
Photosynthetic Protists
6
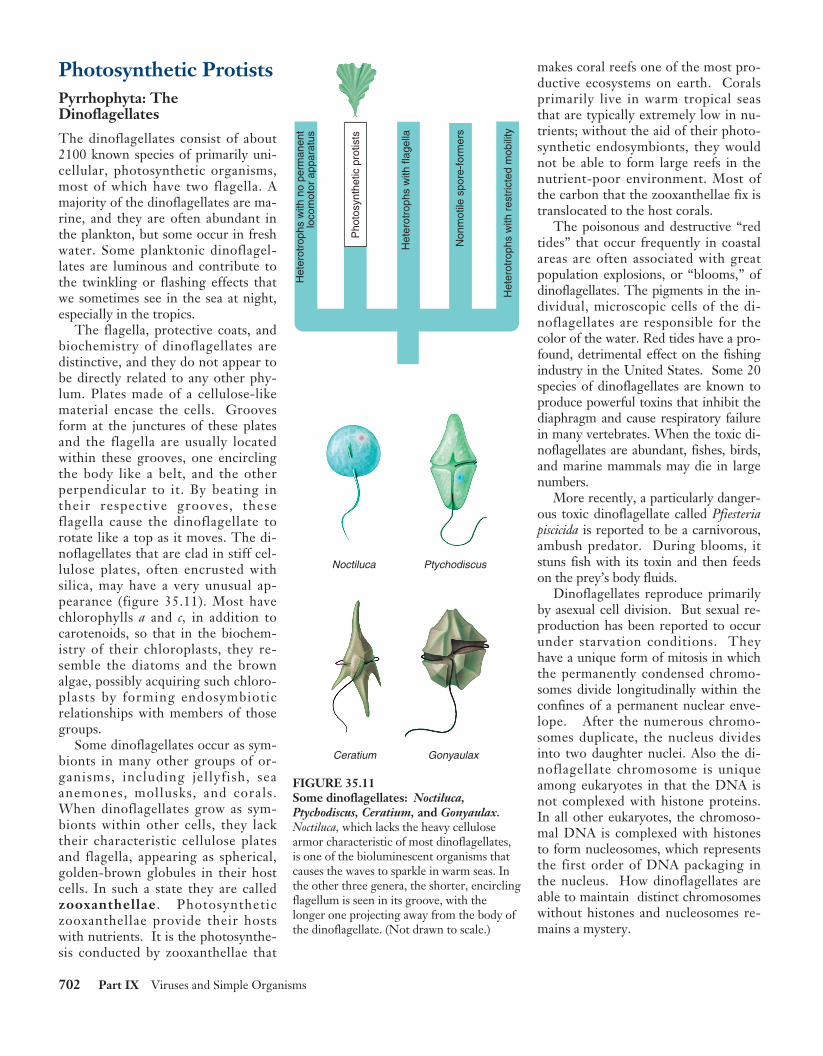
Photosynthetic Protists Pyrrhophyta: The Dinoflagellates The dinoflagellates consist of about 2100 known species of primarily uni- cellular, photosynthetic organisms, most of which have two flagella. A majority of the dinoflagellates are ma- rine, and they are often abundant in the plankton, but some occur in fresh water. Some planktonic dinoflagel- lates are luminous and contribute to the twinkling or flashing effects that we sometimes see in the sea at night, especially in the tropics. The flagella, protective coats, and biochemistry of dinoflagellates are distinctive, and they do not appear to be directly related to any other phy- lum. Plates made of a cellulose-like material encase the cells. Grooves form at the junctures of these plates and the flagella are usually located within these grooves, one encircling the body like a belt, and the other perpendicular to it. By beating in their respective grooves, these flagella cause the dinoflagellate to rotate like a top as it moves. The di- noflagellates that are clad in stiff cel- lulose plates, often encrusted with silica, may have a very unusual ap- pearance (figure 35.11). Most have chlorophylls a and c, in addition to carotenoids, so that in the biochem- istry of their chloroplasts, they re- semble the diatoms and the brown algae, possibly acquiring such chloro- plasts by forming endosymbiotic relationships with members of those groups. Some dinoflagellates occur as sym- bionts in many other groups of or- ganisms, including jellyfish, sea anemones, mollusks, and corals. When dinoflagellates grow as sym- bionts within other cells, they lack their characteristic cellulose plates and flagella, appearing as spherical, golden-brown globules in their host cells. In such a state they are called zooxanthellae . Photosynthetic zooxanthellae provide their hosts with nutrients. It is the photosynthe- sis conducted by zooxanthellae that makes coral reefs one of the most pro- ductive ecosystems on earth. Corals primarily live in warm tropical seas that are typically extremely low in nu- trients; without the aid of their photo- synthetic endosymbionts, they would not be able to form large reefs in the nutrient-poor environment. Most of the carbon that the zooxanthellae fix is translocated to the host corals. The poisonous and destructive “red tides” that occur frequently in coastal areas are often associated with great population explosions, or “blooms,” of dinoflagellates. The pigments in the in- dividual, microscopic cells of the di- noflagellates are responsible for the color of the water. Red tides have a pro- found, detrimental effect on the fishing industry in the United States. Some 20 species of dinoflagellates are known to produce powerful toxins that inhibit the diaphragm and cause respiratory failure in many vertebrates. When the toxic di- noflagellates are abundant, fishes, birds, and marine mammals may die in large numbers. More recently, a particularly danger- ous toxic dinoflagellate called Pfiesteria piscicida is reported to be a carnivorous, ambush predator. During blooms, it stuns fish with its toxin and then feeds on the prey’s body fluids. Dinoflagellates reproduce primarily by asexual cell division. But sexual re- production has been reported to occur under starvation conditions. They have a unique form of mitosis in which the permanently condensed chromo- somes divide longitudinally within the confines of a permanent nuclear enve- lope. After the numerous chromo- somes duplicate, the nucleus divides into two daughter nuclei. Also the di- noflagellate chromosome is unique among eukaryotes in that the DNA is not complexed with histone proteins. In all other eukaryotes, the chromoso- mal DNA is complexed with histones to form nucleosomes, which represents the first order of DNA packaging in the nucleus. How dinoflagellates are able to maintain distinct chromosomes without histones and nucleosomes re- mains a mystery. 702 Part IX Viruses and Simple Organisms Noctiluca Ptychodiscus Ceratium Gonyaulax Heterotrophs with no permanent locomotor apparatus Photosynthetic protists Heterotrophs with flagella Nonmotile spore-formers Heterotrophs with restricted mobility FIGURE 35.11 Some dinoflagellates: Noctiluca, Ptychodiscus, Ceratium, and Gonyaulax. Noctiluca, which lacks the heavy cellulose armor characteristic of most dinoflagellates, is one of the bioluminescent organisms that causes the waves to sparkle in warm seas. In the other three genera, the shorter, encircling flagellum is seen in its groove, with the longer one projecting away from the body of the dinoflagellate. (Not drawn to scale.)
Transcript of Photosynthetic Protists
Photosynthetic ProtistsPyrrhophyta: TheDinoflagellates
The dinoflagellates consist of about2100 known species of primarily uni-cellular, photosynthetic organisms,most of which have two flagella. Amajority of the dinoflagellates are ma-rine, and they are often abundant inthe plankton, but some occur in freshwater. Some planktonic dinoflagel-lates are luminous and contribute tothe twinkling or flashing effects thatwe sometimes see in the sea at night,especially in the tropics.
The flagella, protective coats, andbiochemistry of dinoflagellates aredistinctive, and they do not appear tobe directly related to any other phy-lum. Plates made of a cellulose-likematerial encase the cells. Groovesform at the junctures of these platesand the flagella are usually locatedwithin these grooves, one encirclingthe body like a belt, and the otherperpendicular to it. By beating intheir respective grooves, theseflagella cause the dinoflagellate torotate like a top as it moves. The di-noflagellates that are clad in stiff cel-lulose plates, often encrusted withsilica, may have a very unusual ap-pearance (figure 35.11). Most havechlorophylls a and c, in addition tocarotenoids, so that in the biochem-istry of their chloroplasts, they re-semble the diatoms and the brownalgae, possibly acquiring such chloro-plasts by forming endosymbioticrelationships with members of thosegroups.
Some dinoflagellates occur as sym-bionts in many other groups of or-ganisms, including jellyfish, seaanemones, mollusks, and corals.When dinoflagellates grow as sym-bionts within other cells, they lacktheir characteristic cellulose platesand flagella, appearing as spherical,golden-brown globules in their hostcells. In such a state they are calledzooxanthellae . Photosyntheticzooxanthellae provide their hostswith nutrients. It is the photosynthe-sis conducted by zooxanthellae that
makes coral reefs one of the most pro-ductive ecosystems on earth. Coralsprimarily live in warm tropical seasthat are typically extremely low in nu-trients; without the aid of their photo-synthetic endosymbionts, they wouldnot be able to form large reefs in thenutrient-poor environment. Most ofthe carbon that the zooxanthellae fix istranslocated to the host corals.
The poisonous and destructive “redtides” that occur frequently in coastalareas are often associated with greatpopulation explosions, or “blooms,” ofdinoflagellates. The pigments in the in-dividual, microscopic cells of the di-noflagellates are responsible for thecolor of the water. Red tides have a pro-found, detrimental effect on the fishingindustry in the United States. Some 20species of dinoflagellates are known toproduce powerful toxins that inhibit thediaphragm and cause respiratory failurein many vertebrates. When the toxic di-noflagellates are abundant, fishes, birds,and marine mammals may die in largenumbers.
More recently, a particularly danger-ous toxic dinoflagellate called Pfiesteriapiscicida is reported to be a carnivorous,ambush predator. During blooms, itstuns fish with its toxin and then feedson the prey’s body fluids.
Dinoflagellates reproduce primarilyby asexual cell division. But sexual re-production has been reported to occurunder starvation conditions. Theyhave a unique form of mitosis in whichthe permanently condensed chromo-somes divide longitudinally within theconfines of a permanent nuclear enve-lope. After the numerous chromo-somes duplicate, the nucleus dividesinto two daughter nuclei. Also the di-noflagellate chromosome is uniqueamong eukaryotes in that the DNA isnot complexed with histone proteins.In all other eukaryotes, the chromoso-mal DNA is complexed with histonesto form nucleosomes, which representsthe first order of DNA packaging inthe nucleus. How dinoflagellates areable to maintain distinct chromosomeswithout histones and nucleosomes re-mains a mystery.
702 Part IX Viruses and Simple Organisms
Noctiluca Ptychodiscus
Ceratium Gonyaulax
Het
erot
roph
s w
ith n
o pe
rman
ent
loco
mot
or a
ppar
atus
Pho
tosy
nthe
tic p
rotis
ts
Het
erot
roph
s w
ith fl
agel
la
Non
mot
ile s
pore
-for
mer
s
Het
erot
roph
s w
ith r
estr
icte
d m
obili
ty
FIGURE 35.11Some dinoflagellates: Noctiluca,Ptychodiscus, Ceratium, and Gonyaulax.Noctiluca, which lacks the heavy cellulosearmor characteristic of most dinoflagellates,is one of the bioluminescent organisms thatcauses the waves to sparkle in warm seas. Inthe other three genera, the shorter, encirclingflagellum is seen in its groove, with thelonger one projecting away from the body ofthe dinoflagellate. (Not drawn to scale.)
rav31208_ch35.qxd 7/19/01 12:50 PM Page 702
Euglenophyta: The Euglenoids
Most of the approximately 1000 known species of eugle-noids live in fresh water. The members of this phylumclearly illustrate the impossibility of distinguishing “plants”from “animals” among the protists. About a third of the ap-proximately 40 genera of euglenoids have chloroplasts andare fully autotrophic; the others lack chloroplasts, ingesttheir food, and are heterotrophic. These organisms are notsignificantly different from some groups of zoomastigotes(see next section), and many biologists believe that the twophyla should be merged into one.
Some euglenoids with chloroplasts may become het-erotrophic if the organisms are kept in the dark; thechloroplasts become small and nonfunctional. If they areput back in the light, they may become green within a fewhours. Normally photosynthetic euglenoids may sometimesfeed on dissolved or particulate food.
Individual euglenoids range from 10 to 500 micrometerslong and are highly variable in form. Interlocking proteina-ceous strips arranged in a helical pattern form a flexiblestructure called the pellicle, which lies within the cellmembrane of the euglenoids. Because its pellicle is flexible,a euglenoid is able to change its shape. Reproduction inthis phylum occurs by mitotic cell division. The nuclear en-velope remains intact throughout the process of mitosis.No sexual reproduction is known to occur in this group.
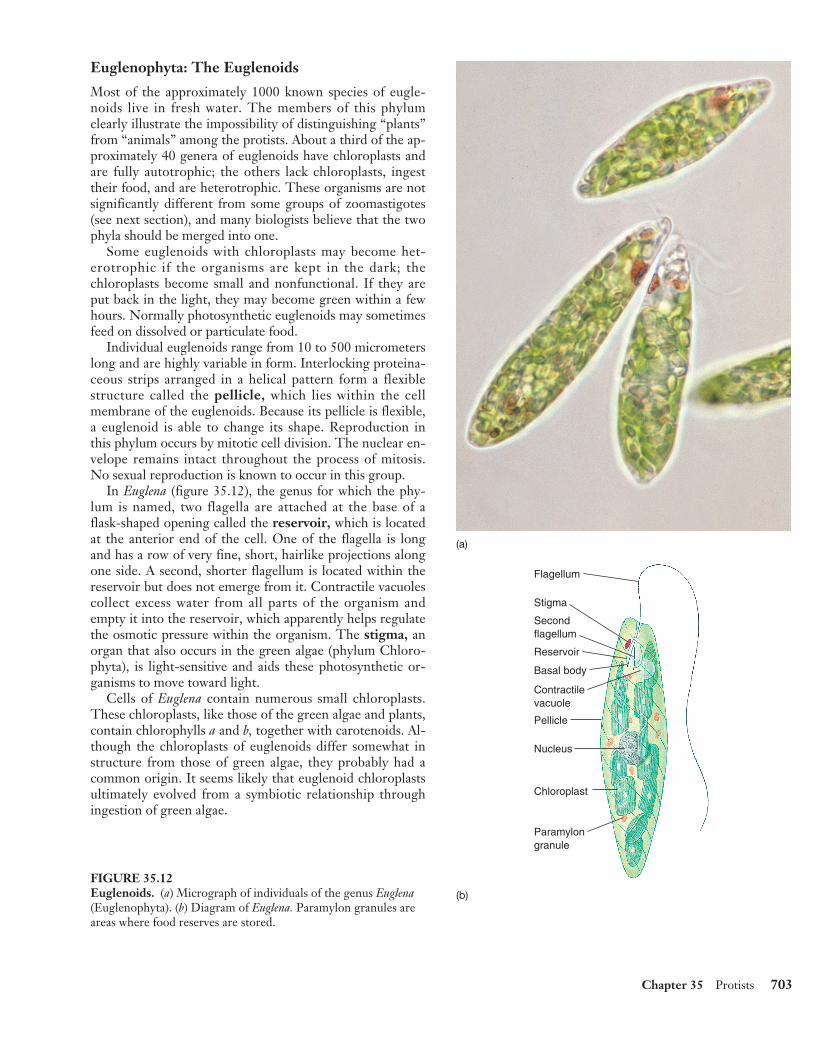
In Euglena (figure 35.12), the genus for which the phy-lum is named, two flagella are attached at the base of aflask-shaped opening called the reservoir, which is locatedat the anterior end of the cell. One of the flagella is longand has a row of very fine, short, hairlike projections alongone side. A second, shorter flagellum is located within thereservoir but does not emerge from it. Contractile vacuolescollect excess water from all parts of the organism andempty it into the reservoir, which apparently helps regulatethe osmotic pressure within the organism. The stigma, anorgan that also occurs in the green algae (phylum Chloro-phyta), is light-sensitive and aids these photosynthetic or-ganisms to move toward light.
Cells of Euglena contain numerous small chloroplasts.These chloroplasts, like those of the green algae and plants,contain chlorophylls a and b, together with carotenoids. Al-though the chloroplasts of euglenoids differ somewhat instructure from those of green algae, they probably had acommon origin. It seems likely that euglenoid chloroplastsultimately evolved from a symbiotic relationship throughingestion of green algae.
Chapter 35 Protists 703
(a)
Reservoir
Pellicle
Basal body
Contractilevacuole
Secondflagellum
Stigma
Flagellum
Nucleus
Chloroplast
Paramylongranule
(b)
FIGURE 35.12Euglenoids. (a) Micrograph of individuals of the genus Euglena(Euglenophyta). (b) Diagram of Euglena. Paramylon granules areareas where food reserves are stored.
rav31208_ch35.qxd 7/19/01 12:50 PM Page 703

Chrysophyta: The Diatoms and Golden Algae
The Diatoms. Diatoms, members of the phylum Chrys-ophyta, are photosynthetic, unicellular organisms withunique double shells made of opaline silica, which are oftenstrikingly and characteristically marked. The shells of di-atoms are like small boxes with lids, one half of the shell fit-ting inside the other. Their chloroplasts, with chlorophyllsa and c, as well as carotenoids, resemble those of the brownalgae and dinoflagellates. In other respects, however, thereare few similarities between these groups, and they proba-bly do not share an immediate common ancestor. Anothermember of the phylum Chrysophyta is the golden algae.Diatoms and golden algae are grouped together becausethey both produce a unique carbohydrate called chrysolam-inarin.
There are more than 11,500 living species of diatoms,with many more known in the fossil record. The shells offossil diatoms often form very thick deposits, which aresometimes mined commercially. The resulting “diatoma-ceous earth” is used as an abrasive or to add the sparklingquality to the paint used on roads, among other purposes.Living diatoms are often abundant both in the sea and infresh water, where they are important food producers. Di-atoms occur in the plankton and are attached to submergedobjects in relatively shallow water. Many species are able tomove by means of a secretion that is produced from a finegroove along each shell. The diatoms exude and perhapsalso retract this secretion as they move.
There are two major groups of diatoms, one with radialsymmetry (like a wheel) and the other with bilateral (two-sided) symmetry (figure 35.13). Diatom shells are rigid, andthe organisms reproduce asexually by separating the twohalves of the shell, each half then regenerating another halfshell within it. Because of this mode of reproduction, thereis a tendency for the shells, and consequently the individualdiatoms, to get smaller and smaller with each asexual re-production. When the resulting individuals have dimin-ished to about 30% of their original size, one may slip outof its shell, grow to full size, and regenerate a full-sized pairof new shells.
Individual diatoms are diploid. Meiosis occurs more fre-quently under conditions of starvation. Some marine di-atoms produce numerous sperm and others a single egg. Iffusion occurs, the resulting zygote regenerates a full-sizedindividual. In some freshwater diatoms, the gametes areamoeboid and similar in appearance.
The Golden Algae. Also included within the Chryso-phyta are the golden algae, named for the yellow andbrown carotenoid and xanthophyll accessory pigments intheir chloroplasts, which give them a golden color. Unicel-lular but often colonial, these freshwater protists typicallyhave two flagella, both attached near the same end of thecell. When ponds and lakes dry out in summer, goldenalgae form resistant cysts. Viable cells emerge from thesecysts when wetter conditions recur in the fall.
704 Part IX Viruses and Simple Organisms
FIGURE 35.13Diatoms (Chrysophyta). Several different centric (radially symmetrical) diatoms.
rav31208_ch35.qxd 7/19/01 12:51 PM Page 704

Rhodophyta: The Red Algae
Along with green algae and brownalgae, red algae are the seaweeds wesee cast up along shores and onbeaches. Their characteristic colors re-sult from phycoerythrin, a type ofphycobilin pigment. Phycobilins areresponsible for the colors of thecyanobacteria. Chlorophyll a also oc-curs with the phycobilins in red algae,just as it does in cyanobacteria. Thesesimilarities with cyanobacteria make itlikely that the rhodophyta evolvedwhen their heterotrophic eukaryoticancestor developed an endosymbioticrelationship with a cyanobacteriawhich eventually gave rise to theirchloroplasts.
The great majority of the estimated4000 species of red algae occur in thesea, and almost all are multicellular.
Red algae have complex bodiesmade up of interwoven filaments ofcells. In the cell walls of many redalgae are sulfated polysaccharides suchas agar and carrageenan, which makethese algae important economically.Agar is used to make gel capsules, asmaterial for dental impressions, and asa base for cosmetics. It is also the basisof the laboratory media on which bac-teria, fungi, and other organisms areoften grown. In addition, agar is usedto prevent baked goods from dryingout, for rapid-setting jellies, and as atemporary preservative for meat andfish in warm regions. Carrageenan isused mainly to stabilize emulsionssuch as paints, cosmetics, and dairyproducts such as ice cream. In additionto these uses, red algae such as Por-phyra, called “nori,” are eaten and, inJapan, are even cultivated as a humanfood crop.
The life cycles of red algae are com-plex but usually involve an alternationof generations (sporic meiosis). Noneof the red algae have flagella or cilia atany stage in their life cycle, and theymay have descended directly from an-cestors that never had them, especiallyas the red algae also lack centrioles. To-gether with the fungi, which also lackflagella and centrioles, the red algaemay be one of the most ancient groupsof eukaryotes.
Phaeophyta: The Brown Algae
The phaeophyta, or brown algae, con-sist of about 1500 species of multicel-lular protists, almost exclusivelymarine. They are the most conspicu-ous seaweeds in many northernregions, dominating rocky shores al-most everywhere in temperate NorthAmerica. In habitats where largebrown algae known as kelps (orderLaminariales) occur abundantly in so-called kelp forests (figure 35.14), theyare responsible for most of the foodproduction through photosynthesis.Many kelps are conspicuously differ-entiated into flattened blades, stalks,and grasping basal portions that an-chor them to the rocks.
Among the larger brown algae aregenera such as Macrocystis, in whichsome individuals may reach 100 me-ters in length. The flattened blades ofthis kelp float out on the surface ofthe water, while the base is anchoredtens of meters below the surface. An-other ecologically important memberof this phylum is sargasso weed, Sar-gassum, which forms huge floatingmasses that dominate the vast Sar-gasso Sea, an area of the AtlanticOcean northeast of the Caribbean.The stalks of the larger brown algaeoften exhibit a complex internal dif-ferentiation of conducting tissuesanalogous to that of plants.
The life cycle of the brown algae ismarked by an alternation of genera-tions between a sporophyte and a ga-metophyte. The large individuals werecognize, such as the kelps, aresporophytes. The gametophytes areoften much smaller, filamentous indi-viduals, perhaps a few centimetersacross. Sporangia, which producehaploid, swimming spores after meio-sis, are formed on the sporophytes.These spores divide by mitosis, givingrise to individual gametophytes.There are two kinds of gametophytesin the kelps; one produces sperm, andthe other produces eggs. If sperm andeggs fuse, the resulting zygotes growinto the mature kelp sporophytes,provided that they reach a favorablesite.
Chapter 35 Protists 705
FIGURE 35.14Brown algae (Phaeophyta). The massive“groves” of giant kelp that occur inrelatively shallow water along the coasts ofthe world provide food and shelter formany different kinds of organisms.
rav31208_ch35.qxd 7/19/01 12:51 PM Page 705

Chlorophyta: The GreenAlgae
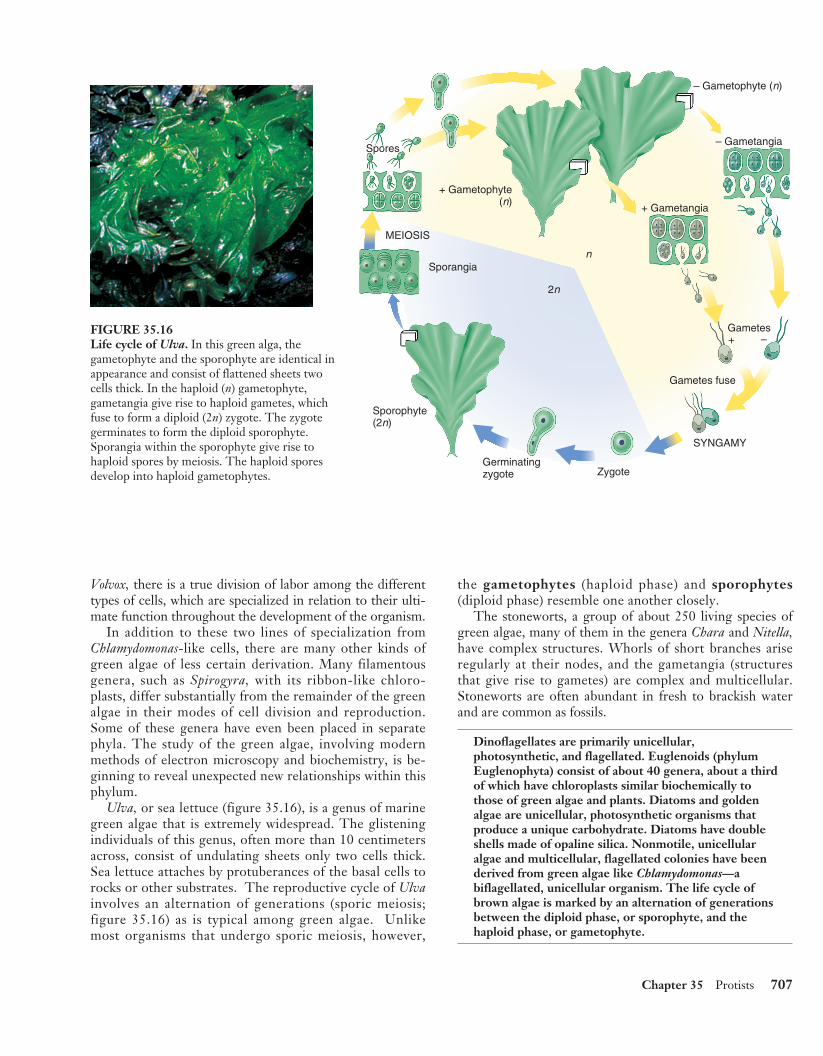
Green algae are an extremely variedgroup of more than 7000 species.The chlorophytes have an extensivefossil record dating back 900 millionyears. They are mostly aquatic, butsome are semiterrestrial in moistplaces, such as on tree trunks or insoil. Many are microscopic and uni-cellular, but some, such as sea let-tuce, Ulva (see figure 35.16), aretens of centimeters across and easilyvisible on rocks and pilings aroundthe coasts.
Green algae are of special inter-est, both because of their unusual di-versity and because the ancestors ofthe plant kingdom were clearly mul-ticellular green algae. Many featuresof modern green algae closely re-semble plants, especially theirchloroplasts which are biochemicallysimilar to those of the plants. Theycontain chlorophylls a and b, as wellas carotenoids. Green algae includea very wide array of both unicellularand multicellular organisms.
Among the unicellular greenalgae, Chlamydomonas (figure 35.15)is a well-known genus. Individualsare microscopic (usually less than 25micrometers long), green, rounded,and have two flagella at the anteriorend. They move rapidly in water by beating their flagella inopposite directions. Each individual has an eyespot, whichcontains about 100,000 molecules of rhodopsin, the samepigment employed in vertebrate eyes. Light received bythis eyespot is used by the alga to help direct its swimming.Most individuals of Chlamydomonas are haploid. Chlamy-domonas reproduces asexually (by cell division) as well assexually. In sexual reproduction, two haploid individualsfuse to form a four-flagellated zygote. The zygote ulti-mately enters a resting phase, called the zygospore, inwhich the flagella disappear and a tough protective coat isformed. Meiosis occurs at the end of this resting period andresults in the production of four haploid cells.
Chlamydomonas probably represents a primitive state forgreen algae and several lines of evolutionary specializationhave been derived from organisms like it. The first is theevolution of nonmotile, unicellular green algae. Chlamy-domonas is capable of retracting its flagella and settlingdown as an immobile unicellular organism if the ponds inwhich it lives dry out. Some common algae of soil and bark,such as Chlorella, are essentially like Chlamydomonas in this
trait, but do not have the ability to form flagella. Chlorella iswidespread in both fresh and salt water as well as soil and isonly known to reproduce asexually. Recently, Chlorella hasbeen widely investigated as a possible food source for hu-mans and other animals, and pilot farms have been estab-lished in Israel, the United States, Germany, and Japan.
Another major line of specialization from cells likeChlamydomonas concerns the formation of motile, colonialorganisms. In these genera of green algae, the Chlamy-domonas-like cells retain some of their individuality. Themost elaborate of these organisms is Volvox (see figure 35.1),a hollow sphere made up of a single layer of 500 to 60,000individual cells, each cell with two flagella. Only a smallnumber of the cells are reproductive. The colony has defi-nite anterior and posterior ends, and the flagella of all of thecells beat in such a way as to rotate the colony in a clockwisedirection as it moves forward through the water. The repro-ductive cells of Volvox are located mainly at the posteriorend of the colony. Some may divide asexually, bulge inward,and give rise to new colonies that initially remain within theparent colony. Others produce gametes. In some species of
706 Part IX Viruses and Simple Organisms
– Strain
– Strain
– Gamete
+ Gamete
SYNGAMY
MEIOSIS
Zygospore (diploid)
Asexualreproduction
+ Strain
+ Strainn
2nPairing ofpositive andnegativestrains
FIGURE 35.15Life cycle of Chlamydomonas (Chlorophyta). Individual cells of this microscopic,biflagellated alga, which are haploid, divide asexually, producing identical copies of themselves.At times, such haploid cells act as gametes—fusing, as shown in the lower right-hand side of thediagram, to produce a zygote. The zygote develops a thick, resistant wall, becoming azygospore; this is the only diploid cell in the entire life cycle. Within this diploid zygospore,meiosis takes place, ultimately resulting in the release of four haploid individuals. Because of thesegregation during meiosis, two of these individuals are called the (+) strain, the other two the(–) strain. Only + and – individuals are capable of mating with each other when syngamy doestake place, although both may divide asexually to reproduce themselves.
rav31208_ch35.qxd 7/19/01 12:51 PM Page 706
Volvox, there is a true division of labor among the differenttypes of cells, which are specialized in relation to their ulti-mate function throughout the development of the organism.
In addition to these two lines of specialization fromChlamydomonas-like cells, there are many other kinds ofgreen algae of less certain derivation. Many filamentousgenera, such as Spirogyra, with its ribbon-like chloro-plasts, differ substantially from the remainder of the greenalgae in their modes of cell division and reproduction.Some of these genera have even been placed in separatephyla. The study of the green algae, involving modernmethods of electron microscopy and biochemistry, is be-ginning to reveal unexpected new relationships within thisphylum.
Ulva, or sea lettuce (figure 35.16), is a genus of marinegreen algae that is extremely widespread. The glisteningindividuals of this genus, often more than 10 centimetersacross, consist of undulating sheets only two cells thick.Sea lettuce attaches by protuberances of the basal cells torocks or other substrates. The reproductive cycle of Ulvainvolves an alternation of generations (sporic meiosis;figure 35.16) as is typical among green algae. Unlikemost organisms that undergo sporic meiosis, however,
the gametophytes (haploid phase) and sporophytes(diploid phase) resemble one another closely.
The stoneworts, a group of about 250 living species ofgreen algae, many of them in the genera Chara and Nitella,have complex structures. Whorls of short branches ariseregularly at their nodes, and the gametangia (structuresthat give rise to gametes) are complex and multicellular.Stoneworts are often abundant in fresh to brackish waterand are common as fossils.
Dinoflagellates are primarily unicellular,photosynthetic, and flagellated. Euglenoids (phylumEuglenophyta) consist of about 40 genera, about a thirdof which have chloroplasts similar biochemically tothose of green algae and plants. Diatoms and goldenalgae are unicellular, photosynthetic organisms thatproduce a unique carbohydrate. Diatoms have doubleshells made of opaline silica. Nonmotile, unicellularalgae and multicellular, flagellated colonies have beenderived from green algae like Chlamydomonas—abiflagellated, unicellular organism. The life cycle ofbrown algae is marked by an alternation of generationsbetween the diploid phase, or sporophyte, and thehaploid phase, or gametophyte.
Chapter 35 Protists 707
Spores
MEIOSIS
Sporangia
Sporophyte(2n)
Germinatingzygote Zygote
SYNGAMY
Gametes+ –
+ Gametangia
+ Gametophyte(n)
– Gametangia
– Gametophyte (n)
2n
n
Gametes fuse
FIGURE 35.16Life cycle of Ulva. In this green alga, thegametophyte and the sporophyte are identical inappearance and consist of flattened sheets twocells thick. In the haploid (n) gametophyte,gametangia give rise to haploid gametes, whichfuse to form a diploid (2n) zygote. The zygotegerminates to form the diploid sporophyte.Sporangia within the sporophyte give rise tohaploid spores by meiosis. The haploid sporesdevelop into haploid gametophytes.
rav31208_ch35.qxd 7/19/01 12:51 PM Page 707


















