
Quantitative Study Antibiotic-Induced Susceptibility ...aac.asm.org/content/34/7/1348.full.pdf ·...
6
Vol. 34, No. 7 ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, July 1990, p. 1348-1353 0066-4804/90/071348-06$02.00/0 Copyright © 1990, American Society for Microbiology Quantitative Study of Antibiotic-Induced Susceptibility to Clostridium difficile Enterocecitis in Hamsters H. E. LARSON* AND S. P. BORRIELLO Microbial Pathogenicity Research Group, Division of Communicable Diseases, Clinical Research Centre, Watford Road, Harrow, Middlesex, England HAI 3UJ Received 27 September 1989/Accepted 13 April 1990 Commonly used antibiotics were compared for their ability to induce Clostridium difficile enterocecitis and death in hamsters. Susceptibility to infection with C. difficile was measured by calculating 50% lethal doses (in CFU) for hamsters for various intervals after antibiotic treatment. Infection occurred after very small doses of C. difficile were given to hamsters treated with clindamycin, ampicillin, flucloxacillin, and cefuroxime; there was little difference between the antibiotics in the degree of susceptibility that they induced. A large difference in the duration of susceptibility was observed, however, with susceptibility being temporary following ampicillin, flucloxacillin, and cefuroxime administration but long-lived following clindamycin administration. A larger dose of ampicillin, multiple doses of ampicillin, and a combination of antibiotics had comparatively small effects on the duration of susceptibility. C. dfficile growth and toxin production in in vitro suspensions of cecal contents were found to correlate closely with in vivo hamster infectivity. A persisting loss of colonization resistance following antibiotic treatment may be a type of postantibiotic effect. Although these results cannot be applied directly to humans, they suggest lines of further investigation into how antibiotics may differ in producing risks of C. difficile infection and pseudomembranous colitis in patients. Prior to the recognition that pseudomembranous colitis (PMC) was caused by Clostridium difficile, it was known to be a risk of antibiotic treatment, especially lincomycin and clindamycin (11). Discovery of the microbial pathogenesis of PMC (18) has permitted a more careful assessment of the relationship between antibiotic treatment and C. difficile infection (17). A large number of antibiotics have been found to facilitate infection in both animals and humans (2, 10). There has been suspicion that some antibiotics are more likely to result in C. difficile-associated disease than others. Aronsson et al. (1) compared the relative risk of antibiotics by determining the number of C. difficile toxin-positive cases relative to antibiotic sales in Sweden. In contrast, Pierce et al. (19) failed to find differences in the rates of antibiotic- associated colitis after the administration of several antibi- otics, although the power of that study to detect significant differences likely was small. Quantitative risk estimations in humans are bedevilled by problems of sample size, uncer- tainties about how much and how often antibiotics are actually consumed, differences in routes of administration, use of combination antibiotic therapies, and ignorance about whether individuals have actually been exposed to C. diff- cile in their environments. We undertook an experimental quantitative comparison of various antibiotics using both in vivo and in vitro methods. It is known that oral vancomycin treatment makes hamsters susceptible to infection with very small doses of C. difficile (18). We determined the 50% lethal dose (LD50) for hamsters of C. difficile spores after administration of single oral doses of various antibiotics and assessed the capacity of suspen- sions of hamster cecal contents to support growth of C. difficile in vitro. * Corresponding author. MATERIALS AND METHODS C. difficile. Two strains (B-1 and H-1) of C. difficile isolated from patients with PMC and known to produce enterocecitis when administered to clindamycin-treated hamsters (17) were used in this study. Both strains produce toxins A and B. Antibiotic MICs were determined by a microdilution technique. For strain B-1 the MICs were as follows: clinda- mycin, 256 ,ug/ml; ampicillin, 0.5 ,ug/ml; flucloxacillin, 4 ,ug/ml; and cefuroxime, 128 ,ug/ml. Strain H-1 was inhibited by 0.13 ,ug of clindamycin per ml and 0.5 ,ug of ampicillin per ml. For inoculation of hamsters, a spore preparation was made from a 5-day Robertson cooked meat medium culture by using alcohol shock. The culture broth was centrifuged at 2,000 x g for 10 min, suspended in 5 ml of brain heart infusion (Difco Laboratories, Detroit, Mich.) with cysteine hydrochloride (BHIC; 0.5% [wt/vol]), mixed with 5 ml of 100% ethanol, held for 1 h at ambient temperature, and centrifuged again as described above. The pellet was washed once in BHIC and suspended in 5 ml of BHIC, and serial 10-fold dilutions in BHIC were prepared. The number of viable spores in the inocula was estimated by seeding 0.5 ml of diluted suspension on a blood agar plate (Difco), incubat- ing it anaerobically at 37°C for 48 h in an anaerobic cabinet (Forma Scientific, Marietta, Ohio), and recording the num- ber of colonies. Animals. Syrian hamsters obtained from breeding colonies at the National Institute for Medical Research, London, England, were individually maintained in sterilized filter- lidded polycarbonate boxes (Stephen Clark Fabrications Ltd., Alva, Scotland) and were given autoclaved bedding, feed, and tap water (17). Wire cage floors were used to prevent coprophagia in one experiment. Antibiotic solutions and suspensions of C. difficile were administered orally in 0.25-ml volumes from the end of a 1-ml syringe. Antibiotics, as supplied for clinical use, were diluted in distilled water to yield 3 mg in 0.25 ml. All manipulations of antibiotic-treated animals were carried out with attention given to the preven- 1348 on July 5, 2018 by guest http://aac.asm.org/ Downloaded from
Transcript of Quantitative Study Antibiotic-Induced Susceptibility ...aac.asm.org/content/34/7/1348.full.pdf ·...

Vol. 34, No. 7ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, July 1990, p. 1348-13530066-4804/90/071348-06$02.00/0Copyright © 1990, American Society for Microbiology
Quantitative Study of Antibiotic-Induced Susceptibility toClostridium difficile Enterocecitis in Hamsters
H. E. LARSON* AND S. P. BORRIELLOMicrobial Pathogenicity Research Group, Division of Communicable Diseases, Clinical Research Centre,
Watford Road, Harrow, Middlesex, England HAI 3UJ
Received 27 September 1989/Accepted 13 April 1990
Commonly used antibiotics were compared for their ability to induce Clostridium difficile enterocecitis anddeath in hamsters. Susceptibility to infection with C. difficile was measured by calculating 50% lethal doses (inCFU) for hamsters for various intervals after antibiotic treatment. Infection occurred after very small doses ofC. difficile were given to hamsters treated with clindamycin, ampicillin, flucloxacillin, and cefuroxime; therewas little difference between the antibiotics in the degree of susceptibility that they induced. A large differencein the duration of susceptibility was observed, however, with susceptibility being temporary followingampicillin, flucloxacillin, and cefuroxime administration but long-lived following clindamycin administration.A larger dose of ampicillin, multiple doses of ampicillin, and a combination of antibiotics had comparativelysmall effects on the duration of susceptibility. C. dfficile growth and toxin production in in vitro suspensionsof cecal contents were found to correlate closely with in vivo hamster infectivity. A persisting loss of colonizationresistance following antibiotic treatment may be a type of postantibiotic effect. Although these results cannotbe applied directly to humans, they suggest lines of further investigation into how antibiotics may differ inproducing risks of C. difficile infection and pseudomembranous colitis in patients.
Prior to the recognition that pseudomembranous colitis(PMC) was caused by Clostridium difficile, it was known tobe a risk of antibiotic treatment, especially lincomycin andclindamycin (11). Discovery of the microbial pathogenesis ofPMC (18) has permitted a more careful assessment of therelationship between antibiotic treatment and C. difficileinfection (17). A large number of antibiotics have been foundto facilitate infection in both animals and humans (2, 10).There has been suspicion that some antibiotics are more
likely to result in C. difficile-associated disease than others.Aronsson et al. (1) compared the relative risk of antibioticsby determining the number of C. difficile toxin-positive casesrelative to antibiotic sales in Sweden. In contrast, Pierce etal. (19) failed to find differences in the rates of antibiotic-associated colitis after the administration of several antibi-otics, although the power of that study to detect significantdifferences likely was small. Quantitative risk estimations inhumans are bedevilled by problems of sample size, uncer-tainties about how much and how often antibiotics areactually consumed, differences in routes of administration,use of combination antibiotic therapies, and ignorance aboutwhether individuals have actually been exposed to C. diff-cile in their environments.We undertook an experimental quantitative comparison of
various antibiotics using both in vivo and in vitro methods. Itis known that oral vancomycin treatment makes hamsterssusceptible to infection with very small doses of C. difficile(18). We determined the 50% lethal dose (LD50) for hamstersof C. difficile spores after administration of single oral dosesof various antibiotics and assessed the capacity of suspen-sions of hamster cecal contents to support growth of C.difficile in vitro.
* Corresponding author.
MATERIALS AND METHODS
C. difficile. Two strains (B-1 and H-1) of C. difficile isolatedfrom patients with PMC and known to produce enterocecitiswhen administered to clindamycin-treated hamsters (17)were used in this study. Both strains produce toxins A andB. Antibiotic MICs were determined by a microdilutiontechnique. For strain B-1 the MICs were as follows: clinda-mycin, 256 ,ug/ml; ampicillin, 0.5 ,ug/ml; flucloxacillin, 4,ug/ml; and cefuroxime, 128 ,ug/ml. Strain H-1 was inhibitedby 0.13 ,ug of clindamycin per ml and 0.5 ,ug of ampicillin perml. For inoculation of hamsters, a spore preparation wasmade from a 5-day Robertson cooked meat medium cultureby using alcohol shock. The culture broth was centrifuged at2,000 x g for 10 min, suspended in 5 ml of brain heartinfusion (Difco Laboratories, Detroit, Mich.) with cysteinehydrochloride (BHIC; 0.5% [wt/vol]), mixed with 5 ml of100% ethanol, held for 1 h at ambient temperature, andcentrifuged again as described above. The pellet was washedonce in BHIC and suspended in 5 ml of BHIC, and serial10-fold dilutions in BHIC were prepared. The number ofviable spores in the inocula was estimated by seeding 0.5 mlof diluted suspension on a blood agar plate (Difco), incubat-ing it anaerobically at 37°C for 48 h in an anaerobic cabinet(Forma Scientific, Marietta, Ohio), and recording the num-ber of colonies.
Animals. Syrian hamsters obtained from breeding coloniesat the National Institute for Medical Research, London,England, were individually maintained in sterilized filter-lidded polycarbonate boxes (Stephen Clark FabricationsLtd., Alva, Scotland) and were given autoclaved bedding,feed, and tap water (17). Wire cage floors were used toprevent coprophagia in one experiment. Antibiotic solutionsand suspensions of C. difficile were administered orally in0.25-ml volumes from the end of a 1-ml syringe. Antibiotics,as supplied for clinical use, were diluted in distilled water toyield 3 mg in 0.25 ml. All manipulations of antibiotic-treatedanimals were carried out with attention given to the preven-
1348
on July 5, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

QUANTITATIVE STUDY OF SUSCEPTIBILITY TO C. DIFFICILE
tion of cross-contamination of infection from the environ-ment of the animal holding unit. Control antibiotic-treatedhamsters were inoculated with sterile broth after the inocu-lations of C. difficile were completed. Observations were
continued for 7 days, at which time surviving animals were
sacrificed. All animals were examined for evidence of ceci-tis.
Infectivity calculation. Susceptibility of hamsters to C.difficile enterocecitis was measured by calculating the num-
ber of CFU required to produce enterocecitis in half of theanimals inoculated with the highest dilution that produceddisease. First, the LD50 was determined by challenginghamsters with log1o dilutions of a spore preparation afterthey were given a single oral dose of antibiotic. Threehamsters were challenged per dilution and observed as
described above. The number of hamsters with enterocecitisat each dilution was recorded, and the Karber method ofendpoint calculation was used to yield the LD50 (12). Sec-ond, the CFU in each dilution of the spore suspension was
estimated as described above. The CFU, as determined bythe estimated number of C. difficile at the endpoint dilution,was divided by the LD50, and this value was rounded to givethe nearest whole CFU per 1 LD50. To conserve animals butstill give a complete picture of the duration of susceptibilityafter various experimental variables, some challenges wereperformed with only a single large dose (104 CFU) ofspores. Intervals after antibiotic treatment were selected forchallenge that defined the duration of susceptibility, indays.
In vitro suspensions. Suspensions of cecal contents for invitro studies were prepared from freshly sacrificed hamsterswho had received antibiotics but not C. difficile, as describedpreviously (6). Briefly, cecal contents were removed fromeach of two hamsters for each day following antibiotictreatment, suspended to 10%o (wt/vol) in prereduced distilledwater, and placed in bijoux in duplicate aliquots of 5.0 mleach. Logarithmic-growth-phase C. difficile was prepared byinoculating an overnight culture into fresh BHIC and incu-bating it anaerobically for 6 h at 37°C. This was centrifuged,washed in 20 ml of fresh broth, centrifuged, and suspendedin 10 ml of fresh broth. The suspension was diluted 1:10, and50 p.l was inoculated into each suspension of cecal contents.This inoculum typically contained 5 x 106 CFU. Suspen-sions of cecal contents were then incubated at 37°C anaero-
bically and examined for C. difficile growth at 48 h by a
micromethod as described elsewhere (6). To test for toxinproduction, suspensions of cecal contents were centrifugedand supernatants were serially diluted twofold. One hundredmicroliters of each dilution was added to confluent mono-
layers of Vero cells in microdilution plates and observed at24 and 48 h for evidence of a C. difficile toxin B cytopathiceffect.To test for the possible presence of residual antibiotic,
suspensions of cecal contents were centrifuged at 12,900 x gfor 30 min, and supernatants were filtered through 0.45-,um-pore-size filters. Filtrates were inoculated, incubated,and tested for growth of C. difficile and toxin production inthe same manner as described previously for suspensions ofcecal contents (6). Some suspensions of cecal contents wereassayed for clindamycin by the method of Roberts et al. (20),for ampicillin by using Sarcina lutea as the test organism,and for P-lactamase by the method of Rolfe and Finegold(21).
TABLE 1. Calculation of LD50 of C. difficile B-1 2 daysafter a single 3-mg dose of clindamycin
Dilution of C. difficile No. with enterocecitis/ No. of CFU/spore suspension no. inoculated 0.5 mlinoculated
10-2 3/3 251lo-3 3/3 41lo-4 3/3 510-5 1/3 110-6 0/3 010-7 0/3 0Broth only 0/2
RESULTS
Hamsters given antibiotics followed by broth and thosehamsters in the holding stocks that did not receive antibiot-ics remained well.
Table 1 records the results of a sample experiment used todetermine susceptibility to C. difficile B-1 2 days after asingle oral, 3-mg dose of clindamycin. The LD50 per 0.25 mlof undiluted challenge inoculum was 4.8 (log1o). There was4.7 CFU (log1o) in 0.25 ml of undiluted challenge inoculum.This yielded 0.8, or approximately 1, CFU/LD50.
Table 2 summarizes the results of a series of similarexperiments conducted at various intervals between antibi-otic treatment and challenge with C. difficile. For up to 2days after treatment, less than 10 CFU was needed toproduce enterocecitis and death for all antibiotics listed inTable 2. After clindamycin treatment, hamsters remainedsusceptible to small doses of C. difficile for at least 14 days;these animals were maintained on wire cage floors. Incontrast, animals treated with ampicillin and cefuroxime didnot remain susceptible and were resistant to large doses ofC. difficile by 3 days after antibiotic treatment. After fluclox-acillin treatment, hamsters were still susceptible at 3 days. Inone experiment, suspensions of C. difficile were adminis-tered immediately following the administration of ampicillin(Table 2, 0 day interval). This produced susceptibility to 6CFU of C. difficile.
Since it appeared that antibiotics were distinguished bythe duration rather than by the degree of susceptibility theyproduced, further experiments were performed by challeng-ing the hamsters with a single large inoculation of C. difficilespores. Table 3 summarizes experiments which further com-pared various antibiotic regimens by challenging the ham-sters with 104 spores at various intervals after antibiotictreatment was completed. Clindamycin-treated animals werestill susceptible 74 days after a single 3-mg dose; thesehamsters were not maintained on wire cage floors. Anampicillin dose of 0.3 mg did not produce susceptibility,whereas a dose of 3 mg produced susceptibility that lasted 2days with evidence of recovery of resistance by 3 days. A
TABLE 2. LD50 of C. difficile B-1 at various intervalsafter a single 3-mg dose of antibiotic
LD50 (CFU) at the followingAntibiotic intervals (days)
0 1 2 3 5 14
Clindamycin 1 2Ampicillin 6 8 >1 x 103 >2 x 103Flucloxacillin 5 6Cefuroxime 7 >1 x 104
VOL. 34, 1990 1349
on July 5, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
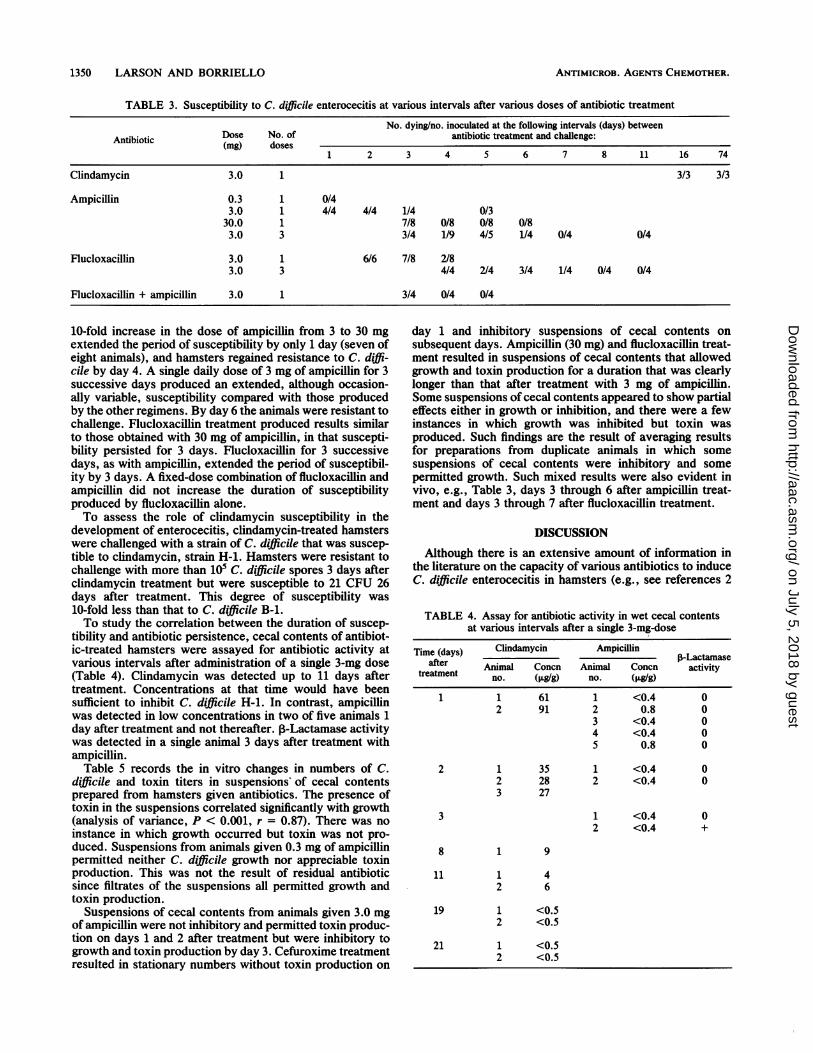
1350 LARSON AND BORRIELLO
TABLE 3. Susceptibility to C. difficile enterocecitis at various intervals after various doses of antibiotic treatment
No. dying/no. inoculated at the following intervals (days) betweenAntibiotic Dose No. of antibiotic treatment and challenge:(mg) doses
1 2 3 4 5 6 7 8 11 16 74
Clindamycin 3.0 1 3/3 3/3
Ampicillin 0.3 1 0/43.0 1 4/4 4/4 1/4 0/3
30.0 1 7/8 0/8 0/8 0/83.0 3 3/4 1/9 4/5 1/4 0/4 0/4
Flucloxacillin 3.0 1 6/6 7/8 2/83.0 3 4/4 2/4 3/4 1/4 0/4 0/4
Flucloxacillin + ampicillin 3.0 1 3/4 0/4 0/4
10-fold increase in the dose of ampicillin from 3 to 30 mgextended the period of susceptibility by only 1 day (seven ofeight animals), and hamsters regained resistance to C. diffi-cile by day 4. A single daily dose of 3 mg of ampicillin for 3successive days produced an extended, although occasion-ally variable, susceptibility compared with those producedby the other regimens. By day 6 the animals were resistant tochallenge. Flucloxacillin treatment produced results similarto those obtained with 30 mg of ampicillin, in that suscepti-bility persisted for 3 days. Flucloxacillin for 3 successivedays, as with ampicillin, extended the period of susceptibil-ity by 3 days. A fixed-dose combination of flucloxacillin andampicillin did not increase the duration of susceptibilityproduced by flucloxacillin alone.To assess the role of clindamycin susceptibility in the
development of enterocecitis, clindamycin-treated hamsterswere challenged with a strain of C. difficile that was suscep-tible to clindamycin, strain H-1. Hamsters were resistant tochallenge with more than 10i C. difficile spores 3 days afterclindamycin treatment but were susceptible to 21 CFU 26days after treatment. This degree of susceptibility was10-fold less than that to C. difficile B-1.To study the correlation between the duration of suscep-
tibility and antibiotic persistence, cecal contents of antibiot-ic-treated hamsters were assayed for antibiotic activity atvarious intervals after administration of a single 3-mg dose(Table 4). Clindamycin was detected up to 11 days aftertreatment. Concentrations at that time would have beensufficient to inhibit C. difficile H-1. In contrast, ampicillinwas detected in low concentrations in two of five animals 1day after treatment and not thereafter. P-Lactamase activitywas detected in a single animal 3 days after treatment withampicillin.Table 5 records the in vitro changes in numbers of C.
difficile and toxin titers in suspensions' of cecal contentsprepared from hamsters given antibiotics. The presence oftoxin in the suspensions correlated significantly with growth(analysis of variance, P < 0.001, r = 0.87). There was noinstance in which growth occurred but toxin was not pro-duced. Suspensions from animals given 0.3 mg of ampicillinpermitted neither C. difficile growth nor appreciable toxinproduction. This was not the result of residual antibioticsince filtrates of the suspensions all permitted growth andtoxin production.
Suspensions of cecal contents from animals given 3.0 mgof ampicillin were not inhibitory and permitted toxin produc-tion on days 1 and 2 after treatment but were inhibitory togrowth and toxin production by day 3. Cefuroxime treatmentresulted in stationary numbers without toxin production on
day 1 and inhibitory suspensions of cecal contents onsubsequent days. Ampicillin (30 mg) and flucloxacillin treat-ment resulted in suspensions of cecal contents that allowedgrowth and toxin production for a duration that was clearlylonger than that after treatment with 3 mg of ampicillin.Some suspensions of cecal contents appeared to show partialeffects either in growth or inhibition, and there were a fewinstances in which growth was inhibited but toxin wasproduced. Such findings are the result of averaging resultsfor preparations from duplicate animals in which somesuspensions of cecal contents were inhibitory and somepermitted growth. Such mixed results were also evident invivo, e.g., Table 3, days 3 through 6 after ampicillin treat-ment and days 3 through 7 after flucloxacillin treatment.
DISCUSSION
Although there is an extensive amount of information inthe literature on the capacity of various antibiotics to induceC. difficile enterocecitis in hamsters (e.g., see references 2
TABLE 4. Assay for antibiotic activity in wet cecal contentsat various intervals after a single 3-mg-dose
Time (days) Clindamycin Ampicillinafter Animal Concn Animal Concn activity
treatment no. (Lg/g) no. (4/g)1 1 61 1 <0.4 0
2 91 2 0.8 03 <0.4 04 <0.4 05 0.8 0
2 1 35 1 <0.4 02 28 2 <0.4 03 27
3 1 <0.4 02 <0.4 +
8 1 9
11 1 42 6
19 1 <0.52 <0.5
21 1 <0.52 <0.5
ANTIMICROB. AGENTS CHEMOTHER.
on July 5, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

QUANTITATIVE STUDY OF SUSCEPTIBILITY TO C. DIFFICILE
..b3 i13:x bo _Q
CDCDC
+
It l+I+ I+1I+1+
0 A'- --
1.~1t,4~~~~~--00 00~~DiC)
~3-j :PWO 'oo'ob1+ R1+1+1+1+1C o
LQLaOb
:I
wD00 00w
A6 )0 zA6 " " 00
'I *N *- * . cl p1+
o) oo)C
t'i04 t 0-0 e
1+ 1+1+1+1+ 1+
z Z 00000z
1+0oo
* **oi7
It
z
c
c
- .
coCD
3
CD
-3_)
CD
0
n
(I0
DQ
0
(D
r-L1vco
co0.CD
DI.
C)airmD0 .
(D,0-0.
00
'000.
Di0
0DiD
0-00Xi0
CD
0_DiD
'0
DiDCD
U'
(D
CD
CD0F
1351
and 10), we are not aware of previous efforts to measuresusceptibility or to use characterized challenge strains. Byand large, previous investigators have allowed antibiotic-treated animals to become infected fortuitously, although nodoubt, with time, the environments of animal holding unitsbecome heavily contaminated with a predominant strain ofC. difficile. The animal containment system we used pre-vents infection from ambient sources and permits challengeexperiments with measured numbers of defined microorgan-isms at specified times. An earlier study, in which contain-ment methods were used, showed that hamsters were sus-ceptibile to a very small number of C. difficile afterprolonged oral vancomycin treatment, whereas untreatedhamsters were uniformly resistant to challenges with largedoses of C. difficile (18). The current study considerablyextended this approach and permits a quantitative measureof susceptibility after treatment with several commonly usedantibiotics. The main findings are that all antibiotics testedproduced susceptibility to very small doses of C. difficile butthat these antibiotics differed to a smaller or greater extent inthe duration of susceptibility after treatment. A 10-foldincrease in the dose or the administration of three doses hada comparatively small effect on the duration of susceptibil-ity. Persistence of susceptibility was not necessarily due tocontinued detectability of the antibiotic. Finally, the state ofsusceptibility was modeled in vitro, and the correlation withsusceptibility in the animal was very strong.An effect on colonization resistance that outlasts detect-
able antibiotic activity may be a type of postantibiotic effect.Both clindamycin and ampicillin are associated with post-antibiotic effects in other systems (8); clindamycin is knownto produce the more prolonged effects and to act this way invivo as well as in vitro. Prolonged excretion of active drug,active metabolites, or both may produce an especially strongeffect. The mechanisms of the postantibiotic effect, andparticularly, how they might apply in the gut, are notimmediately obvious. A "postantibiotic effect" from clinda-mycin may simply be the elimination of the particularspecies responsible for colonization resistance.The data in Table 2 support the idea that susceptibility is
an all-or-nothing state; reversion to resistance occurredrapidly. At some intervals after antibiotic treatment, individ-ual differences in susceptibility were noted, especially inanimals which had received multiple doses of ampicillin orflucloxacillin. It is not clear from the results of these exper-iments why individual animals might vary in their responsesto multiple antibiotic doses. It is impossible from theseexperiments to say whether the apparent small differences inCFU/LD5O between clindamycin (1 and 2, Table 2) and theother antibiotics tested (5 to 8, Table 2) are real.
Residual clindamycin activity in the gut appeared toinhibit infection with clindamycin-susceptible strain H-1.However, this strain had a slightly reduced capacity to infecthamsters even when clindamycin activity was undetectable.Clindamycin-susceptible strains of C. difficile are fully capa-ble of causing illness in humans who receive clindamycin (9),but the onset of infection may be delayed until levels ofantibiotic in the gut have abated. The occurrence of infectionafter treatment with ampicillin, to which all virulent strainsare highly susceptible, has been ascribed to the induction of,B-lactamase in cecal contents (21). However, we found thatsmall doses of C. difficile spores were able to induce diseaseimmediately after a single oral dose of ampicillin, prior to theappearance of ,B-lactamase. Overall, there seems to be nodirect correlation between susceptibility to an antibiotic and
VOL. 34, 1990
()
CD0
CD CD 0:C) 0co
_.DCD
CD
CDQ
*-I
CI0
0.
CD
BCDCD
0
DD0.z
on July 5, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

1352 LARSON AND BORRIELLO
the capacity of a C. difficile strain to produce disease ineither hamsters or humans (9).
Borriello and Barclay (6) have shown in a clinical studythat the growth of C. difficile in fecal suspensions in vitroparallels in vivo colonization. The correlation between theexperiments reported in Tables 3 and 5 is striking. Thissuggests that the capacity of C. difficile to replicate in thefecal stream is a fundamental feature of the pathogenesis ofthe infection. The mechanism of resistance to colonizationby C. difficile in untreated animals postulated by Borrielloand Barclay (5, 6) requires the presence of viable microor-ganisms; it does not appear to be mediated by preformedbacterial products or metabolites that can be detected infecal filtrates (5, 6). The very rapid onset of susceptibilitynoted in the present study is consistent with a requirementfor viable organisms rather than soluble substances. Resto-ration of resistance after antibiotic treatment would then bethe result of their regrowth. Mediation of resistance bymeans of viable microorganisms explains the apparentlysuccessful efforts to manage relapses of PMC by administer-ing fecal suspensions by enema (7, 22).How might these results be applied to clinical situations?
Differences between hamsters and humans limit the clinicalapplicability of our results. One difference concerns speciesvariation in the way in which clindamycin is metabolized. In15 patients given clindamycin intravenously (14) for 48 hperioperatively, levels were still present in feces and, in-deed, were still increasing at day 5. Ten adult volunteersgiven oral clindamycin for 7 days (13) had substantial levelsin feces 2 days after they concluded the course of therapy,but there was no detectable antibiotic activity at 9 days. Thereview of clindamycin by Klainer (16) emphasizes its fecalpersistence and the potential role of this characteristic in thedevelopment of antibiotic-associated colitis. We found thatclindamycin is detectable in hamsters 11 days after a singledose but not at 19 days. The human data suggest somepersistence of the drug in feces, but not for as long as thatwhich we observed in hamsters. A second difference con-cerns the normal gut flora in hamsters and humans respon-sible for excluding C. difficile. It is not known what specificmicroorganisms are responsible for colonization resistancein either hamsters or humans, even at the genus level (4).However, we have successfully restored hamster coloniza-tion resistance with a suspension of human faeces (H. E.Larson, unpublished data). The relevant microorganismsand susceptibilities may not be different in the two species,but this possibility needs to be confirmed.A third difference concerns coprophagia. Prevention of
coprophagia did not shorten the duration of susceptibilityafter clindamycin treatment. We cannot be sure it did nothasten the return of colonization resistance after treatmentwith the other antibiotics. However, hamsters remainedsusceptible to clindamycin even when coprophagia wasallowed. A fourth difference concerns the many more per-mutations of dose, frequency, and route of administrationwhich are features of clinical use and which were not testedby our methods. Our results predict that differences in doseand frequency of administration are less important than thetype of antibiotic, but this prediction might be overcome byvery large doses or very long courses of treatment.One of the most striking of our findings was the very low
infective dose of C. difficile required to initiate infection.After clindamycin treatment, we reproducibly found thatonly one or two microorganisms were needed. It is notknown what dose of C. difficile is required to infect humansubjects. It may be very low since nosocomial C. difficile
infections appear to occur as a consequence of exposure tocontaminated fomites rather than to contaminated food orwater. This is likely to mean ingestion of only a fewmicroorganisms.We are not aware that the duration of susceptibility to C.
difficile infection has previously been assessed as a riskfactor for PMC. Prolonged susceptibility after antibiotictreatment offers more opportunity for exposure to andinfection with C. difficile. It has not usually been possible tocorrelate the risk of C. difficile colitis with dose or durationof antibiotic treatment. This difficulty is most likely due tooverriding differences in the degree to which at-risk patientshave been exposed to environmental sources of C. difficile.Given the low rates of colonization in normal subjects,exposure to the organism is likely to be the single mostimportant factor after antibiotic treatment that determinesthe risk of disease. The risk of exposure, however, is clearlyrelated to the duration of susceptibility. PMC has beenshown to occur after even short courses of cefoxitin given asperioperative infection prophylaxis (3). However, the post-operative duration of prophylactic antibiotic regimens iscontroversial, and PMC is their most important complication(15). We speculate that shorter rather than longer courses ofantibiotic treatment could reduce the risk of complicatingantibiotic-associated colitis.
ACKNOWLEDGMENTS
We thank A. Welch, F. Barclay, and H. Anderson for technicalassistance and E. Goodband for typing the manuscript.
LITERATURE CITED1. Aronsson, A., R. MolHby, and C. E. Nord. 1982. Clostridium
difficile and antibiotic associated diarrhoea in Sweden. Scand. J.Infect. Dis. 35(Suppl.):53-58.
2. Bartlett, J. G., T. W. Chang, N. Moon, and A. B. Onderdonk.1978. Antibiotic-induced lethal enterocolitis in hamsters: studieswith eleven agents and evidence to support the pathogenic roleof toxin-producing clostridia. Am. J. Vet. Res. 39:1525-1530.
3. Block, B. S., L. J. Mercer, M. A. Ismail, and A. H. Moawad.1985. Clostridium difficile-associated diarrhea follows perioper-ative prophylaxis with cefoxitin. Am. J. Obstet. Gynecol.153:835-838.
4. Borriello, S. P. 1989. Influence of the normal flora of the gut onClostridium difficile, p. 239-251. In R. Grubb, T. Midtvedt, andE. Norin (ed.), The regulatory and protective role of the normalmicroflora. Macmillan, London.
5. Borriello, S. P., and F. E. Barclay. 1984. Colonization resistanceto Clostridium difficile infection. Microecol. Ther. 14:75-87.
6. Borriello, S. P., and F. E. Barclay. 1986. An in-vitro model ofcolonisation resistance to Clostridium difficile infection. J. Med.Microbiol. 21:299-309.
7. Bowden, T. A., A. R. Mansberger, and L. E. Lykins. 1981.Pseudomembranous enterocolitis: mechanism of restoring floralhomeostasis. Am. Surg. 47:178-183.
8. Craig, W. A., and B. Vogelman. 1987. The postantibiotic effect.Ann. Intern. Med. 106:900-902.
9. Dzink, J., and J. G. Bartlett. 1980. In vitro susceptibility ofClostridium difficile isolates from patients with antibiotic-asso-ciated diarrhea or colitis. Antimicrob. Agents Chemother. 17:695-698.
10. Fekety, R., J. Silva, R. Toshniwal, M. Allo, J. Armstrong, R.Browne, J. Ebright, and G. Rifkin. 1979. Antibiotic-associatedcolitis: effects of antibiotics on Clostridium difficile and thedisease in hamsters. Rev. Infect. Dis. 1:386-396.
11. Gorbach, S. L., and J. G. Bartlett. 1977. Pseudomembranousenterocolitis: a review of its diverse forms. J. Infect. Dis.135:S89-S94.
12. Hawkes, R. A. 1979. General principles underlying laboratorydiagnosis of viral infections, p. 32-35. In E. H. Lennette and
ANTIMICROB. AGENTS CHEMOTHER.
on July 5, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

QUANTITATIVE STUDY OF SUSCEPTIBILITY TO C. DIFFICILE
N. J. Schmidt (ed.), Diagnostic procedures for viral, rickettsialand chlamydial infections, 5th ed. American Public HealthAssociation, Washington, D.C.
13. Heimdahl, A., and C. E. Nord. 1982. Effect of erythromycin andclindamycin on the indigenous human anaerobic flora and newcolonization of the gastrointestinal tract. Eur. J. Clin. Micro-biol. 1:38-48.
14. Kager, L., L. Liljeqvist, A. S. Mahnborg, and C. E. Nord. 1981.Effect of clindamycin prophylaxis on the colonic microflora inpatients undergoing colorectal surgery. Antimicrob. AgentsChemother. 20:736-740.
15. Kaiser, A. B. 1986. Antimicrobial prophylaxis in surgery. N.Engl. J. Med. 315:1129-1138.
16. Klainer, A. S. 1987. Clindamycin. Med. Clin. North Am.71:1169-1175.
17. Larson, H. E., A. B. Price, and S. P. Borriello. 1980. Epidemi-ology of experimental enterocecitis due to Clostridium difficile.J. Infect. Dis. 142:408-413.
18. Larson, H. E., A. B. Price, P. Honour, and S. P. Borriello. 1978.
Clostridium difficile and the aetiology of pseudomembranouscolitis. Lancet i:1063-1066.
19. Pierce, P. F., R. Wilson, J. Silva, Jr., V. F. Garagusi, G. D.Rifkin, R. Fekety, 0. Nunez-Montiel, Jr., V. R. Dowell, andJ. M. Hughes. 1982. Antibiotic-associated pseudomembranouscolitis: an epidemiologic investigation of a cluster of cases. J.Infect. Dis. 145:269-274.
20. Roberts, A. P., J. B. Eastwood, P. E. Gower, C. M. Fenton, andJ. R. Curtis. 1978. Serum and plasma concentrations of clinda-mycin following a single injection of clindamycin phosphate inmaintenance haemodialysis patients and normal subjects. Eur.J. Clin. Pharmacol. 14:435-439.
21. Rolfe, R. D., and S. M. Finegold. 1983. Intestinal P-lactamaseactivity in ampicillin-induced, Clostridium difficile-associatedileocecitis. J. Infect. Dis. 147:227-235.
22. Schwan, A., S. Sjolin, V. Trottestam, and B. Aronsson. 1984.Relapsing Clostridium difficile enterocolitis cured by rectalinfusion of normal faeces. Scand. J. Infect. Dis. 16:211-215.
VOL. 34, 1990 1353
on July 5, 2018 by guesthttp://aac.asm
.org/D
ownloaded from


















