Protein Sequencing Strategy.docx
9
Step 1: Separation of Polypeptide Chains: If the protein of interest is a heteromultimer (composed of more than one type of polypeptide chain), then the protein must be dissociated and its component polypeptide subunits must be separated from one another and sequenced individually. Subunit associations in multimeric proteins are typically maintained solely by noncovalent forces, and th erefore most multimeric proteins can usually be dissociated by exposure to pH extremes, 8 M urea, 6 M guanidinium hydrochloride, or high salt concentrations. (All of these treatments disrupt polar interactions such as hydrogen bonds both within the protein molecule and between the protein and the aqueous solvent.) Once dissociated, the individual polypeptides can be isolated from one another on the basis of differences in size and/or charge. Occasionally, heteromultimers are linked together by interchain S--S bridges. In such instances, these cross-links must be cleaved prior to dissociation and isolation of the individual chains. The methods described under step 2 are applicable for this purpose. Step 2: Cleavage of Disulfide Bridges A number of methods exist for cleaving disulfides. An important consideration is to carry out these cleavages so that the original or even new S--S links do not form. Oxidation of a disulfide by performic acid results in t he formation of two equivalents of cysteic acid. Because these cysteic acid side chains are ionized SO3 groups, electrostatic repulsion (as well as altered chemistry) prevents S--S recombination. Alternatively, sulfhydryl compounds such as 2-mercaptoethanol readily reduce S--S bridges to re generate two cysteine-SH side chains. However, these SH groups recombine to re-form either the original disulfide link or, if other free Cys-SHs are available, new disulfide links. To prevent this, SOS reduction must be followed by treatment with alkylating agents such as iodoacetate or 3-bromopropylamine, which modify the SH groups and block disulfide bridge formation.
Transcript of Protein Sequencing Strategy.docx

7/27/2019 Protein Sequencing Strategy.docx
http://slidepdf.com/reader/full/protein-sequencing-strategydocx 1/9
Step 1: Separation of Polypeptide Chains:
If the protein of interest is a heteromultimer (composed of more than one type of polypeptide
chain), then the protein must be dissociated and its component polypeptide subunits must be
separated from one another and sequenced individually. Subunit associations in multimeric proteins are typically maintained solely by noncovalent forces, and therefore most multimeric
proteins can usually be dissociated by exposure to pH extremes, 8 M urea, 6 M guanidinium
hydrochloride, or high salt concentrations. (All of these treatments disrupt polar interactions
such as hydrogen bonds both within the protein molecule and between the protein and the
aqueous solvent.) Once dissociated, the individual polypeptides can be isolated from one
another on the basis of differences in size and/or charge. Occasionally, heteromultimers are
linked together by interchain S--S bridges. In such instances, these cross-links must be
cleaved prior to dissociation and isolation of the individual chains. The methods described
under step 2 are applicable for this purpose.
Step 2: Cleavage of Disulfide Bridges
A number of methods exist for cleaving disulfides. An important consideration is to carry out
these cleavages so that the original or even new S--S links do not form. Oxidation of a
disulfide by performic acid results in the formation of two equivalents of cysteic acid.
Because these cysteic acid side chains are ionized SO3 groups, electrostatic repulsion (as well
as altered chemistry) prevents S--S recombination. Alternatively, sulfhydryl compounds such
as 2-mercaptoethanol readily reduce S--S bridges to regenerate two cysteine-SH side chains.
However, these SH groups recombine to re-form either the original disulfide link or, if other
free Cys-SHs are available, new disulfide links. To prevent this, SOS reduction must be
followed by treatment with alkylating agents such as iodoacetate or 3-bromopropylamine,
which modify the SH groups and block disulfide bridge formation.
7/27/2019 Protein Sequencing Strategy.docx
http://slidepdf.com/reader/full/protein-sequencing-strategydocx 2/9
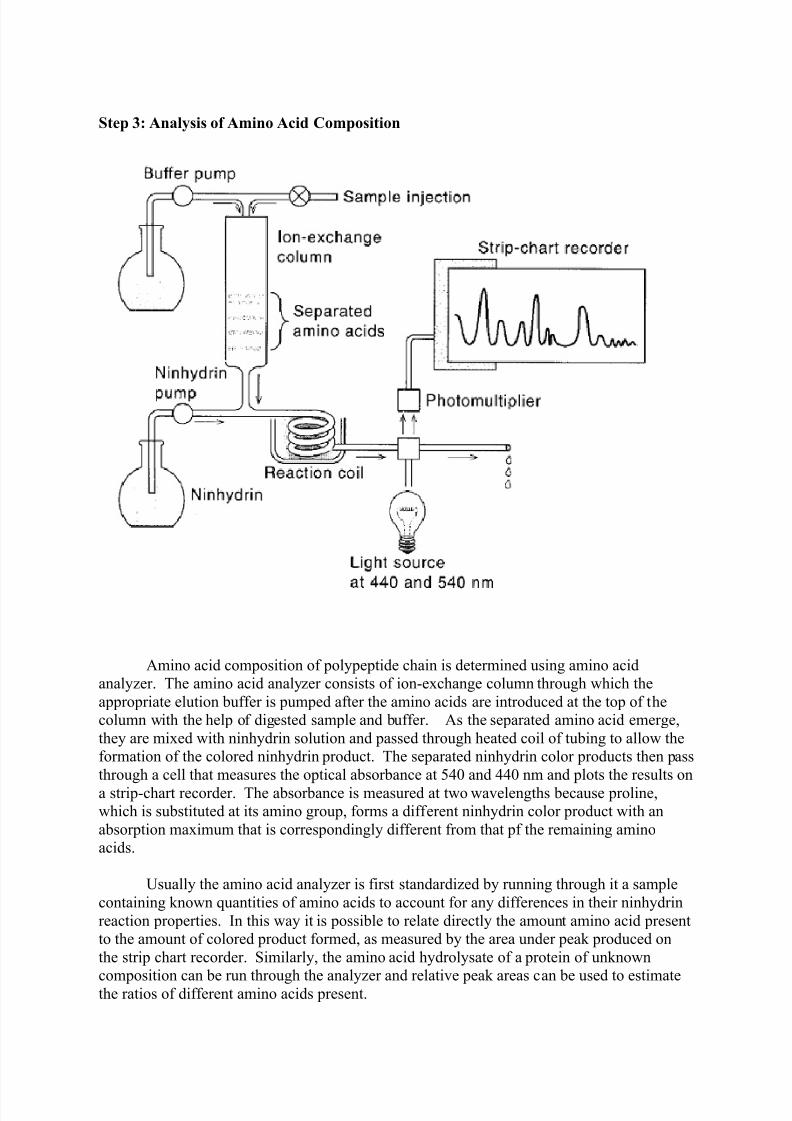
Step 3: Analysis of Amino Acid Composition
Amino acid composition of polypeptide chain is determined using amino acid
analyzer. The amino acid analyzer consists of ion-exchange column through which the
appropriate elution buffer is pumped after the amino acids are introduced at the top of the
column with the help of digested sample and buffer. As the separated amino acid emerge,
they are mixed with ninhydrin solution and passed through heated coil of tubing to allow the
formation of the colored ninhydrin product. The separated ninhydrin color products then pass
through a cell that measures the optical absorbance at 540 and 440 nm and plots the results on
a strip-chart recorder. The absorbance is measured at two wavelengths because proline,
which is substituted at its amino group, forms a different ninhydrin color product with an
absorption maximum that is correspondingly different from that pf the remaining amino
acids.
Usually the amino acid analyzer is first standardized by running through it a sample
containing known quantities of amino acids to account for any differences in their ninhydrin
reaction properties. In this way it is possible to relate directly the amount amino acid present
to the amount of colored product formed, as measured by the area under peak produced on
the strip chart recorder. Similarly, the amino acid hydrolysate of a protein of unknown
composition can be run through the analyzer and relative peak areas can be used to estimatethe ratios of different amino acids present.
7/27/2019 Protein Sequencing Strategy.docx
http://slidepdf.com/reader/full/protein-sequencing-strategydocx 3/9
Step 4: Identification of the N- and C-Terminal Residues
End-group analysis reveals several things. First, it identifies the N- and C-terminal residues in
the polypeptide chain. Second, it can be a clue to the number of ends in the protein. That is, if
the protein consists of two or more different polypeptide chains, then more than one end
group may be discovered, alerting the investigator to the presence of multiple polypeptides.
7/27/2019 Protein Sequencing Strategy.docx
http://slidepdf.com/reader/full/protein-sequencing-strategydocx 4/9
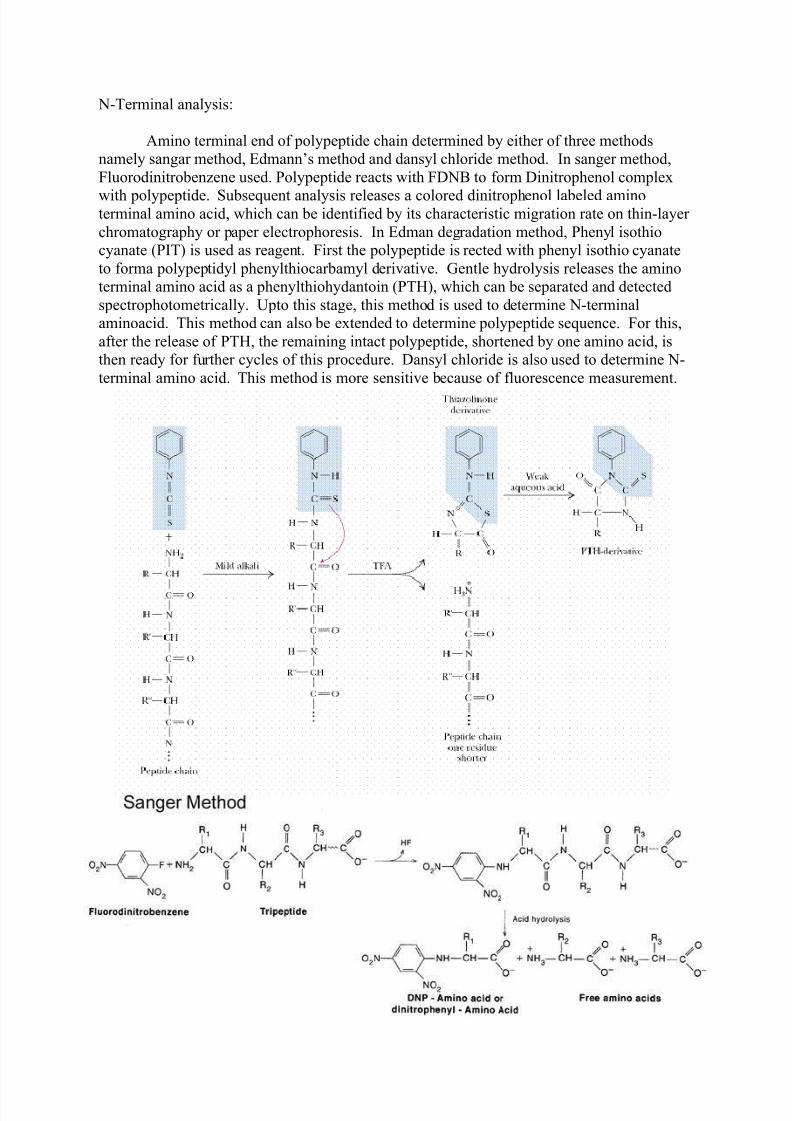
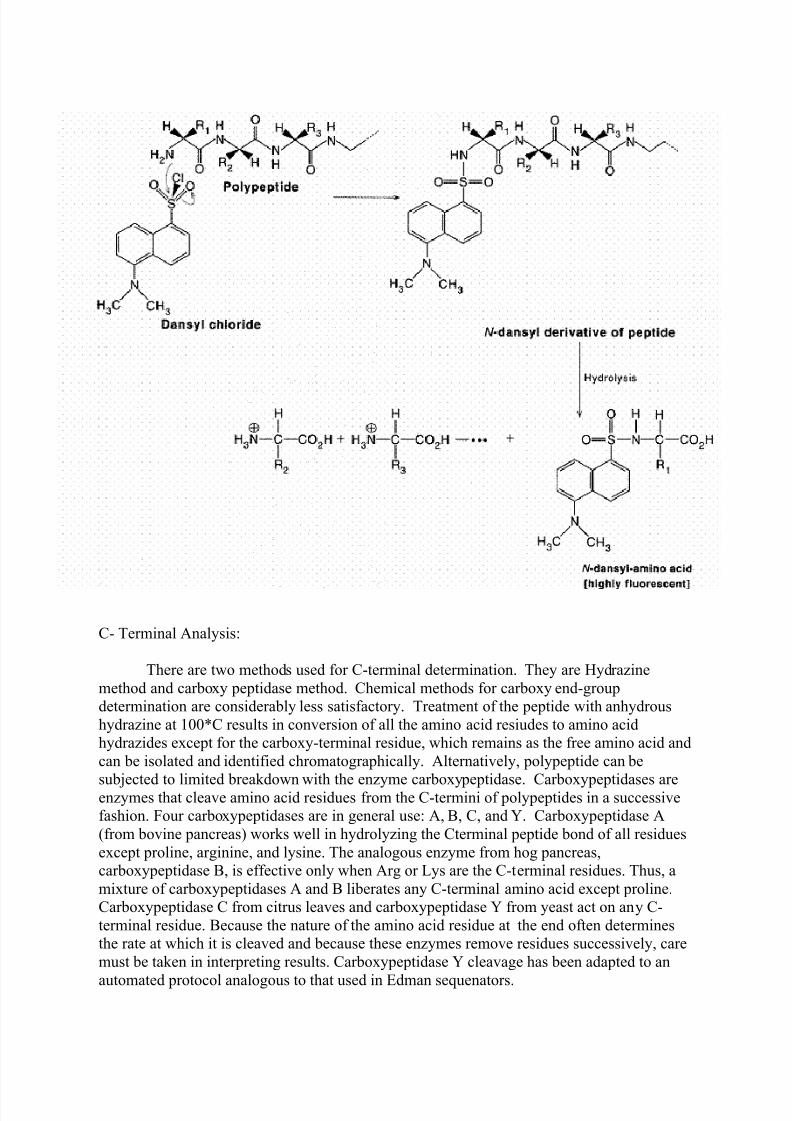
N-Terminal analysis:
Amino terminal end of polypeptide chain determined by either of three methods
namely sangar method, Edmann’s method and dansyl chloride method. In sanger method,
Fluorodinitrobenzene used. Polypeptide reacts with FDNB to form Dinitrophenol complex
with polypeptide. Subsequent analysis releases a colored dinitrophenol labeled aminoterminal amino acid, which can be identified by its characteristic migration rate on thin-layer
chromatography or paper electrophoresis. In Edman degradation method, Phenyl isothio
cyanate (PIT) is used as reagent. First the polypeptide is rected with phenyl isothio cyanate
to forma polypeptidyl phenylthiocarbamyl derivative. Gentle hydrolysis releases the amino
terminal amino acid as a phenylthiohydantoin (PTH), which can be separated and detected
spectrophotometrically. Upto this stage, this method is used to determine N-terminal
aminoacid. This method can also be extended to determine polypeptide sequence. For this,
after the release of PTH, the remaining intact polypeptide, shortened by one amino acid, is
then ready for further cycles of this procedure. Dansyl chloride is also used to determine N-
terminal amino acid. This method is more sensitive because of fluorescence measurement.
7/27/2019 Protein Sequencing Strategy.docx
http://slidepdf.com/reader/full/protein-sequencing-strategydocx 5/9
C- Terminal Analysis:
There are two methods used for C-terminal determination. They are Hydrazine
method and carboxy peptidase method. Chemical methods for carboxy end-group
determination are considerably less satisfactory. Treatment of the peptide with anhydrous
hydrazine at 100*C results in conversion of all the amino acid resiudes to amino acid
hydrazides except for the carboxy-terminal residue, which remains as the free amino acid and
can be isolated and identified chromatographically. Alternatively, polypeptide can be
subjected to limited breakdown with the enzyme carboxypeptidase. Carboxypeptidases areenzymes that cleave amino acid residues from the C-termini of polypeptides in a successive
fashion. Four carboxypeptidases are in general use: A, B, C, and Y. Carboxypeptidase A
(from bovine pancreas) works well in hydrolyzing the Cterminal peptide bond of all residues
except proline, arginine, and lysine. The analogous enzyme from hog pancreas,
carboxypeptidase B, is effective only when Arg or Lys are the C-terminal residues. Thus, a
mixture of carboxypeptidases A and B liberates any C-terminal amino acid except proline.
Carboxypeptidase C from citrus leaves and carboxypeptidase Y from yeast act on any C-
terminal residue. Because the nature of the amino acid residue at the end often determines
the rate at which it is cleaved and because these enzymes remove residues successively, care
must be taken in interpreting results. Carboxypeptidase Y cleavage has been adapted to an
automated protocol analogous to that used in Edman sequenators.

7/27/2019 Protein Sequencing Strategy.docx
http://slidepdf.com/reader/full/protein-sequencing-strategydocx 6/9
Step- 5: Fragmentation of the Polypeptide Chain
The aim at this step is to produce fragments useful for sequence analysis. The cleavage
methods employed are usually enzymatic, but proteins can also be fragmented by specific or
nonspecific chemical means (such as partial acid hydrolysis). Fragmentation can be
achieved by the use of endopeptidases, which are enzymes that catalyze polypeptide chain
cleavage at specific sites in the protein. The specificity of four endopeptidases commonly for
this purpose are Pepsin, Trypsin, Chymotrypsin and Papain. Another specific chemicalmethod for polypeptide chain cleavage involves reaction with cyanogens bromide. This
reaction cleaves specifically at the methionine residues, with the accompanying conversion of
free carboxyl-terminal methionine to homoserine lactone. For sequencing protein minimum
two proteolytic factors should be selected and two sets of fragements should be prepared.

7/27/2019 Protein Sequencing Strategy.docx
http://slidepdf.com/reader/full/protein-sequencing-strategydocx 7/9
Step 6: Sequencing Peptide fragments
Peptides resulting from cleavage of the intact protein are generally separated by
column chromatography. The isolated peptides may then be analyzed to determine their
sequence. There two methods recently used for this purpose, they are Edman degradation
and tandem mass spectrometer. Devices called sequenators are available that automate
Edman degradation procedure. The success of these devices depends in large part on the
technical innovation of covalently linking the peptide to be sequenced to glass
beads. Attachment of peptide through its carboxy-terminal group to this immobile phase
facilitates the complete removal of potentially conataminating reaction procedure during
successive stages of the degradation. Thus with the help of this sequenators, peptide
fragements are sequenced.
The other method used for protein sequencing is tandem mass spectrometer. Tandem
MS (or MS/MS) allows sequencing of proteins by hooking two mass spectrometers in tandem.
The first mass spectrometer is used to separate oligopeptides from a protein digest and then to
select in turn each of these oligopeptides for further analysis. A selected ionized oligopeptideis directed toward the second mass spectrometer; on the way, this oligopeptide is fragmented
by collision with helium or argon gas molecules, and the collection of fragments is analyzed
by the second mass spectrometer. Fragmentation occurs primarily in the peptide bonds
linking successive amino acids in the oligopeptide, so the fragments created represent a
nested set of peptides that differ in size by one amino acid residue. The fragments differ in
mass by 56 atomic mass units (the mass of the peptide backbone atoms (NH-CH-CO)) plus
the mass of the R group at each position, which ranges from 1 atomic mass unit (Gly) to 130
(Trp). MS sequencing has the advantages of very high sensitivity, fast sample processing, and
the ability to work with mixtures of proteins. Subpicomoles (less than 10_12 moles) of
peptide can be analyzed. However, in practice, tandem MS is limited to rather short
sequences (no longer than 15 or so amino acid residues).

7/27/2019 Protein Sequencing Strategy.docx
http://slidepdf.com/reader/full/protein-sequencing-strategydocx 8/9
Step 7: Reconstruction of the Overall Amino Acid Sequence
The sequences obtained for the sets of fragments derived from two or more cleavage
procedures are now compared, with the objective being to find overlaps that establish
continuity of the overall amino acid sequence of the polypeptide chain. Peptides generated
from specific hydrolysis of the polypeptide can be aligned to reveal the overall amino acid
sequence. Such comparisons are also useful in eliminating errors and validating the accuracy
of the sequences determined for the individual fragments.
Step 8: Location of Disulfide Cross-Bridges
Strictly speaking, the disulfide bonds formed between cysteine residues in a protein are not a
part of its primary structure. Nevertheless, information about their location can be obtained
by procedures used in sequencing, provided the disulfides are not broken prior to cleaving the
polypeptide chain. Because these covalent bonds are stable under most conditions used in the
cleavage of polypeptides, intact disulfides link the peptide fragments containing their specific
cysteinyl residues and thus these linked fragments can be isolated and identified within the
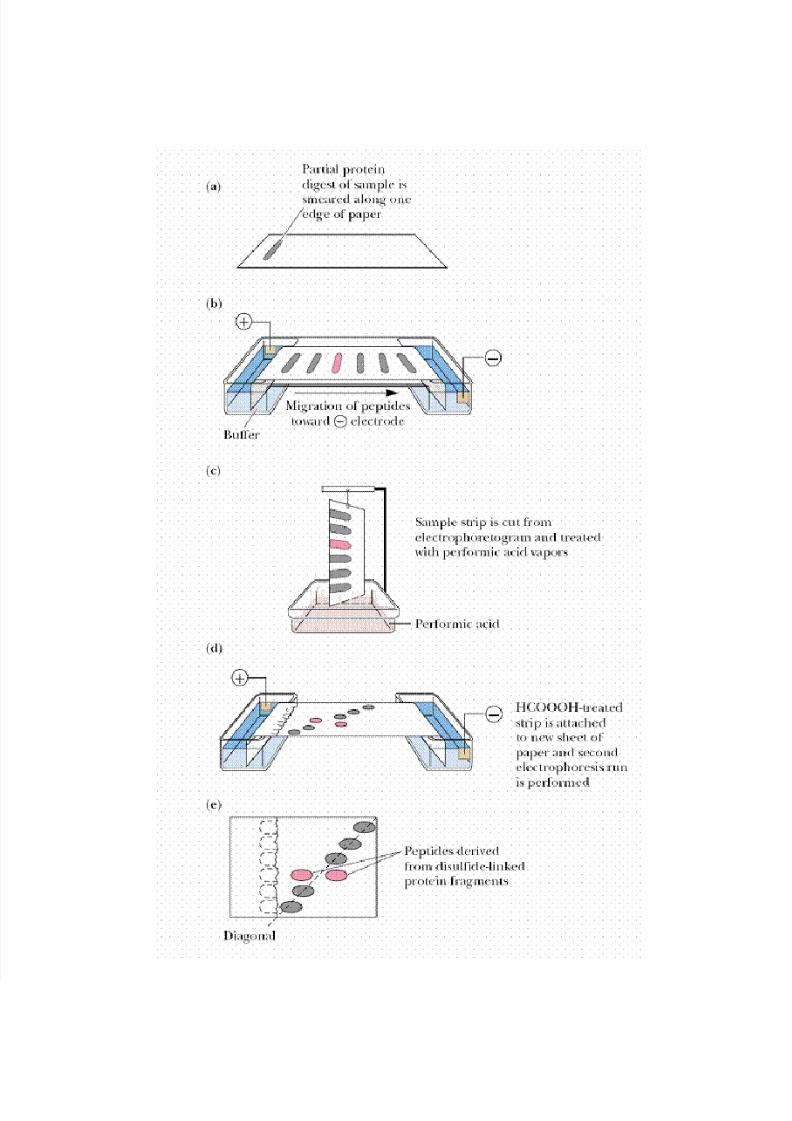
protein digest. An effective way to isolate these fragments is through diagonal
electrophoresis.
A protein digest in which any disulfide bonds remain intact and link their respective Cys-
containing peptides is streaked along the edge of a filter paper and subjected to
electrophoresis. A strip cut from the edge of the paper is then exposed to performic acid
fumes to oxidize any disulfide bridges. Then the paper strip is attached to a new filter paper
so that a second electrophoresis can be run in a direction perpendicular to the first. Peptides
devoid of disulfides experience no mobility change, and thus their pattern of migration
defines a diagonal. Peptides that had disulfides migrate off this diagonal and can be easily
identified, isolated, and sequenced to reveal the location of cysteic acid residues formerly
involved in disulfide bridges. From this information, the positions of the disulfides in the
protein can be stipulated.
7/27/2019 Protein Sequencing Strategy.docx
http://slidepdf.com/reader/full/protein-sequencing-strategydocx 9/9