
On the', Structure of Microtubules, Tau, and Paired ... the Structure of Microt… · On the',...
8
Pergamon 0197-4580(95)00026-7 Neurobiology of Aging,Vol. 16, No. 3, pp. 347-354, 1995 Copyright © 1995ElsevierScienceLtd. Printed in the USA. All rights reserved 0197-4580/95 $9.50 + .00 On the', Structure of Microtubules, Tau, and Paired Helical Filaments E. MANDELKOW, Y.-H. SONG, O. SCHWEERS, A. MARX AND E.-M. MANDELKOW 1 Max-Planck-Unit for Structural Molecular Biology, Notkestrasse 85, D22603 Hamburg, Germany MANDELKOW, E., Y.-H. SONG, O. SCHWEERS, A. MARX AND E.-M. MANDELKOW. On the structure ofmicrotubules, tau, andpairedhelicalfilaments. NEUROBIOL AGING 16(3) 347-354, 1995.-Microtubules and their associated proteins form the basis of axonal transport; they are degraded during the neuronal degeneration in Alzheimer's disease. This article surveys recent results on the st:ructure of microtubules, tau protein, and PHFs. Microtubules have been investigated by electron micros- copy and image processing after labeling them with the head domain of the motor protein kinesin. This reveals the arrange- ment of tubulin subunits in microtubules and the shape of the tubulin-motor complex. Tau protein was studied by electron microscopy, solution X-ray scattering, and spectroscopic methods. It appears as an elongated molecule (about 35 nm) without recognizable secondary structure. Alzheimer PHFs were examined by FTIR and X-ray diffraction; they, too, show evidence for secondary structure such as/~ sheets. Microtubules Alzheimer's disease Paired helical filaments X-ray diffraction Electron microscopy Tau-protein Kinesin Tubulin MICROTUBULES, stabilized by tau protein, serve as tracks for the axonal transport mediated by motor proteins such as kine- sin. The phosphorylation and aggregation of tau into PHFs is accompanied by its detachment from microtubules and their de- cay, leading to the breakdown of axonal transport. To under- stand the basis of these events in physiological and pathologic conditions it is therefore important to define the structures and interactions of the components. Here we describe some recent results on microtubules, tau protein, and PHFs. RESULTSAND DISCUSSION Microtubule Structure and Interaction With the Motor Protein Kinesin Microtubules provide the "tracks" for cytoplasmic transport in axons and dendrites. They are stabilized by microtubule- associated proteins which act as "ties," as it were, and the mo- tion itself is achieved by motor proteins whose force-producing end (the ATPase domain) in'Leracts with microtubules, and the opposite end pulls the "cargo" (e.g., vesicles) (for review, see 52). Unlike a railroad track, the molecular track is discontinuous be- cause it consists of discrete protein subunits (tubulin), and con- sequently, the motor cannot :roll smoothly (like a wheel) but has to step. The arrangement of the "stepping stones" along the microtubule therefore determines the motion of the motor. This raises several basic questions: What determines the direction of the movement? What is the step size of a motor protein? What is the efficiency and force of the motor? How is the movement affected by other proteins attached to microtubules? These questions have been addressed by a number of labo- ratories and some answers can be summarized as follows: The polarity of transport is determined by the intrinsic polarity of the microtubules and the intrinsic polarity of the motors. Micro- tubules have a "minus" and a "plus" end; in most cases, the mi- nus end is at the nucleation site near a microtubule organizing center and the plus endpoints to the cell periphery. A standard example is that of a flagellum where the minus end is at the basal body (proximal end) and the plus end is at the tip (distal end; 1,12). This arrangement is also found in axons where the plus ends uniformly point to the tip; dendrites are different, how- ever, in that they have microtubules pointing in both directions (3). The polarity can be visualized on the molecular level (see below). Most microtubule-organizing centers contain gamma- tubulin which can be labeled by gamma-tubulin antibodies. However, neuronal microtubules differ in that gamma-tubulin is not visible at the minus ends, suggesting that microtubules are originally assembled in the cell body and then transported out- wards (4). The directionality of motor proteins also appears to be an intrinsic property. Generally, kinesins are plus-end directed mo- 1Requests for reprints shouhl be addressed to E.-M. Mandelkow, Max-Planck-Unit for Structural Molecular Biology, Notkestrasse 85, D22603 Hamburg, Germany. 347
Transcript of On the', Structure of Microtubules, Tau, and Paired ... the Structure of Microt… · On the',...

Pergamon
0197-4580(95)00026-7
Neurobiology of Aging, Vol. 16, No. 3, pp. 347-354, 1995 Copyright © 1995 Elsevier Science Ltd. Printed in the USA. All rights reserved
0197-4580/95 $9.50 + .00
On the', Structure of Microtubules, Tau, and Paired Helical Filaments
E. M A N D E L K O W , Y.-H. S O N G , O. S C H W E E R S , A. M A R X A N D E . -M. M A N D E L K O W 1
Max-Planck-Unit f o r Structural Molecular Biology, Notkestrasse 85, D22603 Hamburg, Germany
MANDELKOW, E., Y.-H. SONG, O. SCHWEERS, A. MARX AND E.-M. MANDELKOW. On the structure ofmicrotubules, tau, andpaired helical filaments. NEUROBIOL AGING 16(3) 347-354, 1995.-Microtubules and their associated proteins form the basis of axonal transport; they are degraded during the neuronal degeneration in Alzheimer's disease. This article surveys recent results on the st:ructure of microtubules, tau protein, and PHFs. Microtubules have been investigated by electron micros- copy and image processing after labeling them with the head domain of the motor protein kinesin. This reveals the arrange- ment of tubulin subunits in microtubules and the shape of the tubulin-motor complex. Tau protein was studied by electron microscopy, solution X-ray scattering, and spectroscopic methods. It appears as an elongated molecule (about 35 nm) without recognizable secondary structure. Alzheimer PHFs were examined by FTIR and X-ray diffraction; they, too, show evidence for secondary structure such as/~ sheets.
Microtubules Alzheimer's disease Paired helical filaments X-ray diffraction Electron microscopy
Tau-protein K i n e s i n Tubulin
MICROTUBULES, stabilized by tau protein, serve as tracks for the axonal transport mediated by motor proteins such as kine- sin. The phosphorylation and aggregation of tau into PHFs is accompanied by its detachment from microtubules and their de- cay, leading to the breakdown of axonal transport. To under- stand the basis of these events in physiological and pathologic conditions it is therefore important to define the structures and interactions of the components. Here we describe some recent results on microtubules, tau protein, and PHFs.
RESULTS AND DISCUSSION
Microtubule Structure and Interaction With the Motor Protein Kinesin
Microtubules provide the "tracks" for cytoplasmic transport in axons and dendrites. They are stabilized by microtubule- associated proteins which act as "ties," as it were, and the mo- tion itself is achieved by motor proteins whose force-producing end (the ATPase domain) in'Leracts with microtubules, and the opposite end pulls the "cargo" (e.g., vesicles) (for review, see 52). Unlike a railroad track, the molecular track is discontinuous be- cause it consists of discrete protein subunits (tubulin), and con- sequently, the motor cannot :roll smoothly (like a wheel) but has to step. The arrangement of the "stepping stones" along the microtubule therefore determines the motion of the motor. This
raises several basic questions: What determines the direction of the movement? What is the step size of a motor protein? What is the efficiency and force of the motor? How is the movement affected by other proteins attached to microtubules?
These questions have been addressed by a number of labo- ratories and some answers can be summarized as follows: The polarity of transport is determined by the intrinsic polarity of the microtubules and the intrinsic polarity of the motors. Micro- tubules have a "minus" and a "plus" end; in most cases, the mi- nus end is at the nucleation site near a microtubule organizing center and the plus endpoints to the cell periphery. A standard example is that of a flagellum where the minus end is at the basal body (proximal end) and the plus end is at the tip (distal end; 1,12). This arrangement is also found in axons where the plus ends uniformly point to the tip; dendrites are different, how- ever, in that they have microtubules pointing in both directions (3). The polarity can be visualized on the molecular level (see below). Most microtubule-organizing centers contain gamma- tubulin which can be labeled by gamma-tubulin antibodies. However, neuronal microtubules differ in that gamma-tubulin is not visible at the minus ends, suggesting that microtubules are originally assembled in the cell body and then transported out- wards (4).
The directionality of motor proteins also appears to be an intrinsic property. Generally, kinesins are plus-end directed mo-
1Requests for reprints shouhl be addressed to E.-M. Mandelkow, Max-Planck-Unit for Structural Molecular Biology, Notkestrasse 85, D22603 Hamburg, Germany.
347

348 M A N D E L K O W ET AL.
tors, dyneins are minus-end directed motors, i.e., molecular en- gines do not switch directions by means of a gear box (for review, see 17). Given the uniformity of axonal microtubules, it follows that kinesin always moves toward the synapse, dynein moves to- ward the cell body. In dendrites, these motors could move either way, depending on the polarity of the microtubule they inter- act with.
Thanks to powerful new methods of high resolution video microscopy it has become possible in recent years to measure the step sizes and forces of motors such as myosin (on F-actin tracks) or kinesin and dynein (on microtubule tracks) (for re- view see 52). The forces are typically in the picoNewton range (33). The step sizes tend to agree with the spacings of the step- ping stones. For the case of kinesin, minimal step sizes of 4 nm (15) or 8 nm (47) have been observed; flagellar dyneins have 4 nm or 4 plus 8 nm (26).
These numbers are consistent with the structure of micro- tubules, as determined by X-ray fiber diffract ion, electron microscopy, and image reconstruction (Fig. 1). In order of in- creasing hierarchy, microtubules consist of protein monomers (a or/3 tubulin, molecular weight - 5 0 kD), dimers ( a -~ ) , pro- tofilaments (strings of dimers running along the length), and the tubules made up of 13 protofi laments. The effective "subuni t" or building block of a microtubule is the a-/3-heterodimer. The axial spacing of monomers is 4 nm; the spacing of dimers is therefore 8 nm, consistent with the step sizes mentioned. Because protofilaments run along the length a motor protein could sim- ply adhere to the path of one protofi lament; this seems to be the case for kinesin (27,42), whereas dynein can jump between adjacent protofi laments as is moves along (48).
There has been a protracted debate regarding the arrange- ment of tubulin dimers in a microtubule. In the original model (2) adjacent protofi laments were shifted by 3.1 nm (nearly half a dimer length); this lattice was proposed for flagellar A-tubules, hence the name A-lattice. The alternative model was the B-lattice where adjacent protofilaments are shifted by 0.9 nm (as in Fig. 1; for review see 38). The B-lattice does not allow a fully symmet- ric microtubule to be constructed from 13 protofilaments. It was therefore tacitly assumed that cytoplasmic microtubules had the (symmetric) A-lattice. This issue was difficult to decide because o f the very low intrinsic contrast between ~ and/3 tubulin, but the issue could be settled recently by specific labeling of micro- tubules with the kinesin head domain (Fig. 2).
These results can be summarized as follows (45): The kine- sin head crosslinks to/3 tubulin but not a tubulin. This shows that the two tubulin monomers are not equivalent with respect to the interaction with motor proteins, and that there is a stoi- chiometry of one kinesin head per tubulin dimer. Consistent with this, the kinesin heads have an axial spacing of 8 nm (the dimer spacing). The predominant stagger between adjacent pro- tofilaments is that of the B-lattice (0.9 nm). This means that microtubules must have some helical discontinuity, they cannot be fully symmetric. This discontinuity is shown as a continuous "seam" in Fig. 1, although other less continuous models are con- ceivable as well. The function of the seam is unclear at present. It could be the point where the wall closes up into a tubule, a special at tachment site for MAPs or other proteins, or it could be merely coincidental.
A more detailed view of the kinesin-tubulin interaction can be obtained by image reconstruction of decorated microtubule
B. GTP
®
• GTP
in
Tubulin sheets _+ kinesin head
. - I
, - 0
B.GDP
. G D P
~continuity
, - 1 ,-1/2 , - 0
® FIG. 1. Model of a microtubule surface lattice showing c~ tubulin (dark) and ~-tubulin (light). The axial spacing of monomers is 4 nm, that of dimers is 8 nm. Protofilaments are staggered by about 0.9 nm. This gen- erates quasi-helical lines which are however broken at least once around the circumference ("discontinuity"). The plus-end (top) terminates with a-tubulin, the minus-end with/3-tubulin./3-tubulin with wound GTP is near the plus-end, the interior contains only 13-GDP.
/
FIG. 2. Opened-up microtubule wall (sheet) without (top) and with (bottom) decoration by kinesin head. Note the additional cross-striations generated by the decoration, running at an angle of about 10 ° with re- spect to the horizontal. The optical diffraction patterns (right) show a 4 nm layer line, corresponding to the tubulin monomer repeat along the protofilaments (top), or an additional 8 nm layer line (bottom) arising from the spacing of kinesin heads.

MICROTUBULES, TAU, AND PHFS 349
wails (Fig. 3; 39). It is intere~;ting to note that the kinesin head lies asymmetrically with respect to/3-tubulin. If the microtubule wall is viewed from the outside, with the plus end up, the kine- sin head is on the right edge of fl-tubulin, reaching into the groove between and perhaps even across protofilaments; its shape is reminiscent of a pear or boomerang (depending on the stain distribution).
Anticipating the results discussed later, it is remarkable that it has so far not been possible to identify the interactions between microtubules and MAPs in ~a equivalent fashion. For example, cross-linking between tau and tubulin is not restricted to just one tubulin monomer, and negatively stained images of microtubules decorated with MAPs are only marginally different from undec- orated ones. This suggests that MAPs are more randomly dis- tributed on the microtubule surface and have a less compact structure than kinesin.
Quick freeze-deep etch images of the neuronal cytoskeleton reveal closely spaced filaments with many cross-bridges (23); they make it difficult to imagine how organelles might glide through the maze. On the other hand, in vitro motility assays show that MAPs do not have a major influence on the speed of gliding. One may thus ask, how does kinesin behave when it encounters another protein such as tau or MAP2? Does kine- sin push the MAP out of the way, or are the MAPs overwhelmed by the many kinesins actinl~, in concert? Experimentally this question is difficult to tackle; the evidence available thus far sug- gests that neither tau nor MAP2c have a measurable effect on the motility, whereas MAP2 can disrupt the kinesin-microtubule interaction to some extent, depending on the relative concen- trations (20,35,49). Overall, the simplest interpretation is that MAPs, even though they might bind tightly to the microtubule surface, are very flexible and do not present a major obstacle to a travelling complex of a vesicle drawn by several motor proteins.
One of the most intriguing facets of microtubule behavior is their dynamic instability, that is, their ability to switch ran- domly between phases of growth and shrinkage (catastrophe and rescue) even when the solution is at an apparent equilibrium (40). This behavior can be monitored and analyzed by video micros- copy (51), it occurs in vivo and in vitro and allows the micro- tubule network to rearrange itself, for example during mitosis. Growth is coupled to the hydrolysis of GTP bound to ~-tubulin, whereas shrinkage is autonomous. These two phases are char- acterized by very different conformations of tubulin at the microtubule ends which can be visualized by time-resolved cryo- electron microscopy (37). Growing microtubules have straight protofilaments at their ends which may protrude for different lengths, shrinking ones have coiled protofilaments, creating the appearance of ram's horns (Fig. 4). This implies, among others, that microtubules are not helical structures but rather grow or shrink along protofilaments. Because the "catastrophe" occurs without nucleotide hydrolysis it can be likened to the release of stored chemical energy, and it has been proposed that this energy could be converted into mechanical work, for example, in pull- ing chromosomes apart during mitosis (9,29). Whether this plays a role in axonal transport is not clear at present.
Microtubule-Associated Protein Tau
As mentioned, MAPs are notoriously elusive structures. They are nearly invisible in negative stain electron microscopy or in unstained cryo-EM images. In thin sections, MAP2 molecules (but not tau or MAP2c) are discernible as "projections" (41),
FIG. 3. Image reconstruction (using the Fourier transform method) of a tubulin wall decorated by kinesin heads, showing four tubulin proto- filaments (in contour lines, each subunit displays a pear-shaped struc- ture). The shaded areas represent the kinesin heads bound to/~-tubulin subunits. The axial spacing between the kinesin heads is 8 nm. The view is from outside a microtubule, the plus end is up (arrow).
FIG. 4. Growing microtubules (a, b) and shrinking microtubules (c) in a thin layer of vitrified ice imaged by cryo-electron microscopy. Note that the growing microtubules have straight protof'daments at their ends, sometimes tapering, and shrinking microtubules have coiled protofila- ments at the ends, indicating a conformational switch during the tran- sition between growth and shrinkage. From Mandelkow et al., 1991, reprinted with permission.

350 MANDELKOW ET AL.
A
Htau23 + A B 2 - 4
K12 + A B 2 - 4 FIG. 5. Glycerol-spray rotary-shadowed images of tau molecules (top) and antiparallel dimers of the re- peat domain (construct K 12) labeled with antibodies at both ends. From Wille et al., 1992c, reprinted with permission.
and the quick-freeze deep-etch technique reveals stub-like pro- jections even for tau (24). To image individual MAPs we found the glycerol-spray rotary-shadow technique to be the most use- ful one. All three MAPs studied so far (MAP2, MAP2c, Tau; see 53,54,55) appear highly elongated by this method, with lengths between 35 nm (tau; Fig. 5a) and 97 nm (MAP2). The repeat domain alone accounts for about 22-25 nm.
A common feature of these molecules is that they tend to associate into dimers and higher oligomers. This is particularly notable with tau or shorter constructs comprising the repeat domain of tau. A homogeneous population of dimers can be prepared by chemical crosslinking and labeled with N-terminal or C-terminal antibodies. These particles appears as dumb-bells in the electron microscopy (Fig. 5b), confirming that the two monomers are roughly in register (otherwise the dimer would be significantly longer than the monomer) and antiparallel (be- cause the antibodies label both ends).
MAPs tend to aggregate into thin fibers which can form a three-dimensional network in solution. The substructure and significance of such aggregates is usually difficult to assess. How- ever, the repeat domain of tau, and in particular, the cross-linked antiparallel dimers are prone to form synthetic paired helical filaments in vitro which by all accounts strongly resemble the PHFs isolated from Alzheimer brains (55; Fig. 6). This is sug- gestive of a PHF assembly model in which the dimer features as an intermediate stage, as depicted in Fig. 7. The antiparallel arrangement of tau subunits in Alzheimer PHFs would also be consistent with biochemical data (32). Two features of the syn- thetic PHFs are noteworthy: First, they form much more easily from tau constructs containing only the repeat region than from whole tau; the latter sometimes assembles into related structures called "dashed minifilaments" (10) but more usually just into fibers of indistinct substructure. Second, it has not been possi- ble so far to demonstrate clearly a dependence of the synthetic PHFs on the state of phosphorylation of tau. Thus, for the mo- ment it is not clear whether tau's hyperphosphorylation is a pre- requisite for PHF assembly in Alzheimer's disease or whether is it merely coincidental.
There is a widespread feeling that pathologic fibers are "amy- loidogenic" in nature, meaning that they consist of proteins with predominant B-structure which leads to the irreversible aggre- gation. Examples of this kind are the fibers formed from the AB peptide in Alzheimer's disease (13,21) or the fibers from prion
protein in Creutzfeld-Jacob's disease (14). Structures with B- strands tend to stain with dyes such as Congo Red or Thiofla- vin S, and because Alzheimer PHFs also stain with these dyes to some extent (43), it was assumed that they, too, contain mainly B-stranded components. This prompted us to investigate the solution structure of tau by spectroscopic or X-ray methods (44). Summarizing the results, under no circumstances did we observe the CD-spectrum expected for a protein with 8- conformation; this holds for several tau isoforms or constructs
i i ! ~ i ~ ! i ¸
FIG. 6. Synthetic PHFs assembled from chemically crosslinked anti- parallel dimers of the repeat domain of tau protein (construct K12). From Wille et al., 1992c, reprinted with permission.
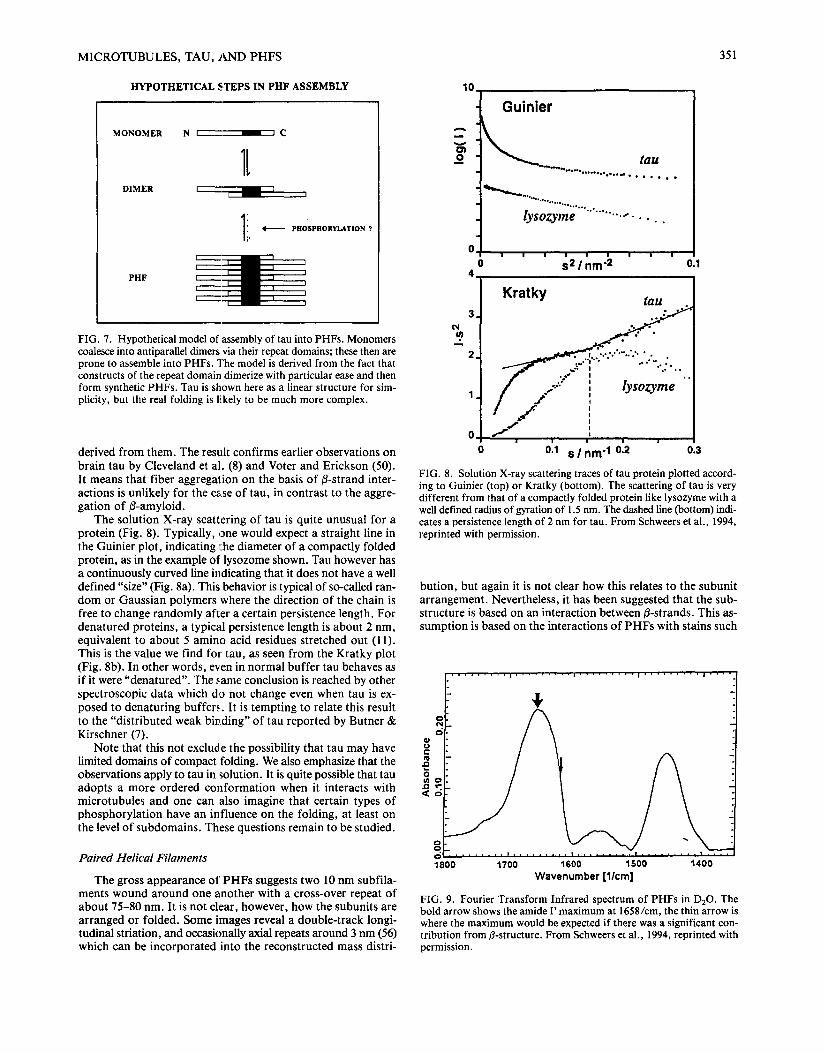
M I C R O T U B U L E S , TAU, A N D P H F S 351
HYPOTHETICAL STEPS IN PHF ASSEMBLY
M O N O M E R
D I M E R
N i I C
1t
( P H O S P ] B O R Y L A T I O N ?
P H F
FIG. 7. Hypothetical model of assembly of tau into PHFs. Monomers coalesce into antiparallel dimers via their repeat domains; these then are prone to assemble into PHFs. The model is derived from the fact that constructs of the repeat domain dimerize with particular ease and then form synthetic PHFs. Tau is shown here as a linear structure for sim- plicity, but the real folding is likely to be much more complex.
derived from them. The result confirms earlier observations on brain tau by Cleveland et al. (8) and Voter and Erickson (50). It means that fiber aggregation on the basis of H-strand inter- actions is unlikely for the ca:se of tau, in contrast to the aggre- gation of 13-amyloid.
The solution X-ray scattering of tau is quite unusual for a protein (Fig. 8). Typically, one would expect a straight line in the Guinier plot, indicating l:he diameter of a compactly folded protein, as in the example of lysozome shown. Tau however has a continuously curved line indicating that it does not have a well defined "size" (Fig. 8a). This behavior is typical of so-called ran- dom or Gaussian polymers where the direction of the chain is free to change randomly after a certain persistence length. For denatured proteins, a typical persistence length is about 2 nm, equivalent to about 5 amino acid residues stretched out (11). This is the value we find for tau, as seen f rom the Kratky plot (Fig. 8b). In other words, even in normal buffer tau behaves as if it were "denatured". The same conclusion is reached by other spectroscopic data which do not change even when tau is ex- posed to denaturing buffers. It is tempting to relate this result to the "distributed weak binding" of tau reported by Butner & Kirschner (7).
Note that this not exclude the possibility that tau may have limited domains of compact folding. We also emphasize that the observations apply to tau in solution. It is quite possible that tau adopts a more ordered conformat ion when it interacts with microtubules and one can also imagine that certain types of phosphorylat ion have an influence on the folding, at least on the level of subdomains. These questions remain to be studied.
Paired Helical Filaments
The gross appearance of PHFs suggests two 10 nm subfila- ments wound around one another with a cross-over repeat of about 75-80 nm. It is not clear, however, how the subunits are arranged or folded. Some images reveal a double-track longi- tudinal striation, and occasionally axial repeats around 3 nm (56) which can be incorporated into the reconstructed mass distri-
10,
Guinier
° % ' ' o . ° , . ° ° O . o , o . ~ • * . • I e o
l y s o z y m e . . . . . . . . . . .
0 ! I i i I I I I I
0 s 2 1 n m - 2 0.1 4.
Kratky M U
c~ ~ u ) 3. ..: " 2. . ,...... ' . '"-.'."" • .
. . . . " B " • o . ° " " .
1 / / /
0 I "~'J
0 ' 011 S / n m "1 012 ' 0.3
FIG. 8. Solution X-ray scattering traces of tat; protein plotted accord- ing to Guinier (top) or Kratky (bottom). The scattering of tau is very different from that of a compactly folded protein like lysozyme with a well defined radius of gyration of 1.5 nm. The dashed line (bottom) indi- cates a persistence length of 2 nm for tau. From Schweers et al., 1994, reprinted with permission.
bution, but again it is not clear how this relates to the subunit arrangement. Nevertheless, it has been suggested that the sub- structure is based on an interaction between/3-strands. This as- sumption is based on the interactions of P H F s with stains such
c 0
o
1800 . . . . . . . . . P . . . . . . . . . 1 , , , - . . . . , I , . . . . . . . . J . . . .
1700 1600 1500 1400 Wavenumber [l/cm] FIG. 9. Fourier Transform Infrared spectrum of PHFs in D20. The bold arrow shows the amide I' maximum at 1658/cm, the thin arrow is where the maximum would be expected if there was a significant con- tribution from/3-structure. From Schweers et al., 1994, reprinted with permission.

352 M A N D E L K O W ET AL.
/!! ~iii ~ iili ~iii
FIG. 10. X-ray patterns of PHFs at two different magnifications. The white spot in the center is the beam stop inter- cepting the direct beam. The diffuse ring w on the left comes from water. The reflections c~ and ~ (smeared out into rings because there is no preferred orientation) correspond to spacings of 9 nm and 3.7 nm. There are no peaks due to a/~ or 13 structure. From Schweers et al., 1994, reprinted with permission.
as Congo Red or Thioflavin S, and on the X-ray reflections typ- ical of/3-structure reported by Kirschner et al. (28).
Considering the lack of 3-structure in soluble tau, this might suggest that tau would adopt a 3-structure during the aggrega- tion process. We therefore wanted to address the issue by two methods. One is FTIR spectroscopy which is sensitive to the stretch vibration of carbonyl bonds and distinguishes between different types of secondary structure. The spectrum is shown in Fig. 9; the maximum is at 1658/cm, the thin arrow indicates where the peak would be expected if there was pronounced /3 structure but this is apparently not the case for PHFs .
Second, we performed X-ray fiber diffraction experiments on PHFs prepared from Alzheimer tissue. X-rays are particu- larly sensitive to periodicities, and different types of secondary structures can be distinguished by their different spacings. For example, c~-helices contain spacings of 0.15 and 0.51 nm due to the axial separation of amino acid residues and the repeat of adjacent gyres, whereas 3-strands are separated by 0.47 nm. A diffraction pattern is shown in Fig. 10. In contrast to most other fibrous structures (we used microtubules for comparison, which contains both c¢ and 13 structure), the P H F pattern is practically f e a t u r e l e s s - n o trace o f either type o f substructure. There is a weak low resolution "reflection" around 9-10 nm (smeared out into a ring because the PHFs lack a preferred orientation) which is probably indicative of the mean diameter of the fibers. There is also a faint reflection around 4 nm which could indicate some axially repeating structure of very low contrast. Taken together, the data are negative in terms of ordered secondary structure but consistent with the results obtained from tau in solution, i.e., even the assembly into PHFs does not introduce noticeable ordering of the structure. To reconcile this with previous data, we note that Congo Red staining does not necessarily require
extended/3 strands; occasional 3-turns could be enough to cause the binding of the dye. Second, the 3-type reflection observed by Kirschner et al. (28) could be ascribed to a contamination of amyloid fibers which were probably present in the preparations used at the time.
In summary, at this moment the substructure of tau and PHFs remains enigmatic. None of the usual tools of structure analy- sis has yielded a reliable indicator of how the subunits are folded and how they are built into the structure. It is equally unclear how phosphorylation affects the assembly of tau into PHFs , or how tau interacts structurally with microtubules. It is becoming clear that the repeat region of tau binds only weakly to micro- tubules (and therefore does not really deserve the name "micro- tubule-binding domain," see 19); rather it requires the presence of the flanking regions which act as "jaws". On the other hand, phosphorylation of a single residue (Ser262) in the repeat region has a surprisingly strong influence on tau's binding to micro- tubules (5), and, finally, the dimerization of the repeat region promotes the subsequent assembly into PHFs (55). How these features are related to one another remains to be seen.
ACKNOWLEDGEMENTS
We thank K. Kosik (Harvard Medical School) for the clone of kine- sin, and M. Goedert (MRC Cambridge) for the clones of tau. Brain tis- sue was generously provided by the Bryan Alzheimer Disease Research Center (Duke University Medical Center, Durham, NC), the Brain Tis- sue Resource Center (McLean Hospital/Harvard Medical School, Bel- mont, MA), and the Alzheimer Research Center (University of Rochester Medical School, Rochester, NY). This work was supported by Bundes- ministerium fiir Forschung und Technologie (BMFT) and the Deutsche Forschungsgemeinschaft (DFG).
REFERENCES
1. Allen, C.; Borisy, G. G. (1974). Structural polarity and directional growth of microtubules of Chlamydomonas flagella. J. Mol. Biol. 90:381-402; 1974.
2. Amos, L. A.; Klug, A. Arrangement of subunits in flageUar micro- tubules. J. Cell Sci. 14:523-549; 1974.
3. Baas, P. W.; Deitch, J. S.; Black, M. M.; Banker, G. A. Polarity orientation of microtubules in hippocampal neurons: Uniformity
in the axon and nonuniformity in the dendrite. Proc. Natl. Acad. Sci. USA 85:8335-8339; 1988.
4. Baas, P. W.; Joshi, H. C. Gamma-tubulin distribution in the neu- ron: Implications for the origins of neuritic microtubules. J. Cell Biol. 119:171-178; 1992.
5. Biernat, J.; Gustke, N.; Drewes, G.; Mandelkow, E.-M.; Man- delkow, E. Phosphorylation of serine 262 strongly reduces the bind-

M I C R O T U B U L E S , TAU, A N D P H F S 353
ing of tau protein to microtubules: Distinction between PHF-like immunoreactivity and microtubule binding. Neuron 11 : 153-163; 1993.
6. Binder, L. I.; Dentler, W. L.; Rosenbaum, J. L. Assembly of chick brain tubulin onto flagellar axonemes of Chlamydomonas and sea urchin sperm. Proc. Natl. Acad. Sci. USA 72:1122-1126; 1975.
7. Butner, K. A.; Kirschner, M. W. Tau-protein binds to microtubules through a flexible array of distributed weak sites. J. Cell Biol. I 15: 717-730; 1991.
8. Cleveland, D. W.; Hwo, S.-Y.; Kirschner, M. W. Physical and chem- ical properties of purified t~tu factor and the role of tau in micro- tubule assembly. J. Mol. Biol. 116:227-247; 1977.
9. Coue, M.; Lombillo, V. A.; McIntosh, J. R. Microtubule depoly- merization promotes particle and chromosome movement in vitro. J. Cell Biol. 112:1165-1175; 1991.
10. Crowther, R. A.; Olesen, O. F.; Smith, M. J.; Jakes, R.; Goedert, M. Assembly of Alzheimer-like filaments from full-length tau pro- tein. FEBS Lett. 337:135-138; 1994.
11. Damaschun, G.; Damaschun, H.; Gast, K.; Misselwitz, R.; Miiller, J. J.; Pfeil, W.; Zirwer, D. (Sold denaturation-induced conforma- tional changes in phosphoglycerate kinase from yeast. Biochem. 32:7739-7746; 1993.
12. Euteneuer, U.; Mclntosh, J. R. Polarity of some motility-related microtubules. Proc. Natl. Acad. Sci. USA 78:372-376; 1981.
13. Fraser, P. E.; Nguyen, J. T.; Surewicz, W. K.; Kirschner, D. A. pH- dependent structural transitions of Alzheimer amyloid peptides. Bio- phys. J. 60:1190-1201; 1991.
14. Gasset, M.; Baldwin, M. A.; Fletterick, R. J.; Prusiner, S. B. Per- turbation of the secondary slructure of the scarpie prion protein un- der conditions that alter infectivity. Proc. Natl. Acad. Sci. USA 90:1-5; 1993.
15. Gelles, J.; Schnapp, B. J.; Sheetz, M. P. Tracking kinesin-driven movements with nanometre-scale precision. Nature (Lond.) 331: 450-453; 1988.
16. Goedert, M.; Spillantini, M.; Jakes, R.; Rutherford, D.; Crowther, R. A. Multiple isoforms of human microtubule-associated protein- tau: Sequences and localization in neurofibrillary tangles of Alzheimers-disease. Neuron 3:519-526; 1989.
17. Goldstein, L. S. B. With apologies to Scheherazade: Tails of 1001 kinesin motors. Annu. Rev. Genetics 27:319-351; 1993.
18. Greenberg, S. G.; Davies, P. A preparation of Alzheimer paired heli- cal filaments that displays distinct tau-proteins by polyacrylamide- gel electrophoresis. Proc. Natl. Acad. Sci. USA 87:5827-5831; 1990.
19. Gustke, N.; Trinczek, B.; Biernat, J.; Mandelkow, E.-M.; Man- delkow, E. Domains of tau protein and interactions with micro- tubules. Biochem. 33:9511--9522; 1994.
20. Heins, S.; Song, Y.-H.; Wille, H.; Mandelkow, E.; Mandelkow, E.-M. Effect of MAP2, MAP2c, and tau on kinesin-dependent microtubule motility. J. Cell Sci. S14:121-124; 1991.
21. Hilbich, C.; Kisters-Woike, B.; Reed, J.; Masters, C.; Beyreuther, K. Substitutions of hydropb obic amino-acids reduce the amyloido- genicity of Alzheimer's disease/3A4 peptides. J. Mol. Biol. 228:460- 473; 1992.
22. Himmler, A.; Drechsel, D.; Kirschner, M.; Martin, D. Tau consists of a set of proteins with repeated C-terminal microtubule-binding domains and variable N-terminal domains. Mol. Cell. Biol. 9:1381- 1388; 1989.
23. Hirokawa, N. Cross-linker system between neurofilaments, micro- tubules, and membranous organelles in frog axons revealed by the quick-freeze, deep-etching method. J. Cell Biol. 94:129-142; 1982.
24. Hirokawa, N.; Shiomura, Y.; Okabe, S. Tau proteins: The molec- ular structure and mode of binding to microtubules. J. Cell Biol. 107:1449-1459; 1988.
25. Jarvis, J. A.; Craik, D. J.; Wilce, M. C. X-ray diffraction studies of fibrils formed from peptide fragments of transthyretin. Biochem. Biophys. Res. Commun. 192:991-998; 1993.
26. Kamimura, S.; Kamiya, R. High-frequency vibration in flagellar axonemes with amplitudes reflecting the size of tubulin. J. Cell Biol. 116:1443-1454; 1992.
27. Kamimura, S.; Mandelkow, E. Tubulin protofilaments and kinesin- dependent motility. J. Cell Biol. 118:865-875; 1992.
28. Kirschner, D. A.; Abraham, C.; Selkoe, D. J. X-ray diffraction from intraneural paired helical filaments and extraneural amyloid fibers in Alzheimer disease indicates cross-/3 conformation. Proc. Natl. Acad. Sci. USA 83:503-507; 1986.
29. Koshland, D. E.; Mitchison, T. J.; Kirschner, M. W. Polewards chromosome movement driven by microtubule depolymerization in vitro. Nature (Lond.) 331:499-504; 1988.
30. Kosik, K.; Orecchio, L.; Binder, L.; Trojanowski, J.; Lee, V.; Lee, G. Epitopes that span the tau molecule are shared with paired heli- cal filaments. Neuron 1:817-825; 1988.
31. Kosik, K. S.; Orecchio, L. D.; Schnapp, B.; Inouye, H.; Neve, R. L. The primary structure and analysis of the squid kinesin heavy-chain. J. Biol. Chem. 265:3278-3283; 1990.
32. Ksiezak-Reding, H.; Yen, S. H. Structural stability of paired heli- cal filaments requires microtubule-binding domains of tau: A model for self-association. Neuron 6:717-728; 1991.
33. Kuo, S. C.; Sheetz, M. P. Force of single kinesin molecules mea- sured with optical tweezers. Science 260:232-234; 1993.
34. Lee, G.; Cowan, N.; Kirschner, M. The primary structure and het- erogeneity of tau protein from mouse brain. Science 239:285-288; 1988.
35. Lopez, L. A.; Sheetz, M. P. Steric inhibition of cytoplasmic dynein and kinesin motility by MAP2. Cell Mot. Cytoskel. 24:1-16; 1993.
36. Mandelkow, E.; Thomas, J.; Cohen, C. Microtubule structure at low resolution by X-ray diffraction. Proc. Natl. Acad. Sci. USA 74:3370-3374; 1977.
37. Mandelkow, E.-M.; Mandelkow, E.; Milligan, R. A. Microtubule dynamics and microtubule caps: A time-resolved cryo-electron mi- croscopy study. J. Cell Biol. 114:977-991; 1991.
38. Mandelkow, E.; Mandelkow, E.-M. Microtubule structure. Curr. Opin. Struct. Biol. 4:171-179; 1994.
39. Marx, A.; Song, Y. H.; Mandelkow, E.-M.; Kamimura, S.; Thor- m~ihlen, M.; Mandelkow, E. Reconstruction of the tubulo-kinesin complex and implications for microtubule motility (submitted for publication).
40. Mitchison, T.; Kirschner, M. Dynamic instability of microtubule growth. Nature (Lond.) 312:237-242; 1984.
41. Murphy, D. B.; Borisy, G. G. Association of high molecular weight proteins with microtubules and their role in microtubule assembly in vitro. Proc. Natl. Acad. Sci. USA 72:2696-2700; 1975.
42. Ray, S.; Meyhofer, E.; Milligan, R. A.; Howard, J. Kinesin fol- lows the microtubules protofilament axis. J. Cell Biol. 121:1083- 1093; 1993.
43. Roher, A.; Palmer, K.; Chau, V.; Ball, M. Isolation and chemical characterization of Alzheimer's disease paired helical filament cyto- skeletons: Differentiation from amyloid plaque core protein. J. Cell Biol. 107:2703-2716; 1988.
44. Schweers, O.; Sch6nbrunn-Hanebeck, E.; Marx, A.; Mandelkow, E. Structural studies of tau protein and Alzheimer paired helical filaments: Extended random configuration without evidence for 13 structure. J. Biol. Chem. (in press).
45. Song, Y.-H.; Mandelkow, E. Recombinant kinesin motor domain binds to/3-tubulin and decorates microtubules with a B-surface lat- tice. Proc. Natl. Acad. Sci. USA 90:1671-1675; 1993.
46. Song, Y. H.; Mandelkow, E. Anatomy of flagellar microtubules: Polarity, seam, junctions, and lattice (submitted for publication).
47. Svoboda, K.; Schmidt, C. F.; Schnapp, B. J.; Block, S. M. Direct observation of kinesin stepping by optical trapping interferometry. Nature 365:721-727; 1993.
48. Vale, R. D.; Toyoshima, Y. Y. Rotation and translocation of micro- tubules in vitro induced by dyneins from Tetrahymena. Cell 52:459- 469; 1988.
49. Von Massow, A.; Mandelkow, E.-M.; Mandelkow, E. Interaction between kinesin, microtubules, and microtubule-associated protein 2. Cell Mot. Cytoskel. 14:562-571; 1989.
50. Voter, W. A.; Erickson, H. P. Electron microscopy of MAP2 (Mic rotubule-associated protein 2). J. Ultrastruct. Res. 80:374-382; 1982.
51. Walker, R.; O'Brien, E.; Pryer, N.; Soboeiro, M.; Voter, W.; Erick- son, H.; Salmon, E. Dynamic instability of individual microtubules analyzed by video light microscopy: Rate constants and transition frequencies. J. Cell Biol. 107:1437-1448; 1988.

354 M A N D E L K O W ET AL.
52. Walker, R.; Sheetz, M. P. Cytoplasmic microtubule-associated mo- tors. Annu. Rev. Biochem. 62:429-451; 1993.
53. Wille, H.; Mandelkow, E.-M.; Dingus, J.; Vallee, R.; Binder, L.; Mandelkow, E. Domain structure and antiparallel dimers of micro- tubule-associated protein 2 (MAP2). J. Struct. Biol. 108:49-61; 1992.
54. Wille, H.; Mandelkow, E.-M.; Mandelkow, E. The juvenile micro- tubule-associated protein MAP2c is a rod-like molecule that forms antiparallel dimers. J. Biol. Chem. 267:10737-10742; 1992.
55. Wille, H.; Drewes, G.; Biernat, J.; Mandelkow, E.-M.; Mandelkow, E. Alzheimer-like paired helical filaments and antiparallel dimers formed from microtubule-associated protein tau in vitro. J. Cell Biol. 118:573-584; 1992.
56. Wischik, C.; Crowther, R.; Stewart, M.; Roth, M. Subunit struc- ture of paired helical filaments in Alzheimer's disease. J. Cell Biol. 100:1905-1912; 1985.


















