
Suppression of c-Fos Protein and mRNA Expression in Pentylenetetrazole-Induced Kindled Mouse Brain...
12
Suppression of c-Fos Protein and mRNA Expression in Pentylenetetrazole-Induced Kindled Mouse Brain by Isoxylitones Shabana Usman Simjee & Farzana Shaheen & M. Iqbal Choudhary & Atta-ur Rahman & Siddiqua Jamall & Syed Uzair Ali Shah & Noureen Khan & Nurul Kabir & Nadeem Ashraf Received: 4 August 2011 /Accepted: 7 November 2011 /Published online: 15 December 2011 # Springer Science+Business Media, LLC 2011 Abstract An early immediate gene c-fos has been proposed as the gene responsible for turning on molecular events that might underlie the long-term neural changes occurring during kindling. We have evaluated the effects of novel anticonvulsant isomeric compounds isoxylitones [(E/Z)-2- propanone-1,3,5,5-trimethyl-2-cyclohexen-1-ylidine] on the c-Fos protein and mRNA expression in the brain samples of kindled mice and compared it with the normal and untreated kindled groups. Kindling was induced in male NMRI mice by repeated administration of sub-convulsive dose (50 mg/kg) of pentylenetetrazole (PTZ) until a seizure score of 4–5 was achieved. The c-Fos expression was quantified by combina- tion of immunohistochemistry and RT-PCR protocols. Both the immunohistochemical and RT-PCR analysis revealed a marked increase in the expression of c-fos mRNA and protein in the brain regions tested in case of PTZ-kindled control group compared to normal control. In contrast, the isoxyli- tone (30 mg/kg)-treated group demonstrated significant reduction of c-Fos expression compared to PTZ-kindled control animals. However, low expression of c-fos mRNA was only detected in the thalamus of the isoxylitone-treated brain samples. Based on these observations, we suggest that isoxylitones may have the capacity to control the seizure pattern by mechanism such as the suppression of c-Fos protein and mRNA levels in different regions of the brain. Further investigations to explore the mechanism of action of these compounds are under process. Keywords Epilepsy . Kindling . Isoxylitones . Delphinium denudatum . c-fos mRNA . c-Fos protein Abbreviations BLA Basolateral amygdala CeL Lateral division of the central nucleus of amygdala CeM Medial division of the central nucleus of amygdala DLG Dorsal lateral geniculate nucleus of thalamus CMT Centromedial nucleus of thalamus PVT Paraventricular nucleus of thalamus IEGs Immediate early genes IDV Integrated density value PBS Phosphate buffer saline PTZ Pentylenetetrazole RT-PCR Reverse transcriptase polymerase chain reaction ROI Region of interest GAPDH Glyceraldehyde-3-phosphate dehydrogenase GABA Gamma aminobutyric acid S. U. Simjee : F. Shaheen : M. I. Choudhary : A.-u. Rahman : S. U. A. Shah : N. Khan : N. Ashraf H.E.J. Research Institute of Chemistry, International Center for Chemical and Biological Sciences (ICCBS), University of Karachi, Karachi 75270, Pakistan S. U. Simjee (*) : M. I. Choudhary : A.-u. Rahman : N. Kabir Dr. Panjwani Center for Molecular Medicine and Drug Research, ICCBS, University of Karachi, Karachi 75270, Pakistan e-mail: [email protected] S. U. Simjee e-mail: [email protected] S. Jamall Department of Biochemistry, University of Karachi, Karachi 75270, Pakistan J Mol Neurosci (2012) 47:559–570 DOI 10.1007/s12031-011-9674-4
-
Upload
nadeem-ashraf -
Category
Documents
-
view
216 -
download
0
Transcript of Suppression of c-Fos Protein and mRNA Expression in Pentylenetetrazole-Induced Kindled Mouse Brain...

Suppression of c-Fos Protein and mRNA Expressionin Pentylenetetrazole-Induced Kindled Mouse Brainby Isoxylitones
Shabana Usman Simjee & Farzana Shaheen & M. Iqbal Choudhary & Atta-ur Rahman &
Siddiqua Jamall & Syed Uzair Ali Shah & Noureen Khan & Nurul Kabir & Nadeem Ashraf
Received: 4 August 2011 /Accepted: 7 November 2011 /Published online: 15 December 2011# Springer Science+Business Media, LLC 2011
Abstract An early immediate gene c-fos has been proposedas the gene responsible for turning on molecular events thatmight underlie the long-term neural changes occurringduring kindling. We have evaluated the effects of novelanticonvulsant isomeric compounds isoxylitones [(E/Z)-2-propanone-1,3,5,5-trimethyl-2-cyclohexen-1-ylidine] on thec-Fos protein and mRNA expression in the brain samples ofkindled mice and compared it with the normal and untreatedkindled groups. Kindling was induced in male NMRI miceby repeated administration of sub-convulsive dose (50 mg/kg)of pentylenetetrazole (PTZ) until a seizure score of 4–5 wasachieved. The c-Fos expression was quantified by combina-tion of immunohistochemistry and RT-PCR protocols. Boththe immunohistochemical and RT-PCR analysis revealed a
marked increase in the expression of c-fos mRNA and proteinin the brain regions tested in case of PTZ-kindled controlgroup compared to normal control. In contrast, the isoxyli-tone (30 mg/kg)-treated group demonstrated significantreduction of c-Fos expression compared to PTZ-kindledcontrol animals. However, low expression of c-fos mRNAwas only detected in the thalamus of the isoxylitone-treatedbrain samples. Based on these observations, we suggest thatisoxylitones may have the capacity to control the seizurepattern by mechanism such as the suppression of c-Fosprotein and mRNA levels in different regions of the brain.Further investigations to explore the mechanism of action ofthese compounds are under process.
Keywords Epilepsy . Kindling . Isoxylitones .Delphiniumdenudatum . c-fos mRNA . c-Fos protein
AbbreviationsBLA Basolateral amygdalaCeL Lateral division of the central nucleus of amygdalaCeM Medial division of the central nucleus of amygdalaDLG Dorsal lateral geniculate nucleus of thalamusCMT Centromedial nucleus of thalamusPVT Paraventricular nucleus of thalamusIEGs Immediate early genesIDV Integrated density valuePBS Phosphate buffer salinePTZ PentylenetetrazoleRT-PCR Reverse transcriptase polymerase chain reactionROI Region of interestGAPDH Glyceraldehyde-3-phosphate dehydrogenaseGABA Gamma aminobutyric acid
S. U. Simjee : F. Shaheen :M. I. Choudhary :A.-u. Rahman :S. U. A. Shah :N. Khan :N. AshrafH.E.J. Research Institute of Chemistry, International Centerfor Chemical and Biological Sciences (ICCBS),University of Karachi,Karachi 75270, Pakistan
S. U. Simjee (*) :M. I. Choudhary :A.-u. Rahman :N. KabirDr. Panjwani Center for Molecular Medicine and Drug Research,ICCBS, University of Karachi,Karachi 75270, Pakistane-mail: [email protected]
S. U. Simjeee-mail: [email protected]
S. JamallDepartment of Biochemistry, University of Karachi,Karachi 75270, Pakistan
J Mol Neurosci (2012) 47:559–570DOI 10.1007/s12031-011-9674-4

Introduction
Kindling is a process in which regular sub-convulsantstimulation of the brain, elicited by chemical convulsant orelectrical current, gradually results in the development ofgeneralized seizure pattern which then leads to tonic–clonicconvulsions (Goddard et al. 1969; Racine 1972; Racine1978). Kindling induces long-lasting changes in neuronalexcitability which becomes almost permanent (Burazin andGundlach 1996; Vourliotis et al. 1998). The neurochemicalchanges that modulate the development of the kindlingprocess still remain unclear; however, changes in thetranscription factors and expression of neuropeptide genesare thought to be involved in the cascade of molecularevents, leading to permanent changes in neuronal activity,associated with kindling/epilepsy. Immediate early genes(IEGs) have been reported to be involved in the neuronalexcitation and are believed to play a role in the develop-ment of the kindling process. The proto-oncogene c-fos isone of the members of IEGs which are present in many tissues(Matsuoka et al. 1997). The basal concentration of c-fos isvery low in brain cells; however, it can be inducedtransiently in response to various stimuli such as seizures(Schreiber et al. 1991). The c-fos gene encodes DNA-binding proteins (c-Fos) that can transactivate other induciblegenes (Kaczmarek et al. 2002; Conversi et al. 2004; Raivichand Behrens 2006; Gass et al. 2004). It has been reportedthat a single administration of a known convulsant pentyle-netetrazole (PTZ) (45 mg/kg) to mice induced c-fosexpression in various brain regions (Dragunow et al. 1988;Sagar et al. 1988). Therefore, it is speculated that theinduction of c-fos expression might play an important role in
the development of seizures, as well as in the excitatoryamino acid induced neurotoxicity (Yoshihisa et al. 2000).Since c-fos expression in neurons is transiently increasedduring seizures, therefore, we have used it as a marker ofneuronal activity in the nervous system during epilepsy.
The isoxylitones ([E/Z]-2-propanone-1,3,5,5-trimethyl-2-cyclohexen-1-ylidine) are novel isomeric compoundscontaining the E/Z isomers in a ratio of 1:2 as determinedby proton NMR spectrum (Fig. 1). These compounds werediscovered and patented by us (US Patent 20080004353)through a bioassay-guided isolation study on a medicinalplant Delphinium denudatum. The extracts prepared fromthe aforementioned plant were studied in acute seizuremodels (Raza et al. 2001a; Raza et al. 2001b; Raza et al.2003). However, the isoxylitones are studied for the firsttime in our group. Based on our pilot studies (unreported)that lead to the finding that isomeric mixture of isoxylitoneshas potent anticonvulsant activity in both the electrical- andchemical-induced seizure models, we decided to examinethe significance of the same in the chemically inducedepileptogenesis process. Correspondingly, we also determinedthe expression of c-Fos protein and c-fos mRNA as a measurefor neuronal activation of various brain regions, i.e.amygdala, cortex, hippocampus and thalamus.
Materials and Methods
scPTZ-Induced Seizure Test and PTZ-Induced Mortality
Anticonvulsant effect of isoxylitones using scPTZ testwas evaluated by administering at least three doses, i.e.
Fig. 1 Structure and proton NMR spectrum of the isomeric mixture of isoxylitones [E/Z]
560 J Mol Neurosci (2012) 47:559–570

15, 20 and 30 mg/kg to groups of six mice at least30 min before subcutaneous administration of convul-sive doses of PTZ (90 mg/kg). After administering PTZ,the mice were isolated and observed for an hour for thepresence or absence of different types of seizurepatterns (Loscher et al. 1991). Protection of testingmaterial against PTZ-induced mortality within 24 was alsoevaluated. In all experiments, diazepam (7.5 mg/kg, i.p.) wasused as positive control.
In Vivo Epileptogenesis Model (scPTZ-Induced Kindlingin Mice)
Kindling was induced according to the modified method ofDe Sarro et al. (2000). Each treatment group consists of 12male NMRI mice ranging from 20–23 g. Except normalcontrol, all other groups were given sub-convulsive dosesof PTZ, i.e. 50 mg/kg subcutaneously once every secondday between 9:30 and 11:00 a.m. The normal control anddrug control groups received daily intraperitoneal dose ofsaline (0.5 mL of 0.9% NaCl) and diazepam (7.5 mg/kg),respectively. Likewise, the test group was administeredwith the isoxylitone mixture (30 mg/kg, i.p.) once daily. Onthe day of PTZ administration, the treatment of saline,diazepam or isoxylitones was given 45–50 min before thePTZ. After each PTZ injection, animals were placed inobservation chambers for 1 h, and behavioral seizureactivity was rated. Animals were scored according to apre-validated scoring scale for the severity of the seizureactivity they show. Seizure patterns during the gradualdevelopment of kindling are classified into five distinctbehavioral stages: stage 0 no response; stage 1 ear andfacial twitching; stage 2 convulsive wave through the body;stage 3 myoclonic jerks; stage 4 clonic–tonic convulsions,turnover into side position; stage 5 generalized clonic–tonicseizures, loss of postural control. The cumulative kindlingscore was then calculated, i.e. average of the individualbehavioral scores and then dividing them with the numberof the animals in order to plot against the number oftreatments. The experiments were terminated once the PTZcontrol group reached the score of 5. The brain sampleswere collected and processed as required.
Transcardiac Perfusion Protocol
Mice were deeply anesthetized and transcardially perfusedwith 50-mL chilled pre-perfusion solution, i.e. phosphatebuffer saline (PBS) containing heparin (for 500 mL of PBS,we added 1 mL of heparin (5000 I.U.)). Brains wereremoved from the cranial cavity and gently rinsed in coldPBS before they were post-fixed in ice cold 2% parafor-maldehyde for 24 h at +4°C and then placed in acryoprotectant, i.e. 30% sucrose in PBS until brain samples
sank at the bottom of the container. The samples were thenstored at −80°C until further processing.
Immunohistochemistry (IHC) for c-Fos Expression
Cryostat sections were prepared by using standard techniques,as described before (Scharfinan et al. 2002; Krsek et al.2004). Using cryostat (Thermo Electron Corporation, UK),frozen brain sections of 30-μm thickness were cut at −20°Cand collected directly on poly-lysine coated slides and storedat −80°C until further processing. The cryosections werekept overnight in a humid chamber at room temperaturecontaining PBS buffer. Care was taken so that the slidehaving cryosections does not come in direct contact withPBS. On the next day, they were re-hydrated by rinsing threetimes (5 min/rinse) with PBS buffer, followed by incubationfor 30 min at 37°C in blocking solution consisting of 2%bovine serum albumin (BSA), 2% normal goat serum and0.1% Tween-20. After this step, the sections were incubatedovernight at 4°C with the c-Fos primary antibody sc-52rabbit polyclonal IgG (Santa Cruz Biotechnology Inc., CA,USA). Following overnight incubation in the primary anti-body, sections were washed three times in PBS and then wereincubated with secondary antibody, i.e. Alexa Fluor 546 goatantirabbit IgG (Invitrogen, Oregon, USA), for 30 min at 37°Cin the dark, followed by a final washing step with PBS.Negative control experiments performed without primary andsecondary antibodies (to rule out the nonspecific tissue bindingof antibodies) exhibited negative results. The stained sectionswere observed under a fluorescent microscope (NikonECLIPSE TE2000-S, Japan). The images were captured andprocessed by using proprietary software supplied by Nikon.
Image Analysis
The schematic diagrams of brain sections adapted from themouse brain atlas (Franklin and Paxinos, 1997) were used asa visual guide for determining the sub-region boundaries.The image processing program ImageJ (National Institutes ofHealth, MD, USA) was used to analyze the images. Thissoftware helps in multiple imaging system data comparisons,taking density (densitometry) in consideration (Collins2007). For each image, background density was determinedand subtracted; the remaining particles were considered torepresent c-Fos expression. Data were obtained from twosections per rat (n=12 animals per group) and presented asmeans±S.E.M. c-Fos immunoreactivity in the amygdala,cortex, dorsal hippocampus and thalamus were centeredapproximately around 3.6 mm posterior to bregma. Withinthe hippocampus region, measurements were performed overthe layer extending from sub-regions CA1–CA3. Within thecortex, a relatively uniform distribution of c-Fos immunore-activity across all of the cortical layers was found.
J Mol Neurosci (2012) 47:559–570 561

Reverse Transcriptase Polymerase Chain Reaction(RT-PCR) Analysis
Region of Interest
The ROI, i.e. BLA, CeL and CeM (basolateral, lateral andmedial division of the central nucleus) of the amygdala,extended region from CA1–CA3 from the hippocampus,cortex and DLG, CMT and PVT (dorsal lateral, centrome-dial nucleus and paraventricular nucleus) of the thalamus,was dissected out from each group of mice (six animalsfrom each group) and used for the PCR analysis.
Total RNA Extraction and Its Quantification
The samples were homogenized by using ULTRA-TURRAX®tissue homogenizer (Netzgerät IKA®-WERKE, Germany).Total RNA was isolated from these regions by usingRNeasy® Mini Kit (Qiagen, Hilden, Germany) according tothe manufacturer's instructions. The isolated RNA samples(1–2 μL) were dissolved in Rnase-free water in a ratio of1:200 and were used to measure the concentration and purityby taking the absorbance at 260 and 280 nm on Uvikonspectrophotometer (Shimadzu, Kyoto, Japan). An absorbanceof 1 unit at 260 nm corresponds to 40 μg of RNA per mL.(A260=1⇒ 40 μg/mL). The concentration of the RNAsample was calculated by multiplying the absorbance at260 nm with the dilution factor and 40 μg/mL of RNA. Next,the total RNA yield was calculated by multiplying theconcentration of the RNA with the volume of the sample inmilliliters. The purity of total RNA was calculated with theratio of A260:A280. The sample was considered pure when anA260/A280 ratio of 2.0±0.15 was obtained; variationsoutside this range were generally considered to be contami-nated. The remaining samples of isolated RNAwere aliquotedinto eppendorf tubes and stored at −80°C for later use.
First Strand cDNA Synthesis
The total RNA of 0.8–1.0 μg was taken from each groupand was reverse-transcribed into first strand cDNA usingSuperScript™ III First Strand Synthesis Kit (Invitrogen,Oregon, USA), using oligo(dT) primers and MMuLV reversetranscriptase according to the manufacturer's instructions. Foreach total RNA sample, a negative control was alsoprepared, in which the reverse transcriptase enzyme wasomitted and rest of the procedure was the same.
Primer/oligonuleotide Design
The sequence for mouse gene specific primers for c-fos wassearched by using the NCBI website (http://www.ncbi.nlm.nih.gov). Once the sequence was found, it was used for
designing the primers using a software program, primer3(V.0.4.0) (http://www.frodo.wi.mit.edu, copyright byWhithead Institute for Biomedical Research). The primerswere synthesized by Bio Basic Inc. (Canada). A pair ofglyceraldehyde-3-phosphate dehydrogenase (GAPDH)primers (Integrated DNA Technology, Inc., USA) wasused as an internal standard. The sequences of eachprimer/oligonuleotide are listed in Table 1.
PCR Amplification of cDNA and Agarose GelElectrophoresis
The thermal cycler was used to amplify the cDNA templatein a reaction mixture containing 1X PCR buffer (containingdithiothreitol, DTT), MgCl2, dNTPs, primers (sense andantisense) and taq DNA polymerase in 0.2-mL thin-walledPCR tubes. Up to 30 cycles were run for each reactionmixture. Optimization of c-fos and GAPDH-specificprimers was performed with separate runs by varyingmagnesium chloride and primer concentrations, amountof cDNA template and annealing temperature for theprimers. The prepared PCR products were analyzed on1% agarose gel electrophoresis. Ethidium bromide (1.5 μL)was added into the gel for the detection of specific bands ofDNA under UV light. DNA standard (GeneRulerTM 100 bpDNA Ladder, Fermentas Life Sciences, Maryland, USA) and10-μL PCR products were loaded in each well. Aftercomplete run, the gel was removed and visualized underUV transluminescence and photographed using Gel-Docsystem (Alpha Innotech, CA, USA). After electrophoresis,the density of each band was quantified. For mRNAanalyses, the density of c-fos product was normalized to thatof GAPDH product. The density of each band was quantifiedby taking a two-dimensional area of each band. Theintegrated density value was obtained through thecorresponding pixel intensity using Spot Density Tools ofthe Gel-Doc system for comparison purposes.
Statistical Analysis
The statistical analysis was performed using StatisticalPackage for the Social Sciences (SPSS). Results are reportedas mean±S.E.M. Data of seizure activity (seizure score) wasanalyzed by nonparametric Mann–WhitneyU tests. The c-fos
Table 1 Sequence of the primers used in the study
Primer/oligonucleotide Sequences
Fos-F 5'-AAGTAGTGCAGCCCGGAGTA-3'
Fos-R 5'-CCAGTCAAGAGCATCAGCAA-3'
GAPDH-F 5'-GGAAAGCTGTGGCGTGATGG-3'
GAPDH-R 5'-GTAGGCCATGAGGTCCACCA-3'
562 J Mol Neurosci (2012) 47:559–570
mRNA level in different treatment groups was analyzed byStudent's t-test and ANOVA (using Bonferron's post-hoctest). The immunostaining data were analyzed by one-wayANOVA with post-hoc Dunnett's multiple comparison test.Sequential differences among means were calculated at thelevel of P<0.05 using SPSS version 10.
Results
scPTZ-Induced Seizure Test and PTZ-Induced Mortality
Dose-related protection from different patterns, such astwitches, body jerks, clonus and generalized seizure ofPTZ-induced seizures in animals, was observed in the
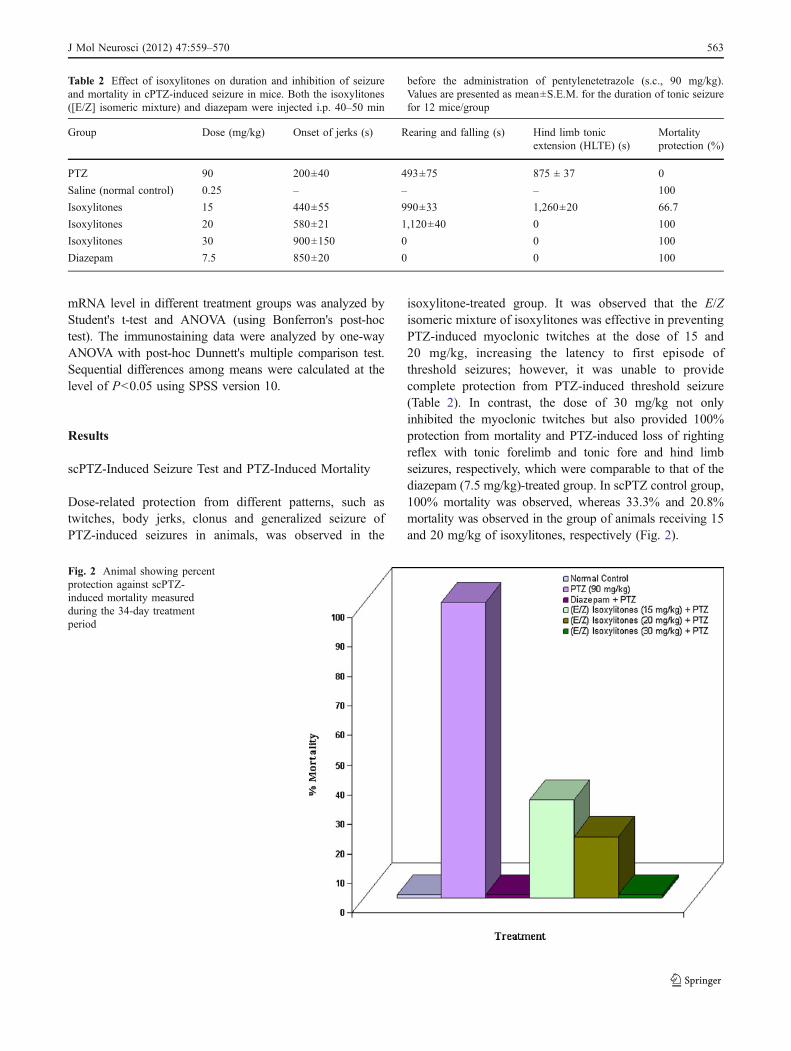
isoxylitone-treated group. It was observed that the E/Zisomeric mixture of isoxylitones was effective in preventingPTZ-induced myoclonic twitches at the dose of 15 and20 mg/kg, increasing the latency to first episode ofthreshold seizures; however, it was unable to providecomplete protection from PTZ-induced threshold seizure(Table 2). In contrast, the dose of 30 mg/kg not onlyinhibited the myoclonic twitches but also provided 100%protection from mortality and PTZ-induced loss of rightingreflex with tonic forelimb and tonic fore and hind limbseizures, respectively, which were comparable to that of thediazepam (7.5 mg/kg)-treated group. In scPTZ control group,100% mortality was observed, whereas 33.3% and 20.8%mortality was observed in the group of animals receiving 15and 20 mg/kg of isoxylitones, respectively (Fig. 2).
Table 2 Effect of isoxylitones on duration and inhibition of seizureand mortality in cPTZ-induced seizure in mice. Both the isoxylitones([E/Z] isomeric mixture) and diazepam were injected i.p. 40–50 min
before the administration of pentylenetetrazole (s.c., 90 mg/kg).Values are presented as mean±S.E.M. for the duration of tonic seizurefor 12 mice/group
Group Dose (mg/kg) Onset of jerks (s) Rearing and falling (s) Hind limb tonicextension (HLTE) (s)
Mortalityprotection (%)
PTZ 90 200±40 493±75 875 ± 37 0
Saline (normal control) 0.25 – – – 100
Isoxylitones 15 440±55 990±33 1,260±20 66.7
Isoxylitones 20 580±21 1,120±40 0 100
Isoxylitones 30 900±150 0 0 100
Diazepam 7.5 850±20 0 0 100
Fig. 2 Animal showing percentprotection against scPTZ-induced mortality measuredduring the 34-day treatmentperiod
J Mol Neurosci (2012) 47:559–570 563

Subcutaneous Pentylenetetrazole (scPTZ)-Induced Kindlingin Mice
The animals in untreated scPTZ control group displayed agradual increase in the seizure score, reaching a score of 5after 18 treatments, with an average seizure score of 4.9.Compared to PTZ-kindled control group, the diazepam-treated group did not exhibit any seizure pattern till the endof the kindling protocol. Based on our earlier results, it wasdecided to use only a 30-mg/kg dose of isoxylitones. At thetested dose, isoxylitones completely retarded the develop-ment of kindling induced by scPTZ administration (Fig. 3).Within the treatment groups, no difference was found in thediazepam- and isoxylitone (E/Z)-treated animals.
c-Fos Immunohistochemistry
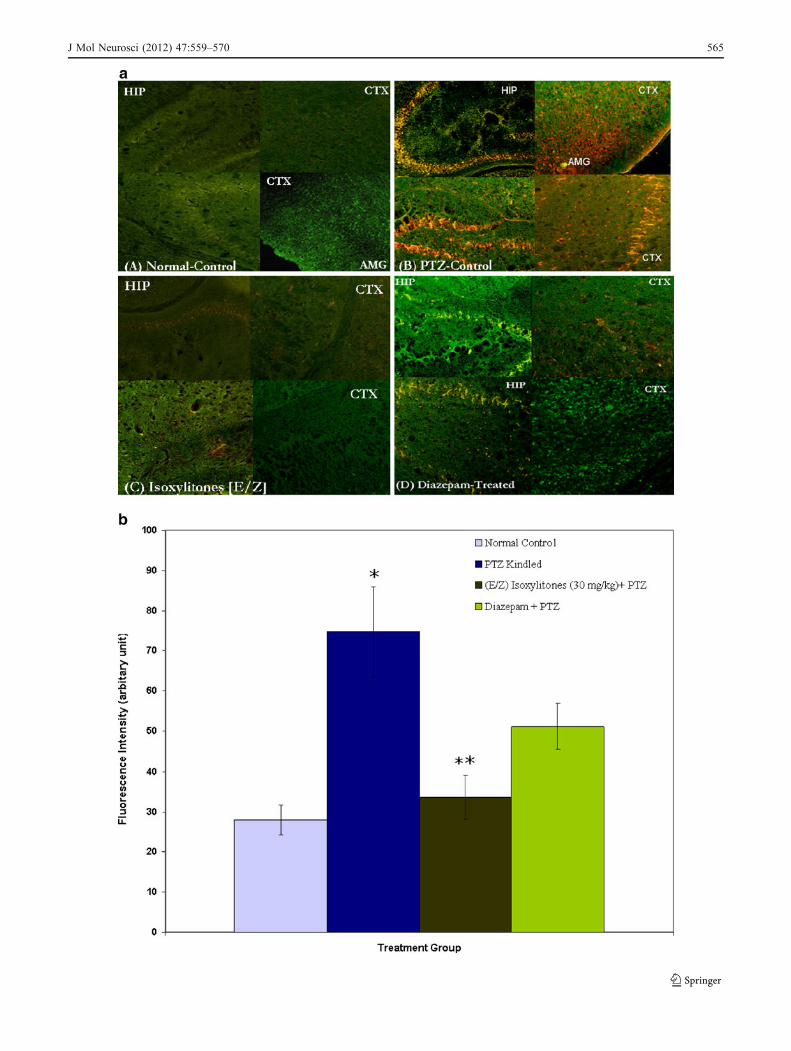
In the control animal without any kindling, there was littleto no immunoreactivity of c-Fos. Immunohistochemicalanalysis revealed a significant increase in the expression ofc-Fos protein in the brain sections of untreated PTZ-kindledmice, which was centered especially in the CA1–CA2hippocampal region (3.6 mm posterior to the bregma) and
thalamus, when compared with the normal brain (Fig. 4a,b). Within the cortex, a relatively uniform distribution of c-Fos across all of the cortical layers was found; however, thecortical sub-regions 1.8 mm posterior to the bregmashowed relatively less c-Fos expression compared to the
Fig. 3 Effect of isoxylitones on scPTZ-induced kindling. Thekindling scores are expressed as the arithmetic mean±S.E.M. of 12animals in each treatment group. The treatment of 30 mg/kg ofisoxylitones did not show any seizure score until the end of theexperiment. The bar under the figure represents the timeline of theexperiments. The rats were treated with PTZ or saline on alternate day.A total of 18 doses were given during the 34-day period when the
animals in the control group were fully kindled. Starting from the fifthinjection of PTZ, the average scores of PTZ-induced seizures weresignificantly higher in the kindled control rats (receiving no othertreatment except PTZ). *p<0.05: significantly higher than normalcontrol revealed by nonparametric Mann–Whitney U tests. Theanimals were sacrificed for immunohistochemistry and RT-PCRanalysis after 34 days
Fig. 4 a Representative photomicrograph of c-Fos immunoreactivity innormal and kindled mice brains with stage 5 seizure activity.Immunohistochemistry was performed in normal or kindled mice withor without treatment with isoxylitones (E/Z). Note the increasedimmunofluorescence in the PTZ-kindled control brain sections (B);AMG, CTX and HIP represents the amygdala, cortex and hippocampus,respectively. The c-Fos positivity was scattered in the entire corticalregion. In contrast, the immunoreactivity of c-Fos in the brain of thekindled animals treated with isoxylitones was markedly reduced (C).The diazepam-treated kindled animals exhibited marked reduction in thelevel of c-Fos expression as compared to PTZ-kindled control group(D); however, the c-Fos positivity was higher than the isoxylitone-treated group. b Graphical representation of the level of expression ofimmunohistochemically detected c-Fos in kindled and unkindled brainsections analyzed by ImageJ software. In comparison to the normalcontrol group (29.47±3.73), the PTZ control animals showed asignificant increase in the c-Fos staining intensity (74.69±17.3, *p<0.024). In contrast, c-Fos expression in the kindled animals treated withisoxylitones (44.94±5.19, **p<0.05) declines significantly
�
564 J Mol Neurosci (2012) 47:559–570
J Mol Neurosci (2012) 47:559–570 565
566 J Mol Neurosci (2012) 47:559–570

cortical sub-regions located in the bregma 3.6 mm. Incontrast, the animals treated with diazepam (7.5 mg/kg, p<0.05) and isoxylitones (30 mg/kg, p<0.03) showed asignificant reduction in the expression of c-Fos protein,which was almost comparable to the normal control group.Interestingly, within the treatment groups, the brain sectionsfrom isoxylitone-treated animals showed far less expressionof c-Fos as compared to the kindled group.
Reverse Transcriptase Polymerase Chain Reaction(RT-PCR) Analysis for c-fos mRNA Expression
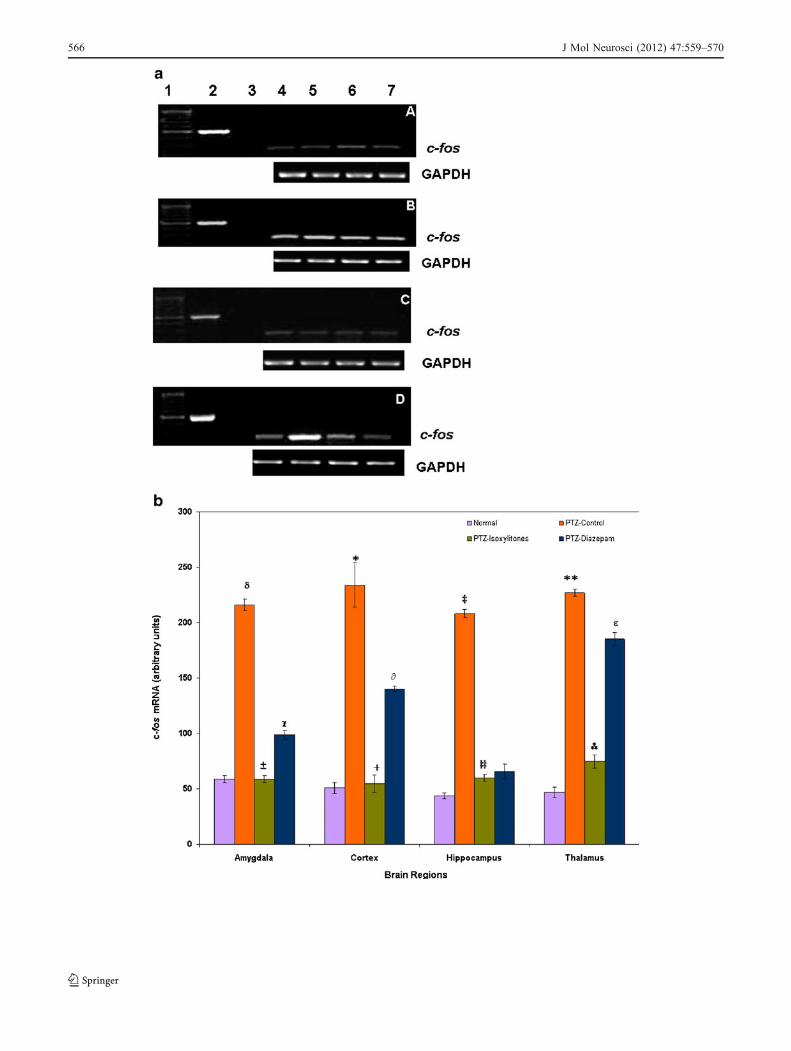
The PTZ-induced kindling caused a marked increase inthe mRNA expression of c-fos. Among the various
regions tested, the highest concentration of c-fos mRNAwas observed in the cortex, followed by the thalamus andamygdala, and finally in the hippocampus (Fig. 5a, b). Thetreatment of isoxylitones significantly inhibited the c-foslevel in the amygdala (p<0.01) and cortex (p<0.005)compared to the PTZ control group; however, in thethalamus, the level of c-fos expression was found to bemarkedly reduced as compared to the control kindledgroup. The diazepam-treated group showed reduced levelsof c-fos mRNA in the cortex, thalamus and amygdalacompared to the PTZ control group; however, whencompared to isoxylitone-treated group, the expression inthese regions was found to be significantly higher (p<0.02, p<0.042, p<0.05 for the cortex, thalamus andamygdala, respectively). In all experiments, the levels ofhousekeeper gene GAPDH mRNAwere equal between thetreated and untreated samples. The PCR products of c-fos(238 bp) and GAPDH (414 bp) were separated andvisualized by electrophoresis on 1% agarose gel containingethidium bromide. The bands were compared to the concur-rent controls, i.e. normal and PTZ-kindled control groups, andthe percent change of each treatment group is shown inTable 3.
Discussion
The (E/Z) isomeric mixture of isoxylitones was identifiedearlier in our laboratory as potent anticonvulsant agents;therefore, it was decided to evaluate its action on theepileptogenesis process in the model of chemical kindlinginduced in mice. Our data demonstrated a potent anti-epileptogenic activity of isoxylitones in scPTZ-inducedkindling model in mice. It was observed that a 30-mg/kgdose of isoxylitones significantly retarded the developmentof seizures as compared to the kindled control group.Pentylenetetrazole (PTZ) most likely produces seizures byinhibiting the gamma aminobutyric acid (GABA) neuro-transmission (De Sarro et al. 2000). Gamma aminobutyricacid (GABA) is a major inhibitory neurotransmitter in the
Fig. 5 a Effects of PTZ and isoxylitones [E/Z] on c-fos mRNA level.Total RNAwas isolated and subjected to RT-PCR analysis, and finally,the PCR products were visualized on 1% agarose gel. Gels A, B, Cand D represent normal, PTZ-kindled control and isoxylitone- (E/Z)and diazepam-treated kindled groups, respectively. Lanes 1–7 repre-sent DNA molecular weight marker, GAPDH (414 bp), RT negativeand expression of c-fos in the hippocampus, thalamus, cortex andamygdala, respectively. The PCR products were quantified bydensitometry. For each treatment group, the house keeping geneGAPDH was also analyzed. The GAPDH (PCR products) of eachregion of the brain from each treatment group is shown below theirrespective gel. The c-fos mRNAs were normalized to their respectiveGAPDH mRNA levels. b Effect of isoxylitones on c-fos mRNAlevels. Following the agarose gel electrophoresis, the bands werequantified by densitometry. Each bar represents the mean±S.E.M.The significantly increased levels of c-fos expression were observed inthe cortex (*p<0.001), thalamus (**p<0.005), amygdala (δp<0.02)and hippocampus (‡p<0.05) of the PTZ-kindled control groupcompared to the normal group. The treatment of isoxylitone givento the PTZ-kindled animals showed a significant reduction in thec-fos expression in all four regions tested (±p<0.01, +p<0.005, ‡‡p<0.01 and p<0.05 for the amygdala, cortex, hippocampus andthalamus, respectively) when compared to the PTZ control group.When the treatment groups were compared with the normal controlgroup, no significant difference was found between the normal andisoxylitones-treated groups; however, a significant increase in the c-fos mRNA was observed in the amygdala (χp<0.05), cortex (∂p<0.01) and thalamus (εp<0.005) region of the diazepam-treated group
�
Table 3 Percent change in the c-fos mRNA expression. FollowingPCR analysis of the bands obtained after agarose gel electrophoresis,both treatment groups, i.e. isoxylitone- [E/Z] and diazepam-treated
kindled groups, were compared with the concurrent controls, i.e.normal and PTZ-kindled control group
Brain regions Amygdala Cortex Hippocampus Thalamus
Treatment Normal PTZ Normal PTZ Normal PTZ Normal PTZ
59 216 51 234 44 208 47 227
% Change from normal or PTZ control
Isoxylitones [E/Z] 0 72.8 (↓) 7.84 (↑) 76.49 (↓) 36.36 (↑) 71.15 (↓) 59.57 (↑) 66.96 (↓)
Diazepam 67 (↑) 54.1 (↓) 174.5 (↑) 40.17 (↓) 50 (↑) 68.26 (↓) 293.61 (↑) 18.50 (↓)
J Mol Neurosci (2012) 47:559–570 567

brain, and the inhibition of GABAergic neurotransmissionhas been thought to be the major underlying factor in thedevelopment of epilepsy (Amabeoku et al. 1998). Theenhancement of the GABAergic neurotransmission hasalready been reported to antagonize seizures, while itsinhibition promotes seizures (Gale 1992; Leonard 2003). Inour study, we have used diazepam (7.5 mg/kg) as areference drug. The protection of mice against PTZ-induced seizures observed in diazepam-treated kindledmice was expected since various authors have shown thatdiazepam exerts its anticonvulsant activity by enhancingGABA-mediated inhibition (Olsen 1981; Amabeoku et al.1998; Leonard 2003; Rang et al. 2003; Shaun et al. 2004).Earlier studies have shown that the systemic administrationof PTZ leads to generalized tonic–clonic seizures andantagonizes the effects of the GABA (Faingold 1987;Bloms-Funke et al. 1996). The suggested reduction ofsynaptic inhibition can contribute to the convulsive effectsof PTZ (MacDonald and Barker 1977; Rehavi et al. 1982;Simmonds 1982; Leweke et al. 1990; Leidenheimer et al.1991; Klee et al. 1992; Maksay et al. 1994). In the light ofthese reports and our observations, i.e. retardation of PTZ-induced kindling by isoxylitones, we suggest that theantiseizure effect of the isoxylitones may be, in part, dueto their ability to attenuate PTZ-induced seizures byinterfering with GABAergic transmission. However, unlikediazepam, we are not yet clear regarding the mechanism ofaction of the isoxylitones. Studies are still ongoing tovalidate the mechanism of action of isoxylitones at thereceptor level.
Among other processes that are thought to contributeto the development of epilepsy, the rapid and transientincrease in the c-fos expression is an extensively reportedphenomenon that underlines the molecular changes occur-ring during the process of epileptogenesis. The expressionof c-fos has been shown to be increased in various brainregions following the administration of PTZ in mice(Morgan et al. 1987; Schreiber et al. 1991; Morgan andCurran 1991; Vourliotis et al. 1998). In agreement withthese reports, our data showed that PTZ (50 mg/kg)increased the c-fos gene level in all four regions studied ofthe mouse brain, with the highest expression found in thethalamus. However, the pre-treatment with isoxylitonescompletely inhibited c-fos level in the amygdala andcortex; however, in the thalamus, the level of c-fosexpression was found to be markedly reduced as com-pared to the control kindled group. The decrease in c-fosgene expression might simply be due to the suppression ofseizures by isoxylitones. The PTZ has been reported toevoke seizures depending on the stimulation of excitatoryamino acid receptors (Velisek et al. 1990; Schröder et al.1993; Schröder et al. 1994; Schroeder et al. 1998), whichalso contribute in the induction of c-fos both in vitro
(Figiel and Kaczmarek 1997; Lauritzen et al. 1997;Griffiths et al. 1997) and in vivo (Berretta et al. 1992;Mitsikostas et al. 1998; Hattori et al. 2001; Hussain et al.2001). Induction of c-fos expression might play animportant role in the development of seizures as well asin the excitatory amino acid induced neurotoxicity(Yoshihisa et al. 2000). Based on these reports and ourdata, we also suggest that the suppression of c-fos geneexpression in the brain by isoxylitones alone mightunderlie its anticonvulsive effects through blocking theexcitatory neurotransmission, although, at this stage, themechanism of these beneficial effects of isoxylitones isstill not clear.
We have also observed that as compared to theisoxylitone treatment, the diazepam treatment did notcompletely inhibit the expression of c-fos in the cortex,thalamus and amygdala. It has been reported that the PTZadministration in the fully kindled animals producedbenzodiazepine-binding decrease in the amygdaloidnuclei and thalamic nuclei, thereby decreasing thebinding capacity of the benzodiazepine (Squires et al.1984; Rocha et al. 1996; Cremer et al. 2009). In light ofthese reports, we hypothesized that the decrease indiazepam binding in these regions might be the reasonwhy it was unable to completely decrease the expressionof the c-fos.
We have also checked the expression of c-fos in acuteseizure model in order to verify whether the c-Fosexpression is altered after the first PTZ treatment or is itrelated with the epileptogenesis process. For this purpose,animals were given acute convulsive dose of PTZ (90 mg/kg) and thereafter evaluated for the seizure pattern. Thesamples collected at the end of these experiments showed aslight increase in the expression of the c-Fos in the PTZcontrol group; however, this increase was neither signifi-cant when compared to the normal control group nor whencompared with the treatment groups, i.e. diazepam- andisoxylitone-treated groups. Therefore, we did not check itfurther at different time frames during the 34-day experi-ments since it would have required more animals to includein the study.
In conclusion, the current study showed that iso-xylitones not only possess powerful antiepileptogenicactivity but they also affect some of the underliningmolecular changes that are induced during the seizures.With the present findings, it can be concluded thatisoxylitones may be useful against the generalizedtonic–clonic. Further studies are required to validatethe proposed mechanism of action at receptor level, i.e.GABA and NMDA receptors which are thought to playa pivotal role in epilepsy. Studies are also ongoing todetermine the refined efficacy of this compound, alongwith toxicity testing on vital organs.
568 J Mol Neurosci (2012) 47:559–570

Acknowledgment This work was supported by grant 20-855/RND/07/540 from the Higher Education Commission, Pakistan, and a grantfrom the Organization for the Prohibition of Chemical Weapons(OPCW; S/228/2000).
References
Amabeoku GJ, Leng MJ, Syce JA (1998) Antimicrobial andanticonvulsant activities of Viscum capense. J Ethnopharmacol61:237–241
Berretta S, Robertson HA, Graybiel AM (1992) Dopamine andglutamate agonists stimulate neuron-specific expression of Fos-like protein in the striatum. J Neurophysiol 68:767–777
Bloms-Funke P, Madeia M, MuBhoff U, Speckmann EJ (1996)Effects of pentylenetetrazol on GABA receptors expressed inoocytes of Xenopus laevis: extra- and intracellular sites of action.Neurosci Lett 205:115–118
Burazin TCD, Gundlach ALo (1996) Rapid and transient increases incellular immediate early gene and neuropeptide mRNAs incortical and limbic areas after amygdaloid kindling seizures inthe rat. Epilepsy Res 26:281–293
Collins TJ (2007) ImageJ for microscopy. Biotechniques 43(1 Suppl):25–30
Conversi D, Orsini C, Cabib S (2004) Distinct patterns of Fosexpression induced by systemic amphetamine in the striatalcomplex of C57BL/JICo and DBA/2JlCo inbred strains of mice.Brain Res 1025(1–2):59–66
Cremer CM, Palomero-Gallagher N, Bidmon H-J, Schleicher A,Speckmann E-J, Zilles K (2009) Pentylenetetrazole-induced sei-zures affect binding site densities for GABA, glutamate andadenosine receptors in the rat brain. Neuroscience 163(1):490–499
De Sarro G, Palma E, Nicola C, Marra R, Gratteri S, De Sarro A,Rotiroti D (2000) Effects of compounds acting on GABABreceptors in the pentylenetetrazole kindling model of epilepsy inmice. J Neuropharmacol 39:2147–2161
Dragunow M, Robertson HA, Robertson GS (1988) Amygdalakindling and c-fos proteins. Exp Neurol 102(2):261–263
Faingold CL (1987) Seizures induced by convulsant drugs. In: JobesPC, Laird HE (eds) Neurotransmitters and epilepsy. HumanaPress, Clifton, pp 215–276
Figiel I, Kaczmarek L (1997) Cellular and molecular correlates ofglutamate-evoked neuronal programmed cell death in the in vitrocultures of rat hippocampal dentate gyrus. Neurochem Int31:229–240
Franklin KBJ, Paxinos G (1997) The mouse brain in stereotaxiccoordinates. San Diego, Academic Press.
Gale K (1992) Role of GABA in the genesis of chemoconvulsantseizures. Toxicol Lett. 64–67 Spec No, 417–428.
Gass P, Fleischmann A, Hvalby O, Jensen V, Zacher C, Strekalova T,Kvello A,Wagner EF, Sprengel R (2004)Mice with a fra-1 knock-ininto the c-fos locus show impaired spatial but regular contextuallearning and normal LTP. Mol Brain Res 130(1–2):16–22
Goddard G, Mclntyre D, Leech C (1969) A permanent change in brainfunction resulting from daily electrical stimulation. Exp Neurol25:295–330
Griffiths R, Malcolm C, Ritchie L, Frandsen A, Schousboe A, ScottM, Rumsby P, Meredith C (1997) Association of c-fos mRNAand excitoxicity in primary cultures of mouse neocortical andcerebellar neurons. J Neurosci Res 15:533–542
Hattori K, Yagi T, Maekawa M, Sato T, Yuasa S (2001) N-methyl-D-aspartate-induced c-Fos expression is enhanced in the forebrainstructures related to emotion in fyn-deficient mice. Brain Res 905(1–2):188–198
Hussain N, Flumerfelt BA, Rajakumar N (2001) Glutamatergicregulation of haloperidol-induced c-fos expression in the ratstriatum and nucleus accumbens. Neuroscience 102(2):391–399
Kaczmarek L, Lapinska-Dzwonek J, Szymczak S (2002) Matrixmetalloproteinases in the adult brain physiology: a link betweenc-Fos, AP-1 and remodeling of neuronal connections? EMBO J21(24):6643–6648
Klee MR, Shirasaki T, Nakaye T, Akaike N, Melikov EN (1992)Interaction of strychnine and bicuculline with GABA- andglycine-induced chloride currents in isolated CAI neurons. In:Epilepsy and inhibition, Speckmann E.-J. and Gutnick M.J., Eds.,Urban und Schwarzenbergz Miinchen, pp. 93–106.
Krsek P, Mikulecka A, Druga R, Kubova H, Hlinak Z, SuchomelovaL, Mares P (2004) Long-term behavioral and morphologicalconsequences of nonconvulsive status epilepticus in rats.Epilepsy Behav 5:180–191
Lauritzen I, De Weille JR, Lazdunski M (1997) The potassiumchannel opener (7)-cromakalim prevents glutamate-induced celldeath in hippocampal neurons. J Neurochem 69:1570–1579
Leidenheimer NJ, Browning MD, Harris RA (1991) GABA Areceptor phosphorylation: multiple sites, actions and artifacts.Trends Pharmacol Sci 12:84–87
Leonard BE (2003) Fundamentals of psychopharmacology, 3rd edn.Chichester, John Wiley and sons, pp 295–318
Leweke FM, Louvel J, Rausche G, Heinemann U (1990) Effects ofpentetrazol on neuronal activity and on extracellular calciumconcentration in rat hippocampal slices. Epilepsy Res 6:187–198
Loscher W, Honack D, Fassbender CP, Nolting B (1991) The role oftechnical, biological and pharmacological factors in the labora-tory evaluation of anticonvulsant drugs. III. Pentylenetetrazoleseizure models. Epi Res 8:171–189
MacDonald RL, Barker JL (1977) Pentylenetetrazol and penicillin areselective antagonists of GABA-mediated post-synaptic inhibitionin cultured mammalian neurons. Nature 267:720–721
Maksay G, Molnar P, Gruber L (1994) Common modes of action ofgamma-butyrolactones and pentylenetetrazol on the GABA Areceptor-ionophore complex. Eur J Pharmacol 288:61–68
Matsuoka M, Matsumura H, Igisu H, Hori H, Tanaka I (1997) Effectsof single exposure to toluene vapor on the expression ofimmediate early genes and GFAP gene in the mouse brain.Arch Toxicol 71:722–723
Mitsikostas DD, Sanchez del Rio M, Waeber C, Moskowitz MA,Cutrer FM (1998) The NMDA receptor antagonist MK-801reduces capsaicin-induced c-fos expression within rat trigeminalnucleus caudalis. Pain 76(1–2):239–248
Morgan JI, Curran T (1991) Proto-oncogene transcription factors andepilepsy. Trends Pharmacol Sci 12(9):343–349
Morgan JI, Cohen DR, Hempstead JL, Curran T (1987) Mappingpattern of c-fos expression in the central nervous system afterseizure. Science 237:192–197
Olsen RW (1981) The GABA postsynaptic membrane receptor-ionophore complex. Site of action of convulsant and anticonvul-sant drugs. Mol Cell Biochem 39:261–279
Racine RJ (1972) Modification of seizure activity by electricalstimulation: II. Motor seizure. Electroencephalogr ClinNeurophysiol 32:281–294
Racine RJ (1978) Kindling: the first decade. Neurosurgery 3(2):234–252Raivich G, Behrens A (2006) Role of the AP-1 transcription
factor c-Jun in developing, adult and injured brain. ProgNeurobiol 78(6):347–363
Rang HP, Dale MM, Ritter JM, Moore PK (2003) Pharmacology, 5thedn. Edinburgh, Churchill Livingstone, pp 550–593
Raza M, Shaheen F, Choudhary MI, Suria A, Rahman AU, Sombati S,DeLorenzo RJ (2001a) Anticonvulsant activities of the FS-1
J Mol Neurosci (2012) 47:559–570 569

subfraction isolated from roots of Delphinium denudatum. PhytoRes 15(5):426–430
Raza M, Shaheen F, Choudhary MI, Sombati S, Rafiq A, Suria A,Rahman A, DeLorenzo RJ (2001b) Anticonvulsant activities ofethanolic extract and aqueous fraction isolated from Delphiniumdenudatum. J Ethnopharmacol 78(1):73–78
Raza M, Shaheen F, Choudhary MI, Rahman AU, Sombati S, Suria A,Rafiq A, DeLorenzo RJ (2003) Anticonvulsant effect of FS-1subfraction isolated from roots of Delphinim denudatum onhippocampal pyramidal neurons. Phyto Res 17(1):38–43
Rehavi M, Skolnick P, Paul SM (1982) Effects of tetrazole derivativeson [3H] diazepam binding in vitro: correlation with convulsantpotency. Eur J Pharmacol 78:353–356
Rocha L, Ackermann RF, Engel J Jr (1996) Chronic and singleadministration of pentylenetetrazol modifies benzodiazepine receptor-binding: an autoradiographic study. Epilepsy Res 24(2):65–72
Sagar SM, Sharp FR, Curran T (1988) Expression of c-fos proteinin brain: metabolic mapping at the cellular level. Science240(4857):1328–1331
Scharfinan HE, Sollas AL, Goodman JH (2002) Spontaneous recurrentseizures after pilocarpine-induced status epilepticus activate calbindin-immunoreactive hilar cells of the rat dentate gyrus. Neurosci 111:71–81
Schreiber SS, Tocco G, Shors TJ, Tompson RF (1991) Activation ofimmediate early genes after acute stress. Neuroreport 2:17–20
Schröder H, Becker A, Lössner B (1993) Glutamate binding to brainmembranes is increased in pentylenetetrazole-kindled rats. JNeurochem 60:1007–1011
Schröder H, Schlichthaar R, Krug M (1994) Changes of glutamatergicsynaptic mechanisms after PTZ kindling and LTP. Neurosci60:337–342
Schroeder H, Becker A, Grecksch G, Schroeder U, Hoellt V (1998)The effect of pentylenetetrazol kindling on synaptic mechanismsof interacting glutamatergic and opioid system in the hippocam-pus of rats. Brain Res 811(1–2):40–46
Shaun FM, Wei-Hua C, Christopher JM (2004) Dorsomedialhypothalamic and brainstem pathways controlling thermogenesisin brown adipose tissue. J Thermal Biol 29(7–8):333–337
Simmonds MA (1982) Classification of some GABA antagonists withregard to site of action and potency in slices of rat cuneatenucleus. Eur J Pharmacol 80:347–358
Squires RF, Saederup E, Crawley JN, Skolnick P, Paul SM (1984)Convulsant potencies of tetrazoles are highly correlated withactions on GABA/benzodiazepine/picrotoxin receptor complexesin brain. Life Sci 35:1439–1444
Velisek L, Kusá R, Kulovaná M, Mares P (1990) Excitatory aminoacid antagonists and pentylenetetrazol-induced seizures duringontogenesis. Life Sci 46:1349–1357
Vourliotis ME, Riechert U, Mayer P, Grecksch G, Höllt V (1998)Pentylenetetrazole (PTZ)-induced c-fos expression in the hippo-campus of kindled rats is suppressed by concomitant treatmentwith naloxone. Brain Res 792:299–308
Yoshihisa I,MasatoM, Hideki I (2000) Suppression of pentylenetetrazole-induced seizures and c-fos expression in mouse brain by L-carnitine.J Occup Health 42:119–123
570 J Mol Neurosci (2012) 47:559–570


















