
Radiazioni ionizzanti a basso ed alto LET: valutazione...
31
Gianluca Ingrosso U.O.C. Radioterapia Policlinico “Tor Vergata”, Roma Radiazioni ionizzanti a basso ed alto LET: valutazione epigenetica e bioumorale del danno
Transcript of Radiazioni ionizzanti a basso ed alto LET: valutazione...

Gianluca Ingrosso U.O.C. Radioterapia Policlinico “Tor Vergata”, Roma
Radiazioni ionizzanti a basso ed alto LET: valutazione epigenetica e bioumorale del danno

Using the PubMed portal with the following search strategy ‘radiation’ AND (‘bystander’ OR ‘genomic instability’) 705 references were identified.
Munro. J Radiol. Prot. 2009
Bystander effect: any biological effect following irradiation that is expressed by a cell whose nucleus has not been directly exposed to ionising radiation.
Clastogenic factor: a substance present in serum or plasma that causes damage to the chromosomes of cells that are exposed to its influence.
Genomic instability: is found when a cell population, surviving after radiation, shows an increased rate of acquisition of delayed changes leading to chromosomal aberrations, gene mutations and delayed cell death in its progeny.
radiobiology

DNA damage that is misrepaired lead to mutations, increasing the probability of transformation and carcinogenesis.
Cells have multiple and complex processes for sensing and repairing changes to their genome to enable future propagation and stability.
Irradiated cells send signals to their neighbours.
bystander

Irradiated cells send signals to their neighbours.
GJ are multimeric protein channels that allow molecules to pass through (Ca2+, nucleotides, peptides)
Many aspects of bystander- mediated signalling and response have close parallels to inflammatory respones.
The modulation of inflammatory response could reduce bystander effect protecting normal tissue from additional damage during RT.
bystander

The modulation of inflammatory response could reduce bystander effect protecting normal tissue from additional damage during RT.
1. Rats irradiated in a region of the lung had micronucleus formation in other non irradiated areas of the lung. Pre-injection of animal with Cu-Zn superoxide dismutase (SOD) or the NOS inihbitor L-NAME led to a reduced response in the shielded area.
2. The partial lung irradiation in rats is accompanied by macrophage activation and production of cytokines (IL-1α, IL-1β, IL-6, TNF-α and TGF-β) in-field and out-field, lasting up to 16 weeks after irradiation.
3. In mouse models activated macrophages and T cells activate 2 bystander effect mechanisms. a. Activated immune cells secrete cytokines leading to NO production. b. Activated macrophages produce NO.
1. Khan. IJROBP. 1998 2. Calveley. IJROBP. 2005 3. De Ridder. IJRBBP. 2008
bystander

Mice irradiation (1 Gy X-rays) of one side of the body led to epigenetic changes in the shielded non-exposed side 1 cm away. It was observed a change in the DNA methyltransferases activity1.
bystander
Chromosome aberration in unirradiated cells induced by plasma from rats exposed to whole-body irradiation2, atomic bomb survivors from Hiroshima3, and salvage personel exposed during cleanup after Chernobyl4.
1. Koturbash. Oncogene. 2006 2. Faguet. Cancer Genet. Cyotgenet. 1984 3. Pant. J. Med. Sci. 1977 4. Emerit. J. Cancer Res. Clin. Oncol. 1994
Extracellular IR-induced DNA-damage factors that persist for prolonged periods after the exposure to IR.
A chromosome breakage factor present in plasma of irradiated individuals that could mimic the direct effect of X-rays on chromosome.
Death inducing effect: chinese hamster-human hybrid GM10115 cells after IR exposure demonstrate genomic instability showing dynamic and persistent generation of chromosomal rearrangements multiple generations after the original insult. The genomic instability is thought to be related to secreting factors into the culture medium. The medium of unstable cells transferred into culture of unirradiated cells leads to cell death5.
clastogenic
5. Nagar. Cancer Res. 2003

Radiation-induced genomic instability (RIGI) is an early event in radiation carcinogenesis and the mechanisms underlying this process are poorly understood. RIGI is observed in cells that survive irradiation at delayed times after exposure and manifest in their progeny over many generations.
To understand the initiation and perpetuation of RIGI studies have evaluated several end-points (DSBs, mutations, gene expression, cell cycle arrest, apoptosis). Recent studies have shown radiation-induced epigenetics changes including DNA methylation and mi-RNA expression.
Epigenetic changes could be the missing link between radiation-exposure, RIGI and radiation-induced carcinogenesis.

Waddington (the strategy of the genes. Allen-Unwin, London, 1957)
Epigenetic deals with how genotypes give rise to phenotypes during development.
Epigenetic is the structural adaptation of chromosomal regions so as to register, signal or perpetuate altered activity states. Epigenetic system would not, under normal circumstances, initiate a change of state at a particular locus but would register a change already imposed by others events. Such event could be the collision of DNA with ionizing radiations or a developmental switch in gene expression.
Epigentetics refers to heritable changes in gene expression that take place without alterations in the DNA sequence
• DNA methylation
• Chromatin remodelling
• Mi-RNA regulated gene expression

DNA methylation
de novo methylation of unmethylated CpG sites in embryonic stem cells and in cancer cells
Mammalian DNA is predominantly methylated at
cytosine bases in cytosine-phosphate-guanine (CpG)
-dinucleotides through the enzymatic activity of
DNA methyltransferases (DNMT).
Studies analysing genome-wide methylation levels
have shown non-CpG methyl. in somatic cells.
Mammals have 4 DNA-methyl-transferase (DNMT).
DNMT1: functions as the manteinance methyltransferase by coping the methyl pattern of
The old DNA strand onto the newly synthesised strand during DNA replication.
DNMT2: is capable of tRNA methyl. in addition to weak DNA metyltransferase activity.
DNMT3a
DNMT3b

In normal cells DNA methyl. is involved in the control of gene expression, especially in terms of tissue-specificity and germ-line gene expression.
Promoter regions of tumor-suppressor genes are hypomethyl. to allow their expression and maintain the normal state of the cell.
Genomic imprinting hypermethyl. ensures suppression of one of the 2 parental alleles of a gene for monoallelic axpression.
Repression of certain repeat elements through hypermethyl. prevent chromosom. instability, translocation, gene disruption.
Cells that have mutations in DNMT lack the stabilizing effect of DNA methyl. and have prominent nuclear abnormalities
DNA methylation

1. C57/BI mice whole-body irradiated with 0.5, 1, 2.5, 5 Gy of X-rays. Methylation analysis was performed in liver, spleen and lung tissues. Dose-dependent DNA hypomethyl. correlated with decreased expression of DNMT1 and DNMT3a/b. This DNA hypomethyl. was associated with increased levels of DNA strand-breaks.
1. Pogribny, Kovalchuk. Bioch Biophys Res Commun. 2004 2. Pogribny, Kovalchuk. Molec. Cancer Res. 2005 3. Kovalchuk, Pogribny. Mutation Res. 2004
2. Mice exposed to fractionated whole-body X-ray irradiation (0.5 Gy). DNA hypomethyl. correlated with decreased expression of DNMT1 and DNMT3a/b, methyl CpG binding protein 2 (MeCP2) and methyl CpG binding domain protein 2 (MBD2). This DNA hypomethyl. was associated with increased levels of DNA strand-breaks.
3. Mice exposed to acute and cronic low-dose whole-body X-ray irradiation. The promoter of the tumor-suppressor p16 was hypermethyl. Chronic low-dose radiation exposure is a more potent inducer of epigenetic effects than the acute exposure.
Chronic exposure leads to greater genome destabilization than acute exposure.
DNA methylation

4. HPV-G cells (human keratinocytes) irradiated with 60Co manifested genomic instability at 20 passages post-irradiation: reproductive cell death, reduced cloning efficency, increased apoptosis, increased proportion of chromosomally aberrant cells. This RIGI was associated with DNA hypermetylation of CpG dinulceotides and peri- centromeric spermidine N1-acetyltransferase 2 (SAT2) satellite sequence.
4. Kaup. Mutation Res. 2006 5. Gronbaek. Acta Patologica 2007 6. Franco. Cancer Letters. 2008
6. ROS production post-irradiation affect changes in DNA methylation that can lead to RIGI. Free radicals scavengers reduce the incidence of instability after irradiation.
5. Aberrant methylation status of repeat elements and of SAT2 satellite sequence affect genomic instability.
DNA methylation
hypomethylation of repeat elements and promoter regions of oncogenes and
hypermethylation of promoter regions of tumor-suppressor genes post-irradiation play an important role in RIGI and radiation-induced carcinogenesis

Chromatin remodelling
A histone octamer consists of 2 copies of each of the four core histone proteins. The octamer assembles when a tetramer, containing two copies of both H3 and H4, complexes with two H2A/H2B dimers. These histones of the histone octamer all contain N-terminal tails.

Chromatin remodelling is involved
in gene expression, DNA replication
and repair, chromosome
condensation, segregation and apoptosis. Gene transcription is controlled by histone
modifications in chromatin.
Open chromatin is associated with transcriptional activity and is characterised by unmethyl.
cytosines, acetylated histone tails by histone acetyltransferases (HAT) and trimethylation of
lysine 4 on histone H3.
Closed chromatin occurs when cytosines become methyl., methyl-binding domains bind to
methyl. cytosines and acetyl groups are removed from the histone deacetylases (HDAC). DNA
becomes coiled into a closed chromatin structure carryng the transcriptional silencing mark
histone H3 lysine 9 trimethylation.
Chromatin remodelling

Chromatin remodelling
Falk. Biochim et Biophys. Acta. 2007
Cells exposed to gamma-rays
15 min after irradiation:
decreased intensity of chromatin labeling increased histone H4 lysine 5 acetylation decreased histone H3 lysine 9 dimethylation
40 min after irradiation:
decreased acetylation of histone H4 lysine 5 increased methylation of histone H3 lysine 9
Chromatin decondensation
Chromatin condensation
DSBs
The need to convert from less to more condensed chromatin after DNA repair

Chromatin remodelling
Park. EMBO J 2006
Inactivation of the SWI/SNF complexes or downregulation of the catalytic core subunits of the complexes both result in inefficient DNA double-strand break (DSB) repair.
DSB-activated ATM initiates S139ph (1) and establishes low levels of g-H2AX on the chromatin around a DSB. g-H2AX triggers the acetylation (2) of H3 within the same and/or neighbouring nucleosomes by recruiting HATs such as Gcn5 to these nucleosomes. SWI/ SNF then binds to g-H2AX nucleosomes by interacting with the acetylated H3 using the BRD of BRG1 (3). Recruited SWI/SNF in turn facilitates ATM-mediated S139ph (4) probably by increasing the accessibility of nucleosomes around the DSB. The increased S139ph leads to further acetylation of H3 and more accumulation of SWI/SNF at g-H2AX nucleosomes. This positive feedback loop among SWI/SNF, S139ph and H3 acetylation eventually establishes the high levels of g-H2AX for efficient DSB response.
1
2
3
4

Chromatin remodelling
Phosphorylation of histone H2AX
is required for phosphorylation
of 53BP1 by ATM and localization
of 53BP1 into nuclear foci.
In turn, 53BP1 function is important
for coupling ATM to several of its
downstream targets, including p53 and
SMC1.
Gamma-H2AX has been used as a measure of DSBs post-irradiation.

DNA damage induce phosphorylation of the KRAB-ZFP–associated protein 1 (KAP1), the dedicated corepressor for Krüppel-associated box (KRAB) zinc finger protein (ZFP) proteins. KAP1 is phosphorylated following DNA damage by the phosphatidylinositol-3kinase–like family of kinases in a ATM-dependent manner.
Phosphorilated-KAP-1 leads to euchromatinisation that is a fundamental for DNA damage-response.
Pro-apoptotic
Ziv. Nature Cell Biol. 2006
Chromatin remodelling
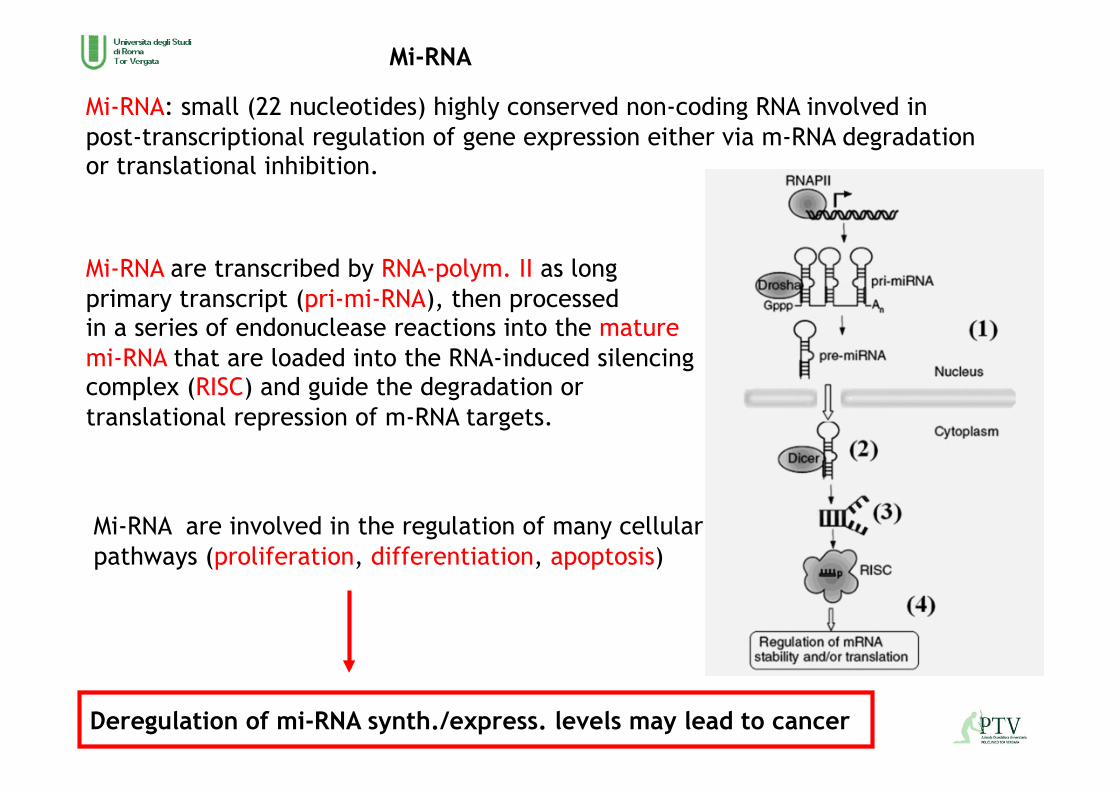
Mi-RNA: small (22 nucleotides) highly conserved non-coding RNA involved in post-transcriptional regulation of gene expression either via m-RNA degradation or translational inhibition.
Mi-RNA are transcribed by RNA-polym. II as long primary transcript (pri-mi-RNA), then processed in a series of endonuclease reactions into the mature mi-RNA that are loaded into the RNA-induced silencing complex (RISC) and guide the degradation or translational repression of m-RNA targets.
Deregulation of mi-RNA synth./express. levels may lead to cancer
Mi-RNA are involved in the regulation of many cellular pathways (proliferation, differentiation, apoptosis)
Mi-RNA

Mi-RNA
1. Exposure of human fibroblast to low-LET x-rays (2 Gy) 0.5 h: down-regulation in mi-RNA levels. 2 h: mi-RNA up-regulation of mi-RNA 579, 608, 548 and 585 that target genes involved in radioresponsive mechanisms, such as cell cycle checkpoints and apoptosis
1. Maes. Molec and Cellular Biol. 2008 2. Cha. Oncology Reports. 2009 3. Cha. Int J of Oncol. 2009
mi-RNA are down-regulated immediately after irradiation to stimulate DNA repair mechanisms and later are up-regulated to suppress apoptosis for cell survival.
2. low-dose of irradiated cells (0.05 Gy): decrease in mi-RNA 20 and 21 that are involved in carcinogenesis as onco-miRNA 3. high of irradiated cells (10 Gy): increase in mi-RNA 197 that is associated with carcinogenesis as onco-miRNA
IR alters mi-RNA profiles in a way that affect: oxidative stress DNA damage and reapir cell cycle regulation apoptosis DNA methylation
RIGI is thought to be linked to inflammatory processes in the radiation response

IR
DNMT1
DNMT3a/3b
MeCP2
MBD2
Hypomethyl.
Chromosomal instability
DNA methylation
Hypermethyl.
Silencing tumor suppressor genes (p16), SAT2 satellite sequences
Chromatin remodelling
H4K trimethyl.
H4K5 acetyl.
H3K9 dimethyl.
Chromatin decondensation
Mi-RNA expression
Mi-RNA
Altered gene expression
DNA methyl. Oxdative stress DNA damage Apoptosis tumorigenesis
Radiation-induced epigenetic changes

inflammation
bystander effect
clastogenic factors
epigenetic changes
genomic insability
ionizing radiation
radiation-induced damage
carcinogenesis

Biomarker: measurable characteristic of a biological system that is indicative of normal function or disease state of the system or its response to an external factor such as a therapeutic intervention.
The study of biomarkers in normal-tissue radiobiology is in its infancy
2 classes of biomarkers:
• Predictive factors: biological or clinical factors (but not treatment-related) observable at baseline, that are statistically associated with the probability of a given outcome of a specific treatment in an individual.
• Response markers: therapy-related changes in biomarkers that are mechani- stically related to treatment outcome at the individual-patient level.
Bentzen. IJROBP 2010
radiation-induced damage

Predictive marker
Predictive markers are aimed at selecting cases for a specific type of therapy or a changing radiotherapy dose-fractionation prescription or a planned dose distribution.
Cytokines and growth factors
1. TGF-beta1: profibrotic cytokine activated by IR strong correlation between the levels of TGF in pre-radiotherapy plasma samples and subsequent developlment of radiation fibrosis in 91 early-stage breast cancer patients.
2. IL-1alfa and IL-6: baseline plasma levels of IL-1alfa and IL-6 are correlated with symptomatic radiation pneumonitis.
1. Li. Int. J Cancer 1999 2. Chen. IJROBP 2005

Predictive marker
Radiogenomics: the study of the possible link between genotypic variation and radiation therapy toxicity.
The aim is to develop tools for biological individualized radiotherapy, taking into consideration the individual risk of radiation-induced morbidity. Single nucleotide polymorphisms (SNPs) represent substitutions in wich an alternate base pair is present at a particular nucleotide location. SNPs can affect protein function altering amino acid composition or affecting the transcriptional or translational control. In human genome SNPs occur on average every 300 bp; with an average gene lenght of 27.000 bp, about 100 SNPs are in a typical gene.
Inter-patient heterogeneity in normal tissue reactions after RT is probably due to genetically determined intrinsic differences in the cellular response to damage produced by IR and in particular the repair of DNA DSBs.
Genome-wide association studies: dense sets of SNPs are genotyped to identified genetic variations associated with a certain phenotype.
The candidate gene approach: analysis restricted to known polymorphisms

Predictive marker
The candidate gene approach: analysis restricted to known polymorphisms
Identify mutations and SNPs in candidate radiosensitivity genes and screen for abnormalities in expression of candidate proteins.
SNPs in genes involved in
ATM: encodes a protein that plays a key role in detection and repair of DNA DSBs. The signaling cascade (activation of p53, Chk2, Mdm2, BRCA1, SMC1, Nbs1, FancD2) results in activation of cell cycle ceckpoints, DNA repair or apoptotis.
XRCC1: encodes for a DNA repair protein required for the repair on DNA damage caused by IR, oxidatve stress and DNA methylating agents.
SOD2: encoded for a manganese superoxide dismutase important for oxidative damage.
DNA damage detection DNA damage repair Pro-fibrotic and inflamm. cytokines Endogenous anti-oxidant enzymes

Response marker
Response markers could serve as individualized in vivo dosimeters and they could form the basis for the biological adaptive radiotherapy
1. The normalization of TGF-beta1 level at the end of radiotherapy yelded a positive predictive value (PPV) of 90% for identifying patients who did not develop pneumonitis.
2. Baseline and weekly measure of TGF-beta1 were not associated with RILT (radiation-induced lung toxicity), whereas the ratio between TGF-beta1 level During and before RT showed a significant association with subsequent RILT.
1. Anscher. IJROBP. 1998 2. Zhao. IJROBP. 2009
EVALUATE: • RT-technique • Dose distribution • Toxicity scoring

35 pazienti consecutivi affeti da adenocarcinoma della prostata
Età media: 71 y mediana: 72 range: 46-78
Stadio: T1a 1 pt T1c 22 pts T2a 8 pts T2c 3 pts T3b 1 pt
Gleason score: 5(3+2) 1 pt 6(3+3) 18 pts 7(3+4) 5 pts 7(4+3) 4 pts 8(4+4) 3 pts 8(3+5) 1 pt 9(4+5) 1 pt 10(5+5) 1 pt 1 pt non determinato il Gleason dall’anatomo-patologo
PSA Mediano 7,9 ng/ml Medio 11,57 ng/m Range 0,53-72,3 ng/ml
Response marker

Radioterapia conformazionale Volume bersaglio: prostata e vescichette seminali DTF 66 Gy su prostata e vescichette DTF 76 Gy su prostata In 10/35 pazienti è stata associata terapia con analogo LH-RH. Prelievi ematici per lo stoccaggio del siero 1° prima dell’inizio della radioterapia 2° a metà trattamento radiante (DTF 38 Gy) 3° a fine radioterapia (DTF 76 Gy) 4° 2 mesi dopo la fine della radioterapia
Molecole purificate di Collagene I, III, IV, V, di Fibronectina, Laminina Sigma(Missouri, USA) e Chemicon (CA, USA).
ELISA del siero 1° e 4° dei 35 pazienti per valutare la presenza di IgG contro epitopi conformazionali dei componenti della matrice extracellulare (MEC). Sono stati utilizzati 40 sieri di controllo da donatori del Centro Trasfusionale dell’Università di Roma “Sapienza”
Analisi statistica: Test di Fisher per valutare la significatività nei due gruppi [di studio e di controllo] per la presenza di auto-Ab contro epitopi conformazionali delle molecole della MEC. T di Student per valutare le differenze nei titoli degli auto-Ab
Response marker

Dopo radioterapia espressione di auto-Ab verso componenti della matrice: 2/35 pts verso Collagene III 4/35 pts verso Collagene IV 2/35 pts verso Collagene I, IV, Laminina, Fibronectina 1/35 pts verso Collagene I, IV, V, Laminina 1/35 pts verso Collagene I, IV e Laminina 2/35 pts verso Collagene I 1/35 pts verso Laminina 1/35 pts verso Collagene IV
Differenza significativa tra pts irradiati e controllo per: Collagene I (p=0,0045) Collagene IV (p=0,043)
Titolo medio degli autoanticorpi prima e dopo RT: Collagene I: 758 ± 196 / 1233 ± 1246 Collagene III: 600 ± 565 / 850 ± 70 Collagene IV: 862 ± 718 / 2225 ± 1573 Collagene V: 1066 ± 375 / 1183 ± 448 Fibronectina: 1300 ± 700 / 575 ± 125 Laminina: 640 ± 476 / 1180 ± 1175
Risultato La radioterapia nei pazienti affetti da adenoca della prostata condiziona la risposta immunologica verso le molecole della matrice extracellulare
Response marker

Dose distribution and biomarkers are mutually confounding factors
Standardized scoring of the grade of reaction
Adequate follow-up time
Prospective storing of the full 3D dose matrix
Development of data analysis strategies to allow correction for the confounding effects of dose distribution as well as patient and treatment-related risk factors
Findings reported in the literature are often inconsistent across studies
Most studies have modest sample sizes


















