
Role in cation translocation of the N-terminus of the ac-subunit of the ...
16
Journal of Physiology (1996), 491.3, pp.579-594 Role in cation translocation of the N-terminus of the ac-subunit of the Na`-K+ pump of Bufo Xinyu Wang, Frederic Jaisser and Jean-Daniel Horisberger * Institute of Pharmacology and Toxicology, University of Lausanne, Bugnon 27, CH-1005 Lausanne, Switzerland 1. We have studied the effects on the physiological properties of the Na+-K+ pump of both 31- and 40-amino acid N-terminal truncated forms of the a-subunit of the Na+-K+-ATPase. 2. Na+-K+ pumps that were moderately ouabain resistant (Ki = 50 /M) were expressed in the Xenopus oocyte by injection of wild-type or truncated variants of the Bufo marinus Na+-K+-ATPase a-subunit cRNA with Bufo ,8-subunit cRNA. The function of the Na+-K+ pump was studied by electrophysiological methods after Na+ loading and inhibition of the endogenous Xenopu,s Na+-K+-ATPase by exposure to a low concentration (0-2 uM) of ouabain. 3. The voltage-dependent potassium activation kinetics of the Na+-K+ pump current and the ouabain-sensitive proton-dependent inward current were studied using the two-electrode voltage-clamp technique. A novel technique involving permeabilization of part of the oocyte membrane with digitonin was developed to enable study of the pre-steady-state current following fast voltage perturbation. 4. By comparison with the wild type, the 40-amino acid N-terminal truncation induced a lower level of Na+-K+ pump current, a 2- to 3-fold reduction in the apparent external K+ affinity when measured in the presence of extracellular Na+, a relative increase in the proton-dependent inward current, and a reduction in the rate constant of the pre-steady- state current following a voltage step towards a positive membrane potential. The 31-amino acid truncation induced changes that were qualitatively similar but of smaller magnitude. 5. We have analysed these results using a kinetic model of the Na+-K+ pump cycle and have shown that all these effects can be explained by the change in a single rate constant in the cycle kinetics, namely a reduction in the rate of the main charge translocating part of the Na+-K+ pump cycle, i.e. the forward El to E2 conformational change, the deocclusion and release of Na+ to the external side. 6. The highly charged N-terminal segment seems to be directly involved in the mechanism that translocates Na ions across the membrane's electrical field. Na+-K+-ATPase is the enzyme responsible for the maintenance of Nae and K+ gradients across the membrane of all animal cells. Because of its central role in the physiology of the cell and its possible involvement in several important clinical conditions, a great deal of effort has been expended in attempts to clarify the molecular physiology of this cation transport system. However, the molecular mechanism responsible for cation translocation across the membrane is still not fully understood. Na+-K+-ATPase is composed of an a- and a fl-subunit. The primary structure of both subunits is known for many species. Currently the most widely accepted model of the membrane topology of the a-subunit of the Na+-K+- ATPase places the N-terminus inside the cell (Horisberger, 1994; Yoon & Guidotti, 1994). The N-terminal part is one of the less-conserved regions of the a-subunit. However, some of its characteristics are common to many species and isoforms. In most cases, it contains a first cluster of five to nine lysine residues and in all the known al-subunits of mammalian, avian and amphibian species (Horisberger, 1994), there is a highly conserved ten-amino-acid segment enclosed between two methionines that contains two * To whom correspondence should be addressed. 4645 579
Transcript of Role in cation translocation of the N-terminus of the ac-subunit of the ...

Journal of Physiology (1996), 491.3, pp.579-594
Role in cation translocation of the N-terminus of theac-subunit of the Na`-K+ pump of Bufo
Xinyu Wang, Frederic Jaisser and Jean-Daniel Horisberger *
Institute of Pharmacology and Toxicology, University of Lausanne,Bugnon 27, CH-1005 Lausanne, Switzerland
1. We have studied the effects on the physiological properties of the Na+-K+ pump of both 31-and 40-amino acid N-terminal truncated forms of the a-subunit of the Na+-K+-ATPase.
2. Na+-K+ pumps that were moderately ouabain resistant (Ki = 50/M) were expressed in theXenopus oocyte by injection of wild-type or truncated variants of the Bufo marinusNa+-K+-ATPase a-subunit cRNA with Bufo ,8-subunit cRNA. The function of the Na+-K+pump was studied by electrophysiological methods after Na+ loading and inhibition of theendogenous Xenopu,s Na+-K+-ATPase by exposure to a low concentration (0-2 uM) ofouabain.
3. The voltage-dependent potassium activation kinetics of the Na+-K+ pump current and theouabain-sensitive proton-dependent inward current were studied using the two-electrodevoltage-clamp technique. A novel technique involving permeabilization of part of theoocyte membrane with digitonin was developed to enable study of the pre-steady-statecurrent following fast voltage perturbation.
4. By comparison with the wild type, the 40-amino acid N-terminal truncation induced alower level of Na+-K+ pump current, a 2- to 3-fold reduction in the apparent external K+affinity when measured in the presence of extracellular Na+, a relative increase in theproton-dependent inward current, and a reduction in the rate constant of the pre-steady-state current following a voltage step towards a positive membrane potential. The31-amino acid truncation induced changes that were qualitatively similar but of smallermagnitude.
5. We have analysed these results using a kinetic model of the Na+-K+ pump cycle and haveshown that all these effects can be explained by the change in a single rate constant in thecycle kinetics, namely a reduction in the rate of the main charge translocating part of theNa+-K+ pump cycle, i.e. the forward El to E2 conformational change, the deocclusion andrelease of Na+ to the external side.
6. The highly charged N-terminal segment seems to be directly involved in the mechanismthat translocates Na ions across the membrane's electrical field.
Na+-K+-ATPase is the enzyme responsible for themaintenance of Nae and K+ gradients across the membraneof all animal cells. Because of its central role in thephysiology of the cell and its possible involvement inseveral important clinical conditions, a great deal of efforthas been expended in attempts to clarify the molecularphysiology of this cation transport system. However, themolecular mechanism responsible for cation translocationacross the membrane is still not fully understood.
Na+-K+-ATPase is composed of an a- and a fl-subunit. Theprimary structure of both subunits is known for many
species. Currently the most widely accepted model of themembrane topology of the a-subunit of the Na+-K+-ATPase places the N-terminus inside the cell (Horisberger,1994; Yoon & Guidotti, 1994). The N-terminal part is oneof the less-conserved regions of the a-subunit. However,some of its characteristics are common to many species andisoforms. In most cases, it contains a first cluster of five tonine lysine residues and in all the known al-subunits ofmammalian, avian and amphibian species (Horisberger,1994), there is a highly conserved ten-amino-acid segmentenclosed between two methionines that contains two
* To whom correspondence should be addressed.
4645 579

X. Wang, F Jaisser and J-D. Horisberger
positively charged lysines and three negatively chargedamino acids:
M (D/E) E L K K E (I/V) (S/T) M.
The two central lysines are highly conserved throughoutthe whole Na+-K+- and H+-K+-ATPase family. Thefunction of N-truncated mutants has been studied with theaid of either the proteolytic property of trypsin, whichrapidly cleaves the N-terminus of the a-subunit (Jorgensen,1975, 1977; Jorgensen & Collins, 1986) (see cleavage site inFig. 1) or of genetically engineered mutants. In most cases,the truncated form was found to be able to sustain ATPaseor ion transport functions (Ohta et al. 1991; Vasilets, Omay,Ohta, Noguchi & Kawamura, 1991; Wierzbicki & Blostein,1993; Vasilets, Ohta, Noguchi, Kawamura & Schwarz, 1993;Shanbaky & Pressley, 1994; Daly, Lane & Blostein, 1994).Surprisingly, truncation of the intracellular N-terminus ofthe a-subunit induced a significant modification of theactivation of the Na+-K+ pump by external K+ (Burgener-Kairuz, Horisberger, Geering & Rossier, 1991; Vasilets et al.1991) and of its inhibition by external Na+ (Vasilets et al.1993). We have previously studied the two forms truncatedat one or other of the two methionines in the Xenopusal -isoform and reported a decrease in the apparent affinityfor external K+ in the larger truncation (Burgener-Kairuzet al. 1991). Other investigators have observed an oppositeeffect with a shorter truncation in the Torpedo Na+-K+pump (Vasilets et al. 1991) and under different experimentalconditions (Nae-free external medium). The half-activationconstant (K;, the K+ concentration that produces a half-maximal turnover rate) is a measure of apparent affinity,a variable that depends not only on the intrinsic affinity(Kd) of the E2 conformation for the cation, but also on thekinetics of the steps preceding and following cationbinding (Lauger, 1991; Jaisser, Jaunin, Geering, Rossier& Horisberger, 1994). Thus, the effect of N-terminaltruncation on the apparent affinity for external K+ could,in principle, be explained by one of two mechanisms. In thefirst, the N-terminal part, although intracellular, extendsfar enough into the membrane to either participate in thestructure of the external K+ binding site or to influence itby an allosteric mechanism (modification of intrinsicaffinity). In the second, N-terminal truncations alter the
kinetics of some other steps in the pump cycle resulting in amodification of the apparent affinity for potassium.
We have examined these hypotheses by studying severalfunctional aspects of cation translocation in wild-type andN-truncated mutants of the a-subunit. For this purpose, wegenerated two N-terminal truncated forms of the a-subunitof Bufo marinus Na+-K+-ATPase. Bufo Na+-K+-ATPaseis moderately resistant to ouabain and is highly expressedwhen a- and f-subunit cRNA is injected into Xenopusoocytes (Jaisser, Canessa, Horisberger & Rossier, 1992).This allows inhibition of the endogenous, highly ouabain-sensitive Xenopus Na+-K+ pump and specific study of theproperties of the exogenous, artificially expressed Na+-K+pump variant (Jaisser et al. 1994). We studied potassiumactivation kinetics in the presence and absence of externalNa+ in the wild type and the two truncated variants.
The Na+-K+ pump moves 3 Na+ out and 2 K+ into the celleach time an ATP molecule is hydrolysed; it thus producesan outward current of one net charge per cycle. There isgood experimental evidence to suggest that the main chargetranslocation occurs during the transport and release to theoutside of Na+ ions (Stiirmer, Biihler, Apell & Lauger, 1991;Gadsby, Rakowski & De Weer, 1993). This phenomenonhas allowed study of the kinetics of the Nae-transportinglimb of the pump cycle by measurement of the pre-steady-state current following fast voltage perturbation inmyocardial cells (Nakao & Gadsby, 1986; Hilgemann, 1994)and in Xenopus oocytes (Rakowski, 1993; Holmgren &Rakowski, 1994; Sagar & Rakowski, 1994). We havedeveloped a new technique that enables the study of thecurrent transients following fast voltage perturbation andwhich allows the study of the Na+-K+ pump-mediated(i.e. ouabain-sensitive) pre-steady-state current in oocytesexpressing the wild-type or mutant Na+-K+ pumps. Thepresent data indicate that N-terminal truncation induces areduction in the forward rate constant of the Na+-translocating part of the cycle. Qualitatively similar resultsthat we obtained in preliminary experiments using the two-electrode voltage-clamp technique and the T40 N-truncatedmutant have already been reported (Horisberger, Jaisser,Canessa & Rossier, 1994).
1 2 Try 2
MGYGAGRDKY EPAATSEHGG KKGKGKGKDR DMEELKKEVT MEDHKMTLEEMEELKKEVT MEDHKMTLEE
MEDHKMTLEE
WTT31T40
Figure 1. N-terminal truncation of the a-subunit of the Bufo Na+-K+ pumpArrows point to sites of N-terminal truncation which have been studied by other investigators. 1 and 2indicate the site of the 2 truncations of the Torpedo a-subunit described by Ohta et al. (1991) and Vasiletset al. (1991, 1993). The position of the trypsin cleavage site (Try2) (Jorgensen & Collins, 1986) is alsoindicated. A truncation at the corresponding Try2 position in the rat al -subunit has been studied byDaly et al. ( 1994). WT, wild type.
580 J Physiol.491.3

Role of N-terminus of Na+-K+ pump a-subunit
Efthymiadis, Rettinger & Schwarz (1993) observed aninward current mediated oy the Na+-K+ pump when theextracellular medium contained no Na+ or K+. We recentlyshowed that this inward current was due to the transportof protons and proposed that it was produced by a protonleak through the Na+-K+ pump in the E2 conformation(Wang & Horisberger, 1995). In the present study, we alsolooked at the effects of N-terminal truncation on thisNa+-K+ pump-mediated inward proton current.
METHODSGeneration of N-terminal deletion mutantsDeletion mutants of the N-terminus of the Bufo marinusal-subunit were prepared using the polymerase chain reaction(PCR), as previously described (Burgener-Kairuz et al. 1991).Briefly, mutated DNA fragments were amplified by PCR betweena 5' sense deletion oligonucleotide (T31 or T40) and a 3' antisenseal-oligonucleotide (B), located downstream of a unique XhoJrestriction site, using the Bufo marinus al-subunit cDNA astemplate (Jaisser et al. 1992).
The oligonucleotides were as follows:
T31 GGAGAATTCGTTATACCGACAGC ATG GAA GAG CTC AAG
-24 -1/+150 +164
T40 GGAGAATTCGTTATACCGACAGC ATG GAA GAC CAT
-24 -1/+177 +188
B CCTTCACTTCCACAAGGTCT
+640 +620
The PCR-mutated fragments were digested with EcoRI (whoserestriction site is encoded at the 5' end of the deletionoligonucleotides) and Xhol (whose restriction site is locatedupstream of the 3' antisense B oligonucleotide). The DNA fragmentwas then subcloned in the psD Bufo al-plasmid (Jaisser et al.1992). Successful introduction of the mutation was confirmed bydideoxysequencing using the Sanger method (Sequenase, USB)
Expression of Bufo Na+-K+ pump in Xenopus oocytesOvarian segments were removed from female Xenopus laeviswhich had been anaesthetized by immersion in MS 222 (2 g F';Sandoz, Basel, Switzerland) and stage V/VI oocytes weredefolliculated as previously described (Geering, Theulaz, Verrey,Hiiuptle & Rossier, 1989). Wild-type and mutant Bufo a-subunitcRNA (7 ng) was injected together with Bufo /61 -subunit cRNA(1 ng), in a total volume of 50 nl, into oocytes as describedpreviously (Horisberger, Jaunin, Good, Rossier & Geering, 1991).Electrophysiological measurements were performed 3-5 days later.All the oocytes were loaded with sodium by a 2 h incubation in aK+-free and Ca2+-free medium containing 90 mm Nae and 0 5 mmEGTA. They were then kept in a K+-free amphibian Ringersolution (with 0 4 mM Ca2+) containing 0-2 /M ouabain for at least2 h (Horisberger et al. 1991). This treatment leads to inhibition ofthe endogenous Xenopus Na+-K+ pump and allows selective studyof the activity of the exogenously expressed Bufo Na+-K+ pumpwhich is moderately ouabain resistant.
Electrophysiological measurements were performed using a DaganTEV oocyte-clamp apparatus (Dagan Corporation, Minneapolis,MN, USA). Data acquisition and analysis were performed using a
TL-1 DMA interface and the pCLAMP software package (AxonInstruments). All the experiments were performed at roomtemperature (22-25 °C).
Steady-state currentSteady-state current was measured using the two-electrodevoltage-clamp method (Jaisser et al. 1994). The current signal waslow-pass filtered at 25 Hz. Current-voltage (I- V) curves wereobtained by recording the current while rectangular voltage pulses(125 ms) of varying amplitude (between the limits +80 and-80 mV) were applied every 1.5 s from a holding membranepotential of -50 mV. The steady-state current was measured100 ms after the start of the voltage step.
The steady-state current activated by 10 mm K+ at -50 mV wasmeasured first in each experiment. We have shown earlier thatthis current yields a reliable estimate of the number of Na+-K+pumps active at the oocyte membrane (Jaunin, Horisberger,Richter, Good, Rossier & Geering, 1992). Since the level ofexpression of the exogenous Na+-K+ pump is variable after cRNA,current values were normalized in each oocyte with respect to thisK+-induced current at -50 mV.
The half-activation constant for external K+ was measured overthe -130 to +30 mV potential range in the absence and in thepresence of 80 mm external Nae, as previously described (Jaisser etal. 1994). Steady-state current I-V curves were derived in thepresence of different concentrations of K+ (0, 20 #m, 100 sm,500 /M and 5 mm for the Nae-free condition and 0, 20 /zm,100 /am, 500 /uM, 2 mm and 10 mm in the presence of Na+). Thepotassium-activated current was obtained by subtracting thecurrent measured in K+-free solution from that measured in thepresence of K+. Activation kinetic parameters, maximal current(Imax) and the half-activation constant (K) were obtained byparameter fitting to the Hill equation using a Hill coefficient of 1'6for the experiments performed in the presence of external Na+ anda Hill coefficient of 1-0 for the experiments performed in theabsence of external Nae, as previously described (Jaisser et al.1994).
The pH-dependent, ouabain-sensitive inward current was measuredas described earlier (Wang & Horisberger, 1995). Briefly, I-Vcurves were obtained before and after addition of 2 mm ouabain ina nominally Nae- and K+-free solution at pH 6-0. This highconcentration of ouabain is necessary to inhibit the relativelyouabain-resistant Bufo Na+-K+ pump (Jaisser et al. 1992). Theouabain-sensitive inward current was obtained by subtraction ofthe current measured after 2 mm ouabain from the current beforeouabain.
Pre-steady-state currentIn order to overcome the limitations imposed by the low timeresolution of the two-electrode voltage-clamp method, a newtechnique was developed to enable fast voltage clamp on the wholeoocyte. This technique, which we name the 'half-permeabilizedoocyte (HPO) technique', was inspired by the 'perfused oocytetechnique' described by Dascal, Chilcott & Lester (1992). Glasspipettes were prepared from Pasteur pipettes cut to a diameter of1'5-2 mm. The tip of the pipette was fire polished to an internaldiameter of 0'5-0-6 mm and the inner surface of the tip coatedwith a thin layer of silicone (Silicone Rhodorcil Pate 430.R,Rhone-Poulenc, Paris). An oocyte was aspirated against the tip ofthe pipette and maintained there by a low, constant negativepressure (5-10 cmH2O) as illustrated in Fig. 2. Current was
J Physiol.491.3

X. Wang, F Jaisser and J-D. Horisberger
Table 1. Steady-state Bufo Na`-K+ pump current expressed in Xenopus oocyte
Ip (2-e) mut/WT ratio Ip (HPO) mut/WT ratio(nA) (nA)
3 0 + 2'3 (7)219+ 22(45)173 + 11 (48)126 + 14**t (41)
561 + 57 (9)0 79 461 ± 41 (12)0-58 334 + 63 * (7)
Forward Na+-K+ pump K+-activated current (Ip) after inhibition of the endogenous Na`-K+ pump byincubation with 0-2 /M ouabain. The table shows results from all experiments using the two-electrodevoltage-clamp technique, Ip (2-e), and those obtained using the half-permeabilized oocyte technique,Ip (HPO). The ratio between the mean value for the N-truncated mutant and that for the wild-typeNa+-K+ pump is indicated by mut/WT ratio. * and ** indicate mean values significantly different fromthat for the wild-type group (P < 0 05 or 0-001, respectively). t indicates a significant difference betweenT31 and T40 truncated-mutant groups (P < 0i01). n is shown in parentheses.
passed between two chloridized silver wires, one in the bath andthe other in the pipette. Membrane voltage was measured betweena 3 M KCl in 3% agar electrode in the bath and a microelectrode(filled with 3 M KCl, resistance about 1 MQ2) impaling the freesurface of the oocyte (see Fig. 2). The pipette solution contained(mM): Na+, 70; K+, 20; Mg2+, 1; Cl, 2; EDTA, 1; gluconate, 90;Hepes, 10; pH 7*4. The electrical resistance of the oocytemembrane facing the inside of the pipette was reduced by additionof digitonin inside the pipette to a final concentration of 8 /M. Inexperiments using the two-electrode voltage-clamp technique, we
observed that this concentration of digitonin decreased the oocytemembrane resistance by more than 100-fold (data not shown). Weestimated the conductance of the membrane exposed to the bathsolution (Gb) and the leak conductance of the pipette seal (Ge) usingthe equivalent circuit analysis described in Fig. 2B and C. Fromten measurements we obtained mean values of 9-1 + 1X0 jS for Gb
and 4 9 + 0-8 juS for C8. The capacitance of the membrane exposedto the bath - estimated using the time integral of the transientcurrent following a 20 mV step after permeabilization and in thepresence of ouabain - was 350 + 10 pF (n = 28). To measure thepre-steady-state current due to the Na+-K+ pump, we recordedcurrent and voltage during a series of eleven 40 ms voltage stepsbetween the limits +50 and -150 mV, with a 100 #as samplingrate. Ten runs of recordings were averaged. The current signal wasfiltered at 5 kHz using an 8-pole Bessel filter (Frequency Devices,Haverhill, MA, USA). The ouabain-sensitive current was obtainedby subtraction of the current measured about 1 min after additionof 2 mm ouabain from that measured before.
Solutions and drugsThe composition of the Nae-free solution used for determiningactivation kinetics of the steady-state current K+ was (mM): Mg2+,0-82; Ca2+, 0-41; Ba2+, 5; tetraethylammonium (TEA), 10; Cl-,22-4; Hepes, 10; N-methyl-D-glucamine (NMDG), 10; sucrose 140;pH 7-4. The composition of the Nae-containing solution useddetermining activation kinetics of the steady-state current K+ andfor the pre-steady-state current measurements was (mM): Na+,92-4; Mg2+,0282; Ba2+, 5; Ca2P, 0-41; TEA, 10; Cl-, 22-4; HCO3-,2-4; Hepes, 10; and gluconate, 80; pH 7*4. The various K+concentrations were obtained by addition of the required amountof potassium gluconate. For proton-current measurements a
solution similar to the Nae-free solution was used, except thatHepes was replaced by Mops and the pH was set to 6-0 (Wang &Horisberger, 1995). Ouabain was obtained from Sigma.
RESULTSExpression of wild-type and mutant Bufo Na+-K+pumps
The mean K+-activated outward current at -50 mV ineach experimental group and in a group of non-injectedoocytes exposed to the same conditions is shown in Table 1.A K+-activated current was barely detectable in non-
injected oocytes indicating that pre-incubation with 0i2 /LM
ouabain had effectively inhibited the Xenopus endogenousNa+-K+ pump. On the other hand, a sizeable current was
measured in each group of oocytes injected with a- andfl-subunit cRNA, demonstrating that the ouabain-resistantBufo Na+-K+ pump was well expressed. These values were
above those due to the endogenous Xenopus Na+-K+ pumpof 40-100 nA that we usually observe in native oocytes(Horisberger et al. 1991; Jaunin et al. 1992; Jaisser et al.1994). The K+-activated Na+-K+ pump current was
significantly smaller in the T40 truncated form than in thewild type (WT) or the T31 truncated form. Table 1 alsoshows the steady-state K+-activated Na+-K+ pump currentmeasured using the half-permeabilized oocyte technique.Although only about half of the oocyte surface contributesto the measurement (see Methods and Fig. 2), the Na+-K+pump current was clearly larger with this technique. Wehave at present no explanation for this difference. However,the ratio of the Na+-K+ pump current for a given truncatedgroup and that for the wild-type group was very similar tothe corresponding ratio derived using the two-electrodetechnique (Table 1) and, as shown below, estimates of theturnover rate were similar to those obtained using the two-electrode technique.
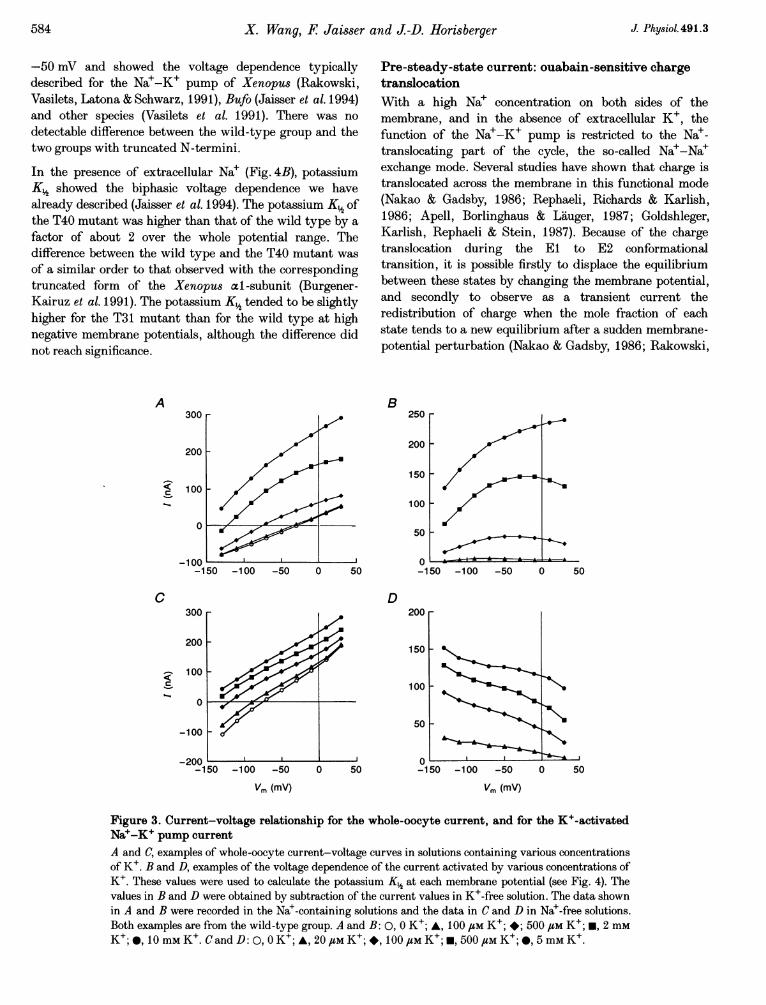
Potassium activation kineticsFigure 3 shows examples of whole-oocyte current-voltagecurves and of K+-activated current-voltage curves in Na+-containing and Na+-free solutions. Figure 4 shows thevoltage dependence of the external potassium half-activation constant (K%). When measured in the absence ofextracellular Nae (Fig. 4A), K% was close to 120 /SM at
Non-injectedWTT31T40
082059
J Physiol.491.3582

J Physiol.491.3 Role of N-terminus of Na+-K+ pump a -subunit 583
A BFluid inlet
Aspiration
Va
Gb
C11 \ t 0 > / Permeabilized 15
membrane
10~~~~~~
----- Intact 'active'membrane
5......--- i - ---------
V2;~~~~~~- :::|_1gs---- -- ---:: : :.
------:-~----~-~-~-~-~-~-------=_V~~~~~~~~~~~ 0
--------------- ---- ------------- ---:-- -- 0 02 0-4 0o60o81-0~e--- ------------- --~--~--~---------- -~-~-~-,-~- ---b(R Rb/ARa+ Rb)
Figure 2. The half-permeabilized oocyte (HPO) techniqueA, schematic (not to scale) illustrating the principles of the half-permeabilized oocyte technique. Theoocyte is aspirated against the tip of a large, fire-polished and silicone-coated Pasteur pipette andmaintained there by constant aspiration (5-10 cmH20). The free surface of the oocyte is impaled by aconventional microelectrode and the membrane potential measured between this intracellular electrode(V2) and an agar bridge placed close to the oocyte (V1). Digitonin (8 uM) can then be added to the pipettesolution through the fluid inlet to permeabilize the part of the membrane exposed to the inside of thepipette (membrane 'a'). The constant negative pressure inside the pipette ensures that no digitonin reachesthe free surface of the oocyte. Chloridized silver wires in the pipette (12) and bath (II) allow current to bepassed and the potential across the intact, 'active' part of the membrane exposed to the external solutionto be clamped (about half of the total membrane area, membrane 'b'). The large area of the permeabilizedmembrane (about a quarter of the total oocyte area) allows the passage of a large current through theoocyte without the problems resulting from use of a point-like source of current and the associatedproblem of inhomogeneity of current density encountered with the classical two-electrode oocyte clamptechnique. B, equivalent electrical circuit used to evaluate the leak resistance (R. = I/G.) and conductanceof the membrane exposed to the pipette solution (Ga = I/Ra) or to the bath (Gb = I/Rb). This analysisfollows the principles used to calculate membrane and paracellular resistance in epithelia after apicalmembrane permeabilization (Broillet & Horisberger, 1991). In this circuit, the following relationships hold:
Ot = Gs + h/(Ra + Rb),
fRb = Rb/(Ra + Rb) =A Vb/(A Va + A Vb),
where fRb is the fractional resistance of membrane 'b'. Therefore, we can write Gt = G8 + GbfRb. Wemeasured G0 (as AI/A V) and fRb before and during permeabilization of membrane 'a' with digitonin, amanoeuvre that primarily modifies Ga. As shown in the example in panel C, expressing Gt as a function offRb during membrane permeabilization gives a straight line which yields G8 at the origin (i.e. extrapolatedfor a 0a value of 0) and C8 + Gb when fRb is close to 1, i.e. after full permeabilization of membrane 'a'.
X. Wang, F Jaisser and J-D. Horisberger
-50 mV and showed the voltage dependence typicallydescribed for the Na+-K+ pump of Xenopus (Rakowski,Vasilets, Latona & Schwarz, 1991), Bufo (Jaisser et al. 1994)and other species (Vasilets et al. 1991). There was nodetectable difference between the wild-type group and thetwo groups with truncated N-termini.
In the presence of extracellular Na+ (Fig. 4B), potassiumK% showed the biphasic voltage dependence we havealready described (Jaisser et al. 1994). The potassium K½ ofthe T40 mutant was higher than that of the wild type by afactor of about 2 over the whole potential range. Thedifference between the wild type and the T40 mutant wasof a similar order to that observed with the correspondingtruncated form of the Xenopus al-subunit (Burgener-Kairuz et al. 1991). The potassium K½ tended to be slightlyhigher for the T31 mutant than for the wild type at highnegative membrane potentials, although the difference didnot reach significance.
A
c
C
c
300
300
200
100
-1 00 - -at
-200-150 -100
Pre-steady-state current: ouabain-sensitive chargetranslocationWith a high Na+ concentration on both sides of themembrane, and in the absence of extracellular K+, thefunction of the Na+-K+ pump is restricted to the Na+-translocating part of the cycle, the so-called Na+-Na+exchange mode. Several studies have shown that charge istranslocated across the membrane in this functional mode(Nakao & Gadsby, 1986; Rephaeli, Richards & Karlish,1986; Apell, Borlinghaus & Liiuger, 1987; Goldshleger,Karlish, Rephaeli & Stein, 1987). Because of the chargetranslocation during the El to E2 conformationaltransition, it is possible firstly to displace the equilibriumbetween these states by changing the membrane potential,and secondly to observe as a transient current theredistribution of charge when the mole fraction of eachstate tends to a new equilibrium after a sudden membrane-potential perturbation (Nakao & Gadsby, 1986; Rakowski,
B250 r
200 F
150 F
100 1
50
50
D
-50 0 50
Vm (mV)
0O-150 -100
200 r
150 F
100 F
50 F
O L
-50
v-150 -100 -50
Vm (mV)
0 50
N011
N.
0 50
Figure 3. Current-voltage relationship for the whole-oocyte current, and for the K+-activatedNa`-K+ pump currentA and C, examples of whole-oocyte current-voltage curves in solutions containing various concentrationsof K+. B and D, examples of the voltage dependence of the current activated by various concentrations ofK+. These values were used to calculate the potassium K½ at each membrane potential (see Fig. 4). Thevalues in B and D were obtained by subtraction of the current values in K+-free solution. The data shownin A and B were recorded in the Na+-containing solutions and the data in C and D in Na+-free solutions.Both examples are from the wild-type group. A and B: 0, 0 K+; A, 100 /M K+; *; 500 FM K+; m, 2 mMK+; *, 10 mm K+. Cand D: 0, 0 K+; A, 20 #M K+; *, 100 uM K+; *, 500 uM K+; *, 5 mm K+.
A A --- A
V".
584 J Physiol.491.3
oll--*.

Role of N-terminus of Na+-K+ pump a-subunit
1993; Holmgren & Rakowski, 1994). With regard to thecharge translocation, we can use the simplified reactionscheme (Nakao & Gadsby, 1986):
kcfW( V)Nai + E1 f E2 + Na+.
kbw(V)
Fast voltage steps result in very large currents because ofthe large capacitance of the oocyte membrane. To be able tostudy the charge translocation due to the Na+-K+ pump itis therefore necessary to record pre-steady-state currentbefore and after inhibition of the Na+-K+ pump withouabain and to obtain the ouabain-sensitive transientcurrent by subtraction, and also to have a good qualityvoltage clamp during the voltage steps before and afterouabain treatment. Figure 5 shows original recordings ofthe voltage and current traces obtained before and after theaddition of 2 mm ouabain. The voltage trace A, recordedbefore addition of ouabain, shows that a stable voltage clampwas obtained within about 1 ms. Trace B, the differencebetween the voltages recorded before and after ouabain,shows that a very good control of the membrane potentialwas obtained during the voltage steps, as the voltagesrecorded before and after ouabain differed by no more thana small fraction of a millivolt even during +100 mV voltagesteps. Trace E, the difference between the currents recordedbefore (trace C) and after (trace D) addition of ouabain,shows the presence of a sizeable ouabain-sensitive transientcurrent with a monoexponential relaxation rate, at least forthe part starting 2 ms after the command of the voltagestep. The shape of the current trace during the first 2 mswas variable, showing sometimes a rising phase, sometimesa fast decaying component with a relaxation rate faster
than 1 ms. As a result of the limited time resolution of ourvoltage-clamp system it was not possible to obtain reliableinformation concerning the fast component. Therefore, allsubsequent analysis of kinetics and charge distribution willbe restricted to the slow component of the ouabain-sensitive pre-steady-state current.
The ouabain-sensitive charge displacement (Qou) is theintegral over time of the ouabain-sensitive current.Assuming an exponential decrease in this current with arate constant kon:
I= I(O) exp(-1kont),where t is time. We can use the values of kin obtained fromthe exponential fit (see below) and the current valueextrapolated to t = 0 to calculate Qou as the integral of thecurrent (I) over time:
Qou = I(0)/kncThe mean values of Qou for the WT, T31 and T40 groupsare shown in Fig. 6. Parameter fitting with the Boltzmannequation:
Qou = Qmin +Qmax
1 + exp(z,(V, - V)(F/RT)'where F is the Faraday constant, R is the Boltzmannconstant and T is the temperature, yielded the maximaldisplaceable charge (Qmax), the mid-point potential (Va) andthe slope factor (zn,). The mean values for these parametersobtained by fitting the Boltzmann equation with the data ofeach individual experiment are shown in Table 2. The slopefactor zI was similar for the three groups, whereas the Qmaxtended to be larger for the truncated forms. Assuming that
No Nao
-100 -50Vm (mV)
B +Nao4 r
2
O L'-1500 50 -100 -50
Vm (mV)
Figure 4. Voltage dependence of the external potassium half-activation constantHalf-activation constants were obtained by fitting the parameters of the Hill equation to the steady-statecurrent induced by increasing the concentration of K+ (see Methods). A, in the absence of external sodium(n = 21, 20 and 14 for the wild type (WT), T31 and T40 truncations, respectively). B, in the presence ofexternal sodium (n = 10, 11 and 11 for the WT, T31 and T40 truncations, respectively). 0, WT; *, T31;A, T40.
A800 -
600
400
200 -
0--150
BAT40
0 50
J. Physiol.491.3 585
3 FA
T31
-w

X. Wang, F Jaisser and J-D. Horisberger
the ouabain-sensitive pre-steady-state current is due to themovement of exactly one charge per pump across themembrane (Nakao & Gadsby, 1986), Qmax yields anestimate of the absolute number of Na+-K+ pumps(np= QmaxF) in the membrane. On the other hand IpFyields the Na+-K+ pump current in elementary chargeunits, which is equal to the number of Na+-K+ pumpstimes the current produced by each pump (nip= IpF).Dividing the Na+-K+ pump current Ip (values given inTable 1) by Qmax yields iE, the number of charges trans-ported by each Na+-K+ pump per second (i.e. the Na+-K+pump turnover rate, as we know that one net charge istransported during each cycle). The mean values forturnover rate are indicated in the last line of Table 2. Theturnover rate obtained in the Bufo wild-type group(59 7 + 9.3 s-') is in good agreement with the turnoverobtained by counting the ouabain binding sites andmeasuring the Na+-K+ pump current (48 + 3 s-') usingthe Xenopus Na+-K+ pump under similar experimentalconditions (Jaunin et al. 1992). This suggests that the
A
B
C
D
E
assumption of the translocation of one charge per pump isvalid. The turnover rates were significantly smaller in bothN-truncated forms. The values for the WT (59 7 s-1) andT40 (16-3 s-1) groups were in very close agreement withthose (47 5 and 14-2 s-', respectively) obtained using thetwo-electrode voltage-clamp technique in preliminaryexperiments (Horisberger et al. 1994).
The ouabain-sensitive charge translocation can also becalculated as the time integral of the ouabain-sensitivecurrent. While this method does not depend on the currenthaving a monoexponential relaxation rate, it does requirethe inclusion of the fast initial component of the transientcurrent. Using the integration routine of the pCLAMPprogram to obtain Qou by integration of the currentbetween 0 and 20 ms after the beginning of the voltagestep yielded results qualitatively similar to those obtainedby the method described above. However, all the Qmaxvalues were slightly larger due to the inclusion of the fastinitial component in the second type of calculation (datanot shown).
Figure 5. Original recordings of voltage and currenttraces obtained using the half-permeabilized oocytetechniqueThe traces were recorded over 100 ms periods with 40 msvoltage steps within the limits -100 to +100 mV. A, voltagerecordings during a series of voltage steps before addition ofouabain. B, difference between the voltages recorded before and1 min after ouabain (note the expanded scale). C and D,recordings of the total current across the oocyte recorded,respectively, before and 1 min after addition of 2 mm ouabain.The ouabain-sensitive current, i.e. the difference between thecurrents shown in C and D, is shown in E (note difference scale).The traces of A to E were recorded in one oocyte of the T31group. The inset shows two superimposed traces of the ouabain-sensitive current during a single voltage step from -50to + 50 mV in an oocyte of the wild type (thin line) and one ofthe T40 group (thick line). Note the large difference between thegroups in relaxation rate for both 'on' and 'off' tail currents.
J Physiol.491.3

Role of N-terminus of Na+-K+ pump a-subunit
Figure 6. Ouabain-sensitive charge translocation (Qu)The charge translocation, calculated as the time integral ofthe pre-steady-state current, is plotted as a function of themembrane potential. The curves are drawn using the bestfitting parameters from the equation:
Qou = Qmin + Qmax/(l + exp(zq( Vq- V)F/RT)).For the wild type (WT; n = 9), the T31 mutant (n = 12) andthe T40 mutant (n = 7) the values of Qmax were, respectively,11-3, 16-5 and 19-8 nC; the values of zI were 0O85, 088 and0-91; and the values of V. were -73, -45 and -22 mV. Thearrows point to the Vq value for each group. Mean values forthe best-fitting parameters in each individual experiment areshown in Table 2. Error bars indicate S.E.M.
15 r
10 H
c
03
5h
-V
-5
-10-150 -100 -50
Vm (mV)
Pre-steady-state current: relaxation ratesThe relaxation rates of the 'on' tail current (kon) and 'off'tail current (k/ff) were determined by fitting a singleexponential equation to the current values starting 2-4 msafter the initial change induced by the command potential.The mean values of kon for each voltage step are shown inFig. 7. The mean values of kff, which were similar for allvoltage steps in a given group, are indicated by the largesymbols at the -50 mV point. In all three groups, konshowed a clear voltage dependence with the rate increasingprogressively at negative potentials.
To analyse the kinetics of charge translocation we can use
the simplified reaction scheme:
Na +E1k
E2+Na+.
iC( V)
The relaxation rate of the transient current (kin( V)) is equalto the sum of the forward and backward rate constants atthe newly set potential (V) (Nakao & Gadsby, 1986):
kon( V) = kfw( V) + 4w( V).
The Qou- V curve (Fig. 6) shows that at -130 mV thecharge distribution is nearly completely displaced towardsEl (Qou t Qmin) indicating that CbW(-130) is much largerthan kfw(-130). Thus, at -130 mV, kon provides a good
Figure 7. Voltage dependence of the relaxation rate consta(k) of the pre-steady-state ouabain-sensitive currentThe mean 'on' relaxation rate for the ouabain-sensitive current atbeginning of a voltage step (from a holding potential of -50 mVexpressed as a function of voltage. The mean values at -130 and+50 mV are also given in Table 2. The mean 'off' relaxation ratethe ouabain-sensitive current at the end of a voltage step is giverthe large symbols at -50 mV. Error bars indicate S.E.M.
estimate of kbw. Similarly, at +50 mV, the chargedistribution is almost completely displaced towards E2(Qou ; Qmin + Qmax), indicating that kfW(+50) is muchlarger than lcbW(+50). Thus, at +50 mV, kon(+50) providesa good estimate of k/fw. The mean values of kon(-130) andkon(+50) are indicated in Table 2.
Figure 7 shows that all values of kon at positive membranepotentials were significantly smaller in the two groups withan N-terminal truncation than in the wild-type group andeven smaller in the T40 group than in the T31 group.
However, there was no significant difference in the values athigh negative membrane potentials. This indicates that k/fw- the rate constant that determines the re-equilibration ofcharges when the membrane potential is changed from a
negative to a positive value by a conformational changefrom the El to the E2 state - was significantly smaller forthe N-terminal truncated forms of the Na+-K+-ATPase.The difference was larger, close to a 4-fold reduction (Table2), for the T40 mutant. No such difference was observed forkbw at high negative membrane potential. In summary themajor difference between the wild-type and N-truncatedforms was that the latter showed a reduction of the rate ofthe slow component of the charge translocation associatedwith the forward El to E2 conformational change.
int 400
U-
-)
300
200
100
0
-150 -100 -50
V. (mV)0 50
T40
~-~---* T31
_WT
0 50
J Physiol.491.3
1.n

X. Wang, F Jaisser and J-D. Horisberger
Table 2. Results from experiments involving pre-steady-state current measurements
n kon(+50 mV) kon(-130 mV)(s-') (s-')
WT 9 248+ 18T31 12 146 + 5***T40 7 68 + 9*** ttt
437 + 53400 + 14407 + 32
Vq(mV)
-74 + 14-45 + 3***-20 + 4*** ttt
Zq
0 93 + 0 050-92 + 0 040-91 + 0-06
Qmax(nC)
115+ 2-316-6 + 1-020-1 + 1*1 * t
IP(K)(nA)
561 + 57461 + 41334 + 63*
Turnover rate(s-')
59 7 + 9'328-7 + 2.6**16-3 + 2-7**tt
*, ** and *** indicate mean values significantly different from that of the wild-type (WT) group(P < 0 05, 0f01 and 0-001, respectively). t, tt and ttt indicate significant difference between T31 andT40 truncated-mutant groups (P < 0 05, 0-01 and 0-001, respectively).
Ouabain-sensitive proton-dependent inward currentIn all three groups sizeable ouabain-sensitive proton-dependent inward currents (lnw,H) were observed innominally Na+- and K+-free solution with a pH of 6-0. Thelnw,H showed a similar voltage dependence in all threegroups (see Fig. 8), though the absolute value of the inwardcurrent was larger in the T40 group. Table 3 shows themean values for the forward K+-activated Na+-K+ pumpcurrent at -50 mV (Ip), for the ouabain-sensitive inwardcurrent at -110 mV and pH 6X0 and for the ratio betweenthese two variables. The combined effect of the reduction inthe forward K+-activated current and of the increase in theinward current resulted in a much larger Iinw,H/Ip ratio inthe T40 group than in the wild-type group. A smaller effecton the lnw,H/Ip ratio was observed in the T31 group.
DISCUSSIONThe transport cycle of the Na+-K+ pump can be dividedinto two main components: a Na+ translocating limb,which, in the normal functional mode, carries 3 Na ionsout of the cell and includes an El to E2 conformationalchange, and a K+ translocating limb, which moves two K+into the cell and reverses the conformation back to El. TheNae translocating limb has been shown to move anelectrical charge across the membrane (Goldshleger et al.1987; Nakao & Gadsby, 1986) and this charge translocationcan be observed as a pre-steady-state ouabain-sensitivecurrent when the membrane potential undergoes rapidperturbation. Our measurements of the pre-steady-stateouabain-sensitive current in the Bufo wild-type Na+-K+
50-
0
-50-
' -100
-150 -
-200 -
-250-150 -100 -50 0 50
pump expressed in Xenopus oocytes show a relationshipbetween the relaxation rate and the membrane voltagesimilar to that observed in other experimental models(Nakao & Gadsby, 1986; Rakowski, 1993; Holmgren &Rakowski, 1994). The relaxation rate tended to be constantat positive membrane potentials and to increase withincreasingly negative membrane potentials. Several pointsin our data show that the N-terminal truncations wereassociated with a slower rate constant for the chargetranslocating step linked to the forward El to E2conformational change. First, all the values determined forthe rate constant at positive potentials were significantlysmaller in the N-truncated groups. Second, there was ashift towards positive potentials of Vq, the potential atwhich the moveable charges associated with the Na+-K+pump are equally distributed across the membrane'selectrical field. This shift is most easily explained by adecrease in the forward rate constant while the backwardrate constant is unaltered (or at least altered less than theforward rate constant).
The outward movement of Na+ ions is an essential part ofthe normal Na+-K+-exchange transport cycle and if areduction in the rate constant were to make thecorresponding step rate limiting, any step in this part ofthe cycle then would have the facility to cause a reductionof the overall turnover rate (Heyse, Wuddel, Appell &Stiirmer, 1994). Indeed, we observed that the truncatedforms of the enzyme showed a lower activity when assessedin terms of forward K+-activated Na+-K+ pump current(Table 1). This lower level of activity could, in principle, also
Figure 8. Ouabain-sensitive proton-dependent inwardcurrentCurrent-voltage relationship for the current sensitive to 2 mmouabain measured in a pH 6-0, nominally Na- and K+-freesolution. Number of measurements was 14, 17 and 16 for theWT, T31 and T40 groups, respectively. Mean values for theforward Na+-K+ pump current at -50 mV (Ip) measured inthe same oocytes are given in Table 3.
Vm (mV)
588 J Physiol.491.3

Role of N-terminus of Na+-K+ pump ax-subunit
Table 3. K+-activated Na+-K+ pump current, Ip, and ouabain-sensitive proton-dependent inward current IinwHn Ipat-50mV
(nA)WT 14 237 + 29T31 17 122 + 11 ***T40 16 63+10***ttt
inw,H at-110 mV -lnw,H/Ip ratio(nA)
-125 + 16-85 + 8*-164 + 19*ttt
0 53 + 002073 + 0 04**2 81 + 0-12***ttt
*, ** and *** indicate mean values significantly different from that of the wild-type group (P < 0 05,0-01 and 0 001, respectively). t, tt and ttt indicate significant difference between T31 and T40truncated-mutant groups (P < 0 05, 0 01 and 0 001, respectively).
result from a lower expression of the truncated proteins inthe Xenopus oocyte. However this seems unlikely to be theexplanation as two other estimates of the density of activeNa+-K+ pumps in the membrane show higher rather thanlower values for the truncated forms (see below). This Na+-translocating limb of the cycle includes several distinctsteps, the binding and occlusion of internal Na+ ions, theEl to E2 conformational change, the release of a first Naeion followed maybe by additional conformational changeand the release of the two other Na+ ions:
3 Nai + El = EINa3 . E(Na3) = E2Na3 =
E2Na2 + 1 Na+ = E2 + 3 Na+.
High time resolution measurements (Hilgemann, 1994) andother techniques (Stiirmer et al. 1991; Heyse et al. 1994)have recently ascribed the main charge translocating eventto the deocclusion and release of the first Na+ ion to theoutside medium. Our data do not allow us to determinewhich of the discrete step(s) of this part of the cycle may bedirectly involved. A reduction in the rate of Na+ deocclusionor of any preceding rate-limiting step(s) could equally wellaccount for our observations. In summary, our data showthat truncation of a-subunit's N-terminus induces areduction in the speed of the outward translocation of Na+that could occur either by the slowing of the rate ofdeocclusion or of an associated rate limiting step.
Our present data on the activation kinetics for external K+associated with the Bufo Na+-K+ pump confirm our earlierreport on the Xenopus pump (Burgener-Kairuz et al. 1991)and indicate that the larger 40-amino-acid truncated forminduced a 2-fold decrease in the apparent affinity forexternal K+ when this was measured in the presence ofexternal Na+. This effect was not apparent in external Na+-free solution. The shorter truncated form (T31) did notinduce a significant change in apparent K+ affinity. Ourmeasurements of the proton-dependent ouabain-sensitiveinward current showed that N-terminal truncation induceda relative increase in this current over and above the K+-activated current that represents the physiological Na+-K+exchange function of the pump.
How can these changes in maximal turnover rate and inapparent affinity for external potassium be related to thedecrease in the forward rate constant of the Na+ trans-locating limb of the pump cycle observed with the
N-truncated form of the enzyme? To address this questionwe used the simplified four-state model described in theAppendix. This model includes four rate constants, oneequilibrium constant, kN, which describes the equilibriumbetween the ElK and ElNa states, and the intrinsicbinding constant for K+ binding to its external site, kd.The forward (,f) and backward (bl) rate constants standfor the Na+-carrying, charge-translocating steps whichinclude the El to E2 conformational change, the de-occlusion and the release of Na+ to the external mediumand are therefore the equivalent of the kfw and b,w rateconstants that were used earlier in our analysis of theexperimental results. The measurements of charge trans-location rate yielded estimates of the values of f1 and b, inthe presence of extracellular Na+. Assuming that thebackward-running cycle can occur only with an inwardmovement of Na+, the parameter b, takes a value of 0 inthe absence of external Na+. In the presence of extracellularNa+, this b, parameter may cover a large range of valuesdepending on the membrane potential. Finally, the /2 andb2 rate constants summarize all the steps that occurbetween the binding of external K+ and its release on theintracellular side.
We have used eqns (A13) and (A17) of the Appendix toexamine the effect of a change in f1 on maximal turnoverrate (Vmax) and on potassium half-activation constant (K1,"),i.e. the external K+ concentration resulting in a half-maximal trunover rate, a measure of the apparent affinityof K+ for its external site. Figure 9A shows the predictedvalues of Vmax and K% as a function of f1 in one examplewith the values of ckx, /2 and b2 given in the legend. In theabsence of external Nae, the backward E2 to El reaction isnot possible and this situation is represented by a b, valueof 0 (open symbols, Fig. 9A). In the presence of Nae, weknow that the backward rate constant has a value of about400-500 s-5 (see koon(-130 mV), Table 2) at high negativepotentials and that it decreases with increasing membranepotential. We have chosen a value for b, of 200 s-' (filledsymbols, Fig. 9A) to represent the situation in the middleof the potential range. This example shows that Vmaxincreases monotonically with f/ in both cases and isindependent of the presence of external Na+. While K%tends to increase slightly in the absence of external Nae,there is a marked decrease in the K% values with increasingf1 in the presence of Na+. We next examined the effect of a
J Physiol.491.3

X. Wang, F Jaisser and J-D. Horisberger
change of f/ from 250 to 70 s-' (these approximate to themean values of klfw observed in the wild-type and T40mutant, respectively, at positive membrane potentials, andwe asssume that they are weakly voltage dependent) over awide range of possible values for kN, f2 and b2, since wehave no a priori knowledge of the real values of thesevariables. To this end we have calculated the K; and Vmaxpredicted when /2 takes a large range of values from 80 to10000 s-' and for three values (1 0, 10 and 100) of kN.From our calculations of turnover rate in the present study(see Table 2) and earlier measurements (Jaunin et al. 1992),we know that the cycle turnover rate in the wild-typeNa+-K+ pump is close to 50 s-'. For each set of kN and /2values, b2 was set so that the resulting Vmax (calculatedfrom eqn (Al 2) of the Appendix) would be equal to 50 s-'when calculated using the wild-type value for the fi rateconstant (250 s-1). The change in Vmax and K4 resultingfrom a 250 to 70 s-f decrease in f/ is illustrated in Fig. 9B.This figure shows that a 250 to 70 s-' reduction in theforward rate constant f/ results in a significant increase inK,f when the b, rate constant has a finite value (200 s-')which is of the same order as the f/ constant, simulatingconditions with external Na+. In contrast, when b1 is set at0 (simulating conditions with no external Na+) the changein apparent affinity is minor, at least when the product/2/e is high enough. These results fit closely with our
observations of the changes in K4 for potassium and Vmax
A5 r
4
3
Vmax
2 _
0 _10
No NaO
100
f, (S 1)1000
200
in the absence and presence of external Na+ and show thatthere is no need to suppose a direct involvement of the K+binding site to account for the observed change in theapparent affinity for K+. In fact, the absence of anydetectable effect on the apparent affinity for external K+over a large range of membrane potentials in the absence ofexternal Nae strongly suggests that the K+ binding siteitself is not significantly modified by N-terminal truncation.This is not surprising in view of the intracellular location ofthe N-terminus of the a-subunit. However, there is adiscrepancy between this conclusion and that of Vasilets etal. (1991) who, using the a-subunit of the Torpedo Na+-K+pump expressed in Xenopus oocytes, observed thatN-terminal truncation induced an increase in the apparentaffinity for external K+, and also an increase in the voltagedependence of this parameter. At present, we cannotprovide any documented explanation for this discrepancy,but both the species difference and the difference in theposition of the truncation are possible factors.
The change in the turnover rate predicted by the model for a250 to 70 s-5 decrease in the fi rate constant is a reduction toabout 30% of control, at least for high /2 values. Thischange is clearly larger than the difference between the meanforward pump currents for wild-type and T40 mutantgroups, the latter showing a value which is 60-70% ofthat for the wild type (Table 1). However, two independent
B3.5 r
- 3-00
150 CM11 2-5CY
Q.t20100 :E N
11 1-5
0.50 ,0
m
0 0
F
100 1000f2 (s ')
10000
Figure 9. Na+-K+ pump four-state model: effect of a change in f1 on the maximal turnover rateand the apparent K+ affinity (see Appendix for definition of kinetic paramneters)A, relationship between f, and Vmax (O) or apparent K;, (A and *) for an intrinsic K+ affinity of 1X0 mm.A, values obtained with a b1 value of 0 s' (no external Na+); *, values obtained with a b1 value of 200 s-(with external Na+). The Vmax values were independent of b, (identical whether or not external Nae waspresent). The values of f2, b2, and kN used in this simulation were 200 s-5, 400 s-' and 10, respectively.B, the graphs represent the relative increase in apparent K+ affinity (K& filled symbols, withb, = 200 s-, open symbols, with b1 = 0 s1) calculated from the eqn (At 7), or the relative decrease inmaximal turnover rate (Vmax, dotted line, no symbols), calculated from eqn (At 3) when the value of theparameter fi is decreased from 250 to 70 s-I. The calculation of K4 was made for three values of theequilibrium constant kN (1P0, 0 and 0; 10, A and A; and 100, * and E) as indicated. The kN value isexpected to be high under our experimental conditions (sodium loading). Data are shown over a largerange of values of the /2 rate constant. The b2 parameter was chosen so that Vmax would be equal to 50 s-1when calculated with the 250 s5 fi parameter. The decrease in Vmax was the same, whether in thepresence or absence of external Nae, and was independent of kN.
O
590 J Physiol.491.3
F1

Role of N-terminus of Nat+-K+ pump cc-subunit
estimates of the number of active Na+-K+ pumps, namelythe ouabain-sensitive charge translocation (Q0u see Table 2)and the ouabain-sensitive proton-dependent inward current(IilnwH, see Table 3), reveal higher values for the truncatedforms. Thus, it seems probable that the truncated formswere indeed expressed in the oocyte at a higher level thanwas the wild type. Because of the relatively low affinity ofouabain for the Bufo Na+-K+ pump, it was not possible toassess the absolute number of Na+-K+ pumps by measuringthe ouabain binding. For this reason, we were unable toobtain an independent estimate of the number of Na+-K+pumps. However, the reduction in turnover rate calculatedfrom the Na+-K+ pump current and Qou (Table 2, last line)fits rather well with the prediction made by the model.
In summary, a coherent interpretation of all our data isthat the T40 N-terminal truncation induces a 3- to 4-folddecrease in the charge-translocating outward Na+-movingstep (fi), and that the consequences of this change are a3-fold reduction in maximal turnover rate and a 2-foldincrease in apparent K%,, for external K+, when measured inthe presence of external Na+. The shorter T31 truncationinduces changes in the same direction, although of smallermagnitude. It is obvious that the experimental datapresented here can also be explained by many possiblemodels with alteration of multiple kinetic parameters. Thedata of Daly et al. (1994) support kinetic alterations in adifferent part of the cycle. However, the apparently complexeffects induced by the N-terminal truncation are reasonablywell explained by the alteration in the single fi rateconstant in the simple model presented in the Appendix.
Trypsin treatment of Na+-K+-ATPase leads to a rapidcleavage of the N-terminus of the a-subunit (Jorgensen &Klodos, 1978; Jorgensen & Collins, 1986) at a site close tothat of the truncation that we have produced by mutagenesis(see Fig. 1). Jorgensen & Karlish (1980) have studied thepump-cycle kinetics of the trypsinized Na+-K+-ATPase, aform they called an 'invalid' enzyme. The 'invalid' Na+-K+-ATPase had a lower turnover rate and the El =E2equilibrium was shifted towards the El conformation. Ourfinding of a shift of Vq towards positive potentials isequivalent to a shift towards the El conformation at anygiven potential, indicating that our N-truncated mutantsexhibit a physiological behaviour similar to the trypsin-treated 'invalid' enzyme. Wierzbicki & Blostein (1993) havestudied the activation kinetics of the Na+-K+-ATPase byATP and K+ using the trypsin-treated enzyme and, morerecently, similar studies have been performed by Daly et al.(1994) on an N-truncated mutant of the rat a-subunit, atruncation corresponding to that produced by trypsincleavage (see Fig. 1). In both cases, the results showed areduction in the rate of K+ deocclusion at low concentrationsof ATP, a condition in which K+ deocclusion is ratelimiting. It is difficult to compare these results with ours,because our measurements were made in vivo, probablywith a high (physiological) concentration of ATP. Moreover,
the N-terminal truncation in the study of Daly et al. (1994)was not in exactely the same position as that in our study(see Fig. 1).
The finding that an N-terminal truncation decreases theforward Na+-K+ pump current but not the proton-dependent inward current (Iinw H) also supports ourproposed interpretation of the nature of lrw,H We haveproposed that lin, H is not due to a transport cycle, inwhich case we should see an effect of a reduction in fl, butrather to proton 'leak' through a defined configuration ofthe Na+-K+ pump, perhaps E2 or some sub-conformationof E2 (Wang & Horisberger, 1995). With the externalsolution Na+ free and K+ free, the Na+-K+ pump is trappedin the E2 cation-free conformation, and the mole fraction ofE2 is large (close to 1) even if the fi rate constant is slow.Therefore, as long as the proton conductance of the E2state is not modified, Inw,H should yield an estimate of thetotal number of active Na+-K+ pumps in the membrane(i.e. those able to adopt the E2 conformation). This is animportant point because the ouabain-sensitive proton-dependent inward current should then allow assessment ofthe presence of Na+-K+ pumps and some of the functionsof mutants that may have much reduced Na+-K+ exchangetransport or ATPase activity.
It is interesting to note that the two truncations used in thepresent study affect the Na+-K+ pump function in aqualitatively similar manner, although the T31 truncationremoves a highly variable domain and a highly positivelycharged segment, while the T40 truncation removes inaddition the conserved MEELKKEVT segment whichcontains three negatively charged and two positively chargedamino acids. The precise role of each of the highly conservedamino acid residues in this segment may be established bysite-directed mutagenesis in future experiments. Becauseof the intracellular localization of the N-terminus it isdifficult to imagine a direct steric effect of this domain onthe mechanism underlying the de-occlusion/external releaseof Nae. Obviously, an allosteric effect cannot be excluded,but a steric effect on a rate-limiting step preceding Na+deocclusion - such as the occlusion of Na+ or interferencewith the energy transduction mechanism that links phos-phorylation to conformational change - appears moreprobable to us.
APPENDIXWe used the simplified kinetic model of the Na+-K+ pumpillustrated in the scheme below to analyse the repercussionsof a modification in one of the cycle rate constants on theapparent affinity for external K+. This model includes fourstates: ElNa, E2, E2K and ElK. The step from ElNa toE2 summarizes in two rate constants, f1 (forward) and b1(backward), all the events that accomplish the translocationof Na+ ions across the membrane and their release into theoutside medium. The E2 and E2K states are linked by aK+-binding equilibrium. The f2 and b2 rate constants
591J Physiol.491.3

X. Wang, F Jaisser and J-D. Horisberger
summarize all the steps involved in occlusion, translocationand deocclusion of K+. The two El states are related byan equilibrium constant kN. This pseudomonomolecularequilibrium constant depends in reality on intracellular K+and Na+. The full reaction cycle for the intracellular part ofthe cycle should be:
ElK [< - El [ ElNa,kcdK, kd,Na,
where [K+]i and [Na+], are the intracellular Na+ and K+concentrations and kd,K, and kd,Na. the intrinsic affinityconstants for the binding of these ions to their intracellularsites. In this reaction scheme, the ratio between the molefractions ofElK and ElNa is given by:
ElK [K ]icdK,EINa [Na ]ikdlNa1
Under our experimental conditions, i.e. after Na+ loading,the mole fraction of the unoccuppied El state should benegligibly small and the intracellular concentrations shouldnot change significantly within the time frame of theexperiment. We can therefore simplify the intracellularpart of the reaction cycle and replace the full reactionscheme by the simple distribution equilibrium:
kNElK < > ElNa,
with the equilibrium constant:
k [K ]ikd,K,[Na]ikd Na,
kN is a constant because the intracellular concentrationsof Na+ and K+ do not change significantly under ourexperimental conditions.
The full scheme of the cycle would be:
fiElNa-
bE2
+ bl +~~
IN kd [K+]o
/2ElK E2K
b2
where [K+]. is the external K+ concentration and kd itsintrinsic affinity constant.
Using ElNa, ElK, E2 and E2K as the mole fraction ofeach corresponding state, the following equations can bewritten:
ElNa + ElK + E2 + E2K = 1,
ElNa = ElKkN,
E2K = E2([K ]o/1kd).
(Al)
(A2)
(A3)
Under steady-state conditions, the rate of change of the
mole fraction of E1 (= ElNa + ElK) is equal to zero:
ElNaf1 + ElKb2 = E2Kf2 + E2bl. (A4)Using eqns (Al), (A2), (A3) and (A4) we can solve for ElKand obtain:
EIK([K+]O/kd)f2+ b1
b1+ b2 + kN(bl + fl) + ([K+]O/kd)(f2 + b2 + CN(fl +f2))
(A5)
We define
and
a = bI + b2+ kN(bl + fi),
/3=f2+ b2+1N(l + A2),and using eqns (A2), (A3), (A4) and (A5) we obtain the molefraction of each state:
ElK = (f2([K ]o/kcd) + bl)/(a + /J([K+]O/kcd)), (A6)
ElNa = kN (f2([K+]O/kd) + b1)/a + # ([K ]O/kd)), (A7)
E2 = (kNfl + b2)/(a + fl([KK]O/kd)), (A8)
E2K = ([K+]O/kd)(kNfl + b2)/(ac + J([K ]O/kd). (A9)
Under steady-state conditions, the turnover rate (V) isgiven by:
V= ElNaf1 - E2bl, (AlO)
and using eqns (A7) and (A8) in (AlO) we can calculate V:
V= (flf2kx([K ]o/kd) - b1b2)/(az +fl([K+]O/kd)). (A1)The maximal steady-state turnover rate (Vmax) occurs when[K+]O/kd tends to high values or, expressed otherwise,when kd/[K+]o tends to 0. Under this condition, E2 = 0and this allows us to determine the value of Vmax from eqn(A 1) as:
Vmax = kNAff2/fl,or, using the value of , given above:
ICNflf2
(A12)
(A13)max f2+ b2 +ki(fl +f2)To determine the K+ concentration ([K+]%) that produces ahalf-maximal turnover rate (V%), we use eqn (Al 1) whichthen becomes:
V% = (flf2kiN([K ]/kd) - b1b2)/(z + f([K+]%/kd)).(A14)
Using eqns (A12) and (A14) in the relation 2 V% = Vmax, weobtain:
2 (f, f2 kN([K+]4/kd) - b, b2)/(a + /8([K ]%/kd)) = kN f, f2)//1,(A15)
and we solve for [K ]4/kd:[K ],/2/kd = a/4 + 2blb2/(kNflf2), (A16)
[K+];(=bkd2b
+ bi+ b2+ fkf f+ b f+kl(A+f) (A17)
592 J Physiol.491.3
V ,-

APELL, H.-J., BORLINGHAUS, R. & LXUGER, P. (1987). Fast chargetranslocations associated with partial reactions of the Na,K-pump:II. Microscopic analysis of transient currents. Journal of MembraneBiology 97, 179-191.
BROILLET, M.-C. & HORISBERGER, J.-D. (1991). Basolateral membraneconductance of A6 cells. Journal of Membrane Biology 124, 1-12.
BURGENER-KAIRUZ, P., HORISBERGER, J.-D., GEERING, K. &ROSSIER, B. C. (1991). Functional expression of N-terminustruncated a-subunits of Na,K-ATPase in Xenopus laevis oocytes.FEBS Letters 290, 83-86.
DALY, S. E., LANE, L. K. & BLOSTEIN, R. (1994). Functionalconsequences of amino-terminal diversity of the catalytic subunit ofthe Na,K-ATPase. Journal of Biological Chemistry 269,23944-23948.
DASCAL, N., CHILCOTT, G. & LESTER, H. A. (1992). Intracellularperfusion of Xenopus oocytes. Methods in Enzymology 207,345-352.
EFTHYMIADIS, A., RETTINGER, J. & SCHWARZ, W. (1993). Inward-directed current generated by the Na+,K+ pump in Na+- and K+-free medium. Cell Biology International 17, 1107-1116.
GADSBY, D. C., RAKOWSKI, R. F. & DE WEER, P. (1993). Extracellularaccess to the Na,K pump: Pathway similar to ion channel. Science260,100-103.
GEERING, K., THEULAZ, I., VERREY, F., HAUPTLE, M. T. & ROSSIER,B. C. (1989). A role for the fl-subunit in the expression of functionalNa+-K+-ATPase in Xenopus oocytes. American Journal ofPhysiology 257, C851-858.
GOLDSHLEGER, R., KARLISH, S. D. J., REPHAELI, A. & STEIN, W. D.(1987). The effect of membrane potential on the mammaliansodium-potassium pump reconstituted into phospholipid vesicles.Journal of Physiology 387, 331-355.
HEYSE, S., WUDDEL, I., APELL, H.-J. & STURMER, W. (1994). Partialreactions of the Na,K-ATPase: Determination of rate constants.Journal of General Physiology 104, 197-240.
HILGEMANN, D. W. (1994). Channel-like function of the Na,K pump
probed at microsecond resolution in giant membrane patches.Science 263, 1429-1432.
HOLMGREN, M. & RAKOWSKI, R. F. (1994). Pre-steady-state transientcurrents mediated by the Na/K pump in internally perfusedXenopus oocytes. Biophysical Journal 66, 912-922.
HORISBERGER, J.-D. (1994). The Na,K-ATPase: Structure-FunctionRelationship. Landes, Austin, TX, USA.
HORISBERGER, J.-D., JAISSER, F., CANESSA, C. M. & ROSSIER, B. C.(1994). Na/K-pump mutant with a slow charge translocating step.In The Sodium Pumrp, ed. BAMBERG, E. & SCHONER, W., pp.
549-552. Steinkopff, Darmstadt, Germany.
HORISBERGER, J.-D., JAUNIN, P., GOOD, P. J., RosSIER, B. C. &GEERING, K. (1991). Coexpression of al with putative /33 subunitsresults in functional Na-K-pumps in Xenopus oocyte. Proceedingsof the National Academy of Sciences of the USA 88, 8397-8400.
JAISSER, F., CANESSA, C. M., HORISBERGER, J.-D. & RoSSIER, B. C.(1992). Primary sequence and functional expression of a novelouabain-resistant Na,K-ATPase. Journal of Biological Chemistry267, 16895-16903.
JAISSER, F., JAUNIN, P., GEERING, K., RoSSIER, B. C. &HORISBERGER, J.-D. (1994). Modulation of the Na -K+ pump
function by the fl-subunit isoforms. Journal of General Physiology103, 605-623.
JAUNIN, P., HORISBERGER, J.-D., RICHTER, K., GOOD, P. J., RoSSIER,B. C. & GEERING, K. (1992). Processing, intracellular transport and
functional expression of endogenous and exogenous a-,83 Na,K-ATPase complexes in Xenopus oocytes. Journal of BiologicalChemistry 267, 577-585.
593
JORGENSEN, P. L. (1975). Purification and characterization of (Na+,K+)-ATPase. V. Conformational changes in the enzyme transitionsbetween the Na-form and the K-form studied with tryptic digestionas a tool. Biochimica et Biophysica Acta 401, 399-415.
JORGENSEN, P. L. (1977). Purification and characterization of(Na+ + K+)-ATPase. VI. Differential tryptic modification ofcatalytic functions of the purified enzyme in presence of NaCl andKCl. Biochimica et Biophysica Acta 466, 97-108.
JORGENSEN, P. L. & COLLINS, J. H. (1986). Tryptic and chymotrypticcleavage sites in sequence of alpha-subunit of (Nae + K+)-ATPasefrom outer medulla of mammalian kidney. Biochimica et BiophysicaActa 860, 570-576.
JORGENSEN, P. L. & KARLISH, S. J. (1980). Defective conformationalresponse in a selectively trypsinized (Nae + K+)-ATPase studiedwith tryptophan fluorescence. Biochimica et Biophysica Acta 597,305-317.
JORGENSEN, P. L. & KLODOS, I. (1978). Purification andcharacterization of (Nae + K+)-ATPase. VII. Tryptic degradationof the Na-form of the enzyme protein resulting in selectivemodification of dephosphorylation reactions of the (Na+ + K+)-ATPase. Biochimica et Biophysica Acta 507, 8-16.
LXUGER, P. (1991). Electrogenic Ion Pumps. Sinauer, Sunderland, MA,USA.
NAKAO, M. & GADSBY, D. C. (1986). Voltage dependence of the Natranslocation by the Na/K pump. Nature 323, 628-630.
OHTA, T., NoGUCHI, S., NAKANISHI, M., MUTOH, Y., HIRATA, H.,KAGAWA, Y. & KAwAMURA, M. (1991). The 'lysine cluster' in the N-terminal region of Na+/K+-ATPase a-subunit is not involved inATPase activity. Biochimica et Biophysica Acta 1059, 157-164.
RAKOWSKI, R. F. (1993). Charge movement by the Na/K pump inXenopusoocytes. Journal of General Physiology 101, 117-144.
RAKOWSKI, R. F., VASILETS, L. A., LATONA, J. &SCHWARZ, W. (1991).A negative slope in the current-voltage relationship of the Na+/K+pump in Xenopus oocytes produced by reduction of external [K+].Journal of Membrane Biology 121, 177-187.
REPHAELI, A., RICHARDS, D. E. & KARLISH, S. J. (1986). Electricalpotential accelerates the ElP(Na)-E2P conforinational transition of(Na,K)-ATPase in reconstituted vesicles. Journal of BiologicalChemistry 261, 12437-12440.
SAGAR, A. & RAKOWSKI, R. F. (1994). Access channel model for thevotage dependence of the forward-running Na+/K+ pump. Journalof General Physiology 103, 869-894.
SHANBAKY, N. M. & PRESSLEY, T. A. (1994). Mammalian at-subunitof Na+-K+-ATPase does not need its amino terminus to maintaincell viability. American Journal of Physiology 267, C590-597.
STURMER, W., BUHLER, R., APELL, H.-J. & LXUGER, P. (1991).Charge translocation by the Na+-K+ pump: II. Ion binding andrelease at the extracellular face. Journal of Membrane Biology 121,163-176.
VASILETS, L. A., OHTA, T., NOGUCHI, S., KAwAMURA, M. & SCHWARZ,W. (1993). Voltage-dependent inhibition of the sodium pump byexternal sodium: Species differences and possible role of the N-terminus of the a-subunit. European Biophysics Journal 21,433-443.
VASILETS, L. A., OMAY, H. S., OHTA, T., NoGUCHI, S., KAwAMURA, M.& SCHWARZ, W. (1991). Stimulation of the Na+/K+ pump byexternal [K+] is regulated by voltage-dependent gating. Journal ofBiological Chemistry 266, 16285-16288.
WANG, X. & HORISBERGER, J.-D. (1995). A conformation of the Na,K-pump is permeable to proton. American Journal of Physiology 268,C590-595.
J Physiol. 491.3 Role of N-terminus of Na+-K+ pump a-subunit

594 X. Wang, F Jaisser and J-D. Horisberger J Physiol.491.3
WIERZBICKI, W. & BLOSTEIN, R. (1993). The amino-terminal segmentof the catalytic subunit of kidney Na,K-ATPase regulates thepotassium deocclusion pathway of the reaction cycle. Proceedings ofthe National Academy of Sciences of the USA 90, 70-74.
YOON, K. L. & GUIDOTTI, G. (1994). Studies on the membranetopology of the (Na,K)-ATPase. Journal of Biological Chemistry269, 28249-28258.
AcknowledgementsWe are grateful to Laurent Schild for his useful comments andsuggestions. This work was supported by the Swiss Fonds Nationalpour la Recherche Scientifique, grant 31-36138.92 to J.-D.H.
Received 31 M/fay 1995; accepted 6 October 1995.


















