
Supporting Online Material for -...
35
www.sciencemag.org/cgi/content/full/science.1219723/DC1 Supporting Online Material for MMS19 Assembles Iron-Sulfur Proteins Required for DNA Metabolism and Genomic Integrity Oliver Stehling, Ajay A. Vashisht, Judita Mascarenhas, Zophonias O. Jonsson, Tanu Sharma, Daili J. A. Netz, Antonio J. Pierik, James A. Wohlschlegel,* Roland Lill* *To whom correspondence should be addressed. E-mail: [email protected]; [email protected] Published 7 June 2012 on Science Express DOI: 10.1126/science.1219723 This PDF file includes: Materials and Methods Figs. S1 to S13 Tables S1 to S4 References
Transcript of Supporting Online Material for -...

www.sciencemag.org/cgi/content/full/science.1219723/DC1
Supporting Online Material for
MMS19 Assembles Iron-Sulfur Proteins Required for DNA Metabolism and
Genomic Integrity
Oliver Stehling, Ajay A. Vashisht, Judita Mascarenhas, Zophonias O. Jonsson, Tanu Sharma, Daili J. A. Netz, Antonio J. Pierik, James A. Wohlschlegel,* Roland Lill*
*To whom correspondence should be addressed. E-mail: [email protected]; [email protected]
Published 7 June 2012 on Science Express
DOI: 10.1126/science.1219723
This PDF file includes: Materials and Methods
Figs. S1 to S13
Tables S1 to S4
References

1
Materials and Methods
Yeast strains, cell growth and genetic manipulation Yeast strains used in this study (Table S1) were grown on YP (yeast extract, peptone) rich or
synthetic complete (SC) minimal medium, containing 2% (w/v) of galactose or glucose and supplemented with required auxotrophic markers (39). Galactose-regulatable strains were grown for 40 or 64 h on glucose-containing minimal medium to reduce expression to physiologically
critical levels. Except for the experiments with deletion strains, which were derived from Saccharomyces cerevisiae BY (Open Biosystems), all experiments used S. cerevisiae strain W303-1A as reference wild-type strain. For deletion of MMS19 the coding region was replaced with the S. pombe HIS3 marker generated through a PCR fragment using pFA6a-HIS3MX6 as a template (40).
Regulatable yeast strains in which the levels of Mms19 can be decreased were generated by exchanging the endogenous promoter of MMS19 for the GAL1-10 promoter or for a truncated version thereof (GalL; (41)). Growth of yeast cells in the presence of galactose leads to expression of MMS19, while cultivation in glucose-containing medium depletes the protein strongly (30). Exchange of the endogenous MMS19 for the various Gal promoters by homologous recombination during construction of the GalL-MMS19, GalL-HA-MMS19 or Gal-GFP-MMS19 strains was accomplished by using a cassette amplified from plasmids pYM-N27, pYM-N28 or pYM-N25 as templates, respectively (41). Primers contained 20 nucleotides of the template and nucleotides 113,756-113,806 or 113,808-113,853 of chromosome IX. Correct integration was confirmed by PCR reactions of chromosomal DNA amplifying the TAO3-MMS19 intergenic and NAT-MMS19 (nourseothricin acetyl transferase) region. For exchange of the RAD3 promoter by the GalL promoter during construction of the GalL-HA-RAD3 strain plasmid pYM-N28 was used as a template to amplify the cassette for homologous recombination. Primers contained 20 nucleotides of the template and nucleotides 526,759-526,808 or 527,078-527,127 of chromosome V. Correct integration was confirmed by PCR amplification of chromosomal DNA in the ADK2-RAD3 intergenic and NAT-RAD3 regions.
The yeast RAD3 gene was PCR amplified from chromosomal DNA with primers introducing NheI and NcoI restriction sites, and cloned into the SpeI and NcoI sites of p424-TDH-TAP (C-terminal) (42). Using human cDNA cloned into pCMV-SPORT6 by the IMAGE consortium (MGC 102762), the XPD gene was PCR-amplified with primers introducing SpeI and BglII sites, and cloned into the SpeI and BamHI sites of p424-TDH-TAP. XPD mutagenesis (see Table S2 for plasmids) was carried out with the p424-TDH-XPD-TAP plasmid as template. Primers designed according to (43) introduced restriction sites with silent mutations given by the Watcut tool (http://watcut.uwaterloo.ca/) from Michael Palmer (University of Waterloo, Canada). All plasmid constructs were sequenced. 55Fe incorporation into Fe/S proteins
Appropriate plasmids were transformed into various S. cerevisiae W303 derivatives. After growth of cells on galactose- or glucose-containing SC medium for 24 h (or 48 h with dilution after 24 h), cells were grown in iron-poor (no added iron) SC medium for 16 h. Δyfh1 cells were only grown on glucose. Precipitation of the 55Fe associated with the Fe/S proteins was performed as described earlier using IgG-Sepharose, Protein A-Sepharose (coated with specific antibodies), or anti-HA Agarose (30). Depletion of proteins and immunoprecipitation from cell extracts was analyzed by SDS-PAGE and immunostaining. Monoclonal antibodies against Myc- or HA-

2
epitopes were from Santa Cruz. Polyclonal antibodies against various yeast proteins were raised in rabbits in the Marburg laboratory (44).
DNA damage reporter assays Hyperphosphorylation of the yeast Rad53 protein was used as a marker for DNA damage
(36). Yeast cells were first depleted for various ISC and CIA proteins by growth in glucose-containing SC medium. During the final growth step MMS (methyl methanesulfonate) was added at the indicated concentrations. Cell extracts were prepared under denaturing conditions (36) and separated by 8% acrylamide SDS-PAGE (50 µg protein/lane). After overnight tank blotting onto PVDF membranes phosphorylated Rad53 was detected by specific monoclonal antibodies (F9) generously supplied by Drs. A. Pellicioli and M. Foiani.
Another yeast DNA damage reporter assay employed Photinus pyralis luciferase under the control of well-characterized yeast DNA-damage inducible promoter regions in centromeric plasmids (p414). Promoter regions (750 bp at the 5’ side of the start codon) of the HUG1 and RNR3 genes were amplified from yeast chromosomal DNA and cloned into the SacI/NcoI site of p414-LUX. The plasmids were transformed into wild-type yeast cells and luminescence from luciferase in cell extracts was determined with a Tecan Infinite M200 microtiter plate reader. To the cell extracts (5 µL, 1-2 µg protein) 20 µL firefly luciferase reagent (Promega) was added. The signal response of the p414-HUG1-LUX and p414-RNR3-LUX reporter plasmids was optimal at 0.25 mM MMS. Luciferase activities (arbitrary luminescence intensities per mg soluble protein) were normalized to the response of wild-type cells grown under identical conditions.
UV- and MMS-induced DNA damage was estimated using human HEK293 cells. Stable shRNA expressing cells (see section describing lentiviral stable knockdown cell lines) were induced with doxycycline for 12 days. Equal numbers of cells were plated for each cell line and grown overnight. Medium was then removed and cells were treated with 20 J/m2 of UV light (CL-1000 Ultraviolet Crosslinker from UVP, LLC, Upland, CA) with the lid removed. Fresh media was added to the cells after UV treatment and cells were grown for 7 days before cell viability was measured using the MTS assay (Promega). Similarly, 20 μM MMS (Acros Organics, New Jersey, USA) was added to the different stable knockdown cell lines and cell viability was assessed using MTS assay 7 days later. The MTS assay reagent was made by dissolving CellTiter 96R Aqueous MTS (G1112; Promega) powder at a concentration of 2 mg/mL in 1x PBS containing 300 μM phenazine ethosulfate (PES) (MP Biomedicals). The MTS assay reagent was added to cells at a final concentration of 20%. Cells were incubated for 3 h at 37°C before absorbance of the sample was read at 490 nm. All absorbance values were averaged for control and treated samples.
Small interfering RNAs Silencer® Select siRNAs against human mRNAs of MMS19 and frataxin (Table S3) as well
as scrambled siRNAs (Silencer® select negative controls #1 and #2) were purchased from Ambion. Double-stranded siRNA si#3 (s34551) against MMS19 contained a mismatch (highlighted in Table S3) and thus served as an internal negative control.
Plasmids used in human tissue culture A plasmid containing MMS19 was kindly provided by Lurdes Queimado. The MMS19 ORF
was using primers containing AttB sites with Phusion Taq polymerase (NEB) and subcloned into pDONR221 vector using the Gateway cloning system (Invitrogen). The Gateway cloning system

3
was also used to transfer genes into Gateway-adapted expression plasmids that encode 3xHA-3xFLAG.
Quantitative real-time PCR Total RNA from snap-frozen HeLa cell samples was prepared using the peqGOLD
RNApure reagent (Peqlab). Complementary DNA (cDNA) was amplified by means of the RevertAid™ H Minus First Strand cDNA Synthesis Kit (Fermentas). Quantification of cDNA was performed using a Cepheid Smart Cycler II (Intas, Göttingen, Germany) and the Maxima® SYBR® Green/ROX qPCR Master Mix (Fermentas). MMS19 expression was quantified using the primer pair CTGTGCTGGCTTCTACACCA / ACACAGCACAGCAAG CATTC, while beta-actin expression was determined by the primer pair CTCTTCCAGCCTT CCTTCCT / AGCACTGTGTTGGCGTACAG.
Antibodies and immunoprecipitation MMS19 antiserum was raised in the Lill laboratory (fig. S5B), as well as obtained from
Proteintech Group, Inc. (Chicago, IL, USA; fig. S12). Antibodies against beta-actin, DPYD, RTEL1, XPB, XPD, and c-Myc came from Santa Cruz biotechnology (Santa Cruz, USA). POLD1 antibodies were obtained from Santa Cruz biotechnology or from Proteintech, and FANCJ antibodies from Bethyl Laboratories; ISCU and Fam96B antibodies were purchased from Proteintech; mouse anti-Complex II (30 kDa Fe/S protein subunit SDHB) mab was purchased from MitoSciences; CIAO1 antiserum (UCLA852) was raised in the Wohlschlegel Laboratory; antibodies against NARFL/IOP1, FLAG-M2, and β-tubulin were bought from Sigma; affinity-purified rabbit anti-murine GPAT serum was kindly provided by H. Puccio, Illkirch Cedex, France; monoclonal mouse anti-IRP1 mab (clone 295B) and rabbit anti-IRP2 serum were kindly provided by R. Eisenstein, Wisconsin, USA; rabbit serum against human MIA40 was kindly provided by J. Herrmann, Kaiserslautern, Germany; rabbit anti-mitochondrial aconitase serum was kindly donated by L. Szweda, USA. Peroxidase-conjugated goat anti-rabbit and anti-mouse antibodies were used as secondary reagents upon immunoblotting.
Immunoprecipitation and immunoblotting was performed as described (35). For FLAG immunoprecipitations, affinity matrices for anti-FLAG M2 were used. For endogenous immunoprecipitation, antibodies were pre-bound to equilibrated GammaBindTM Plus SepharoseTM (GE healthcare) for 1 h and then washed three times with lysis buffer to remove unbound antibody. Equal amounts of protein lysate were added to the beads and incubated for 2 h at 4°C. Beads were washed three times with lysis buffer and then suspended in 2x SDS loading buffer. Samples were further analyzed by SDS-PAGE followed by immunoblotting with relevant antibodies.
Tissue culture, cell lines and transfection Human cervix carcinoma cells (HeLa) were maintained in Dulbecco's Modified Eagles
medium (DMEM) containing 4.5 g/L glucose and supplemented with 7.5% fetal calf serum, 1 mM glutamine, and 100 units/mL penicillin and 100 µg/mL streptomycin. Upon transfection by electroporation 4 to 8 × 106 HeLa cells were resuspended in 525 µL transfection buffer (21 mM Hepes, pH 7.4, 137 mM NaCl, 5 mM KCl, 0.7 mM Na2HPO4, and 6 mM dextrose) and supplemented either with a pool or with individual siRNAs amounting to a total of 15 µg RNA.

4
In order to prolong the time period of mRNA depletion cells were retransfected twice at three day intervals. Protein content of harvested cells was determined by the Bradford method (Biorad).
HEK cell lines were cultured in DMEM containing 10% fetal calf serum (Foundation B FBS from Gemini), 100 units/mL penicillin and streptomycin, and 2 mM glutamine at 37°C in ambient air with 5% CO2. HEK293, HEK293T, Flp-InTM T-RExTM-293 cells were obtained from the American Type Culture Collection (ATCC), Thermo Scientific, and Invitrogen, respectively. Flp-InTM T-RExTM-293 cells stably expressing 3xHA-3xFLAG-MMS19 were generated using the Flp-In system (Invitrogen) according to manufacturer’s instructions. Transient transfections of HEK cells were carried out using either BioT (Bioland, Long Beach, CA) or LipofectamineTM 2000 according to the manufacturer’s directions. The expression of 3xHA-3xFLAG-MMS19 in stable Flp-InTM T-RExTM-293 cells was induced by adding doxycycline overnight to a final concentration of 500 ng/mL. Lentiviral stable shRNA cell lines were treated with doxycycline (1μg/mL) for 12 days as indicated. Doxycycline was added to the media every 24 h.
Lentiviral preparation, lentiviral infection and generation of doxycycline-inducible stable knockdown cell lines
TRIPZ lentiviral inducible shRNAmir vectors encoding shRNAs for control (No target), MMS19 (V3THS_32644), FAM96B (V3THS_382805), NARFL/IOP1 (V3THS_344324) and ISCU (V2THS_253498) were purchased from Thermo Scientific and transfected into 293T cells with lentivirus packaging mix according to the manufacturer’s guidelines (Thermo Scientific). Viruses for each target gene knockdown were collected after 48 and 72 h post transfection. HEK293 cells were infected with viruses as described by the manufacturer (Thermo Scientific). Media were exchanged 6-8 h after infection. Stable integrants were selected by the addition of puromycin (1 μg/mL) 48 h after infection for one week while changing media and adding fresh puromycin at 2-3 day intervals. The selected colonies were induced with doxycycline (1 μg/mL) for 12 consecutive days to turn on shRNA expression and attain sustained depletion of the genes of interest. Fresh doxycycline was added to the culture media every 24 h.
Enzyme activity measurements and HeLa cell fractionation The enzyme activity of dihydropyrimidine dehydrogenase (DPYD) was determined by thin
layer chromatography (TLC) using a combination of several published protocols (45-48). In short, a HeLa cell sample corresponding to 125-250 µg protein was applied to 25-50 µL of a reaction mixture containing 25 mM Tris-HCl (pH 7.5), 0.08% digitonin, 2.5 mM MgCl2, 2 mM DTT, 10-30 µM [4-14C]-thymine (59 mCi/mmol, Hartmann Analytic, Germany), up to 5 mM NADPH, as well as 5 mM glucose-6-phosphate (G6P) and 1 U/mL G6P-dehydrogenase to regenerate NADPH. After 4 h of incubation at 32.5°C the reaction was stopped by addition of 10 µL of perchloric acid (10% v/v). Reaction mixtures were centrifuged at 15,000 ×g for 5 min and supernatants were analyzed by TLC. Plastic TLC sheets pre-coated with 0.1 mm cellulose MN 300 (Macherey-Nagel, Germany) were pre-run with the organic phase of a n-butanol:water:ammonium hydroxide (90:45:15, v/v/v) mixture. Six microliters of the final sample were spotted onto a dried TLC plate, separated by three consecutive developing steps with intermittent drying, and analyzed by autoradiography.
Digitonin-based cell fractionation of HeLa cells, as well as measurements of aconitase, succinate dehydrogenase (SDH), lactate dehydrogenase (LDH), and citrate synthase (CS) activities were performed as described (16, 49). Binding of IRP1 to IREs present in ferritin mRNA was examined by REMSA (16, 50).

5
Affinity purification of human MMS19 for proteomic analysis Twelve 15 cm tissue cultures plates of 3xHA-3xFLAG MMS19-expressing Flp-InTM
TRExTM-293 cells were grown, induced with doxycycline overnight, harvested and lysed in IP buffer (100 mM Tris-HCl pH 8.0, 150 mM NaCl, 5 mM EDTA, 5% glycerol, 0.1% NP-40, 1 mM DTT, 0.5 mM PMSF, 1 μM pepstatin, 1 μM leupeptin and 2 μg/mL aprotinin). 200-300 mg of clarified protein lysate was then incubated with 100 μL of equilibrated anti-FLAG M2 agarose beads (Sigma) for 2 h at 4°C. Beads were washed four times with 1 mL of IP buffer per wash before eluting with two sequential steps of 500 μL using FLAG elution buffer (IP buffer without NP-40 supplemented with 250 μg/mL of 3xFLAG peptide (Sigma)). These eluted fractions were pooled and precipitated by the addition of trichloroacetic acid (TCA) to a final concentration of 20% followed by incubation on ice for 60 min and centrifugation at 16,000 xg for 30 min to collect the precipitate. The final precipitate was washed twice with acetone and air dried prior to further analysis.
Identification of human MMS19 associated protein complexes by MudPIT analysis TCA-precipitated samples of affinity-purified 3xHA-3xFLAG-MMS19 were proteolytically
digested and analyzed by MudPIT, a shotgun proteomic approach in which the entire peptide mixture is fractionated online using a combination of strong cation exchange and reversed phase chromatography and eluted directly into the mass spectrometer where MS/MS spectra are collected (51, 52). An online 5-step multidimensional chromatographic separation was performed and fractionated peptides were eluted directly into a LTQ-Orbitrap XL mass spectrometer (Thermo Fisher) where tandem mass spectra were collected. Peptide mass spectra were analyzed using the SEQUEST and DTASelect algorithms (53). False positive rates were estimated using a decoy database approach and results filtered such that only proteins identified by two different peptides using peptide-level false positive rate of 5% were considered present in the sample (54). Proteins were treated as putative MMS19-associated proteins, if they were identified in the relevant affinity purification but not in a previously generated database of proteins that bind non-specifically to FLAG M2 beads. Table S4A contains a curated list of MMS19-interacting proteins identified in this analysis that are known or putative Fe/S proteins or CIA machinery components. Table S4B contains the complete set of putative MMS19-interacting proteins ranked according to their normalized spectral abundance values (NSAFx105) (55). A detailed description of the multidimensional peptide fractionation protocol, mass spectrometer settings, and bioinformatic workflow has been published elsewhere (56).
Statistics All values reported are expressed as the mean ± standard deviation (SD). Pairwise
comparisons of data were calculated according to Student’s T-test, while comparisons of multiple data sets were analyzed by variance analysis (ANOVA) followed by appropriate post-hoc tests.

6
Supplementary Tables
Table S1. Yeast strains used in this study. Strain name Genotype a Source
W303‐1A (wild‐type) MATa ura3-1 ade2-1 trp1-1 his3-11,15 leu2-3,112 (57)
Δmms19 W303-1A; mms19::HIS3 (PCR fragment obtained with pFA6a-HIS3MX6)
This study
Gal‐GFP‐MMS19 W303‐1A pMMS19::natNT2GALGFP (PCR fragment obtained with pYM‐N25)
This study
GalL‐MMS19 W303‐1A pMMS19::natNT2GALL (PCR fragment obtained with pYM‐N27)
This study
GalL‐HA‐MMS19 W303‐1A pMMS19::natNT2GALL3HA (PCR fragment obtained with pYM‐N28)
This study
GalL‐HA‐RAD3 W303‐1A pRAD3::natNT2GALL3HA (PCR fragment obtained with pYM‐N28)
This study
Gal-NFS1 W303‐1A pNFS1::GAL10HIS3 (PCR fragment obtained with pFA6a-HIS3MX6-Gal1-10)
(58)
Gal-YAH1 W303‐1A pYAH1::GAL10LEU2 (HindIII fragment of Yep51))
(59)
Δyfh1 W303-1A; YFH1::kanMX2 (PCR fragment obtained with pFA6‐kanMX2)
(60)
Gal-SSQ1 W303‐1A pSSQ1::GAL110LEU2 (PCR fragment obtained with pTL26)
(61)
GalL-HA-DRE2 W303‐1A pDRE2::natNT2GALL3HA (PCR fragment obtained with pYM‐N28)
(18)
GalL-HA-TAH18 W303‐1A pTAH18::natNT2GALL3HA (PCR fragment obtained with pYM‐N28)
(18)
Gal-CFD1 W303‐1A pCFD1::GAL110HIS3 (PCR fragment obtained with pFA6a-HIS3MX6-Gal1-10)
(19)
Gal-NBP35 W303‐1A pNBP35::GAL110HIS3 (PCR fragment obtained with pFA6a-HIS3MX6-Gal1-10)
(42)
Gal-NAR1 W303‐1A pNAR1::GAL110HIS3 (PCR fragment obtained with pFA6a-HIS3MX6-Gal1-10)
(20)
Gal-CIA1 W303‐1A pCIA1::GAL110HIS3 (PCR fragment obtained with pFA6a-HIS3MX6-Gal1-10)
(19)
BY4742 (wild-type) MATα ; his3Δ 1; leu2Δ 0; lys2Δ 0; ura3Δ 0
(62); Euroscarf
Δmet1 BY4742; YKR069w::kanMX4
(62); Euroscarf
Δmet5 BY4742; YJR137c::kanMX4
(62); Euroscarf
Δmet10 BY4742; YFR030w::kanMX4
(62); Euroscarf
Δmms19 BY4742; YIL128w::kanMX4
(62); Euroscarf
a In parenthesis the plasmids used for the generation of the PCR fragment needed for gene manipulation is provided.

7
Table S2. Yeast plasmids used in this study.
Plasmid (promoter) Encoding protein Tag and position Use 414 (RNR3) Luciferase none Reporter assay
414 (HUG1) Luciferase none Reporter assay
414 (MET25) Rli1 3HA C 55Fe incorporation
416 (MET25) Cia1 3HA C Immunoprecipitation
424 (TDH3)
Rad3 none Immunoprecipitation
Rad3 TAP C 55Fe incorporation
XPD TAP C 55Fe incorporation
XPD C116A TAP C 55Fe incorporation
XPD R118H TAP C 55Fe incorporation
XPD C134A TAP C 55Fe incorporation
XPD C155A TAP C 55Fe incorporation
XPD C190A TAP C 55Fe incorporation
Nbp35 TAP C 55Fe incorporation
Cfd1 TAP C 55Fe incorporation Bio2 none 55Fe incorporation
424 (endogenous promoter) Rli1 3HA C Immunoprecipitation
425 (TDH3) Dna2 3HA C 55Fe incorporation, immunoprecipitation
Cia2 3HA C Immunoprecipitation
426 (TDH3)
Mms19 none Growth complementation
Met10 3HA C 55Fe incorporation,
immunoprecipitation
Ntg2 3HA C 55Fe incorporation
Abbreviations: C, C-terminal. All promoters and proteins are from yeast, except for luciferase
(firefly) and XPD (man).

8
Table S3. siRNA sequences used in this study.
Target siRNA ID siRNA# Sense Antisense
MMS19 s34552 1 AGGCCCUAGUGCUCAGAUAtt UAUCUGAGCACUAGGGCCUtt
s34553 2 GACUCUGAAUGCUUGCUGUtt ACAGCAAGCAUUCAGAGUCtg
s34551 3 CGGGAGUUUUUGGAACUGAtt UCAGUUCCAAAAACUCCCGca
Frataxin S5360 1 GAACCUAUGUGAUCAACAAtt UUGUUGAUCACAUAGGUUCct
S5361 2 AGAGCUCACUAAAGCCUUAtt UAAGGCUUUAGUGAGCUCUgc
S5362 3 GGAGUGGUGUCUUAACUGUtt ACAGUUAAGACACCACUCCca
9
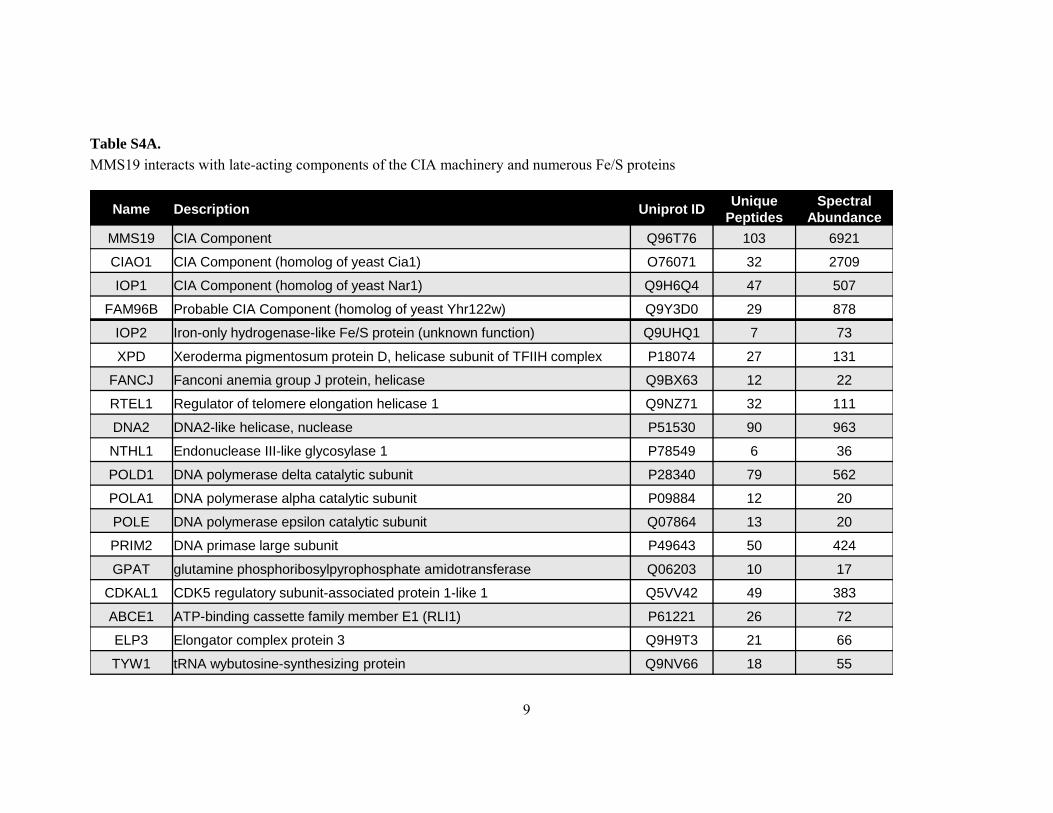
Table S4A. MMS19 interacts with late-acting components of the CIA machinery and numerous Fe/S proteins
Name Description Uniprot ID Unique Peptides
Spectral Abundance
MMS19 CIA Component Q96T76 103 6921
CIAO1 CIA Component (homolog of yeast Cia1) O76071 32 2709
IOP1 CIA Component (homolog of yeast Nar1) Q9H6Q4 47 507
FAM96B Probable CIA Component (homolog of yeast Yhr122w) Q9Y3D0 29 878
IOP2 Iron-only hydrogenase-like Fe/S protein (unknown function) Q9UHQ1 7 73
XPD Xeroderma pigmentosum protein D, helicase subunit of TFIIH complex P18074 27 131
FANCJ Fanconi anemia group J protein, helicase Q9BX63 12 22
RTEL1 Regulator of telomere elongation helicase 1 Q9NZ71 32 111
DNA2 DNA2-like helicase, nuclease P51530 90 963
NTHL1 Endonuclease III-like glycosylase 1 P78549 6 36
POLD1 DNA polymerase delta catalytic subunit P28340 79 562
POLA1 DNA polymerase alpha catalytic subunit P09884 12 20
POLE DNA polymerase epsilon catalytic subunit Q07864 13 20
PRIM2 DNA primase large subunit P49643 50 424
GPAT glutamine phosphoribosylpyrophosphate amidotransferase Q06203 10 17
CDKAL1 CDK5 regulatory subunit-associated protein 1-like 1 Q5VV42 49 383
ABCE1 ATP-binding cassette family member E1 (RLI1) P61221 26 72
ELP3 Elongator complex protein 3 Q9H9T3 21 66
TYW1 tRNA wybutosine-synthesizing protein Q9NV66 18 55

10
Table S4B. MMS19 protein interaction profile (full data set)
Name Description Uniprot ID
Mol. Mass (Da)
pI Spectral Abundance
NSAF x 105
Unique Peptides
% Seq. Cov.
Unique PE SV
CIAO1 CIA Component (homolog of yeast Cia1) O76071 37840 5.0 2709 11336.03 32 58.7 U 1 1 MMS19 CIA Component Q96T76 113289 6.4 6921 9531.98 103 64.1 U 1 2 FAM96B Probable CIA Component (homolog of yeast Yhr122w) Q9Y3D0 17663 5.2 878 7641.15 29 87.7 U 1 1 ELP4 Elongator complex protein 4 Q96EB1 46588 8.5 497 1662.81 40 65.8 U 1 2 C3orf75 UPF0405 protein C3orf75 Q0PNE2 29793 6.8 293 1562.56 23 44.7 U 2 1 IOP1 CIA Component (homolog of yeast Nar1) Q9H6Q4 53020 7.2 507 1510.96 47 72.1 U 1 1 DNA2 DNA2-like helicase, nuclease P51530 120415 7.7 963 1288.76 90 60.8 U 1 3 PRIM2 DNA primase large subunit P49643 58806 7.9 424 1181.68 50 57.8 U 1 2 PRIM1 DNA primase small subunit P49642 49902 8.2 304 1026.78 32 54.5 U 1 1 CDKAL1 CDK5 regulatory subunit-associated protein 1-like 1 Q5VV42 65112 7.5 383 938.37 49 67.2 U 1 1 DERP6 Dermal papilla-derived protein 6 Q8TE02 34841 5.0 203 911.30 15 33.9 U 1 2 CTU1 Cytoplasmic tRNA 2-thiolation protein 1 Q7Z7A3 36450 9.2 200 815.27 24 48.0 U 1 1 CTU2 Cytoplasmic tRNA 2-thiolation protein 2 Q2VPK5 56107 6.3 277 763.00 30 48.9 U 1 1 POLD1 DNA polymerase delta catalytic subunit P28340 123631 7.0 562 720.18 79 57.9 U 1 2 IKBKAP Elongator complex protein 1 O95163 150254 5.9 246 261.99 63 46.2 U 1 3 FMNL3 Formin-like protein 3 Q8IVF7 117213 6.7 189 260.81 47 35.0 U 1 3 XPD Xeroderma pigmentosum protein D, helicase subunit of TFIIH
complex P18074 86909 7.2 131 244.52 27 35.0 U 1 1
FMNL2 Formin-like protein 2 Q96PY5 123321 7.4 176 229.90 35 26.0 U 1 3 IOP2 Iron-only hydrogenase-like Fe/S protein (unknown function) Q9UHQ1 51156 7.1 73 227.10 7 11.6 U 1 1 GLRX3 Glutaredoxin-3 O76003 37432 5.4 47 199.02 14 33.7 U 1 2 ELP3 Elongator complex protein 3 Q9H9T3 62259 8.9 66 171.16 21 40.6 U 1 2 ABCE1 ATP-binding cassette family member E1 (RLI1) P61221 67314 8.3 72 170.51 26 44.7 U 1 1 NTHL1 Endonuclease III-like glycosylase 1 P78549 34390 9.7 36 163.68 6 26.9 U 1 2 NUP53 Nucleoporin NUP53 Q8NFH5 34774 9.1 34 147.95 10 34.7 U 1 1 MAK16 Protein MAK16 homolog Q9BXY0 35369 5.4 30 141.86 10 28.0 U 1 2 RTEL1 Regulator of telomere elongation helicase 1 Q9NZ71 133683 8.3 111 129.17 32 35.0 U 1 2 CNP 2',3'-cyclic-nucleotide 3'-phosphodiesterase P09543 47579 9.1 33 111.19 15 33.5 U 1 2 TYW1 tRNA wybutosine-synthesizing protein Q9NV66 83702 6.9 55 106.59 18 29.2 U 1 2

11
ORAOV1 Oral cancer-overexpressed protein 1 Q8WV07 15354 5.8 10 103.55 5 40.9 U 2 2 MLF2 Myeloid leukemia factor 2 Q15773 28147 6.9 18 102.96 7 31.0 U 1 1 MSL1 Male-specific lethal 1 homolog Q68DK7 67139 8.9 43 99.35 10 24.9 U 1 2 EYA2 Eyes absent homolog 2 O00167 59232 6.5 36 94.92 2 1.3 U 1 2 ALDH3A2 Fatty aldehyde dehydrogenase P51648 54848 7.9 30 87.75 9 21.9 U 1 1 ELP2 Elongator complex protein 2 Q6IA86 92500 6.0 49 84.15 23 32.1 U 1 2 KIF4A Chromosome-associated kinesin KIF4A O95239 139881 6.3 72 82.90 33 29.4 U 1 3 ARL1 ADP-ribosylation factor-like protein 1 P40616 20418 5.7 9 70.54 3 13.8 U 1 1 SIP1 Survival of motor neuron protein-interacting protein 1 O14893 31585 5.6 13 65.86 6 22.5 U 1 1 C7orf36 Uncharacterized protein C7orf36 Q9NRH1 25299 5.6 10 62.77 5 27.0 U 1 1 SHKBP1 SH3KBP1-binding protein 1 Q8TBC3 76344 8.3 28 56.18 10 24.0 U 1 2 DPH2 Diphthamide biosynthesis protein 2 Q9BQC3 52083 5.5 18 52.22 5 15.1 U 1 1 TFPI Tissue factor pathway inhibitor P10646 35015 8.2 10 46.66 5 16.4 U 1 1 GPAT Glutamine phosphoribosylpyrophosphate amido transferase Q06203 57399 6.8 17 46.65 10 22.6 U 1 1 USPL1 Ubiquitin-specific peptidase-like protein 1 Q5W0Q7 120440 6.2 33 42.87 13 19.4 U 2 1 ZNF536 Zinc finger protein 536 O15090 141417 7.3 38 41.47 20 24.4 U 1 3 RANGAP1 Ran GTPase-activating protein 1 P46060 63542 4.7 17 41.08 8 15.7 U 1 1 SPC25 Kinetochore protein Spc25 Q9HBM1 26153 8.0 6 38.00 4 27.2 U 1 1 RFC5 Replication factor C subunit 5 P40937 38497 7.2 9 37.55 6 23.5 U 1 1 GRK6 G protein-coupled receptor kinase 6 P43250 65991 8.0 14 34.48 9 19.8 U 1 2 MRPL40 39S ribosomal protein L40, mitochondrial Q9NQ50 24490 9.6 5 34.43 2 16.5 U 1 1 GEMIN6 Gem-associated protein 6 Q8WXD5 18824 5.1 4 33.98 2 14.4 U 1 1 EIF2S1 Eukaryotic translation initiation factor 2 subunit 1 P05198 36112 5.1 7 31.52 4 16.5 U 1 3 KPNA2 Importin subunit alpha-2 P52292 57862 5.4 11 29.50 6 13.0 U 1 1 MBD3 Methyl-CpG-binding domain protein 3 O95983 32844 5.3 6 29.25 2 10.0 U 1 1 ILVBL Acetolactate synthase-like protein A1L0T0 67868 8.1 13 29.18 6 13.9 U 1 2 FANCJ Fanconi anemia group J protein, helicase Q9BX63 140877 6.9 22 24.99 12 14.4 U 1 1 TRIP13 Thyroid receptor-interacting protein 13 Q15645 48551 6.1 7 22.99 5 13.4 U 1 2 DPH1 Diphthamide biosynthesis protein 1 Q9BZG8 48805 8.2 7 22.42 4 10.4 U 1 2 WNK1 Serine/threonine-protein kinase WNK1 Q9H4A3 250753 6.3 37 22.03 19 13.2 U 1 1 TMEM43 Transmembrane protein 43 Q9BTV4 44876 8.1 6 21.28 3 10.2 U 1 1 SURF4 Surfeit locus protein 4 O15260 30394 7.8 4 21.09 3 15.2 U 1 3 USP39 U4/U6.U5 tri-snRNP-associated protein 2 Q53GS9 65381 8.9 8 20.09 6 13.1 U 1 2 EIF3K Eukaryotic translation initiation factor 3 subunit K Q9UBQ5 25060 4.9 3 19.52 3 17.9 U 1 1

12
POLA1 DNA polymerase alpha catalytic subunit P09884 165912 5.8 20 19.41 12 8.2 U 1 2 FMNL1 Formin-like protein 1 O95466 121853 5.7 15 19.34 3 3.0 U 1 3 GEMIN8 Gem-associated protein 8 Q9NWZ8 28637 6.8 3 17.59 2 9.9 U 1 1 SMCR8 Smith-Magenis syndrome chromosomal region candidate gene 8
protein Q8TEV9 105022 5.6 11 16.65 7 13.4 U 1 2
PRPSAP1 Phosphoribosyl pyrophosphate synthase-associated protein 1 Q14558 39394 7.2 4 15.94 3 8.4 U 1 2 RPN2 Dolichyl-diphosphooligosaccharide--protein glycosyltransferase
subunit 2 P04844 69284 5.7 7 15.74 3 6.7 U 1 3
ARF1 ADP-ribosylation factor 1 P84077 20697 6.8 2 15.67 2 13.8 group-13 1 2 ARF3 ADP-ribosylation factor 3 P61204 20601 7.4 2 15.67 2 13.8 group-13 1 2 GNL2 Nucleolar GTP-binding protein 2 Q13823 83655 9.2 8 15.52 6 11.6 U 1 1 WDR41 WD repeat-containing protein 41 Q9HAD4 51728 5.4 5 15.45 4 15.3 U 2 3 LARP4B La-related protein 4B Q92615 80552 6.9 8 15.38 3 7.7 U 1 3 APOBEC3C Probable DNA dC- dU-editing enzyme APOBEC-3C Q9NRW3 22826 7.6 2 14.93 2 13.7 U 1 2 TOP3B DNA topoisomerase 3-beta-1 O95985 96662 8.1 9 14.81 7 12.4 U 1 1 IDH3B Isocitrate dehydrogenase [NAD] subunit beta, mitochondrial O43837 42184 8.5 4 14.74 2 10.6 U 1 2 CDIPT CDP-diacylglycerol--inositol 3-phosphatidyltransferase O14735 23539 8.0 2 13.32 2 11.3 U 1 1 CSE1L Exportin-2 P55060 110417 5.8 9 13.15 7 9.8 U 1 3 None Eukaryotic translation initiation factor 2 subunit 2-like protein A6NK07 37787 6.4 3 12.90 2 8.3 group-19 1 1 EIF2S2 Eukaryotic translation initiation factor 2 subunit 2 P20042 38388 5.8 3 12.90 2 8.1 group-19 1 2 SNRPB2 U2 small nuclear ribonucleoprotein B'' P08579 25486 9.7 2 12.61 2 11.6 U 1 1 THOC6 THO complex subunit 6 homolog Q86W42 37535 7.4 3 12.48 2 5.6 U 1 1 POLE DNA polymerase epsilon catalytic subunit A Q07864 261515 6.4 20 12.41 13 7.4 U 1 5 PRPF6 Pre-mRNA-processing factor 6 O94906 106925 8.3 8 12.06 4 5.3 U 1 1 DHCR7 7-dehydrocholesterol reductase Q9UBM7 54489 8.7 4 11.95 2 4.6 U 1 1 DNAJB11 DnaJ homolog subfamily B member 11 Q9UBS4 40514 6.2 3 11.89 2 8.4 U 1 1 EIF3M Eukaryotic translation initiation factor 3 subunit M Q7L2H7 42503 5.6 3 11.38 3 11.0 U 1 1 DUSP9 Dual specificity protein phosphatase 9 Q99956 41868 6.1 3 11.08 2 8.3 U 1 1 SLFN11 Schlafen family member 11 Q7Z7L1 102836 7.7 7 11.02 4 7.2 U 1 1 CDK17 Cell division protein kinase 17 Q00537 59582 9.0 4 10.85 3 5.4 U 1 2 ABCD3 ATP-binding cassette sub-family D member 3 P28288 75476 9.4 5 10.76 4 8.2 U 1 1 KCTD3 BTB/POZ domain-containing protein KCTD3 Q9Y597 88984 7.0 6 10.44 3 6.6 U 1 2 MUTYH A/G-specific adenine DNA glycosylase Q9UIF7 60069 8.7 4 10.39 3 6.6 U 1 1 DCP2 mRNA-decapping enzyme 2 Q8IU60 48457 8.0 3 10.13 2 6.2 U 1 2 GTF2E2 Transcription initiation factor IIE subunit beta P29084 33044 9.7 2 9.75 2 6.9 U 1 1

13
ATP1A1 Sodium/potassium-transporting ATPase subunit alpha-1 P05023 112896 5.5 7 9.71 5 5.8 U 1 1 FANCI Fanconi anemia group I protein Q9NVI1 149323 6.7 9 9.61 6 6.4 U 1 4 HSD17B12 Estradiol 17-beta-dehydrogenase 12 Q53GQ0 34324 9.3 2 9.09 2 12.8 U 1 2 AP2B1 AP-2 complex subunit beta P63010 104553 5.4 6 9.08 4 5.1 U 1 1 POLD2 DNA polymerase delta subunit 2 P49005 51289 5.6 3 9.07 3 8.1 U 1 1 SLC25A11 Mitochondrial 2-oxoglutarate/malate carrier protein Q02978 34062 9.9 2 9.04 2 6.7 U 1 3 ERLIN2 Erlin-2 O94905 37840 5.6 2 8.37 2 5.9 U 1 1 PRPF3 U4/U6 small nuclear ribonucleoprotein Prp3 O43395 77529 9.5 4 8.31 3 5.4 U 1 2 SMARCD1 SWI/SNF-related matrix-associated actin-dependent regulator of
chromatin subfamily D member 1 Q96GM5 58233 9.2 3 8.26 2 5.4 U 1 2
RBMXL3 RNA-binding motif protein, X-linked-like-3 Q8N7X1 114938 9.1 6 7.98 2 3.9 U 2 2 BAT4 Protein BAT4 O95872 39314 8.2 2 7.97 2 7.3 U 2 1 IRAK1 Interleukin-1 receptor-associated kinase 1 P51617 76537 6.6 4 7.97 3 6.3 U 1 2 HLA-A HLA class I histocompatibility antigen, A-69 alpha chain P10316 40977 6.9 2 7.75 2 10.1 group-18 1 2 HLA-B HLA class I histocompatibility antigen, B-73 alpha chain Q31612 40435 6.2 2 7.75 2 10.2 group-18 1 1 HLA-C HLA class I histocompatibility antigen, Cw-17 alpha chain Q95604 41238 6.8 2 7.75 2 9.9 group-18 1 1 HLA-A HLA class I histocompatibility antigen, A-2 alpha chain P01892 40922 7.0 2 7.75 2 10.1 group-18 1 1 HLA-C HLA class I histocompatibility antigen, Cw-7 alpha chain P10321 40649 6.0 2 7.75 2 10.1 group-18 1 3 ADD1 Alpha-adducin P35611 80955 5.8 4 7.70 3 6.4 U 1 2 MLH1 DNA mismatch repair protein Mlh1 P40692 84601 5.7 4 7.51 3 4.6 U 1 1 CCNK Cyclin-K O75909 64240 8.4 3 7.34 3 10.5 U 1 2 WTAP Pre-mRNA-splicing regulator WTAP Q15007 44244 5.2 2 7.16 2 6.3 U 1 2 POLA2 DNA polymerase alpha subunit B Q14181 65948 5.2 3 7.12 2 6.9 U 1 2 RANBP2 E3 SUMO-protein ligase RanBP2 P49792 358201 6.2 16 7.04 12 5.0 U 1 2 MAGED2 Melanoma-associated antigen D2 Q9UNF1 64954 9.3 3 7.02 2 4.6 U 1 2 TRMT2A tRNA (uracil-5-)-methyltransferase homolog A Q8IZ69 68726 7.9 3 6.81 2 4.6 U 1 2 ABCF1 ATP-binding cassette sub-family F member 1 Q8NE71 95926 6.8 4 6.72 3 3.9 U 1 2 RAF1 RAF proto-oncogene serine/threonine-protein kinase P04049 73052 9.2 3 6.57 2 5.2 U 1 1 BAG5 BAG family molecular chaperone regulator 5 Q9UL15 51200 6.0 2 6.35 2 5.6 U 1 1 SFRS15 Splicing factor, arginine/serine-rich 15 O95104 125869 9.6 5 6.18 3 3.8 U 1 3 FRYL Protein furry homolog-like O94915 339598 5.6 13 6.12 9 4.3 U 1 2 ALG1 Chitobiosyldiphosphodolichol beta-mannosyltransferase Q9BT22 52518 7.2 2 6.11 2 7.3 U 1 2 QSOX2 Sulfhydryl oxidase 2 Q6ZRP7 77529 7.7 3 6.10 2 3.9 U 1 3 C9orf72 Uncharacterized protein C9orf72 Q96LT7 54328 6.2 2 5.90 2 4.4 U 2 2 SMC3 Structural maintenance of chromosomes protein 3 Q9UQE7 141541 7.2 5 5.83 4 5.1 U 1 2

14
PIGS GPI transamidase component PIG-S Q96S52 61656 6.5 2 5.11 2 5.6 U 1 3 AATF Protein AATF Q9NY61 63133 4.9 2 5.07 2 5.5 U 1 1 MSL2 Male-specific lethal 2 homolog Q9HCI7 62541 7.8 2 4.92 2 7.8 U 1 2 ZNF281 Zinc finger protein 281 Q9Y2X9 96915 8.5 3 4.75 3 5.1 U 1 1 DDX52 Probable ATP-dependent RNA helicase DDX52 Q9Y2R4 67466 9.7 2 4.74 2 5.3 U 1 2 RPN1 Dolichyl-diphosphooligosaccharide--protein glycosyltransferase
subunit 1 P04843 68569 6.4 2 4.67 2 4.3 U 1 1
PWP2 Periodic tryptophan protein 2 homolog Q15269 102452 6.1 3 4.63 2 2.7 U 1 2 PPP1R10 Serine/threonine-protein phosphatase 1 regulatory subunit 10 Q96QC0 99058 9.2 3 4.53 2 4.6 U 1 1 SLC3A2 4F2 cell-surface antigen heavy chain P08195 67994 5.0 2 4.50 2 3.2 U 1 3 EDEM1 ER degradation-enhancing alpha-mannosidase-like 1 Q92611 73768 6.9 2 4.32 2 3.2 U 1 1 ARHGEF2 Rho guanine nucleotide exchange factor 2 Q92974 111543 7.3 3 4.32 2 3.9 U 1 4 XPO1 Exportin-1 O14980 123386 6.1 3 3.97 3 3.6 U 1 1 RBBP6 Retinoblastoma-binding protein 6 Q7Z6E9 201563 9.6 5 3.96 2 1.6 U 1 1 XPO5 Exportin-5 Q9HAV4 136311 5.8 3 3.53 2 2.9 U 1 1 HDAC6 Histone deacetylase 6 Q9UBN7 131419 5.3 3 3.50 3 4.9 U 1 2 MTUS1 Microtubule-associated tumor suppressor 1 Q9ULD2 141396 7.5 3 3.35 2 2.0 U 1 2 NOP14 Nucleolar protein 14 P78316 97668 7.6 2 3.31 2 2.9 U 1 3 RNF160 RING finger protein 160 O94822 200524 6.3 4 3.21 2 1.4 U 1 5 WDR36 WD repeat-containing protein 36 Q8NI36 105322 7.5 2 2.98 2 2.0 U 1 1 AP2A1 AP-2 complex subunit alpha-1 O95782 107546 7.0 2 2.90 2 3.4 U 1 3 ZNF318 Zinc finger protein 318 Q5VUA4 251110 7.2 4 2.49 3 2.1 U 1 2 MON2 Protein MON2 homolog Q7Z3U7 190486 6.1 3 2.48 2 2.2 U 1 2 EIF5B Eukaryotic translation initiation factor 5B O60841 138827 5.5 2 2.33 2 1.6 U 1 4 DICER1 Endoribonuclease Dicer Q9UPY3 218680 5.7 3 2.21 2 1.3 U 1 3 DST Bullous pemphigoid antigen 1 Q03001 860679 5.2 6 1.12 3 0.5 U 1 4 Abbreviations: NSAF, normalized spectral abundance factor: PE, protein existence; SV, sequence version.

15
Supplementary Figures
Fig. S1. Interaction of yeast Mms19 with CIA factors and Fe/S proteins. (A) Gal-GFP-MMS19 yeast cells were transformed with plasmids encoding Cia2-HA, Cia1-HA, Nar1-HA, or no insert (-). Cell extracts (CE) prepared from an overnight culture grown in SC galactose medium were immunoprecipitated using anti-HA beads. Cell extracts and immunoprecipitates (IP:HA) were separated by SDS-PAGE, and Mms19 was immunostained using its GFP-tag (IB:GFP). (B) GalL-MMS19 cells were transformed with plasmids encoding Rli1-HA, Ntg2-HA, or Cia1-HA. The non-Fe/S protein Ymr134w-HA (see (19)) was used as a negative control. Extracts from overnight cultures grown in SC galactose medium were used for immunoprecipitation (IP) with anti-HA beads. Cell extracts (CE) and immunoprecipitates (IP:HA) were analyzed by immunostaining for the indicated proteins with specific antibodies (anti-HA, top; anti-Mms19, bottom).

16
Fig. S2. Cytosolic enzyme activities in methionine biosynthesis-defective and CIA machinery-depleted cells. (A) Wild-type (WT) and the deletion yeast strains met1, met5, met10 and mms19 (in BY strain background) were grown in glucose-containing minimal medium supplemented with methionine. Enzyme activities of isopropylmalate isomerase (Leu1) were measured in cell extracts. (B) Wild-type (WT) and the indicated galactose-regulatable CIA strains were grown overnight in SC medium containing galactose (Gal) or glucose (Glc), and spotted on SC galactose (SC-Gal-Bi) or glucose (SC-Glc-Bi) plates containing bismuth ammonium citrate and sodium sulfite. Cells were grown for 3 days. Sulfide produced by sulfite reductase yields a brown precipitate (Bi2S3).
17
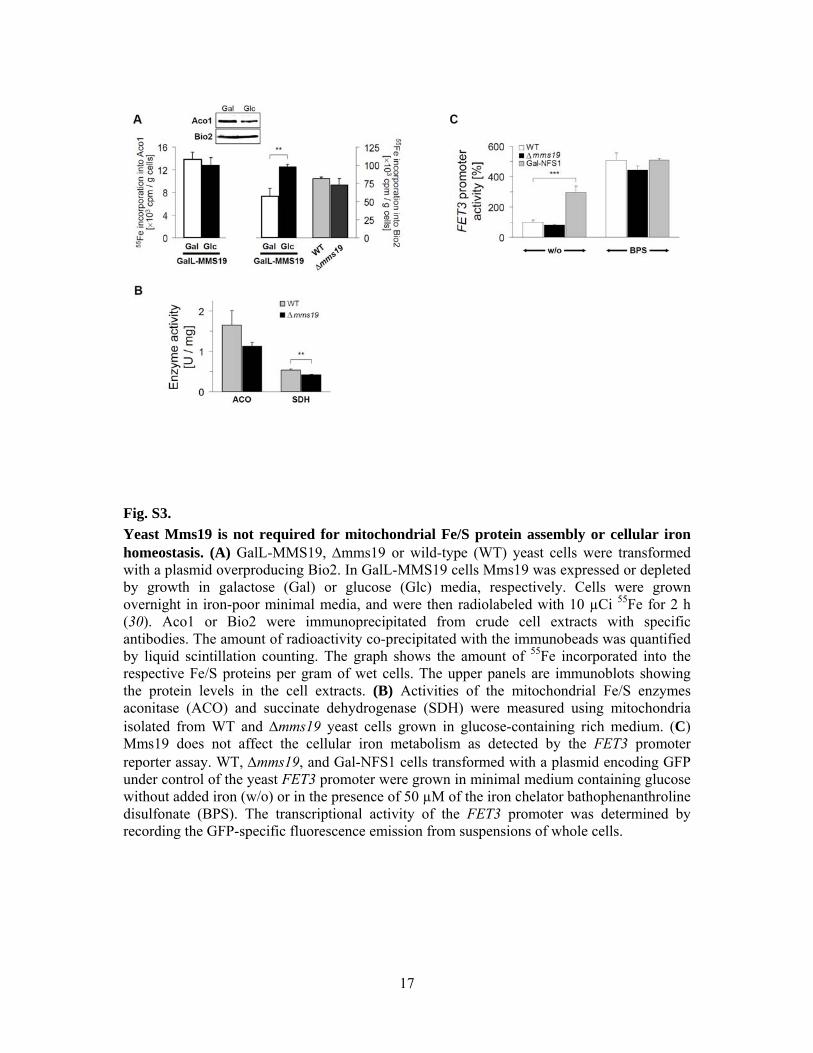
Fig. S3. Yeast Mms19 is not required for mitochondrial Fe/S protein assembly or cellular iron homeostasis. (A) GalL-MMS19, Δmms19 or wild-type (WT) yeast cells were transformed with a plasmid overproducing Bio2. In GalL-MMS19 cells Mms19 was expressed or depleted by growth in galactose (Gal) or glucose (Glc) media, respectively. Cells were grown overnight in iron-poor minimal media, and were then radiolabeled with 10 µCi 55Fe for 2 h (30). Aco1 or Bio2 were immunoprecipitated from crude cell extracts with specific antibodies. The amount of radioactivity co-precipitated with the immunobeads was quantified by liquid scintillation counting. The graph shows the amount of 55Fe incorporated into the respective Fe/S proteins per gram of wet cells. The upper panels are immunoblots showing the protein levels in the cell extracts. (B) Activities of the mitochondrial Fe/S enzymes aconitase (ACO) and succinate dehydrogenase (SDH) were measured using mitochondria isolated from WT and Δmms19 yeast cells grown in glucose-containing rich medium. (C) Mms19 does not affect the cellular iron metabolism as detected by the FET3 promoter reporter assay. WT, Δmms19, and Gal-NFS1 cells transformed with a plasmid encoding GFP under control of the yeast FET3 promoter were grown in minimal medium containing glucose without added iron (w/o) or in the presence of 50 µM of the iron chelator bathophenanthroline disulfonate (BPS). The transcriptional activity of the FET3 promoter was determined by recording the GFP-specific fluorescence emission from suspensions of whole cells.

18
Fig. S4. 55Fe incorporation into XPD and Dna2 Fe/S cluster-containing helicases. (A) The human DNA helicase XPD (WT) and the indicated mutant proteins were TAP tagged and expressed in W303 wild-type yeast cells. After growth in galactose-containing medium cells were radiolabeled with 55Fe. XPD-TAP and mutant proteins were immunoprecipitated from cell extracts, and bound 55Fe was quantified by scintillation counting. The immunoblot shows XPD in whole cell extracts. For all depletions p<0.001. (B) Mms19 is required for Fe/S protein maturation of yeast Dna2. GalL-MMS19 cells were transformed with a plasmid encoding Dna2-HA. 55Fe incorporation (as in A) was measured in cells grown in galactose- (Gal) and glucose- (Glc) containing media. An immunostain of Dna2-HA in cell extracts is shown.

19
Fig. S5. Effective depletion of human MMS19 by siRNAs. (A) HeLa cells were transfected thrice at a three day interval without or with individual (diamonds, triangles, and squares) or pooled (▬ ± SD) siRNAs directed against different regions of MMS19 mRNA. 3, 6, and 9 days after the first transfection residual mRNA (left) or protein (right) levels of MMS19 were determined by qRT-PCR or immunoblotting, respectively. The data were normalized to beta-actin, and these ratios to the levels of mock-transfected cells (set to 100%). Note that the mismatch-containing siRNA si#3 was ineffective in depleting MMS19 thus serving as an internal control (see Table S3). (B) A representative example of the data in (A) is presented for HeLa cells treated for 9 days. Cells were permeabilized with digitonin, and the lysate was separated into supernatant (Cytosol) and pellet (Organelles) fractions showing a predominant cytosolic localization of MMS19. The cytosolic protein β-actin and mitochondrial intermembrane space protein MIA40 were used as controls. Abbreviations: -, mock-treated cells, +, cells treated with the pool of three siRNAs directed against MMS19.

20
Fig. S6. Depletion of human MMS19 does not affect the Fe/S cluster status of IRP1. HeLa cells were treated with MMS19-targeting siRNAs (cf. fig. S5), or with two scrambled siRNAs not targeting any known human RNA (used individually or as a pool) for 3, 6, and 9 days as in fig. S5. (A) IRP1 (cytosolic aconitase) enzyme activities were measured using the cytosolic fractions (cf. fig. S5B) by an isocitrate dehydrogenase-coupled NADP+-dependent spectrophotometric assay. Data were normalized to LDH activity, and expressed relative to the activities in mock-transfected cells (set to 100%; dashed line). (B) Total cell lysates were immunostained and the ratio of IRP1 and β-actin was normalized to the respective ratio of mock-transfected cells (dashed line). (C) The IRP1 binding to 32P-labeled iron-responsive element (IRE) of human ferritin mRNA was analyzed by native gel electrophoresis and subsequent phosphorimaging. IRP2 was super-shifted by specific antibodies to obtain the IRP1 signal only. The IRP1-IRE binding (- β-ME) was calculated relative to the maximum IRP1 binding activity determined in the presence of 2% β-mercaptoethanol (+ β-ME), and a representative example is shown for 9 days of treatment (bottom panel). Data (± SD) were normalized to the respective ratios in mock-transfected cells (dashed line).

21
Fig. S7. Depletion of human MMS19 does not affect GPAT protein levels. HeLa cells were depleted for MMS19 as in Figs. 3A and S5A, and analyzed by immunostaining for GPAT and beta-actin. Data were analyzed and quantitated as in fig. S6.

22
Fig. S8. DPYD enzyme activity and protein levels are severely decreased upon human MMS19 depletion. HeLa cells were depleted for MMS19 as in Figs. 3B and S5A and analyzed for (A) DPYD-dependent conversion of [4-14C]-thymine ([4-14C]-T) to [4-14C]-dihydrothymine ([4-14C]-DHT) by thin layer chromatography and autoradiography. The product of DPYD function, [4-14C]-DHT, is virtually absent in samples of HeLa cells treated for 9 days with pooled siRNAs for depletion of MMS19. Formation of [4-14C]-DHT was not affected in lysates of cells transfected with scrambled siRNA. (B-C) Normalization of specific DPYD activity to lactate dehydrogenase (LDH) activity (B) and presentation of the data relative to the ratios in mock-transfected cells (set to 100%; dashed lines) demonstrated that formation of [4-14C]-DHT was least efficient in samples of HeLa cells transfected with pooled MMS19-directed siRNAs, yet was hardly affected in HeLa cells transfected with the scrambled siRNAs or the ineffective MMS19-targeting siRNA si#3 (grey squares; cf. fig. S5A). (D) Total cell lysates from (B) were immunostained and the ratio of protein-associated chemiluminescence for DPYD and β-actin was normalized to the respective ratio of mock-transfected cells (set to 100%; dashed line).

23
Fig. S8 (ctd.). (E) HeLa cells were RNAi-depleted for the ISC assembly components NFS1 and frataxin (FXN) and the CIA factor NBP35 (16, 63, 64). Total cell extracts were immunostained for DPYD and beta-actin and data analyzed as in (D). For all depletions p<0.01, except for day 3 of FXN siRNA treatment.

24
Fig. S9. POLD1 protein levels are diminished upon human MMS19 depletion. (A) HeLa cells were depleted for MMS19 as in Figs. 3D and S5A and analyzed by immunostaining for POLD1, the catalytic subunit of DNA polymerase δ. Normalization of the data was done using beta-actin, and the ratios were presented relative to those in mock-transfected cells (set to 100%; dashed lines). POLD1 levels were strongly affected in cells depleted for MMS19 for 9 days with pooled siRNAs, and virtually no effect was obtained for the non-effective siRNA si#3 (grey squares; cf. fig. S5A). (B) HeLa cells were RNAi-depleted for the ISC assembly components NFS1 and frataxin (FXN) and the CIA factor NBP35 (16, 63, 64). Total cell extracts were immunostained for POLD1 and beta-actin and data analyzed as in (A).

25
Fig. S10. Depletion of human MMS19 does not affect mitochondrial Fe/S enzymes. HeLa cells were depleted for MMS19 as in Figs. 3A and S5A and analyzed for (A) protein levels and (B) enzyme activities of mitochondrial aconitase (mtAco), or (C) protein levels (SDHB subunit) and (D) enzyme activities of mitochondrial succinate dehydrogenase (SDH). Protein levels were determined by immunostaining of total cell lysates, and quantitated relative to β-actin levels. Data were normalized to the respective ratio in mock-transfected cells. Aconitase and SDH enzyme activities were measured in the organellar fraction of digitonin-lysed cells, and quantitated relative to citrate synthase (CS) activity. Data were expressed relative to the activities in mock-transfected cells (dashed lines), and presented as in fig. S6.

26
Fig. S11. XPD associates with MMS19 at endogenous concentrations. Endogenous XPD protein was immunoprecipitated from protein extracts derived from HEK293 cells using XPD antibodies. Whole cell extracts (WCE) and immunoprecipitates (IP) were immunostained with specified antibodies.

27
Fig. S12. Doxycycline-inducible depletion of MMS19 in HEK293 cells. HEK293 cells stably expressing a doxycycline-inducible control non-targeting shRNA construct (No target) or shRNA constructs designed to silence expression of MMS19, FAM96B, IOP1, or ISCU were generated the using the Thermo Scientific TRIPZ Lentiviral shRNAmir system. Cells were induced continuously with doxycycline (1 µg/mL) for 12 days to maintain sustained depletion of the proteins of interest. Cell lysates were prepared and immunostained with the specified antibodies to assess the degree of silencing.

28
Fig. S13. Model for MMS19 function in maturation of Fe/S proteins involved in DNA metabolism. Fe/S protein biogenesis requires the mitochondrial ISC assembly machinery, the ABC transporter ABCB7 and the CIA machinery. The CIA component MMS19 is part of the ‘CIA targeting complex’ that transfers Fe/S clusters from the CIA scaffold complex CFD1-NBP35 to specific Fe/S apoproteins including members involved DNA metabolism. The putative roles of human CIAO1 and FAM96B in the assembly process remain to be elucidated.

1
References
1. M. A. McMurray, D. E. Gottschling, Aging and genetic instability in yeast. Curr. Opin.
Microbiol. 7, 673 (2004). doi:10.1016/j.mib.2004.10.008 Medline
2. J. H. Hoeijmakers, DNA damage, aging, and cancer. N. Engl. J. Med. 361, 1475 (2009).
doi:10.1056/NEJMra0804615 Medline
3. A. Ciccia, S. J. Elledge, The DNA damage response: Making it safe to play with knives.
Mol. Cell 40, 179 (2010). doi:10.1016/j.molcel.2010.09.019 Medline
4. J. R. Veatch, M. A. McMurray, Z. W. Nelson, D. E. Gottschling, Mitochondrial
dysfunction leads to nuclear genome instability via an iron-sulfur cluster defect. Cell
137, 1247 (2009). doi:10.1016/j.cell.2009.04.014 Medline
5. G. Karthikeyan, L. K. Lewis, M. A. Resnick, The mitochondrial protein frataxin prevents
nuclear damage. Hum. Mol. Genet. 11, 1351 (2002). doi:10.1093/hmg/11.11.1351
Medline
6. R. Thierbach et al., The Friedreich’s ataxia protein frataxin modulates DNA base excision
repair in prokaryotes and mammals. Biochem. J. 432, 165 (2010).
doi:10.1042/BJ20101116 Medline
7. J. Rudolf, V. Makrantoni, W. J. Ingledew, M. J. Stark, M. F. White, The DNA repair
helicases XPD and FancJ have essential iron-sulfur domains. Mol. Cell 23, 801
(2006). doi:10.1016/j.molcel.2006.07.019 Medline
8. S. Klinge, J. Hirst, J. D. Maman, T. Krude, L. Pellegrini, An iron-sulfur domain of the
eukaryotic primase is essential for RNA primer synthesis. Nat. Struct. Mol. Biol. 14,
875 (2007). doi:10.1038/nsmb1288 Medline
9. J. T. Yeeles, R. Cammack, M. S. Dillingham, An iron-sulfur cluster is essential for the
binding of broken DNA by AddAB-type helicase-nucleases. J. Biol. Chem. 284, 7746
(2009). doi:10.1074/jbc.M808526200 Medline
10. D. J. Netz et al., Eukaryotic DNA polymerases require an iron-sulfur cluster for the
formation of active complexes. Nat. Chem. Biol. 8, 125 (2012).
doi:10.1038/nchembio.721 Medline
11. A. Sheftel, O. Stehling, R. Lill, Iron-sulfur proteins in health and disease. Trends
Endocrinol. Metab. 21, 302 (2010). doi:10.1016/j.tem.2009.12.006 Medline

2
12. M. F. White, M. S. Dillingham, Iron-sulphur clusters in nucleic acid processing enzymes.
Curr. Opin. Struct. Biol. 22, 94 (2012). doi:10.1016/j.sbi.2011.11.004 Medline
13. R. Lill, Function and biogenesis of iron-sulphur proteins. Nature 460, 831 (2009).
doi:10.1038/nature08301 Medline
14. H. Ye, T. A. Rouault, Human iron-sulfur cluster assembly, cellular iron homeostasis, and
disease. Biochemistry 49, 4945 (2010). doi:10.1021/bi1004798 Medline
15. A. Roy, N. Solodovnikova, T. Nicholson, W. Antholine, W. E. Walden, A novel
eukaryotic factor for cytosolic Fe-S cluster assembly. EMBO J. 22, 4826 (2003).
doi:10.1093/emboj/cdg455 Medline
16. O. Stehling et al., Human Nbp35 is essential for both cytosolic iron-sulfur protein
assembly and iron homeostasis. Mol. Cell. Biol. 28, 5517 (2008).
doi:10.1128/MCB.00545-08 Medline
17. U. Mühlenhoff et al., Cytosolic monothiol glutaredoxins function in intracellular iron
sensing and trafficking via their bound iron-sulfur cluster. Cell Metab. 12, 373 (2010).
doi:10.1016/j.cmet.2010.08.001 Medline
18. D. J. Netz et al., Tah18 transfers electrons to Dre2 in cytosolic iron-sulfur protein
biogenesis. Nat. Chem. Biol. 6, 758 (2010). doi:10.1038/nchembio.432 Medline
19. J. Balk, D. J. Aguilar Netz, K. Tepper, A. J. Pierik, R. Lill, The essential WD40 protein
Cia1 is involved in a late step of cytosolic and nuclear iron-sulfur protein assembly.
Mol. Cell. Biol. 25, 10833 (2005). doi:10.1128/MCB.25.24.10833-10841.2005
Medline
20. J. Balk, A. J. Pierik, D. J. Netz, U. Mühlenhoff, R. Lill, The hydrogenase-like Nar1p is
essential for maturation of cytosolic and nuclear iron-sulphur proteins. EMBO J. 23,
2105 (2004). doi:10.1038/sj.emboj.7600216 Medline
21. D. Song, F. S. Lee, A role for IOP1 in mammalian cytosolic iron-sulfur protein
biogenesis. J. Biol. Chem. 283, 9231 (2008). doi:10.1074/jbc.M708077200 Medline
22. L. Prakash, S. Prakash, Three additional genes involved in pyrimidine dimer removal in
Saccharomyces cerevisiae: RAD7, RAD14 and MMS19. MGG 176, 351 (1979).
doi:10.1007/BF00333097 Medline

3
23. S. H. Askree et al., A genome-wide screen for Saccharomyces cerevisiae deletion
mutants that affect telomere length. Proc. Natl. Acad. Sci. U.S.A. 101, 8658 (2004).
doi:10.1073/pnas.0401263101 Medline
24. S. Ito et al., MMXD, a TFIIH-independent XPD-MMS19 protein complex involved in
chromosome segregation. Mol. Cell 39, 632 (2010). doi:10.1016/j.molcel.2010.07.029
Medline
25. H. Kou, Y. Zhou, R. M. Gorospe, Z. Wang, Mms19 protein functions in nucleotide
excision repair by sustaining an adequate cellular concentration of the TFIIH
component Rad3. Proc. Natl. Acad. Sci. U.S.A. 105, 15714 (2008).
doi:10.1073/pnas.0710736105 Medline
26. A. C. Gavin et al., Proteome survey reveals modularity of the yeast cell machinery.
Nature 440, 631 (2006). doi:10.1038/nature04532 Medline
27. S. Ben-Aroya et al., Toward a comprehensive temperature-sensitive mutant repository of
the essential genes of Saccharomyces cerevisiae. Mol. Cell 30, 248 (2008).
doi:10.1016/j.molcel.2008.02.021 Medline
28. E. Weerapana et al., Quantitative reactivity profiling predicts functional cysteines in
proteomes. Nature 468, 790 (2010). doi:10.1038/nature09472 Medline
29. See supplementary materials on Science Online.
30. A. J. Pierik, D. J. A. Netz, R. Lill, Analysis of iron-sulfur protein maturation in
eukaryotes. Nat. Protoc. 4, 753 (2009). doi:10.1038/nprot.2009.39 Medline
31. F. Li, R. Martienssen, W. Z. Cande, Coordination of DNA replication and histone
modification by the Rik1-Dos2 complex. Nature 475, 244 (2011).
doi:10.1038/nature10161 Medline
32. T. Seroz et al., Cloning of a human homolog of the yeast nucleotide excision repair gene
MMS19 and interaction with transcription repair factor TFIIH via the XPB and XPD
helicases. Nucleic Acids Res. 28, 4506 (2000). doi:10.1093/nar/28.22.4506 Medline
33. O. Stehling et al., Investigation of iron-sulfur protein maturation in eukaryotes. Methods
Mol. Biol. 372, 325 (2007). doi:10.1007/978-1-59745-365-3_24 Medline

4
34. K. D. Schnackerz, D. Dobritzsch, Y. Lindqvist, P. F. Cook, Dihydropyrimidine
dehydrogenase: A flavoprotein with four iron-sulfur clusters. Biochim. Biophys. Acta
1701, 61 (2004). Medline
35. A. A. Vashisht et al., Control of iron homeostasis by an iron-regulated ubiquitin ligase.
Science 326, 718 (2009). doi:10.1126/science.1176333 Medline
36. A. Pellicioli et al., Activation of Rad53 kinase in response to DNA damage and its effect
in modulating phosphorylation of the lagging strand DNA polymerase. EMBO J. 18,
6561 (1999). doi:10.1093/emboj/18.22.6561 Medline
37. M. G. Benton, N. R. Glasser, S. P. Palecek, The utilization of a Saccharomyces cerevisiae
HUG1P-GFP promoter-reporter construct for the selective detection of DNA damage.
Mutat. Res. 633, 21 (2007). Medline
38. A. V. Goldberg et al., Localization and functionality of microsporidian iron-sulphur
cluster assembly proteins. Nature 452, 624 (2008). doi:10.1038/nature06606 Medline
39. F. Sherman, Getting started with yeast. Methods Enzymol. 350, 3 (2002).
doi:10.1016/S0076-6879(02)50954-X Medline
40. A. Wach, A. Brachat, C. Alberti-Segui, C. Rebischung, P. Philippsen, Heterologous HIS3
marker and GFP reporter modules for PCR-targeting in Saccharomyces cerevisiae.
Yeast 13, 1065 (1997). doi:10.1002/(SICI)1097-0061(19970915)13:11<1065::AID-
YEA159>3.0.CO;2-K Medline
41. C. Janke et al., A versatile toolbox for PCR-based tagging of yeast genes: New
fluorescent proteins, more markers and promoter substitution cassettes. Yeast 21, 947
(2004). doi:10.1002/yea.1142 Medline
42. A. Hausmann et al., The eukaryotic P loop NTPase Nbp35: An essential component of
the cytosolic and nuclear iron-sulfur protein assembly machinery. Proc. Natl. Acad.
Sci. U.S.A. 102, 3266 (2005). doi:10.1073/pnas.0406447102 Medline
43. L. Zheng, U. Baumann, J. L. Reymond, An efficient one-step site-directed and site-
saturation mutagenesis protocol. Nucleic Acids Res. 32, e115 (2004).
doi:10.1093/nar/gnh110 Medline
44. E. Harlow, D. Lane, Using Antibodies: A Laboratory Manual (Cold Spring Harbor
Laboratory, Cold Spring Harbor, NY, 1998).

5
45. T. W. Traut, S. Loechel, Pyrimidine catabolism: Individual characterization of the three
sequential enzymes with a new assay. Biochemistry 23, 2533 (1984).
doi:10.1021/bi00306a033 Medline
46. D. J. Porter, T. Spector, Dihydropyrimidine dehydrogenase. Kinetic mechanism for
reduction of uracil by NADPH. J. Biol. Chem. 268, 19321 (1993). Medline
47. S. Sadahiro et al., The stability of dihydropyrimidine dehydrogenase activity in colorectal
cancer tissue after resection. Oncol. Rep. 10, 867 (2003). Medline
48. A. B. Van Kuilenburg, H. Van Lenthe, A. Tromp, P. C. Veltman, A. H. Van Gennip,
Pitfalls in the diagnosis of patients with a partial dihydropyrimidine dehydrogenase
deficiency. Clin. Chem. 46, 9 (2000). Medline
49. O. Stehling, A. D. Sheftel, R. Lill, Controlled expression of iron-sulfur cluster assembly
components for respiratory chain complexes in mammalian cells. Methods Enzymol.
456, 209 (2009). doi:10.1016/S0076-6879(08)04412-1 Medline
50. A. D. Sheftel et al., Human ind1, an iron-sulfur cluster assembly factor for respiratory
complex I. Mol. Cell. Biol. 29, 6059 (2009). doi:10.1128/MCB.00817-09 Medline
51. M. P. Washburn, D. Wolters, J. R. Yates 3rd, Large-scale analysis of the yeast proteome
by multidimensional protein identification technology. Nat. Biotechnol. 19, 242
(2001). doi:10.1038/85686 Medline
52. D. A. Wolters, M. P. Washburn, J. R. Yates 3rd, An automated multidimensional protein
identification technology for shotgun proteomics. Anal. Chem. 73, 5683 (2001).
doi:10.1021/ac010617e Medline
53. D. L. Tabb, W. H. McDonald, J. R. Yates 3rd, DTASelect and Contrast: Tools for
assembling and comparing protein identifications from shotgun proteomics. J.
Proteome Res. 1, 21 (2002). doi:10.1021/pr015504q Medline
54. J. E. Elias, S. P. Gygi, Target-decoy search strategy for increased confidence in large-
scale protein identifications by mass spectrometry. Nat. Methods 4, 207 (2007).
doi:10.1038/nmeth1019 Medline
55. L. Florens et al., Analyzing chromatin remodeling complexes using shotgun proteomics
and normalized spectral abundance factors. Methods 40, 303 (2006).
doi:10.1016/j.ymeth.2006.07.028 Medline

6
56. J. A. Wohlschlegel, Identification of SUMO-conjugated proteins and their SUMO
attachment sites using proteomic mass spectrometry. Methods Mol. Biol. 497, 33
(2009). doi:10.1007/978-1-59745-566-4_3 Medline
57. A. Tzagoloff, C. L. Dieckmann, PET genes of Saccharomyces cerevisiae. Microbiol. Rev.
54, 211 (1990). Medline
58. U. Mühlenhoff et al., Functional characterization of the eukaryotic cysteine desulfurase
Nfs1p from Saccharomyces cerevisiae. J. Biol. Chem. 279, 36906 (2004).
doi:10.1074/jbc.M406516200 Medline
59. H. Lange, A. Kaut, G. Kispal, R. Lill, A mitochondrial ferredoxin is essential for
biogenesis of cellular iron-sulfur proteins. Proc. Natl. Acad. Sci. U.S.A. 97, 1050
(2000). doi:10.1073/pnas.97.3.1050 Medline
60. F. Foury, Low iron concentration and aconitase deficiency in a yeast frataxin homologue
deficient strain. FEBS Lett. 456, 281 (1999). doi:10.1016/S0014-5793(99)00961-8
Medline
61. U. Mühlenhoff, J. Gerber, N. Richhardt, R. Lill, Components involved in assembly and
dislocation of iron-sulfur clusters on the scaffold protein Isu1p. EMBO J. 22, 4815
(2003). doi:10.1093/emboj/cdg446 Medline
62. C. Baker Brachmann et al., Designer deletion strains derived from Saccharomyces
cerevisiae S288C: A useful set of strains and plasmids for PCR-mediated gene
disruption and other applications. Yeast 14, 115 (1998). doi:10.1002/(SICI)1097-
0061(19980130)14:2<115::AID-YEA204>3.0.CO;2-2 Medline
63. A. Biederbick et al., Role of human mitochondrial Nfs1 in cytosolic iron-sulfur protein
biogenesis and iron regulation. Mol. Cell. Biol. 26, 5675 (2006).
doi:10.1128/MCB.00112-06 Medline
64. O. Stehling, H. P. Elsässer, B. Brückel, U. Mühlenhoff, R. Lill, Iron-sulfur protein
maturation in human cells: Evidence for a function of frataxin. Hum. Mol. Genet. 13,
3007 (2004). doi:10.1093/hmg/ddh324 Medline


















