
Selectively reduced expression of thick ascending limb Tamm-Horsfall protein in hypothyroid kidneys
9
Histochem Cell Biol (2004) 121:319–327 DOI 10.1007/s00418-004-0638-4 ORIGINAL PAPER Roland Schmitt · Thomas Kahl · Kerim Mutig · Sebastian Bachmann Selectively reduced expression of thick ascending limb Tamm-Horsfall protein in hypothyroid kidneys Accepted: 17 February 2004 / Published online: 9 March 2004 # Springer-Verlag 2004 Abstract Hypothyroidism causes major changes in the kidney by affecting its growth, structure, function, and biosynthesis of its specific gene products. As a conse- quence, the nephron shows a selective hypotrophy in the medullary portion of the thick ascending limb (TAL). Contrastingly, we have shown earlier that the abundance of the major ion transporter of the TAL, the furosemide- sensitive Na-K-2Cl cotransporter (NKCC2) was increased when related to kidney function in the hypothyroid organism. Synthesis of Tamm-Horsfall protein (THP), the most abundant urinary protein produced in the TAL, has been suggested to interfere with TAL function. We therefore studied the localization and synthesis rate of THP in the hypothyroid kidney. We used rats chronically treated with methimazole. Kidneys were processed for western blotting and histochemical evaluation. Morpho- logically the hypothyroid TAL displayed a selectively reduced epithelial cell height of its medullary portion. This was paralleled by decreased THP immunostaining and mRNA abundance. Western blotting indicated a 40% reduction in renal THP content (P<0.005), and daily urinary THP excretion was 68% lower than in controls (P<0.05). T3 substitution restored these parameters. To further confirm that changes were specific for THP and not merely the consequence of reduced kidney growth, the abundance of barttin, another distal tubular protein related to chloride transport, was tested as well. Barttin was increased by 43% in the hypothyroid rats. Together with our previous results showing increased NKCC2 expression in hypothyroidism, these results demonstrate a selective decrease in medullary THP synthesis. We suggest a potential involvement of THP in the renal functional changes associated with hypothyroidism. Keywords Tamm-Horsfall protein · Hypothyroidism · Barttin · Thick ascending limb Introduction Hypothyroidism is a common clinical disorder that exerts marked effects on various organs including kidney. Studies in man and in experimental animals have revealed functional alterations such as a decreased glomerular filtration rate (GFR) and diminished urinary concentrating and diluting ability along with altered tubular sodium reabsorption (review in Bradley et al. 1974). T3 depriva- tion during adolescence further results in decreased kidney growth and distinct structural changes of the proximal and distal parts of the nephron (Bradley et al. 1982; Davis et al. 1983; Bentley et al. 1985; Schmitt et al. 2003). Changes in the diluting segment (thick ascending limb; TAL) are prominent and are located chiefly in its medullary portion (mTAL). Although the molecular background of these changes has not been evaluated, a resulting functional impairment of the mTAL may well interfere with the defective urine concentration in the hypothyroid state (Bentley et al. 1985). Besides express- ing a variety of transport-related proteins, the TAL is the exclusive site of expression of the highly abundant Tamm-Horsfall protein (THP; Hoyer et al. 1979; Bach- mann et al. 1985, 1990). Although the physiological role of THP is presently still unclear, an association with functional properties of the TAL epithelium appears plausible (reviewed in Kokot and Dulawa 2000). The normal TAL epithelium constitutively synthesizes THP. Under pathological conditions such as diabetic nephrop- athy, hyperprostaglandin E syndrome, polycystic kidney disease, and active lupus nephritis, however, the biosyn- thesis or urinary excretion of THP was altered (Lynn and Marshall 1984; SchrɆter et al. 1993; Rasch et al. 1995; R. Schmitt · T. Kahl · K. Mutig · S. Bachmann ( ) ) Institut fɒr Anatomie, Campus CharitȖ-Mitte, CharitȖ–UniversitȨtsmedizin Berlin, Philippstrasse 12, 10115 Berlin, Germany e-mail: [email protected] Tel.: +49-30-450528001 Fax: +49-30-450528922 R. Schmitt Campus CharitȖ-Buch, HELIOS Kliniken Berlin, Wiltbergstrasse 50, 13125 Berlin, Germany
-
Upload
roland-schmitt -
Category
Documents
-
view
215 -
download
2
Transcript of Selectively reduced expression of thick ascending limb Tamm-Horsfall protein in hypothyroid kidneys

Histochem Cell Biol (2004) 121:319–327DOI 10.1007/s00418-004-0638-4
O R I G I N A L P A P E R
Roland Schmitt · Thomas Kahl · Kerim Mutig ·Sebastian Bachmann
Selectively reduced expressionof thick ascending limb Tamm-Horsfall proteinin hypothyroid kidneysAccepted: 17 February 2004 / Published online: 9 March 2004� Springer-Verlag 2004
Abstract Hypothyroidism causes major changes in thekidney by affecting its growth, structure, function, andbiosynthesis of its specific gene products. As a conse-quence, the nephron shows a selective hypotrophy in themedullary portion of the thick ascending limb (TAL).Contrastingly, we have shown earlier that the abundanceof the major ion transporter of the TAL, the furosemide-sensitive Na-K-2Cl cotransporter (NKCC2) was increasedwhen related to kidney function in the hypothyroidorganism. Synthesis of Tamm-Horsfall protein (THP), themost abundant urinary protein produced in the TAL, hasbeen suggested to interfere with TAL function. Wetherefore studied the localization and synthesis rate ofTHP in the hypothyroid kidney. We used rats chronicallytreated with methimazole. Kidneys were processed forwestern blotting and histochemical evaluation. Morpho-logically the hypothyroid TAL displayed a selectivelyreduced epithelial cell height of its medullary portion.This was paralleled by decreased THP immunostainingand mRNA abundance. Western blotting indicated a 40%reduction in renal THP content (P<0.005), and dailyurinary THP excretion was 68% lower than in controls(P<0.05). T3 substitution restored these parameters. Tofurther confirm that changes were specific for THP andnot merely the consequence of reduced kidney growth,the abundance of barttin, another distal tubular proteinrelated to chloride transport, was tested as well. Barttinwas increased by 43% in the hypothyroid rats. Togetherwith our previous results showing increased NKCC2expression in hypothyroidism, these results demonstrate a
selective decrease in medullary THP synthesis. Wesuggest a potential involvement of THP in the renalfunctional changes associated with hypothyroidism.
Keywords Tamm-Horsfall protein · Hypothyroidism ·Barttin · Thick ascending limb
Introduction
Hypothyroidism is a common clinical disorder that exertsmarked effects on various organs including kidney.Studies in man and in experimental animals have revealedfunctional alterations such as a decreased glomerularfiltration rate (GFR) and diminished urinary concentratingand diluting ability along with altered tubular sodiumreabsorption (review in Bradley et al. 1974). T3 depriva-tion during adolescence further results in decreasedkidney growth and distinct structural changes of theproximal and distal parts of the nephron (Bradley et al.1982; Davis et al. 1983; Bentley et al. 1985; Schmitt et al.2003). Changes in the diluting segment (thick ascendinglimb; TAL) are prominent and are located chiefly in itsmedullary portion (mTAL). Although the molecularbackground of these changes has not been evaluated, aresulting functional impairment of the mTAL may wellinterfere with the defective urine concentration in thehypothyroid state (Bentley et al. 1985). Besides express-ing a variety of transport-related proteins, the TAL is theexclusive site of expression of the highly abundantTamm-Horsfall protein (THP; Hoyer et al. 1979; Bach-mann et al. 1985, 1990). Although the physiological roleof THP is presently still unclear, an association withfunctional properties of the TAL epithelium appearsplausible (reviewed in Kokot and Dulawa 2000). Thenormal TAL epithelium constitutively synthesizes THP.Under pathological conditions such as diabetic nephrop-athy, hyperprostaglandin E syndrome, polycystic kidneydisease, and active lupus nephritis, however, the biosyn-thesis or urinary excretion of THP was altered (Lynn andMarshall 1984; Schr�ter et al. 1993; Rasch et al. 1995;
R. Schmitt · T. Kahl · K. Mutig · S. Bachmann ())Institut f�r Anatomie, Campus Charit�-Mitte,Charit�–Universit�tsmedizin Berlin,Philippstrasse 12, 10115 Berlin, Germanye-mail: [email protected].: +49-30-450528001Fax: +49-30-450528922
R. SchmittCampus Charit�-Buch,HELIOS Kliniken Berlin,Wiltbergstrasse 50, 13125 Berlin, Germany

Tsai et al. 2000), and recent studies have identified linksbetween mutations in the THP gene and renal failureassociated with medullary cystic kidney disease, familialjuvenile hyperuricemic nephropathy, and volume con-traction (Hart et al. 2002; Rampoldi et al. 2003).
In the present study we addressed the question whetherhypothyroidism in rats is associated with changes in theexpression or cellular distribution of THP to advance theunderstanding of mechanisms potentially involved inhypothyroid functional impairment. To this end rats wererendered hypothyroid by adding methimazole to thedrinking water (Schmitt et al. 2003). THP expression andurinary excretion were analyzed by western blot, immu-nohistochemistry, and in situ hybridization. In order toestablish the specificity of changes in THP expression,another product of the distal nephron, barttin, the b-subunit of the ClC-K chloride channels (Birkenhager etal. 2001; Estevez et al. 2001), was studied.
Materials and methods
Experimental animals
The animals used in this study were the same as used in ourpreviously published study on sodium transporters in hypothyroidrats (Schmitt et al. 2003). Briefly, male Sprague-Dawley rats(Sch�nwalde, Berlin, Germany) weighing 150 g were dividedrandomly into two groups (each n=8). Both groups were fed thesame standard chow and had free access to drinking water.Hypothyroidism was induced by adding 0.05% methimazole to thewater in one group. After 7 weeks, urine was collected for 20 h byplacing the rats into individual metabolic cages. Thereafter, the ratswere in vivo perfusion-fixed for immunohistochemistry (n=6; seebelow). For biochemical analysis of serum and kidney parametersrats (n=10) were killed by an overdose of pentobarbital. Blood wascollected by puncture of the abdominal aorta. Both kidneys werequickly removed, weighed, decapsulated, cut into small pieces,frozen in liquid nitrogen, and stored at �80�C. To exclude a directeffect of methimazole, an additional group of rats (n=5) receiving0.05% methimazole for 4 weeks was substituted with T3 bysubcutaneous implantation of 0.05 mg T3-releasing pellets (IRA,Sarasota, FL, USA); pellets were implanted in the second week todeliver T3 during the following 3 weeks.
Blood and urine analysis
Thyroid hormones were measured in plasma samples using achemiluminescent immunoassay (Centaur Analyzer; Bayer, Ger-many). Serum and urine electrolytes were determined by indirection-selective electrode analysis (Modular Analytics; Roche Diag-nostics, Mannheim, Germany). Creatinine concentrations weredetermined by the kinetic method of Jaff� (Roche Diagnostics).Creatinine clearance and fractional sodium excretion were calcu-lated using standard equations.
Tissue preparation for immunoblotting
The tissue was homogenized for 3 min using a tissue homogenizer(Heidolph Diax 600, Germany) in ice-cold homogenization solutioncontaining 250 mM sucrose, 10 mM triethanolamine, and onetablet/50 ml of protease inhibitor cocktail (Complete; RocheDiagnostics). The kidney homogenates were centrifuged at1,000 g for 10 min to remove nuclei and incompletely homogenizedmembrane fragments. The supernatant was then centrifuged at
200,000 g for 1 h, producing a pellet containing membranefractions enriched in plasma membranes and intracellular vesicles.The 1,000-g centrifugation was carried out using a Sigma 3K15refrigerated centrifuge. The 200,000-g spin was carried out with aBeckman LE 80 ultracentrifuge. The resulting pellets were resus-pended in homogenization solution, and total protein concentrationwas measured using the Pierce BCA Protein Assay reagent kit(Pierce, Rockford, IL, USA).
Antibodies
Rabbit polyclonal anti-THP antibody was a gift of J.R. Hoyer(Philadelphia) and has been used in previous studies (Bachmann etal. 1991). Anti-barttin antibody was characterized previously(Estevez et al. 2001).
Protein electrophoresis and immunoblotting
Proteins of the tissue preparations were added to Laemmli’s bufferand then solubilized at 65�C for 10 min. Twenty micrograms ofeach sample was loaded on 8–10% polyacrylamide gels. Equity inprotein loading and blotting was verified by membrane stainingusing 0.1% Ponceau red. Urine samples were centrifuged at 5,000 gfor 5 min, the pellets discarded, and supernatants mixed with equalamounts of Laemmli’s buffer. Ten microliters of each sample wereapplied to 8–10% polyacrylamide gels, electrophoresed, andtransferred to PVDF membranes. Membranes were probed withanti-THP or anti-barttin antibody at room temperature for 2 h andthen exposed to horseradish peroxidase-conjugated secondaryantibody (Dako, Glostrup, Denmark) for 45 min. Immunoreactivebands were detected by chemiluminescence, using an EnhancedChemiluminescence kit (Amersham Pharmacia, Freiburg, Ger-many) before exposure to X-ray films (Hyperfilm; Amersham). Fordensitometric evaluation of the resulting bands films were scannedand analyzed using BIO-PROFIL Bio-1D image software (VilberLourmat, France). To calculate relative amounts of THP in urinethe densitometry intensities from immunoblots were normalized forthe individual urine volumes.
Tissue preparation for LM and EM immunohistochemistry
The animals were killed by in vivo perfusion fixation underpentobarbital anesthesia. The kidneys were perfused retrogradelythrough the abdominal aorta using PBS adjusted to 330 mosmol/kgH2O with sucrose, pH 7.4, for 20 s, followed by 3% paraformal-dehyde in PBS for 5 min, removed, and then cryoprotected in800 mosmol/kg H2O in PBS overnight and shock-frozen in liquidnitrogen-cooled isopentane. Alternatively, fine slices of the kidneyswere processed for embedding in LR White (London Resin,Reading, UK) for immunoelectron microscopy. Tissue was cryosub-stituted in a Leica AFS freeze-substitution unit, equilibrated over4 days in pure ethanol at temperatures gradually increasing from�90�C to �45�C, and infiltrated with LR White for 11 h at �15�Cand for 48 h at �10�C. After polymerization 70-nm sections werecut with an ultramicrotome (Reichert Ultracut S) and placed onnickel grids.
LM immunohistochemistry
Immunohistochemical staining was performed on cryostat sectionsthat were pretreated with 5% skim milk in PBS, pH 7.4. Afterincubating the sections with the primary antibody for 2 h at roomtemperature or at 4�C overnight, bound antibody was detected by a1-h incubation with swine anti-rabbit antibody followed by perox-idase-coupled anti-swine IgG (Dako). Signal was generated with0.1% diaminobenzidine and 0.02% H2O2 in PBS for standardizedduration. Sections were counterstained with hematoxylin andanalyzed using a Leica DMRB microscope.
320
Immunoelectron microscopy
Grids were preincubated in PBS containing 5% skim milk–PBS,pH 7.4, followed by incubation for 1 h with rabbit anti-THPantibody at room temperature and then overnight at 4�C. Primaryantibody was visualized using gold-labeled goat anti-rabbit IgG(AuroProbe, EM Gar-G5; Amersham). After further rinsing, gridswere fixed in 5% glutaraldehyde for 10 min, and then silver-impregnated (AuroProbe) before examination in a LEO 906 elec-tron microscope (Oberkochen, Germany).
In situ hybridization
To generate a THP riboprobe, a cDNA fragment was generated byRT-PCR using a sequence from position 1170 to 1662 of mouseTHP. The identity of the cDNA probe was verified by sequencing.The generated fragment was subcloned into the EcoRV site ofpBluescript KS+ vector (Stratagene, La Jolla, USA). To generatesense and antisense riboprobes the vector was linearized either byHindIII or by BamHI, respectively. RNA probes were synthesizedand labeled by in vitro transcription using digoxigenin (DIG)-labeled UTP and T3 or T7 RNA polymerases (Roche) to obtaineither sense (control) or antisense transcripts, respectively. In situhybridization on 6-�m-thick cryostat sections was essentially doneas described (Schmitt et al. 1999). Briefly, 6 ng DIG-labeled probesper microliter of hybridization mixture were applied. Hybridizationwas performed at 40�C for 18 h. After washing and subsequentblocking (2% normal sheep serum, 0.5% BSA, 3% Triton X-100 inbuffer I), sheep anti-DIG alkaline phosphatase-conjugated antibodywas administered to the sections. After removal of excess antibody,substrate (nitro blue tetrazolium salt containing 5-bromo-4-chloro-3-indolyl phosphate) was added and the reaction developed for 4–20 h at 4�C.
Presentation of data and statistical analysis
Quantitative data are presented as mean € standard error of mean(SEM). For statistical comparison the unpaired t-test (whenvariances were the same) or the Mann-Whitney rank sum test(when variances differed significantly between groups) was em-ployed. P values of less than 0.05 were considered statisticallysignificant.
Results
Clinical parameters and gross histology
A complete description of the clinical parameters of therats used in this study has been previously reported indetail; tissues and urine samples were from the sameanimals (Schmitt et al. 2003). In brief, the changescomprised markedly decreased serum levels of free T3, adrastic lack of weight gain, and a low kidney-to-bodyweight ratio as compared to the control rats. Urinaryoutput was roughly doubled in the hypothyroid rats, andurine osmolality was three times lower than in controls.The creatinine clearance was significantly reduced indi-cating a lowered GFR. These changes were largelyreversed by exogenous substitution of T3 in the treatedgroup. Histology revealed a marked reduction in corticalthickness and a more moderate decrease of the medullaryvolume. In agreement with a previous ultrastructuralreport (Bentley et al. 1985), the medullary portion of theTAL epithelium displayed a conspicuous, disproportion-
ate decrease in tubular perimeter and cell height, whereasqualitative changes at the subcellular level were notobserved.
Quantification of renal and urinary THP levels
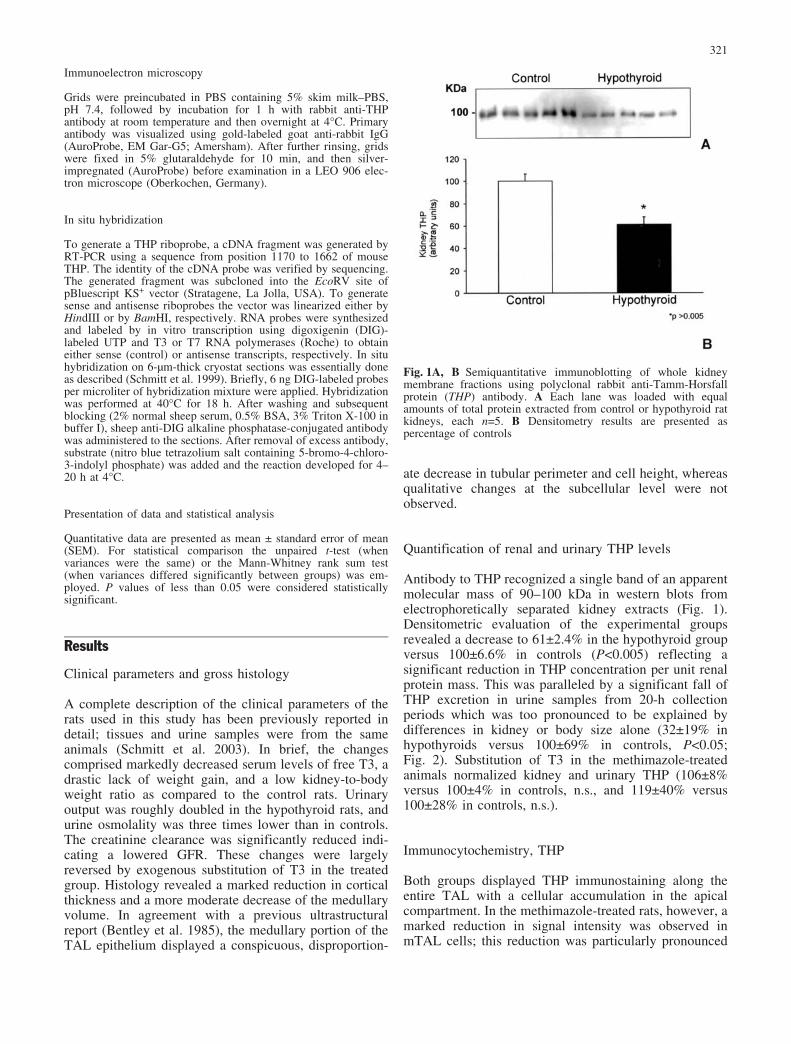
Antibody to THP recognized a single band of an apparentmolecular mass of 90–100 kDa in western blots fromelectrophoretically separated kidney extracts (Fig. 1).Densitometric evaluation of the experimental groupsrevealed a decrease to 61€2.4% in the hypothyroid groupversus 100€6.6% in controls (P<0.005) reflecting asignificant reduction in THP concentration per unit renalprotein mass. This was paralleled by a significant fall ofTHP excretion in urine samples from 20-h collectionperiods which was too pronounced to be explained bydifferences in kidney or body size alone (32€19% inhypothyroids versus 100€69% in controls, P<0.05;Fig. 2). Substitution of T3 in the methimazole-treatedanimals normalized kidney and urinary THP (106€8%versus 100€4% in controls, n.s., and 119€40% versus100€28% in controls, n.s.).
Immunocytochemistry, THP
Both groups displayed THP immunostaining along theentire TAL with a cellular accumulation in the apicalcompartment. In the methimazole-treated rats, however, amarked reduction in signal intensity was observed inmTAL cells; this reduction was particularly pronounced
Fig. 1A, B Semiquantitative immunoblotting of whole kidneymembrane fractions using polyclonal rabbit anti-Tamm-Horsfallprotein (THP) antibody. A Each lane was loaded with equalamounts of total protein extracted from control or hypothyroid ratkidneys, each n=5. B Densitometry results are presented aspercentage of controls
321
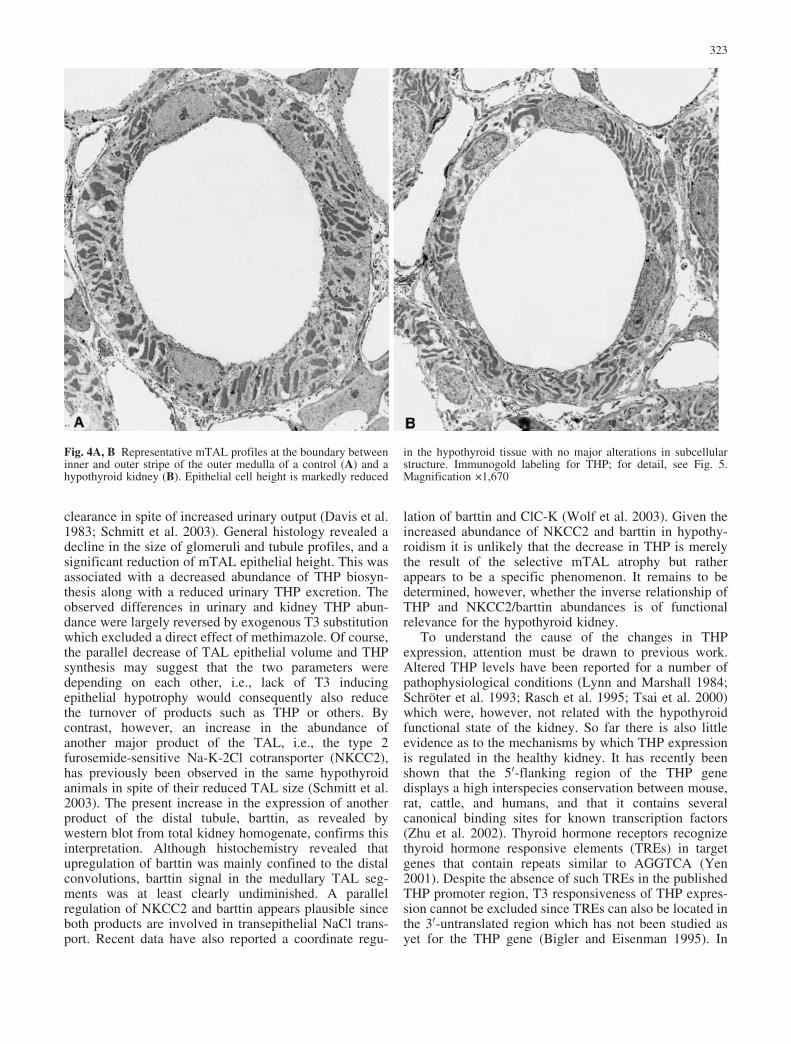
in the inner stripe (Fig. 3C, D). No differences in theintensity of THP immunostaining were observed in thecortical TAL between groups (Fig. 3A, B). At theultrastructural level immunogold labeling of THP wasconsistently observed over the TAL epithelium, where itwas concentrated in the apical vesicle compartment(Figs. 4, 5). Basolateral membranes showed little stainingin both groups. Labeling in the mTAL of hypothyroidkidneys was clearly diminished (Figs. 4, 5). This was notthe result of a reduced total number of apical vesicles, butrather of a smaller fraction of THP-positive vesicles(Fig. 5A–D).
In situ hybridization, THP
In agreement with the immunohistochemical results, insitu hybridization revealed a robust abundance of THPmRNA signal in the cortical TAL of both groups and amarked reduction in signal intensity in mTALs ofhypothyroid kidneys (Fig. 6).
Quantification of renal barttin levels
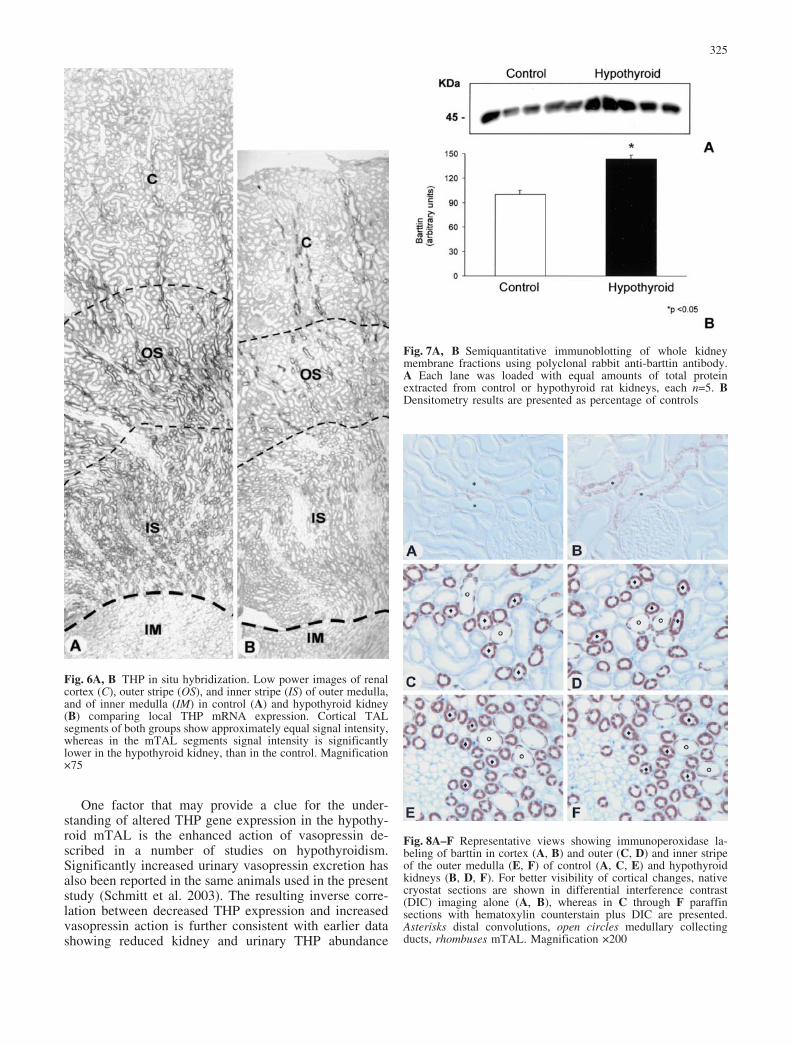
Antibody to barttin recognized a single band of anapparent molecular mass of 40–45 kDa in western blotsfrom electrophoretically separated kidney extracts(Fig. 7). Densitometric evaluation of the experimentalgroups revealed an increase to 143€11.4% in the hypo-thyroid group versus 100€11.3% in controls (P<0.05)reflecting a significant increase in barttin concentrationper unit renal protein mass.
Immunocytochemistry, barttin
Both groups displayed significant barttin immunostainingof the basolateral membrane in several tubular portions.In the cortex of controls, signal was located in the TAL,distal convoluted tubule (DCT), and connecting tubule(CNT) cells. Intercalated cells of CNT and corticalcollecting duct (CCD) were also positive. In the outermedulla, strong staining was prevalent in the TAL and,less strikingly, in collecting duct intercalated cells(Fig. 8A, C, E). In the methimazole-treated rats, a markedincrease in signal intensity was observed in the distalconvolutions (Fig. 8B), whereas the TAL segments incortex and outer medulla revealed no obvious changes instaining (Fig. 8D, F). Medullary collecting duct principalcells were not obviously altered either.
Discussion
In agreement with previous reports, hypothyroid ratsshowed a lack of growth that affected kidneys dispropor-tionately and that was associated with reduced creatinine
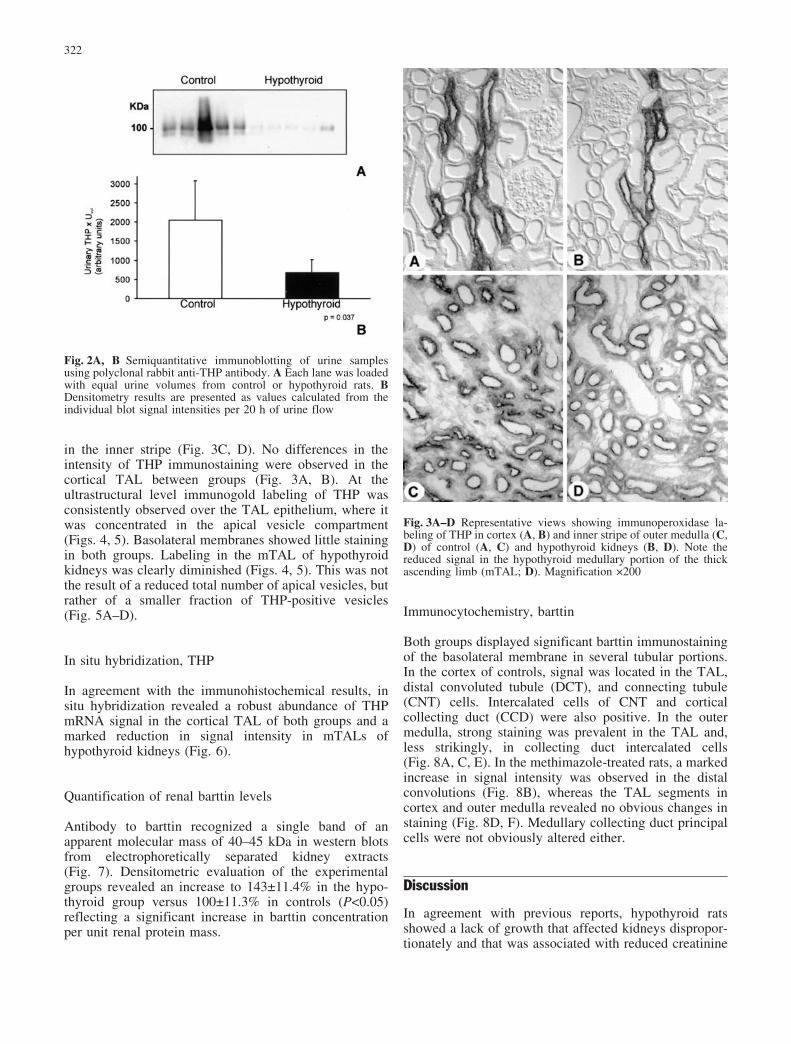
Fig. 2A, B Semiquantitative immunoblotting of urine samplesusing polyclonal rabbit anti-THP antibody. A Each lane was loadedwith equal urine volumes from control or hypothyroid rats. BDensitometry results are presented as values calculated from theindividual blot signal intensities per 20 h of urine flow
Fig. 3A–D Representative views showing immunoperoxidase la-beling of THP in cortex (A, B) and inner stripe of outer medulla (C,D) of control (A, C) and hypothyroid kidneys (B, D). Note thereduced signal in the hypothyroid medullary portion of the thickascending limb (mTAL; D). Magnification 200
322
clearance in spite of increased urinary output (Davis et al.1983; Schmitt et al. 2003). General histology revealed adecline in the size of glomeruli and tubule profiles, and asignificant reduction of mTAL epithelial height. This wasassociated with a decreased abundance of THP biosyn-thesis along with a reduced urinary THP excretion. Theobserved differences in urinary and kidney THP abun-dance were largely reversed by exogenous T3 substitutionwhich excluded a direct effect of methimazole. Of course,the parallel decrease of TAL epithelial volume and THPsynthesis may suggest that the two parameters weredepending on each other, i.e., lack of T3 inducingepithelial hypotrophy would consequently also reducethe turnover of products such as THP or others. Bycontrast, however, an increase in the abundance ofanother major product of the TAL, i.e., the type 2furosemide-sensitive Na-K-2Cl cotransporter (NKCC2),has previously been observed in the same hypothyroidanimals in spite of their reduced TAL size (Schmitt et al.2003). The present increase in the expression of anotherproduct of the distal tubule, barttin, as revealed bywestern blot from total kidney homogenate, confirms thisinterpretation. Although histochemistry revealed thatupregulation of barttin was mainly confined to the distalconvolutions, barttin signal in the medullary TAL seg-ments was at least clearly undiminished. A parallelregulation of NKCC2 and barttin appears plausible sinceboth products are involved in transepithelial NaCl trans-port. Recent data have also reported a coordinate regu-
lation of barttin and ClC-K (Wolf et al. 2003). Given theincreased abundance of NKCC2 and barttin in hypothy-roidism it is unlikely that the decrease in THP is merelythe result of the selective mTAL atrophy but ratherappears to be a specific phenomenon. It remains to bedetermined, however, whether the inverse relationship ofTHP and NKCC2/barttin abundances is of functionalrelevance for the hypothyroid kidney.
To understand the cause of the changes in THPexpression, attention must be drawn to previous work.Altered THP levels have been reported for a number ofpathophysiological conditions (Lynn and Marshall 1984;Schr�ter et al. 1993; Rasch et al. 1995; Tsai et al. 2000)which were, however, not related with the hypothyroidfunctional state of the kidney. So far there is also littleevidence as to the mechanisms by which THP expressionis regulated in the healthy kidney. It has recently beenshown that the 50-flanking region of the THP genedisplays a high interspecies conservation between mouse,rat, cattle, and humans, and that it contains severalcanonical binding sites for known transcription factors(Zhu et al. 2002). Thyroid hormone receptors recognizethyroid hormone responsive elements (TREs) in targetgenes that contain repeats similar to AGGTCA (Yen2001). Despite the absence of such TREs in the publishedTHP promoter region, T3 responsiveness of THP expres-sion cannot be excluded since TREs can also be located inthe 30-untranslated region which has not been studied asyet for the THP gene (Bigler and Eisenman 1995). In
Fig. 4A, B Representative mTAL profiles at the boundary betweeninner and outer stripe of the outer medulla of a control (A) and ahypothyroid kidney (B). Epithelial cell height is markedly reduced
in the hypothyroid tissue with no major alterations in subcellularstructure. Immunogold labeling for THP; for detail, see Fig. 5.Magnification 1,670
323

general, the TAL has not been explicitly established as atarget for T3. Published data regarding the segmentalexpression of T3 receptors along the nephron epitheliumare inconclusive with respect to the TAL (Luo et al. 1989;
Tagami et al. 1990). Indirect information derived fromstudies investigating T3 action on Na,K-ATPase rathersuggest a lack of responsiveness for T3 within thissegment (Barlet and Doucet 1986).
Fig. 5A–F High resolution immunoelectron microscopy showingmTAL in control (A, C, E) and hypothyroid kidneys (B, D, F).Labeling in the apical vesicle compartment in the hypothyroid
tissue is markedly reduced as compared to the control in supranu-clear (A–D) and apical cytoplasmic portions (E, F) of the cell.Magnifications 7,600 (A, B); 32,800 (C, D); 21,500 (E, F)
324
One factor that may provide a clue for the under-standing of altered THP gene expression in the hypothy-roid mTAL is the enhanced action of vasopressin de-scribed in a number of studies on hypothyroidism.Significantly increased urinary vasopressin excretion hasalso been reported in the same animals used in the presentstudy (Schmitt et al. 2003). The resulting inverse corre-lation between decreased THP expression and increasedvasopressin action is further consistent with earlier datashowing reduced kidney and urinary THP abundance
Fig. 6A, B THP in situ hybridization. Low power images of renalcortex (C), outer stripe (OS), and inner stripe (IS) of outer medulla,and of inner medulla (IM) in control (A) and hypothyroid kidney(B) comparing local THP mRNA expression. Cortical TALsegments of both groups show approximately equal signal intensity,whereas in the mTAL segments signal intensity is significantlylower in the hypothyroid kidney, than in the control. Magnification75
Fig. 7A, B Semiquantitative immunoblotting of whole kidneymembrane fractions using polyclonal rabbit anti-barttin antibody.A Each lane was loaded with equal amounts of total proteinextracted from control or hypothyroid rat kidneys, each n=5. BDensitometry results are presented as percentage of controls
Fig. 8A–F Representative views showing immunoperoxidase la-beling of barttin in cortex (A, B) and outer (C, D) and inner stripeof the outer medulla (E, F) of control (A, C, E) and hypothyroidkidneys (B, D, F). For better visibility of cortical changes, nativecryostat sections are shown in differential interference contrast(DIC) imaging alone (A, B), whereas in C through F paraffinsections with hematoxylin counterstain plus DIC are presented.Asterisks distal convolutions, open circles medullary collectingducts, rhombuses mTAL. Magnification 200
325

upon vasopressin administration (Bachmann et al. 1991;Kim et al. 1999).
It has been well established that in hypothyroidismthere is decreased capacity of the kidney to achievemaximal urinary concentration and dilution. Severalfactors have been suggested to account for this phenom-enon, i.e., reduced delivery of glomerular filtrate to thedistal tubule, impaired ion reabsorption by the TAL, andenhanced ADH activity in the collecting duct (Holmes etal. 1970; Michael et al. 1972, 1976; Emmanouel et al.1974; Katz et al. 1975). Since it has been postulated thatTHP contributes to salt reabsorptive properties of theTAL (Hoyer et al. 1979; Mattey and Naftalin 1992;Rampoldi et al. 2003), decreased THP abundance in thehypothyroid state might be relevant in this context.Interestingly, THP deficiency in mice appears to reducethe kidney’s concentrating capacity (unpublished resultsfrom our group). This finding is in agreement with thedescribed phenotype in patients with medullary cystickidney disease (Rampoldi et al 2003). However, sincepotential interactions between THP and ion transportproteins have not been investigated at a molecular level, arole of decreased THP abundance in hypothyroid renaldysfunction remains speculative. Further studies areneeded to elucidate potential relations between THP andtransepithelial transport in physiological and pathologicalstates.
In summary, we show that THP expression is specif-ically decreased in the mTAL of rats rendered hypothy-roid by methimazole treatment, and that THP abundancewas normalized by exogenous T3 substitution. We sug-gest that decreased THP levels are indirectly related to theenhanced action of vasopressin. Reduced THP is inverse-ly correlated with increased NKCC2 and barttin abun-dance. These changes may be relevant in the impairedfunction of the hypothyroid TAL resulting in defectiveion transport and urinary dilution and concentration.
Acknowledgements We wish to express our gratitude to T. Jentsch,ZMNH Hamburg, for providing us with anti-barttin antibody, Dr.John R. Hoyer, Children’s Hospital, Philadelphia, for the THPantibody, Ursula M�ller for technical assistance, and FranziskaTheilig for producing Fig. 8.
References
Bachmann S, Koeppen-Hagemann I, Kriz W (1985) Ultrastructurallocalization of Tamm-Horsfall glycoprotein (THP) in rat kidneyas revealed by protein A-gold immunocytochemistry. Histo-chemistry 83:531–538
Bachmann S, Metzger R, Bunnemann B (1990) Tamm-Horsfallprotein-mRNA synthesis is localized to the thick ascendinglimb of Henle’s loop in rat kidney. Histochemistry 94:517–523
Bachmann S, Dawnay AB, Bouby N, Bankir L (1991) Tamm-Horsfall protein excretion during chronic alterations in urinaryconcentration and protein intake in the rat. Ren PhysiolBiochem 14:236–245
Barlet C, Doucet A (1986) Kinetics of triiodothyronine action onNa-K-ATPase in single segments of rabbit nephron. Pfl�gersArch 407:27–32
Bentley AG, Madsen KM, Davis RG, Tisher CC (1985) Responseof the medullary thick ascending limb to hypothyroidism in therat. Am J Pathol 120:215–222
Bigler J, Eisenman RN (1995) Novel location and function of athyroid hormone response element. EMBO J 14:5710–5723
Birkenhager R, Otto E, Schurmann MJ, Vollmer M, Ruf E-M,Maier-Lutz I, Beekmann F, Fekete A, Omran H, Feldmann D,Milford DV, Jeck N, Konrad M, Landau D, Knoers NVM,Antignac C, Sudbrak R, Kispert A, Hildebrandt F (2001)Mutation of BSND causes Bartter syndrome with sensorineuraldeafness and kidney failure. Nat Genet 29:310–314
Bradley SE, Stephan F, Coelho JB, Reville P (1974) The thyroidand the kidney. Kidney Int 6:346–365
Bradley SE, Coelho JB, Sealey JE, Edwards KD, Stephan F (1982)Changes in glomerulotubular dimensions, single nephronglomerular filtration rates and the renin-angiotensin system inhypothyroid rats. Life Sci 30:633–639
Davis RG, Madsen KM, Fregly MJ, Tisher CC (1983) Kidneystructure in hypothyroidism. Am J Pathol 113:41–49
Emmanouel DS, Lindheimer MD, Katz AI (1974) Mechanism ofimpaired water excretion in the hypothyroid rat. J Clin Invest54:926–934
Estevez R, Boettger T, Stein V, Birkenhager R, Otto E, HildebrandtF, Jentsch TJ (2001) Barttin is a Cl� channel beta-subunitcrucial for renal Cl� reabsorption and inner ear K+ secretion.Nature 414:558–561
Hart TC, Gorry MC, Hart PS, Woodard AS, Shihabi Z, Sandhu J,Shirts B, Hy L, Zhu H, Barmada MM, Bleyer AJ (2002)Mutations in UMOD gene are responsible for medullary cystickidney disease 2 and familial juvenile hyperuricaemic neph-ropathy. J Med Genet 39:882–892
Holmes EW Jr, DiScala VA (1970) Studies on the exaggeratednatriuretic response to a saline infusion in the hypothyroid rat. JClin Invest 49:1224–1236
Hoyer JR, Sisson SP, Vernier RL (1979) Tamm-Horsfall glyco-protein: ultrastructural immunoperoxidase localization in ratkidney. Lab Invest 41:168–173
Katz AI, Emmanouel DS, Lindheimer MD (1975) Thyroid hormoneand the kidney. Nephron 15:223–224
Kim GH, Ecelbarger CA, Mitchell C, Packer RK, Wade JB,Knepper MA (1999) Vasopressin increases Na-K-2Cl cotrans-porter expression in thick ascending limb of Henle’s loop. Am JPhysiol 276:F96–F103
Kokot F, Dulawa J (2000) Tamm-Horsfall protein updated.Nephron 85:97–102
Luo M, Faure R, Tong YA, Dussault JH (1989) Immunocyto-chemical localization of the nuclear 3,5,30-triiodothyroninereceptor in the adult rat: liver, kidney, heart, lung and spleen.Acta Endocrinol (Copenh) 120:451–458
Lynn KL, Marshall RD (1984) Excretion of Tamm-Horsfallglycoprotein in renal disease. Clin Nephrol 22:253–257
Mattey M, Naftalin L (1992) Mechanoelectrical transduction, ionmovement and water stasis in uromodulin. Experientia 48:975–980
Michael UF, Barenberg RL, Chavez R, Vaamonde CA, Papper S(1972) Renal handling of sodium and water in the hypothyroidrat. Clearance and micropuncture studies. J Clin Invest51:1405–1412
Michael UF, Kelley J, Alpert H, Vaamonde CA (1976) Role ofdistal delivery of filtrate in impaired renal dilution of thehypothyroid rat. Am J Physiol 230:699–705
Rampoldi L, Caridi G, Santon D, Boaretto F, Bernascone I,Lamorte G, Tardanico R, Dagnino M, Colussi G, Scolari F,Ghiggeri GM, Amoroso A, Casari G (2003) Allelism ofMCKD, FJHN and GCKD caused by impairment of uromod-ulin export dynamics. Hum Mol Genet 15:3369–3384
Rasch R, Torffvit O, Bachmann S, Jensen PK, Jacobsen NO (1995)Tamm-Horsfall glycoprotein in streptozotocin diabetic rats: astudy of kidney in situ hybridization, immunohistochemistry,and urinary excretion. Diabetologia 38:525–535
Schmitt R, Ellison DH, Farman N, Rossier BC, Reilly RF, ReevesWB, Oberb�umer I, Tapp R, Bachmann S (1999) Develop-
326

mental expression of sodium entry pathways in rat nephron.Am J Physiol 276:F367–F381
Schmitt R, Klussmann E, Kahl T, Ellison DH, Bachmann S (2003)Renal expression of sodium transporters and aquaporin-2 inhypothyroid rats. Am J Physiol 284:F1097–F1104
Schr�ter J, Timmermans G, Seyberth HW, Greven J, Bachmann S(1993) Marked reduction of Tamm-Horsfall protein synthesis inhyperprostaglandin E-syndrome. Kidney Int 44:401–410
Tagami T, Nakamura H, Sasaki S, Mori T, Yoshioka H, Yoshida H,Imura H (1990) Immunohistochemical localization of nuclear3,5,30-triiodothyronine receptor proteins in rat tissues studiedwith antiserum against C-ERB A/T3 receptor. Endocrinology127:1727–1734
Tsai CY, Wu TH, Yu CL, Lu JY, Tsai YY (2000) Increasedexcretions of beta2-microglobulin, IL-6, and IL-8 and de-
creased excretion of Tamm-Horsfall glycoprotein in urine ofpatients with active lupus nephritis. Nephron 85:207–214
Wolf K, Meier-Meitinger M, Bergler T, Castrop H, Vitzthum H,Riegger GA, Kurtz A, Kramer BKJ (2003) Parallel down-regulation of chloride channel CLC-K1 and barttin mRNA inthe thin ascending limb of the rat nephron by furosemide.Pflugers Arch 446:665–671
Yen PM (2001) Physiological and molecular basis of thyroidhormone action. Physiol Rev 81:1097–1142
Zhu X, Cheng J, Gao J, Lepor H, Zhang ZT, Pak J, Wu XR (2002)Isolation of mouse THP gene promoter and demonstration of itskidney-specific activity in transgenic mice. Am J Physiol282:F608–F617
327


















