
Novel and Expanded Roles for MAPK Signaling in Arabidopsis...
12
Novel and Expanded Roles for MAPK Signaling in Arabidopsis Stomatal Cell Fate Revealed by Cell Type–Specific Manipulations C W Gregory R. Lampard, a,1 Wolfgang Lukowitz, b Brian E. Ellis, c and Dominique C. Bergmann a,2 a Department of Biology, Stanford University, Stanford, California 94305 b Department of Plant Biology, University of Georgia, Athens, Georgia 30602 c Michael Smith Laboratory, University of British Columbia, Vancouver, British Columbia, Canada V6T 1Z4 Mitogen-activated protein kinase (MAPK) signaling networks regulate numerous eukaryotic biological processes. In Arabidopsis thaliana, signaling networks that contain MAPK kinases MKK4/5 and MAPKs MPK3/6 function in abiotic and biotic stress responses and regulate embryonic and stomatal development. However, how single MAPK modules direct specific output signals without cross-activating additional downstream processes is largely unknown. Studying relation- ships between MAPK components and downstream signaling outcomes is difficult because broad experimental manip- ulation of these networks is often lethal or associated with multiple phenotypes. Stomatal development in Arabidopsis follows a series of discrete, stereotyped divisions and cell state transitions. By expressing a panel of constitutively active MAPK kinase (MAPKK) variants in discrete stomatal lineage cell types, we identified a new inhibitory function of MKK4 and MKK5 in meristemoid self-renewal divisions. Furthermore, we established roles for MKK7 and MKK9 as both negative and (unexpectedly) positive regulators during the major stages of stomatal development. This has expanded the number of known MAPKKs that regulate stomatal development and allowed us to build plausible and testable subnetworks of signals. This in vivo cell type–specific assay can be adapted to study other protein families and thus may reveal insights into other complex signal transduction pathways in plants. INTRODUCTION Mitogen-activated protein kinase (MAPK) signaling networks are found in all eukaryotic organisms and regulate fundamental aspects of biology, including but not limited to cell division, initiation of developmental pathways, response to abiotic and biotic stresses, and triggering programmed cell death (reviewed in Widmann et al., 1999; Chen and Thorner, 2007; Colcombet and Hirt, 2008). In plants, MAPK networks regulate a similar array of processes, but genomic sequence data have revealed that, in comparison to other eukaryotes, plant genomes encode en- larged gene families of MAPK kinase kinases (MAPKKKs), MAPK kinases (MAPKKs), and MAPKs (Ichimura et al., 2002; Hamel et al., 2006). Furthermore, large-scale gene expression studies indicate that many of these genes are broadly expressed throughout the plant (Schmid et al., 2005; Schmidt, 2007). These extended gene families may have evolved to allow plants, which are sessile, to sense and respond to a continuous flux of environmental conditions. In support of this, the majority of plant MAPK signaling components studied to date have been asso- ciated with responses to abiotic and biotic stresses (Colcombet and Hirt, 2008). Constitutive and ectopic modulation of MAPK signaling path- ways in plants is typically associated with pleiotropic phenotypes and/or is lethal (Jin et al., 2003; Liu et al., 2003; Popescu et al., 2009). These effects may be the result of indiscriminate activa- tion of multifunctional kinases that have discrete functions in different cell types. Therefore, to both characterize specific functions of MAPK networks in plants and learn how signal integrity is maintained within these networks, it was necessary to devise a system that allows for cell type–specific modulation of MAPK signaling while providing an accessible means to analyze the effects of these changes. Stomatal development is an ideal system to study discrete aspects of MAPK signaling networks. Stomata are specialized structures found in the epidermis of aerial tissues of land plants and are the primary conduit for gas and water exchange. MAPK signaling has roles in conveying intrinsic developmental cues to regulate stomatal development and relaying extrinsic environ- mental signals that influence stomatal physiology and develop- ment. For example, MAPK signaling networks are positive regulators of environmental stress–induced stomatal closure and negative regulators of stomatal development (Bergmann et al., 2004; Wang et al., 2007; Neill et al., 2008). Besides influencing stomatal behavior, environmental conditions are also capable of influencing stomatal development (Coupe et al., 2006; 1 Current address: Department of Biology and Health Sciences, Pace University, 861 Bedford Road, Pleasantville, NY 10570. 2 Address correspondence to [email protected]. The author responsible for distribution of materials integral to the findings presented in this article in accordance with the policy described in the Instructions for Authors (www.plantphysiol.org) is: Dominique C. Bergmann ([email protected]). C Some figures in this article are displayed in color online but in black and white in the print edition. W Online version contains Web-only data. www.plantcell.org/cgi/doi/10.1105/tpc.109.070110 This article is a Plant Cell Advance Online Publication. The date of its first appearance online is the official date of publication. The article has been edited and the authors have corrected proofs, but minor changes could be made before the final version is published. Posting this version online reduces the time to publication by several weeks. The Plant Cell Preview, www.aspb.org ã 2009 American Society of Plant Biologists 1 of 12
Transcript of Novel and Expanded Roles for MAPK Signaling in Arabidopsis...

Novel and Expanded Roles for MAPK Signalingin Arabidopsis Stomatal Cell Fate Revealed by CellType–Specific Manipulations C W
Gregory R. Lampard,a,1 Wolfgang Lukowitz,b Brian E. Ellis,c and Dominique C. Bergmanna,2
a Department of Biology, Stanford University, Stanford, California 94305b Department of Plant Biology, University of Georgia, Athens, Georgia 30602cMichael Smith Laboratory, University of British Columbia, Vancouver, British Columbia, Canada V6T 1Z4
Mitogen-activated protein kinase (MAPK) signaling networks regulate numerous eukaryotic biological processes. In
Arabidopsis thaliana, signaling networks that contain MAPK kinases MKK4/5 and MAPKs MPK3/6 function in abiotic and
biotic stress responses and regulate embryonic and stomatal development. However, how single MAPK modules direct
specific output signals without cross-activating additional downstream processes is largely unknown. Studying relation-
ships between MAPK components and downstream signaling outcomes is difficult because broad experimental manip-
ulation of these networks is often lethal or associated with multiple phenotypes. Stomatal development in Arabidopsis
follows a series of discrete, stereotyped divisions and cell state transitions. By expressing a panel of constitutively active
MAPK kinase (MAPKK) variants in discrete stomatal lineage cell types, we identified a new inhibitory function of MKK4 and
MKK5 in meristemoid self-renewal divisions. Furthermore, we established roles for MKK7 and MKK9 as both negative and
(unexpectedly) positive regulators during the major stages of stomatal development. This has expanded the number of
known MAPKKs that regulate stomatal development and allowed us to build plausible and testable subnetworks of signals.
This in vivo cell type–specific assay can be adapted to study other protein families and thus may reveal insights into other
complex signal transduction pathways in plants.
INTRODUCTION
Mitogen-activated protein kinase (MAPK) signaling networks are
found in all eukaryotic organisms and regulate fundamental
aspects of biology, including but not limited to cell division,
initiation of developmental pathways, response to abiotic and
biotic stresses, and triggering programmed cell death (reviewed
in Widmann et al., 1999; Chen and Thorner, 2007; Colcombet
and Hirt, 2008). In plants, MAPK networks regulate a similar array
of processes, but genomic sequence data have revealed that, in
comparison to other eukaryotes, plant genomes encode en-
larged gene families of MAPK kinase kinases (MAPKKKs), MAPK
kinases (MAPKKs), and MAPKs (Ichimura et al., 2002; Hamel
et al., 2006). Furthermore, large-scale gene expression studies
indicate that many of these genes are broadly expressed
throughout the plant (Schmid et al., 2005; Schmidt, 2007). These
extended gene families may have evolved to allow plants, which
are sessile, to sense and respond to a continuous flux of
environmental conditions. In support of this, the majority of plant
MAPK signaling components studied to date have been asso-
ciated with responses to abiotic and biotic stresses (Colcombet
and Hirt, 2008).
Constitutive and ectopic modulation of MAPK signaling path-
ways in plants is typically associatedwith pleiotropic phenotypes
and/or is lethal (Jin et al., 2003; Liu et al., 2003; Popescu et al.,
2009). These effects may be the result of indiscriminate activa-
tion of multifunctional kinases that have discrete functions in
different cell types. Therefore, to both characterize specific
functions of MAPK networks in plants and learn how signal
integrity is maintained within these networks, it was necessary to
devise a system that allows for cell type–specific modulation of
MAPK signaling while providing an accessible means to analyze
the effects of these changes.
Stomatal development is an ideal system to study discrete
aspects of MAPK signaling networks. Stomata are specialized
structures found in the epidermis of aerial tissues of land plants
and are the primary conduit for gas and water exchange. MAPK
signaling has roles in conveying intrinsic developmental cues to
regulate stomatal development and relaying extrinsic environ-
mental signals that influence stomatal physiology and develop-
ment. For example, MAPK signaling networks are positive
regulators of environmental stress–induced stomatal closure
and negative regulators of stomatal development (Bergmann
et al., 2004; Wang et al., 2007; Neill et al., 2008). Besides
influencing stomatal behavior, environmental conditions are also
capable of influencing stomatal development (Coupe et al., 2006;
1Current address: Department of Biology and Health Sciences, PaceUniversity, 861 Bedford Road, Pleasantville, NY 10570.2 Address correspondence to [email protected] author responsible for distribution of materials integral to thefindings presented in this article in accordance with the policy describedin the Instructions for Authors (www.plantphysiol.org) is: Dominique C.Bergmann ([email protected]).CSome figures in this article are displayed in color online but in blackand white in the print edition.WOnline version contains Web-only data.www.plantcell.org/cgi/doi/10.1105/tpc.109.070110
This article is a Plant Cell Advance Online Publication. The date of its first appearance online is the official date of publication. The article has been
edited and the authors have corrected proofs, but minor changes could be made before the final version is published. Posting this version online
reduces the time to publication by several weeks.
The Plant Cell Preview, www.aspb.org ã 2009 American Society of Plant Biologists 1 of 12

Casson and Gray, 2008; Casson et al., 2009). In Arabidopsis
thaliana, increased carbon dioxide levels typically decrease the
overall stomatal density (number of stomata per unit area)
(Coupe et al., 2006), whereas high light intensities increase the
stomatal index (number of stomata relative to the total number of
cells per unit area; Casson et al., 2009). The MAPK network that
contains MKK4/5 and MPK3/6 regulates both the responses to
environmental conditions and overall stomatal development
(Bergmann et al., 2004; Wang et al., 2007; Colcombet and Hirt,
2008; Lampard et al., 2008). Linking a stress-activated MAPK
module to the negative regulation of developmental processes is
not surprising; plants arrest development in response to abiotic
and biotic stresses as evidenced by stress-induced downregu-
lation of developmentally associated gene expression (Kultz,
2005; Baena-Gonzalez et al., 2007; Baena-Gonzalez and Sheen,
2008). Thus, using a common MAPK module to regulate both
stress responses and stomatal development could allow rapid
modulation of developmental processes in response to stresses.
Arabidopsis stomatal development follows a stereotyped
pathway regulated by several receptor proteins, putative ligands,
a MAPK signaling module consisting (at a minimum) of a
MAPKKK,YODA (YDA), twoMAPKKs, MKK4/5, and twoMAPKs,
MPK3/6, anda series of transcription factors (Figure 1) (Bergmann
et al., 2004; Ohashi-Ito and Bergmann, 2006; Bergmann and
Sack, 2007; Hara et al., 2007; MacAlister et al., 2007; Pillitteri
et al., 2007; Wang et al., 2007; Kanaoka et al., 2008). This
particular MAPK module, which we refer to as the YDA MAPK
module for simplicity, negatively regulates stomatal develop-
ment, as illustrated by findings showing that artificial activation of
the YDA MAPK module at both MAPKKK and MAPKK levels
results in the plant creating an epidermis consisting only of
pavement cells (Bergmann et al., 2004;Wang et al., 2007).
Conversely, inhibition of the MAPK module using null alleles or
inducible RNA interference constructs correlates with stomatal
overproliferation and clustering (Bergmann et al., 2004; Wang
et al., 2007). These results indicate that the YDA MAPK module
functions to inhibit entry into the stomatal lineage.
More recent work indicates that the YDA MAPK module
prevents entry into the stomatal lineage by regulating the phos-
phorylation state of the basic helix-loop-helix (bHLH) protein
SPEECHLESS (SPCH) (Lampard et al., 2008). After entry into the
stomatal lineage, stomatal development is regulated by the
related and sequentially expressed bHLHs, MUTE and FAMA
(Figures 1A to 1C), which like SPCH, function as positive
regulators of development (Ohashi-Ito and Bergmann, 2006;
MacAlister et al., 2007; Pillitteri et al., 2007; Lampard et al., 2008).
Figure 1. Diagram of Stomatal Lineage Development and Gene Expression Patterns.
Arabidopsis stomatal development follows a three-step stereotyped pathway that involves a series of asymmetric and symmetric cell divisions. Entry
into the stomatal lineage is negatively regulated by a MAPK module containing YDA (MAPKKK), MKK4 and MKK5 (MAPKKs), and MPK3 and MPK6
(MAPKs). Progression through the developmental pathway is positively influenced by sequentially acting bHLH transcription factors, SPCH, MUTE, and
FAMA, which regulate entry (1), progression through (2), and terminal differentiation of guard cell development (3), respectively. The expression of green
fluorescent protein (GFP)-tagged transcriptional reporters of each of SPCH (A), MUTE (B), and FAMA (C) coincides with each major developmental
transition. Bars = 50 mm.
2 of 12 The Plant Cell

While Wang et al. (2007) identified components that function
downstream of YDA (MKK4/5 and MPK3/6) to regulate stomatal
development, the approach of non-cell type–specific and simul-
taneous induction ofMAPK activity did not enable them to assign
discrete MAPK functions to specific stomatal lineage cell types.
Here, we describe a targeted approach to address the issue of
cell type specificity in MAPK signaling. We have used the
promoters of the genes encoding the stomatal bHLH proteins
SPCH, MUTE, and FAMA to individually express a constitutively
active (CA) YDA variant (CA-YDA) and a panel of CA-MAPKKs
beginning in either meristemoid mother cells (MMCs), meriste-
moid cells, or guard mother cells (GMCs). This strategy enabled
us to activate MAPK signaling in specific stomatal lineage cell
types and has resulted in our identification of functions for MAPK
signaling as both a negative and a positive regulator during the
major stages of stomatal development. The 26 separate MAPK
pathway manipulations described here have both expanded the
repertoire of known MAPKKs affecting stomatal development
and have allowed us to propose plausible and testable subnet-
works of signal components. Because the cell type–specific in
vivo assay used in this study can readily be adapted to the study
of other protein families, it has the potential to deconvolve other
similarly complex signal transduction pathways in plants.
RESULTS
Macroscopic yda Seedling Phenotypes Are Separable
A major limitation of studying MAPK signaling in plants has been
that the typical modes of analysis (observing phenotypes asso-
ciated with either loss-of-function mutants or those arising from
plants with constitutively activatedMAPK signaling networks) do
not allow cell-specific resolution of function. In addition, ubiqui-
tous perturbation of broadly expressed, multifunctional proteins
can induce misleading phenotypic defects that may accumulate
over time. For example, yda loss-of-function plants show dra-
matic stomatal clustering phenotypes (Bergmann et al., 2004;
Wang et al., 2007). However, these plants also display hyper-
activation of MPK3/6, embryonic malformations, and severe
dwarfism (G.R. Lampard, unpublished data; Bergmann et al.,
2004; Lukowitz et al., 2004). Therefore, to identify specific
functions of YDA (and associated downstream signaling mod-
ules) in regulating stomatal development, we needed to separate
the range of phenotypes associated with altering YDA signaling.
Mosaic approaches, in which genes are selectively over- or
inactivated in specific tissues, have aided in the resolution of
complex phenotypes (for example, in Drosophila melanogaster;
Xu and Rubin, 1993; Blair, 2003), but these techniques are both
technically challenging in plants and ill-suited for the specific
lineage relationships among the stomatal precursors.
We hypothesized that we could analyze the effects of dimin-
ished YDA signaling specifically in cells about to enter the
stomatal lineage (MMCs) using the SPCH promoter to express
a dominant-negative YDA construct (SPCHpro:DN-YDA; see
Methods). Phenotypic analysis of 10-d-old seedlings revealed
that like yda null plants (Figure 2B), SPCHpro:DN-YDA plants also
have excessive and clustered stomata (Figure 2C). However, as
would be predicted from the SPCH expression pattern (Figure
1A), not all cells in the epidermis are affected, and the seedlings
do not show the dwarfism associated with a systemic lack of
YDA (Figure 2A). When YDA signaling was activated beginning in
MMCs using a constitutively active variant of YDA (SPCHpro:CA-
YDA), the resulting transgenic plants created an epidermis
devoid of guard cells (Figure 2E). This result was identical to
the phenotype produced by YDApro:CA-YDA (Bergmann et al.,
2004). However, the additional developmental phenotypes as-
sociated with broad activation of YDA, such as partially fused
cotyledons, were not observed in the SPCHpro:CA-YDA plants,
indicating that induced YDA signaling was confined to the
stomatal lineage (Figure 2A) (Bergmann et al., 2004; Lukowitz
et al., 2004). Individuals within these transgenic populations
display variability in the strength of this phenotype, and the
strongest phenotypic classes (completely lacking stomata) die
before producing progeny. Therefore, in this and all subsequent
experiments, we characterized phenotypes in large T1 popula-
tions where every individual represented an independent trans-
formation event. In SPCHpro:CA-YDA T1 transgenics, 16/79
plants had no stomata, while the remaining plants had reduced
or normal numbers of stomata. These data suggested that
manipulation of MAPK signaling by expressing dominant-
negative or constitutively active kinase variants under the control
of cell type–specific promoters could allow us to study discrete
aspects of MAPK signaling without inducing pleiotropic pheno-
types.
Cell Type–Specific Activation of the YDA Signaling Pathway
Reveals YDA Functions in Each Stage of
Stomatal Development
Because the genes that have been reported to comprise the YDA
signaling module (YDA, MKK4/5, and MPK3/6) are expressed
throughout the stomatal lineage (Bergmann et al., 2004), we
sought to determine if the YDA signaling module was capable of
regulating additional aspects of stomatal development. To ad-
dress this, we also used the MUTE and FAMA promoters to
initiate expression of CA-YDA in meristemoids and GMCs,
respectively.
Plants expressing the MUTEpro:CA-YDA construct also fail to
produce mature stomata (Figure 2F). Whereas activation of YDA
signaling via SPCHpro:CA-YDA blocked entry into the stomatal
lineage, activation of YDA beginning in meristemoids (MUTEpro:
CA-YDA) arrests stomatal development at a later stage. Here,
while being devoid of guard cells, the epidermis is comprised of
both epidermal pavement cells and smaller, meristemoid-like
cells (Figure 2F). As with SPCHpro:CA-YDA expression, a range
of phenotypes was associated with MUTEpro:CA-YDA expres-
sion, and complete inhibition of stomatal development occurred
in 12/68 T1 transgenics. We attempted to study the phenotype
associatedwith diminished YDA activity in these cells by express-
ing a DN-YDA variant beginning in meristemoids (MUTEpro:
DN-YDA). However, we were unable to recover transformants
among >100,000 seeds from three independent transformations.
The phenotypes resulting fromYDA activation inmeristemoids
(MUTEpro:CA-YDA) were consistent with previously reported
functions of YDA as a negative regulator of stomatal
MAPK Regulation of Stomatal Cell Fate 3 of 12
development. The expanded capability of YDA to regulate addi-
tional stages of stomatal development led us to hypothesize that
YDA activity in GMCs would also inhibit stomatal development.
However, contrary to our predictions, activation of YDA begin-
ning in GMCs via the expression of a FAMApro:CA-YDA trans-
gene promoted excess guard cell formation. Small clusters of
guard cells were observed throughout the epidermis, which
appeared otherwise normal (Figure 2G). These surprising results
were confirmed by reducing YDA signaling with FAMApro:DN-
YDA. Consistent with the results obtained when YDA signaling
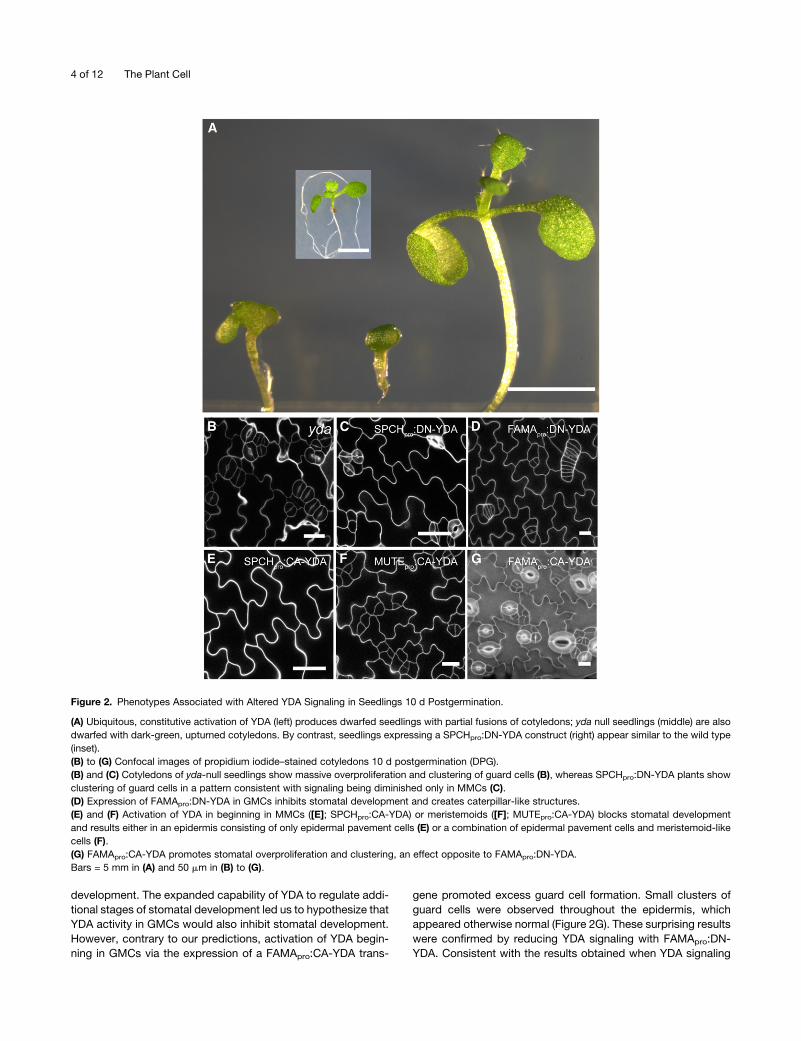
Figure 2. Phenotypes Associated with Altered YDA Signaling in Seedlings 10 d Postgermination.
(A) Ubiquitous, constitutive activation of YDA (left) produces dwarfed seedlings with partial fusions of cotyledons; yda null seedlings (middle) are also
dwarfed with dark-green, upturned cotyledons. By contrast, seedlings expressing a SPCHpro:DN-YDA construct (right) appear similar to the wild type
(inset).
(B) to (G) Confocal images of propidium iodide–stained cotyledons 10 d postgermination (DPG).
(B) and (C) Cotyledons of yda-null seedlings show massive overproliferation and clustering of guard cells (B), whereas SPCHpro:DN-YDA plants show
clustering of guard cells in a pattern consistent with signaling being diminished only in MMCs (C).
(D) Expression of FAMApro:DN-YDA in GMCs inhibits stomatal development and creates caterpillar-like structures.
(E) and (F) Activation of YDA in beginning in MMCs ([E]; SPCHpro:CA-YDA) or meristemoids ([F]; MUTEpro:CA-YDA) blocks stomatal development
and results either in an epidermis consisting of only epidermal pavement cells (E) or a combination of epidermal pavement cells and meristemoid-like
cells (F).
(G) FAMApro:CA-YDA promotes stomatal overproliferation and clustering, an effect opposite to FAMApro:DN-YDA.
Bars = 5 mm in (A) and 50 mm in (B) to (G).
4 of 12 The Plant Cell

was activated in GMCs, diminished YDA signaling arrested
stomatal development prior to guard cell formation; the epider-
mis of T3 progeny contained caterpillar-like structures strongly
reminiscent of those observed in fama null or flp myb88mutants
(Figure 2D) (Lai et al., 2005; Ohashi-Ito and Bergmann, 2006).
Activation of MKK4 and MKK5 Inhibits Stomatal
Development at Multiple Stages
Given these additional roles of YDA in regulating stomatal
development identified here, we next questioned which down-
stream MAPKKs regulate specific stages (entry, progression,
and terminal differentiation of stomata). First, we sought to
determine the extent towhichMKK4 andMKK5activity regulates
each developmental stage. Constitutively active versions of
MKK4 and MKK5 (CA-MKK4 and CA-MKK5, respectively) were
created by substituting the phosphorylatable S/T residues with
phosphomimic E/D residues (see Methods; Popescu et al.,
2009). Each of CA-MKK4 and CA-MKK5 was expressed in
wild-type Columbia-0 (Col-0) plants during discrete stages of
stomatal development using the SPCH, MUTE, or FAMA pro-
moters. As with YDA, activation of either MKK4 (SPCHpro:CA-
MKK4) or MKK5 (SPCHpro:CA-MKK5) beginning in MMCs pre-
vents entry into the stomatal lineage (Figures 3A and 3D),
and expression of either construct beginning in meristemoids
(MUTEpro:CA-MKK4 or MUTEpro:CA-MKK5) results in a buildup
of arrested meristemoid-like cells (Figures 3B and 3E). These
results are consistent with previously described functions of a
Nicotiana tabacum MEK2 (the putative tobacco ortholog of
MKK4 and MKK5) transgene and mutations in each of MKK4
and MKK5 in regulating stomatal development downstream of
YDA (Wang et al., 2007).
We next expressed CA-MKK4 and CA-MKK5 beginning in
GMCs (FAMApro:CA-MKK4 and FAMApro:CA-MKK5); here, we
expected that activation of MKK4 or MKK5 in GMCs would, like
YDA activation, induce guard cell overproliferation and cluster-
ing. However, expression of neither transgene affected guard
cell development (Figures 3C and3F). For each construct, >75 T1
lines were screened and despite verification of transgene ex-
pression (as detected by yellow fluorescent protein [YFP] fluo-
rescence in GMCs and young guard cells; see Supplemental
Figure 1 online), only wild-type stomatal patterns were detected.
Figure 3. MKK4 and MKK5 Activity Influences Multiple Stages of Stomatal Development.
Activation of MKK4 and MKK5 signaling at specific stages of stomatal development using the SPCH, MUTE, and FAMA promoters revealed additional
inhibitory functions of MKK4 andMKK5 on stomatal development. Gray-shaded cells in the top panel show the expression pattern of each of the SPCH,
MUTE, and FAMA promoters, which are active in MMCs, meristemoids, and GMCs, respectively.
(A) to (F) Confocal images of propidium iodide–stained 10-DPG cotyledons. SPCHpro:CA-MKK4 (A) and SPCHpro:CA-MKK5 (D) blocked entry into the
stomatal lineage, and MUTEpro:CA-MKK4 (B) and MUTEpro:CA-MKK5 (E) result in cells arresting with meristemoid morphology. Expression of neither
FAMApro:CA-MKK4 (C) nor FAMApro:CA-MKK5 (F) affected stomatal development. Bars = 50 mm.
MAPK Regulation of Stomatal Cell Fate 5 of 12

Design and Construction of a CA-MAPKK Panel
Although we have shown that YDA can both inhibit and promote
specific transitions during stomatal development, MKK4 and
MKK5 appear to be downstream kinases only during the first two
(inhibitory) stages. These results raise two important questions.
First, do additional MAPKKs function downstream of YDA to
regulate early stages of stomatal development? Second, if other
MAPKKs are involved in stomatal development, which ones
function downstream of YDA in GMCs?
To answer these questions, we expanded our MAPKK test
panel to include MAPKKs whose broad expression patterns
included developing leaf tissues, that had previously been dem-
onstrated to mediate abiotic stress responses, and for whom
cognate downstream MAPKs had been described. Database
queries of publicly available microarray and MPSS data sets
indicated that each of MKK1, MKK2, MKK4, MKK5, MKK7, and
MKK9 are expressed in leaf tissue (see Supplemental Table
1 online) and have been implicated in stress responses. Inter-
estingly, these MAPKKs are all capable of phosphorylating
MPK3 and/or MPK6 (Colcombet and Hirt, 2008; Popescu et al.,
2009). As an additional test of the specificity of this system, we
also included MKK6, which has not been reported to be capable
of phosphorylating MPK3 or MPK6 in vivo. MKK6 allowed us to
determine whether expressing any CA-MAPKK would create
abnormal stomatal phenotypes. As with the other kinases in this
panel, MKK6 is broadly expressed in leaf tissue (see Supple-
mental Table 1 online).
Additional MAPKKs Can Inhibit Stomatal Development at
Multiple Stages
To examine the extent of MAPKK regulation over stomatal
development, each member of our CA-MAPKK panel was ex-
pressed beginning in MMCs and meristemoids using the SPCH
and MUTE promoters, respectively. Initiating MKK7 and MKK9
overactivity in MMCs prevents guard cell formation as reflected
by an epidermis consisting only of epidermal pavement cells
(similar toMKK4andMKK5; Figures 4Jand4M).ExpressionofCA-
MKK6 in MMCs had no effect on guard cell development (Figure
4G). Thus, the relationship betweenMAPK signaling and guard cell
development does not appear to be due to nonspecific MAPK
signaling defects within the stomatal lineage but, interestingly,
correlates with the ability to phosphorylate MPK3 and MPK6.
Due to their relationships with stress signaling involving MPK3
and MPK6, we speculated that MKK1 or MKK2 activity in MMCs
would also prevent stomatal development. However, we found
that plants expressing either MKK1 or MKK2 with the SPCH
promoter retained normal guard cell patterning in each of >50 T1
lines scored for each construct (Figures 4A and 4D). Transgene
expression was verified by the appearance of YFP fluorescence
in MMCs (see Supplemental Figure 1 online).
When expressed beginning in meristemoids, CA-MKK7 and
CA-MKK9, like CA-MKK4 and CA-MKK5, cause the plant to
create an epidermis consisting of epidermal pavement cells and
clusters of small cells that appear morphologically similar to
meristemoids (Figures 4K and 4N). However, expression of
MUTEpro:CA-MKK1 or MUTEpro:CA-MKK2, as confirmed by
the presence of YFP fluorescence (see Supplemental Figure
1 online), had no effect on guard cell development (Figures 4B
and 4E). Similarly, MUTEpro:MKK6 expression did not impair
stomatal patterning (Figure 4H). Thus, we identified two addi-
tional MAPKKs (MKK7 and MKK9) that are capable of inhibiting
the first two stages of stomatal development.
MKK7 and MKK9 Positively Influence the GMC to Guard
Cell Transition
We then turned to the outstanding question of which MAPKK(s)
could act downstream of YDA in promoting the differentiation of
guard cells. As seen in earlier stages, MKK1, MKK2, or MKK6
activity again appears to have no function in guard cell develop-
ment. The stomatal pattern in transgenic plants expressing
FAMApro:CA-MKK1, FAMApro:CA-MKK2, or FAMApro:CA-MKK6
appeared wild-type (Figures 4C, 4F, and 4I). In all cases, ex-
pression was confirmed by observation of YFP fluorescence in
GMCs and young guard cells (see Supplemental Figure 1 online).
By contrast, expression of FAMApro:CA-MKK7 or FAMApro:
CA-MKK9 resulted in a phenotype resembling themost severe of
the FAMApro:CA-YDA plants: gross overproduction of stomata
and the formation of guard cell clusters that protrude from the
epidermis of the leaves (Figures 4L and 4O). Thus, it appears that
like YDA, MKK7 and MKK9 can influence development at the
GMC to guard cell stages and, moreover, their activity promotes
rather than inhibits guard cell proliferation at the terminal stages
of stomatal development. Transgene expression was verified by
fluorescence from the CA-MKK7-YFP construct within GMCs
and young guard cells (see Supplemental Figure 1 online).
Guard Cell Tumor Formation Caused by Activation of MKK9
Results in SPCH Transcription
The guard cell overproliferation phenotype observed upon acti-
vation of YDA, MKK7, or MKK9 in GMCs is consistent with a
positive role of the YDA MAPK module in regulating terminal
guard cell development. Because misexpression of MUTE can
result in stomatal overproliferation independently of SPCH ac-
tivity (essentially bypassing this first step in the pathway;
MacAlister et al., 2007; Pillitteri et al., 2007), we tested whether
the guard cells that comprise the MKK9-induced stomatal clus-
ters develop by following the normal stomatal development
pathway. We assayed this by examining if the developing guard
cells displayed SPCH expression using a SPCH transcriptional
reporter (SPCHpro:nGFP; MacAlister et al., 2007). Because of the
similarity in the phenotypes produced by YDA, MKK7, and
MKK9, we followed only the effects of CA-MKK9. FAMApro:CA-
MKK9 plants displayed SPCHpro:nGFP reporter activity in the
clusters of cells that would become guard cells (Figure 5A). This
suggests that the guard cell overproliferation phenotype involves
resetting of the program back to the beginning of the normal
stomatal development pathway.
MAPKs Acting in the Pathway Downstream of MAPKKs
MPK3 and MPK6 function downstream of MKK4 and MKK5 in
broadly regulating stomatal development (Wang et al., 2007). As
6 of 12 The Plant Cell
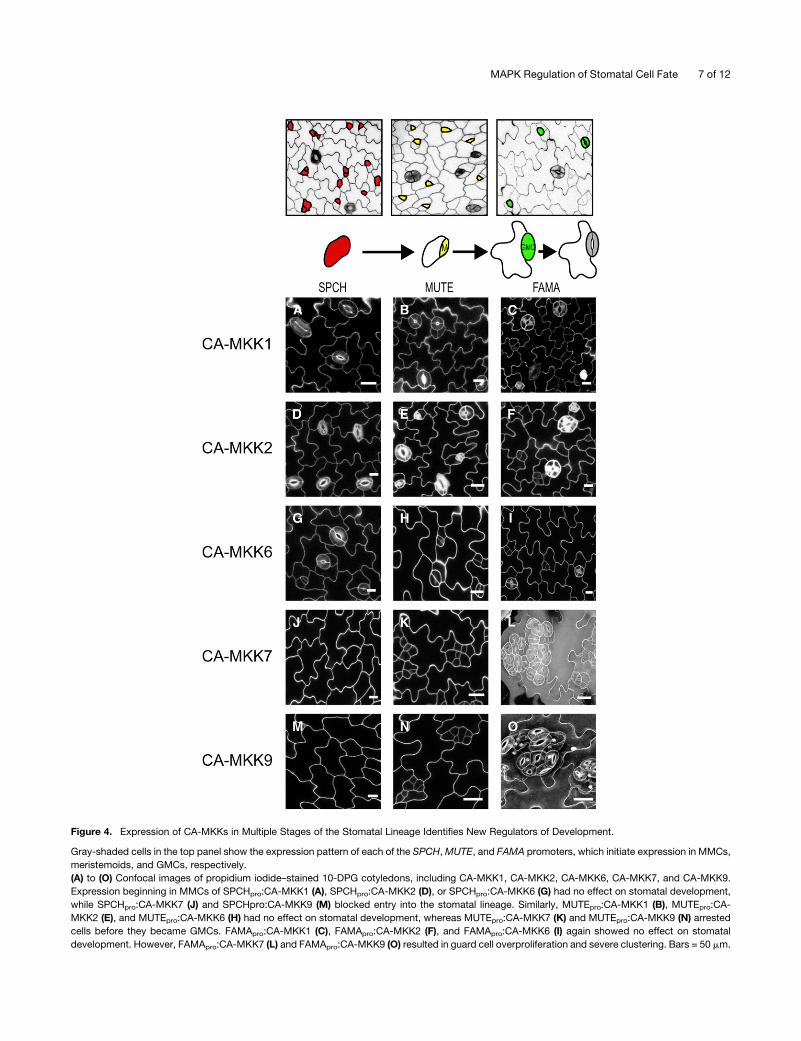
Figure 4. Expression of CA-MKKs in Multiple Stages of the Stomatal Lineage Identifies New Regulators of Development.
Gray-shaded cells in the top panel show the expression pattern of each of the SPCH,MUTE, and FAMA promoters, which initiate expression in MMCs,
meristemoids, and GMCs, respectively.
(A) to (O) Confocal images of propidium iodide–stained 10-DPG cotyledons, including CA-MKK1, CA-MKK2, CA-MKK6, CA-MKK7, and CA-MKK9.
Expression beginning in MMCs of SPCHpro:CA-MKK1 (A), SPCHpro:CA-MKK2 (D), or SPCHpro:CA-MKK6 (G) had no effect on stomatal development,
while SPCHpro:CA-MKK7 (J) and SPCHpro:CA-MKK9 (M) blocked entry into the stomatal lineage. Similarly, MUTEpro:CA-MKK1 (B), MUTEpro:CA-
MKK2 (E), and MUTEpro:CA-MKK6 (H) had no effect on stomatal development, whereas MUTEpro:CA-MKK7 (K) and MUTEpro:CA-MKK9 (N) arrested
cells before they became GMCs. FAMApro:CA-MKK1 (C), FAMApro:CA-MKK2 (F), and FAMApro:CA-MKK6 (I) again showed no effect on stomatal
development. However, FAMApro:CA-MKK7 (L) and FAMApro:CA-MKK9 (O) resulted in guard cell overproliferation and severe clustering. Bars = 50 mm.
MAPK Regulation of Stomatal Cell Fate 7 of 12

with MKK4 and MKK5, MKK7 and MKK9 each phosphorylate
MPK3 andMPK6 and are thus capable of signaling through these
kinases to regulate entry and progression through the stomatal
lineage. However, no positive roles for MPK3 or MPK6 in regu-
lating stomatal development have been reported. To determine
whether MPK3 and MPK6 function downstream of the YDA
module in promoting terminal guard cell development, we
expressed CA-MKK9 under the control of the FAMA promoter
in previously established mpk3 (SALK_100651) and mpk6
(SALK_074003) T-DNA insertion lines. Since MKK7 and MKK9
both phosphorylate MPK3 and MPK6 and are very similar in
amino acid sequence (79.5% identity; 88.8% similarity), we
chose to further examine the effects of only MKK9 activity in
GMCs. There were no differences in the ability of the CA-MKK9
transgene to promote guard cell overproliferation inmpk3 (38/40)
ormpk6 (38/38) null lines relative to wild-type Col (32/32). This is
consistent with MPK3 and MPK6 functioning redundantly in the
regulation of guard cell development but also with MKK9 signal-
ing through different or additional MAPKs in GMCs.
To establish whether additional MAPKs were involved in
stomatal development downstream of MKK9 in GMCs, we
assayed the stomatal clustering phenotype induced by MKK9
activity in GMCs in plant lines carrying T-DNA insertions within
coding regions of 14/20 of the Arabidopsis MAPK genes (Table
1). Guard cell clustering was not blocked in any of these trans-
genic lines, which may be due in part to functional redundancy
among downstreamMAPKs. Because these results do not allow
us to distinguish between MPK3/MPK6 functioning redundantly
downstream of MKK9 in GMCs and/or additional MAPKs func-
tioning downstream of MKK9, it remains unclear which MAPKs
function downstream of MKK9 during the guard mother cell to
guard cell transition and subsequent guard cell differentiation.
Targets of MAPKs
SPCH regulates the first cell state transition in the stomatal
lineage and was shown to be a target of MPK3 and MPK6
(Lampard et al., 2008). In these assays, however, neither FAMA
nor MUTEwas found to be an in vitro substrate of either MPK3 or
MPK6 (Lampard et al., 2008). Given the novel role of MAPK
signaling in regulating the GMC to guard cell transition, we
directed our experiments toward identifying targets of the MAPK
module during this last stage of stomatal development.
FLP and MYB88 are related MYB transcription factors, func-
tion at approximately the same stage as FAMA, and are both in
vitro substrates of MPK3 andMPK6 (Feilner et al., 2005; data not
shown). flp myb88 double mutants display defective terminal
guard cell differentiation, and they form caterpillar-like stomatal
lineage structures, often with a single stoma at one end of the
chain of cells (Lai et al., 2005). The FAMApro:CA-MKK9 construct
was introduced into flp myb88 double mutant plants (Figures 5C
and 5D). While guard cell formation was blocked in these plants,
consistent with the pattern observed in flp myb88 plants, there
were differences in the caterpillar structures generated by
Figure 5. Analysis of Stomatal Development Regulated by MKK9 Ac-
tivity in GMCs.
(A) Confocal images of propidium iodide–stained 5-DPG cotyledons.
Immature cells that will eventually become mature guard cell clusters
express SPCHpro:nGFP.
(B) to (D) Confocal images of propidium iodide–stained 10-DPG coty-
ledons.
(B) flp myb88 double mutants display caterpillar-like structures that tend
to contain parallel division planes and a single stomate at one end.
(C) and (D) flp myb88 FAMApro:CA-MKK9 plants produce caterpillar-like
structures that both divide in multiple orientations (C) and are found in
clusters (D).
Bars = 50 mm.Table 1. Single T-DNA Insertion Lines Tested for Their Ability to Block
the Phenotypic Consequences of FAMApro:CAMKK9 Expression
Gene T-DNA Insertion Line Accession Number
MPK1 SALK_63847C AT1G10210
MPK1 SALK_122198C AT1G10210
MPK2 SALK_047422C AT1G59580
MPK3 SALK_100651 AT3G45640
MPK6 SALK_074003 AT2G43790
MPK7 SALK_038863 AT2G18170
MPK8 SALK_219553C AT1G18150
MPK10 SALK_026099C AT3G59790
MPK11 SALK_049352C AT1G01560
MPK14 SALK_018940C AT4G36450
MPK15 SALK_046143C AT1G73670
MPK16 SALK_059737C AT5G19010
MPK18 SALK_069399C AT1G53510
MPK19 SALK_075213C AT3G14720
MPK20 SALK_090004 AT2G42880
The FAMApro:CA-MKK9 transgene was introduced into plants homozy-
gous for each T-DNA insertion listed to determine which, if any, potential
mpk-mutant lines blocked the ability of MKK9 activity to promote the
formation of guard cell clustering and overproliferation. Neither guard
cell overproliferation nor patterning defects was blocked in any line.
8 of 12 The Plant Cell

excess cell division in the epidermis of these plants. In flp myb88
doublemutants, these structures generally show parallel division
planes (Figure 5B), whereas in flp myb88 FAMApro:CA-MKK9
plants, the division planes within the structures were oriented in
numerousdirections (Figure 5C). Furthermore,multiple caterpillar-
like structures were clustered next to each other (Figures 5C and
5D), consistent with an additive effect of the FAMApro:CA-MKK9
transgene on the flp myb88 double mutant phenotype.
DISCUSSION
Using Cell Type–Specific Promoters to Activate MAPK
Signaling Networks
Large-scale genomic and proteomic studies have illustrated that
eukaryotic signaling pathways are complex and vastly intercon-
nected. Plant MAPK signaling networks are no exception. Each
of the MAPK components used in this study is multifunctional
and broadly expressed: YDA has functions in stomatal and
embryonic patterning (Bergmann et al., 2004; Lukowitz et al.,
2004; Wang et al., 2007), MKK1 and MKK2 are generally asso-
ciated with cold stress and pathogen responses (Teige et al.,
2004; Meszaros et al., 2006; Brader et al., 2007; Gao et al., 2008;
Qiu et al., 2008), andMKK4, MKK5, MKK7, andMKK9 have each
been reported to function in hormone, stress, and pathogen
signaling networks (Dai et al., 2006; Zhang et al., 2007; Colcombet
and Hirt, 2008; Xu et al., 2008; Yoo et al., 2008). In addition,
MPK3 and MPK6 are downstream targets of each of the afore-
mentioned MAPKKs and can be activated in response to a wide
range of stimuli (reviewed in Colcombet and Hirt, 2008). Because
of the lethal effects of broadly activating MAPKKs and/or the
multiple phenotypes associated with altered MAPK signaling in
multiple or ectopic cell types, it has been particularly difficult to
identify how these signaling components function in specific
physiological and/or developmental contexts.
Chemically inducible gene expression techniques that allow
quantitative induction of transgene expression have allowed
some circumnavigation around the lethal effects of widely acti-
vating MAPK networks (Wang et al., 2007). In fact, they have
been instrumental in providing details regarding MAPK signaling
and its role in stomatal development: A dexamethasone-inducible
MEK2 (the putative tobacco ortholog of MKK4 and MKK5)
construct was used to activate MPK3 and MPK6 signaling in
Arabidopsis, revealing inhibitory functions of MKK4/MKK5 and
MPK3/MPK6 in regulating stomatal development (Wang et al.,
2007). However, dexamethasone-induced activation of N. taba-
cum MEK2 arrested stomatal development by inhibiting entry
into the stomatal lineage, which prevented the identification of
the additional regulatory functions of MKK4 and MKK5 in regu-
lating stomatal development.
Here, we have shown that MAPK networks can be dissected
by modulating the expression of individual components in spe-
cific stomatal lineage cell types at both theMAPKKK andMAPKK
levels. For example, expression of a DN-YDA construct using the
SPCH promoter results in stomatal clustering and overprolifera-
tion but does not show the severe growth defects characteristic
of yda-null plants (Figure 2). Activation of MKK4 or MKK5 in
MMCs inhibited entry into the stomatal lineage (likely due to
SPCH phosphorylation by MPK3 and MPK6) without inducing
the rapid hypersensitive response–like cell death typically asso-
ciated with general overexpression of MKK4/5 activity (Jin et al.,
2003; Popescu et al., 2009). The revelation that discrete effects
of MAPKK activation can be effectively studied by inducing
activity in specific cell types provided the impetus to analyze the
roles of MKK4, MKK5, as well as other MKKs in regulating
stomatal development.
We have also used this system to investigate both functional
specificity among MAPK networks and the extent to which
MAPK signaling impacts stomatal development. Besides entry
into the stomatal lineage, MKK4, MKK5, MKK7, and MKK9 can
each negatively regulate the meristemoid-to-GMC transition.
However, only MKK7 and MKK9 activity beginning in GMCs is
capable of phenocopying the stomatal clustering and overprolif-
eration associated with YDA overactivity at the same stage. This
suggests that while each of the aforementioned MAPKKs can
function downstreamof YDA, signal integrity in specific cell types
may bemaintained via additional mechanisms, such as tethering
via scaffold proteins. Since activity of MKK1 or MKK2, which
each phosphorylate MPK3 and/or MPK6, does not influence
stomatal development, it is reasonable to conclude that in
addition to the specificity occurring at the MAPK substrate level
(Lampard et al., 2008), MAPK signaling specificity occurs at the
MAPKKK-MAPKK level. Therefore, this report also validates the
usefulness of perturbingMAPK signaling in specific stomatal cell
types to identify novel functions of MAPK signaling in vivo.
Negative Regulation of Stomatal Development by
MAPK Signaling
The effector(s) downstream of YDA in meristemoids remains
unknown. MUTE is not an in vitro substrate of MPK3 or MPK6,
suggesting it is not the direct target of the YDAmodule (Lampard
et al., 2008). Recently it was reported that MUTE is an in vitro
target of another stress and pathogenesis-associated MAPK,
MPK4, and that MPK4 is phosphorylated by MKK1 and MKK2
(Popescu et al., 2009). We demonstrated that expression of CA-
MKK1 or CA-MKK2 had no effect on the transition regulated by
MUTE. Therefore, either MPK4 is not a target of MKK1 or MKK2
in stomatal lineage cells or MUTE phosphorylation status does
not correlate with the arrested stomatal development phenotype
observed upon activation of YDA, MKK4, MKK5, MKK7, or
MKK9.
By revealing that additional stress-associated MAPKKs
(MKK7 and MKK9) can regulate stomatal development, we
have highlighted an important question: why is stomatal devel-
opment inhibited by stress-associated MAPKs? Several reports
indicate that the expression of genes linked to metabolism and
growth is downregulated upon exposure of plants to stress and
pathogens (reviewed in Baena-Gonzalez et al., 2007; Baena-
Gonzalez and Sheen, 2008), suggesting that plants divert
resources from these processes to better manage stress re-
sponses. It is plausible that activation of the MPK3/6 module by
environmental or pathogenic stimuli would be coupled with
arrested stomatal development. The rapid (and potentially re-
versible) block to development made possible by employing
MAPK Regulation of Stomatal Cell Fate 9 of 12

broadly expressed, stress-responsive kinases may, in fact, be
especially well suited to fine-tune the stomatal lineage. The
stomata on a mature leaf develop from many independent
precursor cells; it is estimated that two-thirds of the cells in the
Arabidopsis leaf epidermis are capable of producing stomata
(Geisler et al., 2000). Each of these cells has the potential to
transit through the precursor stages with independent timing.
Transient stresses can block development of subsets of the
lineage without compromising the ability of the leaf to ultimately
make stomata and be photosynthetically active, while chronic
stresses will eventually be able to affect the entire lineage.
A Novel, Positive Role for MAPK Signaling in Regulating
Stomatal Development
We unexpectedly observed that activation of YDA in GMCs
correlated with the appearance of clustered guard cells. Con-
sistent with this, we also observed that inhibition of YDA signaling
in GMCs corresponded with the appearance of cell patterns
similar to those seen in fama null plants and a general lack of
mature guard cells in the epidermis. Activation ofMKK7 orMKK9
but not MKK4 or MKK5 resulted in the formation of large clusters
of mature guard cells. Therefore, it appears that there is a branch
point in the YDA MAPK signaling module with MKK7 and MKK9
specifically functioning downstream of YDA in GMCs (Figure 6).
We attempted to assay loss of MKK7/9 function by engineering
changes in the MKK9 kinase domain equivalent to those shown
in the DN version of tobacco NQK1 (Soyano et al., 2003).
However, thesemodifications did not result in phenotypic effects
nor did expression of a synthetic microRNA construct dually
targeting MKK7 and MKK9 in GMCs. It remains to be seen
whether these results point to additional genetic redundancy in
the signaling module or reflect technical limitations in our ability
to eliminateMKK7 and/orMKK9 activity in stomatal lineage cells.
The specificity within the MAPK module whereby MKK7 and
MKK9 but not MKK4 or MKK5 activity promotes terminal differ-
entiation of guard cells may lie in the substrate specificity for
each of the MAPKKs, or it could be due to the presence of
additional specificity-determining factors, such as scaffold pro-
teins. In trying to determine if additional MAPKs function down-
stream ofMKK7 and/orMKK9 in GMCs, we assayed the ability of
MKK9 activity to promote guard cell clustering in single putative
loss of function mutants for 14 of the 20 MAPKs (Table 1). Guard
cell clustering was not blocked in any of these lines, suggesting
that either multiple MAPKs can function at least partially redun-
dantly in promoting guard cell development, or this process is
regulated by a yet to be identified MAPK. We note that this
developmental question may only be addressed by cell type–
specific manipulations because reduction of MPK3 and MPK6
expression throughout the plant (and therefore early in the
stomatal lineage where the MAPK signals are inhibitory) will
induce stomatal clusters (Wang et al., 2007).
An intriguing and unexpected result revealed by our studies is
the flip in behavior of the MAPK signaling system between the
negative regulation of the first stages and the positive regulation
of the final stage of stomatal development. This change can be
considered from several perspectives. At a physiological level,
the inverted behavior could be tied to a threshold effect. Upon
encountering a biotic or abiotic challenge, it may be favorable to
complete guard cell development once GMCs have formed
instead of just arresting their development. At the level of
signaling cascades, the ultimate development-promoting tar-
gets of the MAPKs in the early stages (e.g., SPCH; Lampard
et al., 2008) may be repressed by phosphorylation, whereas the
targets at later stages may be activated. Finally, in terms of cell
fate and commitment, it is possible that cells that have transi-
tioned through the early stages of stomatal development are now
differently competent to respond to MAPK signaling. This last
Figure 6. Model of MAPK Regulation of Stomatal Development.
The activation of four MKKs, MKK7 MKK9, MKK4, and MKK5, can
prevent entry into the stomatal lineage, likely by signaling through MPK3
and MPK6 to influence the phosphorylation status of SPCH. Each of the
aforementioned MAPKKs is also capable of blocking stomatal develop-
ment at the meristemoid-to-GMC transition. Activation of YDA, MKK7, or
MKK9 beginning in GMCs results in stomatal clustering and overprolif-
eration, highlighting a novel, positive function of MAPK signaling in
regulating terminal guard cell differentiation. Because each of these
MAPKKs mediates abiotic and/or biotic stress responses, there is a
potential for crosstalk between environmental and developmental cues.
The integration point could lie at the level of upstream activators,
downstream targets, or interaction partners. Alternatively, the MAPKKs
functioning at each step of stomatal development may be regulated
solely by developmental cues. While MPK3 and MPK6 are likely down-
stream MAPKs, this has only been experimentally verified for the entry
stage, and it is probable that additional MAPKK participate, especially at
the guard cell differentiation stage. The substrates of the MAPK modules
in meristemoids and GMCs remain to be identified.
10 of 12 The Plant Cell

hypothesis fits with the observation that the GMCs induced to
restart the pathway express SPCH but do not then immediately
arrest and will instead continue developing into guard cells. In
addition, a diminished or differential ability to respond to normal
developmental cues could explain why these reset cells form
clusters and do not adhere to the one-cell-spacing rule of normal
guard cell development.
We have described a novel in vivo assay to address specificity
issues inherent in many signaling networks. This has resulted in
the identification of new roles for specific MAPK members at
multiple time points throughout plant epidermal (stomatal) de-
velopment. This system is readily adaptable to study additional
signaling networks in Arabidopsis and complements the pro-
teome scale in vitrowork on kinases and substrates (for example,
Popescu et al., 2009). Ultimately, we will benefit from developing
other tools to activate and inactivate genes during stomatal
development, possibly by incorporating chemically inducible
gene expression systems and by a thorough characterization of
how stomatal development is affected following acute and
chronic stresses.
METHODS
Plant Materials
Col-0 was used as the wild type, and all transgenic plants were created
using this background unless otherwise noted. Plant lines containing
T-DNA inserts in coding regions of genes encodingMAPKs are described
in Table 1. Additional lines and alleles used in this work are as follows:
yda-Y295 (Bergmann et al., 2004) and flp myb88 (Lai et al., 2005).
Construction of CA-MKKs and Promoter-Specific
Expression Constructs
cDNA sequences corresponding to MKK2, MKK5, MKK7, and MKK9 in
pCR8 (Lee et al., 2008) were used as starting templates. Stop codons for
each gene were removed by amplifying the coding sequences using
Accuprime Pfx (Invitrogen) and the oligonucleotides listed in Supplemen-
tal Table 2 online. Following amplification, each amplicon was cloned into
pENTR-D-TOPO (Invitrogen). Constitutively active variants of each
MAPKK were constructed by conversion of the S/TXXXXXS/T MAPKK
activation motif to E/DXXXXXE/D using the QuikChange II XL site-
directed mutagenesis kit (Stratagene) according to the manufacturer’s
protocols using oligonucleotides listed in Supplemental Table 2 online
and as described (Popescu et al., 2009). cDNA sequences encoding each
of CA-MKK1, CA-MKK4, andCA-MKK6were containedwithin pCR8, and
the stop codons from each sequence were removed as described.
For expression with the SPCH, MUTE, and FAMA promoters, we
ligated the previously described 2.5-, 1.5-, and 2.5-kb sequences into the
NotI site within the pENTR-D-TOPOplasmid immediately upstream of the
cDNA sequences (Ohashi-Ito and Bergmann, 2006; MacAlister et al.,
2007). Each promoter:CA-MKK construct was then recombined into
pHGY (Kubo et al., 2005) usingGateway LR recombinase II (Invitrogen) for
transformation and subsequent expression in transgenic Arabidopsis
thaliana plants. The same procedure was performed to create promoter-
specific expression constructs of CA-YDA and DN-YDA, each of which
initially was contained within pENTR-D-TOPO. The CA-YDA sequence
contains a deletion of amino acids 185 to 322 and is a cDNA version of the
previously described DNB89 CA-YDA construct (Lukowitz et al., 2004),
and the DN-YDA variant contains a K429R substitution to eliminate the
catalytic site of the kinase.
Microscopy
Confocal images were collected using a Leica SP5 confocal microscope
with excitation/emission spectra of 514/520 to 540 for YFP and 565/580
to 610 for propidium iodide counterstaining. Images were processed in
ImageJ (NIH).
Accession Numbers
Accession numbers for YODAandMPKs andMKKs used in this study can
be found in Table 1 and Supplemental Table 1 online.
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Figure 1. Verification of Expression of CA-MKKs in
Multiple Stages of the Stomatal Lineage.
Supplemental Table 1. Gene Expression Data for MAPKKs Used in
This Study.
Supplemental Table 2. Oligonucleotides Used in This Study.
ACKNOWLEDGMENTS
We thank David Ehrhardt (Carnegie Department of Plant Biology) for the
use of the confocal microscopes, Kyoko Ohashi-Ito (University of Tokyo,
Japan) for building the CA- and DN-YDA constructs, and Marcus
Samuel (University of Calgary, Canada) for building the CA-MKK1,
CA-MKK4, and CA-MKK6 constructs. We also thank the current and
past members of the Bergmann lab for their comments on and insights
into this study. This work was supported by DOE-FG02-06ER15810.
Received July 14, 2009; revised September 9, 2009; accepted October
15, 2009; published November 6, 2009.
REFERENCES
Baena-Gonzalez, E., Rolland, F., Thevelein, J.M., and Sheen, J.
(2007). A central integrator of transcription networks in plant stress
and energy signalling. Nature 448: 938–942.
Baena-Gonzalez, E., and Sheen, J. (2008). Convergent energy and
stress signaling. Trends Plant Sci. 13: 474–482.
Bergmann, D.C., Lukowitz, W., and Somerville, C.R. (2004). Stomatal
development and pattern controlled by a MAPKK kinase. Science
304: 1494–1497.
Bergmann, D.C., and Sack, F.D. (2007). Stomatal development. Annu.
Rev. Plant Biol. 58: 163–181.
Blair, S.S. (2003). Genetic mosaic techniques for studying Drosophila
development. Development 130: 5065–5072.
Brader, G., Djamei, A., Teige, M., Palva, E.T., and Hirt, H. (2007). The
MAP kinase kinase MKK2 affects disease resistance in Arabidopsis.
Mol. Plant Microbe Interact. 20: 589–596.
Casson, S., and Gray, J. (2008). Influence of environmental factors on
stomatal development. New Phytol. 178: 9–23.
Casson, S.A., Franklin, K.A., Gray, J.E., Grierson, C.S., Whitelam,
G.C., and Hetherington, A.M. (2009). Phytochrome B and PIF4
regulate stomatal development in response to light quantity. Curr.
Biol. 19: 229–234.
Chen, R.E., and Thorner, J. (2007). Function and regulation in MAPK
signaling pathways: Lessons learned from the yeast Saccharomyces
cerevisiae. Biochim. Biophys. Acta 1773: 1311–1340.
MAPK Regulation of Stomatal Cell Fate 11 of 12

Colcombet, J., and Hirt, H. (2008). Arabidopsis MAPKs: A complex
signalling network involved in multiple biological processes. Biochem.
J. 413: 217–226.
Coupe, S.A., Palmer, B.G., Lake, J.A., Overy, S.A., Oxborough, K.,
Woodward, F.I., Gray, J.E., and Quick, W.P. (2006). Systemic
signalling of environmental cues in Arabidopsis leaves. J. Exp. Bot.
57: 329–341.
Dai, Y., Wang, H., Li, B., Huang, J., Liu, X., Zhou, Y., Mou, Z., and Li,
J. (2006). Increased expression of MAP KINASE KINASE7 causes
deficiency in polar auxin transport and leads to plant architectural
abnormality in Arabidopsis. Plant Cell 18: 308–320.
Feilner, T., et al. (2005). High throughput identification of potential
Arabidopsis mitogen-activated protein kinases substrates. Mol. Cell.
Proteomics 4: 1558–1568.
Gao, M., Liu, J., Bi, D., Zhang, Z., Cheng, F., Chen, S., and Zhang, Y.
(2008). MEKK1, MKK1/MKK2 and MPK4 function together in a mito-
gen-activated protein kinase cascade to regulate innate immunity in
plants. Cell Res. 18: 1190–1198.
Geisler, M., Nadeau, J., and Sack, F.D. (2000). Oriented asymmetric
divisions that generate the stomatal spacing pattern in arabidopsis are
disrupted by the too many mouths mutation. Plant Cell 12: 2075–2086.
Hamel, L.P., et al. (2006). Ancient signals: Comparative genomics of
plant MAPK and MAPKK gene families. Trends Plant Sci. 11: 192–198.
Hara, K., Kajita, R., Torii, K.U., Bergmann, D.C., and Kakimoto, T.
(2007). The secretory peptide gene EPF1 enforces the stomatal one-
cell-spacing rule. Genes Dev. 21: 1720–1725.
Ichimura, K., Shinozaki, K., Tena, G., Sheen, J., Henry, Y., Champion,
A., Kreis, M., Zhang, S., and Hirt, H. (2002). Mitogen-activated protein
kinase cascades in plants: a new nomenclature. Trends Plant Sci. 7:
301–308.
Jin, H., Liu, Y., Yang, K.Y., Kim, C.Y., Baker, B., and Zhang, S. (2003).
Function of a mitogen-activated protein kinase pathway in N gene-
mediated resistance in tobacco. Plant J. 33: 719–731.
Kanaoka, M.M., Pillitteri, L.J., Fujii, H., Yoshida, Y., Bogenschutz,
N.L., Takabayashi, J., Zhu, J.K., and Torii, K.U. (2008). SCREAM/ICE1
and SCREAM2 specify three cell-state transitional steps leading to
Arabidopsis stomatal differentiation. Plant Cell 20: 1775–1785.
Kubo, M., Udagawa, M., Nishikubo, N., Horiguchi, G., Yamaguchi,
M., Ito, J., Mimura, T., Fukuda, H., and Demura, T. (2005). Tran-
scription switches for protoxylem and metaxylem vessel formation.
Genes Dev. 19: 1855–1860.
Kultz, D. (2005). Molecular and evolutionary basis of the cellular stress
response. Annu. Rev. Physiol. 67: 225–257.
Lai, L.B., Nadeau, J.A., Lucas, J., Lee, E.K., Nakagawa, T., Zhao, L.,
Geisler, M., and Sack, F.D. (2005). The Arabidopsis R2R3 MYB
proteins FOUR LIPS and MYB88 restrict divisions late in the stomatal
cell lineage. Plant Cell 17: 2754–2767.
Lampard, G.R., MacAlister, C.A., and Bergmann, D.C. (2008). Arabi-
dopsis stomatal initiation Is controlled by MAPK-mediated regulation
of the bHLH SPEECHLESS. Science 322: 1113–1116.
Lee, J.S., Huh, K.W., Bhargava, A., and Ellis, B.E. (2008). Compre-
hensive analysis of protein-protein interactions between Arabidopsis
MAPKs and MAPK kinases helps define potential MAPK signaling
modules. Plant Signal. Behav. 3: 1037–1041.
Liu, Y., Jin, H., Yang, K.Y., Kim, C.Y., Baker, B., and Zhang, S. (2003).
Interaction between two mitogen-activated protein kinases during
tobacco defense signaling. Plant J. 34: 149–160.
Lukowitz, W., Roeder, A., Parmenter, D., and Somerville, C. (2004). A
MAPKK kinase gene regulates extra-embryonic cell fate in Arabidop-
sis. Cell 116: 109–119.
MacAlister, C.A., Ohashi-Ito, K., and Bergmann, D.C. (2007). Tran-
scription factor control of asymmetric cell divisions that establish the
stomatal lineage. Nature 445: 537–540.
Meszaros, T., Helfer, A., Hatzimasoura, E., Magyar, Z., Serazetdinova,
L., Rios, G., Bardoczy, V., Teige, M., Koncz, C., Peck, S., and
Bogre, L. (2006). The Arabidopsis MAP kinase kinase MKK1 partic-
ipates in defence responses to the bacterial elicitor flagellin. Plant J.
48: 485–498.
Neill, S., Barros, R., Bright, J., Desikan, R., Hancock, J., Harrison, J.,
Morris, P., Ribeiro, D., and Wilson, I. (2008). Nitric oxide, stomatal
closure, and abiotic stress. J. Exp. Bot. 59: 165–176.
Ohashi-Ito, K., and Bergmann, D.C. (2006). Arabidopsis FAMA con-
trols the final proliferation/differentiation switch during stomatal de-
velopment. Plant Cell 18: 2493–2505.
Pillitteri, L.J., Sloan, D.B., Bogenschutz, N.L., and Torii, K.U. (2007).
Termination of asymmetric cell division and differentiation of stomata.
Nature 445: 501–505.
Popescu, S.C., Popescu, G.V., Bachan, S., Zhang, Z., Gerstein, M.,
Snyder, M., and Dinesh-Kumar, S.P. (2009). MAPK target networks
in Arabidopsis thaliana revealed using functional protein microarrays.
Genes Dev. 23: 80–92.
Qiu, J.L., Zhou, L., Yun, B.W., Nielsen, H.B., Fiil, B.K., Petersen, K.,
Mackinlay, J., Loake, G.J., Mundy, J., and Morris, P.C. (2008).
Arabidopsis mitogen-activated protein kinase kinases MKK1 and
MKK2 have overlapping functions in defense signaling mediated by
MEKK1, MPK4, and MKS1. Plant Physiol. 148: 212–222.
Schmid, M., Davison, T.S., Henz, S.R., Pape, U.J., Demar, M.,
Vingron, M., Scholkopf, B., Weigel, D., and Lohmann, J.U.
(2005). A gene expression map of Arabidopsis thaliana development.
Nat. Genet. 37: 501–506.
Schmidt, E.E. (2007). The origins of polypeptide domains. Bioessays
29: 262–270.
Soyano, T., Nishihama, R., Morikiyo, K., Ishikawa, M., and Machida,
Y. (2003). NQK1/NtMEK1 is a MAPKK that acts in the NPK1 MAPKKK-
mediated MAPK cascade and is required for plant cytokinesis. Genes
Dev. 17: 1055–1067.
Teige, M., Scheikl, E., Eulgem, T., Doczi, R., Ichimura, K., Shinozaki,
K., Dangl, J.L., and Hirt, H. (2004). The MKK2 pathway mediates cold
and salt stress signaling in Arabidopsis. Mol. Cell 15: 141–152.
Wang, H., Ngwenyama, N., Liu, Y., Walker, J.C., and Zhang, S.
(2007). Stomatal development and patterning are regulated by envi-
ronmentally responsive mitogen-activated protein kinases in Arabi-
dopsis. Plant Cell 19: 63–73.
Widmann, C., Gibson, S., Jarpe, M.B., and Johnson, G.L. (1999).
Mitogen-activated protein kinase: Conservation of a three-kinase
module from yeast to human. Physiol. Rev. 79: 143–180.
Xu, J., Li, Y., Wang, Y., Liu, H., Lei, L., Yang, H., Liu, G., and Ren, D.
(2008). Activation of MAPK kinase 9 induces ethylene and camalexin
biosynthesis and enhances sensitivity to salt stress in Arabidopsis.
J. Biol. Chem. 283: 26996–27006.
Xu, T., and Rubin, G.M. (1993). Analysis of genetic mosaics in
developing and adult Drosophila tissues. Development 117: 1223–
1237.
Yoo, S.D., Cho, Y.H., Tena, G., Xiong, Y., and Sheen, J. (2008). Dual
control of nuclear EIN3 by bifurcate MAPK cascades in C2H4 signal-
ling. Nature 451: 789–795.
Zhang, X., Dai, Y., Xiong, Y., DeFraia, C., Li, J., Dong, X., and Mou, Z.
(2007). Overexpression of Arabidopsis MAP kinase kinase 7 leads to
activation of plant basal and systemic acquired resistance. Plant J.
52: 1066–1079.
12 of 12 The Plant Cell


















