophisticated statistical programs can pick up significant differences between phytoliths produced in one tissue type and those produced in another (Ball et al., 1993). Silica bodies sampled from a single plant tissue may not necessarily be representative of those produced by the plant as a whole. The SEM has also been used in conjunction with X-ray microanalysis to locate silica and assess silica content (for reviews, see Parry et al., 1984; Harvey, 1986; Hodson & Sangster, 1989a, 1989b; Larcher et al., 1991; Dorweiler & Doebley, 1997; Gattuso et al., 1998). For example, silica was localized in specific wall layers of the stomatal apparatus of sugarcane by Sakai and Thom (1979). The technique relies on the production of a characteristic X-ray wave- length for each element in the sample. If the wavelengths are recorded, they yield a list of the elements present. Thus, images of only the areas producing a characteristic X-ray wavelength are localizations of a particular element within the sample. Similarly, silica has been detected in the leaves of various grasses using two analytical electron-microscopical techniques, Electron- Energy-Loss-Spectroscopy (EELS) and Electron-Spectroscopic-Imaging (ESI) (Bode et al., 1994). Brandenburg et al. (1985) visualized silica bodies in grass leaves using the SEM with

382 THE BOTANICAL REVIEW
backscattered electron imaging; previously this technique had mainly been used by materials scientists and biomedical researchers. Hodson and Sangster (1989a) used this technique to locate silica deposition in the inflorescence bracts of wheat (Triticum aestivum), and Whang et al. (1998) studied the variation of epidermal silica bodies in rice (Oryza). The technique relies upon the intensity of a backscattered electron signal being proportional to the average atomic number of the sample irradiated, the higher the atomic number the brighter the signal. Areas of high silica deposition would appear brighter on the image than areas of low silica deposition. However, if other elements of higher atomic number, such as calcium, phosphorus, potassium, sodium or sulphur, were also present in significant quantities these areas would also appear bright and could thus be confused with the silica signal.
Wavelength dispersive electron probe microanalysis provides electron images of a speci- men and also a corresponding X-ray distribution image for the particular element of interest. Numerous studies have been carried out using this technique, including that of Sangster and Parry (1976), who investigated the occurrence of silica in relation to endodermal thickening and meristematic zones in Sorghum roots. Parry and Hodson (1982) and Parry et al. (1986) mapped silica distribution in the caryopsis and inflorescence bracts of Setaria italica and leaves of Bidens pilosa respectively, in relation to implications of plant silica in esophageal cancer.
Ultrastructural experiments using transmission electron microscopy (TEM) to examine up- take of silicic acid and its subsequent incorporation into cell-wall silica were carried out by Sowers and Thurston (1979). They grew Urtica pilulifera plants, which bear silicified stinging cells on their leaves, in hydroponic solutions with and without supplements of silicic acid to determine whether silicon starvation would affect plant growth. Ultrathin sections of plant tissues were micrographed before and after treatment with hydrofluoric acid for comparison. Similar experiments were undertaken by Chen and Lewin (1969) on Equisetum; they found that successive reductions in available silicon in the growth solution resulted in an increasingly stunted habit. Ultrastructural localization of soluble silicon has been carried out using freeze substitution to preserve and embed specimen material (Ashton & Jones, 1976). This technique reduces the loss of soluble components from the cells. Radioisotopes have been used in several studies of silicon metabolism in higher plants as labels for silicic acid (Rothbuhr & Scott, 1957). Silica bodies or phytoliths that have been released into the soil through plant decay can be radiocarbon-dated by utilizing the carbon trapped within the body. Amorphous silica gel has very high X-ray absorptivities; Agarie et al. (1996) used soft X-ray analysis to determine silica body distribution and relative frequency in whole leaves of Oryza sativa. Dried leaves were irradiated with X rays on soft X-ray film; silica bodies appeared as black spots on the film.
Scanning transmission electron microscopy (STEM) has also been used (Hodson et al., 1984; Hodson & Sangster, 1993) to localize silica. For instance, Hodson and Sangster (1993) used the technique to look at the interaction between silicon and aluminum in the freeze-dried roots of Sorghum bicolor. The higher resolution of the STEM meant that the authors could localize the aluminum/silicon deposit of the root epidermis to the outer tangential wall.
VI. Uptake and Deposition
Lewin and Reismann (1969) and Raven (1983) reviewed the evidence for passive uptake of monosilicic acid in certain plants. A mechanism of this type would allow prediction of the amount of deposited silicon in a plant from the concentration of silicic acid in the growth medium, as Jones and Handreck (1965) inferred in oat plants. However, there is also evidence for active uptake of soluble silica in some species. For example, in rice shoots, silicic acid can pass into xylem sap even against a concentration gradient (Okuda & Takahashi, 1964). Hodson

SILICA BODIES IN MONOCOTYLEDONS 383
and Evans (1995) found that more silica is taken up by wetland grasses than would be predicted from the quantity of silica present in the soil solution and the transpiration rate, thereby imply- ing an active uptake mechanism. Piperno (1988) suggested that both active and passive absorp- tion may occur in different species or even, sometimes, in different regions of the same plant.
Silica accumulation (sensu Takahashi & Miyake, 1977) is characteristic of some plant fami- lies, whereas others produce little or no silica. Species with little silica control the amount of silicic acid that enters the root or passes from the root to the aerial tissues of the plants (Piperno, 1988). Jones and Handreck (1969) proposed a hypothetical barrier in root epidermal cells of clover (Trifolium incarnatum) that restricted the flow of silicic acid into the transpiration stream. Experiments undertaken by Parry and Winslow (1977) on pea seedlings (Pisum sativum) showed that there is some mechanism at the root surface which disallows the passage of monosilicic acid into the root. In other species, such as Viciafabia and Ricinus communis, a layer of fatty substance on the root-hair surface may form the barrier (Parry & Winslow, 1977).
In monocotyledons, relatively few studies on opaline silica bodies have considered devel- opmental aspects, especially using modem methods. Grob (1896), Frohnmeyer (1914) and Prat (1931) examined silica deposition in Poaceae. They found differing types, ranging from a fine-grained silica network, with accumulating silica continuing to fill the cell centripetally (Grob, 1896), to the formation of a "silica ring" around the cell periphery enclosing the cell contents, which later increases, restricting the cell lumen (Frohnmeyer, 1914). Mature silica bodies have characteristic vesicular cavities within them; Grob (1896) related the position of these cavities to the stage of silica deposition. In Saccharum officinarum, silica deposition occurs rapidly, and there is layering or stratification in the peripheral silica (Blackman, 1969). Frohnmeyer (1914) suggested that rapid deposition of silica results in homogeneous silica bodies, whereas slow deposition results in non-homogeneous silica masses. In Arundo donax a thin silica ring is formed around the periphery of the cell, and the cell lumen is then filled either by formation of a fine network or by growth of a silica ring, a form of deposition intermediate between the two previous types. Silica may also be deposited on a dispersed organic matrix within the cell, since broken silica bodies have a porous internal structure (Grob, 1896; Sangster, 1968). Prat (1931) suggested that transparent silica bodies originate from an opaque silica gel stage and that granular inclusions within them are the remains of the nucleus and cytoplasmic contents. In addition to infillings of the cell lumen, silica may also be laid down as deposits within the cell wall or between the cellulose wall and the plasma membrane (cell-wall deposi- tion or membrane silicification: Drum, 1968), or in cortical intercellular spaces (Montgomery & Parry, 1979). Cell-wall incrustations are common in dicotyledons, whereas infilling of the cell lumen occurs more frequently in monocotyledons (Piperno, 1988).
Developing silica cells in the leaf sheath of wheat (Triticum aestivum) have an apparently normal cuticle but differ from surrounding cells in having smaller nucleoli and thinner outer cellulose walls (Blackman, 1969). Thin outer cellulose walls may result in a higher rate of transpiration, facilitating an influx of silica as monosilicic acid. Jones and Handreck (1967), Sangster and Parry (1971) and Raven (1983) all cited transpiration or water loss as a major factor in silica polymerization. In some species, greater amounts of silica are deposited in those regions of the plant where water loss is highest. However, this is not always the case, since silica is often deposited in tissues that restrict water loss, such as sclerenchyma. Indeed, the association between development of silica bodies and sclerenchyma requires further explora- tion, since the two are often associated; for example, in orchids (M0ller & Rasmussen, 1984). Parry et al. (1984) considered that a passive, transpiration-based process cannot account for many cases of localized silica deposition. Such highly localized silica distribution patterns indicate genetic control that is phylogenetically mediated.

384 THE BOTANICAL REVIEW
Nuclei and other cell contents degenerate before silica bodies are formed, resulting in "empty" cells possessing no apparent cell contents or silica bodies. The small size and rapid degenera- tion of nuclei indicate that protein synthesis in the cell is at a minimum (Lowary & Avers, 1965). As the concentration of monosilicic acid in the cell increases, the solution becomes supersaturated, changing to a sol form and then to a highly polymerized gel (SiO2.nH2O). This process could result from a reduction in cellular pH to 5.0-6.0, probably caused by the break- down of the cellular buffering system as the cytoplasm degenerates (Iler, 1955). The polymer- ization and gelling processes may also be catalyzed by the presence of organic molecules. The mechanism of silica deposition is also thought to involve other mineral ions (Perry et al., 1984a, 1984b; Hodson & Bell, 1986). Often the polymerized silicic acid fills the cell lumen and binds to the cellulose cell walls, forming a silico-cellulose membrane; thus cell walls can be silicified (Lewin & Reismann, 1969; Schwarz, 1973). The increasing concentration of silica in the cell results in highly refractive mature silica bodies that typically fill the whole cell (Blackman, 1969).
The genetic control of silica deposition is also under investigation. For example, a single Mendelian locus located on maize chromosome 4, teosinte glume architecture I (tgal), has a major influence on several aspects of cupulate fruit-wall morphology, both in maize (Zea mays ssp. mays) and its wild progenitor, teosinte (Zea mays ssp. parviglumis). Dorweiler and Doebley (1997) demonstrated that the teosinte allele (tgal + teosinte) increases cellular deposition of silica. They suggested that tgal regulates which cells of the glume and rachis epidermis become silicified, since it apparently activates silica deposition in certain cells or represses it in others.
VII. Morphology and Location
There are several morphological forms of silica in monocot tissues, and the number of silica bodies per cell may also vary. Classification of phytoliths is invariably based on their sizes and shapes; for example, Bertoldi de Pomar (1971) proposed two morphological groups: microsilicophytoliths and macrosilicophytoliths. The most common type of silica body in mono- cots is the "druse-like" spherical, spherical-rugose (nodular) or spherical-spinulose type (Fig. 2B), usually a single body per cell but sometimes more. Other forms include the "hat- shaped" type (sometimes called a "truncated conical," Fig. 2A), trough-shaped (Fig. 3C) and an amorphous, fragmentary type (silica sand, Figs. 2E, 2F). These shapes are consistent enough within genera or tribes to be used as characters in taxonomic studies (e.g., Stebbins, 1956; Metcalfe, 1960) and as an aid in identification (e.g., Wilding & Drees, 1968; Dormaar & Lutwick, 1969; Lutwick & Johnston, 1969; Twiss et al., 1969).
Another variable feature is the relative location of silica bodies in plant tissues. They are most commonly found either in the epidermis (e.g., in grasses, commelinas and sedges) or in the sheath cells of vascular bundles (e.g., in palms, bananas and orchids). Silica-bearing cells are most commonly associated with sclerenchyma, either subepidermal sclerenchyma (in the case of epidermal silica) or bundle-sheath sclerenchyma, although there are exceptions to this.
VIII. Ecology
A number of ecological factors, including climate, soil variability, moisture availability and plant age, affect silica body development by regulating the concentrations of dissolved silica that is available to plants (for a review, see Pipemo, 1988). In many species, leaves of older plants contain greater amounts of silica than do their younger counterparts, possibly due in part to cellular changes required to obtain silica and in part to an increased availability of deposition sites (Bezeau et al., 1966; Blackman, 1968, 1969; Lanning & Eleuterius, 1985). In some

SILICA BODIES IN MONOCOTYLEDONS 385
species, an increase in the amount of dissolved silica in a growth medium increases the amount of plant silica (measured as a percentage of dry weight) in direct proportion (Okuda & Takahashi, 1961; Jones & Handreck, 1965). Weathering of silicate minerals is thought to be accelerated by wet climates, thereby liberating greater quantities of soluble silica into the soil than in dry climates (Dunne, 1978), partly explaining the relatively greater concentrations of silica in wet tropical soils (Siever, 1967). Tropical plant root systems are extremely efficient in absorbing any soil nutrients present, resulting in high concentrations of silica in these plants (Lovering, 1959; Riquier, 1960; Pipemo, 1985).
Plants of the same species grown in different soils can contain different amounts of silica. Substances such as iron and aluminum oxides, which may be present in the soil, are known to interact with silica under appropriate conditions, producing colloidal complexes that are not taken up by biological systems (Birchall, 1990; Exley & Birchall, 1992, 1993), thereby reduc- ing the amount of soluble silica available. Indeed, the amount of aluminum and silica that plants transport in their tissues varies considerably, and there is some evidence that very high aluminum accumulation and very high silica accumulation are mutually exclusive (Hodson & Evans, 1995). Exley and Birchall (1993) postulated that silicic acid inhibits the nucleation of aluminum hydrox- ide by forming hydroxyaluminosilicate complexes and that this process was found to increase with increasing pH. Independent of these substances, soluble silica concentration has been shown to reach a maximum at a soil pH of 8-9 (McKeague & Cline, 1963), with the result that more soluble silica is available to plants growing in acid soils. However, silica solubility rises con- siderably above pH 9.0 due to silicate ion formation (Hodson & Evans, 1995).
Increased soil water content may result in increased silica uptake (Jones & Handreck, 1967). For example, grasses grown in the floodwater fields of Egypt had greater silica deposition than did grasses grown in areas where rainfall agriculture was practiced (Miller, 1980). The amount of solid silica in the plant decreases when high concentrations of nitrogen and phosphorus are present in the soil. Soluble silica also increases with an increase in the amount of decomposed organic material in the soil. On the other hand, silica bodies are often present in epiphytic orchids, which do not have access to soluble silica in groundwater but instead obtain it from rainwater. Terrestrial orchid species often lack silica (see section XI, "Systematic Distribution").
Although silica uptake and concentration are affected by environmental factors, they are primarily under genetic control (Pipemo, 1988). Families considered to be non-accumulators do not accumulate silica regardless of the environmental conditions. Plants that show different morphological forms of silica retain their individual silica morphologies when grown under identical environmental conditions.
IX. Functions
Silica is considered to be an important factor for normal growth and development (Agarie et al., 1996). The study of silicon metabolism is beyond the scope of the present review. We are concemed here with deposits of silica, which have sometimes been regarded as a waste product or as a form of storage, from which silicon can be mobilized if needed to interact with other elements; e.g., aluminum (Hodson & Evans, 1995). It has been suggested that epidermal silica bodies may act to reduce transpiration in leaves (Yoshida, 1965; Lanning & Eleuterius, 1983), thereby improving water use efficiency (Yoshida et al., 1959). Hutton and Norrish (1974) found a direct correlation between the silicon percentage in wheat husks and the quantity of water transpired.
The "window" hypothesis postulated that the presence of epidermal silica bodies facilitates the transmission of light through the epidermis to the photosynthetic mesophyll or to stem cortical tissue, consequently increasing photosynthesis and plant growth (e.g., Takeoka et al.,

386 THE BOTANICAL REVIEW
1979). However, Kaufman et al.'s (1979, 1981, 1985) investigations did not support the sug- gestion, and Agarie et al. (1996) disproved this hypothesis in rice.
Silica may help to maintain rigidity in stems and linear leaves (Ishizuka, 1971), although leaf stiffness may also be related to the degree of lignification (Matsuda et al., 1983). Silica has been shown to improve lodging resistance in wheat (Gartner et al., 1984).
Silica content may be related to grazing of grasses by herbivores (Vicari & Bazely, 1993). McNaughton and Tarrants (1983) and McNaughton et al. (1985) found that silica content was higher in heavily grazed grasslands than in others and that more silica was present in plant tissues produced early in the growing season. In a feeding experiment, prairie voles (Microtus ochrogaster) preferentially ate grasses with low silica content (Gali Muhtasib et al., 1992). Wadham and Parry (1981) investigated the relationship between silica content and slug resis- tance.
Djamin and Pathak (1967) tested several rice varieties and found that those with high silica content showed greater resistance to Asiatic stem borer (Chilo suppressalis) than others, prob- ably because the silica interfered with boring and feeding of the larvae; selecting rice varieties with a high silica content was more economical than applying silicate to the soil. Hanifa et al. (1974) studied the role of silica in the resistance of rice to the leaf roller (Cnaphalocrocis medinalis), and Moore (1984) examined the relationship of silica to stem-borer infection by Oscinella species in Lolium.
Silica body content may be correlated with resistance to fungal diseases; for instance, brown spot disease of rice (Nanda & Gangopadhyay, 1984) and blast disease of rice (Suzuki, 1937).
X. Applications
A. SOILS AND ARCHAEOLOGY
Silica bodies and other silicified plant parts, such as silicified hairs and cell walls, occurring in soils, dust, etc., are termed "phytoliths," or sometimes "silicophytoliths," "opal phytoliths" or "plant opal." The term "phytolith" may also be applied to other mineral structures of plant origin, such as calcium oxalate crystals, but is more usually restricted to silica particles. Deflandre (1963) reported that Ehrenberg (1841) was one of the first scientists to recognize that these particles were, at least in part, of plant origin; Ehrenberg studied them in soils and classified them by shape, terming them "phytolitharia." The study of phytoliths in soils and archaeologi- cal material has proved a valuable aid for determining vegetation types in a particular locality at various ages in the past and thus reconstructing palaeoenvironments, as well as for investi- gating ancient agricultural practices and crop-plant evolution. Phytoliths are extremely durable in soil and may not migrate through it to the same extent as do other microfossils, such as pollen and diatoms. However, while phytoliths derived from grasses and sedges may be abun- dant, those from some vegetation types may be underrepresented or absent; for example, gym- nosperms and fems (Horrocks et al., 2000).
This is a vast subject, and we refer the reader to relevant texts for more information (Rovner, 1986; Pipemo, 1988; Mulholland, 1989; Pearsall, 1989; Rapp & Mulholland, 1992; Wang & Lu, 1993; Pinilla et al., 1997). As an example of this work, we can cite Blinnikov (1994), who found different phytolith assemblages in changing alpine plant communities in the northwest- em Caucasus, and Madella (1997), who analyzed the phytolith content of sediments from mod- ern and palaeosols in Tadjikistan and was able to show that at certain periods grasses predominated, whereas at others there were also gymnosperms and dicot trees in the vegeta- tion. Runge and Runge (1997) demonstrated rain-forest degradation and vegetation changes in East Africa by examination of phytoliths in sediments and comparison with those extracted

SILICA BODIES IN MONOCOTYLEDONS 387
from species composing different present-day plant communities. Changes in forest and grass- land vegetation in Arizona were examined by Kems et al. (2001) and Kems (2001) and in Washington State by Blinnikov et al. (2002). The spread of grasslands in the Late Tertiary of Nebraska was investigated by Stromberg (2002). Such studies rely on a comprehensive refer- ence collection of plants from the area of study, as has been provided by Palmer and others for EastAfrican grasses (Stewart, 1965; Palmer & Tucker, 1981, 1983; Palmer et al., 1985; Palmer & Gerberth Jones, 1986,1988), Brown (1984), Mulholland (1989), Fredlund and Tieszen (1994) and Pipemo and Pearsall (1998) for the USA, and Kealhofer and Pipemo (1998) for the South- east Asian flora.
Archaeological studies are very varied, ranging from analysis of the phytoliths or wear associated with prehistoric or recent human (Puech et al., 1983; Lalueza et al., 1996; Cummings & Magennis, 1997; Juan-Tresserras et al., 1997; Gugel et al., 2001) or animal (Baker et al., 1959; Armitage, 1975; Acuna-Mesen & Garcia-Diaz, 1998) teeth and the information they provide about diet to investigations of ancient settlements (e.g., Kajale & Eksambekar, 1997) and basketry (Ollendorf et al., 1988). Rosen (1992) demonstrated that it was possible to distin- guish between cereal straw and husks in archaeological settlements by the use of silica skel- etons of epidermal cells.
The occurrence of palm remains, including phytoliths, at New World archaeological sites provides evidence for palm dispersal by humans (Morcote-Rios & Bemal, 2001). Stegmata in fossil palm stems have been demonstrated by Ancibor (1995).
B. AGRICULTURAL CROPS AND THEIR EVOLUTION
An important field of research deals with changes in agricultural practices over time and with ancient crop plants and their evolution. There is a large body of literature on maize (e.g., Pipemo, 1988; Bush et al., 1989; Russ & Rovner, 1989; Pearsall & Pipemo, 1990; Doolittle & Frederick, 1991; Bozarth, 1993; Pipemo & Pearsall, 1993; Dorweiler & Doebley, 1997; Pipemo & Flannery, 2001), rice (Fujiwara & Sasaki, 1978; Fujiwara et al., 1990; Sato et al., 1990; Udatsu & Fujiwara, 1993; Cailin et al., 1994; Pearsall et al., 1995; Whang et al., 1998; Zhao et al., 1998; Huang & Zhang, 2000) and wheat (Ball et al., 1993). Musa phytoliths (distinguished from Ensete) have been found in refuse pits dated to the first millennium B.C. in Cameroon and provide evidence for early banana cultivation and contacts with Asia, where Musa is native (Mbida Mindzie et al., 2001); edible bananas do not produce pollen or seeds, so in this case phytolith evidence is particularly useful.
C. MEDICAL STUDIES
The identification of drug plants may be aided by the study of phytoliths (Umemoto & Hozumi, 1971a, 1971b; Umemoto et al., 1973). Silica particles in dust from plant fragments in the atmosphere have been found to affect the health of workers (Baker, 1961; Hodson & Sangster, 1988, 1989a), while silicified particles in cereal foods have been implicated in causing esoph- ageal cancer (Bennett & Parry, 1981; Hodson et al., 1982; O'Neill et al., 1982, 1986; Parry & Hodson, 1982; Newman & Mackay, 1983; Sangster et al., 1983). Silica of plant origin was reported in human intestines as long as 150 years ago (Quekett, 1852), and it may be the cause of urinary complaints in cattle (Bezeau et al., 1966) and sheep (Baker et al., 1961).
D. ANIMAL DIETS
Analysis of phytoliths from teeth or in the digestive system or feces has been used to help determine the diet of herbivorous mammals (Baker et al., 1961; Cherouvrier et al., 1975; Chapuis,

388 THE BOTANICAL REVIEW
Acorus Alismatales Petrosaviaceae Dioscoreales Pandanales Liliales
L__ other Asparagales Asparagales 6 ~Orchidaceae I
Arecaceae Dasypogonaceae Zingiberales commelinids Hanguana Commelinales Poales
Fig. 1. Generalized diagram of monocot relationships based on Chase et al. (2000), showing silica distribution.
1980), insects (Gueguen et al., 1975) and snails (Chevalier et al., 2001). It may even be used to study the diet of extinct species such as the moa (Kondo et al., 1994) and American mastodon (Gobetz & Bozarth, 2001). The digestibility of animal foodstuffs may be related to the silica content (Van Soest & Jones, 1968; Harbers et al., 1981; Balasta et al., 1989).
E. OTHER APPLICATIONS
Comparison of phytolith assemblages in soils from different sites may be useful in forensic investigations (Marumo & Yanai, 1986). Silica extracted from plants has been used in the construction of solar cells (Amick, 1982). Rice husks have been utilized in the manufacture of composite board (Cote, 1974) and as a filler to enhance properties of natural rubber vulcanizates (Sae-Oui et al., 2002). The silica extracted from rice husks has been tested for use in high- performance concrete (Chandrasekhar et al., 2002) and as a filler in epoxy resin for embedding material in electronic devices (Suwanprateeb & Hatthapanit, 2002).
XI. Systematic Distribution
Recent analyses of molecular sequence data (e.g., Chase et al., 2000; Stevenson et al., 2000) have identified several well-supported major monocot clades. However, even combined multigene analyses (e.g., Chase et al., 2000) have yielded an unresolved polytomy between six of these clades (perhaps representing a relatively deep, rapid radiation): the four lilioid orders (Asparagales, Liliales, Dioscoreales and Pandanales), the large commelinid clade and the small bigeneric family Petrosaviaceae (Fig. 1). The major clades can be grouped for convenience into: 1) basal monocots (Acorus and Alismatales); 2) a polyphyletic assemblage of five clades of lilioid monocots (Asparagales, Liliales, Dioscoreales, Pandanales and Petrosaviaceae); and 3) a monophyletic group encompassing all the commelinid monocots, including bromeliads, grasses, sedges, rushes, palms and gingers (Fig. 1). Silica bodies are not found in basal mono- cots or lilioid monocots (apparent reports are based on confusion with crystals); with the single exception of the asparagoid Orchidaceae, which are thus the only non-commelinid monocots with silica bodies (Table I).

SILICA BODIES IN MONOCOTYLEDONS 389
Table I Distribution of silica bodies in families of monocotyledons
Order, family Silica bodies
Basal monocots
Acoraceae Absent Alismatales
Araceae, Alismataceae, Aponogetonaceae, Butomaceae, Absent Cymodoceaceae, Hydrocharitaceae, Juncaginaceae, Lilaeaceae, Limnocharitaceae, Najadaceae, Potamogetona- ceae, Scheuchzeriaceae, Tofieldiaceae, Zannichelliaceae, Zosteraceae
Lilioid monocots
Petrosaviaceae Absent Pandanales
Cyclanthaceae, Pandanaceae, Stemonaceae, Triuridaceae, Absent Velloziaceae
Dioscoreales Burmanniaceae (incl. Thismiaceae), Dioscoreaceae (incl. Absent
Stenomeridaceae, Taccaceae, Trichopodaceae), Nartheciaceae Asparagales
Higher asparagoids:. Agapanthaceae, Agavaceae, Alliaceae, Absent Amaryllidaceae, Anemarrhenaceae, Anthericaceae, Aphyl- lanthaceae, Asparagaceae, Behniaceae, Hyacinthaceae, Lax- manniaceae, Ruscaceae (incl. Eriospermaceae, Convallariaceae, Dracaenaceae, Nolinaceae), Themidaceae
Lower asparagoids: Asphodelaceae, Asteliaceae, Blandfordi- Absent aceae, Boryaceae, Doryanthaceae, Hypoxidaceae, Iridaceae, Ixioliriaceae, Lanariaceae, Tecophilaeaceae, Xanthorrhoea- ceae, Xeronemataceae
Orchidaceae Present or absent (Table II) Liliales
Alstroemeriaceae, Campynemataceae, Colchicaceae, Corsiaceae, Absent Liliaceae, Luzuriagaceae, Melanthiaceae, Philesiaceae, Smila- caceae
Commelinid monocots
Arecaceae (Palmae) (c. 190 genera, 2000 spp.) Present (Table III) Dasypogonaceae (4 genera, 8 spp.) Present (Table X) Hanguanaceae (1 genus, 5+ spp.) Present (Table IV)
Commelinales Commelinaceae (incl. Cartonemataceae) (2 subfamilies, 4 tribes, Present or absent (Table IV)
41 genera, 650 spp.) Haemodoraceae (2 subfamilies, 13 genera, 100 spp.) Present or absent (Table IV) Philydraceae (4 genera, 5 spp.) Absent Pontederiaceae (9 genera, 33 spp.) Absent
Poales Anarthriaceae (1 genus, 7 spp.) Absent Bromeliaceae (56 genera, 2600+ spp.) Present (Table VI) Centrolepidaceae (3 genera, 35 spp.) Present or absent (Table VI) Cyperaceae (104 genera, 5000+ spp.) Present (Table IX) Ecdeiocoleaceae (2 genera, 2 spp.) Present (Table VI) Eriocaulaceae (10 genera, 700-1400 spp.) Absent Flagellariaceae (1 genus, 4 spp.) Present (Table VI) Hydatellaceae (2 genera, 10 spp.) Absent Joinvilleaceae (1 genus, 2 spp.) Present (Table VI)

390 THE BOTANICAL REVIEW
Table I, continued
Order, family Silica bodies
Commelinid monocots, continued
Poales, continued Juncaceae (8 genera, 350 spp.) Present or absent (Table VI) Mayacaceae (1 genus, 4-10 spp.) Absent Poaceae (Gramineae) (700+ genera, 10,000+ spp.) Present (Table VI) Rapateaceae (16 genera; 80 spp.) Present (Table VIII) Restionaceae (55 genera; 490 spp.) Present or absent (Table VII) Thumiaceae (1 genus; 3 spp.) Present (Table VI) Typhaceae (incl. Sparganiaceae) (2 genera; 16-30 spp.) Absent Xyridaceae (5 genera; 300 spp.) Absent
Zingiberales Cannaceae (1 genus, 25 spp.) Present (Table V) Costaceae (4 genera, 100 spp.) Present (Table V) Heliconiaceae (1 genus, 200 spp.) Present (Table V) Lowiaceae (1 genus, 10 spp.) Present (Table V) Marantaceae (31 genera, 550 spp.) Present (Table V) Musaceae (2 genera, 40 spp.) Present (Table V) Strelitziaceae (3 genera, 7 spp.) Present (Table V) Zingiberaceae (50 genera, 1300 spp.) Present (Table V)
Recent molecular analyses have found Orchidaceae to be monophyletic and placed in the order Asparagales (sensu APQ 1998), either as the sole sister taxon to all other Asparagales (Fay et al., 2000) or as sister to the earliest-diverging clade of Asparagales (in analyses of rbcL alone: e.g., Chase et al., 1 995a; Rudall et al., 1997), although they are at best weakly supported in these positions. On present phylogenetic evidence, the presence of silica bodies in both commelinids and orchids in contrast to all other monocots must therefore be interpreted as a homoplasy; i.e., de novo development of silica bodies in two closely related taxa, as M0ller and Rasmussen (1984) suggested. This homoplasy may be adaptive, as a result of similar environ- mental constraints, or it could have resulted from an iterative gene mutation at a time of rapid monocot radiation. Silica body structure and distribution within tissues are closely similar in orchids and putatively "basal" commelinids such as palms. Evolutionary events, such as a secondary loss of silica bodies in some groups and their apparent regain in others, presumably reflect underlying differences in cell chemistry.
A. ORCHIDACEAE
The most widely used orchid classification is still that of Dressler (1993). However, even this classification requires some revision following a recent analysis of molecular data from rbcL (Cameron et al., 1999) that produced five major orchid clades, roughly corresponding to the five traditional subfamilies Epidendroideae (including Vandeae, Dendrobieae, Neottieae and other tribes), Orchidoideae (including Diurideae, Cranichideae, Diseae and Spirantheae), Vanilloideae, Cypripedioideae and Apostasioideae. Subfamilies Apostasioideae, Vanilloideae and Cypripedioideae are all relatively poor in numbers of genera and species. Most of the taxonomic diversity in orchids is encompassed by Orchidoideae and especially Epidendroideae. The bigeneric subfamily Apostasioideae (Apostasia and Neuwiedia) is traditionally regarded as the sister group to other orchids (reviewed by Dressler, 1993). Furthermore, all of the most recent molecular and morphological analyses support this basal position (Cameron et al., 1999; Freudenstein & Rasmussen, 1999; Cameron & Chase, 2000; Freudenstein et al., 2000).

SILICA BODIES IN MONOCOTYLEDONS 391
Following Apostasioideae, the next-branching orchid subfamily is either Cypripedioideae or Vanilloideae (cf. Cameron et al., 1999; Pridgeon et al., 2001; Cameron & Chase, 2000; Molvray et al., 2000; K. Cameron, pers. comm., 2001).
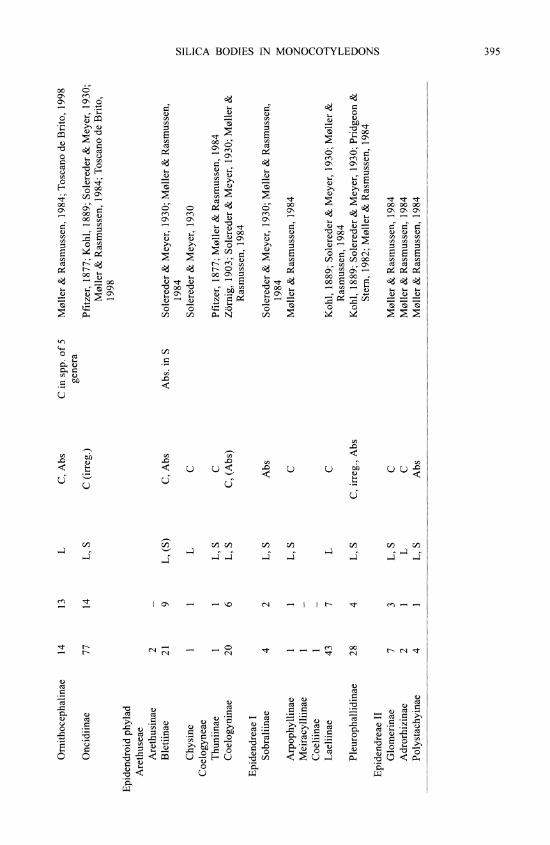
M0ller and Rasmussen (1984) reviewed early work on silica bodies in orchids and added many original observations. In orchids, silica bodies occur in stegmata overlying sclerenchy- matous vascular bundle sheaths, especially adjacent to the phloem, or over independent fiber strands in leaves and stems (including rhizomes and pseudobulbs), absent from the epidermis, although they may be subepidermal. In the leaf of a few Maxillaria species silica cells have been recorded in hypodermal or chlorenchyma cells independent of sclerenchyma (Davies, 1999). In stems the stegmata are on the outer surface of a sclerenchyma ring, in which vascular bundles may be embedded, or in connection with vascular bundles scattered in the ground tissue. The only reports for roots are in three Cymbidium spp. (Cyrtopodiinae) (Yukawa & Stem, 2002), one Maxillaria sp. (Maxillariinae) and a few species of Lycastinae (Holtzmeier et al., 1998; Yukawa & Stem, 2002). Interestingly, Cameron et al. (1999) suggested that tribes Cymbidieae and Maxillarieae may be closely related, based on rbcL sequence data, and Yukawa & Stem (2002) considered that root stegmata might be a synapomorphy in the Cymbidieae- Maxillarieae clade.
Silica bodies are entirely absent from two orchid subfamilies, Vanilloideae and Orchidoideae (Table II). In Cypripedioideae they are either conical, rod-like or absent. Silica bodies are also absent from most tribes or subtribes of Epidendroideae but present in Apostasioideae, which are the putative sister group to all other Orchidaceae. The most parsimonious interpretation is therefore that silica bodies are secondarily lost in groups that lack them entirely. The altemative would require multiple origins of silica bodies within orchids, which seems less plausible. In some (but by no means all) of the subtribes lacking silica bodies, sclerenchyma is also absent (M0ller & Rasmussen, 1984).
Orchid stegmata are small cells that are usually arranged in continuous or discontinuous axial files; in some cases a file may border one axially elongated air canal. There is generally one silica body per cell. Silica bodies are of two types in orchids: 1) conical or hat shaped, sometimes spiny (Fig. 2A), or 2) spherical, often with a rough, warty or spiny surface (Fig. 2B). The conical type is by far the most common in orchids. Silica and crystals frequently occur in the same plant. Spherical silica bodies are restricted to three groups within orchids, one species of Apostasioideae (see below) and two groups of Epidendroideae: 1) subtribes Eriinae, Podochilinae and Dendrobiinae of tribe Podochileae, and 2) all three subtribes of Vandeae. In a few of these groups conical bodies also occur occasionally, but the only group in which M0ller and Rasmussen (1984) recorded both conical and spherical types was Podochilinae, in which Agrostophyllum had conical bodies and the other two genera examined had spherical ones. However, Agrostophyllum has since been transferred to Glomerinae in Dressler's system, in which all the genera have conical silica bodies. Yan Peng Ng (pers. comm., 2000) also reported records of both conical and spherical silica bodies in the large and possibly polyphyl- etic genus Eria. Moller and Rasmussen (1984) and Rasmussen (1986) noted that spherical bodies occur only in epiphytic orchids, whereas conical ones are present in both epiphytic and terrestrial species. They suggested that the conical type and terrestrial state might be ancestral and that spherical bodies might have evolved at least twice. Given our current knowledge of the phylogenetic distribution of the two types in orchids, their assessment is certainly possible, despite the fact that in commelinids the spherical type is apparently the plesiomorphic condi- tion. However, the putatively basal subfamily Apostasioideae is interesting in this context be- cause the single orchid species so far known to possess both conical and spherical bodies is Apostasia wallichii, which has spiny conical bodies in the leaf and spiny spherical bodies in the
(Text continues on p. 397)

Table II Distribution of silica bodies in Orchidaceae (classification of Dressler, 1993).
Key to location: L leaf, R root; S = stem (including rhizome and pseudobulb. Key to type: C = conical; Sph spherical; Abs absent. Brackets indicate few seen or variable.
Location / No. of No. organs
Subfamily, tribe, subtribe genera studied studied Type Notes References
Apostasioideae 2 2 L, S L: C; S: Sph, C, Abs C (L) & Sph (S) in M0ller & Rasmussen, 1984; Stern et al., 1993a I sp. (Stern et al.) H
Cypripedioideae 4 3 L (C), (rod), Abs Solereder & Meyer, 1930; M0ller & Rasmussen, 1984 w
Spiranthoideae 0
Diceratosteleae 1 1 L, S C Stern et al., 1993b z Tropidieae 2 2 L, S C M0ller & Rasmussen, 1984; Stern et al., 1993b Cranichideae
Goodyerinae 35 13 L, S Abs Moller & Rasmussen, 1984; Stern et al., 1993b Prescottiinae 7 2 L, S Abs M01ler & Rasmussen, 1984; Stern et al., 1993b Spiranthinae 41 6 L, S Abs M0ller & Rasmussen, 1984; Stern et al., 1993b Manniellinae 1 1 L Abs Stern et al., 1993b Pachyplectroninae 1 1 L Abs Stern et al., 1993b Cranichidinae 9 2 L Abs Moller & Rasmussen, 1984; Stern et al., 1993b
Orchidoideae Diurideae
Chloraeinae 6 1 L Abs Moller & Rasmussen, 1984 Caladeniinae 10 9 L Abs Pridgeon, 1994 Drakaeinae 5 5 L Abs Moller & Rasmussen, 1984; Pridgeon, 1994 Pterostylidinae 1 1 L Abs Moller & Rasmussen, 1984 Acianthinae 5 1 L Abs Moller & Rasmussen, 1984 Cryptostylidinae 2 1 L, S Abs Stern et al., 1993b

Diuridinae 3 Thelymitrinae 2 1 L Abs Moller & Rasmussen, 1984 Rhizanthellinae 1 -
Prasophyllinae 3 1 L Abs Moller & Rasmussen, 1984 Orchideae
Orchidinae 35 13 L, S, R Abs Kohl, 1889; Solereder & Meyer, 1930; Moller & Rasmussen, 1984; Stern, 1997a
Habenariinae 24 4 L, S, R Abs Moller & Rasmussen, 1984; Stern, 1997b Diseae
Huttonaeinae I I L, S Abs Kurzweil et al., 1995 Satyriinae 3 3 L, S Abs Moller & Rasmussen, 1984; Kurzweil et al., 1995 Coryciinae 5 4 L, S Abs Moller & Rasmussen, 1984; Kurzweil et al., 1995 0 Disinae 5 5 L, S Abs Moller & Rasmussen, 1984; Kurzweil et al., 1995
Epidendroideae Neottieae
Limodorinae 4 2 L C, Abs C in 1 sp. Kohl, 1889; Molisch, 1920; Solereder & Meyer, 1930; Moller & Rasmussen, 1984 z
Listerinae 2 2 L Abs Kohl, 1889; Solereder & Meyer, 1930 Palmorchideae I - 0 Triphoreae 3 - Vanilleae
Galeolinae 4 4 (L), S, R Abs Stern & Judd, 1999, 2000 0 Vanillinae 5 4 L, S, R Abs Kohl, 1889; Solereder & Meyer, 1930; Moller & Z
Rasmussen, 1984; Stern & Judd, 2000 Lecanorchidinae 1 1 S, R Abs Stern & Judd, 2000
Gastrodieae Gastrodiinae 6 2 L, S, R Abs Moller & Rasmussen, 1984; Stern, 1999 Epipogiinae 3 1 S Abs Moller & Rasmussen, 1984 Wullschlaegeliinae 1 1 S, R Abs Moller & Rasmussen, 1984; Stern, 1999
Nervilieae 1 1 L, S Abs Moller & Rasmussen, 1984
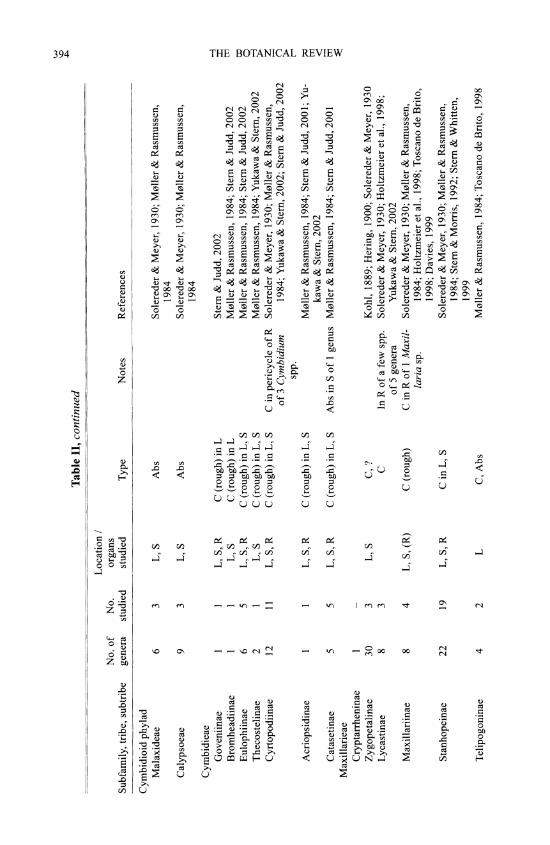
Table I1, continued
Location / No. of No. organs
Subfamily, tribe, subtribe genera studied studied Type Notes References
Cymbidioid phylad Malaxideae 6 3 L, S Abs Solereder & Meyer, 1930; M0ller & Rasmussen,
1984 Calypsoeae 9 3 L, S Abs Solereder & Meyer, 1930; M0ller & Rasmussen,
1984 Cymbidieae
Goveniinae 1 1 L, S, R C (rough) in L Stem & Judd, 2002 Bromheadiinae 1 1 L, S C (rough) in L M0ller & Rasmussen, 1984; Stem & Judd, 2002 f
Eulophiinae 6 5 L, S, R C (rough) in L, S M0ller & Rasmussen, 1984; Stem & Judd, 2002 Thecostelinae 2 1 L, S C (rough) in L, S M0ller & Rasmussen, 1984; Yukawa & Stem, 2002 g
Cyrtopodiinae 12 11 L, S, R C (rough) in L, S C in pericycle of R Solereder & Meyer, 1930; M0oler & Rasmussen, z of 3 Cymbidium 1984; Yukawa & Stem, 2002; Stem & Judd, 2002 ;;
spp. Acriopsidinae 1 1 L, S, R C (rough) in L, S M0ller & Rasmussen, 1984; Stem & Judd, 2001; Yu-
kawa & Stem, 2002 Catasetinae 5 5 L, S, R C (rough) in L, S Abs in S of 1 genus M0oler & Rasmussen, 1984; Stem & Judd, 2001
Maxillarieae Cryptarrheninae 1 -
Zygopetalinae 30 3 L, S C, ? Kohl, 1889; Hering, 1900; Solereder & Meyer, 1930 Lycastinae 8 3 C In R of a few spp. Solereder & Meyer, 1930; Holtzmeier et al., 1998;
of 5 genera Yukawa & Stem, 2002 Maxillariinae 8 4 L, S, (R) C (rough) C in R of 1 Maxil- Solereder & Meyer, 1930; M0ller & Rasmussen,
laria sp. 1984; Holtzmeier et al., 1998; Toscano de Brito, 1998; Davies, 1999
Stanhopeinae 22 19 L, S, R C in L, S Solereder & Meyer, 1930; M0ller & Rasmussen, 1984; Stem & Morris, 1992; Stem & Whitten, 1999
Telipogoninae 4 2 L C, Abs M0ller & Rasmussen, 1984; Toscano de Brito, 1998
Ornithocephalinae 14 13 L C, Abs C in spp. of 5 M0ller & Rasmussen, 1984; Toscano de Brito, 1998 genera
Oncidiinae 77 14 L, S C (irreg.) Pfitzer, 1877; Kohl, 1889; Solereder & Meyer, 1930; M0ller & Rasmussen, 1984; Toscano de Brito, 1998
Epidendroid phylad Arethuseae
Arethusinae 2 -
Bletiinae 21 9 L, (S) C, Abs Abs. in S Solereder & Meyer, 1930; M0ller & Rasmussen, 1984
Chysine 1 1 L C Solereder & Meyer, 1930 Coelogyneae
Thuniinae 1 1 L, S C Pfitzer, 1877; M0ller & Rasmussen, 1984 Coelogyninae 20 6 L, S C, (Abs) Zormig, 1903; Solereder & Meyer, 1930; M0oler& ?
Rasmussen, 1984 Epidendreae I
Sobraliinae 4 2 L, S Abs Solereder & Meyer, 1930; M0ller & Rasmussen, 1984
Arpophyllinae 1 1 L, S C M0ller & Rasmussen, 1984 Meiracylliinae 1 - ? Coeliinae 1 - o Laeliinae 43 7 L C Kohl, 1889; Solereder & Meyer, 1930; M0ller &
Rasmussen, 1984 Pleurophallidinae 28 4 L, S C, irreg., Abs Kohl, 1889; Solereder & Meyer, 1930; Pridgeon& 0
Stem, 1982; Moller & Rasmussen, 1984 Epidendreae II
Glomerinae 7 3 L, S C M0ller & Rasmussen, 1984 Adrorhizinae 2 1 L C M0ller & Rasmussen, 1984 Polystachyinae 4 1 L, S Abs Moller & Rasmussen, 1984
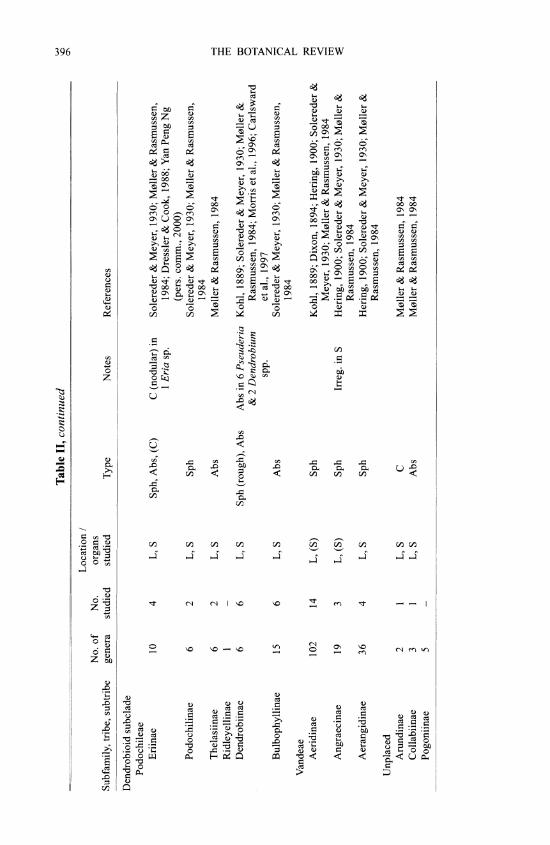
Table II, continued
Location / No. of No. organs
Subfamily, tribe, subtribe genera studied studied Type Notes References
Dendrobioid subclade Podochileae
Eriinae 10 4 L, S Sph, Abs, (C) C (nodular) in Solereder & Meyer, 1930; M0ller & Rasmussen, 1 Eria sp. 1984; Dressler & Cook, 1988; Yan Peng Ng
(pers. comm., 2000) Podochilinae 6 2 L, S Sph Solereder & Meyer, 1930; M0ller & Rasmussen,
1984 Thelasiinae 6 2 L, S Abs M0ller & Rasmussen, 1984 Ridleyellinae 1 - Dendrobiinae 6 6 L, S Sph (rough), Abs Abs in 6 Pseuderia Kohl, 1889; Solereder & Meyer, 1930; M0ller &
& 2 Dendrobium Rasmussen, 1984; Morris et al., 1996; Carlsward ? spp. etal., 1997 z
Bulbophyllinae 15 6 L, S Abs Solereder & Meyer, 1930; M0ller & Rasmussen, 1984 >
Vandeae x Aeridinae 102 14 L, (S) Sph Kohl, 1889; Dixon, 1894; Hering, 1900; Solereder & t1
Meyer, 1930; M0ller & Rasmussen, 1984 Angraecinae 19 3 L, (S) Sph Irreg. in S Hering, 1900; Solereder & Meyer, 1930; M0ller &
Rasmussen, 1984 Aerangidinae 36 4 L, S Sph Hering, 1900; Solereder & Meyer, 1930; M0ller &
Rasmussen, 1984 Unplaced
Arundinae 2 1 L, S C M0ller & Rasmussen, 1984 Collabiinae 3 1 L, S Abs M0ller & Rasmussen, 1984 Pogoniinae 5 -

SILICA BODIES IN MONOCOTYLEDONS 397
Fig. 2. Various silica body morphologies found in Orchidaceae, Arecaceae and the order Comme- linales. A. Cephalanthera pal/ens (Orchidaceae), conical silica bodies with truncated tops (hat shaped) adjacent to phloem cells (bar =10 /,am). B. Angraecum chevalieri (Orchidaceae), spherical bodies overlying sclerenchymatous bundle-sheath cells (bar = 1 0 trm). C. Drymophloeus beguinil (Arecaceae), irregularly spherical bodies in vascular bundle-sheath cells (bar = 1 0 Arm). D. Cyanotis arachnoidea (Commelinaceae), small, spherical, spinulose bodies in epidermnal cells, apparently following the cell wall (bar = 20 ptm). E. Conostylis bract eata (Haemodoraceae), large quantities of silica sand in vascu- lar bundle-sheath cells (bar = 10 gin). F. Anigozanthos flavida (Haemodoraceae) epidermal cells con- taining silica sand (bar = 10 grn).
stem (Judd et al., 1993; Stem et al., 1 993a). Since their basal position makes apostasioids critical in evolutionary assessments of orchid structures, this record of both types of silica body in a single species (A. wallichii) indicates that both types originally occurred in the family and that either both types or the spherical type alone were later lost, with a subsequent regain of the spherical type in two epidendroid groups. However, independent origin of both types in Apostasia is equally plausible.

398 THE BOTANICAL REVIEW
B. COMMELINIDS
The commelinids (or commelinoids) have been identified as a monophyletic clade in sev- eral successive molecular and combined morphological/molecular analyses (e.g., Chase et al., 1995a, 1995b, 2000; Stevenson et al., 2000). Some highly consistent non-molecular (anatomi- cal) characters support the commelinid clade: 1) the presence of cell-wall ferulates, almost entirely absent from non-commelinid monocotyledons (Harris & Hartley, 1980; Rudall & Caddick, 1994); 2) the presence of silica bodies in many commelinid taxa, absent from most non-commelinid monocotyledons except orchids; 3) the presence of Strelitzia-type surface waxes (long, often curly, extruded wax ribbons) in some commelinids, virtually absent from non- commelinids (Barthlott & Frolich, 1983; Frolich & Barthlott, 1988); and 4) stomatal develop- ment by non-oblique cell division, as opposed to oblique division in most non-commelinids (Tomlinson, 1974; Rudall, 2000).
Several taxa formerly had controversial systematic placement in monocotyledons. Pan- danaceae, Cyclanthaceae and Velloziaceae were previously considered commelinid (for ex- ample, by Dahlgren et al., 1985), but recent molecular systematic studies (e.g., Chase et al., 1995b, 2000) have conclusively demonstrated that they are more closely related to non- commelinid groups. Apparent reports of silica bodies are based on confusion with crystals; e.g., Lim and Stone (1971) for Freycinetia. They all lack silica bodies (Table I), which is consistent with a non-commelinid placement (although, admittedly, absence of silica bodies is non-informative for systematics in this context, since it is the plesiomorphic condition for this character within monocotyledons). On the other hand, the presence of silica bodies in Dasypo- gonaceae (Rudall & Chase, 1996), Hanguanaceae (Solereder & Meyer, 1929; Smithson, 1956; Tomlinson, 1969; Tillich & Sill, 1999) and Haemodoraceae (Prychid et al., 2003) supports their inclusion in the commelinid clade, together with other morphological and molecular data (see below).
Although Dasypogonaceae remain unplaced among the commelinids, three other major commelinid clades have been identified (e.g., Chase et al., 2000; Stevenson et al., 2000): 1) Arecales (Arecaceae); 2) a broadly circumscribed Poales; and 3) a "ZHC" clade consisting of Zingiberales, Commelinales and Hanguana. However, phylogenetic relationships between these clades remain unresolved, pending further analyses. A stable phylogeny is critical in evolutionary assessments of characters such as presence of silica bodies. For example, if Arecaceae are sister to other commelinids, as tentatively indicated by some analyses (e.g., Givnish et al., 1999; Chase et al., 2000), it would seem likely that the presence of silica bodies is a synapomorphy for the commelinid clade and that its absence from some taxa (Table III) represents one or more secondary reversals.
1. Arecaceae
The earliest work on silica in palms was mentioned above (see section III, "Historical Re- view"). The first comprehensive anatomical study of palms was that of Tomlinson (1961b), who described the types and distribution of silica bodies in all the major taxonomic groups. Silica bodies occur in stegmata in continuous or discontinuous longitudinal files adjacent to fibers sheathing vascular bundles or independent fiber strands. They are most frequent in leaves and stems but are also seen next to cortical sclerenchymatous strands in roots. They do not occur in the epidermis, but hypodermal stegmata may occasionally intrude between epidermal cells and therefore wrongly appear to be epidermal (e.g., in Borassus: Eberwein, 1903).
There are two types of silica body and silica cell in palms, in both cases with a single silica body per cell: 1) conical or hat-shaped bodies with a smooth base, a rough or spiny surface and

Table III Distribution of silica bodies in leaf of Arecaceae (classification of Uhl & Dransfield, 1987).
Key: cont. = continuous; discont. = discontinuous; Hat = hat shaped; incl. = including; occ. = occasionally; Sph = spherical; vb(s) = vascular bundles(s). Brackets indicate rare or in few taxa.
No. of No. Subfamily, tribe genera studied Location of silica in leaf Type Notes References
Coryphoideae Corypheae 31 20 Over fibers, vb or not + Sph, spiny In cont. or discont. files, Solla, 1884 (fruit); Pfister, 1892;
(incl. transverse vbs) basal wall thick; also Tomlinson, 1961b; Ghose & stem, fruit Johri, 1987; Killmann & Hong,
1992 (stem) Phoeniceae 1 1 Over fibers, vb or not + Sph In cont. files, basal wall Rosanoff, 1871 (root); Tomlinson, >
thick; also stem, root 1961b; Ginieis, 1964; Ghose & Johri, 1987 O
Borasseae 7 6 Over fibers, vb or not Sph, spiny (+ Hat In ? cont. files, basal Tomlinson, 196 ib; Ghose & Johri, (incl. transverse vbs) in Lodoicea) wall thick; also stem, 1987 '/
root
Calamoideae Calameae 19 13 Over fibers, vb or not + Sph, spiny In cont. or discont. files, Tomlinson, 196 lb; Ghose & Johri,
(occ. transverse vbs) basal wall thick; also 1987; Weiner & Liese, 1990 0 stem (absent from (stem); Weiner, 1992 (stem); o root) Schmitt et al., 1995
Lepidocaryeae 3 2 Over fibers, vb or not + Sph, spiny In discont. files (basal Tomlinson, 196 ib; Killmann & (incl. transverse vbs wall thick in 1 ge- Hong, 1992 (stem) in 1 genus) nus); also stem 0
Nypoideae 1 1 Over fibers, vb or not Hat In discont. files, basal Tomlinson, 1961b cI
wall scarcely thick- ened
Ceroxyloideae Cyclospatheae 1 1 Over fibers, vb or not Sph, large In ? cont. files, basal Tomlinson, 1961b
wall ? unthickened Ceroxyleae 5 1 Over fibers, vb + Sph In cont. or discont. files, Tomlinson, 1961b
basal wall thick Hypophorbeae 5 4 Over fibers, mainly vb Hat (+ sand in In cont. files, basal wall Tomlinson, 1961b; Ginieis, 1964
mesophyll of scarcely thickened; (perianth) 1 genus) also stem in 1 genus

Table III, continued
No. of No. Subfamily, tribe genera studied Location of silica in leaf Type Notes References
Arecoideae Caryoteae 3 3 Over fibers, vb or not Hat In discont. files, basal Tomlinson, 1961b; Ghose & Johri,
wall scarcely thick- 1987; Killmann & Hong, 1992 ened; also stem, root (stem)
Triarteae 6 4 Over fibers, vb (not Hat In cont. files, basal wall Tomlinson, 1961b transverse vbs) scarcely thickened
Podococceae 1 0 No data Areceae, except 86 35 Over fibers, vb or not ? Sph or ellipti- In cont. or discont. files, Tomlinson, 1961b; Ghose & Johri,
2 subtribes: (not transverse vbs) cal (rarely basal wall thick; also 1987; Killmann & Hong, 1992 (also mesophyll in 1 sand in hypo- stem (stem) genus, epidermis in 1 dermis) genus)
Oraniinae 2 1 Over fibers, vb or not + Hat to elliptical In discont. files, basal Tomlinson, 1961b wall slightly thick- ened
Sclerosper- 2 1 Over fibers, vb or not Hat In discont. files, basal Tomlinson, 1961b matinae wall ? unthickened
Cocoeae, except 22 8 Over fibers, vb or not + Sph (+ sand in Most in discont. files, ba- Rosanoff, 1871 (root); Molisch, 1913 1 subtribe: (incl. transverse vbs) 1 genus in hy- sal wall thick; also (fruit); Tomlinson, 1961b; Ghose
podermis) stem, root, fruit & Johri, 1987; Killmann & Hong, 1992 (stem)
Bactridinae 6 5 Over fibers, vb or not (& Hat In cont. or discont. files, Tomlinson, 1961b over sclereids in 1 basal wall scarcely genus) thickened; also stem,
root Geonomeae 6 5 Over fibers, vb or not + Sph (rarely el- In cont. or discont. files, Tomlinson, 1961b
liptical) basal wall mostly thick
Phytelephantoideae 3 1 Over fibers, vb or not Sph In cont. files, basal wall Molisch, 1913 (fruit); Tomlinson, thick; also stem, fruit 1961b

SILICA BODIES IN MONOCOTYLEDONS 401
a basal cell wall that is only slightly thickened and not conspicuously pitted; and 2) spherical bodies, usually rather irregular, sometimes more or less ellipsoidal, also with a rough or spiny surface (Fig. 2C), with a basal cell wall that is thickened, often pitted, and sometimes lignified or suberized and with other walls that are thin. The development of the spherical type has been studied by Schmitt et al. (1995) in Calamus auxillaris. These two types have not been found to occur together except in Lodoicea, where most bodies are spherical but where large, hat-shaped ones up to 23 ,um in diameter are present adjacent to transverse veins in the lamina (Tomlinson, 1961b). Raphide crystals are also common in all parts of palms.
Tomlinson (196 lb) noted that conical or hat-shaped silica bodies are present in the informal groups, which he designated as "bactroid," "caryotoid," "chamaedoroid," "iriartoid" and "nypoid" palms, while spherical bodies occur in the arecoid, borassoid, cocoid, lepidocaryoid, phoenicoid, phytelephantoid and sabaloid palms and in some isolated genera. In Table III his data have been rearranged according to the classification of Uhl and Dransfield (1987). Each subtribe usually possesses only one type of silica body, apart from Lodoicea of Borasseae. Subfamilies Coryphoideae, Calamoideae and Phytelephantoideae all have spherical silica bod- ies, and Nypoideae have hat-shaped silica bodies. Ceroxyloideae comprise two tribes with spherical and one with hat-shaped bodies, while Arecoideae have some tribes or subtribes with spherical and others with hat-shaped bodies.
2. ZHC Clade (Zingiberales, Commelinales, and Hanguana)
This clade is polymorphic for presence or absence of silica bodies. In the taxonomically isolated genus Hanguana (Hanguanaceae), silica bodies are present as irregular granular de- posits, mainly in or near the foliar bundle-sheath cells rather than the epidermis (Solereder & Meyer, 1929; Smithson, 1956; Tomlinson, 1969; Rudall et al., 1999; Tillich & Sill, 1999). Hanguana belongs in a clade together with the orders Zingiberales and Commelinales, but its precise relationships within this clade remain disputed. Morphological data indicate an affinity with Zingiberales (e.g., Rudall et al., 1999), whereas most recent analyses of molecular data place it within Commelinales (e.g., Chase et al., 2000; APG II, 2003).
Apart from Hanguana, there are four families in the order Commelinales: Haemodoraceae, Commelinaceae, Philydraceae and Pontederiaceae (Table IV). Silica bodies are absent from Philydraceae and Pontederiaceae and present in only some genera of Commelinaceae and Haemodoraceae. Evans et al. (2000) did not include presence or absence of silica bodies in their morphological cladistic analysis of Commelinaceae. In Commelinaceae, silica bodies had been thought to be restricted to the tribe Tradescantieae of the subfamily Commelinoideae (Tomlinson, 1969), where they represented a relatively consistent potential synapomorphy for this group, although silica bodies are absent from some genera that were embedded within Tradescantieae in the morphological analysis, such as Murdannia (Faden & Inman, 1996). However, Faden (pers. comm., 2003) has also observed silica bodies in Dictyospermum, a genus of the tribe Commelineae, which appear as large, silica infillings of the cell. These bod- ies differ greatly from the epidermal (several per cell) small, spherical spinulose bodies of the Tradescantieae that are often embedded in the outer cell wall (Fig. 2D). One exception to these spherical bodies is a possible observation of epidermal silica sand in Zebrina pendula. This mirrors observations in Haemodoraceae, where the silica almost always takes the form of silica sand, the only exception to this being rare sightings of spherical bodies, possibly formed by sand coalescence, in two genera that also possess sand. Presence of silica sand is restricted to the subfamily Conostylidoideae (Prychid et al., 2003) and is mainly located in the vascular bundle-sheath cells (Fig. 2E), resembling the case in Hanguana. Only one genus (Anigozanthos) has exclusively epidermal silica sand (Fig. 2F).
(Text continues on p. 406)
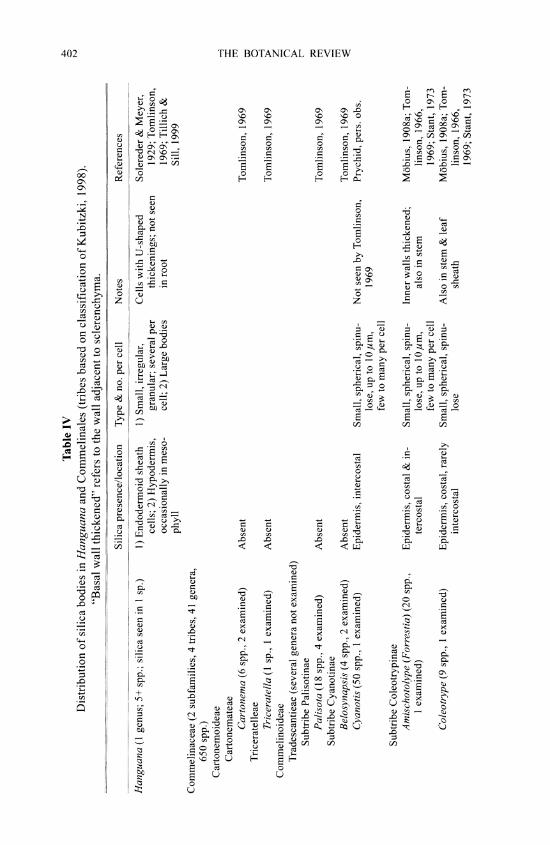
Table IV Distribution of silica bodies in Hanguana and Commelinales (tribes based on classification of Kubitzki, 1998).
"Basal wall thickened" refers to the wall adjacent to sclerenchyma.
Silica presence/location Type & no. per cell Notes References
Hanguana (1 genus; 5+ spp.; silica seen in 1 sp.) 1) Endodermoid sheath 1) Small, irregular, Cells with U-shaped Solereder & Meyer, cells; 2) Hypodermis, granular; several per thickenings; not seen 1929; Tomlinson, occasionally in meso- cell; 2) Large bodies in root 1969; Tillich & phyll Sill, 1999
Commelinaceae (2 subfamilies, 4 tribes, 41 genera, 650 spp.)
Cartonemoideae H
Cartonemateae Cartonema (6 spp., 2 examined) Absent Tomlinson, 1969 w
Triceratelleae ? Triceratella (1 sp., 1 examined) Absent Tomlinson, 1969 >
Commelinoideae Tradescantieae (several genera not examined) >
Subtribe Palisotinae Palisota (18 spp., 4 examined) Absent Tomlinson, 1969 m
Subtribe Cyanotinae Belosynapsis (4 spp., 2 examined) Absent Tomlinson, 1969 Cyanotis (50 spp., 1 examined) Epidermis, intercostal Small, spherical, spinu- Not seen by Tomlinson, Prychid, pers. obs.
lose, up to lOum, 1969 few to many per cell
Subtribe Coleotrypinae Amischotolype (Forrestia) (20 spp., Epidermis, costal & in- Small, spherical, spinu- Inner walls thickened; Mobius, 1908a; Tom-
1 examined) tercostal lose, up to 1 Oum, also in stem linson, 1966, few to many per cell 1969; Stant, 1973
Coleotrype (9 spp., 1 examined) Epidermis, costal, rarely Small, spherical, spinu- Also in stem & leaf Mobius, 1908a; Tom- intercostal lose sheath linson, 1966,
1969; Stant, 1973
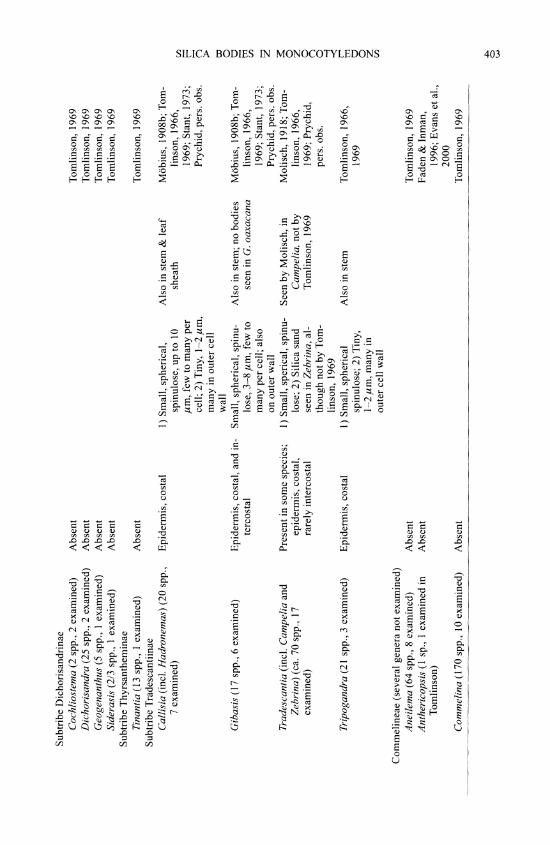
Subtribe Dichorisandrinae Cochliostema (2 spp., 2 examined) Absent Tomlinson, 1969 Dichorisandra (25 spp., 2 examined) Absent Tomlinson, 1969 Geogenanthus (5 spp., 1 examined) Absent Tomlinson, 1969 Siderasis (2/3 spp., 1 examined) Absent Tomlinson, 1969
Subtribe Thyrsantheminae Tinantia (13 spp., 1 examined) Absent Tomlinson, 1969
Subtribe Tradescantiinae Callisia (incl. Hadronemas) (20 spp., Epidermis, costal 1) Small, spherical, Also in stem & leaf Mobius, 1908b; Tom-
7 examined) spinulose, up to 10 sheath linson, 1966, ,um, few to many per 1969; Stant, 1973; r cell; 2) Tiny, 1-2 ,m, Prychid, pers. obs. r many in outer cell wall
Gibasis (17 spp., 6 examined) Epidermis, costal, and in- Small, spherical, spinu- Also in stem; no bodies Mobius, 1908b; Tom- U tercostal lose, 3-8 ,m, few to seen in G. oaxacana linson, 1966,
many per cell; also 1969; Stant, 1973; on outer wall Prychid, pers. obs.
Tradescantia (incl. Campelia and Present in some species; 1) Small, sperical, spinu- Seen by Molisch, in Molisch, 1918; Tom- < Zebrina) (ca. 70 spp., 17 epidermis, costal, lose; 2) Silica sand Campelia, not by linson, 1966, z examined) rarely intercostal seen in Zebrina, al- Tomlinson, 1969 1969; Prychid, 0
though not by Tom- pers. obs. 0 linson, 1969
Tripogandra (21 spp., 3 examined) Epidermis, costal 1) Small, spherical Also in stem Tomlinson, 1966, spinulose; 2) Tiny, 1969 0 1-2,um, many in z outer cell wall
Commelineae (several genera not examined) Aneilema (64 spp., 8 examined) Absent Tomlinson, 1969 Anthericopsis (1 sp., 1 examined in Absent Faden & Inman,
Tomlinson) 1996; Evans et al., 2000
Commelina (170 spp., 1O examined) Absent Tomlinson, 1969

Table IV, continued
Silica presence/location Type & no. per cell Notes References
Commelinaceae, continued Commelinoideae, continued
Commelineae, continued Dictyospermum (5 spp., 1 examined) Large, silica infillings of Faden, pers.com.
the cell Floscopa (20 spp., 2 examined) Absent Tomlinson, 1969 Murdannia (50 spp., 5 examined) Absent Tomlinson, 1969 Pollia (17 spp., 5 examined) Absent Tomlinson, 1969 Polyspatha (3 spp., 2 examined) Absent Tomlinson, 1969 Stanfleldiella (4 spp., 1 examined) Absent Tomlinson, 1969
Haemodoraceae (2 subfamilies, 14 genera, ca. 81 spp.) 0
Conostylidoideae Anigozanthos (ca. 10 spp., 7 exam- Epidermis Sand Sand more numerous in Prychid et al., 2003 z
ined) epidermal cells of bifacial leaf sheath; none seen in A. hu- milis
Blancoa (I sp., I examined) 1) Bundle-sheath cells in Sand leaf blade; 2) Adaxial epidermal cells of leaf sheath
Conostylis (ca. 25 spp., 14 examined) 1) Bundle sheath cells; Sand Occasionally, in C. beali- Prychid et al., 2003 2) Palisade meso- ana, vascular bundle- phyll cells, in some sheath sand appears spp.; 3) Spongy pa- to form a spherical renchyma cells, in body some spp.; 4) Rarely, unlignified adaxial epidermal cells of leaf sheaths
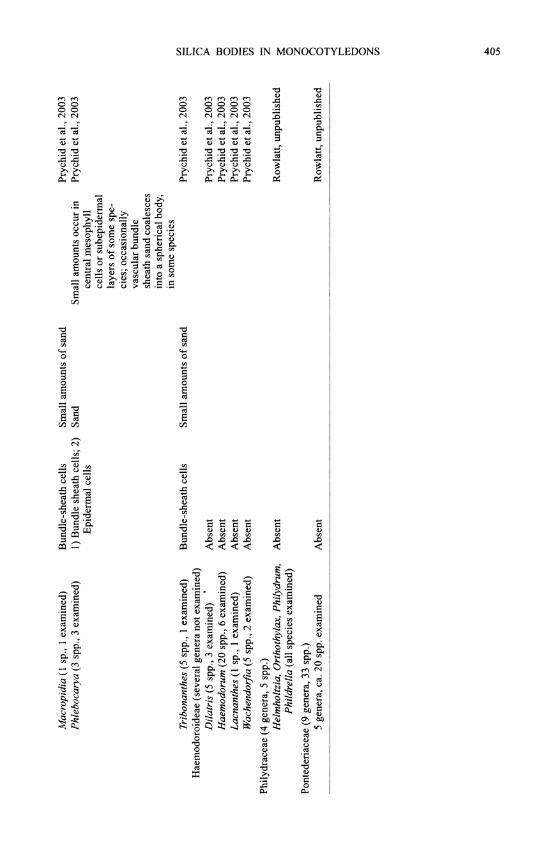
Macropidia (1 sp., I examined) Bundle-sheath cells Small amounts of sand Prychid et al., 2003 Phlebocarya (3 spp., 3 examined) 1) Bundle sheath cells; 2) Sand Small amounts occur in Prychid et al., 2003
Epidermal cells central mesophyll cells or subepidermal layers of some spe- cies; occasionally vascular bundle sheath sand coalesces into a spherical body, in some species
Tribonanthes (5 spp., 1 examined) Bundle-sheath cells Small amounts of sand Prychid et al., 2003 Haemodoroideae (several genera not examined)
Dilatris (5 spp., 3 examined) Absent Prychid et al., 2003 >
Haemodorum (20 spp., 6 examined) Absent Prychid et al., 2003 Lacnanthes (I sp., I examined) Absent Prychid et al., 2003 U
Wachendorfia (5 spp., 2 examined) Absent Prychid et al., 2003
Philydraceae (4 genera, 5 spp.) Helmholtzia, Orthothylax, Philydrum, Absent Rowlatt, unpublished g
Phildrella (all species examined) 0 z Pontederiaceae (9 genera, 33 spp.)
5 genera, ca. 20 spp. examined Absent Rowlatt, unpublished o
0

406 THE BOTANICAL REVIEW
Within the order Zingiberales, silica bodies are present in all families (Table V) (Tomlinson, 1969), mostly restricted to the vascular bundle-sheath cells (in Cannaceae (Fig. 3A), Costaceae (Fig. 3B), Heliconiaceae (Figs. 3C, 3D), Lowiaceae (Fig. 3E), Marantaceae (Figs. 3F, 3G), Musaceae (Figs. 4A, 4B), and some genera of Strelitziaceae (Figs. 4C, 4D), often in a hypoder- mal region adjacent to bundle-sheath sclerenchyma. There are occasional records of silica bod- ies in other mesophyll cells (Heliconiaceae, Marantaceae, Zingiberaceae), and they are also present in the epidermis in Phenakospermum (Strelitziaceae) and most Zingiberaceae (Figs. 4F, 4G) (Tomlinson, 1969). Silica bodies in Zingiberales are druse-like (i.e., spherical with a rugose surface) in most genera, but with some exceptions: they are more or less conical in Orchidantha (Lowiaceae) (Fig. 3E) and some Marantaceae, "trough shaped" (i.e., rectangular with a central shallow depression) in most Heliconiaceae and Musaceae and present as epider- mal silica sand in some Zingiberaceae, rarely in Marantaceae, Strelitziaceae and Musaceae (Fig. 3H). Kress et al. (2001) discussed evolutionary relationships of Zingiberales families using molecular and morphological data. They concluded that Zingiberales are monophyletic, with three sets of sister families: Strelitziaceae + Lowiaceae, Costaceae + Zingiberaceae and Cannanceae + Marantaceae. Of these, Costaceae and Marantaceae have broadly similar silica body types and locations, Lowiaceae and Strelitziaceae have bodies in similar locations but different shapes, Costaceae and Zingiberaceae differ in that Costaceae have bodies over scleren- chymatous bundle sheaths, while Zingiberaceae have predominantly sand or small bodies in epidermal cells.
3. Poales
A broad circumscription of the order Poales is now widely adopted (e.g., APG, 1998; Chase et al., 2000; APG II, 2003). There are two well-defined major clades, the sedge clade (Cyperaceae, Juncaceae, Thumiaceae and Prioniaceae) and the grass clade (Poaceae, Restionaceae and their allies), plus several other families (Bromeliaceae, Hydatellaceae, Mayacaceae, Rapateaceae, Sparganiaceae, Typhaceae and Xyridaceae) whose relationships remain unresolved. Silica bodies are absent from several families of Poales (Table VI), and this may represent a synapomorphy for some groups. In particular, the absence of silica bodies in Hydatellaceae may support a putative relationship between them and Xyridaceae, Eriocaulaceae and Mayacaceae (Michelangeli et al., 2003), all of which lack silica bodies. In contrast to many other commelinids (especially Zingiberales and palms), in most Poales that possess silica bodies they are restricted to the epidermis, with the exception of some species of Bromeliaceae, Ecdeiocoleaceae, Flagellariaceae, Joinvilleaceae, Juncaceae, Restionaceae (Table VII) and Thumiaceae, in which they occur in other tissues, especially the vascular bundle sheath. In Rapateaceae, all genera possess characteristic spherical silica bodies in the epidermis (Table VIII) (Carlquist, 1966; Carlquist in Tomlinson, 1969), rather similar to those of Commelinaceae-Tradescantieae, al- though this must be regarded as a homoplasy, since the two groups belong in different clades within the commelinids.
In the sedge clade, silica bodies are common in most groups of Cyperaceae (Table IX), in which they possess a conical form, often with small spines (satellites) around the base (Figs. 5A, 5B), but rare or absent in some genera of the sedge tribe Hypolytreae, which is the putative sister group to the rest of the family (Muasya et al., 1998, 2000). The distribution and types of silica bodies in leaves of Cyperaceae have been described by Metcalfe (1971), who summa- rized early work on the family; subsequent research has extended the observations but not altered the main conclusions. The most characteristic type of silica body is conical, with the base resting on the usually thickened, inner periclinal wall of an epidermal cell (rarely on anticlinal or outer periclinal walls). There may be one to numerous bodies per cell on a single
(Text continues on p. 422)

Table V Distribution of silica bodies in Zingiberales (classification from Kubitzki, 1998).
Key: LS = longitudinal section; par = parenchyma; scl = sclerenchyma; vb(s) = vascular bundle(s). Basal wall refers to wall adjacent to sclerenchyma. v:
Silica presence / location in leaf Type & no. per cell Notes References
Cannaceae (Igenus, 10-25 spp.) 0 Canna (4 spp. examined) Over vb scl sheath; hypodermis Druse-like, 1 per cell, in In petiole, stem, rhizome, Tomlinson, 1961 a,
thin-walled cells, in files not root 1969 in LS
Costaceae (4 genera, 100 spp.) Costus (3 spp. examined); Over vb scl sheath Druse-like to irregularly In petiole, stem, rhizome, Tomlinson, 1956, 0
Tapeinochilus (1 sp. exam- spiny, 1 per cell; cells not root 1962, 1969 Z ined) often forming complete 0
layer around vbs
Heliconiaceae (1 genus, 200 spp.) Heliconia (14 spp. examined) 1) Over vb sheath fibers & over par 1) Trough shaped; spines Basal wall thickened; cells Tomlinson, 1959,
sheath; 2) Palisade of 1 sp. projecting from base into in files in LS. Also in 1969; Prychid, 0 pits in wall; 1 per cell; 2) stem & rhizome, not in pers. obs. irregularly spherical, 1 root per cell
Lowiaceae (1 genus, 10 spp.) Orchidantha (3-4 spp. exam- Over vb fibers + Conical; top may be trun- Basal wall thickened; cells Tomlinson, 1959,
ined) cated, I per cell; small in files in LS; also in 1969 fingers of silica may petiole, rhizome, not root project from the base of the body into the cell wall

Table V, continued 00 00
Silica presence / location in leaf Type & no. per cell Notes References
Marantaceae (31 genera, 550 spp.; no currently accepted subfamilies or tribes) 17 genera, 37 spp. examined 1) Over vb scl sheath (rarely hypo- 1) Hat shaped, base flat; 1) Basal wall thickened; Petersen, 1893; Tom-
dermis); 2) Over scl of trans- nodular or spiny, up to cells in files in LS; linson, 1961 c, verse vbs; 3) Mesophyll, below 20,um, 1 per cell; 2) As 2) Basal wall thickened; 1969 palisade in 1), but smaller, 1 per cells in files (rarely clus-
cell; 3) Druse-like, up to ters) in LS; 1) & 2) Also 35 m, 1 per cell in stem, not root; 3) In
large, thin-walled cells.Some bodies appear very irregular in Thalia -
geniculata. T welwitchii and Sarcophrynium mac- rostachyum have possi- ble sand (Prychid, pers. obs.)
Calathea, Ctenanthe, Ischnosi- Mesophyll, usually in palisade Druse-like, or elongated, Possible sand in Maranta Tomlinson, 1961 c, > phon, Maranta, Mono- most small, 3-5 gm (up oligantha (Prychid, pers. 1969 tagma, Stromanthe, etc. to 20 ,um in Ctenanthe) obs.)
Donax, Saranthe Sand
Musaceae (2 genera, 40 spp.) Ensete (4 spp. examined); 1) Over vb sheath fibers; 2) In pa- 1) Rectangular in surface In files in LS; also in stem Tomlinson, 1959,
Musa (ca. 25 spp. exam- renchymatous sheath of trans- view, with central shal- and seed, not root; epi- 1969; Graven et ined) verse vbs & occasionally in low depression (trough dermal silica sand also al., 1996; Pry-
transverse septa shaped) ; spines project- seen in M. schizocarpa chid, pers. obs. ing from base into pits in and M. sapientum wall, 1 per cell; 2) Ir- regularly spherical, 1 per cell

Strelitziaceae (3 genera, 6 spp.) Phenakospermum (1 sp. exam- 1) Over vb sheath fibers & girders; 1) & 2) Spherical, spiny, in In files in LS; also in stem, Tomlinson, 1959,
ined) also in transverse septa; 2) Epi- thin-walled cells, 1 per not in root 1960, 1969 dermis over scl cell; 1) Sand may also be
present Ravenala (1 sp. examined) 1) Over vb sheath fibers & girders; 1) & 2) Spherical, spiny, in Tomlinson, 1959,
also in transverse septa; 2) Epi- thin-walled cells, 1 per 1960, 1969 dermis over scl cell; 1) Sand may also be
present Strelitzia (4-5 spp. examined) 1) Over vb sheath fibers & girders; 1) & 2) Spherical, spiny, in Tomlinson, 1959,
also in transverse septa; 2) Epi- thin-walled cells, 1 per 1960, 1969 dermis over scl cell; 1) Sand may also be
present > Zingiberaceae (50 genera, ca. 1300 spp.)
Alpinieae (21 genera, ca. 700 Epidermis over scl (internal in 1) Small, irregularly spheri- Sand in other organs, not in Tomlinson, 1956, spp.) (8 genera, 15 spp. Hornstedtia conica only) cal, 1 per cell; 2) Sand root; spherical, warty in 1962, 1969; Webb tri examined) Elettaria seed & Amott, 1982
Hedychieae (19 genera, ca. 300 Epidermis (rarely mesophyll, vb Sand; Kaempferia: small, ir- Tomlinson, 1956, spp.) (9 genera, 23 spp. sheath) (internal in Kaempferia regularly spherical, 1 per 1962, 1969 examined) only) cell 0
Zingibereae (1 genus, ca. 100 Epidermis over scl 1) Small, irregularly spheri- Tomlinson, 1956, ? spp.) (Zingiber: 2 spp. cal, 1 per cell; 2) Sand 1962, 1969 0 examined)
Globbeae (4 genera, ca. 110 Vb sheath Sand Small, irregularly spherical Tomlinson, 1956, spp.) (Globba: 4 spp. costal and rarely inter- 1962, 1969 examined) costal bodies observed in 0
Globba leucantha C (Prychid, pers. obs.)

410 THE BOTANICAL REVIEW
oooi _ ~~~~~~~~~~~~~~~~~~~~
Fig. 3. Various silica body morphologies found in the order Zingiberales. A. Canna edulis (Cannaceae), druse-like silica bodies over vascular bundle sheath (bar 1 I0 a_m). B. Costus englerianus (Costaceae), druse- like silica bodies over bundle-sheath cells (bar =20 Am). C. Heliconia psittacorum (Heliconiaceae), trough- shaped silica bodies over vascular bundle-sheath fibers (bar = 20 Am). D. Heliconia aff. tortuosa (Heliconiaceae), trough-shaped silica bodies with silica fingers projecting from the base into the cell wall (bar = 10,4.m). E. Orchidantha sp. (Lowiaceae), truncated conical silica bodies overlying a vascular bundle (bar =
10 /Ltm). F. Maranta arundinacea (Marantaceae), costal silica bodies in mesophyll cells (bar = 10 nm). G. Marantochloa purpurea (Marantaceae), a druse-like intercostal silica body in a mesophyll cell (bar = 20 akm). H. Musa schizocarpa (Musaceae), intercostal, epidermal silica sand (bar = 10 Atm).
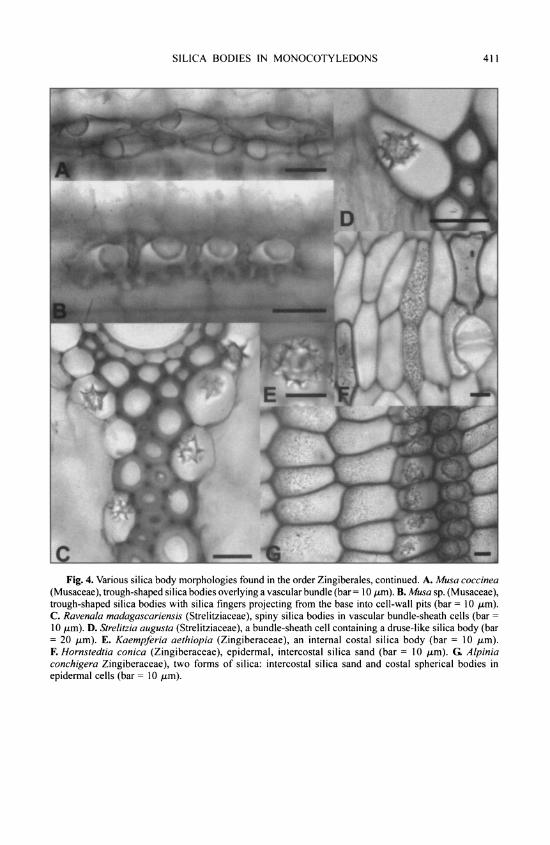
SILICA BODIES IN MONOCOTYLEDONS 411
Fig. 4. Various silica body morphologies found in the order Zingiberales, continued. A. Musa coccinea (Musaceae), trough-shaped silica bodies overlying a vascular bundle (bar 1 I0 /tm). B. Musa sp. (Musaceae), trough-shaped silica bodies with silica fingers projecting from the base into cell-wall pits (bar = 10 gm). C. Ravenala madagascariensis (Strelitziaceae), spiny silica bodies in vascular bundle-sheath cells (bar =
10 /.tm). D. Strelitzia augusta (Strelitziaceae), a bundle-sheath cell containing a druse-like silica body (bar = 20 gtm). E. Kaempferia aethiopia (Zingiberaceae), an internal costal silica body (bar = 10 gtm). F. Hornstedtia conica (Zingiberaceae), epidermal, intercostal silica sand (bar = 10 /.tm). G Alpinia conchigera Zingiberaceae), two forms of silica: intercostal silica sand and costal spherical bodies in epidermal cells (bar = 10 gm).

Table VI Distribution of silica bodies in Poales.
Key: LS = longitudinal section; scl = sclerenchyma; vb = vascular bundle.
Silica presence / location Type & no. of silica bodies per cell Notes References
Anarthriaceae (1 genus, 7 spp.) Anarthria (5 spp. examined) Absent Cutler, 1969
Bromeliaceae (56 genera, 2600+ spp.) Most genera (37 genera, Spherical, spinulose, up to 10,um, Outer wall thin, inner wall Tomlinson, 1969;
106 spp. examined) Epidermis, intercostal & 1 per cell; bodies larger over scl, thickened; never in sto- see also costal may be absent from cells over mata; also present in Bulitsch, 1892;
thin-walled hypodermis stem Linsbauer, 1911
Cryptanthus (1 sp. examined) Epidermis, intercostal & Spherical, spinulose, aggregates of Tomlinson, 1969 costal several per cell ?
Tillandsia (20 spp. examined) Epidermis, intercostal Silica sand Present in hypodermis in Baumert, 1907; > T andicola (Baumert, Tomlinson, 1907); spherical, spinu- 1969 > lose bodies seen in T stricta and possible bodies in T usneoides (Prychid pers. obs.)
8 genera (some spp.) Outer bundle sheath Silica sand Tomlinson, 1969
Centrolepidaceae (3 genera, 35 spp.) Aphelia (5 spp., 5 examined) (Epidermis) (Irregular, in 1 sp.) Silica-like? Not seen in culm Cutler, 1969 Centrolepis (c. 28 spp., 10 (Epidermis, ground tissue) (Particles or sand in 2 spp.) Silica? Also in culm of Cutler, 1969
examined) 3 spp. Gaimardia (3 spp., 3 exam- (Epidermis) (Rectangular, in 1 sp.) Silica? Cutler, 1969
ined)
Cyperaceae (104 genera, 5000+ spp.) Most genera & 86 spp. exam- Epidermis, costal (rarely in- Conical +/- satellites (rarely nodu- See Table IX for details Metcalfe, 1971
ined tercostal) lar, bridges, other types)

Ecdeiocoleaceae (2 genera, 2 spp.) Ecdeiocolea (1 sp., I exam- Chlorenchyma (culm); epi- Sand (culm) None seen in leaf Cutler, 1969;
ined) dermis, costal (culm) Prychid, pers. obs.
Eriocaulaceae (10 genera, 700-1400 spp.) 9 genera (6 examined) Absent N/A Tomlinson, 1965,
1969
Flagellariaceae (1 genus, 4 spp.) Flagellaria (1 sp. examined) Above & below vb fibrous Small, irregular, in small cells; Never epidermal; cells usu- Tomlinson, 1969
sheaths I per cell ally in longitudinal files Hydatellaceae (2 genera, 10 spp.)
Hydatella (2 spp., 1 examined) Absent N/A Cutler, 1969 >
Trithuria (3 spp., 2 examined) Absent N/A Cutler, 1969
Joinvilleaceae (1 genus, 2 spp.) Joinvillea (2 spp. examined) 1) Epidermis, short cells; 1) Smooth cubical bodies, Short cells alternating with Tomlinson, 1969
2) Vb fiber sheath 1 per cell, filling lumen; long cells in epidermis; 2) Irregular nodular bodies, silicification of meso- 1 per cell; cells in files in LS phyll cells 0
Juncaceae (7 genera, 350 spp.) ? Marsippospermum, Rostkovia, Absent N/A Cutler, 1969 8
Distichia, Patosia, Oxy- chloe, Luzula
Juncus 1) Vb fiber sheath; 2) Rarely Sand Prychid pers. obs. U mesophyll z
Mayacaceae (1 genus, 4-10 spp.) Mayaca (4 spp., 3 examined) Absent N/A Tomlinson, 1969
Poaceae (700+ genera, 10,000+ spp.) Epidermis Many shapes, not usually conical or Absent from some genera & Metcalfe, 1960
spherical spp; distribution will be discussed in another pub- lication

Table VI, continued
Silica presence / location Type & no. of silica bodies per cell Notes References
Prioniaceae (1 genus) Prionium Absent N/A Cutler, 1969
Rapateaceae (16 genera; 80 spp.) Epidermis Spherical; see Table VIII for details See Table VIII for details Carlquist, 1966;
Carlquist, in Tomlinson, 1969
Restionaceae (55 genera; 490 spp.) 1) Bundle sheaths; 2) Paren- 1) Spherical-nodular; 2) Irregular or Silica present in culms Cutler, 1969
chyma sheath, ground granular (most genera leafless); tissue see Table VII for t
details ?
Thumiaceae (I genus; 3 spp.) Z Thurnia (2 spp. examined) Epidermis (1 sp.); Paren- Nodular, several per cell, over scl; Also present in culm Cutler, 1965, 1969
chyma sand > (1 sp.)
Typhaceae (2 genera; 16-30 spp.) Sparganium (14 spp., Absent N/A Solereder &
10 examined) Meyer, 1933; Kaul, 1972; Rowlatt, un- published
Typha (8-13 spp., 8 examined) Absent N/A Meyer, 1933; Row- latt, unpub- lished
Xyridaceae (5 genera; 300 spp.) Abolboda, Achlyphila, Aratity- Absent N/A Tomlinson, 1969
opea, Orectanthe, Xyris

Table VII Distribution of silica bodies in culm of Restionaceae (classification of Linder et al., 1998).
Key: Abs = absent; chlor = chlorenchyma; epid = epidermis; par = parenchyma; scl = sclerenchyma; Sph = spherical-nodular. Brackets indicate rarely present or present in a few taxa.
Genera for which no data are available have been omitted from the table.
Genus No. of species No. studied Distribution Type and location in culm Notes References
Hopkinsia 2 1 Australia Abs Now Hopkinsiaceae a Cutler, 1969 separate family
Lyginia 3 1 or 2 Australia Abs Crystals present. Now a Solereder & Meyer, 1929; r separate family, Cutler, 1969 Hyginiaceae
Staberoha ca. 9 5 South Africa Abs Solereder & Meyer, 1929; 0 Cutler, 1969
Ischyrolepis ca. 48 1+? South Africa + Sph in par sheath Cutler, 1969 v: & ground tissue
Elegia ca. 35 21 South Africa Abs Cutler, 1969 Chondropetalum, ca. 24 16 South Africa Abs Cutler, 1969 0
Dovea, Askido- 0 sperma
Platycaulos, Restio ca. 100 ca. 60 South Africa, Sph in par sheath (& Restio groups 3-6 in Cut- Solereder & Meyer, 1929; ? Zaire, Mada- chlor); granular in ler; all his Australian Cutler, 1969 gascar par sheath &/or spp. now in other m
ground tissue, protec- genera o tive cells, chlor; Abs Z in many spp.
Calopsis ca. 23 8 South Africa Abs in 5 spp.; Sph in par South African Leptocar- Cutler, 1969 sheath or chlor pus spp. in Cutler (2 spp.); irregular in protective cells (1 sp.)
Thamnochortus ca. 34 20 South Africa Abs (irregular in epid in Granular bodies in T flori- Solereder & Meyer, 1929; T bachmannii only) bundus; rough, Sph Cutler, 1969
body in T insignis. Possible sand in T scabridus (Prychid, pers. obs.)

Table VII, continued 4
Genus No. of species No. studied Distribution Type and location in culm Notes References
Rhodocoma 7 2 South Africa Abs Cutler, 1969 Ceratocarya ca. 6 1? South Africa Sph in par sheath Cutler, 1969 Cannomois ca. 7 6 South Africa Sph in par sheath & scl Cutler, 1969
ribs Anthochortus ca. 15 2+? South Africa Sph in par sheath Incl. Phyllocomos & Cutler, 1969
Hypolaena p. p. Mastersiella 3 3? South Africa Sph in scl sheath; granular Incl. Hypolaena p. p. Cutler, 1969
in ground tissue or Abs
Hypodiscus ca. 15 ca. 9 South Africa Sph in scl sheath or Abs Solereder & Meyer, 1929; =
Cutler, 1969 Willdenowia ca. 12 7 South Africa Sph in par sheath or scl Solereder & Meyer, 1929;
sheath; granular in Cutler, 1969 ground tissue z
Lepyrodia 22 ca. 9 Australia Sph, 1 per cell in epid; Lepyrodia group 2 in Solereder & Meyer, 1929; z granular in ground tis- Cutler Cutler, 1969 > sue (some spp.) & par sheath (1 sp.)
Sporadanthus 7 5 Australia, New granular in epid (2 spp.); Lepyrodia group 1 & Spo- Cutler, 1969 Zealand Abs (3 spp.) radanthus in Cutler
Calorophus 2 1 Australia Abs See also Empodisma Cutler, 1969 Empodisma 2 2 Australia, New Sph in par sheath; granu- 2 spp. separated from Cutler, 1969; Johnson &
Zealand lar in ground tissue Calorophus Cutler, 1973 Coleocarya 1 1 Australia Sph in par sheath Cutler, 1969 Desmocladus 16 ? Australia Incl. in Hypolaena in Cut- Cutler, 1969
ler Harperia 4 1 Australia Sph in scl sheath; irregu- Cutler, 1969
lar & granular in epid & ground tissue
Onychosepalum 3 1 Australia Sph in par sheath, 1 per Cutler, 1969 cell
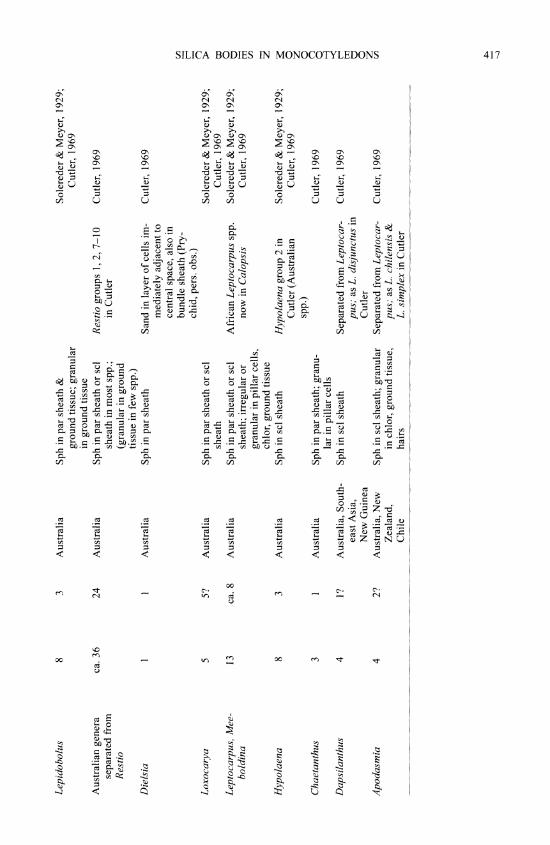
Lepidobolus 8 3 Australia Sph in par sheath & Solereder & Meyer, 1929; ground tissue; granular Cutler, 1969 in ground tissue
Australian genera ca. 36 24 Australia Sph in par sheath or scl Restio groups 1, 2, 7-10 Cutler, 1969 separated from sheath in most spp.; in Cutler Restio (granular in ground
tissue in few spp.) Dielsia 1 1 Australia Sph in par sheath Sand in layer of cells im- Cutler, 1969
mediately adjacent to central space, also in bundle sheath (Pry- chid, pers. obs.)
Loxocarya 5 5? Australia Sph in par sheath or scl Solereder & Meyer, 1929; > sheath Cutler, 1969 w
Leptocarpus, Mee- 13 ca. 8 Australia Sph in par sheath or scl African Leptocarpus spp. Solereder & Meyer, 1929; ? boldina sheath; irregular or now in Calopsis Cutler, 1969
granular in pillar cells, chlor, ground tissue
Hypolaena 8 3 Australia Sph in scl sheath Hypolaena group 2 in Solereder & Meyer, 1929; g
Cutler (Australian Cutler, 1969 spp.) 0
Chaetanthus 3 1 Australia Sph in par sheath; granu- Cutler, 1969 g lar in pillar cells
Dapsilanthus 4 1? Australia, South- Sph in scl sheath Separated from Leptocar- Cutler, 1969 east Asia, pus; as L. disjunctus in New Guinea Cutler 0
Apodasmia 4 2? Australia, New Sph in scl sheath; granular Separated from Leptocar- Cutler, 1969 '7
Zealand, in chlor, ground tissue, pus; as L. chilensis & Chile hairs L. simplex in Cutler

Table VIII Distribution of silica bodies in Rapateaceae (14 genera; 80 spp.)
(data from Carlquist, 1966; Carlquist, in Tomlinson, 1969; Prychid, pers. obs.)
Type and location Notes
Saxofridericioideae Saxofridericieae: Epidryos (1 sp. examined), Present in leaf epidermis; small, spherical, rough or Silica absent from stem
Phelpsiella (1 sp. examined), Saxofridericia spiny; several per cell (2 spp. examined), Stegolepis (2 spp. exam- ined), Amphiphyllum (1 sp. examined), Marahuacea not examined
Rapateoideae Rapateae: Rapatea (7 spp. examined), Cephalo- Present in leaf epidermis, sometimes over fibers in Silica sand and or small silica bodies in
stemon (1 sp. examined), Duckea (3 spp. Rapatea; in Rapatea, Cephalostemon and Duckea stem in Spathanthus and Monotremeae examined), Spathanthus (2 spp. examined) spherical, rough or spiny, several to many per cell;
in Spathanthus large, rough, 1 per cell or aggre- gates of sand
Monotremeae: Monotrema (4 spp. examined), Present in leaf epidermis; spherical, rough or spiny, Maschalocephalus (1 sp. examined), Potaro- several to many per cell, and/or silica sand phytum (1 sp. examined), Windsorina not examined
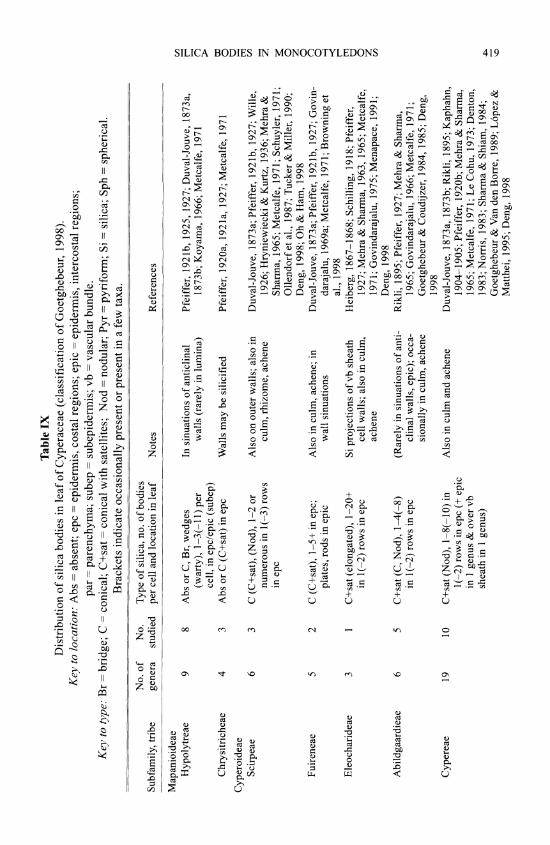
Table IX Distribution of silica bodies in leaf of Cyperaceae (classification of Goetghebeur, 1998).
Key to location: Abs = absent; epc = epidermis, costal regions; epic = epidermis, intercostal regions; par = parenchyma; subep = subepidermis; vb = vascular bundle.
Key to type: Br = bridge; C = conical; C+sat = conical with satellites; Nod = nodular; Pyr = pyriform; Si = silica; Sph spherical. Brackets indicate occasionally present or present in a few taxa.
No. of No. Type of silica, no. of bodies Subfamily, tribe genera studied per cell and location in leaf Notes References
Mapanioideae Hypolytreae 9 8 Abs or C, Br, wedges In sinuations of anticlinal Pfeiffer, 1921b, 1925, 1927; Duval-Jouve, 1873a,
(warty), 1-3(-l 1) per walls (rarely in lumina) 1873b; Koyama, 1966; Metcalfe, 1971 cell, in epc/epic (subep)
Chrysitricheae 4 3 Abs or C (C+sat) in epc Walls may be silicified Pfeiffer, 1920a, 1921 a, 1927; Metcalfe, 1971
Cyperoideae Scirpeae 6 3 C (C+sat), (Nod), 1-2 or Also on outer walls; also in Duval-Jouve, 1873a; Pfeiffer, 1921b, 1927; Wille, t
numerous in 1(-3) rows culm, rhizome, achene 1926; Hryniewiecki & Kurtz, 1936; Mehra & in epc Sharma, 1965; Metcalfe, 1971; Schuyler, 1971; g
Ollendorf et al., 1987; Tucker & Miller, 1990; O Deng, 1998; Oh & Ham, 1998 O
Fuireneae 5 2 C (C+sat), 1-5+ in epc; Also in culm, achene; in Duval-Jouve, 1873a; Pfeiffer, 1921b, 1927; Govin- g plates, rods in epic wall sinuations darajalu, 1969a; Metcalfe, 1971; Browning et i
al., 1998 Eleocharideae 3 1 C+sat (elongated), 1-20+ Si projections of vb sheath Heiberg, 1867-1868; Schilling, 1918; Pfeiffer,
in 1(-2) rows in epc cell walls; also in culm, 1927; Mehra & Sharma, 1963, 1965; Metcalfe, 0 achene 1971; Govindarajalu, 1975; Menapace, 1991; U
Deng, 1998 Abildgaardieae 6 5 C+sat (C, Nod), 1-4(-8) (Rarely in sinuations of anti- Rikli, 1895; Pfeiffer, 1927; Mehra & Sharma,
in 1(-2) rows in epc clinal walls, epic); occa- 1965; Govindarajalu, 1966; Metcalfe, 1971; sionally in culm, achene Goetghebeur & Coudijzer, 1984, 1985; Deng,
1998 Cypereae 19 10 C+sat (Nod), 1-8(-10) in Also in culm and achene Duval-Jouve, 1873a, 1873b; Rikli, 1895; Kaphahn,
1(-2) rows in epc (+ epic 1904-1905; Pfeiffer, 1920b; Mehra & Sharma, in 1 genus & over vb 1965; Metcalfe, 1971; Le Cohu, 1973; Denton, sheath in 1 genus) 1983; Norris, 1983; Sharma & Shiam, 1984;1
Goetghebeur & Van den Borre, 1989; L6pez & C Matthei, 1995; Deng, 1998

Table IX, continued
No. of No. Type of silica, no. of bodies Subfamily, tribe genera studied per cell and location in leaf Notes References
Cyperoideae, continued Dulichieae 3 2 C (C+sat), 1-5+ in epc Also in culm Pfeiffer, 1927; Hryniewiecki & Kurtz, 1936; Met-
calfe, 1971; Deng, 1998 Schoeneae 29 24 C (C+sat), (Nod, Pyr), Rods etc. in wall sinuations; Duval-Jouve, 1873a, 1873b; Kaphahn, 1904-1905;
1-6-numerous in epc also C on outer walls; Pfeiffer, 192 1b, 1921 c, 1927; Wille, 1926; (epic); C, cubical, rods, warty may be in pairs on Peisl, 1957; Norton, 1967; Govindarajalu, warty, spicules (wedges), opposite sides of anticli- 1969b, 1975; Metcalfe, 1971; Ragonese et al., 1-3+ in epic; (C, 2+ in nal walls; also in culm, 1984; Seberg, 1988; Browning & Gordon Gray, par vb sheath) rhizome, root, achene 1995; Ernst et al., 1995
Sclerioideae Cryptangieae 4 4 C (C+sat), (domed), 1-4 in Pfeiffer, 1921a, 1922, 1927; Metcalfe, 1971
1(-2) rows in epc/epic Trilepideae 4 4 C (C+sat), (Nod), 1-numer- May be 2 sizes; also in culm Pfeiffer, 1927; Chermezon, 1933; Metcalfe, 1971
ous in 1(-3) rows in epc z (epic)
Sclerieae 1 1 C (C+sat); warty, 1-3(-7) Sph in pairs on opposite Crilger, 1857; Pfeiffer, 1927; Mehra & Sharma, > in epc; Sph etc., 1-2 in sides of anticlinal walls; 1965; Metcalfe, 1971; Govindarajalu, 1975; epic Si particles in wall sinua- Franklin, 1979, 1981; Deng, 1998
tions; also in culm & achene
Bisboeckelereae 6 4 C (C+sat), (Nod), 1-3+ in Sph etc. in pairs on opposite Pfeiffer, 1921b, 1927; Koyama, 1967; Metcalfe, epc; Sph, warty, 1-2(-3), sides of anticlinal walls; 1971 & particles in epic also Si in mesophyll
Caricoideae Cariceae 5 5 C+sat, (Nod), 1-6(+) in Rarely over sclerenchyma of Duval-Jouve, 1873a; Wilczek, 1892; Pfeiffer,
1(-2) rows in epc (epic) vb sheath; also in culm, 1921b, 1925, 1927; Wille, 1926; Hryniewiecki achene & Kurtz, 1936; Mehra & Sharma, 1965;
Kukkonen, 1967; Metcalfe, 1971; Le Cohu, 1973; Walter, 1975; Toivonen & Timonen, 1976; Tallent & Wujek, 1983; Menapace & Wujek, 1987; Crins & Ball, 1988; Waterway, 1990; Lucefio, 1992; Deng, 1998, 2002; Oh & Lee, 2001; Oh et al., 2001; Starr & Ford, 2001

SILICA BODIES IN MONOCOTYLEDONS 421
6, ~ ~ . ,
Fig. 5. Various silica body morphologies found in the order Poales and in Dasypogonaceae. A. Carex intermedia (Cyperaceae), lateral view of a conical silica body with tiny spines projecting near the base (bar 10 Axm). B. Abildgaardia monostachya (Cyperaceae), conical bodies with satellites in epidermnis (bar 10 Arn). C. Juncus inflexus (Juncaceae), silica sand in bundle-sheath cells (bar = 20 /am). D. Juncus arabicus (Juncaceae), silica sand in vascular bundle-sheath cells (bar = 1 0 Atm). E. Thamnochortus floribundus (Restionaceae), an irregular or granular formn of silica observed in epidermnal cells (bar =10 Am). F. Anthochortus ecklonii (Restionaceae), spherical silica bodies overlying the sclerenchymatous bundle sheath (bar 10 IO m) G. Thurnia j'enmanil. (Thumiaceae), numerous small spherical/nodular bodies in epidermal cells (bar 1 I0 Axm). H. Kingia australis (Dasypogonaceae), spherical silica bodies with a rugose surface in epidermal cells (bar = 10 Atm). J. Dasypogon bromeliifolius (Dasypogonaceae), epidermal silica sand (bar = 20 Am).

422 THE BOTANICAL REVIEW
basal plate (Le Cohu, 1973), most frequently in one row per cell as seen in surface view but sometimes in two or three rows. Silica cells often occur in longitudinal files; there is no differ- entiation into long cells and short cells characteristic of Poaceae, with rare exceptions (bract of Epischoenus villosus and Mesomelaena stygia). The number of bodies and rows per cell may be diagnostic for a species, but there is always a range of values, and variation due to age of leaf, environmental conditions, etc. must be taken into account (see Ollendorf, 1992). Each large cone may be surrounded by a circle of small cones, termed "satellites," as seen in surface view; the extent to which satellites are developed tends to vary within a species.
A conical body may have two or more peaks, when it is transitional to the warty or nodular type, which may be conical, spherical or irregularly shaped. In certain genera and species, smooth or warty hemispherical bodies are formed on anticlinal walls of adjoining epidermal cells; e.g., in Rhynchospora and Scleria (Govindarajalu, 1969b, 1975; Metcalfe, 1971). Other types of body have been described as wedge shaped, often based on sinuations in the anticlinal walls, or bridge shaped, when they traverse the cell lumina; these types are characteristic of Hypolytreae. In certain genera, small silica particles, rods, plates or cubical bodies may occur, especially in intercostal regions, where individual cells, including stomatal guard or subsidiary cells and prickles, may be completely silicified.
Silica cells occur both adaxially and abaxially in the leaf epidermis, but the frequency varies according to the number of subepidermal sclerenchyma strands or girders, which in some gen- era are nearly equally numerous on both surfaces, while in others they are more numerous abaxially (rarely adaxially). Silica bodies are sporadically present in intercostal regions of the epidermis and rarely in mesophyll or diaphragm cells (e.g., in the leaf of Cladium articulatum: Peisl, 1957). Kaphahn (1904-1905) noted silica bodies in parenchymatous cells overlying the sclerenchymatous vascular bundle sheath and at borders of sclerenchyma girders in the leaf of Cladium germanicum. They were seen in a similar position by Pfeiffer (1921b) in leaf and culm of Cyperus mariscus.
Silica bodies, usually of similar type to those in the leaf, are present in the epidermis of leaf sheath and culm and, rarely, in other positions; for example, in cells overlying vascular bundles. Silica projections from parenchymatous bundle-sheath cell walls bordering air cavities were observed in Eleocharis plantaginea culm by Schilling (1918) and Mehra and Sharma (1963). Bodies have been observed infrequently in the rhizome (Wille, 1926) and rarely in the root (e.g., in some exodermal cells of Rhynchospora spp.: Govindarajalu, 1975).
Wilczek (1892) was one of the first to investigate achenes of Cyperaceae for silica bodies. He examined seven genera but found bodies only in Carex species, where there was one coni- cal body per epidermal cell. More recently, several taxonomic studies have included observa- tions of bodies in achenes, often using SEM; e.g., Bulbostylis (Goetghebeur & Coudijzer, 1985), Carex (Walter, 1975; Toivonen & Timonen, 1976; Wujek & Menapace, 1986; Menapace & Wujek, 1987; Crins & Ball, 1988; Standley, 1990; Waterway, 1990; Lucenfo, 1992; Oh & Lee, 2001; Oh et al., 2001; Starr & Ford, 2001), Eriophorum and Scirpus (Schuyler, 1971; Tucker & Miller, 1990), Lipocarpha and related genera (Goetghebeur & Van den Borre, 1989), Rhynchospora (Ragonese et al,. 1984) and Uncinia (Starr et al., 2003), although none was seen in 3 species of Abildgaardia or 19 species of Fimbristylis (Goetghebeur & Coudijzer, 1984).
Table VIII shows the type and numbers of silica bodies in the leaf of tribes of Cyperaceae. The data are taken mainly from Metcalfe (1971), but the observations for individual genera have been regrouped according to Goetghebeur's classification in Kubitzki (1998), to which the numbers of genera refer. The conical type (+ satellites) is dominant in epidermal cells overlying sclerenchyma in all tribes except Hypolytreae and Chrysitricheae, which together constitute the Mapanioideae; here silica bodies are absent, restricted to small cones in sinuations

SILICA BODIES IN MONOCOTYLEDONS 423
of anticlinal walls, or wedge or bridge shaped, rarely of the typical conical type. The composi- tion of the Mapanioideae has not changed since Clarke's (1908) classification, except that Syntrinema, with conical bodies, has been removed and included in Rhynchospora. The Cariceae have also remained almost unaltered and are characterized by 1-5 or more conical bodies with satellites per cell, sometimes in two rows.
An alternative suprageneric classification, based on cladistic analyses of 122 genera using more than 300 morphological and other characters, was proposed by Bruhl (I995); he included silica body type and position in culm and leaf among his characters, and his article should be consulted for a full discussion of the differences between his classification and that of Goetghebeur (1986).
Conical silica bodies are typical for most Cyperaceae, but they are not confined to this family; for example, they are also present in some Orchidaceae and Arecaceae, although in the two latter families there is usually only one body per cell, and they are never epidermal.
Although Prionium was traditionally placed either in Juncaceae or in its own family, Prioniaceae (Munro & Linder, 1998), analyses of molecular data place it as sister to Thumiaceae (e.g., Chase et al., 2000; Michelangeli et al., 2003). However, silica bodies are apparently absent from Prionium and all Juncaceae apart from Juncus, which possesses silica sand in vascular bundle-sheath cells (Figs. 5C, SD), but present in Thurnia (Cutler, 1965, 1969) as small, spherical bodies (Fig. SG). Thus, this distribution of silica bodies may indicate two separate losses of silica bodies in the sedge clade.
There is also at least one loss of silica bodies in the grass clade. Flagellaria, which pos- sesses silica bodies, is sister to two subclades, one comprising Restionaceae and their allies; the other, grasses and their allies. In the first subclade, three genera (Anarthria, Hopkinsia and Lyginia) form a clade that is sister to Restionaceae. Hopkinsia and Lyginia are former Restionaceae that were recently proposed as separate families, Hopkinsiaceae and Lyginiaceae (Briggs & Johnson, 2000), since a molecular analysis (Briggs et al., 2000) found them to be isolated from Restionaceae and sister to Anarthria (Anarthriaceae). Interestingly, all three gen- era (Anarthria, Hopkinsia and Lyginia) apparently lack silica bodies.
Solereder and Meyer (1929) were the first to recognize silica bodies in Restionaceae, but the first major anatomical study was by Cutler (1969); his observations were confirmed by Linder (1984). Table VII is based on Cutler's (1969) data, rearranged according to the classifi- cation of Linder et al. (1998). Many genera are leafless, and most available data are for culms. Silica bodies occur in the leaf or leaf base of a few species and occasionally in rhizomes but have not been seen in roots. The recent taxonomy of Restionaceae is complex, with many genera and species transferred to different taxa (Linder, 2000). Cutler's (1969) work showed that there were anatomical differences between African and Australian species that had previ- ously been placed in the same genera; e.g., Restio and Leptocarpus. Certain species have sub- sequently been transferred to other genera, so that there is now no genus with a distribution in both South Africa and Australia. Silica is absent from several genera and from some species in other genera. When present, silica is found in two main forms in Restionaceae: 1) granular silica, usually filling cell lumina, in cells of parenchymatous sheath and central ground tissue and occasionally in pillar cells, protective cells (see Cutler, 1969, for definitions of these terms) and chlorenchyma but rarely in the epidermis (Fig. 5E); and 2) spherical-nodular bodies in unspecialized cells or in stegmata, forming part of the parenchymatous bundle sheath or the outer layer of the sclerenchymatous sheath (Fig. SF). Stegmata have thickened, lignified or suberized inner and anticlinal walls and thin outer walls. Both types may be present in one species, and they occur in both South African and Australian species, as does absence of silica. Cutler (1969) divided Lepyrodia species into two main groups based on anatomical characters,

424 THE BOTANICAL REVIEW
Fig. 6. Variousilicaboymorphoogiesfondinthe rl cs of P . A. A a s costal dumbbell-shaped silica bodies (bar 10lpm). B. Brachiariajubata, a_ form of silica _termediate
lines6 (bariou20silmca F.dAstoebhaogiquarrosa saddle-shapednsilicas bodiesc(bar A.10risLm)a GeAgropron
elongatum, a conical silica body (bar 20 Am)
one of which was the presence of granular silica alone in one group and spherical-nodular bodies in the other group; species in the group with only granular silica have since been trans- ferred to Sporadanthus. Silica bodies have not been observed in any species examined of Staberoha or in the large genera Elegia and Chondropetalum.
In the other grass subclade (grasses and their allies), the bigeneric family Ecdeiocoleaceae is putatively sister to Poaceae, with Joinvillea sister to this pair (Michelangeli et al., 2003). Silica is present in all three families, although only as silica sand in culms of Ecdeiocolea (Ecdeiocoleaceae). Silica bodies are present in most species of Poaceae, including Anomochloa, Streptochaeta and Pharus, which are putatively sister to the rest of the family (GPWG, 200 1). T'here is a diverse range of silica bodies in Poaceae, including dumbbell-shaped, cross-shaped

SILICA BODIES IN MONOCOTYLEDONS 425
Table X Distribution of silica bodies in Dasypogonaceae
No. of Genus species Type and location of silica References
Baxteria 1 Spherical silica bodies with rugose surface pres- Fahn, 1954; Rudall & ent in leaf epidermal cells, generally one per Chase, 1996 cell
Calectasia 3 Fine silica sand or amorphous crystals present in Rudall & Chase, 1996 leaf epidermis
Dasypogon 3 Fine silica sand or amorphous crystals present in Rudall & Chase, 1996 leaf epidermis
Kingia 1 Spherical silica bodies with rugose surface, Fahn, 1954; Rudall & present in the leaf epidermal cells, generally Chase, 1996 one per cell
intermediates between these two types, horizontally elongated shapes with smooth or sinuous outlines, saddle shaped, conical shaped and numerous others (Figs. 6A-6G) (see Metcalfe, 1960). The distribution and structure of silica bodies in Poaceae will be discussed in more detail in a separate publication.
4. Dasypogonaceae
The phylogenetic position of Dasypogonaceae within the commelinid clade remains equivo- cal. For example, a relationship with palms was tentatively indicated by molecular data, but without bootstrap support (Chase et al., 1995a, 2000; Givnish et al., 1999). Rudall and Chase (1996) noted that habit and method of growth would support a relationship between Dasypogonaceae and Arecaceae, but ovule structure and silica position and morphology indi- cate an affinity with Rapateaceae. Within Dasypogonaceae (Table X), calcium oxalate crystals are absent, but silica is present in all four genera (Rudall & Chase, 1996). In Kingia and Baxteria, spherical (druse-like) silica bodies with a rugose surface are present in the leaf epidermal cells, generally one per cell (Fig. 5H). These were previously reported by Fahn (1954) as druses (clustered crystals of calcium oxalate), but X-ray and SEM examination effectively demon- strated their silicaceous nature. Silica is also present in the epidermis of Dasypogon and Calectasia in the form of fine silica sand or amorphous crystals, present in most epidermal cells in Dasypogon (Fig. 5J) but less frequent in Calectasia.
XII. Literature Cited
Acuna-Mesen, R. & E. Garcia-Diaz. 1998. New Cuvieronius hyodon (Proboscidea: Gomphotheriidae) from the Pleistocene of Costa Rica. Revista Biol. Trop. 46: 1167-1172.
Agarie, S., W. Agata, H. Uchida, F. Kubota & P. Kaufman. 1996. Function of silica bodies in the epidermal system of rice (Oryza sativa L.): Testing the window hypothesis. J. Exp. Bot. 47: 655- 660.
Allingham, M. M., J. M. Cullen, C. H. Giles, S. K. Jain & J. S. Woods. 1958. Adsorption at inorganic surfaces, II. Adsorption of dyes and related compounds by silica. J. Appl. Chem. 8: 108-116.
Amick, J. 1982. Purification of rice hulls as a source of solar grade silicon for solar cells. J. Electrochem. Soc. 129: 864-866.
Ancibor, E. 1995. Palmeras f6siles del Cretacico Tardio de la Patagonia Argentina (Bajo de Santa Rosa, Rio Negro). Ameghiniana 32: 287-299 (Spanish; English summary).
APG (Angiosperm Phylogeny Group). 1998. An ordinal classification for the families of flowering plants. Ann. Missouri Bot. Gard. 85: 531-553.

426 THE BOTANICAL REVIEW
APG II (Angiosperm Phylogeny Group II). 2003. An update of the Angiosperm Phylogeny Group clas- sification for the orders and families of flowering plants: APG II. Bot. J. Linn. Soc. 141: 399-436.
Armitage, P. L. 1975. The extraction and identification of opal phytoliths from the teeth of ungulates. J. Archaeol. Sci. 2: 187-197.
Ashton, M. J. & M. M. Jones. 1976. A study of the transpiration surfaces of Avena sterilis L. var. Algerian leaves using monosilicic acid as a tracer for water movement. Planta 130: 121-129.
Baker, G 1961. Opal phytoliths and adventitious mineral particles in wheat dust. Mineral Investigations Technical Paper No. 4. CSIRO, Melbourne, Australia.
L. H. P. Jones & I. D. Wardrop. 1959. Cause of wear in sheep's teeth. Nature 184: 1583-1584. ,~ & . 1961. Opal phytoliths and mineral particles in the rumen of the sheep. Aus-
tral. J. Agric. Res. 12: 462-473. Balasta, M. L. F. C., C. M. Perez, B. 0. Juliano, C. P. Villareal, J. N. A. Lott & D. B. Roxas. 1989.
Effects of silica level on some properties of Oryza sativa straw and hull. Canad. J. Bot. 67: 2356-2363. Ball, T. B., J. D. Brotherson & J. S. Gardner. 1993. A typologic and morphometric study of variation in
phytoliths from einkorn wheat (Triticum monococcum). Canad. J. Bot. 71: 1182-1192. Barthlott, W. & D. Frolich. 1983. Mikromorphologie und Orientierungsmuster epicuticularer Wachs-
Kristalloide: Ein neues systematisches Merkmal bei Monokotylen. P1. Syst. Evol. 142: 171-185. Baumert, K. 1907. Experimentelle Untersuchungen uiber Lichtschutzeinrichtungen an griunen Blattern.
Beitr. Biol. Pfl. 9: 83-162 (also diss., Erlangen, 1907). Bennett, D. M. & D. W. Parry. 1981. Electron-probe microanalysis studies of silicon in the epicarp hairs
of the caryopses of Hordeum sativum Jess., Avena sativa L., Secale cereale L. and Triticum aestivum L. Ann. Bot. 48: 645-654.
Bertoldi de Pomar, H. 1971. Ensayo de clasificaci6n morfol6gica de los silicofitolitos. Ameghiniana 8: 317-328 (English summary).
Bezeau, L. M., A. Johnston & S. Smoliak. 1966. Silica and protein content of mixed prairie and fescue grassland vegetation and its relationship to the incidence of silica urolithiasis. Canad. J. PI. Sci. 46: 625-631.
Bienfait, A., L. Waterkeyn & L. Ermin. 1985. Importance des verrues foliaires silicifiees dans la systematique des Selaginella: Observations en microscopie electronique a balayage (MEB). Bull. Jard. Bot. Belg. 55: 73-81.
Birchall, J. D. 1990. The role of silicon in biology. Chemistry in Britain 26: 141-144. Blackman, E. 1968. The pattern and sequence of opaline silica deposition in rye (Secale cereale L.). Ann.
Bot., n.s., 32: 207-218. . 1969. Observations on the development of the silica cells of the leaf sheath of wheat (Triticum
aestivum). Canad. J. Bot. 47: 827-838. . 1971. Opaline silica bodies in the range grasses of southern Alberta. Canad. J. Bot. 49: 769-781.
Blinnikov, M. S. 1994. Phytolith analysis and holocene dynamics of alpine vegetation. Veroff. Geobot. Inst. Rtibel 115: 23-40 (English).
, A. Busacca & C. Whitlock. 2002. Reconstruction of the late Pleistocene grassland of the Co- lumbia basin, Washington, USA, based on phytolith records in loess. Palaeogeog. Palaeoclim. Palaeoecol. 177: 77-101.
Bode, E., S. Kozik, U. Kunz & H. Lehmann. 1994. Comparative electron-microscopic studies on the process of silicification in leaves of 2 different grass species. Wochenschrift 101: 367-372.
Bozarth, S. R. 1993. Maize (Zea mays) cob phytoliths from a Central Kansas Great Bend aspect archaeo- logical site. Plains Anthropologist 38: 279-286.
Brady, N. C. 1990. The nature and properties of soils. Ed. 10. Macmillan, New York. Brandenburg, D. M., S. D. Russell, J. R. Estes & W. F. Chissoe. 1985. Backscattered electron imaging
as a technique for visualizing silica bodies in grasses. Scan. Electron Microscop. 1985(4): 1509-1517. Briggs, B. GC & L. A. S. Johnson. 2000. Hopkinsiaceae and Lyginiaceae, two new families of Poales in
Western Australia, with revision of Hopkinsia and Lyginia. Telopea 8: 477-502. , A. D. Marchant, S. Gilmore & C. L. Porter. 2000. A molecular phylogeny of Restionaceae and
allies. Pp. 661-671 in K. L. Wilson & D. A. Morrison (eds.), Monocots: Systematics and evolution. Vol. 1. CSIRO, Melbourne, Australia.
Brown, D. A. 1984. Prospects and limits of a phytolith key for grasses in the central United States. J. Archaeol. Sci. 11: 345-368.

SILICA BODIES IN MONOCOTYLEDONS 427
Browning, J. & K. D. Gordon Gray. 1995. Studies in Cyperaceae in southern Africa, 26: Glume epider- mal silica deposits as a character in generic delimitation of Costularia and Cyathocoma as distinct from Tetraria and other allies. S. Afr. J. Bot. 61: 66-71.
, ,~I~ S. Galen Smith & J. van Staden. 1998. Bolboschoenus glaucus (Cyperaceae), with emphasis upon Africa. Nordic J. Bot. 18: 475-482.
Bruhl, J. J. 1995. Sedge genera of the world: Relationships and a new classification of the Cyperaceae. Austral. Syst. Bot. 8: 125-305.
Bulitsch, A. 1892 [1894]. Zur Anatomie der Bromeliaceae, II. Ausscheidung von Kieselerde in den Blattepidermiszellen einiger Bromeliaceen. [In Russian] Uebers. Leist. Bot. Russland 1892, St. Pe- tersburg 37-38; 1894. [See Just's Jber. 21 (1): 539-540, No. 31, 1893]
Bush, M. B., D. R. Piperno & P. A. Colinvaux. 1989. A 6,000 year history of Amazonian maize cultiva- tion. Nature 340: 303-305.
Cailin, W., H. Fujiwara, T. Udatsu & T. Linghua. 1994. Morphological features of silica bodies from motor cells in local and modern cultivated rice (Oryza sativa L.) from China. Ethnobotany 6: 77-86.
Cameron, K. M. & M. W. Chase. 2000. Nuclear 18s rDNA sequences of Orchidaceae confirm the subfamilial status and circumscription of Vanilloideae. Pp.457-464 in K. L. Wilson & D. A. Morrison (eds.), Monocots: Systematics and evolution. Vol. 1. CSIRO, Melbourne, Australia.
, - , W. M. Whitten, P. J. Kores, D. C. Jarell, V. A. Albert, T. Yukawa, H. G Hills & D. H. Goldman. 1999. A phylogenetic analysis of the Orchidaceae: Evidence from rbcL nucleotide se- quences. Amer. J. Bot. 86: 208-224.
Carlquist, S. 1966. Anatomy of Rapateaceae-Roots and stems. Phytomorphology 16: 17-38. Carlsward, B. S., W. L. Stern, W. S. Judd & T. W. Lucansky. 1997. Comparative leaf anatomy and
systematics in Dendrobium, sections Aporum and Rhizobium (Orchidaceae). Int. J. PI. Sci. 158: 332-342.
Chandrasekhar, S., P. M. Pramada, P. Raghavan, K. G Satyanarayana & T. N. Gupta. 2002. Microsilica from rice husk as a possible substitute for condensed silica fume for high performance concrete. J. Mater. Sci. Lett. 21: 1245-1247.
Chapuis, J. L. 1980. Methodes d'etude du regime alimentaire du lapin de Garenne, Oryclolagus cunicu- lus (L.) par l'analyse micrographique des feces. Terre et Vie 34: 159-198.
Chase, M. W., M. R. Duvall, H. G Hills, J. G Conran, A. V. Cox, L. E. Eguiarte, J. Hartwell, M. F. Fay, L. R. Caddick, K. M. Cameron & S. Hoot. 1 995a. Molecular phylogenetics of Lilianae. Pp. 109-137 in P. J. Rudall, P. J. Cribb, D. F. Cutler & C. J. Humphries (eds.), Monocotyledons: Sys- tematics and evolution. Royal Botanic Gardens, Kew.
, D. W. Stevenson, P. Wilkin & P. J. Rudall. 1995b. Monocot systematics: A combined analysis. Pp. 685-730 in P. J. Rudall, P. J. Cribb, D. F. Cutler & C. J. Humphries (eds.), Monocotyledons: Systematics and evolution. Royal Botanic Gardens, Kew.
, D. S. Soltis, P. S. Soltis, P. J. Rudall, M. F. Fay, W. H. Hahn, S. Sullivan, J. Joseph, T. Givnish, K. J. Sytsma & C. Pires. 2000. Higher-level systematics of the monocotyledons: An assessment of current knowledge and a new classification. Pp. 3-16 in K. L. Wilson & D. A. Morrison (eds.), Monocots: Systematics and evolution. Vol. 1. CSIRO, Melbourne, Australia.
Chen, C. & J. C. Lewin. 1969. Silicon as a nutrient element for Equisetum arvense. Canad. J. Bot. 47: 125-131.
Chermezon, H. 1933. Observations sur le genre Microdracoides. Bull. Soc. Bot. Fr. 80: 90-97. Cherouvrier, A., A. Gueguen & J.-C. Lefeuvre. 1975. Essai de determination du regime alimentaire
d'animaux herbivores a l'aide des phytolithes siliceux des Graminees et des Cyperacees: Descrip- tion, apres etude en microscopie electronique a balayage, des principaux types de phytolithes rencontres. Compt. Rend. Hebd. Acad. Sci., Paris 281: 839-842.
Chevalier, L., C. Desbuquois, J. Le Lannic & M. Charrier. 2001. Poaceae in the natural diet of the snail Helix aspersa Muller (Gastropoda, Pulmonata). Compt. Rend. Hebd. Acad. Sci., ser. III, Sci. Vie, 324: 979-987.
Clarke, C. B. 1908. New genera and species of Cyperaceae. Kew Bull., addit. ser., 8: 1-196. C6ot, W. A. 1974. Rice husk characterization using SEM and EDXA. J. Indian Acad. Wood Sci. 5: 3-17. Crins, W. J. & P. W. Ball. 1988. Sectional limits and phylogenetic considerations in Carex Section
Ceratocystis (Cyperaceae). Brittonia 40: 38-47. Cruger, H. 1857. Westindische Fragmente, 9. El Cauto. Bot. Zeitung 15: 281-292, 297-308.

428 THE BOTANICAL REVIEW
Cummings, L. S. & A. Magennis. 1997. A phytolith and starch record of food and grit in Mayan human tooth tartar. Pp. 211-218 in A. Pinilla, J. Juan-Tresserras & M. J. Machado (eds.), Estado actual de los estudios de fitolitos en suelos y plantas. Monografias del Centro de Ciencias Medioambientales, 4. Consejo Superior de Investigaciones Cientificas and Centro de Ciencias Medioambientales, Madrid.
Cutler, D. F. 1965. Vegetative anatomy of Thurniaceae. Kew Bull. 19: 431-441. . 1969. Anatomy of the monocotyledons, IV. Juncales. Clarendon Press, Oxford.
Dahlgren, R. M. T., H. T. Clifford & P. F. Yeo. 1985. The families of the monocotyledons: Structure, evolution, and taxonomy. Springer-Verlag, Berlin.
Davies, K. L. 1999. A preliminary survey of foliar anatomy in Maxillaria. Lindleyana 14: 126-135. Davis, K. M. C., J. A. Deuchar & D. A. Ibbitson. 1973. Adsorption of phenols from non-polar solvents
onto silica gel. J. Chem. Soc. Faraday Trans., I, 69: 1117-1126. Davy, H. 1814. Elements of agricultural chemistry: In a course of lectures for the Board of Agriculture.
Ed. 2. J. G. Barnard, London. Dayanandan, P. 1983. Localization of silica and calcium carbonate in plants. Scan. Electron Microscop.
1983(3): 1519-1524. Deflandre, G 1963. Les phytolithaires (Ehrenberg): Nature et signification micropaleontologique, pedo-
logique et geologique. Protoplasma 57: 234-259. Deng, D. 1998. The studies on phytolith system of Cyperaceae. Guihaia 18: 204-208 + 3 plates (Chinese;
English summary). . 2002. Studies on phytolith system of Kobresia (Cyperaceae). Guihaia 22: 394-398 + 3 plates
(Chinese; English summary). Denton, M. F. 1983. Anatomical studies of the Luzulae group of Cyperus (Cyperaceae). Syst. Bot. 8:
250-262. Dixon, H. H. 1894. On the vegetative organs of Vanda teres. Proc. Roy. Irish Acad., ser. 3, 3: 441-458. Djamin, A. & M. Pathak. 1967. Role of silica in resistance to the Asiatic rice borer, Chilo suppressalis
(Walker), in rice varieties. J. Econ. Entomol. 60: 347-351. Doolittle, W. E. & C. D. Frederick. 1991. Phytoliths as indicators of prehistoric maize (Zea mays subsp.
mays, Poaceae) cultivation. P1. Syst. Evol. 177: 175-184. Dormaar, J. F. & L. E. Lutwick. 1969. Infrared spectra of humic acids and opal phytoliths as indicators
of palaeosols. Canad. J. Soil Sci. 49: 29-37. Dorweiler, J. E. & J. Doebley. 1997. Developmental analysis of teosinte glume architecture. 1: A key
locus in the evolution of maize (Poaceae). Amer. J. Bot. 84: 1313-1322. Dressler, R. L. 1993. Phylogeny and classification of the orchid family. Dioscorides Press, Portland, OR.
& S. L. Cook. 1988. Conical silica bodies in Eriajavanica. Lindleyana 3: 224-225. Drum, R. W. 1968. Electron microscopy of opaline phytoliths in Phragmites and other Gramineae. Amer.
J. Bot. 55: 713 (abstract). Dunne, T. 1978. Rates of chemical denudation of silicate rocks in tropical catchments. Nature 274: 244-246. Duval-Jouve, J. 1873a. Sur une forme de cellules epidermiques qui paraissent propres aux Cyperacees.
Bull. Soc. Bot. Fr. 20: 91-95. . 1873b. Dieselbe Arbeit mit einem Nachsatze. Mem. Acad. Sci. Lett. Montpellier 8: 227.
Eberwein, R. 1903. Zur Anatomie des Blattes von Borassus flabelliformis. Sitzungsber. Akad. Wiss. Wien 112: 67-76.
Ehrenberg, C. G 1841. Nachtrag zu dem Vortrage uber Verbreitung und Einfluss des mikroskopischen Lebens in Sud- und Nordamerika. Monatsber. Preuss. Akad. Wiss. Berlin, 139-144.
Ernst, W. H. O., R. D. Vis & F. Piccoli. 1995. Silicon in developing nuts of the sedge Schoenus nigricans. J. PI. Physiol. 146: 481-488.
Espinoza de Pernia, N. 1987. Cristales y silice en maderas dicotiled6neas de Latinoamerica. Pittieria 15: 13-65.
Evans, T. M., R. B. Faden, M. G Simpson & K. J. Sytsma. 2000. Phylogenetic relationships in the Commelinaceae, I: A cladistic analysis of morphological data. Syst. Bot. 25: 668-691.
Exley, C. & J. D. Birchall. 1992. Hydroxyaluminosilicate formation in solutions of low total aluminium concentration. Polyhedron 11: 1901-1907.
& . 1993. A mechanism of hydroxyaluminosilicate formation. Polyhedron 12: 1007-1017. Faden, R. B. & K. E. Inman. 1996. Leaf anatomy of the African genera of Commelinaceae: Anthericopsis
and Murdannia. Pp. 464-471 in L. J. G. van der Maesen, X. M. van der Burgt & J. M. van Medenbach

SILICA BODIES IN MONOCOTYLEDONS 429
de Rooy (eds.), The biodiversity of African plants: Proceedings, XIVth AETFAT Congress, 22-27 August 1994, Wageningen, The Netherlands. Kluwer Academic, Dordrecht, Germany.
Fahn, A. 1954. The anatomical structure of the Xanthorrhoeaceae Dumort. J. Linn. Soc., Bot. 55: 158-184. Fay, M. F., P. J. Rudall, S. Sullivan, K. L. D. Stobart, A. Y. Bruijn, G Reeves, F. Qamaruz Zaman,
W.-P. Hong, J. Joseph, W. J. Hahn, J. G Conran & M. W. Chase. 2000. Phylogenetic studies of Asparagales based on four plastid DNA regions. Pp. 360-371 in K. L. Wilson & D. A. Morrison (eds.), Monocots: Systematics and evolution. Vol. 1. CSIRO, Melbourne, Australia.
Franklin, E. F. 1979. A note on the hairy achenes of four African species of Scleria Bergius (Cyperaceae). Bot. J. Linn. Soc. 79: 333-341.
. 1981. SEM examination of silica (Si02) deposits isolated from achenes of Scleria (Cyperaceae). Proc. Electron Microsc. Soc. S. Afr. 11: 147-148.
Fredlund, G G & L. T. Tieszen. 1994. Modern phytolith assemblages from the North American Great Plains. J. Biogeog. 21: 321-335.
Freudenstein, J. V. & F. N. Rasmussen. 1999. What does morphology tell us about orchid relationships? A cladistic analysis. Amer. J. Bot. 86: 225-248.
, D. M. Senyo & M. W. Chase. 2000. Mitochondrial DNA and relationships in the Orchidaceae. Pp. 421-429 in K. L. Wilson & D. A. Morrison (eds.), Monocots: Systematics and evolution. Vol. 1. CSIRO, Melbourne, Australia.
Frohnmeyer, M. 1914. Die Entstehung und Ausbildung der Kieselzellen bei den Gramineen. Biblioth. Bot. 21, Heft 86: 1-41.
Frolich, D. & W. Barthlott. 1988. Mikromorphologie der epicuticularen Wachse und das System der Monokotylen. Trop. Subtrop. Pflwelt 63: 1-135 (English summary).
Fujiwara, H. & A. Sasaki. 1978. Fundamental studies in plant opal analysis (2): The shape of the silica bodies of Oryza. Archaeol. Nat. Sci. 11: 9-20.
, Y. I. Sato, H. Kaidama & T. Udatsu. 1990. Studies on the historical change of rice strains by the morphological analysis of plant opal. J. Archaeol. Soc. Nippon 75: 93-102.
Gali Muhtasib, H. U., C. C. Smith & J. J. Higgins. 1992. The effect of silica in grasses on the feeding behavior of the prairie vole, Microtus ochrogaster. Ecology 73: 1724-1729.
Gartner, S., C. Charlot & N. Paris-Pireyre. 1984. Microanalyse de la silice et resistance A la verse mecanique de ble tendre. Physiol. Veg. 22: 811-820.
Gattuso, M. A., S. J. Gattuso & A. M. Ferri. 1998 [1999]. Anatomical study on the origin and develop- ment of the crown and silica deposition in Johnsongrass (Sorghum halepense (L.) Pers.). Phyto- morphology 48: 357-370.
Ghose, M. & B. M. Johri. 1987. Cell inclusions in vegetative structure of young palms. Proc. Indian Natl. Sci. Acad., B, 53: 193-196.
Ginieis, C. 1964. Les stegmates: Leur origine, leur developpement, leur repartition. Bull. Soc. Linn. Lyon 33: 282-294, 304-307.
Givnish, T., T. M. Evans, J. C. Pires & K. J. Sytsma. 1999. Polyphyly and covergent morphological evolution in Commelinales and Commelinidae: Evidence from rbcL sequence data. Molec. Phylog. Evol. 12: 360-385.
Gobetz, K. E. & S. R. Bozarth. 2001. Implications for late Pleistocene mastodon diet from opal phytoliths in tooth calculus. Quaternary Res. 55: 115-122.
Goetghebeur, P. 1986. Genera Cyperacearum: Een bijdrage tot de kennis van de morfologie, systematiek en fylogenese van de Cyperaceae-genera. Thesis, Rijksuniversiteit Gent.
. 1998. Cyperaceae. Pp. 141-190 in K. Kubitzki (ed.), The families and genera of vascular plants, IV. Flowering plants: Monocotyledons: Alismatanae and Commelinanae (except Gramineae). Springer- Verlag, Berlin.
& J. Coudijzer. 1984. Studies in Cyperaceae, 3: Fimbristylis and Abildgaardia in Central Africa. Bull. Jard. Bot. Belg. 54: 65-89.
& . 1985. Studies in Cyperaceae, 5: The genus Bulbostylis in Central Africa. Bull. Jard. Bot. Belg. 55: 207-259.
& A. Van den Borre. 1989. Studies in Cyperaceae, 8: A revision of Lipocarpha, including Hemicarpha and Rikliella. Wageningen Agric. Univ. Pap. 89(1): 1-87.
Goldblatt, P., J. E. Henrich & P. Rudall. 1984. Occurrence of crystals in Iridaceae and allied families and their phylogenetic significance. Ann. Missouri Bot. Gard. 71: 1013-1020.

430 THE BOTANICAL REVIEW
Govindarajalu, E. 1966. The systematic anatomy of South Indian Cyperaceae: Bulbostylis Kunth. J. Linn. Soc., Bot. 59: 289-304.
. 1969a. The systematic anatomy of South Indian Cyperaceae. Fuirena Rottb. Bot. J. Linn. Soc. 62: 27-40.
. 1969b. Observations on new kinds of silica deposits in Rhynchospora spp. Proc. Indian Acad. Sci., B, 70: 28-36.
. 1975. The systematic anatomy of South Indian Cyperaceae: Eleocharis R.Br., Rhynchospora Vahl and Scleria Bergius. Adansonia, ser. 2, 14: 581-632.
GPWG (Grass Phylogeny Working Group). 2001. Phylogeny and subfamilial classification of the grasses (Poaceae). Ann. Missouri Bot. Gard. 88: 373-457.
Graven, P., C. G de Koster, J. J. Boon & F. Bouman. 1996. Structure and macromolecular composition of the seed coat of the Musaceae. Ann. Bot. 77: 105-122.
Grob, A. 1896. Beitraige zur Anatomie der Epidermis der Gramineenbltitter. Biblioth. Bot. 7(36): 1-64. Gueguen, A., A. Cherouvrier & J. C. Lefeuvre. 1975. Essai de determination du regime alimentaire
d'animaux herbivores a l'aide des phytolithes siliceux des Graminees et des Cyperacees, II: Applica- tion a l'etude du regime alimentaire des Orthopteres. Compt. Rend. Hebd. Acad. Sci., Paris 281: 929-932.
Gugel, I. L., G Grupe & K. H. Kunzelmann. 2001. Simulation of dental microwear: Characteristic traces by opal phytoliths give clues to ancient human dietary behavior. Amer. J. Phys. Anthropol. 114: 124-138.
Hanifa, A. M., T. R. Subramaniam & B. W. X. Ponnaiya. 1974. Role of silica in resistance to the leaf roller, Cnaphalocrocis medinalis Guenee, in rice. Indian J. Exp. Biol. 12: 463-465.
Harbers, L. H., D. J. Raiten & G M. Paulsen. 1981. The role of plant epidermal silica as a structural inhibitor of rumen microbial digestion in steers. Nutr. Rep. Int. 24: 1057-1066.
Harris, P. J. & R. D. Hartley. 1980. Phenolic constituents of the cell walls of monocotyledons. Biochem. Syst. Ecol. 8: 153-160.
Harvey, D. M. R. 1986. Applications of X-ray microanalysis in botanical research. Scanning Electron Microsc. 1986/3: 953-973.
Hayward, D. M. & D. W. Parry. 1980. Scanning electron microscopy of silica deposits in the culms, floral bracts and awns of barley (Hordeum sativum Jess.). Ann. Bot. 46: 541-548.
Heiberg, P. 1867-1868. Morphologisk-anatomisk beskrivelse of Eleocharis palustris. Bot. Tidsskr. 2: 157-225.
Hering, L. 1900. Zur Anatomie der monopodialen Orchideen. Bot. Zentralbl. 84: 1-11, 35-45, 73-81, 113-122, 145-152, 177-184.
Hodson, M. J. & A. Bell. 1986. The mineral relations of the lemma of Phalaris canariensis L., with particular reference to its silicified macrohairs. Israel J. Bot. 35: 241-253.
& D. E. Evans. 1995. Aluminium/silicon interactions in higher plants. J. Exp. Bot. 46: 161- 171.
& A. G Sangster. 1988. Silica deposition in the inflorescence bracts of wheat (Triticum aestivum), I: Scanning electron microscopy and light microscopy. Canad. J. Bot. 66: 829-838.
& . 1989a. Silica deposition in the inflorescence bracts of wheat (Triticum aestivum), II: X-ray microanalysis and backscattered electron imaging. Canad. J. Bot. 67: 281-287.
& . 1989b. X-ray microanalysis of the seminal root of Sorghum bicolor with particular reference to silicon. Ann. Bot. 64: 659-667.
& . 1993. The interaction between silicon and aluminium in Sorghum bicolor (L.) Moench: Growth analysis and X-ray microanalysis. Ann. Bot. 72: 389-400.
9 & D. W. Parry. 1982. Silicon deposition in the inflorescence bristles and macrohairs of Setaria italica (L.) Beauv. Ann. Bot. 50: 843-850.
& . 1984. An ultrastructural study on the development of silicified tissues in the lemma of Phalaris canariensis L. Proc. Roy. Soc. London, B, 222: 413-425.
, S. E. Williams & A. G Sangster. 1997. Silica deposition in the needles of the gymnosperms, 1: Chemical analysis and light microscopy. Pp. 123-133 in A. Pinilla, J. Juan-Tresserras & M. J. Machado (eds.), Estado actual de los estudios de fitolitos en suelos y plantas. Monografias del Centro de Ciencias Medioambientales, 4. Consejo Superior de Investigaciones Cientificas and Centro de Ciencias Medioambientales, Madrid.

SILICA BODIES IN MONOCOTYLEDONS 431
Holtzmeier, M. A., W. L. Stern & W. S. Judd. 1998. Comparative anatomy and systematics of Senghas's cushion species of Maxillaria (Orchidaceae). Bot. J. Linn. Soc. 127: 43-82.
Horrocks, M., Y. Deng, J. Ogden & D. G Sutton. 2000. A reconstruction of the history of a Holocene sand dune on Great Barrier Island, northern New Zealand, using pollen and phytolith analyses. J. Biogeog. 27: 1269-1277.
Hryniewiecki, B. & W. Kurtz. 1936. La repartition des cones siliceux dans les cellules des Cyperacees et leur correlation. Bull. Int. Acad. Pol. Sci. Math. Nat., ser. B1, 33-52.
Huang, F. & M. Zhang. 2000. Pollen and phytolith evidence for rice cultivation during the Neolithic at Longquizhang, eastern Jianghuai, China. Veget. Hist. Archaeobot. 9: 161-168 (English).
Hutton, J. T. & K. Norrish. 1974. Silicon content of wheat husks in relation to water transport. Austral. J. Agric. Res. 25: 203-212.
Iler, R. K. 1955. The colloid chemistry of silica and the silicates. Cornell Univ. Press, Ithaca, NY. Ishizuka, Y. 1971. Physiology of the rice plant. Adv. Agron. 23: 241-315. Jiang, X. M. & Y. Zhou. 1989. SEM observation on crystals and silica in wood species of Chinese
Gymnospermae. Acta Bot. Sin. 31: 835-840 + 1 plate (Chinese; English summary). Johnson, L. A. S. & D. F. Cutler. 1973 [1974]. Empodisma. A new genus of Australasian Restionaceae.
Kew Bull. 28: 381-385. Jones, R. L. & A. H. Beavers. 1963. Some mineralogical and chemical properties of plant opal. Soil Sci.
96: 375-379. Jones, L. H. P. & K. A. Handreck. 1965. Studies of silica in the oat plant, III: Uptake of silica from soils
by the plant. Plant & Soil 23: 79-96. & . 1967. Silica in soils, plants, and animals. Adv. in Agron. 19: 107-149. & . 1969. Uptake of silica by Trifolium incarnatum in relation to the concentration in the
external solution and to transpiration. Plant & Soil 30: 71-80. , A. A. Milne & J. V. Sanders. 1966. Tabashir: An opal of plant origin. Science 151: 464-466.
Juan-Tresserras, J., C. Lalueza, R. Albert & M. Calvo. 1997. Identification of phytoliths from human dental remains from the Iberian Peninsula and the Balearic Islands. Pp. 197-203 in A. Pinilla, J. Juan- Tresserras & M. J. Machado (eds.), Estado actual de los estudios de fitolitos en suelos y plantas. Monografias del Centro de Ciencias Medioambientales, 4. Consejo Superior de Investigaciones Cientificas and Centro de Ciencias Medioambientales, Madrid.
Judd, W. S., W. L. Stern & V. I. Cheadle. 1993. Phylogenetic position of Apostasia and Neuwiedia (Orchidaceae). Bot. J. Linn. Soc. 113: 87-94.
Kajale, M. D. & S. P. Eksambekar. 1997. Application of phytolith analyses to a neolithic site at Budihal, district Gulbarga, South India. Pp. 219-229 in A. Pinilla, J. Juan-Tresserras & M. J. Machado (eds.), Estado actual de los estudios de fitolitos en suelos y plantas. Monografias del Centro de Ciencias Medioambientales, 4. Consejo Superior de Investigaciones Cientificas and Centro de Ciencias Medioambientales, Madrid.
Kaphahn, S. 1904-1905. Beitrage zur Anatomie der Rhynchosporeenblatter und zur Kenntnis der Verkieselungen. Beih. Bot. Zentralbl. 18(1): 233-272.
Kaufman, P. B., L. B. Petering & J. G Smith. 1970. Ultrastructural development of cork-silica cell pairs in Avena internodal epidermis. Bot. Gaz. 131: 173-185.
, W. C. Bigelow, R. Schmid & N. S. Ghosheh. 1971. Electron microprobe analysis of silica in epidermal cells of Equisetum. Amer. J. Bot. 58: 309-316.
, Y. Takeoka, T. J. Carlson, W. C. Bigelow, J. D. Jones, P. H. Moore & N. S. Ghosheh. 1979 [1980]. Studies on silica deposition in sugarcane (Saccharum spp.) using scanning electron micros- copy, energy-dispersive X-ray analysis, neutron activation analysis, and light microscopy. Phytomorphology 29: 185-193.
, P. Dayanandan, Y. Takeoka, W. C. Bigelow, J. D. Jones & R. I1er. 1981. Silica in shoots of higher plants. Pp. 409-449 in T. L. Simpson & B. E. Volcani (eds.), Silicon and siliceous structures in biological systems. Springer-Verlag, New York.
I - , C. I. Franklin & Y. Takeoka. 1985. Structure and function of silica bodies in the epidermal system of grass shoots. Ann. Bot. 55: 487-507.
Kaul, R. B. 1972. Adaptive leaf architecture in emergent and floating Sparganium. Amer. J. Bot. 59: 270-278. Kealhofer, L. & D. R. Piperno. 1998. Opal phytoliths in Southeast Asian flora. Smithsonian Contrib.
Bot. 88: 1-39.

432 THE BOTANICAL REVIEW
Keating, R. C. 2003. Anatomy of the monocotyledons, IX. Acoraceae and Araceae. Clarendon Press, Oxford.
Kerns, B. K. 2001. Diagnostic phytoliths for a ponderosa pine-bunchgrass community near Flagstaff, Arizona. SW Naturalist 46: 282-294.
, M. M. Moore & S. C. Hart. 2001. Estimating forest-grassland dynamics using soil phytolith assemblages and delta C-13 of soil organic matter. Ecoscience 8: 478-488.
Killmann, W. & L. T. Hong. 1992. Some observations on the stegmata of palm trees. Pp. 424-429 in J. P. Rojo, J. U. Aday, E. R. Barile, R. K. Araral & W. M. America (eds.), Proc. 2nd Pacific Regional Wood Anatomy Conf. 1989. For. Prod. Dev. Inst., College, Laguna, Philippines.
Kohl, F. G 1889. Anatomisch-physiologische Untersuchungen der Kieselsaure und Kalksalze in der Pflanze. Marburg.
Kondo, R., C. Childs & I. Atkinson. 1994. Opal phytoliths of New Zealand. Manaaki Whenua Press, Lincoln, New Zealand.
Konstanty, E. C. 1926. Ueber der Entstehung der Kristallzellreihen mit besonderer Beriicksichtigung der Drogenpflanzen. Bot. Archiv 15: 131-186.
Koyama, T. 1966. The systematic significance of leaf structure in the Cyperaceae-Mapanieae. Mem. New York Bot. Gard. 15: 136-159.
. 1967. The systematic significance of leaf structure in the tribe Sclerieae (Cyperaceae). Mem. New York Bot. Gard. 16: 46-70.
Kress, W. J., L. M. Prince, W. J. Hahn & E. A. Zimmer. 2001. Unraveling the evolutionary radiation of the families of the Zingiberales using morphological and molecular evidence. Syst. Biol. 50: 926- 944.
Kubitzki, K (ed.). 1998. The families and genera of vascular plants, IV. Flowering plants: Monocotyle- dons: Alismatanae and Commelinanae (except Gramineae). Springer-Verlag, Berlin.
Kukkonen, I. 1967. Vegetative anatomy of Uncinia (Cyperaceae). Ann. Bot., n.s., 31: 523-544. Kurzweil, H., H. P. Linder, W. L. Stern & A. M. Pridgeon. 1995. Comparative vegetative anatomy and
classification of Diseae (Orchidaceae). Bot. J. Linn. Soc. 117: 171-220. Ktister, E. 1897. Ober die anatomischen Charaktere der Chrysobalaneen, insbesondere ihre Kieselablage-
rungen. Bot. Zentralbl. 69: 46-54,97-106, 129-139, 161-169, 193-202,225-234 (also diss., Cassel). Lalueza, C., J. Juan & R. M. Albert. 1996. Phytolith analysis on dental calculus, enamel surface and
burial soil: Information about diet and paleoenvironment. Amer. J. Phys. Anthropol. 101: 101-113. Lanning, F. C. 1960. Nature and distribution of silica in strawberry plants. Proc. Amer. Soc. Hort. Sci.
76: 349-358. & L. N. Eleuterius. 1983. Silica and ash in tissues of some coastal plants. Ann. Bot. 51: 835-
850. & . 1985. Silica and ash in tissues of some plants growing in the coastal areas of Missis-
sippi, U.S.A. Ann. Bot. 56: 157-172. & . 1989. Silica deposition in some C3 and C4 species of grasses, sedges and composites in
the USA. Ann. Bot. 64: 395-410. , T. L. Hopkins & J. C. Loera. 1980. Silica and ash content and depositional patterns in tissues of
mature Zea mays L. plants. Ann. Bot. 45: 549-554. Larcher, W., U. Meindl, E. Ralser & M. Ishikawa. 1991. Persistent supercooling and silica deposition
in cell walls of palm leaves. J. P1. Physiol. 139: 146-154. Laroche, J. 1968. Contribution a l'etude de l'Equisetum arvense L, III: Recherches sur la nature et la
localisation de la silice chez le sporophyte. Rev. Gen. Bot. 75: 65-116. Lawton, J. R. 1980. Observations on the structure of epidermal cells, particularly the cork and silica
cells, from the flowering stem internode of Lolium temulentum L. (Gramineae). Bot. J. Linn. Soc. 80: 161-177.
Le Cohu, M. C. 1973. Examen au microscope electronique a balayage, des cones de silice chez les Cyperacees. Compt. Rend. Hebd. Acad. Sci., Paris, D, 277: 1301-1303.
Le Coq, C., C. Guervin, J. Laroche & D. Robert. 1991. Modalites d'excretion de la silice chez deux Pteridophytes. Bull. Soc. Bot. Fr., 138: Act. Bot. (2), 231-234.
Lewin, J. & B. E. F. Reismann. 1969. Silica and plant growth. Ann. Rev. P1. Physiol. 20: 289-304. Lim, L. L. & B. C. Stone. 1971. Notes on systematic foliar anatomy of the genus Freycinetia (Pandanaceae).
J. Jap. Bot. 46: 207-220.

SILICA BODIES IN MONOCOTYLEDONS 433
Linder, H. P. 1984. A phylogenetic classification of the African Restionaceae. Bothalia 15: 11-76. . 2000. Vicariance, climate change, anatomy and phylogeny of Restionaceae. Bot. J. Linn. Soc.
134: 159-177. , B. G Briggs & L. A. S. Johnson. 1998. Restionaceae. Pp. 425-445 in K. Kubitzki (ed.), The
families and genera of vascular plants, IV. Flowering plants: Monocotyledons: Alismatanae and Commelinanae (except Gramineae). Springer-Verlag, Berlin.
Linsbauer, K. 1911. Zur physiologischen Anatomie der Epidermis und des Durchluftungsapparates der Bromeliaceen. Sitzungsber. Akad. Wiss. Wien 120: 319-348.
L6pez, P. & 0. Matthei. 1995. Micromorfologia del aquenio en especies del genero Cyperus L. (Cyperaceae), Chile. Gayana, Bot. 52: 67-75 (Spanish; English summary).
Lovering, T. S. 1959. Significance of accumulator plants in rock weathering. Bull. Geol. Soc. Amer. 70: 781-800.
Lowary, P. A. & C. J. Avers. 1965. Nucleolar variation during differentiation of Phleum root epidermis. Amer. J. Bot. 52: 199-203.
Lucenlo, M. 1992. Estudios en la seccion Spirostachyae (Drejer) Bailey del genero Carex, I: Revalorizacion de C. helodes Link. Anal. Jard. Bot. Madrid 50: 73-81.
Lutwick, L. E. & A. Johnston. 1969. Cumulic soils of the rough fescue prairie popular transition region. Canad. J. Soil Sci. 49: 199-203.
Madella, M. 1997. Phytoliths from a Central Asia loess-paleosol sequence and modem soils: Their taphronomical and palaeoecological implications. Pp. 49-57 in A. Pinilla, J. Juan-Tresserras & M. J. Machado (eds.), Estado actual de los estudios de fitolitos en suelos y plantas. Monografias del Centro de Ciencias Medioambientales, 4. Consejo Superior de Investigaciones Cientificas and Centro de Ciencias Medioambientales, Madrid.
Mann, S., S. B. Parker, C. C. Perry, M. D. Ross, A. J. Skarnulis & R. J. P. Williams. 1983a. Problems in the understanding of biominerals. Pp. 171-183 in P. Westbroek & E. W. de Jong (eds.), Biomineralisation and biological metal accumulation. D. Reidel Publishing Company.
, C. C. Perry, R. J. P. Williams, C. A. Fyfe, G C. Gobbi & G. J. Kennedy. 1983b. The characterisation of the nature of silica in biological systems. J. Chem. Soc. Chem. Commun. 168-170.
Marumo, Y. & H. Yanai. 1986. Morphological analysis of opal phytoliths for soil discrimination in forensic science investigation. J. Forensic Sci. 31: 1039-1049.
Matsuda, T., H. Kawahara & N. Chonan. 1983. Histological studies on breaking resistance of lower internodes in rice culm, II: Ultrastructural and histochemical observations on the secondary wall formation. Jap. J. Crop Sci. 52: 84-93 (Japanese; English summary).
Mbida Mindzie, C., H. Doutrelepont, L. Vrydaghs, R. L. Swennen, R. J. Swennen, H. Beeckman, E. de Langhe & P. de Maret. 2001. First archaeological evidence of banana cultivation in central Africa during the third millennium before present. Veget. Hist. Archaeobot. 10: 1-6.
McKeague, J. A. & M. G Cline. 1963. Silica in soil solutions, II: The absorption of monosilicic acid by soil and by other substances. Canad. J. Soil Sci. 43: 83-96.
McNaughton, S. J. & J. L. Tarrants. 1983. Grass leaf silicification: Natural selection for an inducible defense against herbivores. Proc. Natl. Acad. U.S.A. 80: 790-791.
1 ,~9 M. M. McNaughton & R. H. Davis. 1985. Silica as a defense against herbivory and a growth promotor in African grasses. Ecology 66: 528-535.
Mehra, P. N. & 0. P. Sharma. 1963. Anatomy of Eleocharis plantaginea R. Br. Res. Bull. Panjab Univ., n.s., 14: 289-305.
& . 1965. Epidermal silica cells in the Cyperaceae. Bot. Gaz. 126: 53-58. Menapace, F. J. 1991. A preliminary micromorphological analysis of Eleocharis (Cyperaceae) achenes
for systematic potential. Canad. J. Bot. 69: 1533-1541. & D. E. Wujek. 1987. The systematic significance of achene micromorphology in Carex retrorsa
(Cyperaceae). Brittonia 39: 278-283. Metcalfe, C. R. 1960. Anatomy of the monocotyledons. I. Gramineae. Clarendon Press, Oxford.
1971. Anatomy of the monocotyledons. V. Cyperaceae. Clarendon Press, Oxford. & L. Chalk. 1983. Anatomy of the dicotyledons. II. Wood structure and conclusion of the general
introduction. Ed. 2. Clarendon Press, Oxford. Mettenius, G H. 1864. Ober die Hymenophyllaceae. Abhandl. Kon. Sachs. Ges. Wiss., Math-Phys. Cl.,
7: 403-504.

434 THE BOTANICAL REVIEW
Meyer, F. J. 1933. Beitrage zur vergleichenden Anatomie der Typhaceen (Gattung Typha). Beih. Bot. Zentralbl. 51(1): 335-376.
Michelangeli, F. A., J. I. Davis & D. W. Stevenson. 2003. Phylogenetic relationships among Poaceae and related families as inferred from morphology, inversions in the plastid genome, and sequence data from the mitochondrial and plastid genomes. Amer. J. Bot. 90: 93-106.
Miller, A. 1980. Phytoliths as indicators of farming techniques. Paper presented at the 45th annual meet- ing of the Society for American Archaeology, Philadelphia.
M6bius, M. 1908a. Uber die Festlegung der Kalksalze und Kieselkorper in der Pflanzenzellen. Ber. Deutsch. Bot. Ges. 26A: 29-37.
. 1908b. Uber ein eigentumliches Vorkommen von Kieselkorpem in der Epidermis und der Bau des Blattes von Callisia repens. Wiesner Festschrift, Vienna.
Molisch, H. 1913. Mikrochemie der Pflanze. G. Fischer, Jena, Germany (Ed. 3, 1923). . 1918. Beitrage zur Mikrochemie der Pflanze, 12 und 13, 12: Uber Riesenkieselkorper im Blatte
von Arundo donax. Ber. Deutsch. Bot. Ges. 36: 474-481. . 1920. Aschenbild und Pflanzenverwandtschaft. Sitzungsber. Akad. Wiss. Wien, Math-Nat. KI. I,
129: 261-294. M0ller, J. D. & H. Rasmussen. 1984. Stegmata in Orchidales: Character state distribution and polarity.
Bot. J. Linn. Soc. 89: 53-76. Molvray, M., P. J. Kores & M. W. Chase. 2000. Polyphyly of mycoheterotrophic orchids and functional
influences on floral and molecular characters. Pp. 441-448 in K. L. Wilson & D. A. Morrison (eds.), Monocots: Systematics and evolution. Vol. 1. CSIRO, Melbourne, Australia.
Montgomery, D. J. & D. W. Parry. 1979. The ultrastructure and analytical microscopy of silicon depo- sition in the intercellular spaces of the roots of Molinia caerulea (L.) Moench. Ann. Bot. 44: 79-84.
Moore, D. 1984. The role of silica in protecting Italian ryegrass (Lolium multiflorum) from attack by dipterous stem-boring larvae (Oscinellafrit and other related species). Ann. Appl. Biol. 104: 161- 166.
Morcote-Rios, G & R. Bernal. 2001. Remains of palms (Palmae) at archaeological sites in the New World: A review. Bot. Rev. 67: 309-350.
Morris, M. W., W. L. Stern & W. S. Judd. 1996. Vegetative anatomy and systematics of subtribe Dendrobiinae (Orchidaceae). Bot. J. Linn. Soc. 120: 89-144.
Muasya, A. M., D. A. Simpson, M. W. Chase & A. Culham. 1998. An assessment of suprageneric phylogeny in Cyperaceae using rbcL DNA sequences. P1. Syst. Evol. 211: 257-271.
, J. J. Bruhl, D. A. Simpson, A. Culham & M. W. Chase. 2000. Suprageneric phylogeny of Cyperaceae: A combined analysis. Pp. 593-601 in K. L. Wilson & D. A. Morrison (eds.), Monocots: Systematics and evolution. Vol. 1. CSIRO, Melbourne, Australia.
Mulholland, S. C. 1989. Phytolith shape frequencies in North Dakota, U.S.A., grasses: A comparison to general patterns. J. Archaeol. Sci. 16: 489-512.
Munro, S. L. & H. P. Linder. 1998. The phylogenetic position of Prionium (Juncaceae) within the order Juncales based on morphological and rbcL sequence data. Syst. Bot. 23: 43-55.
Nanda, H. P. & S. Gangopadhyay. 1984. Role of silicated cells in rice leaf on brown spot disease incidence by Bipolaris oryzae. Int. J. Trop. P1. Dis. 2(2): 89-98.
Netolitzky, F. 1929. Die Kieselkorper. Die Kalksalze als Zellinhaltskorper by F. Netolitzky. Calciumoxalat- monohydrat und trihydrat by A. Frey. Vol. III/la of Handbuch der Pflanzenanatomie. Berlin.
Newman, R. H. & A. L. Mackay. 1983. Silica spicules in canary grass. Ann. Bot. 52: 927-929. Norris, F. M. G 1983. Anatomy of the genus Kyllinga in South Africa. Bothalia 14: 809-817. Norton, B. E. 1967. Occurrence of silica in Lepidosperma limicola Wakefield. Austral. J. Sci. 29: 371-
372. Oh, Y. C. & E. J. Ham. 1998. A taxonomic study on Scirpus Linne (Cyperaceae) of Korea. Korean J. P1.
Taxon. 28: 217-247. & H. J. Lee. 2001. A taxonomic study on section Acutae of Carex L. in Korea (Cyperaceae).
Korean J. P1. Taxon. 31: 183-222 (Korean; English summary). , C. S. Lee & K. J. Ryu. 2001. A taxonomic study on section Atratae of Carex L. in Korea
(Cyperaceae). Korean J. P1. Taxon. 31: 223-251 (Korean; English summary). Okuda, A. & E. Takahashi. 1961. The effect of various amounts of silicon supply on the growth of the
rice plant and nutrient uptake, part 3. J. Sci. Soil Manure, Japan 32: 533-537.

SILICA BODIES IN MONOCOTYLEDONS 435
& . 1964. The role of silicon. Pp. 123-146 in The mineral nutrition of the rice plant: Proceedings of the symposium of the International Rice Research Institute. John Hopkins Press, Baltimore, MD.
Ollendorf, A. L. 1992. Toward a classification scheme of sedge (Cyperaceae) phytoliths. Pp. 91-111 in G. Rapp & S. C. Mulholland (eds.), Phytolith systematics: Emerging issues. Advances in Archaeo- logical and Museum Science, 1. Plenum Press, New York & London.
, S. C. Mulholland & G Rapp. 1987. Phytoliths from some Israeli sedges. Israel J. Bot. 36: 125- 132.
9 & . 1988. Phytolith analysis as a means of plant identification: Arundo donax and Phragmites communis. Ann. Bot. 61: 209-214.
O'Neill, C., Q. Q. Pan, G Clarke, F. S. Liu, G Hodges, M. Ge, P. Jordan, Y. M. Chang, R. Newman & E. Toulson. 1982. Silica fragments from millet bran in mucosa surrounding oesophageal tumours in patients in northern China. Lancet 82-83: 1202-1206.
, P. Jordan, T. Bhatt & R. Newman. 1986. Silica and oesophageal cancer. CIBA Foundation Symposia 121: 214-230.
Palmer, P. G & S. Gerbeth Jones. 1986. A scanning electron microscope survey of the epidermis of East African grasses, IV. Smithsonian Contrib. Bot. 62: 1-120.
& . 1988. A scanning electron microscope survey of the epidermis of East African grasses, V, and West African supplement. Smithsonian Contrib. Bot. 67: 1-157.
& A. E. Tucker. 1981. A scanning electron microscope survey of the epidermis of East African grasses, I. Smithsonian Contrib. Bot. 49: 1-84.
& . 1983. A scanning electron microscope survey of the epidermis of East African grasses, II. Smithsonian Contrib. Bot. 53: 1-72.
S. Gerbeth Jones & S. Hutchison. 1985. A scanning electron microscope survey of the epider- mis of East African grasses, III. Smithsonian Contrib. Bot. 55: 1-136.
Parr, J. F., V. Dolic, G Lancaster & W. E. Boyd. 2001. A microwave digestion method for the extraction of phytoliths from herbarium species. Rev. Palaeobot. Palynol. 116: 203-212.
Parry, D. W. & M. J. Hodson. 1982. Silica distribution in the caryopsis and inflorescence bracts of foxtail millet [Setaria italica (L.) Beauv.] and its possible significance in carcinogenesis. Ann. Bot. 49: 531-540.
& F. Smithson. 1958. Techniques for studying opaline silica in grass leaves. Ann. Bot., n.s., 22: 543-549.
& A. Winslow. 1977. Electron-probe microanalysis of silicon accumulation in the leaves and tendrils of Pisum sativum (L.) following root severance. Ann. Bot. 41: 275-278.
, M. J. Hodson & A. G Sangster. 1984. Some recent advances in studies of silicon in higher plants. Phil. Trans. Roy. Soc. London, B, 304: 537-549.
, C. O'Neill & M. J. Hodson. 1986. Opaline silica deposits in the leaves of Bidens pilosa L. and their possible significance in cancer. Ann. Bot. 58: 641-647.
Pearsall, D. M. 1989. Paleoethnobotany. A handbook of procedures. Academic Press, San Diego. & D. R. Piperno. 1990. Antiquity of maize cultivation in Ecuador: Summary and reevaluation of
the evidence. Amer. Antiq. 55: 324-337. I ,~q E. H. Dinan, M. Umlauf, Z. Zhao & R. A. Benfer. 1995. Distinguishing rice (Oryza
sativa Poaceae) from wild Oryza species through phytolith analysis: Results of preliminary research. Econ. Bot. 49: 183-196.
Peisl, P. 1957. Die Binsenform. Ber. Schweiz. Bot. Ges. 67: 99-213. Perry, C. C., S. Mann & R. J. P. Williams. 1984a. Structural and analytical studies of the silicified
macrohairs from the lemma of the grass Phalaris canariensis L. Proc. Roy. Soc. London, B, 222: 427-438.
9 ,~9 , F. Watt, G W. Grime & J. Takacs. 1984b. A scanning proton microprobe study of macrohairs from the lemma of the grass Phalaris canariensis L. Proc. Roy. Soc. London, B, 222: 439-445.
Petersen, 0. G 1893. Bidrag til Scitamineernes anatomi. K. Danske Vidensk. Selsk. Skr. (6)8: 337-418. Pfeiffer, H. 1920a. Zur Systematik der Gattung Chrysithrix L. und anderer Chrysithrichinae. Ber. Deutsch.
Bot. Ges. 38: 6-10. . 1920b. Revision der Gattung Ficinia Schrad. Bremen.

436 THE BOTANICAL REVIEW
. 1921 a. Beitrage zur Morphologie und Systematik der Gattungen Lagenocarpus und Cryptangium I. Ber. Deutsch. Bot. Ges. 39: 125-134.
. 192 lb. Der heutige Stand unsere Kenntnisse von den Kegelzellen der Cyperaceen. Ber. Deutsch. Bot. Ges. 39: 353-364.
. 1921c. Die Kegelzellen innerhalb der Gefassbundelscheide bei Cladium mariscus R. Br. Beih. Bot. Zentralbl. 38(1): 401-404.
. 1922. Vergleichende Anatomie der Blatter der Lagenocarpus-Arten. Beih. Bot. Zentralbl. 39(2): 436-445.
. 1925. Aus der Entwicklungsgeschichte der Kegelzellen der Cyperaceen. Ber. Deutsch. Bot. Ges. 43: [26]-[32].
. 1927. Untersuchungen zur vergleichenden Anatomie der Cyperaceen, I: Die Anatomie der Bliitter. Beih. Bot. Zentralbl. 44(1): 90-176.
Pfister, R. 1892. Beitrag zur vergleichenden Anatomie der Sabaleen-Blatter. Diss., Zurich. Pfitzer, E. 1877. Beobachtungen uber Bau und Entwicklung epiphytischer Orchideen, III: Uber das
Vorkommen von Kieselscheiben bei den Orchideen. Flora 60: 245-248. Pinilla, A., J. Juan-Tresserras & M. J. Machado (eds.). 1997. Estado actual de los estudios de fitolitos
en suelos y plantas. Monografias del Centro de Ciencias Medioambientales, 4 Consejo Superior de Investigaciones Cientificas and Centro de Ciencias Medioambientales, Madrid.
Piperno, D. R. 1985. Phytolith analysis and tropical paleo-ecology: Production and taxonomic signifi- cance of siliceous forms in New World plant domesticates and wild species. Rev. Palaeobot. Palynol. 45: 185-228.
. 1988. Phytolith analysis: An archaeological and geological perspective. Academic Press, San Diego.
. 1989. The occurrence of phytoliths in the reproductive structures of selected tropical angiosperms and their significance in tropical paleoecology, paleoethnobotany and systematics. Rev. Palaeobot. Palynol. 61: 147-173.
& K. V. Flannery. 2001. The earliest archaeological maize (Zea mays L.) from highland Mexico: New accelerator mass spectrometry dates and their implications. Proc. Natl. Acad. U.S.A. 98: 2101-2103.
& D. M. Pearsall. 1993. Phytoliths in the reproductive structures of maize and teosinte: Implica- tions for the study of maize evolution. J. Archaeol. Sci. 20: 337-362.
& . 1998. The silica bodies of tropical American grasses: Morphology, taxonomy, and impli- cations for grass systematics and fossil phytolith identification. Smithsonian Contrib. Bot. 85: 1-40.
Prat, H. 1931. L'Epiderme des Graminees: Etude anatomique et systematique. Thesis, Paris. Pridgeon, A. M. 1994. Systematic leaf anatomy of Caladeniinae (Orchidaceae). Bot. J. Linn. Soc. 114:
31-48. & W. L. Stern. 1982. Vegetative anatomy of Myoxanthus (Orchidaceae). Selbyana 7: 55-63. P. J. Cribb, M. W. Chase & F. N. Rasmussen. 2001. Genera Orchidacearum. 2. Orchidoideae
(part 1). Oxford Univ. Press, Oxford. Prychid, C. J. & P. J. Rudall. 1999. Calcium oxalate crystals in Monocotyledons: A review of their
structure and systematics. Ann. Bot. 84: 725-739. & . 2000. Distribution of calcium oxalate crystals in monocotyledons. Pp. 159-162 in
K. L. Wilson & D. A. Morrison (eds.), Monocots: Systematics and evolution. Vol. 1. CSIRO, Melbourne, Australia.
, C. A. Furness & P. J. Rudall. 2003. Systematic significance of cell inclusions in Haemodoraceae and allied families: Silica bodies and tapetal raphides. Ann. Bot. 92: 571-580.
Puech, P.-F., C. Serratrice & F. F. Leek. 1983. Tooth wear as observed in ancient Egyptian skulls. J. Human Evol. 12: 617-629.
Quekett, J. 1852. Lectures on histology. Bailliere, London. Ragonese, A. M., E. R. Guaglianone & C. Dizeo de Strittmatter. 1984. Desarrollo del pericarpio con
cuerpos de silice de dos especies de Rhynchospora Vahl (Cyperaceae). Darwiniana 25: 27-41 (En- glish summary).
Rapp, G & S. C. Mulholland (eds.). 1992. Phytolith systematics: Emerging issues. Advances in Ar- chaeological and Museum Science, 1. Plenum Press, New York & London.
Rasmussen, H. 1986. An aspect of orchid anatomy and adaptationism. Lindleyana 1: 102-107. Raven, J. A. 1983. The transport and function of silicon in plants. Biol. Rev. 58: 179-207.

SILICA BODIES IN MONOCOTYLEDONS 437
Rikli, M. 1895. Beitrage zur vergleichenden Anatomie der Cyperaceen mit besonderer Beriicksichtigung der inneren Parenchymscheide. Jahrb. Wiss. Bot. 27: 485-580.
Riquier, G 1960. Les phytoliths de certains sols tropicaux et des podzals. Trans. Int. Congr. Soil Sci. 4: 425-431.
Rolleri, C., A. M. Deferrari & M. de las M. Ciciarelli. 1987. Epidermis y estomatogenesis en Marattia- ceae (Marattiales-Eusporangiopsida). Revista Mus. La Plata, n.s., 14, Bot. 94: 129-147.
Rosanoff, S. 1871. Ober Kieselsaureablagerungen in einigen Pflanzen. Bot. Ztg. 29: 749-753, 765-769. Rosen, A. M. 1992. Preliminary identification of silica skeletons from Near Eastern archaeological sites:
An anatomical approach. Pp. 129-147 in G. Rapp & S. C. Mulholland (eds.), Phytolith systematics: Emerging issues. Advances in Archaeological and Museum Science, 1. Plenum Press, New York & London.
Rothbuhr, L. & F. Scott. 1957. A study of the uptake of silicon and phosphorus by wheat plants, with radiochemical methods. Biochem. J. 65: 241-245.
Rovner, I. (ed.). 1986. Plant opal phytolith analysis in archaeology and paleoecology: Proceedings of the 1984 Phytolith Research Workshop, North Carolina State University, Raleigh, North Carolina. Oc- casional Paper of the Phytolitharien, 1. North Carolina State Univ., Raleigh.
& J. C. Russ. 1992. Darwin and design in phytolith systematics: Morphometric methods for miti- gating redundancy. Pp. 253-276 in G. Rapp & S. C. Mulholland (eds.), Phytolith systematics: Emerg- ing issues. Advances in Archaeological and Museum Science, 1. Plenum Press, New York & London.
Rudall, P. J. 1994. Anatomy and systematics of Iridaceae. Bot. J. Linn. Soc. 114: 1-21. . 2000. 'Cryptic' characters in monocotyledons: Homology and coding: Revisiting old characters
in the light of new data and new phylogenies. Pp. 114-123 in R. Scotland & T. Pennington (eds.), Homology and Systematics. Taylor & Francis, London & New York.
& L. R. Caddick. 1994. Investigation of the presence of phenolic compounds in monocotyledon- ous cell walls, using UV fluorescence microscopy. Ann. Bot. 74: 483-491.
& M. W. Chase. 1996. Systematics of Xanthorrhoeaceae sensu lato. Evidence for polyphyly. Telopea 6: 629-647.
, C. A. Furness, M. W. Chase & M. F. Fay. 1997. Microsporogenesis and pollen sulcus type in Asparagales (Lilianae). Canad. J. Bot. 75: 408-430.
, D. W. Stevenson & H. P. Linder. 1999. Structure and systematics of Hanguana, a monocotyle- don of uncertain affinity. Austral. Syst. Bot. 12: 311-330.
Runge, F. & J. Runge. 1997. Opal phytoliths in East African plants and soils. Pp. 71-81 in A. Pinilla, J. Juan-Tresserras & M. J. Machado (eds.), Estado actual de los estudios de fitolitos en suelos y plantas. Monografias del Centro de Ciencias Medioambientales, 4. Consejo Superior de Investigaciones Cientificas and Centro de Ciencias Medioambientales, Madrid.
Russ, J. C. & I. Rovner. 1989. Stereological identification of opal phytolith populations from wild and cultivated Zea. Amer. Antiq. 54: 784-792.
Sae-Oui, P., C. Rakdee & P. Thanmathorn. 2002. Use of rice husk ash as a filler in natural rubber vulcanizates: In comparison with other commercial fillers. J. Appl. Polymer Sci. 83: 2485-2493.
Sakai, W. S. & M. Thom. 1979. Localization of silicon in specific cell wall layers of the stomatal appa- ratus of sugarcane by use of energy dispersive X-ray analysis. Ann. Bot. 44: 245-248.
Sangster, A. G 1968. Studies of opaline silica deposits in the leaf of Sieglingia decumbens L. 'Bernh.' using the scanning electron microscope. Ann. Bot. 32: 237-240.
& D. W. Parry. 1971. Silica deposition in the grass leaf in relation to transpiration and the effect of Dinitrophenal. Ann. Bot. 35: 667-677.
& . 1976. The ultrastructure and electron-probe microassay of silica deposits in the endo- dermis of the seminal roots of Sorghum bicolor (L.) Moench. Ann. Bot. 40: 447-459.
, M. J. Hodson & D. W. Parry. 1983. Silicon deposition and anatomical studies in the inflores- cence bracts of four Phalaris species with their possible relevance to carcinogenesis. New Phytol. 93: 105-122.
, S. E. Williams & M. J. Hodson. 1997. Silica deposition in the needles of the Gymnosperms, 2: Scanning electron microscopy and x-ray microanalysis. Pp. 135-145 in A. Pinilla, J. Juan-Tresserras & M. J. Machado (eds.), Estado actual de los estudios de fitolitos en suelos y plantas. Monografias del Centro de Ciencias Medioambientales, 4. Consejo Superior de Investigaciones Cientificas and Centro de Ciencias Medioambientales, Madrid.

438 THE BOTANICAL REVIEW
Sato, Y. I, H. Fujiwara & T. Udatsu. 1990. Morphological differences in silica body derived from motor cell of indica and japonica in rice. Jap. J. Breed. 40: 495-504 (Japanese; English summary).
Schilling, E. 1918. Eigentuimliche Ausgestaltung der Gefassbuindelscheide bei Eleocharis plantaginea. Z. Bot. 10: 512-516.
Schmitt, U., G Weiner & W. Liese. 1995. The fine structure of the stegmata in Calamus axillaris during maturation. IAWA J1 16: 61-68.
Schuyler, A. E. 1971. Scanning electron microscopy of achene epidermis in species of Scirpus (Cyperaceae) and related genera. Proc. Acad. Nat. Sci. Philadelphia 123(2): 29-52.
Schwarz, K. 1973. A bound form of silicon and glycosamino-glycans and polysaccharides matrix/con- nective tissue. Proc. Natl. Acad. U.S.A. 70: 1608-1612.
Seberg, 0. 1988. Leaf anatomy of Oreobolus R.Br. and Schoenoides Seberg (Cyperaceae). Bot. Jahrb. Syst. 110: 187-214.
Sharma, 0. P. & R. Shiam. 1984. Epidermal structures of culm in Cyperus with a discussion of silica bodies in Cyperaceae. Bangladesh J. Bot. 13(1): 16-24.
Siever, R. 1967. The silica budget in the sedimentary cycle. Amer. Minerologist 42: 821-841. Smithson, E. 1956 [1957]. The comparative anatomy of the Flagellariaceae. Kew Bull. 491-501. Solereder, H. & F. J. Meyer. 1928-1933. Systematische Anatomie der Monokotyledonen. Bomtraeger,
Berlin (Heft 1, 155 pp., 1933. Heft 3, 175 pp., 1928. Heft 4, 176 pp., 1929. Heft 6, 242 pp., 1930) Solla, R. F. 1884. Sui cristdlli di silice in serie perifasciali nelle palme. Nota preliminare. Nuovo G. Bot.
Ital. 16: 50-51. Sowers, A. E. & E. L. Thurston. 1979. Ultrastructural evidence for uptake of silicon-containing silicic
acid analogs by Urtica pilulifera and incorporation into cell wall silica. Protoplasma 101: 11-22. Standley, L. A. 1990. Anatomical aspects of the taxonomy of sedges (Carex, Cyperaceae). Canad. J. Bot.
68: 1449-1456. Stant, M. Y. 1973. Scanning electron microscopy of silica bodies and other epidermal features in Gibasis
(Tradescantia) leaf. Bot. J. Linn. Soc. 66: 233-244. Starr, J. R. & B. A. Ford. 2001. The taxonomic and phylogenetic utility of vegetative anatomy and fruit
epidermal silica bodies in Carex section Phyllostachys (Cyperaceae). Canad. J. Bot. 79: 362-379. , S. A. Harris & D. A. Simpson. 2003. The relevance of fruit epidermal silica body variation in Uncinia
Pers. (tribe Cariceae) to taxonomic and phylogenetic studies in the Cyperaceae. Abstracts, Monocots III. Stebbins, G L. 1956. Cytogenetics and evolution of the grass family. Amer. J. Bot. 43: 890-905. Sterling, C. 1967. Crystalline silica in plants. Amer. J. Bot. 54: 840-844. Stern, W. L. 1997a. Vegetative anatomy of subtribe Orchidinae (Orchidaceae). Bot. J. Linn. Soc. 124:
121-136. 1997b. Vegetative anatomy of subtribe Habenariinae (Orchidaceae). Bot. J. Linn. Soc. 125: 211-227. 1999. Comparative vegetative anatomy of two saprophytic orchids from tropical America:
Wullschlaegelia and Uleiorchis. Lindleyana 14: 136-146. & W. S. Judd. 1999. Comparative vegetative anatomy and systematics of Vanilla (Orchidaceae).
Bot. J. Linn. Soc. 131: 353-382. & . 2000. Comparative anatomy and systematics of the orchid tribe Vanilleae excluding
Vanilla. Bot. J. Linn. Soc. 134: 179-202. & . 2001. Comparative anatomy and systematics of Catasetinae (Orchidaceae). Bot. J. Linn.
Soc. 136: 153-178. & . 2002. Systematic and comparative anatomy of Cymbidieae (Orchidaceae). Bot. J. Linn.
Soc. 139: 1-27. & M. W. Morris. 1992. Vegetative anatomy of Stanhopea (Orchidaceae) with special reference
to pseudobulb water-storage cells. Lindleyana 7: 34-53. & W. M. Whitten. 1999. Comparative vegetative anatomy of Stanhopeinae (Orchidaceae). Bot.
J. Linn. Soc. 129: 87-103. , V. I. Cheadle & J. Thorsch. 1993a. Apostasiads, systematic anatomy, and the origins of
Orchidaceae. Bot. J. Linn. Soc. 111: 411-455. , M. W. Morris, W. S. Judd, A. M. Pridgeon & R. L. Dressier. 1993b. Comparative vegetative
anatomy and systematics of Spiranthoideae (Orchidaceae). Bot. J. Linn. Soc. 113: 161-197. Stevenson, D. W., J. I. Davis, J. V. Freudenstein, C. R. Hardy, M. P. Simmonds & C. D. Specht. 2000.
A phylogenetic analysis of the monocotyledons based on morphological and molecular character

SILICA BODIES IN MONOCOTYLEDONS 439
sets, with comments on the placement of Acorus and Hydatellaceae. Pp. 17-24 in K. L. Wilson & D. A. Morrison (eds.), Monocots: Systematics and evolution. Vol. 1. CSIRO, Melbourne, Australia.
Stewart, D. R. M. 1965. The epidermal characters of grasses, with special reference to East African plains species. Bot. Jahrb. Syst. 84: 63-116, 117-174.
Stromberg, C. A. E. 2002. The origin and spread of grass-dominated ecosystems in the late Tertiary of North America: Preliminary results concerning the evolution of hypsodonty. Palaeogeog. Palaeoclim. Palaeoecol. 177: 59-75.
Struve, G A. 1835. De silicia in plantis nonnullis. Diss., Berolini. Suwanprateeb, J. & K. Hatthapanit. 2002. Rice-husk-ash-based silica as a filler for embedding com-
posites in electronic devices. J. Appl. Polymer Sci. 86: 3013-3020. Suzuki, H. 1937. Studies on the relation between the anatomical characters of the rice plant and its
susceptibility to blast disease. J. Coll. Agric. Tokyo Univ. 14: 181-264. Takahashi, E. & Y. Miyake. 1977. Silicon and plant growth: Proceedings of the International Seminar on
Soil Environment and Fertility Management in Intensive Agriculture. Takeoka, Y., P. B. Kaufman & 0. Matsumura. 1979 [1980]. Comparative microscopy of idioblasts in lemma
epidermis of some C3 and C grasses (Poaceae) using SUMP method. Phytomorphology 29: 330-337. Tallent, R. C. & D. E. Wujek. 1983. Scanning electron microscopy an aid to taxonomy of sedges
(Cyperaceae: Carex). Micron Microsc. Acta 14: 271-272. Tillich, H.-J. & E. Sill. 1999. Morphologische und anatomische Studien an Hanguana Blume (Hangua-
naceae) und Flagellaria L. (Flagellariaceae), mit der Beschreibung einer neuen Art, Hanguana bogneri spec. nov. Sendtnera 6: 215-238 (English summary).
Toivonen, H. & T. Timonen. 1976. Perigynium and achene epidermis in some species of Carex, subg. Vignea (Cyperaceae), studied by scanning electron microscopy. Ann. Bot. Fenn. 13: 49-59.
Tomlinson, P. B. 1956. Studies in the systematic anatomy of the Zingiberaceae. J. Linn. Soc., Bot. 55: 547-592.
1959. An anatomical approach to the classification of the Musaceae. J. Linn. Soc., Bot. 55: 779-809. 1960. The anatomy of Phenakospermum (Musaceae). J. Arnold Arbor. 41: 287-297. 1961a. The anatomy of Canna. J. Linn. Soc., Bot. 56: 467-473. 1961 b. Anatomy of the monocotyledons. II. Palmae. Clarendon Press, Oxford. 1961c. Morphological and anatomical characteristics of the Marantaceae. J. Linn. Soc., Bot. 58:
55-78. . 1962. Phylogeny of the Scitamineae-Morphological and anatomical considerations. Evolution
16: 192-213. . 1965. Notes on the anatomy of Aphyllanthes (Liliaceae) and comparison with Eriocaulaceae.
J. Linn. Soc., Bot. 59: 163-173. 1966. Anatomical data in the classification of Commelinaceae. J. Linn. Soc. 59: 371-395. 1969. Anatomy of the Monocotyledons. III. Commelinales-Zingiberales. Clarendon Press, Oxford. 1974. Development of the stomatal complex as a taxonomic character in the monocotyledons.
Taxon 23: 109-128. Toscano de Brito, A. L. V. 1998. Leaf anatomy of Ornithocephalinae (Orchidaceae) and related subtribes.
Lindleyana 13: 234-258. Tucker, G C. & N. G. Miller. 1990. Achene microstructure in Eriophorum (Cyperaceae): Taxonomic
implications and paleobotanical applications. Bull. Torrey Bot. Club 117: 266-283. TIwiss, P. C., E. Suess & R. M. Smith. 1969. Morphological classification of grass phytoliths. Proc. Soil
Sci. Soc. Amer. 33: 109-115. Udatsu, T. & H. Fujiwara. 1993. Application of the discriminant function to subspecies of rice (Oryza
sativa) using the shape of motor cell silica body. Ethnobotany 5: 107-116. Uhl, N. W. & J. Dransfield. 1987. Genera palmarum: A classification of palms based on the work of
Harold E. Moore, Jr. The L. H. Bailey Hortorium and the International Palm Society. Allen Press, Lawrence, KS.
Umemoto, K. & K. Hozumi. 1971a. Applications of low-temperature ashing with high-frequency oxy- gen plasma in pharmacognostical studies: Observations of silicon bodies in ashed tissues of leaves of Bambusa multiplex Raeuschel and stems of Equisetum hyemale L. and Equisetum ramosissimum Desf. var. japonicum Milde. Yakugaku Zasshi 91: 850-854 (Japanese; English summary). [Biol. Abstr. 853 (1972) No. 44793]

440 THE BOTANICAL REVIEW
& . 197 lb. Applications of low-temperature ashing with high-frequency oxygen plasma in pharmacognostical studies: Method for observation of mineral microstructures. Yakugaku Zasshi 91: 890-895 (Japanese; English summary). [Biol. Abstr. 853 (1972) No. 44797]
, M. Hutoh & K. Hozumi. 1973. Identification of the plant source of the Chinese crude drug "Dan-zhu-ye" using the low-temperature plasma ashing technique. Mikrochim. Acta (2): 301-313.
Van Soest, P. J. & L. H. P. Jones. 1968. Effect of silica in forages upon digestibility. J. Dairy Sci. 51: 1644-1648.
Vicari, M. & D. R. Bazely. 1993. Do grasses fight back? The case for antiherbivore defences. Trends Ecol. Evol. 8 (4): 137-141.
Von Mohl, H. 1861. Uber das Kieselskelett lebender Pflanzenzellen. Bot. Ztg. 19: 209-215, 217-221, 225-231, 305-308.
Wadham, M. D. & D. W. Parry. 1981. The silicon content of Oryza sativa L. and its effect on the grazing behaviour of Agriolimax reticulatus Muller. Ann. Bot. 48: 399-402.
Walter, K. S. 1975. A preliminary study of the achene epidermis of certain Carex (Cyperaceae) using scanning electron microscopy. Michigan Bot. 14: 67-72.
Wang, Y. & H. Lu. 1993. The study of phytolith and its application. China Ocean Press, n.p. Waterway, M. J. 1990. Systematic implications of achene micromorphology in Carex section
Hymenochlaenae (Cyperaceae). Canad. J. Bot. 68: 630-639. Webb, M. A. & H. J. Arnott. 1982. A survey of calcium oxalate crystals and other mineral inclusions in
seeds. Scan. Electron Microscop. 1982(3): 1109-1131. Weiner, G 1992. Zur Stammanatomie der Rattanpalmen. Diss., Hamburg.
& W. Liese. 1990. Rattans-Stem anatomy and taxonomic implications. IAWA Bull., n.s., 11: 61-70. Welle, B. J. H. ter. 1976. Silica grains in woody plants of the Neotropics, especially Surinam. Pp. 107-
142 in P. Baas, A. J. Bolton & D. M. Catling (eds.), Wood structure in biological and technological research. Leiden Botanical Series, 3. Leiden Univ. Press, Leiden, Netherlands.
Whang, S. S., K. Kim & W. M. Hess. 1998. Variation of silica bodies in leaf epidermal long cells within and among seventeen species of Oryza (Poaceae). Amer. J. Bot. 85: 461-466.
Wieler, A. 1893. Ueber das Vorkommen von Verstopfungen in den Gefassen mono- und dicotyler Pflanzen. Medeel. Proefstation Midden-Java 1-41.
. 1897. Beitrage zur Anatomie des Stockes von Saccharum. Beitr. Wiss. Bot. 2: 141. Wiesner, J. 1867. Einleitung in die technische Mikroskopie. Vienna. Wilczek, E. 1892. Beitrage zur Kenntniss des Baues der Frucht und des Samens der Cyperaceen. Diss.,
Zurich (also Bot. Zentralbl. 51: 129-138, 193-201, 225-233, 257-265). Wilding, L. P. & L. R. Drees. 1968. Biogenetic opal in soils as an index of vegetative history in the
Prairie Peninsula. Pp. 99-103 in R. E. Bergstrom (ed.), The Quatemary of Illinois: A symposium in observance of the centennial of the University of Illinois. Special Publ. No. 14. Univ. of Illinois, College of Agriculture, Urbana.
& . 1974. Contributions of forest opal and associated crystalline phases to fine silt and clay fractions of soils. Clays and Clay Minerals 22: 295-306.
, N. E. Smeck & L. R. Drees. 1977. Silica in soils: Quartz, cristobalite, tridymite and opal. Pp. 471-552 in Minerals in soil environments. Soil Science Society of America, Madison.
Wille, F. 1926. Beitrage zur Anatomie des Cyperaceenrhizoms. Beih. Bot. Zentralbl. 43(1): 267-309. Wujek, D. E. & F. J. Menapace. 1986. Taxonomy of Carex Section Folliculatae using achene morphol-
ogy. Rhodora 88: 399-403. Yoshida, S. 1965. Chemical aspects of the role of silicon in physiology of the rice plant. Bull. Natl. Inst.
Agric. Sci., Japan, ser. B, 15: 1-58 (Japanese; English summary). [Biol. Abstr. 48 (1967) No. 66506] , Y. Ohnishi & K. Kitagishi. 1959. Role of silicon in rice nutrition. Soil Sci. P1. Nutr. 9: 49-53. , A. F. Douglas, H. C. James & A. G Kwanchai. 1976. Laboratory manual for physiological
studies of rice. IRRI Press, Los Bafios, Philippines. Yukawa, T. & W. L. Stern. 2002. Comparative vegetative anatomy and systematics of Cymbidium
(Cymbidieae: Orchidaceae). Bot. J. Linn. Soc. 138: 383-419. Zhao, Z., D. M. Pearsall, R. A. Benfer & D. R. Piperno. 1998. Distinguishing rice (Oryza sativa Poaceae)
from wild Oryza species through phytolith analysis, II: Finalized method. Econ. Bot. 52: 134-145. Zornig, H. 1903. Beitrage zur Anatomie der Coelogyninen. Bot. Jahrb. Syst. 33: 618-741 (also diss.,
Heidelberg).


















