ORIENTATION AND SURVIVAL OF HATCHLINGS AND REPRODUCTIVE...
91
( ( ( ORIENTATION AND SURVIVAL OF HATCHLINGS AND REPRODUCTIVE ECOLOGY OF THE COMMON SNAPPING TURTLE (Chelydra serpentina) IN SOUTHERN QUEBEC by Candace Robinson . A Thesis submltted to the Faeulty of Graduate Studles and Researeh of MeGlll University in partial fulfillment of the requlrements for the degree of Master of Science Wlldllfe Resoutees, Department of Renewable Resourees, Macdonald College of McGIII University, Montreal. Quebee, Canada. e CANDACE ROBINSON September 1989
Transcript of ORIENTATION AND SURVIVAL OF HATCHLINGS AND REPRODUCTIVE...

(
(
(
ORIENTATION AND SURVIVAL OF HATCHLINGS AND REPRODUCTIVE
ECOLOGY OF THE COMMON SNAPPING TURTLE
(Chelydra serpentina)
IN SOUTHERN QUEBEC
by
Candace Robinson
. A Thesis submltted to the Faeulty of Graduate Studles
and Researeh of MeGlll University in partial
fulfillment of the requlrements for
the degree of Master of Science
Wlldllfe Resoutees,
Department of Renewable Resourees,
Macdonald College of McGIII University,
Montreal. Quebee, Canada.
e CANDACE ROBINSON September 1989

(
(
ABSTRACT
The o~Ientatlon and survlval of neonate enapping
tu~tles (CheJvdca secpentIna) were studled ln 1986 and
1987. Orientation mechanisms were tested ln an outdoo~
parabo1ic arena. These experlments suggested that
positIve geotaxis was a domInant eue direeting neonate
snapp1ng turtles to water.
Hatch Il ng surv 1 va 1 and movement from nest to
wate~ we~e studied ln Calumet, Ouebec uslng Isotope
tagglng.
A hlgh1y slgnlficant number of hatchJIngs (95%)
released on sJoped nest sites wer~ positiveJy
geotactlc. Hatchllngs released on leveJ ground moved ln
random dIrections.
Eighty-two percent of hatchllngs from nests
withln 18 m of the shorel In~ we~e successfuJ ln
reachlng water. IncIdence of mortallty was greatest for
hatchl1ngs from nests between 121 and 165 m from water.
Morta11ty resulted from vehlcles and p~edatlon by
bullfrogs (&ànà katesbieana) and meadow voles (Mlcrotus
~...cnsy 1 yan 1 eus) .
Female carapace length was posItlvely correlated
wlth clutch size and mean egg dlameter. Egg dlameters
were posltlvely correlated wlth hatchllng welghts.
II

o C~utch slzes of neete more than 100 m from water w~re
larger than thoS6 closer to water •
........
-l
III

(
( "
.~
ABREGE
L'o~ientatlon et la survie d~s to~tues
serpentInes (Chelydra serpentIijj) ap~ès l'~closIon,
furent étudIées en 1986 et 1987. Les méchanismes
d'orientatIon ont été étudIés dans une arène
extérieure. Le mouvement des to~tue~ dans cette arène
parabolique semblait indiquer que la géotaxie positive
~tait un facteur domInant dans l'orientation des Jeunes
tQrtues ve~s l'eau.
La survie et les déplacements des Jeunes tortues
du nid ~ l'eau ont été examinés à Calumet, Québec,
chaque Individu étant marqué à l'aide d'un Isotope
l"adloac. tif.
Quatre-vingt quinze pourcent des Jeunes tortues
sorpentines qui ont emergé des nids sur les sites
inclln~s ont montré une géotaxie positIve. Les tortues
qul ont emergé des nids en te~~alns plats se sont
dlspe~sées sans direction privilégiée.
Quatre-vingt deux pourcent des tortues qul ont
emergé des nlds à moins de 18 m de l'eau ont atteInt
l'eau avec succès. Le taux de mortallté fut plus grand
pour les tortues qui ont emergé des nids situés l 121
et 165 m de l'eau. Les causes de mo~tâl1té furent les
impacts causés par des véhIcules et la prédation par

o les grenoull les d/Am~rlque <BADA çatesbleoQA) et les
campagnols (Mlcrotus pennsylvanlcus).
Une corrélation positive fut établie entre la
taIlle de la couvée, la diamètre moyen des oeufs, et la
longeur de la carapace de 10 femelle. Une corrélation
positive fut aussi trouv~e entre les diamètres des
oeufs et le poids des Jeunes tortues. Les nids situés
~ plus de 100 m de l'eau contenaient plus d'oeufs que
ceux Eltués près de l'eau.

(
(
------------------~---------------------
TABLE OF CONTENTS
ACXNOWLEDGEMENTS .......... " .................... . VIII
LI ST OF T1\BLES ..........••....•............ f'I • • •• IX
LIST OF FIGURES •.•••.•..••....•••..• f, ••••••••••• X
PREFACE .......................••................ 1
REGULATION OF THESIS PRESENTATION ••••••••••••••• 3
SECTION 1. ORIENTATION eUES USED BY
NEONATE SNAPPING TURTLES
CCbeJydra serpentlna) ••••••••••••.•• 4
NOTE ••••••.••••••••..•••••••••••••.•••••••••• 5
LITERATURE CITED ......•.....•......•.•....•.• 11
FIGURE •.•..•••.••.....•..•••••••••••••••••••• 15
CONNECTING STATEMENT .•••........ ~ . . . . . . . . . . .. 16
SECTION 2. ORIENTATION AND SURVIVAL OF
HATCHLING SNAPPING TURTLES
CCbelydra serpentlna)
AS THEY HOVE FROM THE NEST
TC WATER •••••••••••••••••••••••••••• 17
ABSTRACT •••••.•••.•.••••••••••••••••••••••••• 1 e 1 NTRODUCT 1 ON ••••••••••••••••••••••••••••••••• 20
MATERIALS AND HETHODS •••..••••••••••••• ~ ••••• 23
RESULTa .••.••••••••..••••.••••••••.•••••••••• 27
DISCUSSION ..•.......••••...••.••••••••.•.•.•• 33
LITERATURE CITED •••...••...•••••••••••••••••• 40
VI

-~ ........
-------------------------
TABLES •..•••••••••••.•••••.••••••.••••••••••. 45
FIGURES •..•••••..•..••••..•.•••••.••••.•.•••• 47
CONNECTING STATEMENT ••••..••••••••••.•••••••• 57
SECTION 3~ SOME ASPECTS OF REPRODUCTIVE
ECOLOGY IN RELATION TO NEST DISTANCE
TO WATER OF A NORTHERN POPULATION
OF THE COMMON SNAPPING TURTLE
(Chelydra aecpentlnA) ••••••••••••••••• 58
ABSTRACT •••••••• '............................ 59
1 tlTRODUCT ION ..•.•.....•......•...•••.•....... 61
MATER 1 A LS AND METHOnS •••.•.•••••.••••••••••.• 63
RESULTS , .••• ,................................ 65
DI SCUSS 1 ON ••••••••••••••••••••••••••••••••••• 67
LITERATURE CITED ••...•••....••...••.•••.•..•• 71
FI GURES •..•••••••••.•••••..••••..••••••••••.• 74
GENERAL CONCLUS ION •..•••....••..••..•••.....• 79
VII

c
c
ACKNOWLEDGEMENTS
1 wou 1 d II ke to express my grat 1 tude and
apprec 1 at 1 on ta ,11 those who he 1 ped and adv 1 sed me
throughou t th 1 s proJ ec t • 1 wou 1 d II ke t 0 thank my
thesls dlrector, Dr. J.R. Bider, for hls support and
for edl t l ng the rough draf ts and fi na 1 manuscr 1 pts. 1
am also grateful to hlm and Jean Philippe Auvray for
construction of the parabollc test arena.
Thanks go to my frlend and col1e:ague, Monica
Borobl a, for her encouragement and construct 1 ve
commente on the manuecrlpts.
Michel Richard of the Centre Ecologique de
Harrlngton (C.I.P. Inc.> most klndly granted me access
to the Calumet ml 11 site. Harold and Mary Dzlelak,
Regi na 1 d and Hea ther Schef fie 1 d and Den 1 e Champagne
also generously allowed me access to thelr propertles.
1 thank Simon Nadeau for hie assistance ln
translatlng the abstract lnto french. 1 would also
llke to thank Robert Blssonnette for hie constant
support and assistance.
The Natura 1 Sc 1 ence and Engl neer 1 ng Research
Councll of Canada provlded fundlng for thls research.
VIII

o LIST OF TABLES
SECTION 2
TABLE 1. SUMMARY OF FATE OF PROTECTED
NESTS AND EGGS AT CALUMET, QUEBEC~ •••. 45
TABLE 2. THE FATE OF HATCHLING
Chelvdra serpentlna FROM NESTS
LOCATED WITHIN 18 M AND BETWEEN 122 .
AND 167 M FROM THE WATER .............. 46
.......
IX

(~
(
, ___ =_ .. _ .. ~ JI Il''.WW -sv.
L,dT OF FIGURES
SECTION 1.
FIGURE 1. H~TCHLING DISPERSAL PATTERNS FROM
SECTION 2.
FIGURE 1.
FIGURE 2a.
FIGURE 2b.
FIGURE 2c.
FIGURE 3.
THE 4 RELEASE POINTS WITHIN THE
PARABOLIC ARENA ..........•........••• 15
LOCATION OF THE STUDY AREA AND
NESTING SITES IN RELATION TO THE
OTTAWA RIVER AT CALUMET, QUEBEC ..... . 47
HATCHLING DISPERSAL PATTERNS
FROM SLOPED NEST SITES LOCATED
BETWEEN 122 AND 167 M FROM WATER .... . 49
HATCHLING DISPERSAL PATTERNS
FROM 2 NESTS LOCATED ON LEVEL
GROUND ••••••••••••••••••••••••••••••• 51
HATCHLING DISPERSAL PATTERN
FROM SLOPED NEST SITES
LOCATED WITHIN 18 M OF WATER ........ . 53
THE RELATIONSHIP BETWEEN THE
ANGLE OF HATCHLING DISPERSAL
FROM THE NEST AND THE

SECTION 3.
FIGURE 1. THE RELATIONSHIP BETWEEN CLUTCH
SIZE AND FEMALE CARAPACE LENGTH •..... 73
FIGURE 2. THE RELATIONSHIP BETWEEN MEAN EGG
DIAMETER AND FEMALE CARAPACE LENGTH ... 75
FIGURE 3. THE FREQUENCY DISTRIBUTION OF
CLUTCH SIZE FOR NESTS LOCATED
< 100 M FROM WATER AND NESTS
LOCATED> 100 M FROM WATER .•••••..••• 77

(
(~
( '" ,~
~---~-~------ ------------,-----------
PREFACE
The common snapping turtle (Chelydra serpentina)
ls widely distrlbuted ln many permanent freshwater
habitats ln North America. Whlle many aspects of the
basIc blology of thla species have been studied,
knowledge of the orIentation and survival of hatchlings
as they emerge from the nee:t and move to wa ter i s
limited. In addition, whlle the characterlstlcs of nest
al tes have been frequent 1 y descr ibed, the ef fect of
nest site selection on reproductive success ia poorly
understood. The primary objectives of thls study
were:l) to examine the orientatIon cues used by neonate
snapplng turtles ln a setting where environmental
factors could be controlled; 2) to Investlgate the
orIentation and aurvival of neonate snapping turtles in
the field, as they travel trom the nest to water; and
3) to gain a better understanding of the reproductive
strategies of this species by examlnlng some female
reproductive characterlstics as they relate to nest
distance from water.
In 1986 and 1987 field data were collected on
the extent of nest failure, timing and patterns of
hatchling emeroence, hatchllng movement and
orientation and hatchling mortality. Radioactive
1

o Isotopes we~e used to tag hatchllngs ln o~de~ to follow
thel~ movements f~om the nest to wate~. Most data on
female size, clutch slze, egg slze, hatchllng size and
nest dIstance to wate~ were eollected ln 1987.
As pe~mItted by the Faculty of G~aduate Studles,
thls thesls contains the texts of th~ee manusc~lpts to
be submltted to Jou~nals fo~ publication. The flrst,
presented in Section 1, deals wlth the o~Ientatlon eues
used by neonate snapplng tu~tles ln an outdoor
parabol:~ arena. The second, p~esented ln Section 2,
examInes the o~lentation and su~vlval of hatehling
snapp 1 ng tu~t 1 es 1 n the fie 1 d as they move from the
nest to wate~.- The thl~d, p~esented ln Section 3,
examines sorne aspects of ~eproductlve ecol ogy ,
Includlng female slze and clutch size, as they ~elate
to nest dIstance f~om wate~. The flret manuscript will
be sent to the Jou~nal of Herpetol ogy , the second to
Copela, and the thlrc! to Herpetologlca, with Dr J.R.
Bider as eo-autho~. The style of eaeh manuscrlpt le
that ol the Journal to whleh It will be submltted.
Data collection and analysls were ear~led out
1 ndependant 1 y by the ~uthor. Re f erences. t ab 1 es and
figures appea~ after the manuec~lpt ln whlch they a~e
clted.
2

(
(
~~~-- ------------.------------------------------------............ ..
TH!SIS OFFICE STATEMENT
The candidate has the option, subJect to the
approva 1 of the Deptartment, of 1 nc 1 udl ng as part -of
the thesle the text of an original paper, or paper;e,
sultable for submlsslon to learned Journals for
publication. In thle case the thesls must still
conform to aIl other; requlrements explalned ln thle
document, and addltlonal materlal, (e.g. experlmental
data, detalle of equlpment and experimental design) May
need to be provlded. In Any case abstract, full
Introductlon and cnncluslon must be Included, and where
more than one manuecr;lpt appear;e, connectlng texte and
common abstract Introduction and conclusions ar;e
requlred. A mere co] 1 ect Ion of manuscr 1 pts 1 s not
ac'.:eptable~ nor can reprints of publlshed papers be
accepted.
Whlie the Inclusion of manuscrlpts co-author;ed
by the Candidate and others Is not prohlbl ted for a
test perlod, the candidate Is warned to make an
expllclt statement on who contrlbuted to such work and
to what extent, and supervisors and others will have to
bear wltness to the accuracy of such clalms befor;e the
oral commlttee. It should also be noted that the task
of the External Examiner 18 much more dlfflcult ln such
cases.
3

-----------------
o
SECTION 1
ORIENTATION CUES USED BY NEONATE SNAPPING TURTLES
• (Che)vdra lerpentlna)
-,.,,) . , , ' .....
* Note for submllslon to the Journal of Herpetology

(
(
-
The maJorlty of etudies on the orientation of
hatch Il ng turt 1 es from the nest to water have
concentrat~d on marine specles. Research on the
orientatIon mechanlsm~ of the loggerhead turtle
(Caretta caretta) and green turtle (Chelonla rnvdae)
have focussed prlmarlly on vlsual eues InvolvIn~ llght
Intenslty, color and horIzon effeets (Hooker, 1908;
Parker, 1922; Daniel and SmIth, 1947; Caldwell and
Ca 1 dwe 11, 1962; MeFar 1 ane, 1962; Ehrenfe 1 d and Carr,
1957; Mrosovsky and Carr, 1967; Ehrenfeld, 1968:
Mrosovsky and Shettleworth, 1968; Mrosovsky, 1972).
OrIentatIon to magnetle fIelds has been ruled out as a
meehanlsm ln loggerhead hatehllngs (Parker, 1922).
Reports on the effeets of sI ope on hateh Il ng
orIentation are eontradietory. Parker (1922) found
loggerhead hatehllngs to be posltlvely geotaetle.
Other studies of sea turtles lndleated that lmpalrment
of vIs ion resu 1 t sIn a loss of or 1 en t a t Ion ab 1 II t Y
regardless of slope (Caldwell and Caldwell, 1962;
Ehre~feld and Carr, 1967; Ehrenfeld, 1968).
Terrestrlal orientation in freshwater hatehllnge
may oeeur over greater dIstances and ln the absence of
simple eues sueh as lhose assoelated wlth beaeh
topography (Ehrenfeld, 1979). Noble and Breslau (1938)
demonstrated that snapplng turtle hatchllngs exhlblted
negative geotaxis ln the dark. In a study of 3
5

freshwater speeles, Anderson (1958) suggested that
orIentation proeesses might involve a negative response
to dark masses. A positive response to areas of bright
li ght, the posi t i r.:>n of the sun and hum! di ty grad! ents
have also been proposed as important eues for snapplr;
turtle hatehllngs, following laboratory tests (Noble
and B~eslau, 1938).
Five possible orIentation meehanisms have been
dlseu9sed ln the 1 Iterature: (1) visual eues, e.g.: the
slght of water, horizon effeets and Jight lntenslty,
(2) olfaetory eues, (3) orientation to magnet'e fields,
(4) a response to moi sture gradients, and (S) geotaxls.
In view of the mlx~d opinions, and prelimlnary tests on
a sma11 slope board whieh indleated that snapping
turt1es might be positively geotaetie, we sought to
study hatehling movement in an open (outdoor) arena
where at least geotaxis, orientatIon to magnetie flelQs
and the role of sorne visual eues might be evaluated.
The tests were eondueted ln a way that elimlnated the
vlsual eues associated with an open or bJoeked horizon
wlthout blIndlr.g the animaIs. In this !!I~ .. udy the term
positive ~eotaxls i9 defined as moving down a gravitv
gradient. The defininltlon of this term has
oecasionaJly been eonfused ln the literature.
Twenty snapplng turtle hatehllngs were observed
ln an outdoor arena loeated in the Macdonald College
wil dli fe area at Ste-Anne-de-Be 1 J evue! Gluebee. The

(
(~
(
parabolle clreular arena had a dlameter of 12 m and was
1 m deep (Flg.l). The maximum slope wae 100 whlle at
the base of the parabola, the slope decreaeed ta 20 to
form a relatlvely fIat area, 50 cm wlde. A fIat area
30 cm w 1 de ene 1 rc 1 ed the upper edge of the parabo 1 a.
The area was enclosed by a galvanlzed steel wall 0.5 m
ln helght. A chlld's plastic ewlrrmlng pool, 1.5 m ln
dlameter was eunk Into the center of the parabola. The
pool was filled with pond water and eorne aquatlc
veget:at Ion. Cl umps of 10w, herbaceous growth covered
approxlmately 60% of the arena.
Snapplng turtle eggs frorn 2 nests were collected
at Calumet, Quebec CLat. 450 38' Long. 740 39')
(Robinson and Bider. 1988) and were Incubated Indoors.
Ten emergl ng hateh II ngs were ee 1 eeted at randorn from
eaeh and an Identification number was palnted on the
baek of each hatchllng. Groups of 5 hatchJ Inge were
then reburled ln molst vermlcullte ln 4 Inch (10.16cm)
flower pots and the pots were burled Into the rlm ln
the aide of the parabola 4 m from the pool ln each of
tha 4 cardinal directions. The dIstance and direction
of each hatchling from the release poInt were recorded
every 5 min. untll It reached the water. Experimente
were conducted ln the late afternoon on clear sunny
days ln early September wlth temperatures ranglng
between 18-220 C.
7

o
.. "
Upon emergl ng frcm the pote a 11 hatch li nge
paused and appeared to scan thelr surroundlngs. Each
ha tch 1 1 ng moved 1 ndependen t 1 Y of the others. AlI of
the hatchllngs moved downhll1 trom the pot towards the
pool, pauslng occaslonally. Thlrteen reached the water
dlrectly wlthout bypasslng the pool. Seven hatchllngs
passed wlthln 0.3 m of the pool edge, and contlnued up
the other sI de. Upon reach 1 ng the- ou ter edge of the
arena they elther turned around and contlnued down to
the pool, or they followed the arena wall for several
meters and then descended to the pool (Fig.l).
AlI hatchllngs eventually reached the water, and
once 1 n the poo 1 tended to rema 1 n submerged unt 1 1
removed hours 1 ater. The maximum and mInimum tlmes
taken to reach water were 161 and 6 min., respectlvely.
The mean t i me t aken by those wh i ch en tered the POv l
dlrectly was 13 mins (n=13; 5.E.=2.05) and the Mean
tlme taken by those whlch passed the pool was 72 mins
(n=7; S.E.=21.11). The average speed of aIl hatchlings
was 0.39 ml mIn. (S.E.=0.03).
Slnce aIl hatchllngs went downhl11 from the
release pOints and 35% did not go dlrectly to water, a
good deal of Inference may be made wlth regard to the 5
orientatIon mechanlsms mentloned in the 11terature.
The low parabo 1 a wa II created a constant,
unlnterrupted horizon, thus the horIzon was not bJocked
8

(
(
nor was there a brlght open horizon assoclated wlth the
water. Neverthe 1 ess, aIl of the hatch II ngs r-eaehed
water. Whl1e these results do nct rule out the use of
the vlsual cues of blocked or open horlz~ns when they
oceur ln natural situations, they do Indleate that
hatehl1ngs can orient to water ln thelr absence. The
direct vlsual eue uf seelng the water had no bearlng on
orientation, sinee 7 of the 20 hatchllngs passed wlthln
a few centlmeters (and ln clear vlew) of the pool, yet
cont 1 nued up the other ~1 de of the parabol a wl thout
stopplng.
The 01 factory cues of water appeared to have
llttle effect on orientation, given the proxlmlty of
the 7 hatehllngs that bypassed the pool. This supports
the flndlngs of Noble and Breslau (1938) who noted that
snapplng turtle hatchl1ngs exhlblted no slgniflcant
r-esponse to the odor of river water or food.
Desplte the dlfferent release tlmes and
locations of the 4 groups tested, aIl hatchllngs moved
downh III 1 n the dl rect Ion of the poo 1 . These resu 1 te
suggest that snapplng turt'e hatehllngs are not uslng
orientation to magnetic fields as a cue in thelr
approach to water , nor Is thelr movement affected by
the position of the sun. S!nce the hatchllngs
orlglnated from 2 nests loeated approxlmately 100 km
9

o
[}
from the test si te, or lentat ion to water cannot be
related to any Innate knowledge of the nestlng area.
Nob Je and Bres 1 au (1938) noted tha t enapp 1 ng
turtle hatchllngs dlsplayed a tendency to move towards
an area of greater huml dl ty. In thls experlment the
molsture gradient waS s11mlnated and yet hatchl1ngs
orlented successfully to water.
Noble and Breslau (1938) found that hatchJlngs
of thls species showed a marked tendency to move uphll1
ln the dark, a behavlor that mlght facllitate thelr
escape from the nest. We dld not test hatchl1ngs ln
the dark, however ln the dayllght aIl hatchllngs moved .
downhlll from the release poInt. In the re 1 at 1 ve 1 y
fIat areas at the base and around the upper rlm of the
parabola hatchl1ngs frequently changed direction,
moving away from the water.
In summary. our data are not consistent wlth the
use of visual eues, olfactory eues, orientation to
magnetlc fields, or molsture gradients as ori~lltatlon
mechaniems. Gravit y ln dayllght appears to be the
dominant eue dlrectlng snapplng turtles downhll1 to
water.
10

(~
(~
(
LITERATURE CITED
Anderson, P.K. 1958. The photlc responses and
water-approach behavlour of hatchllng turtles.
Copela 1958: 211-215
Caldwell, M.C. and O.K. Caldwell. 1962. Factors ln the
abillty of the northeastern PacIfie green turtle to
orient toward the sea fram the land, a possible
coordlnate ln long range navigation. Contrlb. Sel.
60: 1-27.
Daniel, R.S. and K.U. SmIth. 1947. The sea-approach
behavlor of the neonate loggerhead turtle (Caretta
caretta). J. Comp. Physlol. Psychol.40: 413-420.
Ehrenfeld, D.W. 1968. The role of vIsIon ln the
sea-flndlng orientation of the green turtle
(Chelonia mydas) 2. OrIentation mechanlsm and
range of spectral sensltlvity. Anim. Behav. 16:
281-287.
Ehrenfeld, D.W. 1979. Behavlor assoclated wlth nestlng.
In: Turtles: perspectives and research. Edlted by M.
Harless and H. Morlock. John Wlley and Sons. New
York. pp.417-434.
Ehrenfeld. D.W. and A. Carro 1967. The role of vision
11

o ln the sea-flndlng orientation of the green turtle
(Che)QclA mydas). Anlm. Behav. 15:25-36.
Hooker,D. 1908. Prellmlnary observations on the
behavlor of seme newly hatched loggerhead turtles
(Tha)asso~eJya caretta). Yearbook CarnegIe Inst.
Wash. 6: 111-112.
McFarlane, R.W. 1962. Dlsorl~ntatlon of 10ggerhead
hatchllngs by artlflclaJ road 11ghtlng. Copela 1963:
153.
Hr.osovsky, N. 1972. The water-flndlng abllity of sea
turtles. Behavloral studles and physlologlcal
speculations. BraIn Behav. Evol.5: 202-225.
Hrosovsky, N. and A. Carr.1967. Preference for 1lght of
short wavelengths ln hatchllng green sea turtle
Cheloola mydas. tested on thelr natural nestlng
beaches. Behavlour 28: 217-231.
Hrosovsky, N. and S.J. Shettleworth. 1968. Wavelength
preferences and brIghtness cues ln the waterflndlng
behavlour of sea turtles. Behavlour 32: 211-257.
Noble, G.K. and A.M. Breslau. 1938. The senses Involved
ln the mIgration of young fresh-water turtle~ after
hatchlng. J. Camp. Psychol. 25: 175-193.
12

(
(
Parker, G.X. 1922. The crawllng of young )oggerhead
turtJes toward the sea. J. Exp. Zoo). 36: 323-331.
Robinson, C. and J.R. Blder.1988. Nestlng synchrony - A
stategy to decrease predation of snapplng turtle
CChelvdra serpeotlna) nests. J. Herp. 22:470-473.
13

·0
-- ---------- ---_ .. _--------------------
FIGURE 1. HATCHLING DISPERSAL PATTERNS FRCM
THE 4 RELEASE POINTS WITHIN THE
PARABOLIC ARENA
14
·--1 1

(
, - - - - - - -.6 m. - - - - - 1 l:m .... -
(
o 5 J ,
meters
R.P.-Release Point
o Pool

o
.....
CONNECTING STATEMENT
Section 1 descrlbed the orientation cues used by
hatchllng snapplng turtles ln an experlmental parabollc
arena.
Section 2 examines orientation, movement, and
survlval of hatchllng snapplng turtles ln the field •
16

(
(
(
SECTION 2
ORIENTATION AND SURVIVAL OF HATCHLING SNAPPING TURTLES
(Cbelydro serpentIna) AS THEY MOVE FROM THE NEST TO
WATER
* For submlsslon to Copela
17

o
~, .: ~' ...
ABSTRACT
The orIentation and survlval of hatchllng snapping
turtles was studied in 1986 and 1987 at Calumet.
Duebec. Seventy-f 1 ve percent of neste protected from
predators were partially successful wlth sorne
hatchllngs emerglng. while 25% falled completely.
Flfty-slx percent of eggs produced hatchllngs that
emerged successfully, 31% of embryos dled durlng
development and 12% of e~gs were Infertile.
The maJority of clutches (80%) emerged ln
September. The average number of days' trom egg
deposltlon to emergence was 96. Nlnety-flve percent of
hatchllngs emerged between 0700-1800 hours .
One hundred and flfty-slx hatchllngs (95%) whlch
emerged from sloped nest sItes moved downhill. There
was a slgnlflcant negatlve Ilnear relatlonshlp between
the angle of hatchllng dispersal from the nest and the
slope of the ne~t site. Vegetation dld not affect the
dl rect Ion of hatch II ng movement. Hatch Il ngs re 1 eased
on level ground dlspersed randomly from the nest.
Elghty-two percent of hatchlings frcm nests located
wlthln 18 m of the shorellne were successful ln
reachlng water. Mortallty was hlghest for hatchllngs
travelling from distant nests and was caused by
18

(
c
preda t Ion and veh 1 c 1 es • !WO hatchllngs were preyed
upon by bullfrogs (RAnA catesbelAna) At the water's
edge and 8 hatchllngs were preyed upon by meadow voles
(Mlccotys pennsylYAnlcys) ln a grassy fIeld.
19

o 1 NTRODUCTI ON
The common snapplng turtle (ChelydrA .erpentina)
ls a long Ilved (Gibbons, 1987) and wldely dlstrlbuted
specles, ranglng Into 80uthern Canada west to the
Rockles and south to the Gulf of Mexico (Conant, 1975).
It occure ln many permanent freehwater habitats (Ernst
and Barbour, 1972; Conant, 1975;) and plays an
Important role ln marsh ecology (Hammer, 1969). Many
aspects of snapplng turtle blology have been studled,
lncludlng: adult movement and migration (Moslmann and
Bider, 1960; Hammer, 1969; Obbard And Brooks, 1980;
Obbard and BrC1oks, 1981b; Ga 1 bra 1 th et al., 1987) ,
reproductive ecology (Yntema., 1970: White and Murphy,
1973; Congdon et alo, 1987), neetlng ecology (Hamilton,
1940, Hammer, 1969; Wllhoft et al., 1979; Petokas and
Alexander, 1980; Obbard and Brooks, 1981A~ Congdon et
aJ., 1987; Robinson and BIder, 1988), population
denslty (Major, 1975; Galbraith et al., 1988), and
habitat preferences (Sexton, 1958; Froese, 1978; Obbard
and Brooks, 1981b). Few etudies, however, have
Investlgated the behavlor of turtle hatchllngs as the y
emerge fram the nest and make thelr way to water
(Burger, 1976). Whlle predation on nests appeaes to be
a major determinant of neet survlval (Ernst and
Barbou~, 1972; Tlnkle et al., 1981; Robinson and Bider,
20

(
(
(
1988) other factors that mlght Influence nest success
and hatchllng survlval followlng emergence have
recelved Infrequent evaluatlon. Hatchllng emergence
was 1 ess than 20 " for undl sturbed nests 1 n South
Dakota (Hanmer, 1969) and an e.verage of 4.14 eggs or
embryos dled ln nests that esc4ped predation ln
MiChigan (Congdon et al., 1987). Obbard and Brooks
(1981a) have examlned the fate of overwintered clutches
ln an OntarIo populatIon and reported that of 129
clutches that dld not emerge ln the fall and escaped
predatIon, only one overwlntered successful1y.
Research examl n 1 ng ha tch 11 ng movement and
orientation to water has focussed prlmarlly on marine
specles. Knowledge of thls toplc concernlng freshwater
hatchllngs Is rudlmentary (Ehrenfeld, 1979). Noble and
Breslau (1938) tested a number of potentlal
water-flndlng cues under laboratory conditIons, us!ng
snapplng turtle, stlnkpot (SternotherYs odoratus), and
palnted turtle (Chrvsemys plcta) hatchllngs and
demonstrated a positive response to are as of brlght
Illumination and humidity gradIents. Two signiflcant
field studies ln thls area are those of Burger (1976)
on the diamondback terrapln (Malaclemys teerApln~. and
Anderson (1958), on the smooth sof tshe 1 1 (Trignyx
mut 1 eus). the Al abama map turt 1 e (GroptemYI!!I PY] chea),
and the ri nged eawback (Graptemys oey] 1 fera) • Burger
21

o
0' ... .}
( 1976), examl ned the emergence pat terne of hatch Il ngs
by followlng thelr tracks on the sand dunes. On sloped
sItes hatchlings moved downhlll, however the location
of the nearest vegetatIon was a stronger orientational
st Imu 1 us than 1 nc li ne. Anderson ( 1958) tested
or lentat Ion mechan 1 SlnS w i th mazes and by observ 1 ng
hatch 11 ng tracks 1 n the sand. The resu 1 ts of th 1 s
study suggested that hatchllngs exhiblted a negatlve
response to dat"k masses. These studl es re Il ed on the
observation of tracks or the direct and constant
observa t i on of each hatch 1 1 ng. Ne 1 ther study reported
on the success of hatchllngs in reaching the water, nor
on the type or extent of pr-edation suffered after
leavlng the nest site •
The object 1 ves of the present study were to
Investlgate: (1) the extent of nest fallures when
predators are excluded: (2) the t!mlng and patter-ns of
hatchl1ng emer-gence; (3) the behavlor, mo"ement, and
orientation of hatchllngs over differ-ent terraIn and at
va['lous dlst~nces from the water; (4) the extent and
causes of mor-ta lit Y as hatch 11 ngs make the 1 [' way to
wate['. ThIs resea['ch Is the fi ['st IntensIve field
study of the o['lentatlon and sur-vlval of snapplng
tu['t 1 e ha tch 1 1 ngs as they move f['om the nest to water
and fIl 19 a gap ln our knowledge of the Ilfe hlstory of
thls specles.
22

(
(
(
MATERIALS AND HETHOnS
This study was carrled out at Calumet, OUebec
(1 at. 45 38' , long. 74 39' >. The si te Includes a
narrow, T shaped, malnly fIat plece of land (m111 site)
approxlmate1y 1 km ln length, that extends eouthward
Into the Ottawa RIver (Fig. 1>. To the north of the
mlll site the land slopes upward gradually. This area
extends east about 1 kl10meter to the flret houses ln
the vIllage. The land near the rallroad tracke Is sandy
and sparee 1 y covered w 1 th grasses, wh II e most of the
sI ope down t 0 the river cone 1 st s of ml xed hardwood
forest, endlng wlth a dense band of wll10w (Sallx mL.,)
and aIder (Alnys ~) at the water's edge. Much of
the old mlll sIte le covered wlth woody and herbaceous
pioneer vegetation. &,oad fIat areas of woodchlps wlth
little or no vegetation except lichen are present ln
the north-central area.
Fleld work wae conducted from 15 May to 5
Oc;-tober 1 n 1986 and 1987. The site was v 1 si ted da Il y
untll nestlng began, after which It was vlslted at
leaet twlce dally untll the end of the nesting season.
Each year, 10 nests were located and protected
tram predators uslng heavy metal screens that measured
60 by 90cm. The nest sites were se1ected on the basle
of thelr substrates, slopes and dIstance fram the
23

,0 river. Nest sites have been found either close to the
water « lBm) or far from lt (121-165m) and the sample
reflects the relatIve proportIon of nests at each sIte.
Where nests cou 1 d not be adeq'.Jate 1 y protected from
vehlcles, they were collected and lncubated Indoors at
240 C, ln flowerpots fl11ed wlth vermlcullte.
After the nestlng season, the protected nests
were checked every second day. Durlng the second and
th 1 rd weeks of August the screens were rep 1 aced wl th
cyllndrlcal baskets (30 cm dlameter, 16 cm helght)
constructed of 0.5 inch (1.27cm) rnesh hardware clotho
Each cyllnder was burled to a depth of 8-10 cm ln order
to contaln emerglng hatchl1ngs. Followlng the f lrst
si gn of emergence, each nest was checked da Il yin the
mornlng and at nlght. The date and tlrne of day when
hatchllngs emerged were recorded and they were rernoved
from the enc 1 osure. Nests were aIl excavated dur 1 ng
the fi rst week of October and 2 unhatched eggs from
each were dlssected. If these eggs were found to
con ta 1 n lIve embryos the nest was rebur 1 ed and 1 ef t
Intact untll the following sprlng. When no live
embryos were found, the remalnlng eggs were dlssected.
The number of i nfert Il e eggs, dead embryos and dead
hatchllngs was recorded for each nest.
24

c
(~
(
AB many aB 10-15 individu.la were aelected at
random from each nest group and identification numbers
were palnted ente their carapaces. Snapplng turt 1 e
hatchl ings are very cryptic and ln thlck vegetatIon
they become virtually Invisible to the human observer.
For th 1 s reason ha t ch 1 1 ngs were a 1 se marked w!th the
radl oact 1 ve 1 sotope, Zn 65. The tag was prepared by
comblnlng a smal1 quantlty of Metal based palnt with
radIoactive ZnCI dry salts. The mixture was then
applled onto a strlp of plastic wrap. The strlp was
wrapped around a spool and small chIps were cut off as
needed. A chlp measurlng approxlmately 3 mm2
contalning 0.02 mlcrocurles of radIoactive material was
attached to the dorsal surface of a marginal scute on
each hatchling wi th acryllc adheslve. This procedure
took approxlmately 10 mIn durlng whlch tlme hatchlings
were handled as little as possible.
Marked hatch 1 i ngs were re 1 eased at the nest
site. The initial direction in whlch each was faced
for release was chosen at random by slldlng hatchllngs
out of a container; no partlcular directIon was
favoured. Every 10 minutes durlng the day each
hatchllng was located uslng a sclntilator counter,
(Ludlum 3 survey meter) wlth a 2 in (5.08cm) dlameter
prObe (mode 1 4410) • Slnce hatchllngs remalned
25

o
-; ~ -
re 1 at 1 ve 1 y 1 nact 1 ve at ni ght, they were on 1 y tracked
from 0700-1800 hrs. Wherever possible, hatchllngs were
monltored for 2 days after the y entered the water. The
detect Ion range of the counter was approx Imate 1 y 1 m
and ha tch Il ngs were f ound by sweep 1 ng the area 1 n an
1 ncreas 1 ng arc. Once il hatch! Ing was located, the
position was marked wlth a ~take and the distance and
dl~ectlen from the nest were recorded. AlI hatchl1ngs
were fo)) owed unt Il they el ther r~ache:d the water, were
kllled o~ were lest. Wheneve~ possible the causes of
mortallty were determlned. In 2 Instances, 3 Longworth
traps were set out ln an attempt to identlfy pr~dators
whlch had killed and carried hatchllngs underground.
26

(
(
(
RESULTS
Hest Survlval
Of a total of 20 nests protected from predators,
5 falled completely and none hatched aIl of the eggs
wlthln (Table 1>' Of the falled nests, 3 located on
woodchlp substrate contalned embryos that died early ln
the 1 r deve 1 opment and 2 located on grave 1 substrate
contalned eggs wlth living embryos that still had not
hatched by J ate fa 1 J and presumabJ y dl ed over wl nter.
One nest observed ln 1986 contained embryos that were
almost fully developed ln October. The fol10wlng
sprlng 2 live hatchllngs ernerged trom thls nest.
Overa Il, emergence 1 ncreased f rom 47.4% 1 n 1986
te 64.2% ln 1987. Most deaths (30.8%) occurred ln the
embryonlc stages. Infertl10 eggs accounted for 12% of
losses and 1.3% of eggs hatched but did not emerge.
Hatchllng Emergence
The Mean numbeL of days from the deposltlon of
eggs 1 n the spr 1 ng to ha tch Il ng emergence in the fa 11
was 96 (N=14: range 82-107: SE= 1.09). On 1 y one nest
produced hatchllngs ln the sprlng. Emergence fram the
nest that overwlntered eccurred sornet Ime prlor to 18
May. The t Ime taken by hatch Il ngs to emerge from
27

o
.......
""----------------
Indlvldual nesta varled. Synchronoua emergence la
deflned here as aIl hatchllngs havlng emerged wlthln a
perlod of 24 hrs while asynchronous emergence refers to
hatchllngs having emerged over a perlod greater than
24h. Hatchlings emerged asynchronously from 9 (64.3%)
nests over a period of 2-4 days, whlle emergence from 5
(35.7%) nests was synchronous. There was no
slgnlflcant difference ln the proportion of nests from
which emergence was synchronous and asynchronous (X2 =
1.14 P> 0.05). Most hatchl1ngs emerged between
0700-1800 hours dur i ng day Il ght • Only 16 hatchllngs
(4%) from 3 of the 14 nests emerged between 1800-0700 .
Movement and OrIentatIon of Hatchllngs
A total of 180 hatch I1ngs were marked and
followed from the ne st sI te. Dispersal patterns from
17 nests were mapped (Fig 2-a.b,c). Three of these
nests were 1 ncubated 1 ndoors but hatch 11 ngs were
released at the original nest sites. Flfteen (88%) of
the nest si tes were on sI opes rangl ng from 40 to 380 •
A hlghly slgniflcant number of hatchllngs, 156 (95.1%),
observed on sloped nest sites were posltlvely
geotactlc. (X2 =133.56: P<O.OOl).
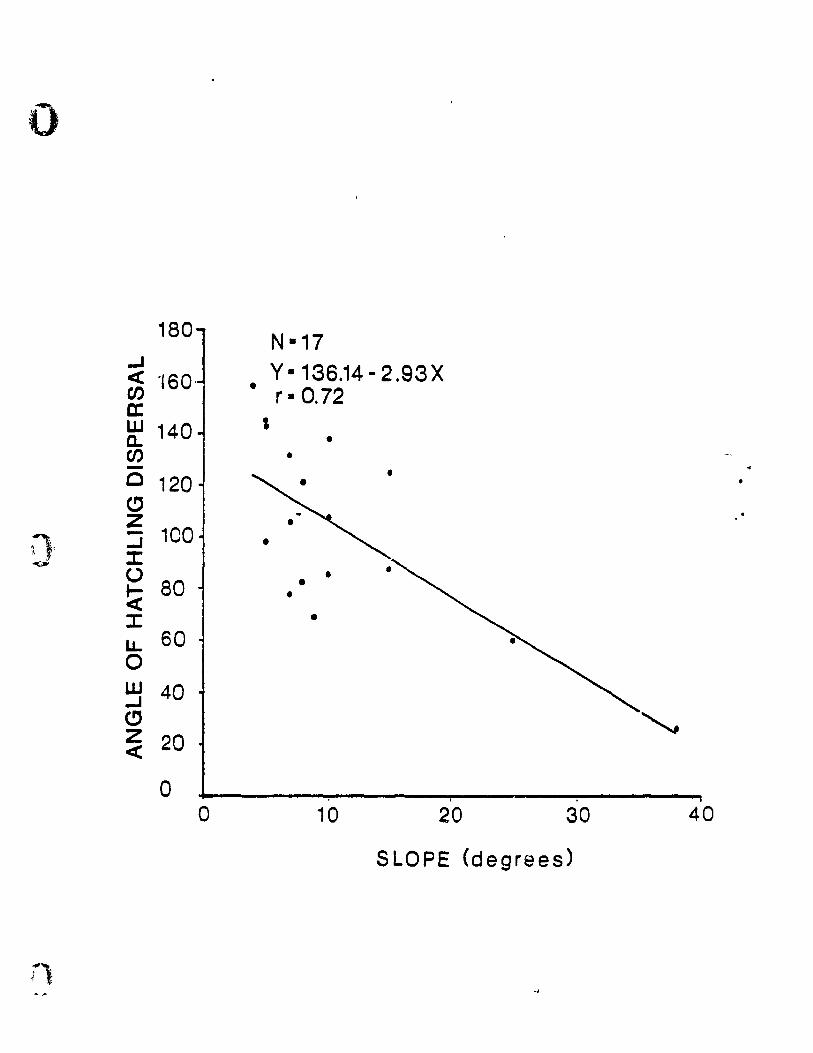
There was a slgnlflcant negatlve llnear
relatlonshlp between the angle of hatchllng dlspersal
28

(
(
(
fram the nest wlthln the flret 10 m, and the slope at
the nest site CSlmple Ilnear regreaelon; F 1,1S- 15.76:
P<O.OI; Fig. 3). !wo of the sloped nest sites were
characterlzed by land sloplng away ln 2 directions
(FIg.2a). thus when measur 1 ng the angl e of hatch II ng
dlepersal, each slope was consldered separately.
The 16 hatchllngs released on fIat nest sites
moved ln aIl dlrectl~ns CFIg.2b). Calculatlons of the
mean angle of dlsplacement were followed by evaluatlon
of the angular dispersion Cclrcular standard
devlatlon>. Uslng Raylelgh~s test lt was determlned
that there was no elgnlflcant mean dlrectlonallty ln
the dlpersal of hatchllngs on each of the 2 fIat nest
sites (Rayleigh/a test, nest 1 z-O.242 P>0.05; ne st 2
z-O. 972. P>O. 05,. Hatch II ngs from 2 nests 1 n 1986
dlspersed wldely as they crossed a fIat section of road
CFlg 2a>.
Hatchllngs were most active between 0700-1800
hours and very Ilttle movement was observed after dark.
When fi ret re 1 eased at the nest si te, a JI hatch 11 nge
remalned immobile, extended thelr necks and appeared to
scan the area. This stop and scan behavlor wae
exhlblted perlodlcally, and was observed whl)e
hatch II ngs weie both 1 n the open or under cover. 1 t
appeared to be performed most frequently when there was
movement ln the hatchllngs~ llne of vision. The hlgh
29

o
--------_._---------------------------- ---
denslty of vegetation around many neste made It
dlfflcult to determlne whether or not hatchllngs were
movlng towards the nearest coyer. The 2 neete on leve1
ground, however, were located ln woodchlp fields wlth
only eparse clumps of vegetation, and as prevlously
lndlcated, hatchllngs at theee sites exhibited no
preference for Any specific direction.
The river was not visible frorn a hatchllng's
perspectlv~ for 6 of the 17 nests and Indlvlduals frorn
9 nests passed through dense vegetat Ion through wh 1 ch
ne 1 ther the water nor a br 1 ght open hor 1 zon cou 1 d be
seen.
When hatch II ngs stopped mov 1 ng for extended
periode of tlme <usually overnlght), the y burrowed Into
the ground and were vlrtually Impossible to see.
Movemen t was observed dur 1 ng heavy prec 1 p 1 ta t Ion, as
we lIas on Wl.rm sunny and coo 1 overcast days. Sorne
hatchllngs occaslonally stopped movlng for several
hours, and euch behavlor was observed on both wet and
dry days, and through a wlde range of temperatures.
The average tlme taken by hatchllngs to reach
water was 2 hrs. 56 mine. for nests located less than
18 m from water, and 65 hrs. 15 mlns. for nests
sltuated between 121-167 m fram water.
After f Irst entering the river, hatchllngs
remalned ln ehallow areas « 15cm deep) and elther

(
(
burrowed Into the substrate or moved lnto nearby
aquatlc vegetation. The maJorlty of hatchllngs (82%)
etayed wlthln 10m of where they entered the water for
the 2 days they were monltored.
Hatchllng Survlval
The fate of hatchllngs from nests located less
than 18 m fram the water was determlned (Table 2). A
maJorlty of hatchl Inge from these neste (72.5% and
82.5% ln 1986 and 1987 respectlvely) were successful ln
reachlng water. Wh1le no Instances of predatlon were
observed 1 n 1986, 2 ha t ch 11 ngs were preyed upon by
bu llfrogs CRAnA catesbe 1 ana) 1 n 1987. 1 n both cases
hatchllngs were preyed upon as they entered the water.
On another occasIon, although no predation occurred, a
) arge garter snake (Thamnophls Sirta"ils alrtalls) was
observed followlng a hatchl1ng for several meters
through dense vegetation. Great blue herons (Ardea
herod1as) were frequently seen feedlng ln the shallows
where hatchllngs dlsappeared fram, but emlgratlon
cannet be [yled out.
The fate of hatch 11 ngs from nests located
between 122-167 m fram water le summarlzed ln Table 2.
Hatchllngs from these nests suffered hlgher mortallty
th an those from nests that were close to water.
31

o
:tJ
Mortall ty was caused by both predat Ion and paeslng
vehlcles as turtles crossed the road. AlI Instances of
predation ln 1986 occurred overnlght whlle hatchllngs
were wlthln 20 m of the r.est sites. Seven partlal1y
eaten hatchllng carcasses were found ln a network of
burl"ows 'C.hat wel"e 1 ocated 1 n an sandy meadow. Three
longworth traps, balted wlth dead hatchllngs, were set
near the burrow entrances for 2 nlghts. One meadow vole
(Mlcrotys pennsylyanlcus) was caught durlng thls
perlod. Only ~ne hatchl1ng was preyed upon ln 1987 and
th ls predat ion occurred overn 1 ght 40 m fran the nest
site on a graesy slope. The remalns of thls hatchllng
were also found ln a burrow. The condition of the
carcass was slmllar to those found ln 1986, however,
nothlng was caught ln 4 nlghts of trapping. These 1987
nest sites were sltuated on the gravel shoulder of a
road, over 60 m east of the 1986 nest sites. In 1986,
4 hatch 11 ngs were run over as they crossed a fIat
section of the road on thelr way to water. Nests ln
1987 were J ocated al ong a sloped sect 1 on of the road
and 4 hatchllngs fram these nests were kl11ed as they
moved down thls slope.
The fate of hatchllngs that were lost by the
observer- ln thlck vegetation Is llsted as unknown ln
Table 2.
32

(
(
(
DISCUSSION
Nest Survlval
Hatchllngs emerged from 15 of the 20 protected
neste whlch Is a conslderably hlgher euccess rate than
was reported for South Dakota (Hammer, 1969) wher-e
hatch Il ngs elnerged from on 1 y B of 22 protected nests.
Nest predatIon ls an important factor reduclng
nest surv 1 va lin th 1 s popu 1 at Ion of snapp 1 ng tur-t 1 es
(RobInson and BIder. 1988), and It le apparent that the
pr-esence of lnfertlle eggs, as weIl as the mortal1ty of
developlng embryos and hatchl1ngs also serve to
decrease nestlng success.
Two types of complete nest fallur-e were observed
ln th 1 s study. In the f 1 ret type embryos appeared to
have dl ed ear) y dur 1 ng the 1 ncubat Ion peI' 1 od. Slnce
only nests deposlted ln the woodchlp substrate suffered
th 1 s type of f a Il ure, 1 t seems Il ke) y tha t the
substrate was ln sorne way r-esponslble. Decomposing
s01,twood chIps are acldlc and perlodlc examlnatlon of
th i s substrate throughout the sunmer revea) ed that 1 t
remalned very wet below 4 cm.
In the second type of nest fal1ur-e, embryos wer-e
a Il ve al though not fu Il y deve 1 oped ln the fa Il and
33

o mor-ta 11 ty was pr-esumabl y caused by fr-eez 1 ng 1 n the
wlnter. Embryos ln these nests may have had
Insufflclent tlme to develop slnce the partlally
shaded condl t Ions at the nest si te may have lowered
temperatures Inslde the nest. These results are
comparable to those of Congdon et al.(1987), who
suggested that low Incubation temperatures related to
fully shaded nests caused 11.8% of nest tallures ln
those nests that escaped predation.
Hatchllng Emergence
The average number of days fram egg deposltlon
to hatchllng emergence ln the Calumet populatIon (96
days) was slmllar to that reported for- southern
MichIgan (93.2 days; Congdon et al. 1987) and the range
of 90-119 days reported for 3 hests in Ontario (Obbard
and Brooks, 1981b). AIl but one clutch emerged ln the
fall and the clutch that emerged ln the sprlng pr-oduced
on 1 y Z II ve hatch II ngs. These hatch Il ngs may have
avolded fr-eezlng because the nest was located at the
slde of the rallr-oad tracks where snow Is pl1ed deep ln
wlnter-. Obbard and Brooks (1981a) clted a slmllar- hlgh
snow accumulation for the 1 nest that successfully
overwlntered ln thelr study. While delayed emer-gence
has been reported for thls specles (Toner, 1940;

(
(
(
Bleakney, 1963; Ernst, 1966; GIbbons and Nelson, 1978,
Obbard and Brooks, 1981a) Most hatchllngs ln Ontario
(Obbard and Brooks, 1981a), Michigan (Sexton,1957;
Congdon et al., 1987) and South Dakota (Hanmer, 1969) ,
emerge ln the fall.
Hatchllngs ln thls study emerged synchronously
and asynchronously as dld hatchllngs ln Michigan
(Congdon et al., 1987). Ninety-six percent of
hatchlings ln this study emerged durlng dayllght hours.
Burger (1976) reported simllar emergence times for
dlamonclLack terrapln hntchllngs. In contrast, green
sea turtle ~Chelonla mvdas) hatchlings exhlblt
nocturnal emergence, controlled by thermal lnhlbl tlon
of activity (Morovsky, 1968).
Movement and OrientatIon of Hatchl1ngs
Hatchlings fram nests on slopes moved downh111
whlle hatchllngs on fIat la~ld moved randanly,
exhlbItlng no dlrectlonal preference. Burger (1976)
documented slmi lar dispersal patterns for dlmondback
terrapln hstchl1ngs, but noted that the location of
nearest vegetation was a stronger directional stimulu~
than 1 nc Il ne. Snapp 1 ng turt 1 e hatch Il ngs showed no
preference for the directIon of the nearest vegetation
ln fIat areas, and It therefore seems unl1kely that the
35

o
1', : ,.
location of cover ls Important ln the orientation of
thls specles. Noble and Breslau (1938) conducted
laboratory tests on the e-ffect of slope on snapplng
turtle hatchllngs ln the dark and noted that hatchllngs
exhlblted a marked tendency to move uphill. Whl1e thls
behavlor may facllitate emergence fram the nest (Noble
and Breslau. 1938>. lt would appear to have llttle
effeet on hatchllngs as they move to water. slnee Most
hatch Il ngs emerged 1 n day Il ght and ceased mov 1 ng by
ni gh t fa Il • In contrast, dlamondback terrapln
hatehllngs emerged durlng the day but made thelr way
rapld1y to the nearest vegetation where they remalned
untll nlghtfall (Burger, 1976).
The angle of snapplng turtle hatchllng dispersal
from the nest varled Inversely wlth the slope at the
nest site. Hatchllngs dlspersed fram as much as 1580
on cl slope of 40 to as llttle as 26° on a slope of
38°. Slmllarly, Burger (1976) reported that
dlamondback terrapln hatchllngs dld nct disperse more
than 300 on slopes greater than 100 , whereas, on s10pes
of less than 100 the y dlspersed more than 750 •
Noble and Breslau (1938) demonstrated that
anapplng turtle hatchllngs were attraeted to areaa of
hlgh humldlty and brlght Illumination. They euggested
that an open hor 1 zon, the presence of humldl ty
gradients and the position of the~ sun mlght be

(
(
(
important eues for orientatIon. A negatlve response to
dark masses of vegetation may also be Involved in the
orientation of sorne freshwater hatchllngs (Anderson,
1958). Humldlty gradients dld not appear to be
neceesary for orientation in thle etudy, eince
hatchllngs moved successfully to water through
eaturated areas, and durlng heavy precipitation. Over
half of the hatchllngs passed through dense vegetation
which obscured any vlew of the water, the sun or the
open horizon. Nevertheless, hatchlings contlnued to
orient slJccessfully to water. The results of thls
study suggest that positive geotaxls 115 the most
Important factor dlrectlng snapplng turtle hatchllngs
to water.
Once ln the water hatchllngs tended to remaln ln
the shal10ws whlch ls consistent wlth the 11mlted
swlmmlng abllity reported for thle age group (Hammer,
1969). Hatchllngs elther burrowed Into the substrate
or moved to aquat 1 c vegetat 1 on. Such behavlor may
provlde enhanced protection, concealment (Froese, 1978)
and ln the case of vegetation, physlcal support
(Sexton, 1958).
37

o
_. -- ._ .... _._---------.,
Hatchllng Survlval
The type and extent a'! hatch 11 ng marta II ty
appeared ta be lnfluenced by at lea!lt 3 factorts: (1)
the amount of tlme that hatchllngs were vulnerable to
predation, (2) the type of habitat through whlch
hatchllngs travel1ed and the predators assoclated wlth
those habl tats, and (3) the presence of roads near
nestlng areas. Hatchllngs fram distant nests spent at
1 east one n 1 gh t on 1 and and a II preda t 1 on on these
Indlvlduals was nocturnal. Hatchl1nga fram nests that
were close ta water usua 11 y reached the river before -nlghtfal1 and were therefore not vulnerable ta
nocturnal terrestrlal predators. AIl nacturna 1
predation, apparently by meadow voles, occurred ln
sandy, weIl dralned fields, whereas diurnal predation
by bullfrogs occurred along the water's edge. Due to
the presence of the observer, the resulte of thls study
may underestlmate the extent of diurnal predation. lt
1 saI sa possi bl e that sorne of the hatch II ngs lost 1 n
thlck vegetation were preyed upon. Law diurnal
predation however, would be consistent wlth the d1urnal
emergence and actlvlty patterns observed for most
hatch Il ngs.
38

(
(
(
Vehlcles passlng on a road accounted for some
hatchllng mortallty ln each of the 4 nests located near
1 t. Hatch Il ngs mov 1 ng on the road decreased the 1 r
chances of surv! va 1 by stopp 1 ng and scann 1 ng the area
every tlme a vehlcle passed, and thereby remalnlng on
the road for a considerable length of tlme. Whlle It
la dlfflcult to determine the purpoae of the stop and
scan behavlor, It appeared to be assoclated Most often
wlth movement ln the hatchl1ngs' llne of vision and
thus, mlght be related to predator avoldance.
In conclusion, the survlvorship of neonate
snapplng turtles was affected by a number of factors
related to female ne st site selection includlng: nest
dl stance from water, the type of habl tat surroundl ng
the nest and the presence of nearby roade. Further
research le needed however, to underetand the factors
whlch affect survlval of hatchllngs once they reach the
water.
39

o LITERATURE CITED
Ande~son, P.l. 1958. The wate~-app~oach behavlor of
hatchllng tu~tles. Copela 1958: 211-215.
Bleakney. J.S. 1963. Notes on the dlst~lbutlon and life
hlsto~les of tu~tles ln Nova Scotla. Cano FIeld-Nat.
77: 67-76.
Bu~ge~, J. 1976. BehavioL of hatchllng diamondback
te~~aplns (Malaclemys te~rapln) ln the fIeld.
Copela 1976: 742-748.
Conant, R. 1975. A FIeld GuIde to Reptiles and
Amphlblans of Easte~n and Cent~al No~th Ame~lca. 2nd
Edition. Houghton Mlfflln, Boston.
Congdon, J.O., G.L. Breitenbach, R.C. Van Loben Sels,
and D.W. Tlnkle. 1987. Rep~oduction dnd nestlng
ecology of snapplng tu~tles (Chelydra serpentlna)
ln southeaste~n Michigan. He~petologlca 43: 39-54.
Ehrenfeld. D.W. 1979. Behavlor assoclated wlth nestlng.
In: Tu~tles: pe~spectlves and ~esea~ch. Edlted by
M. Ha~less and H. Mo~lock. John Wlley and Sons. ~ew
Yo~k. pp.417-434.
40

(
(
(
• ft SI'
Ernst, C.H. 1966. Overwlnterlng of hatchllng Chelydra
serpentlna ln southeastern PennsylvanIa.
PhIl. He~petol. Soc. Bull .14: 8-9.
Ernst, C.H. and R.W. Barbour. 1972. Turtles ~f the
United States. Unlvet'sity Pt'ess of Toronto, Toronto.
Ft'oese, A.D. 1978. HabItat preferences of the common
snapplng tut'tle, Chelydra set'pentlna (Reptilla,
Testudlnes, Chelydrldae). J. Herpetol. 12: 53-58.
Galbra1th, D.A., C.A. B1shop, R.J. Brooks, W.L. Slmser
and K.P. Lampman. 1988. Facto~s affectlng the denslty
of populations of common snapplng turtles (Chelydra
aorpentina serpeutlna). Cano J. 2001. 66:
1233-1240.
GalbraIth, D.A., M.W. Chandler and R.J. Brooks. 1987.
The fIne structut'e of home ranges of male Cbelydra
serpentina: are snapplng tut'tles terrItorial? Cano
J. Zool. 65: 2623-2629.
GIbbons, J.W. 1987. Why do turtles lIve so long?
BloSclence 37: 262-267.
GIbbons, J.W. and D.H. Nels~n. 1978. ~he evolutlonary
slgnlflcance of delayed emergence ft'am the nest. by
hatchllng turtles. EvolutIon 32: 297-303.
41

o Hamilton, W.J., J~ .1940. Observations on the
reproductive behavlor of the snapping tu~tle.
Copela 1940: 124-126.
H~e~, D.A. 1969. Parametere of a marsh snapplng
turtle population, Lacreek Refuge, South Dakota. J.
WIldl. Manage. ~3:995-1005.
MaJor, P.D. 1975. Denslty of enapplng turtles,
Chelvdra serpentlna, ln western West Vlrglnla.
He~petologlca 31: 332-335.
Moslmann, J.E. and J.R. Bider. 1960. Variation, sexual
dlmorphlsm, and maturlty ln a Quebec populatIon of
the common snapplng turtle, Chelvdra serpentina.
Cano J. Z~ol. 38: 19-38.
Morovsky, N. 1968. Nocturnal eme~gence of hatchl1ng sea
tu~tles: control by thermal Inhibition of actlvlty.
Nature ~20: 1338-1339.
Noble, G.K. and A.M. Breslau. 1938. The senses Involved
ln the migration of young fresh-water turtles after
hatchlng. J. Comp. Psychol. 25: 175-193.
Obbard, M.E. and R.J. Brooks. 1980. Nestlng migrations
of the common snapplng turtle Chelvdra
aerpentlna. Herpetologica 36: 158-162.
42

(
(
_____ • 1981a. Fate of overwlntered clutches of the
commen snapplng turtle (CheJydr~ serpentlnl) ln
Algonquin Park, Ontario. Cano Field-Nat. 95:
350-352.
_____ . 1981b. A radlo-telemetry and mark-recapture
study of actlvlty ln the eommon snapplng turtle.
~~ .erpentlna. Copela 1981: 630-637.
Petokas, P.J. and M.M. Alexander. 1980. The nestlng of
Cbelydra .erpentlna ln northern New York. J.
Herpetol. 14: 239-244.
Robinson, C. and J.R. Bider. 1988. Nestlng synchrony- a
strategy to decrease predation of snapplng turtle
(Cbelydra .erpentlna) nests. J. Herpetol. 22:
470-473.
Sexton, O.J. 1957. Notes eoncernlng turtle hatchllngs.
Capela 1957: 229-230 •
. 1958. The relatlonshlp between the habitat
preferences of hatchllng Cbelydra serpentIn. and
the physlcal structure of the vegetation. Ecology 39:
751-754.
Tlnkle, D.W., J.D. Congdon. and P.C. Rosen. 1981.
Nestlng frequency and suceess: Implications fer the
demography of painted turtles. EcoJogy 62: 1426-1432.
43

o Toner, G.C. 1940. Delayed hatchlng ln the snapplng
turtle. Capela 1940: 265.
White, J.B. and G.G. Murphy. 1973. The reproductive
cycle and sexual dlmorphlsm of the common snapplng
turtle, Chelydra .erpentlna serpentlna.
Herpetologlca 29: 240-246.
Wllhoft, D.C., M.G. Del Bagllvo and M.D. Del Bagllvo.
1979. Observations on mammallan predatlon of snapplng
turtle neste (Reptllla, Testudlnes, Chelydrldae). J.
Herpetol. 13: 435-438.
Yntema, C.L. 1970. Observations on females and eggs of
the common enapplng turtle Chetydra serpentlna.
Am. MidI. Nat. 84: 69-76.
44

(
~ 1
1
" f
,
f , (: l-
f" f . 1-
~ ,
(
TABLE 1: SUMMARY OF FATE OF PROTECTED NESTS ~ND
EGGS AT CALUMET, QUEBEC
1986 1987
TOTAL NUMBER OF NESTS PROTECTED 10 10
NUMBER OF FAILED NESTS 3 2
TOTAL NUMBER OF EGGS 350 360 IN ALL NESTS
N % N %
HATCHLINGS EMERGED 166 47.4 231 64.2
INFERTILE EGGS 26 7.4 59 16.4
DEAD EMBRYOS 149 42.6 70 19.4
DEAD HATCHLINGS (IN NEST) 9 2.6 0 0.0
45
TOTAL
20
5
710
N %
397 55.9
85 12.0
219 30.8
9 1 .3

o
f) ........
TABLE 2: THE FATE OF HATCHLING Cbelydra serpentlna
FROM NESTS LOCATED WITHIN 18 M AND BETWEEN
122 AND 167 M FROM THE WATER
NUMBER 'OF HATCHLINGS FROM NESTS < 18 M FROM WATER
1986
58
N " REACHED WATER 48 72.5
PREYED UPON 0 0.0
UNKNOWN 10 27.5
NUMBER OF HATCHLINGS FROM NESTS 23 BETWEEN 122-167 M FROM WATER
N " REACHED WATER 6 26.0
PREYED UPON 7 30.5
KILLED BY VEHICLES -4 17.5
UNKNOWN 6 26.0
46
1987 TOTAL
79 137
N " N " 64 82.5 112 81.7
2 2.5 2 1.5
13 15.0 23 16.8
20 43
N " N " 15 75.0 21 48.8
1 5.0 8 18.6
4 20.0 8 18.6
0 0.0 6 14.0

c
(
(
FIGURE 1. LOCATION OF THE STUDY AREA AND
NESTING SITES IN RELATION TO THE
OTTAWA RIVER AT CALUMET, QUEBEC
47


(
(
(
FIGURE 2a. HATCHLING DISPERSAL PATTERNS
FROM SLOPED NEST SITES LOCATED
BETWEEN 122 AND 167 M FROM WATER
(Each 11ne Indlcates the path of
one hatchl1ng)
49

o
t)
w a.. ~=7#~=~
, ,
, , ,
w C-o ..J (/)
, ,
, ,
" , .. -
, .... / ..
.....
SLOPE----Ir~ v~
- ~/'-- ~ (./ 1'- ~ _ _ u... - .... -.......... , ..................... /"" ~- .... _ ..... .
ln
... .. .. .. ... :'
....
".
o 50 , , meters
ROAD

(
(
(
FIGURE 2b. HATCHLING DISPERSAL PATTERNS
FROM 2 NESTS LOCATED ON LEVEL
GROUND
(Each l1ne Indlcates the path of
one hatch 11 ng)
51

o
\ , ,
o 50 1 ~--:--_..JI
meters
o , meters
10 1

(
(
(
FIGURE 2c. HATCHLING DISPERSAL PATTERNS
FROM SLOPED NEST SITES
LOCATED WITHIN 18 M OF WATER
(Each 11ne Indlcates the path of
one hatchl1ng)
53

o
w Cl.. o ..J rn
w C-O ..J rn
10"
o 10 , , meters
o 5 1 1
meters

l f 1 }
l !
(
(
(
FIGURE 3. THE RELATIONSHIP BETWEEN THE
ANGLE OF HATCHLING DISPERSAL
FROM THE NEST AND THE
SLOPE AT THE NEST SITE
o
180 N -17
..J Y·136.14 - 2.93X < 160·
(/) • r - 0.72 a: w 140
, c.. • (/) • -c 120 C) Z
100 ;j, -..J d J:
U 80 ~ ::c 11.. 60 0 W 40 ..J C) z 20 <
0 0 10 20 30 40
SLOPE (degrees)

(
(~
r.i ,
(
CONNECTING STATEMENT
Section 2 consldered the orientation, movement
and survlval of hatchllng snapplng turtles ln the
field. The results obtalned durlng the flrst field
season of th 1 s study, al ong \ri lth genera 1 observa t ions
made durlng prevlous years, Indlcated that 20% to 25%
of females nest at distances greater than 100 m from
water. The advantages of thls behavior, If Any, are
unknown.
Section 3 deals with selected aspects of female
snapplng turtle reproductive ecology including clutch
size and reproductive effort ln relation to nest
dIstance from water.
57

o
SECTION 3
SOME ASPECTS OF REPRODUCTIVE ECOLOGY IN RELATION TO
NEST DISTANCE TO WATER OF A NORTHERN POPULATION OF THE
COMMON SNAPPING TURTLE (Cbelydrl serpentinl)
*For submlsslon to Herpetologlca

(~
ABSTRACT
Aspects of the reproductIve ecology of a
i'opulatlon of common snapplng turtles (Chelydra
aerpentlna) were studied from May to October (1987) ln
Calumet, Ouebec. During 3 years of observatIon It was
noted that 20-25% of snapplng turtle nests were
deposited at distances greater than 100m from water.
The reasons for dIstant nesting were not apparent,
wh1ch led to the examlnatlon of some of the
reprOductive characteristlcs of nestlng females. The
nests wer-e protected from predators. Hatchllngs wer-e
we 1 ghed upon emergence in the fa Il • The carapace
1 engths of 19 repr-oduct 1 ve fema les r-anged fr-om 211 -
348 mm. The mean clutch size was 33.78 while the mean
egg dlameter was 27.20 mm. Clutch size and Mean egg
dlameter- wer-e posItlvely related to female carapace
length. There was a slgnlficant positIve correlation
between egg dl ameter and hatch Il ng we 1 ght. The mean
straight Ilne dIstance from nests to the nearest
aquatlc habitat was 45 m, (range 4.86 - 173.00 m). The
maJorlty of nests (74%) were located wlthln 42 m of the
water whlle 26% of nests were 10cated between 121 - 173
m. Four of fi ve dl stant nesters were 1 arge fema 1 es.
There was a slgnUlcant dlfference between the mean
59

o cJutch size of nests located close to water and those
Jocated far fram lt, wlth distant nests havlng Jarger
clutches.

( INTRODUCTION
The character 1 st 1 cs of snapp 1 ng turt 1 e nest
sItes have been descrlbed ln the literature (Lonke and
Obbard, 1977: Petokas and Alexander, 1980; Congdon et
al. , 1987; Rob i nson and Bi der, 1988). Neverthe 1 ess,
the relationship between nest site selection factors
and reprOductive success are poorlY understood. Whi1e
sorne females nest relatlvely far from water (as far as
181m in Michigan) these nests are not reported to have
Increased survlvorshlp by decreaslng nest predatIon
(Congdon et al.,1987). Over several years It has been
observed that between 20% to 25% of females at Calumet,
nest far (> 100m) from water. The advantages of th 1 s
are not apparent, however the costs of travel could be
balanced by reproductive characterlstlcs such as better
incubation conditions at the nest site, increased
hatchl1ng survlval following emergence, or safety from
floodlng.
Whlle the common snapping turtle has been wldely
studled (White and Murphy, 1972) it ls a specles for
whlch many parameters of reproduction are poorly known
CPetokas and Alexander, 1980). Mean clutch sizes for
different reglons vary from 19.9 to 49 eggs/ clutch
CHanmer, 1969: Yntema 1970: White and Murphy, 1973:
61

o
----------------------
Lonke and Obbard, 1977: Petokas and Alexander, 1980;
Congdon and Gibbons, 1985: Congdon et al., 1987) and
posl t 1 ve re 1 at lonsh Ips ex 1 st between adu 1 t body slze
and the clutch size of Many polkl10therms (Bluewelss et
al., 1978>' A positive relatlonshlp between female
body size and clutch slze has been demonstrated ln a
number of snapplng turtle populations (Vntema, 1970:
White and Murphy, 1973: Congdon and Gibbons, 1985:
Congdon et al., 1987), as weIl as a positive
relationshlp between femaJe slz~ and egg size (Yntema,
1970; Congdon and Gibbons, 1985; Congdon et al., 1987).
There are very few publlshed data documentlng hatchllng . sI ze for th 1 s spec 1 es. Morr 1 s et al. (1983) observed
that larger eggs tended to produce larger hatchllngs ln
the labot"atot"y.
The purpose of this study was: (1) to examine
the t"elatlonships between female size and clutch sIze,
egg size and hatchllng slze, (2) to examine nest
distance from water as it t"elates to clutch size and
femaJe reprOductive effort.
62

(
(~
(
METHODS AND MATERIALS
The study was conducted ln 1987 at Calumet,
Quebec (Lat. 45 38', Long. 74 39'). The study site was
vlslted dally from mld May untll the onset of the
nestlng aeason. Durlng the nestlng season fram 29 May
to 20 June the site was vlslted 3 tlmes dally, before
0800 hours, at noon and 1 hour before dusk. Followlng
the nestln; season, the nest sites were vlsited every
second day untll the first sion of hatchling emergence,
after whlch nest sites were checked mornlng and evenlng
untll mid October.
When a female had completed nestln;, she was
captured and carapace len;th and wldth vere measut'ed
uslng a metric ruler. Eggs were carefully removed and
counted. and egg dlameters wet'e measured to the nea~est
0.01mm uslng callpers. Followlng excavation eggs vere
returned ta the egg chamber and the solI vas replaced.
Four of the 19 nests were not excavated because females
nested synchronousl y and the obset'ver vas unabl e to
excavate some of these nesta dlrectly after deposltlon.
It wes feared that handllng the eggs from these nests
1ater on mlght reduce hatchabl1lty. Heavy metal
ecreens measur 1 ng 90 by 60 cm were p laced over the
nests. The distance fram the nest to the nearest water

o was recorded. For analysls of the relatlonshlp between
clutch size and nest distance fran water, addltiona}
data gathered ln 1986 were comblned wlth data fram thls
study ln order to obtaln a larger sample.
1 n ml d August the screens were removed and
rep 1 aced wl th _cy II ndr 1 ca 1 baskets constructed of 0.5
Inch (1.27 cm) hardware cloth measurlng 30 cm ln
dlameter and 18 cm ln helght. Cyllnder walls were
burled to a depth of 8 to 10 cm ln order to prevent
emerglng hatchllngs from escaplng. Upon emergence,
hatchllngs were welghed uslng a triple beam balance . accurate to 0.1 gm.

(
(
(
RESULTS
Female Size and Clutch Parameters
Female carapace length averaged 279mm and ranged
from 211II1I'I to 348nvn (N 1: 19: SE == 8.89). The mean
carapace wldth was 233.6mm ranglng fram 179 - 295mm (N
- 19; SE == 7.05). The mean clutch slze was 33.78 (N =
19: range = 18 - 52: SE = 2.32). A slgnlflcant
positive llnear relatlonshlp was found to exlst between
clutch slze and female carapace length (Simple llnear
regresslon: Fl.17 • 41.33: P (0.01. Flg.1).
The overall mean for egg dlameters fram 15 clutches
was 27. 20nvn ( N = 521; range == 23.50 - 32.31; SE -
0.05). Mean egg diameter exhlblted a slgnlflcant
positive 11near relatlonshlp wlth female carapace
length (simple llnear regresslon: F1,13 e 10.44: P (
0.01. Fig. 2>-
Hatchllng welghts from 12 clutches had a mean of
8.1g (N 1: 291: range z 4.5 - 10.3g: SE = 0.09). There
wae a slgnlflcant correlatIon between egg dlameter and
hatchllng welght (Spearman's r = 0.62; 0.01 < p <
0.05) •
The maJorlty (63%) of clutches contalned between 20
to 40 eggs whlle 10.5% had fewer than 20 eggs and 26%,
more than 40.

o
-; i. " l
Nest DIstance From Water
The mean straight line dIstance fram nests to
the nearest permanent water was 45m (N = 19: range =
4.86 - 173.00; SE = 13.21), The major 1 ty of nests
(74%) were 10cated wlthln 42m of the water, whi1e the
remalning mests (26%) were situated between 121 to 173m
from water.
The relatlonshlp between female size and nest
distance from water was not 1inear, however 4 of the 5
most distant nesters were large fema1es whi1e those
nestlng close to water were bath large and small. Sorne
data gathered in 1986 on clutch slze and nest dIstance
from water for 12 nests were comblned wl th the data
from thls study ln order to obtaln a 1arger eamp1e.
There was a slgnl flcant dlfference between the mean
clutch elze of nests located wlthin 100 m of the water
and the mean clutch slze of those located further than
100 m from water CStudent"s t: t= -2.41, p<O.05>.
Distant nests contained primarlly large clutches white
both 1 arge and sma Ile 1 utches were located close to
water (Fig. 3). Given the positive relatlonshlp
between clutch elze and female slze, It le probable
that the distant nesters were large females.
66

(
(~
(
DISCUSSION
Female elze and Clutch Pa~amete~s
The mean clutch slze found ln thls study (33.8)
1 s compa~abJ e to thoee repo~ted for othe~ northern
localltles Includlng: 33.9 ln Ontario (Lonke and
Obba~d, 1977); 30.9 ln northe~n New York (Petokas and
Alexander, 1980) and 27.9 ln Michigan (Congdon et aJ.,
1987>' Fu~ther south howeve~, clutches appear to be
smaller wl th reported mean clutch slzes of 19.9 ln
Tennessee (White and Mu~phy, 1973) and 23.6 ln No~th
Carol Ina (Congdon and Gibbons, 1985).
A elgnlflcant positive reJatlonshlp between female
body slze and clutch size was found ln this study whlch
corresponds wlth prevlous reports of thls relatlonshlp
ln enapplng turtJes (Congdon et aJ., 1987; Yntema,
1973; Congdon and GI bbons, 1985: Wh 1 te and Murphy,
1973), and ln othe~ freshwater turtle specles (Gibbons,
1982>'
The mean egg dlameter (27.20 mm) le smaller than
the means of 28.4 mm reported by Petokas and Alexander
(1980) ln New York, and 31.00 mm reported by Congdon et
al., (1987) ln Michigan, but greater than the means
26.8 mm reported by Yntema (1973), and 25.78 mm
reported by Congdon and Gibbon (1985) in No~th

Carol Ina. The demographlc slgnlflcance of these
measurements ls unclear. Mean egg dlameter exh1blted a
slgn1f1cant positive relatlonship with female carapace
length. This ls slmllar to reports by Congdon et al
(1987), Congdon and Gibbons (1985), and Yntema (1970) ,
al1 of whom found a positive relatlonshlp between
female bOdy size and egg size but used dlfferent
measurements (body Maas, egg Maas and plastron length)
for their comparisons.
The '.lean hatchllng we1ght of 8.1 g. ls slmllar
to the Mean hatchl1ng welght of 8.9 g. reported by
Congdon et al. (1987) ln Michigan. Larger eggs tended
to produce 1 arger hatch II ngs and th i S corresponds to
the findings of Morris et al. (1983) who measured these
trends under control1ed laboratory conditions. Morris
et al., (1983) suggested that hatchl1ng size could be
Influenced by the slze of the eggs at ovlposltlon, and
by the hydr 1 c condl t Ion of the nest substrate chosen.
In the present study the dlfference ln hatchl1ng slze
due to egg size and female size was observed regardless
of the variable hydrie conditions of the weIl dralned
up 1 and nest si tes and the mol st nest si tes located
closer to water. Thus hydrie conditions at the nest
si tes di d not appear to compensate for the observed
differences ln hatchllng slze related to egg slze.
l

(
(~
(
Nest Distance from Water
Whlle the maJorlty of females nested close to .
the water_ seme travelled much farther and over rougher
terrain to nest <Robinson and Blder,1988). The results
of thls study Indlcate that while both large and small
clutches are deposited close to water, lt Is prlmarl1y
large clutches, presumably lald by large females, that
are located far from water. Larger females produce
larger cJutches and Jarger eggs whlch, ln turn, produce
larger hatchlings. It seems llkely that larger females
and larger hatchllngs would be best able to cope wlth
the rlsks assoclated wlth dIstant neBts. Females expend
more energy and lncur a greater rlsK from predation and
water loss ln these longer trips as do the hatchllngs
that emerge from these nests. It therefore seems
llkely that some beneflt could be derlved to compensate
for these costs. However, nests that are both near and
far from the water are equaJly susceptible to predatIon
(Congdon et al., 1987). Hatchlln~ survlval does not
appear to be lncreased at distant nest sItes and
hatchllngs fram distant nests suffer higher mortality
caused by both predators and vehlcles (Robinson and
BIder, SectIon 2). The advantages of distant nestlng
may be re 1 ated to lmproved 1 ncubat Ion' condl t Ions at
these well-drained open sites.

o
-
Inte~estingly, at least 3 females we~e killed
between 1984 and 1987 on a 0.5 km st~etch of road whlle
crosslng durlng the nestlng season. In vlew of thls
mortallty and that of emerglng hatchllngs (Robinson and
BideL, Section 2) due to tLafflc, It 19 appa~ent that
the road has a steady negative Impact on tULtles
nest1ng faL from wateL. Thus, although rlghts of way
create good, open nest1ng habitat at sorne distance from
wateL, they may be ecological tLaps that negate Any
advantages that could be galned fram better incubation
conditions.

(
---- -- ------____ ., __ ._il:lM .... ,-.... sr. _________________ •
LITERATURE CITED
Blueweles, L., H. Fox, V. Xudzna, D. Nakashlma, P.
Petera, and S. Sams. 1978. Relatlonshlps between
body slze and some 11fe hlstory parameters.
Oecologla 37: 257-272.
Congdon, J.D., G.L. BreItenbach, R.C. Van Loben Sels,
and D.W. Tlnkle. 1987. Reproduction and nestlng
ecology of snapplng turtles Chelvdra .erpentlna
ln southeastern Michigan. Herpetologlca 43: 39-54.
Congdon, J.D. and J.W. Gibbons. 1985. Egg components
and reproductive characterlstlcs of turtles:
retationships to body size. Herpetologlca 41:
194-205.
Gibbons, J.W. 1982. Reproductive patterns ln freshwater
turtles. Herpetologlca 38: 222-227.
Hammer, D.~. 1969. Parameters of a marsh snapplng
turtle population: Lacreek Refuge South Dakota. J.
Wlldllfe Mgmt. 33: 995-1005.
Lonke, D.J. and M. Obbard. 1977. Tag success,
dImensions, clutch slze and nestlng fldellty for the
anapplng turtle, Chelydra B.C,2entlna, ln
Algonquin Park, OntarIo, Canada. J. Herpetol. 11:
242-244.
71

o Morris, K.A .• G.e. Packard, T.J. Boardman. G.L.
Paukstis and M.J. Paekard. 1983. Effeet of the hydrie
envlronment on growth of embryonle enapplng turtles
(Chelvdra serpentlna). Herpetologlea 39:
272-285.
Petokas, P.J. and M.M. Alexander. 1980. The nestlng of
Chelvdra serpentina ln northern New York. J.
Herpetol. 14: 239-244.
Robinson, C. and J.R. Bider. 1988. Nesting eynehrony -
a etrategy to deer&ase predation of snapping turtle
(Chelydra serpentlnl) nests. J. Herpetol.
22:470-473.
White, J.B. and G.G. Murphy. 1973. The reproductive
cycle and sexual dlmorphlsm of the common snapplng
turtle. Herpetologlea 29: 240-246.
Yntema, C.L. 1970. Observations on females and eggs of
the common snapplng turt1e, Chelydra serpentlna.
Am. MidI. Nat. 84: 69-76.

(
FIGURE 1. THE REL~TIONSHIP BETWEEN CLUTCH
SI2E (y) ~ND FEMALE CARAPACE LENGTH (X)
(~
( 73

o
(1) N v; .!:. U -.2 U
.....
--
4J
3J
y= -27.55 + O.22x r = 0.84 n= 19
• •
10 ~--r-~--T-~--~~--~~--~~---~~I----~I--~
220 240 200 320 ~
Carapace Length (mm)

FIGURE 2. THE RELATIONSHIP BETWEEN MEAN EGG
DIAMETER (y) AND FEMALE CARAPACE LENGTH ex)
(~
(
75

o
'29 Y = 2.34 + O.OO13x r = 0.66 - n = 1:5 E • • • E 28 • ....." ...
(l) --Q.) • E • • 0 27 -. • -- • ëS ........
f 0> ---.. ".. 0 UJ • C • 0 26 • Q.) • ~
25 1 1
2X) 220 24J 2éO 200 lX) 320
Carapace Length (mm)

(
(~
FIGURE 3. THE FREQUENCY DISTRIBUTION OF
CLUTCH SIZE FOR NESTS LOCATED
< 100 M FROM WATER AND NESTS
LOCATED > 100 M FROM WATER
77

o
Nests located < lOOm frem water 5
4 n=22
3
2
0 o 10 15 Al 25 ::0 35 4) 45 5.J 55 8J - >-
i~ (J "" .. c
Q) ... .... cr Q) ~
LI..
Nests Iccated > 100m from WOTer
5 n=9
4
3
2
o~--------------~--~~~~--~--~-----5 10 15 2) 25 J) 35 4) 45 5J 55 éD
Clutch Size

(
(~
-, ft
GENERAL CONCLUSION
In a test ar'ena, positive geotaxls ln dayllght
eeemed to be the dominant eue dir'ectlng snapplng tUr'tle
hateh Il ngs dO\Jnh 111 to water' wh II e v 1 sua 1 and 01 faetor-y
eues, orientation by magnetic fields and molstur-e
gradl ei1ts appear'ed to be of li t t 1 e Impor'tance. The
maJor-ltyof hatehl1ngs movlng from sloped nest sites ln
the field wer'e also posltlvely geotaetie. In fIat
areas hateh Il ngs di sper'sed 1 n ail dl r'eet Ions and were
not attracted to near'by vegetation.
Infertile eggs and the death of hatehllngs and
developlng embr'Yos r'esulted ln the failure of almost
half of aIl eggs observed and ther-efore appear to be
Important factor's in overall nest sUr'vlvor-ship.
Furthermore, the type and extent of hatehling
mortality fol lowlng emer-gence appear-ed to be Influeneed
by the distance of the nest from water and thus, the
amount of tlme hatchlings wer-e vulne["able to p["edatlon.
the typP. of habItat through whleh hatchllngs tr'avelled,
the pr-edato["s assoclated wl th those habl tats and the
pr'esence of r-oads near the nestlng Ar-ea.
An examlnatlon of sorne female r'epr-oduetlve
characterlstics lndieated that clutch slze and Mean egg
diameter' wer-e positlvely related to female car'apace
79

o
-\ ~ "
-l,
length. There was a positive correlation between egg
d1emeter and hatchl1ng welght. Whlle the maJorlty of
females nest close to water, approxlmately 20" to 25"
nest farther than 100 m from water. The mean clutch
si ze of dl stant nests () 100 m) was 1 arger than the
mean cl utch si ze of nests located cl oser to water.
Glven the relatlonshlp between clutch elze and fernale
slze, distant nesters were probably large females.
80