
Sistem Muskuloskeletal - Stovit2008's Blog | blog … Muskuloskeletal Semua fungsi fisik tubuh...
16
Sistem Muskuloskeletal Yuliati Departemen Biologi Oral
Transcript of Sistem Muskuloskeletal - Stovit2008's Blog | blog … Muskuloskeletal Semua fungsi fisik tubuh...

82 Unit II Membrane Physiology, Nerve, and Muscle
contraction may be as little as one half its strength 10to 50 muscle twitches later. That is, the strength of con-traction increases to a plateau, a phenomenon called thestaircase effect, or treppe.
Although all the possible causes of the staircase effectare not known, it is believed to be caused primarily byincreasing calcium ions in the cytosol because of therelease of more and more ions from the sarcoplasmicreticulum with each successive muscle action potentialand failure of the sarcoplasm to recapture the ionsimmediately.
Skeletal Muscle Tone. Even when muscles are at rest, acertain amount of tautness usually remains. This iscalled muscle tone. Because normal skeletal musclefibers do not contract without an action potential tostimulate the fibers, skeletal muscle tone results entirelyfrom a low rate of nerve impulses coming from thespinal cord. These, in turn, are controlled partly bysignals transmitted from the brain to the appropriatespinal cord anterior motoneurons and partly by signalsthat originate in muscle spindles located in the muscleitself. Both of these are discussed in relation to musclespindle and spinal cord function in Chapter 54.
Muscle Fatigue. Prolonged and strong contraction of amuscle leads to the well-known state of muscle fatigue.Studies in athletes have shown that muscle fatigueincreases in almost direct proportion to the rate ofdepletion of muscle glycogen. Therefore, fatigue resultsmainly from inability of the contractile and metabolicprocesses of the muscle fibers to continue supplying thesame work output. However, experiments have alsoshown that transmission of the nerve signal through theneuromuscular junction, which is discussed in Chapter7, can diminish at least a small amount after intense pro-longed muscle activity, thus further diminishing musclecontraction. Interruption of blood flow through a con-tracting muscle leads to almost complete muscle fatiguewithin 1 or 2 minutes because of the loss of nutrientsupply, especially loss of oxygen.
Lever Systems of the Body. Muscles operate by applyingtension to their points of insertion into bones, and thebones in turn form various types of lever systems. Figure6–14 shows the lever system activated by the bicepsmuscle to lift the forearm. If we assume that a large
biceps muscle has a cross-sectional area of 6 squareinches, the maximum force of contraction would beabout 300 pounds. When the forearm is at right angleswith the upper arm, the tendon attachment of the bicepsis about 2 inches anterior to the fulcrum at the elbow,and the total length of the forearm lever is about 14inches. Therefore, the amount of lifting power of thebiceps at the hand would be only one seventh of the 300pounds of muscle force, or about 43 pounds. When thearm is fully extended, the attachment of the biceps ismuch less than 2 inches anterior to the fulcrum, and theforce with which the hand can be brought forward isalso much less than 43 pounds.
In short, an analysis of the lever systems of the bodydepends on knowledge of (1) the point of muscle inser-tion, (2) its distance from the fulcrum of the lever, (3)the length of the lever arm, and (4) the position of thelever. Many types of movement are required in thebody, some of which need great strength and others of which need large distances of movement. For thisreason, there are many different types of muscle; someare long and contract a long distance, and some areshort but have large cross-sectional areas and canprovide extreme strength of contraction over short dis-tances. The study of different types of muscles, leversystems, and their movements is called kinesiologyand is an important scientific component of humanphysioanatomy.
“Positioning” of a Body Part by Contraction of Agonist and Antag-onist Muscles on Opposite Sides of a Joint—“Coactivation” of Antagonist Muscles. Virtually all body movements arecaused by simultaneous contraction of agonist andantagonist muscles on opposite sides of joints. This is called coactivation of the agonist and antagonistmuscles, and it is controlled by the motor control centersof the brain and spinal cord.
The position of each separate part of the body, suchas an arm or a leg, is determined by the relative degreesof contraction of the agonist and antagonist sets ofmuscles. For instance, let us assume that an arm or a legis to be placed in a midrange position. To achieve this, agonist and antagonist muscles are excited aboutequally. Remember that an elongated muscle contractswith more force than a shortened muscle, which wasdemonstrated in Figure 6–9, showing maximum strengthof contraction at full functional muscle length andalmost no strength of contraction at half normal length.Therefore, the elongated muscle on one side of a jointcan contract with far greater force than the shortermuscle on the opposite side. As an arm or leg movestoward its midposition, the strength of the longermuscle decreases, whereas the strength of the shortermuscle increases until the two strengths equal eachother. At this point, movement of the arm or leg stops.Thus, by varying the ratios of the degree of activationof the agonist and antagonist muscles, the nervoussystem directs the positioning of the arm or leg.
We learn in Chapter 54 that the motor nervoussystem has additional important mechanisms to com-pensate for different muscle loads when directing thispositioning process.
Remodeling of Muscle
to Match Function
All the muscles of the body are continually beingremodeled to match the functions that are required of
Figure 6–14
Lever system activated by the biceps muscle.
Sistem Muskuloskeletal
YuliatiDepartemen Biologi Oral
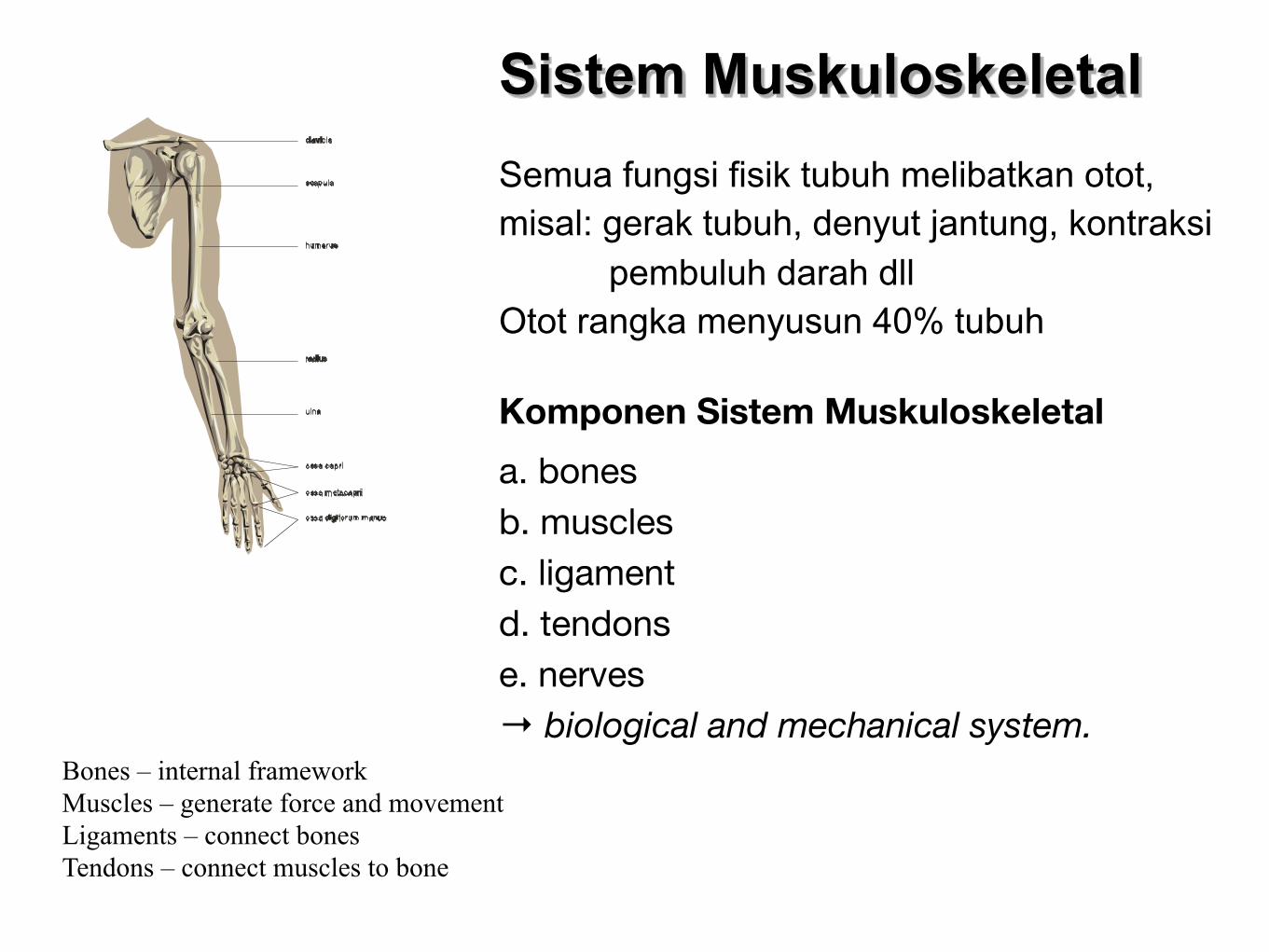
Sistem MuskuloskeletalSemua fungsi fisik tubuh melibatkan otot, misal: gerak tubuh, denyut jantung, kontraksi pembuluh darah dllOtot rangka menyusun 40% tubuh
Komponen Sistem Muskuloskeletal
a. bonesb. musclesc. ligamentd. tendonse. nerves→ biological and mechanical system.
Bones – internal frameworkMuscles – generate force and movementLigaments – connect bonesTendons – connect muscles to bone

Fungsi Sistem muskuloskeletal
1. Memberi bentuk tubuh,
2. Melindungi organ dalam,
3. Menyimpan mineral kalsium dan fosfat,
4. Tempat melekatnya otot,
5. Sebagai pengungkit (tuas) dalam pergerakan sendi,
6. Mengandung sumsum tulang tempat pembentukan sel darah,
7. Menghasilkan panas tubuh
Chapter 6 Contraction of Skeletal Muscle 73
interdigitate with the myosin filaments. The Z disc,which itself is composed of filamentous proteins dif-ferent from the actin and myosin filaments, passescrosswise across the myofibril and also crosswise frommyofibril to myofibril, attaching the myofibrils to oneanother all the way across the muscle fiber. Therefore,the entire muscle fiber has light and dark bands, as dothe individual myofibrils.These bands give skeletal andcardiac muscle their striated appearance.
The portion of the myofibril (or of the whole musclefiber) that lies between two successive Z discs is calleda sarcomere. When the muscle fiber is contracted, asshown at the bottom of Figure 6–4, the length of the sarcomere is about 2 micrometers. At this length,the actin filaments completely overlap the myosin filaments, and the tips of the actin filaments are just
beginning to overlap one another. We will see laterthat, at this length, the muscle is capable of generatingits greatest force of contraction.
What Keeps the Myosin and Actin Filaments in Place?
Titin Filamentous Molecules. The side-by-side rela-tionship between the myosin and actin filaments is dif-ficult to maintain. This is achieved by a large numberof filamentous molecules of a protein called titin. Eachtitin molecule has a molecular weight of about 3million, which makes it one of the largest protein mol-ecules in the body. Also, because it is filamentous, it isvery springy. These springy titin molecules act as aframework that holds the myosin and actin filamentsin place so that the contractile machinery of the sar-comere will work. There is reason to believe that the
G-Actin molecules
F-Actin filament
Myofilaments
Myosin filament
Myosin molecule
Lightmeromyosin
Heavymeromyosin
Muscle fasciculus
SKELETAL MUSCLE
Muscle
Muscle fiber
Myofibril
Hband
Zdisc
Aband
Sarcomere Z
H
D
E
F G H I
N
M
L
K
J
B
C
Z
Iband
A
Figure 6–1
Organization of skeletal muscle,from the gross to the molecularlevel. F, G, H, and I are crosssections at the levels indicated.(Drawing by Sylvia ColardKeene. Modified from FawcettDW: Bloom and Fawcett: A Text-book of Histology. Philadelphia:WB Saunders, 1986.)
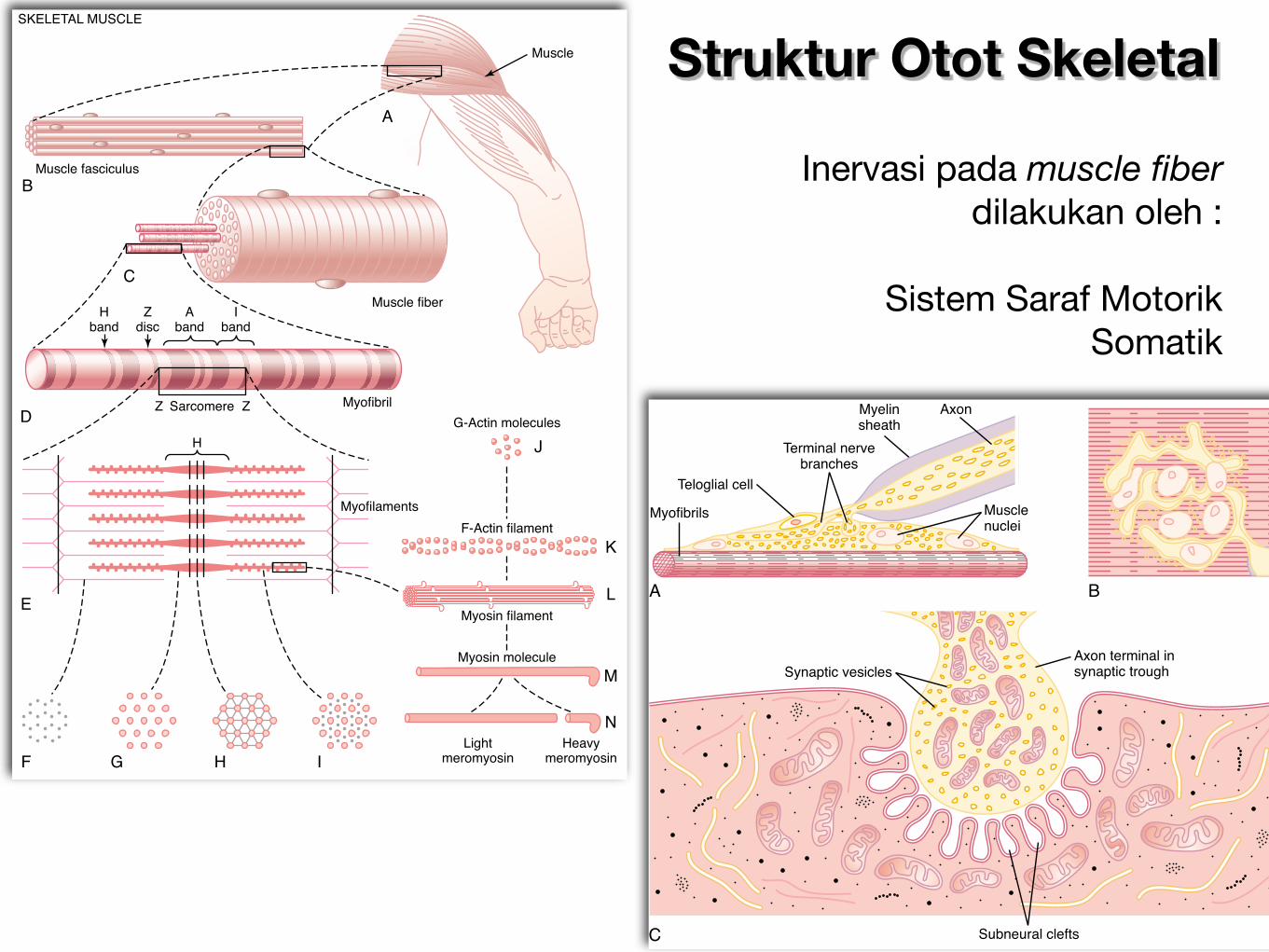
Struktur Otot Skeletal
86 Unit II Membrane Physiology, Nerve, and Muscle
On the inside surface of the neural membrane arelinear dense bars, shown in cross section in Figure 7–2.To each side of each dense bar are protein particlesthat penetrate the neural membrane; these are voltage-gated calcium channels. When an action potentialspreads over the terminal, these channels open andallow calcium ions to diffuse from the synaptic spaceto the interior of the nerve terminal. The calcium ions,in turn, are believed to exert an attractive influence onthe acetylcholine vesicles, drawing them to the neuralmembrane adjacent to the dense bars. The vesiclesthen fuse with the neural membrane and empty theiracetylcholine into the synaptic space by the process ofexocytosis.
Although some of the aforementioned details arespeculative, it is known that the effective stimulus forcausing acetylcholine release from the vesicles is entryof calcium ions and that acetylcholine from the vesi-cles is then emptied through the neural membraneadjacent to the dense bars.
Effect of Acetylcholine on the Postsynaptic Muscle Fiber Mem-brane to Open Ion Channels. Figure 7–2 also shows manyvery small acetylcholine receptors in the muscle fibermembrane; these are acetylcholine-gated ion channels,and they are located almost entirely near the mouthsof the subneural clefts lying immediately below thedense bar areas, where the acetylcholine is emptiedinto the synaptic space.
Each receptor is a protein complex that has a totalmolecular weight of 275,000.The complex is composed
Axon
Myofibrils
A
C
B
Teloglial cell
Terminal nervebranches
Musclenuclei
Myelinsheath
Synaptic vesiclesAxon terminal insynaptic trough
Subneural clefts
Figure 7–1
Different views of the motor endplate. A, Longitudinal sectionthrough the end plate. B, Surfaceview of the end plate. C, Electronmicrographic appearance of thecontact point between a singleaxon terminal and the musclefiber membrane. (Redrawn fromFawcett DW, as modified fromCouteaux R, in Bloom W, FawcettDW: A Textbook of Histology.Philadelphia: WB Saunders,1986.)
Calciumchannels
Neuralmembrane
Musclemembrane
Releasesites
Vesicles
Dense bar
Basal laminaandacetylcholinesterase
Acetylcholinereceptors
Subneural cleft
Figure 7–2
Release of acetylcholine from synaptic vesicles at the neuralmembrane of the neuromuscular junction. Note the proximity ofthe release sites in the neural membrane to the acetylcholinereceptors in the muscle membrane, at the mouths of the subneuralclefts.
Inervasi pada muscle fiber dilakukan oleh :
Sistem Saraf Motorik Somatik
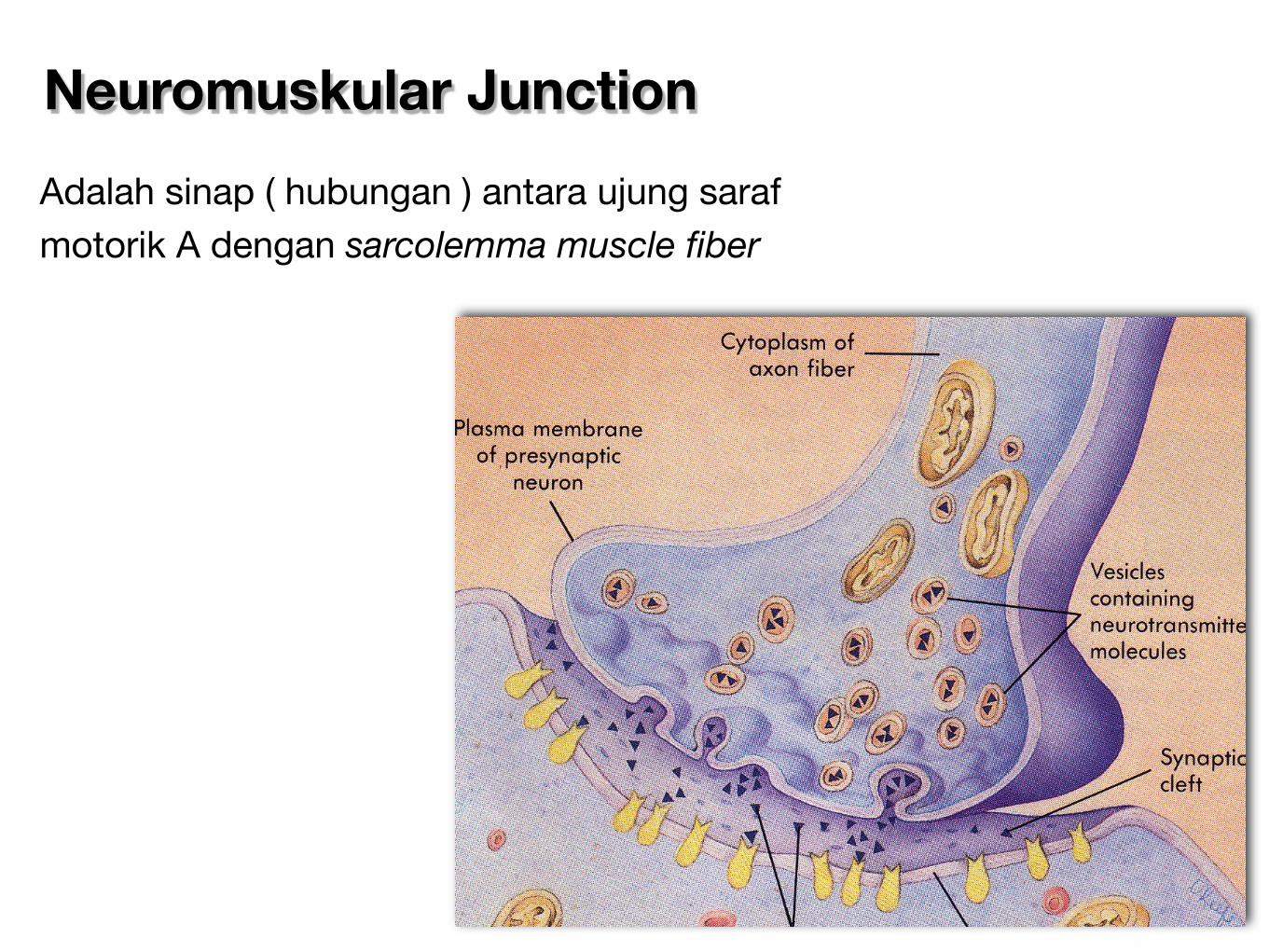
Neuromuskular Junction
Adalah sinap ( hubungan ) antara ujung saraf motorik A dengan sarcolemma muscle fiber
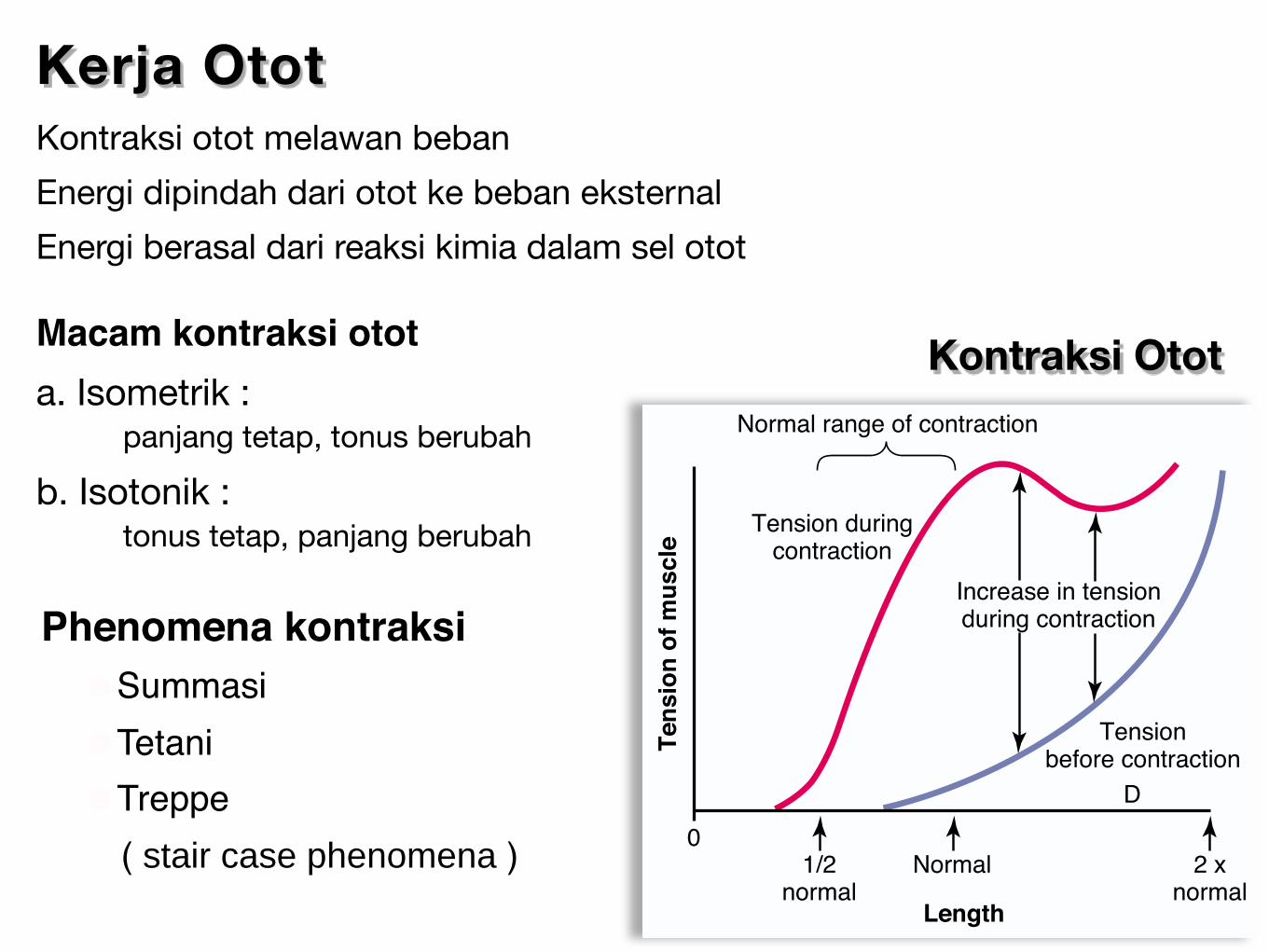
Kontraksi Otot
Kerja Otot Kontraksi otot melawan beban
Energi dipindah dari otot ke beban eksternal
Energi berasal dari reaksi kimia dalam sel otot
Macam kontraksi otota. Isometrik : panjang tetap, tonus berubah
b. Isotonik : tonus tetap, panjang berubah
78 Unit II Membrane Physiology, Nerve, and Muscle
further shortening, the sarcomere maintains fulltension until point B is reached, at a sarcomere lengthof about 2 micrometers. At this point, the ends of thetwo actin filaments begin to overlap each other inaddition to overlapping the myosin filaments. As thesarcomere length falls from 2 micrometers down toabout 1.65 micrometers, at point A, the strength ofcontraction decreases rapidly. At this point, the two Zdiscs of the sarcomere abut the ends of the myosin fil-aments. Then, as contraction proceeds to still shortersarcomere lengths, the ends of the myosin filamentsare crumpled and, as shown in the figure, the strengthof contraction approaches zero, but the entire musclehas now contracted to its shortest length.
Effect of Muscle Length on Force of Contraction in the WholeIntact Muscle. The top curve of Figure 6–9 is similar tothat in Figure 6–8, but the curve in Figure 6–9 depictstension of the intact, whole muscle rather than of asingle muscle fiber. The whole muscle has a largeamount of connective tissue in it; also, the sarcomeresin different parts of the muscle do not always contractthe same amount. Therefore, the curve has somewhatdifferent dimensions from those shown for the individual muscle fiber, but it exhibits the same generalform for the slope in the normal range of contraction,as noted in Figure 6–9.
Note in Figure 6–9 that when the muscle is at itsnormal resting length, which is at a sarcomere lengthof about 2 micrometers, it contracts upon activationwith the approximate maximum force of contraction.However, the increase in tension that occurs duringcontraction, called active tension, decreases as the
muscle is stretched beyond its normal length—that is,to a sarcomere length greater than about 2.2 microm-eters. This is demonstrated by the decreased length ofthe arrow in the figure at greater than normal musclelength.
Relation of Velocity of Contractionto Load
A skeletal muscle contracts extremely rapidly when itcontracts against no load—to a state of full contractionin about 0.1 second for the average muscle. When loadsare applied, the velocity of contraction becomes pro-gressively less as the load increases, as shown in Figure6–10. That is, when the load has been increased to equalthe maximum force that the muscle can exert, the veloc-ity of contraction becomes zero and no contractionresults, despite activation of the muscle fiber.
This decreasing velocity of contraction with load iscaused by the fact that a load on a contracting muscleis a reverse force that opposes the contractile forcecaused by muscle contraction. Therefore, the net forcethat is available to cause velocity of shortening is correspondingly reduced.
Energetics of MuscleContraction
Work Output During MuscleContraction
When a muscle contracts against a load, it performswork. This means that energy is transferred from themuscle to the external load to lift an object to a greaterheight or to overcome resistance to movement.
In mathematical terms, work is defined by the following equation:
3 4210
100
50
0
Length of sarcomere (micrometers)
A
B CD
C
B
A
D
Ten
sio
n d
evelo
ped
(per
cen
t)
Figure 6–8
Length-tension diagram for a single fully contracted sarcomere,showing maximum strength of contraction when the sarcomere is2.0 to 2.2 micrometers in length. At the upper right are the rela-tive positions of the actin and myosin filaments at different sar-comere lengths from point A to point D. (Modified from GordonAM, Huxley AF, Julian FJ: The length-tension diagram of singlevertebrate striated muscle fibers. J Physiol 171:28P, 1964.)
0
Length
Ten
sio
n o
f m
uscle
1/2normal
2 xnormal
Normal
Increase in tensionduring contraction
Tension duringcontraction
Normal range of contraction
Tensionbefore contraction
D
Figure 6–9
Relation of muscle length to tension in the muscle both before andduring muscle contraction.
Phenomena kontraksi ☻Summasi ☻Tetani ☻Treppe ( stair case phenomena )

Mikroskopis tulang : bahan organik (30%) & mineral (70%)
1. Bahan organik tulang
a. Matrik tulang : 98% bhn organik tulang kolagen (95%) protein non-kolagen (5%) b. Sel tulang : sebesar 2% osteoblas, osteosit dan osteoklas
Komponen Pembentuk Tulang
2. Mineral
95% terdiri dari hidroksiapatit kristal kalsium fosfat.
Tulang
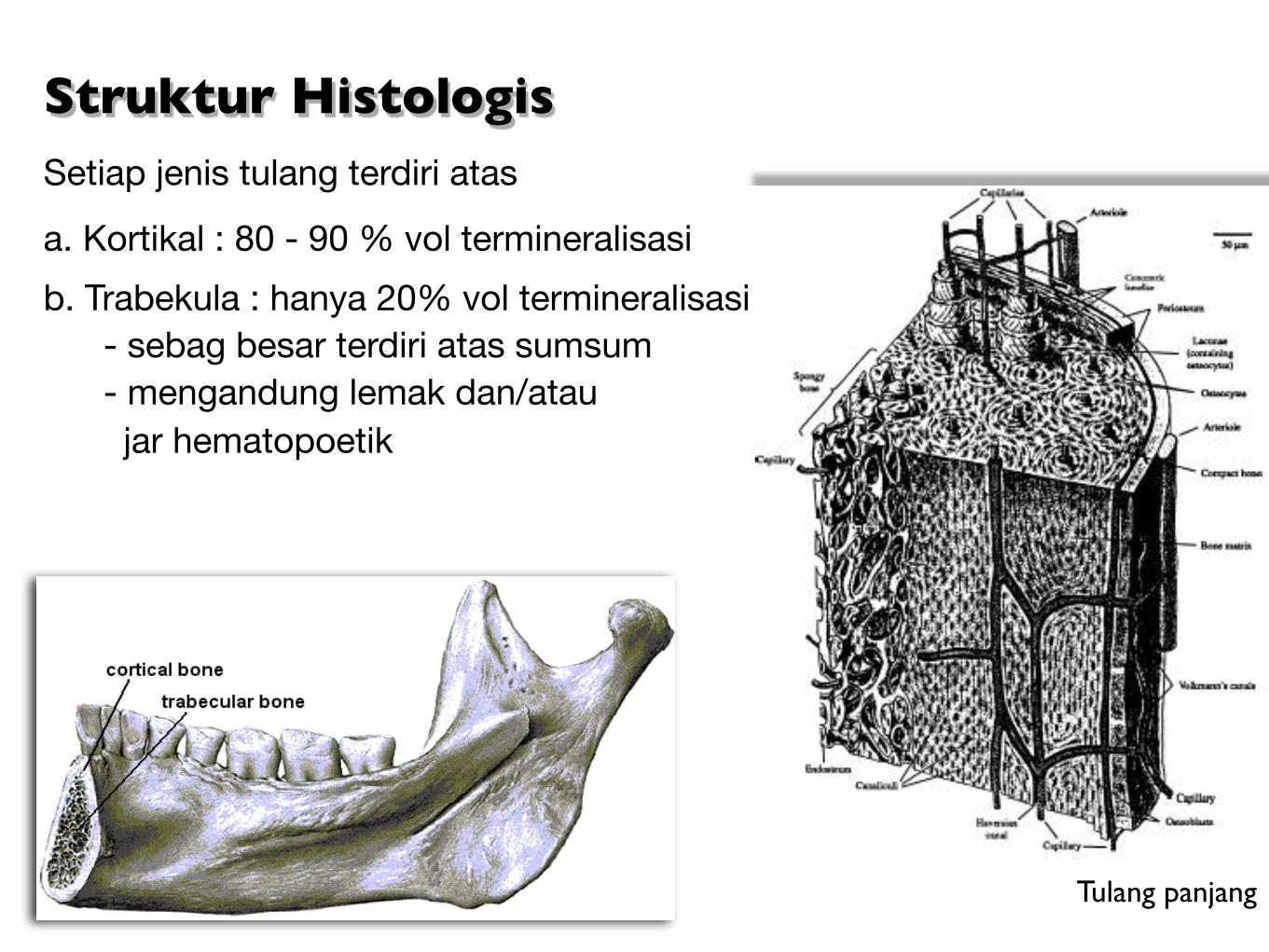
Struktur HistologisSetiap jenis tulang terdiri atas
a. Kortikal : 80 - 90 % vol termineralisasi
b. Trabekula : hanya 20% vol termineralisasi - sebag besar terdiri atas sumsum - mengandung lemak dan/atau jar hematopoetik
Tulang panjang
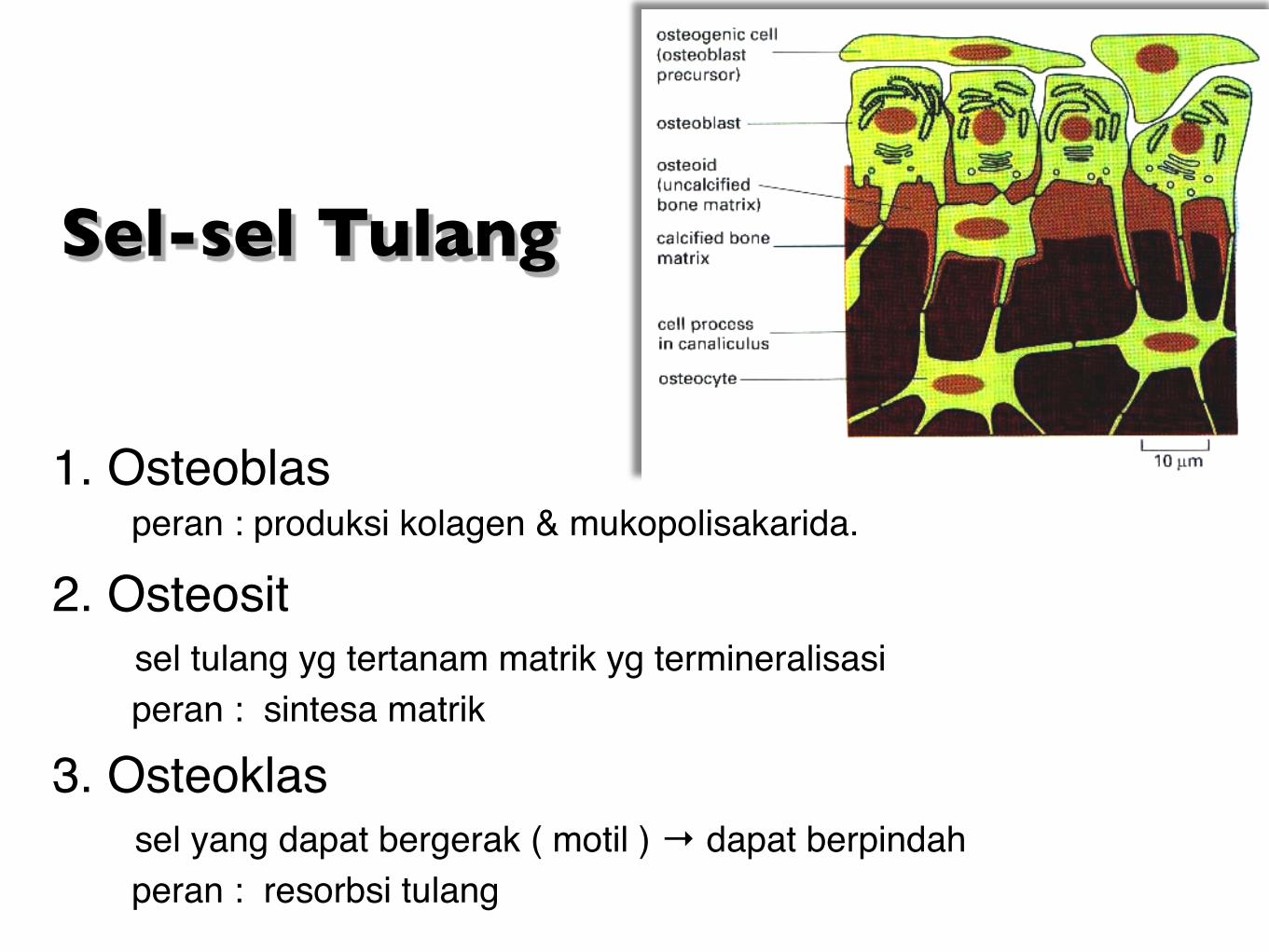
Sel-sel Tulang
1. Osteoblas peran : produksi kolagen & mukopolisakarida.
2. Osteosit sel tulang yg tertanam matrik yg termineralisasi peran : sintesa matrik 3. Osteoklas sel yang dapat bergerak ( motil ) → dapat berpindah peran : resorbsi tulang
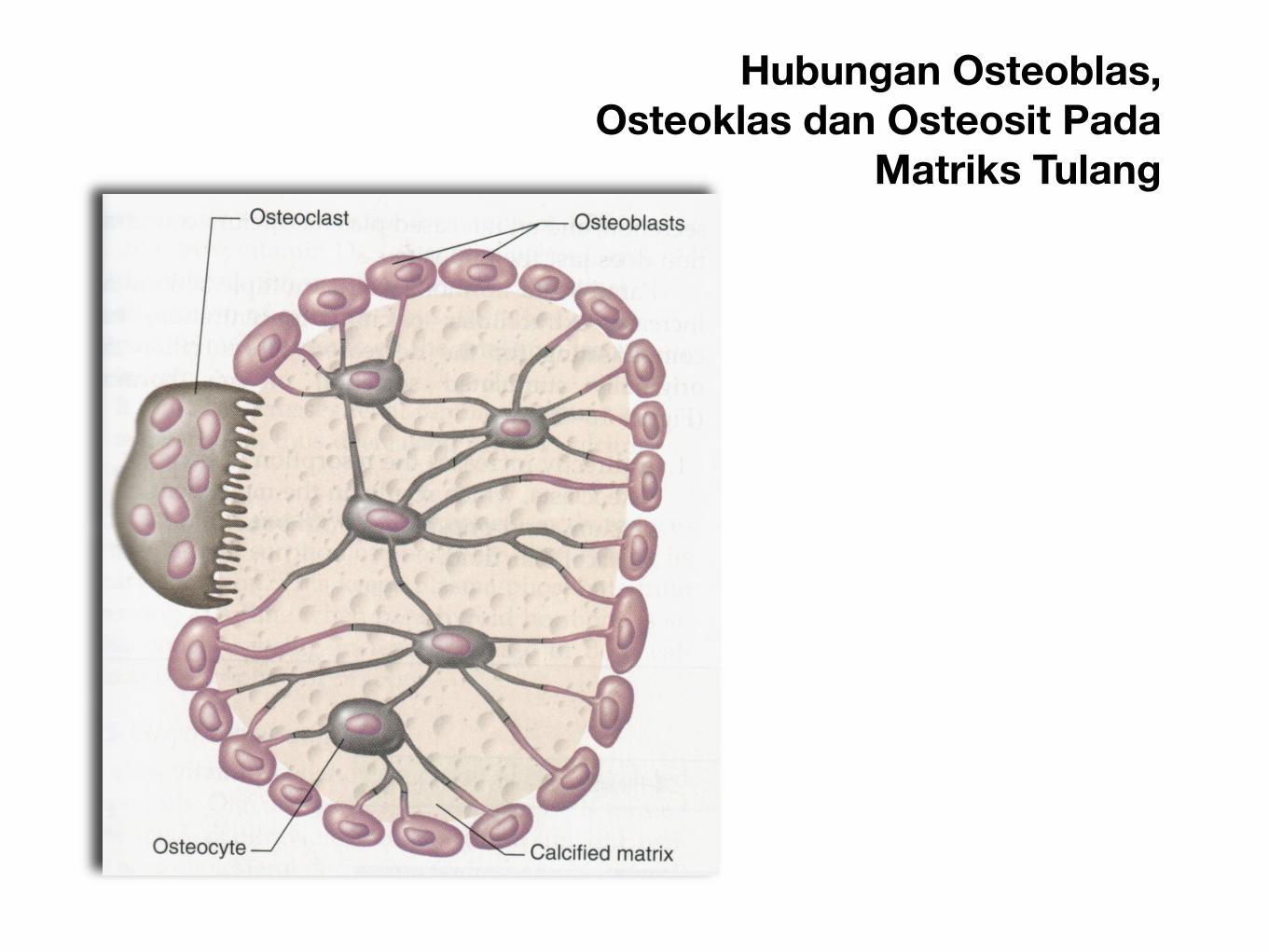
Hubungan Osteoblas, Osteoklas dan Osteosit Pada
Matriks Tulang
Chapter 84 Sports Physiology 1057
still another 7300 calories become available. Removalof the first phosphate converts the ATP into adenosinediphosphate (ADP), and removal of the second convertsthis ADP into adenosine monophosphate (AMP).
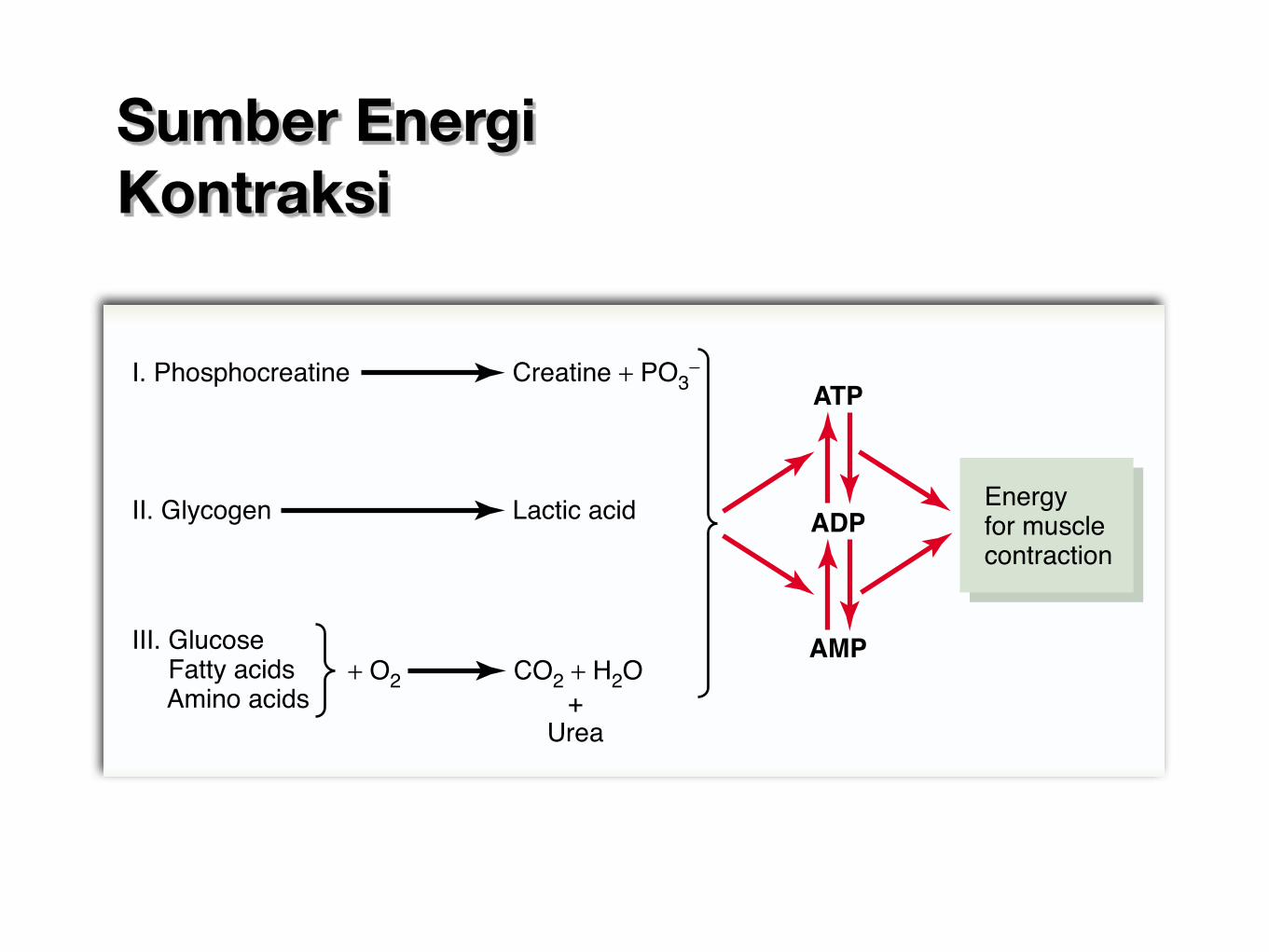
The amount of ATP present in the muscles, even in awell-trained athlete, is sufficient to sustain maximalmuscle power for only about 3 seconds, maybe enoughfor one half of a 50-meter dash. Therefore, except for afew seconds at a time, it is essential that new ATP beformed continuously, even during the performance ofshort athletic events. Figure 84–1 shows the overallmetabolic system, demonstrating the breakdown ofATP first to ADP and then to AMP, with the release ofenergy to the muscles for contraction.The left-hand sideof the figure shows the three metabolic systems thatprovide a continuous supply of ATP in the muscle fibers.
Phosphocreatine-Creatine System
Phosphocreatine (also called creatine phosphate) isanother chemical compound that has a high-energyphosphate bond, with the following formula:
Creatine ~ PO3-
This can decompose to creatine and phosphate ion, asshown to the left in Figure 84–1, and in doing so releaselarge amounts of energy. In fact, the high-energy phos-phate bond of phosphocreatine has more energy thanthe bond of ATP, 10,300 calories per mole in compari-son with 7300. Therefore, phosphocreatine can easilyprovide enough energy to reconstitute the high-energybond of ATP. Furthermore, most muscle cells have twoto four times as much phosphocreatine as ATP.
A special characteristic of energy transfer from phos-phocreatine to ATP is that it occurs within a small frac-tion of a second. Therefore, all the energy stored in themuscle phosphocreatine is almost instantaneously avail-able for muscle contraction, just as is the energy storedin ATP.
The combined amounts of cell ATP and cell phos-phocreatine are called the phosphagen energy system.These together can provide maximal muscle power for8 to 10 seconds, almost enough for the 100-meter run.Thus, the energy from the phosphagen system is used formaximal short bursts of muscle power.
Glycogen-Lactic Acid System. The stored glycogen inmuscle can be split into glucose and the glucose thenused for energy. The initial stage of this process, called
glycolysis, occurs without use of oxygen and, therefore,is said to be anaerobic metabolism (see Chapter 67).During glycolysis, each glucose molecule is split into twopyruvic acid molecules, and energy is released to formfour ATP molecules for each original glucose molecule,as explained in Chapter 67. Ordinarily, the pyruvic acid then enters the mitochondria of the muscle cells and reacts with oxygen to form still many more ATP molecules. However, when there is insufficientoxygen for this second stage (the oxidative stage) ofglucose metabolism to occur, most of the pyruvic acidthen is converted into lactic acid, which diffuses out ofthe muscle cells into the interstitial fluid and blood.Therefore, much of the muscle glycogen is transformedto lactic acid, but in doing so, considerable amounts ofATP are formed entirely without the consumption ofoxygen.
Another characteristic of the glycogen-lactic acidsystem is that it can form ATP molecules about 2.5 times as rapidly as can the oxidative mechanism of themitochondria. Therefore, when large amounts of ATPare required for short to moderate periods of musclecontraction, this anaerobic glycolysis mechanism can beused as a rapid source of energy. It is, however, onlyabout one half as rapid as the phosphagen system.Under optimal conditions, the glycogen-lactic acidsystem can provide 1.3 to 1.6 minutes of maximalmuscle activity in addition to the 8 to 10 seconds pro-vided by the phosphagen system, although at somewhatreduced muscle power.
Aerobic System. The aerobic system is the oxidation offoodstuffs in the mitochondria to provide energy. Thatis, as shown to the left in Figure 84–1, glucose, fatty acids,and amino acids from the foodstuffs—after some inter-mediate processing—combine with oxygen to releasetremendous amounts of energy that are used to convertAMP and ADP into ATP, as discussed in Chapter 67.
In comparing this aerobic mechanism of energysupply with the glycogen-lactic acid system and thephosphagen system, the relative maximal rates of powergeneration in terms of moles of ATP generation perminute are the following:
I. Phosphocreatine Creatine + PO3-
II. Glycogen Lactic acid
ATP
Energyfor musclecontraction
ADP
AMP
+Urea
III. Glucose Fatty acids Amino acids
CO2 + H2O+ O2Figure 84–1
Important metabolic systems that supply energy for musclecontraction.
Moles of ATP/min
Phosphagen system 4Glycogen–lactic acid system 2.5Aerobic system 1
Sumber Energi Kontraksi

Metabolisme Aerob
• Tjd peristiwa oksidasi reaksi dg O2
• Energi hasil reaksi oksidasi tiap bhn mknan ( kh, prot., lemak ) dg 1 liter O2 : a. karbohidrat 5 kkal b. lemak 4,7 kkal c. protein 4,5 kkal
Reaksi : glukosa CO2 + H2O + 36 ATP O2

• Metabolisme tanpa O2
• Fungsi : penyediaan energi mendesak
• Hanya berlangsung pd karbohidrat ATP lebih <
Reaksi : glukosa as.laktat + 2 ATP
Asam Laktat
• M’pengaruhi pH cair tubuh kerja enzim terganggu Diubah as.Piruvat
- dibakar/oksidasi (20%) - diubah jadi glukosa (80%) energi metab.aerob
Metabolisme Anaerob

Kelelahan Otot Muscle Fatique
• Otot bekerja maksimal : posisi keseimbangan, misalnya : kepala, leher dan tulang belakang berada pd satu garis lurus
→ pengaruh gaya gravitasi
• Akibat kontraksi otot terus menerus dalam waktu yang lama.
• Kontraksi makin lemah akibat kekurangan ATP

Tipe kelelahan
• Transmission fatigue : kegagalan hub saraf-otot dalam menghantarkan impuls. Penyebab : turunnya sekresi neurotransmiter atau daya ikat reseptor neurotransmiter
• Central fatigue : sistem saraf pusat tidak dapat menyalurkan impuls pada saraf motorik
• Muscle fatigue : kurangnya energi untuk kontraksi ( ATP, kreatin fosfat ), terjadi akumulasi asam laktat

Sampai Bertemu lagi..


















