
Newt Squamous Carcinoma Proves Phylogenetic Conservation ... · Newt Squamous Carcinoma Proves...
9
[CANCER RESEARCH 52, 4858-4865, September 15, 1992] Newt Squamous Carcinoma Proves Phylogenetic Conservation of Tumors as Caricatures of Tissue Renewall Nikolaos P. Zilakos, Panagiotis A. Tsonis, Katia Del Rio-Tsonis, and Ralph E. Parchment2 Division of Pharmacology and Toxicology, Hippie Cancer Research Center, 4100 South Kettering Blvd., Dayton, OH 45439-2092 [N. P. Z., R. E. P.], and Laboratory of Molecular Biology, Department of Biology, University of Dayton, Dayton, OH 45469-2320 [P. A. T., K. D. R-TJ ABSTRACT Although regeneration-competent newts like Notophthalmus virìde- scens have been reported to be resistant to carcinogenesis, we have been able to induce transplantable epidermal squamous cell carcinomas with 10-20% incidence by implanting 20-methylcholanthrene s.c. into the scapular region, a tissue that cannot regenerate. As soon as 1 week after exposure to this carcinogen, malignant cells were present in the treated skin, and after 4 weeks, macroscopic tumors of infiltrating squamous carcinoma cells positive for Type IV collagenase and/or ras"" p21 had dissolved areas of the epidermal basement membrane and colonized the dermis. Analysis of Ki-67 expression revealed that these tumors grow via a high growth fraction rather than a short cell cycle time. Morpho logical and immunohistochemical analyses showed that these tumors caricature the biology of the renewing epidermis: the presence of basal- like cells; differentiating cells; apoptotic cells; and keratinized horn pearls with an exaggerated or overabundant stem cell compartment as compared to the differentiated cell compartment. Immunochemical analyses indicated that the squamous carcinomas arose from the epi dermis rather than the mucous glands. Thus, the principle that malig nant tumors caricature the process of tissue renewal originally estab lished in rodent tumors appears to be valid down the phylogenetic tree at least to regeneration-competent amphibia. Such a broad conservation indicates that the caricature principle also holds in human tumors. INTRODUCTION Carcinogenesis was originally thought to transform differen tiated cells into immature, embryonic ones, whose rapid prolif eration formed the tumor ( 1). The conversion of a cell of mature phenotype to embryonic phenotype was termed dedifferentia- tion (2). This model did explain the relative lack of differenti ated tissue in the tumor. However, recent data from rodent models indicate that a stem cell model should replace the de- differentiation model (1). Neoplasms occur only in cell popu lations with proliferative ability ("renewable" tissues); postmi- totic, terminally differentiated cell populations do not become malignant. If any differentiated characteristics are evident in a tumor, they will always resemble those of the tumor's tissue of origin, since this is the basis of pathological diagnosis. Thus, the target cell in carcinogenesis is a cell capable of tissue re newal and determined for a particular differentiation, and the only cell that has these characteristics is the stem cell (3). This stem cell model was confirmed by Pierce et al. (l, 4), who showed that differentiated cells in a tumor are postmitotic even though they are produced by malignant stem cells; areas con taining only differentiated tumor cells did not transplant the tumor, whereas areas containing undifferentiated cells did, and these tumors included differentiated cells. Received 12/5/91; accepted 7/7/92. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accord ance with 18 U.S.C. Section 1734 solely to indicate this fact. 1 Supported in part by Grant CA51325 from the National Cancer Institute; by benefactors of the Hippie Cancer Research Center such as the Dayton Exchange Club, the Phi Beta Psi Sorority, the Moraine Lyceum Society, and Kettering Fair mont High School; and by the Research Institute of the University of Dayton. The Developmental Studies Hybridoma Bank is maintained by a contract from the NICHO (NO1-HD-2-3144). 2 To whom requests for reprints should be addressed, at Mod- l,Rm2023,ORR, CDER, FDA, 8301 Muirkirk Road, Laurel, MD 20708. Carcinogenesis (i.e., the malignant transformation of normal stem cells or, in the case of clonal malignancy, perhaps a single stem cell) in renewing tissues such as epithelia results in an exaggeration or "caricature" of tissue renewal (5). It is a cari cature because of the exaggerated production of stem and pro genitor cells in relationship to differentiated cells, and the im mature cells accumulate, even under conditions when normal stem cells would be quiescent. Malignant stem cells respond abnormally to steady-state mechanism(s) in their tissues of or igin, and thus tissue renewal is no longer balanced to cell loss. However, other regulatory mechanisms continue to operate. For example, both physiological cell death by apoptosis (6) and terminal differentiation occur in malignant epidermal tissue, as evidenced by the presence of apoptotic cells and keratinized horn pearls. In addition, some tissue organization is also main tained after malignant transformation, as evidenced by the com- partmentalization of proliferating and terminally differentiated cells. The stem cell basis for malignancy (i.e., the "caricature" model) is an important principle of tumor biology with ramifi cations for developing curative, nontoxic cancer therapy in hu mans (5), but only if this principle is conserved phylogeneti- cally. To challenge the phylogenetic conservation of this principle, we have chosen a primitive amphibian, the newt Notophthalmus viridescens, as a model. The results show that malignant tumors in N. viridescens obey the caricature principle just like rodent tumors. MATERIALS AND METHODS Animals. Adult newts (amphibia Urodela), N. viridescens, of both sexes were used in this study under protocols approved by the Institu tional Animal Care and Use Committee at the Hippie Cancer Research Center. The animals were purchased from Charles D. Sullivan Com pany, Inc. (Nashville, TN) and were maintained in plastic aquaria at a room temperature of 22-25°C. The animals were fed live Newberry shrimp or frozen beef liver given every other day. After each feeding the water of the aquarium was changed. Administration of 20-Methylcholanthrene. In preparation for sur gery, the newts were anesthetized with MS222 (Tricaine methane- sulfonate). Then a 10-15-Mg crystal of 20-MCA3 was implanted under neath the epidermis just above the scapula. All recipients of the carcinogen were examined every week for the appearance of tumor. When tumors reached approximately 3-5 mm in diameter, the animals were sacrificed and the tumors were excised. Tumor Transplantation. Animals with tumors of 3-5 mm diameter were anesthetized by immersion in a 1 mg/ml solution of MS222 for 5 min. The surgical area was irrigated with a gentamicin-penicillin-strep- tomycin-fungizone solution, and then the tumor was resected with a scalpel, placed in a sterile Petri dish containing McCoy's 5A medium (Gibco), and minced with scalpels into the appropriately sized pieces (usually 1-2-mm cubes) for transplantation. Tumor was transferred with forceps into a second newt after antibiotic irrigation as described above. After recovering from anesthesia, the tumor recipients were placed into a 10-liter tank with water to a 1-inch depth. This water was changed every hour for 6 h, and the newts were not fed during these 6 h to control infection rates. 3 The abbreviation used is: 20-MCA, 20-methylcholanthrene. 4858 on June 6, 2020. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Transcript of Newt Squamous Carcinoma Proves Phylogenetic Conservation ... · Newt Squamous Carcinoma Proves...

[CANCER RESEARCH 52, 4858-4865, September 15, 1992]
Newt Squamous Carcinoma Proves Phylogenetic Conservation of Tumors asCaricatures of Tissue Renewall
Nikolaos P. Zilakos, Panagiotis A. Tsonis, Katia Del Rio-Tsonis, and Ralph E. Parchment2
Division of Pharmacology and Toxicology, Hippie Cancer Research Center, 4100 South Kettering Blvd., Dayton, OH 45439-2092 [N. P. Z., R. E. P.], andLaboratory of Molecular Biology, Department of Biology, University of Dayton, Dayton, OH 45469-2320 [P. A. T., K. D. R-TJ
ABSTRACT
Although regeneration-competent newts like Notophthalmus virìde-scens have been reported to be resistant to carcinogenesis, we have beenable to induce transplantable epidermal squamous cell carcinomas with10-20% incidence by implanting 20-methylcholanthrene s.c. into thescapular region, a tissue that cannot regenerate. As soon as 1 week afterexposure to this carcinogen, malignant cells were present in the treatedskin, and after 4 weeks, macroscopic tumors of infiltrating squamouscarcinoma cells positive for Type IV collagenase and/or ras"" p21 had
dissolved areas of the epidermal basement membrane and colonized thedermis. Analysis of Ki-67 expression revealed that these tumors growvia a high growth fraction rather than a short cell cycle time. Morphological and immunohistochemical analyses showed that these tumorscaricature the biology of the renewing epidermis: the presence of basal-like cells; differentiating cells; apoptotic cells; and keratinized hornpearls with an exaggerated or overabundant stem cell compartment ascompared to the differentiated cell compartment. Immunochemicalanalyses indicated that the squamous carcinomas arose from the epidermis rather than the mucous glands. Thus, the principle that malignant tumors caricature the process of tissue renewal originally established in rodent tumors appears to be valid down the phylogenetic treeat least to regeneration-competent amphibia. Such a broad conservationindicates that the caricature principle also holds in human tumors.
INTRODUCTION
Carcinogenesis was originally thought to transform differentiated cells into immature, embryonic ones, whose rapid proliferation formed the tumor ( 1). The conversion of a cell of maturephenotype to embryonic phenotype was termed dedifferentia-tion (2). This model did explain the relative lack of differentiated tissue in the tumor. However, recent data from rodentmodels indicate that a stem cell model should replace the de-differentiation model (1). Neoplasms occur only in cell populations with proliferative ability ("renewable" tissues); postmi-
totic, terminally differentiated cell populations do not becomemalignant. If any differentiated characteristics are evident in atumor, they will always resemble those of the tumor's tissue of
origin, since this is the basis of pathological diagnosis. Thus,the target cell in carcinogenesis is a cell capable of tissue renewal and determined for a particular differentiation, and theonly cell that has these characteristics is the stem cell (3). Thisstem cell model was confirmed by Pierce et al. (l, 4), whoshowed that differentiated cells in a tumor are postmitotic eventhough they are produced by malignant stem cells; areas containing only differentiated tumor cells did not transplant thetumor, whereas areas containing undifferentiated cells did, andthese tumors included differentiated cells.
Received 12/5/91; accepted 7/7/92.The costs of publication of this article were defrayed in part by the payment of
page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1Supported in part by Grant CA51325 from the National Cancer Institute; bybenefactors of the Hippie Cancer Research Center such as the Dayton ExchangeClub, the Phi Beta Psi Sorority, the Moraine Lyceum Society, and Kettering Fairmont High School; and by the Research Institute of the University of Dayton. TheDevelopmental Studies Hybridoma Bank is maintained by a contract from theNICHO (NO1-HD-2-3144).
2 To whom requests for reprints should be addressed, at Mod- l,Rm2023,ORR,CDER, FDA, 8301 Muirkirk Road, Laurel, MD 20708.
Carcinogenesis (i.e., the malignant transformation of normalstem cells or, in the case of clonal malignancy, perhaps a singlestem cell) in renewing tissues such as epithelia results in anexaggeration or "caricature" of tissue renewal (5). It is a cari
cature because of the exaggerated production of stem and progenitor cells in relationship to differentiated cells, and the immature cells accumulate, even under conditions when normalstem cells would be quiescent. Malignant stem cells respondabnormally to steady-state mechanism(s) in their tissues of origin, and thus tissue renewal is no longer balanced to cell loss.However, other regulatory mechanisms continue to operate.For example, both physiological cell death by apoptosis (6) andterminal differentiation occur in malignant epidermal tissue, asevidenced by the presence of apoptotic cells and keratinizedhorn pearls. In addition, some tissue organization is also maintained after malignant transformation, as evidenced by the com-partmentalization of proliferating and terminally differentiatedcells.
The stem cell basis for malignancy (i.e., the "caricature"
model) is an important principle of tumor biology with ramifications for developing curative, nontoxic cancer therapy in humans (5), but only if this principle is conserved phylogeneti-cally. To challenge the phylogenetic conservation of thisprinciple, we have chosen a primitive amphibian, the newtNotophthalmus viridescens, as a model. The results show thatmalignant tumors in N. viridescens obey the caricature principlejust like rodent tumors.
MATERIALS AND METHODS
Animals. Adult newts (amphibia Urodela), N. viridescens, of bothsexes were used in this study under protocols approved by the Institutional Animal Care and Use Committee at the Hippie Cancer ResearchCenter. The animals were purchased from Charles D. Sullivan Company, Inc. (Nashville, TN) and were maintained in plastic aquaria at aroom temperature of 22-25°C. The animals were fed live Newberry
shrimp or frozen beef liver given every other day. After each feeding thewater of the aquarium was changed.
Administration of 20-Methylcholanthrene. In preparation for surgery, the newts were anesthetized with MS222 (Tricaine methane-sulfonate). Then a 10-15-Mg crystal of 20-MCA3 was implanted under
neath the epidermis just above the scapula. All recipients of thecarcinogen were examined every week for the appearance of tumor.When tumors reached approximately 3-5 mm in diameter, the animalswere sacrificed and the tumors were excised.
Tumor Transplantation. Animals with tumors of 3-5 mm diameterwere anesthetized by immersion in a 1 mg/ml solution of MS222 for 5min. The surgical area was irrigated with a gentamicin-penicillin-strep-tomycin-fungizone solution, and then the tumor was resected with ascalpel, placed in a sterile Petri dish containing McCoy's 5A medium
(Gibco), and minced with scalpels into the appropriately sized pieces(usually 1-2-mm cubes) for transplantation. Tumor was transferredwith forceps into a second newt after antibiotic irrigation as describedabove. After recovering from anesthesia, the tumor recipients wereplaced into a 10-liter tank with water to a 1-inch depth. This water waschanged every hour for 6 h, and the newts were not fed during these 6h to control infection rates.
3 The abbreviation used is: 20-MCA, 20-methylcholanthrene.
4858
on June 6, 2020. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

NEWT SQUAMOUS CARCINOMA CARICATURES TISSUE RENEWAL
Histology. MCA-affected tissues were fixed in Bouin's solution or
with 10% neutral buffered formalin and then embedded in paraffin.Sections of 5-7 /im were cut using a rotary microtome. Paraffin sectionswere stained with hematoxylin-eosin or a modified trichrome Mallory-
azan method (7) that accentuates the basement membrane below theepidermis of the skin.
Immunohistochemistry. In preparation for indirect immunofluores-cent studies, the tissue was embedded in optimal cutting temperature(OCT) medium, and frozen sections were cut at 6-10 Mmin a cryostatat -18°C. Primary antibodies were applied at 37°Cfor 60 min. The
slides were washed 3x10 min in phosphate-buffered saline and thenreacted with fluorescein isothiocyanate-conjugated goat anti-rabbit oranti-mouse antibody. The primary antibodies used were (a) Ki-67(AMAC, Inc., Westbrook, Maine), a mouse IgG, monoclonal antibodythat reacts with a nuclear antigen expressed in the nucleus of cyclingcells predominantly during the S phase of the cell cycle (8); (b) anti-typeI keratin polyclonal rabbit anti-serum that reacts with a peptide sequence from the carboxyl terminus of the Xenopus K81A type I keratin(no. 1920; a gift from Dr. T. Sergant and Dr. I. David); (e) WE3(purchased from the Developmental Studies Hybridoma Bank, deposited by R. A. Tassava), a mouse IgG monoclonal antibody that rea >with secretory epithelial cells in mucous glands of the skin (9); (d)anti-ras" monoclonal antibody produced against amino acids 96-118of the v-ras" sequence that reacts with both c-ras" and v-ras11proteins
(Quality Biotech, Trenton, NJ). The results of the negative controlsconsisting of secondary antibody alone, which uniformly gave no im-munostaining, are not shown.
In addition, S-^m sections from formalin-fixed, paraffin-embeddedspecimens were incubated overnight at 4°Cwith 10 Mg/mlmurine IgG2a
monoclonal antibody against Type IV collagenase (Zymed Labs, Inc.)in 1% normal goat serum in phosphate-buffered saline, washed threetimes in phosphate-buffered saline, incubated for 60 min at room temperature with goat anti-mouse IgG secondary antibody labeled withhorseradish peroxidase (Vector Labs), washed again three times inbuffer, and then developed with the HistoBlack detection system(Vector Labs) and counterstained lightly with 0.1% Metanil yellow.
RESULTS
Similar to other vertebrates, the normal skin in the scapularregion of N. viridescens is composed of an epidermal layer ofkeratinocytes approximately 4-6 cell layers thick, separatedfrom the underlying dermis by a basement membrane (Fig. I, Aand li). The epidermis contains a superficial layer called thecorneum striatum, which contains the keratinized remains ofterminally differentiated keratinocytes. Basal cells and kerati-nocyte stem cells are located at the bottom of the epidermis,near the basement membrane, and cells move through the spinal layer into the corneum striatum as they differentiate. Mi-totic figures were very infrequent in the epidermis. Fig. IBshows a higher magnification of normal skin stained with Mal-lory-azan to accentuate the collagen fibrils and basement mem
brane. Note that the cells with the largest nuclearcytoplasmicratio are localized near the basement membrane. Also note thepresence of apoptotic cells (small arrow) in the intermediatelayers as well as in the corneum striatum. Thus the generalfeatures of epidermal cell biology in rodents, such as basallysituated stem cells, apoptotic bodies, and production of corni-fied, terminally differentiated cells, are all present in newt epidermis.
Melanocytes with easily visible melanotic granules were concentrated at the basement membrane, but, in contrast to mammals, they are found exclusively on the dermal side (Fig. 1, Aand B, large arrows). The dermis is composed of a thin layer ofsparse collagen fibrils containing infrequent leukocytes and mucous glands lined with secretory epithelium. As the dermis approached the subcutaneous muscle layer, the collagen fibrils
become denser. Antibody against Type I keratins did not stainany of the cell types in the dermis or underlying tissues (datanot shown).
The limbs of newts show strong regenerative capability, andprevious work has shown that chemical carcinogens or ionizingradiation do not produce tumors in regeneration-competenttissue, even though they produce stable mutations in the genome (10-13). Spontaneous tumors do not occur in these regions either (13). To bypass this obstacle to tumor formation,we implanted 20-methylcholanthrene in the subcutaneous tissue in the scapular region. This area shows no capacity forregeneration (14), so we reasoned that tumor formation wouldbe maximized there.
Fig. 1C shows a section through scapular tissues after 2weeks of 20-MCA exposure. The epidermis appeared normal,with retention of the corneum striatum, a middle spinal layer,and basal layer. However, the dermis was now almost completely devoid of mucous glands, and the deeper layers had lostthe dense collagen matrix and musculature. Inflammatory cellswere common throughout the deep tissue sites and the dermis.Fig. 1C shows one suspicious area (*), where basal cells and
dermal melanocytes appeared to be in abnormally close contact.Fig. \D shows a higher magnification of this area. There werea few cells in the dermis that are morphologically identical tobasal keratinocytes. These keratinocyte nests in the dermis werefound as early as 1 week after 20-MCA implantation.
To determine if these early appearing keratinocyte nests inthe dermis were composed of malignant cells, we determinedwhether transplanted skin exposed to 20-MCA for only 1 weekwould form tumors in the flank of untreated animals. Three of10 of the transplants (30%) formed squamous carcinomas aftertransplantation (Table 1). In contrast, untreated scapular regions did not form tumors after transplantation (Table 1).These data suggested that the keratinocyte-like cells appearingin the dermis after 1 week of exposure to 20-MCA were likelythe first manifestation of malignant transformation.
Three weeks after implanting 20-MCA, small nodules of 1-2mm diameter were observed under the epidermis in the scapularregions of some of the treated animals. By the end of 4 weeks,macroscopic tumors of 3-5 mm were visible in the scapularregion (Fig. 2, A and B). The tumors appeared s.c. and causedsome mild focal ulcérationconsistent with deep invasion. Nosuperficial tumors or papillomas were observed. After a total of8 weeks, no additional tumors were observed in animals thatwere negative for tumor at the end of 4 weeks, and no animalswere found to have multiple tumor nodules. Autopsy of tumor-bearing animals revealed no obvious métastasesto liver, lungs,or pleural cavity. Tumor incidence scored after 4 weeks in foursequential experiments was 18%, 10%, 18%, and 10%, respectively (Table 2). The animals with tumor exhibited no decreasein activity or appetite.
These tumors were excised, and portions were examined his-tologically (Fig. 2, C and D). Examination revealed tumorscontaining tightly associated, atypical/irregular epithelial cellswith pleiomorphic nuclear and cytoplasmic shape and activecytoplasm, which had penetrated the basement membrane andproliferated downward into the dermis (Fig. 4A). Horn pearlformation by spindle cells was particularly noticeable in centralized areas of tumor nests (Fig. 2, C and D). Apoptotic cellswere found throughout the tumor (Fig. 1C). Surrounding thekeratinized pearls were large cells with nuclei, showing pleiomorphic size, shape, and chromatin condensation, that resembled cells in the overlying epidermis, although the nuclei of thetumor cells contained less heterochromatin, and nucleoli were
4859
on June 6, 2020. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
NEWT SQUAMOUS CARCINOMA CARICATURES TISSUE RENEWAL
B
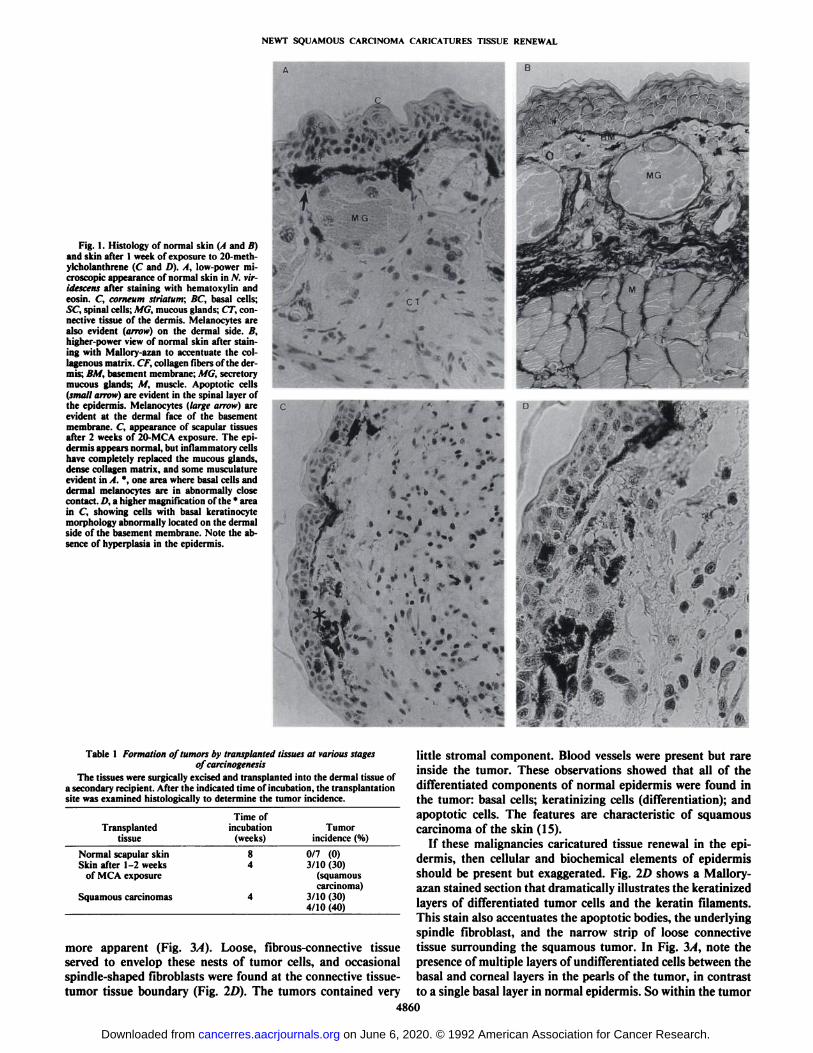
Fig. 1. Histology of normal skin (I and //)and skin after I week of exposure to 20-meth-ylcholanthrene (C and D). A, low-power microscopic appearance of normal skin in N. vir-idescens after staining with hematoxylin andeosin. C, corneum strlatum; BC, basal cells;SC, spinal cells; MG, mucous glands; CT, connective tissue of the dermis. Melanocytes arealso evident (arrow) on the dermal side. B,higher-power view of normal skin after staining with Mallory-azan to accentuate the col-lagenous matrix. CF, collagen fibers of the dermis; BM, basement membrane; MG, secretorymucous glands; M, muscle. Apoptotic cells(small arrow) arc evident in the spinal layer ofthe epidermis. Melanocytes (large arrow) areevident at the dermal face of the basementmembrane. C, appearance of scapular tissuesafter 2 weeks of 20-MCA exposure. The epidermis appears normal, but inflammatory cellshave completely replaced the mucous glands,dense collagen matrix, and some musculatureevident in A. *, one area where basal cells and
dermal melanocytes are in abnormally closecontact. /-'. a higher magnification of the * areain C, showing cells with basal keratinocytemorphology abnormally located on the dermalside of the basement membrane. Note the absence of hyperplasia in the epidermis.
t / >"fs**"!:
•¿� f •¿�* *">/•*•.w* •¿�
Table 1 Formation of tumors by transplanted tissues at various stagesof carcinogenesis
The tissues were surgically excised and transplanted into the dermal tissue ofa secondary recipient. After the indicated time of incubation, the transplantationsite was examined histologically to determine the tumor incidence.
TransplantedtissueNormal
scapular skinSkin after 1-2 weeks
of MCAexposureSquamous
carcinomasTime
ofincubation
(weeks)g
44Tumor
incidence(%)0/7
(0)3/10 (30)
(squamouscarcinoma)
3/10 (30)4/10 (40)
more apparent (Fig. 3/4). Loose, fibrous-connective tissueserved to envelop these nests of tumor cells, and occasionalspindle-shaped fibroblasts were found at the connective tissue-tumor tissue boundary (Fig. 2D). The tumors contained very
little stromal component. Blood vessels were present but rareinside the tumor. These observations showed that all of thedifferentiated components of normal epidermis were found inthe tumor: basal cells; keratinizing cells (differentiation); andapoptotic cells. The features are characteristic of squamouscarcinoma of the skin (15).
If these malignancies caricatured tissue renewal in the epidermis, then cellular and biochemical elements of epidermisshould be present but exaggerated. Fig. ID shows a Mallory-azan stained section that dramatically illustrates the keratinizedlayers of differentiated tumor cells and the keratin filaments.This stain also accentuates the apoptotic bodies, the underlyingspindle fibroblast, and the narrow strip of loose connectivetissue surrounding the squamous tumor. In Fig. 3/4, note thepresence of multiple layers of undifferentiated cells between thebasal and corneal layers in the pearls of the tumor, in contrastto a single basal layer in normal epidermis. So within the tumor
4860
on June 6, 2020. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

NEWT SQUAMOUS CARCINOMA CARICATURES TISSUE RENEWAL
Fig. 2. Squamous carcinomas in the scapular regions of N. viridescens 4 weeks after implantation of 20-methylcholanthrene crystalsinto the underlying dermis. No superficial tumors or papillomas were observed. I. dorsalmacroscopic view of a tumor; B, lateral macroscopic view of a tumor showing ulcérationatthe tumor's surface; C and D, histological sec
tions of squamous carcinomas stained withhematoxylin and eosin or Mallory-azan. Alltumors exhibit identical histological characteristics which identify them as squamous carcinomas (IS): tightly associated, atypical/irregular epithelial cells with pleiomorphicnuclear and cytoplasmic shape and active cytoplasm, producing keratinized horn pearls(HP), which are evidence of terminal differentiation. Melanocytes (arrow) often accompanythe infiltrating tumor. Apoptotic cells AC, arefound throughout the tumor. The tumors contain very little stromal component, and bloodvessels are rare. Note the exaggeration of normal tissue renewal by the tumor, which indicates the caricature principle.
.
Table 2 W-Methylcholanîhrenecarcinogenesis in the scapular regionof Nolophlhalmus viridescens
"Location" refers to the anatomical site where the majority of tumor residedand not to the site of origin of the tumor. 20-MCA crystals (10-15 Mg)wereimplanted into the scapular region of newts (both males and females), and tumorincidence was scored after 4 weeks. No additional tumors appeared after 4 weeks.
Animals No. ofExperiment treated tumors ("<>> Histology Location
1234303040685(18)3(10)7(18)7(10)SquamouscarcinomaSquamouscarcinomaSquamouscarcinomaSquamous
carcinomaSubcutaneousSubcutaneousSubcutaneousSubcutaneous
the number of undifferentiated cells and differentiated cells wasunbalanced. In normal skin, there was usually one basal cell percorneal cell, but in the tumor usually there were six undifferentiated cells per keratinizing cell. This overproduction of immature (stem?) cells is a common feature of tumors whichcaricature the process of tissue renewal in their tissues of origin.
However, evidence of differentiation was retained, such as cellsmorphologically similar to the normal cells in the corneumstriatum layer of normal newt skin.
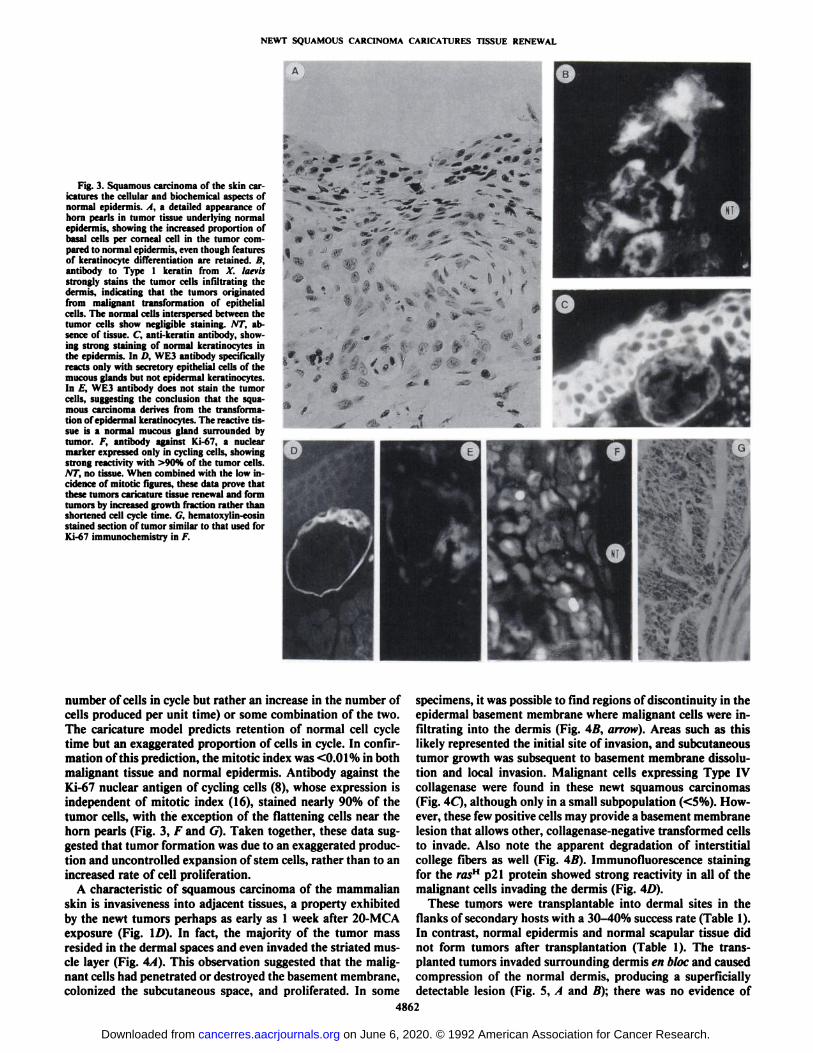
Biochemical characteristics of the tumor reflected its caricature of epidermal renewal. The malignant cells, but not thesurrounding normal dermal components, stained strongly positive with antibody against Xenopus Type I keratin (Fig. 3Ä).This same keratin antibody strongly stained epidermal kerati-
nocytes of the normal skin and the secretory epithelium of themucous glands (Fig. 3C). Furthermore, WE3 antibody specificfor secretory epithelia of the mucous glands (Fig. 3D) did notstain the squamous carcinomas (Fig. 3£).This result suggestedthe conclusion that the squamous carcinomas arose from theepidermis rather than the mucous glands.
Accumulation of malignant tissue could be due to either increased growth fraction (an increase in the number of cells inthe cell cycle) or decreased cell cycle time (no change in the
4861
on June 6, 2020. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
NEWT SQUAMOUS CARCINOMA CARICATURES TISSUE RENEWAL
Fig. 3. Squamous carcinoma of the skin caricatures the cellular and biochemical aspects ofnormal epidermis. . I. a detailed appearance ofhorn pearls in tumor tissue underlying normalepidermis, showing the increased proportion ofbasal cells per corneal cell in the tumor compared to normal epidermis, even though featuresof keratinocyte differentiation are retained. B,antibody to Type 1 keratin from X. laevisstrongly stains the tumor cells infiltrating thedermis, indicating that the tumors originatedfrom malignant transformation of epithelialcells. The normal cells interspersed between thetumor cells show negligible staining. AT, absence of tissue. C, anti-keratin antibody, showing strong staining of normal keratinocytes inthe epidermis. In /'. WE3 antibody specificallyreacts only with secretory epithelial cells of themucous glands but not epidermal keratinocytes.In /:'. WE3 antibody does not stain the tumor
cells, suggesting the conclusion that the squa-mous carcinoma derives from the transformation of epidermal keratinocytes. The reactive tissue is a normal mucous gland surrounded bytumor. /. antibody against Ki-67, a nuclearmarker expressed only in cycling cells, showingstrong reactivity with >90% of the tumor cells.NT, no tissue. When combined with the low incidence of mitotic figures, these data prove thatthese tumors caricature tissue renewal and formtumors by increased growth fraction rather thanshortened cell cycle time. G, hematoxylin-eosinstained section of tumor similar to that used forKi-67 immunochemistry in F.
number of cells in cycle but rather an increase in the number of specimens, it was possible to find regions of discontinuity in thecells produced per unit time) or some combination of the two.The caricature model predicts retention of normal cell cycletime but an exaggerated proportion of cells in cycle. In confirmation of this prediction, the mitotic index was <0.01% in bothmalignant tissue and normal epidermis. Antibody against theKi-67 nuclear antigen of cycling cells (8), whose expression isindependent of mitotic index (16), stained nearly 90% of thetumor cells, with the exception of the flattening cells near thehorn pearls (Fig. 3, F and G). Taken together, these data suggested that tumor formation was due to an exaggerated production and uncontrolled expansion of stem cells, rather than to anincreased rate of cell proliferation.
A characteristic of squamous carcinoma of the mammalianskin is invasiveness into adjacent tissues, a property exhibitedby the newt tumors perhaps as early as 1 week after 20-MCAexposure (Fig. ID). In fact, the majority of the tumor massresided in the dermal spaces and even invaded the striated muscle layer (Fig. 4/4). This observation suggested that the malignant cells had penetrated or destroyed the basement membrane,colonized the subcutaneous space, and proliferated. In some
epidermal basement membrane where malignant cells were infiltrating into the dermis (Fig. 4B, arrow). Areas such as thislikely represented the initial site of invasion, and subcutaneoustumor growth was subsequent to basement membrane dissolution and local invasion. Malignant cells expressing Type IVcollagenase were found in these newt squamous carcinomas(Fig. 4C), although only in a small subpopulation (<5%). However, these few positive cells may provide a basement membranelesion that allows other, collagenase-negative transformed cellsto invade. Also note the apparent degradation of interstitialcollege fibers as well (Fig. 4B). Immunofluorescence stainingfor the rasH p21 protein showed strong reactivity in all of the
malignant cells invading the dermis (Fig. 4D).These tumors were transplantable into dermal sites in the
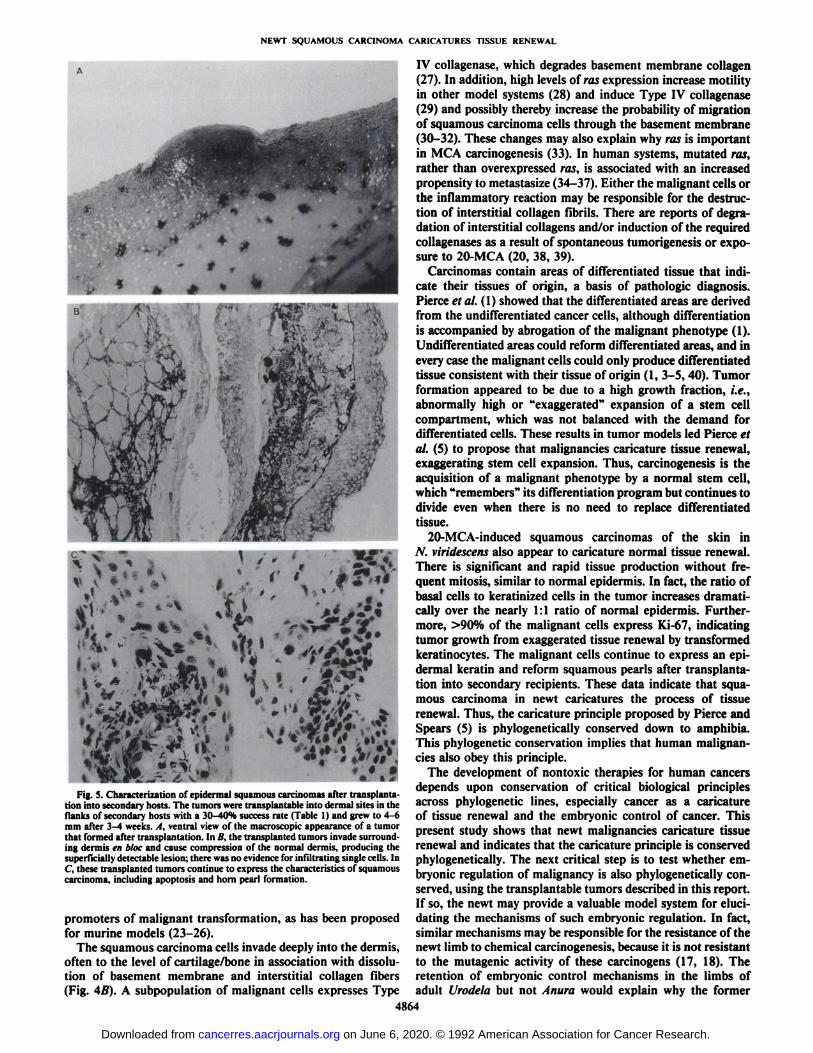
flanks of secondary hosts with a 30-40% success rate (Table 1).In contrast, normal epidermis and normal scapular tissue didnot form tumors after transplantation (Table 1). The transplanted tumors invaded surrounding dermis en bloc and causedcompression of the normal dermis, producing a superficiallydetectable lesion (Fig. 5, A and A); there was no evidence of
4862
on June 6, 2020. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

NEWT SQUAMOUS CARCINOMA CARICATURES TISSUE RENEWAL
Fig. 4. Mechanism of invasion of 20-meth-ylcholanthrene-induced squamous carcinomasinto the dermis and subdermal tissue. A, deeptissue invasion is a commonly occurring characteristic of squamous carcinoma (IS). In JV.viridescens, some tumors have invaded thedeep muscle layers under the dermis by 4-5weeks after administration of 20-MCA. B,Mallory-azan stained section showing a discontinuity in the basement membrane {arrow)and dissolution of interstitial collagen associated with tumor infiltration into the dermis. C,antibody to mammalian Type IV collagenasestains some of the tumor cells in the dermalspace. The very dark cells are melanocytes. D,tumor cells invading the dermis express highlevels of ras" p21 protein, which in mammalian tumors is associated with increased cellmotility and increased metastatic behavior(28-37).
infiltrating single cells. These transplanted tumors continued toexpress the characteristics of squamous carcinoma, includinghorn pearl formation (Fig. 5Q.
DISCUSSION
Many previous studies have shown that limbs from Urodelaamphibia including N. viridescens are resistant to chemical car-cinogenesis (10-13, 17, 18) and spontaneous tumorigenesis(13), but these studies were attempting to challenge the hypothesis that a regeneration-competent tissue is resistant to chemical carcinogenesis, not that chemical carcinogenesis does notoccur in newts. Because we previously identified a newt with aspontaneous melanoma in the scapular region (19), an areawithout regenerative capability, we suspected that the key toexperimentally producing cancers in this species might be toimplant carcinogens in regeneration-incompetent tissue like thescapular region (14). Our results confirmed this idea. These
squamous malignancies are distinct from spontaneously regressing skin tumors (20, 21). These data focus the hypothesisof amphibian resistance to carcinogenesis upon mechanismsspecific to regeneration-competent tissues. This conclusion isconsistent with the higher incidence of tumors found in amphibia like Animi that undergo metamorphosis and loose theirregenerative ability (13, 14).
This is the first report of chemically induced, transplantablesquamous carcinoma of the skin in Urodela. These malignancies occur via rapid transformation of epidermal keratinocytes(and not secretory epithelial cells of the mucous glands), incontrast to the prolonged exposure time required to inducemelanomas with 20-MCA in Triturus cristatus (22). It is interesting that 100% of the induced tumors were squamous carcinoma, especially when a promoting agent, such as phorbol ester, was not coadministered. Perhaps oxygen free radicalsproduced by inflammatory cells at the site of carcinogenimplantation (Fig. 1C) were functioning as endogenous
4863
on June 6, 2020. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
NEWT SQUAMOUS CARCINOMA CARICATURES TISSUE RENEWAL
,m^ -i .w _ •¿�* '
Fig. S. Characterization of epidermal squamous carcinomas after transplantation into secondary hosts. The tumors were transplantable into dermal sites in theflanks of secondary hosts with a 30-40% success rate (Table 1) and grew to 4-6mm after 3-4 weeks. A, ventral view of the macroscopic appearance of a tumorthat formed after transplantation. In B, the transplanted tumors invade surrounding dermis en bloc and cause compression of the normal dermis, producing thesuperficially detectable lesion; there was no evidence for infiltrating single cells. InC, these transplanted tumors continue to express the characteristics of squamouscarcinoma, including apoptosis and horn pearl formation.
promoters of malignant transformation, as has been proposedfor murine models (23-26).
The squamous carcinoma cells invade deeply into the dermis,often to the level of cartilage/bone in association with dissolution of basement membrane and interstitial collagen fibers(Fig. 4B). A subpopulation of malignant cells expresses Type
IV collagenase, which degrades basement membrane collagen(27). In addition, high levels of ras expression increase motilityin other model systems (28) and induce Type IV collagenase(29) and possibly thereby increase the probability of migrationof squamous carcinoma cells through the basement membrane(30-32). These changes may also explain why ras is important
in MCA carcinogenesis (33). In human systems, mutated ras,rather than overexpressed ras, is associated with an increasedpropensity to metastasize (34-37). Either the malignant cells orthe inflammatory reaction may be responsible for the destruction of interstitial collagen fibrils. There are reports of degradation of interstitial collagens and/or induction of the requiredcollagenases as a result of spontaneous tumorigenesis or exposure to 20-MCA (20, 38, 39).
Carcinomas contain areas of differentiated tissue that indicate their tissues of origin, a basis of pathologic diagnosis.Pierce et al.(l) showed that the differentiated areas are derivedfrom the undifferentiated cancer cells, although differentiationis accompanied by abrogation of the malignant phenotype (1).Undifferentiated areas could reform differentiated areas, and inevery case the malignant cells could only produce differentiatedtissue consistent with their tissue of origin (1, 3-5, 40). Tumorformation appeared to be due to a high growth fraction, i.e.,abnormally high or "exaggerated" expansion of a stem cell
compartment, which was not balanced with the demand fordifferentiated cells. These results in tumor models led Pierce etal. (S) to propose that malignancies caricature tissue renewal,exaggerating stem cell expansion. Thus, carcinogenesis is theacquisition of a malignant phenotype by a normal stem cell,which "remembers'" its differentiation program but continues to
divide even when there is no need to replace differentiatedtissue.
20-MCA-induced squamous carcinomas of the skin inN. viridescens also appear to caricature normal tissue renewal.There is significant and rapid tissue production without frequent mitosis, similar to normal epidermis. In fact, the ratio ofbasal cells to keratinized cells in the tumor increases dramatically over the nearly 1:1 ratio of normal epidermis. Furthermore, >90% of the malignant cells express Ki-67, indicatingtumor growth from exaggerated tissue renewal by transformedkeratinocytes. The malignant cells continue to express an epidermal keratin and reform squamous pearls after transplantation into secondary recipients. These data indicate that squamous carcinoma in newt caricatures the process of tissuerenewal. Thus, the caricature principle proposed by Pierce andSpears (5) is phylogenetically conserved down to amphibia.This phylogenetic conservation implies that human malignancies also obey this principle.
The development of nontoxic therapies for human cancersdepends upon conservation of critical biological principlesacross phylogenetic lines, especially cancer as a caricatureof tissue renewal and the embryonic control of cancer. Thispresent study shows that newt malignancies caricature tissuerenewal and indicates that the caricature principle is conservedphylogenetically. The next critical step is to test whether embryonic regulation of malignancy is also phylogenetically conserved, using the transplantable tumors described in this report.If so, the newt may provide a valuable model system for elucidating the mechanisms of such embryonic regulation. In fact,similar mechanisms may be responsible for the resistance of thenewt limb to chemical carcinogenesis, because it is not resistantto the mutagenic activity of these carcinogens (17, 18). Theretention of embryonic control mechanisms in the limbs ofadult Urodela but not Anura would explain why the former
4864
on June 6, 2020. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

NEWT SQUAMOUS CARCINOMA CARICATURES TISSUE RENEWAL
show increased resistance to chemical carcinogenesis and spontaneous tumorigenesis (10, 13).
ACKNOWLEDGMENTS
The authors wish to acknowledge the expert assistance in manuscriptpreparation by Ann Barhorst and technical assistance for histology byStephanie Redwine.
REFERENCES1. Pierce, G. B., Shikes, R., and Fink, L. M. Cancer: A Problem of Develop
mental Biology, pp. 68-87. Englewood Cliffs, NJ: Prentice-Hall, 1978.2. Hay, E. D., and riseli man. D. A. Origin of the blastema in regenerating limbs
of the newt Trilurus viridescens, Dev. Biol., 3: 26-59, 1961.3. Pierce, G. B., and Johnson, L. D. Differentiation and cancer. In Vitro (Rock-
ville), 7: 140-145, 1971.4. Pierce, G. B., Dixon, F. J., and Verney, E. L. Teratocarcinogenic and tissue
forming potentials of the cell types comprising neoplastic embryoid bodies.Lab. Invest., 9: 583-602, 1960.
5. Pierce, G. B., and Speers, W. C. Tumors as caricatures of the process of tissuerenewal: prospects for therapy by directing differentiation. Cancer Res., 48:1996-2004, 1988.
6. Kerr, J. F. R., Wyllie, A. H., and Currie, A. R. Apoptosis: a basic biologicalphenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer,20:239-257, 1972.
7. Delides, G. S. Practical Histológica! Techniques. Athens, GA: University ofAthens Press, 1978.
8. Bruno, S., Crissman, H. A., Bauer, K. D., and Darzynkiewicz, Z. Changes incell nuclei during S phase: progressive chromatin condensation and alteredexpression of the proliferation-associated nuclear proteins ki 67, cyclin(PCNA), plOS, and p34. Exp. Cell Res., 196: 99-106, 1991.
9. Tassava, R. A., Johnson-Wint, B., and Gross, J. Regenerate epithelium andskin glands of the adult newt react to the same antibody. J. Exp. Zool., 239:229-240, 1986.
10. Tsonis, P. A. Effects of carcinogens on regenerating and non-regeneratinglimbs in amphibia. Anticancer Res., 3: 195-202, 1983.
11. Tsonis, P. A. Studies of the effects of cancer inducing and promoting chemicals on newt regenerating and non-regenerating limbs. Arch. Biol., 96: 331-336, 1985.
12. Zavanella, T., and Losa, M. Skin damage in adult amphibians after chronicexposure to ultraviolet radiation. Photochem. Photobiol., 34:487-492,1981.
13. Tsonis, P. A., and Del Rio-Tsonis, K. Spontaneous neoplasms in amphibia.Tumor Biol., 9: 221-224, 1988.
14. Wallace, H. Vertebrate limb regeneration. Chichester, England: John Wileyand Sons, 1981.
15. Kwan, T. H., and Mihm, M. C., Jr. The skin. In: S. L. Robbins and R. S.Cotran (eds.), Pathologic Basis of Disease, pp. 1426-1427. Philadelphia: W.B. Saunders Company, 1979.
16. Ostmeier, H., and Suter, L. The Ki-67 antigen in primary humanmelanomas—its relationship to mitotic rate and tumor thickness and itsstability. Arch. Dermatol. Res., 281: 173-177, 1989.
17. Tsonis, P. A., and Eguchi, G. Abnormal limb regeneration without tumorformation in adult newts directed by carcinogens 20-methylcholanthrene andbenzo(a)pyrene. Dev. Growth Differ., 24: 183-190, 1982.
18. Zilakos, N. P. The regeneration of Trituras limbs deformed by a carcinogen.In Vivo (Athens), 5: 381-384, 1991.
19. Zilakos, N. P., and Tsonis, P. A. A spontaneous melanoma-like tumor in theadult newt Triturus cristatus. Tumor Biol., 12: 120-124, 1991.
20. Wirl, G. Collagenolytic activity and cancerogenesis in the skin of the newtTriturus cristatus. Arch. Geschwulstforsch., 40: 111-115, 1972.
21. Pfeiffer, C. J., Nagai, T., Fujimura, M., and Tobe, T. Spontaneous regressiveepitheliomas in the Japanese newt, Cynops pyrrhogaster. Cancer Res., 39:1904-1910, 1979.
22. Leone, V. G., and Zavanella, T. Some morphological and biological characteristics of a tumor of the newt Triturus cristatus laur. In: M. Mizell (ed.),Biology of Amphibian Tumors, pp. 184-194. New York: Springer-Verlag,1969.
23. Bhisey, R. A., and Sirsat, S. M. Selective promoting activity of phorbolmyristate acetate in experimental skin carcinogenesis. Br. J. Cancer, 34:661-665, 1976.
24. Birnboim, H. C. Superoxide aniónmay trigger DNA strand breaks in humangranulocytes by acting at a membrane target. Ann. N.Y. Acad. Sci., 551:83-94, 1988.
25. Swauger, J. E., Dolan, P. M., and Kensler, T. W. Role of free radicals intumor promotion and progression by benzoyl peroxide. Prog. Clin. Biol.Res., 340D: 143-52, 1990.
26. Kensler, T. W., Egner, P. A., Taffe, B. G., and Trush, M. A. Role of freeradicals in tumor promotion and progression. Prog. Clin. Biol. Res., 298:233-48, 1989.
27. Thorgeirsson, U. P., Turpeenniemi-Hujanen, T., and Liotta L. A. Cancercells, components of basement membranes, and proteolytic enzymes. Int.Rev. Exp. Pathol., 27: 203-34, 1985.
28. Partin, A. W., Isaacs, J. T., Treiger, B., and Coffey, D. S. Early cell motilitychanges associated with an increase in metastatic ability in rat prostate cancercells transfected with the v-Harvey-ros oncogene. Cancer Res., 48: 6050-6053, 1988.
29. Garbisa, S., Negro, A., Kalebic, T., Pozzatti, R., Muschel. R., Saffiotti, U.,and Liotta, L. A. Type IV collagenolytic activity linkage with the metastaticphenotype induced by ras transfection. Adv. Exp. Med. Biol., 233: 179-186,1988.
30. Treiger, B., and Isaacs, J. T. Expression of a transfected v-Harvey-ros oncogene in a Dunning rat prostate adenocarcinoma and the development of highmetastatic ability. J Urol., 140: 1580-1586, 1988.
31. Schwarz, L. C., Gingras, M. C., Goldberg, G., Greenberg, A. H., and Wright,J. A. Loss of growth factor dependence and conversion of transforminggrowth factor >'!,inhibition to stimulation in metastatic H-ras-transformedmurine fibroblasts. Cancer Res., 48: 6999-7003, 1988.
32. Waghorne, C„Kerbel, R. S., and Breitman, M. L. Metastatic potential ofSP1 mouse mammary adenocarcinoma cells is differentially induced by activated and normal forms of c-H-ros. Oncogene, /: 149-155, 1987.
33. Brown, K., Buchmann, A., and Balmain, A. Carcinogen-induced mutations inthe mouse c-Ha-ros gene provide evidence of multiple pathways for tumorprogression. Proc. Nati. Acad. Sci. USA, 87: 538-542, 1990.
34. Yoshida, K., Hamatani, K., Koide, H., Abe, Y., Ikeda, H., Tsuchiyama, H.,Nakayama, E., and Shiku, H. Analysis of ras gene expression in stomachcancer by anti-ras p21 monoclonal antibodies. Cancer Detect. Prev., 12:369-376, 1988.
35. Sheng, Z. M., Barrois, M., Klijanienko, J., Micheau, C., Richard, J. M., andRiou, G. Analysis of the c-Ha-ros-l gene for deletion, mutation, amplificationand expression in lymph node métastasesof human head and neck carcinomas. Br. J. Cancer, 62: 398-404, 1990.
36. Deng, G. R., Liu, X. H., and Wang, J. R. Correlation of mutations ofoncogene r I lu KMat codon 12 with metastasis and survival of gastric cancerpatients. Oncogene Res., 6: 33-38, 1991.
37. Slebos, R. J. C., Kibbelaar, R. E., Dalesio, O., Kooistra, A., Stam, J., Meijer,C. J. L. M., Wagenaar, S. S., Vanderschueren, R. G. J. R. A., van Zandwijk,N., Mooi, W. J., Bos, J. L., and Rodenhuis, S. K-RAS oncogene activation asa prognostic marker in adenocarcinoma of the lung. N. Engl. J. Med., 323:561-565, 1990.
38. Seilern-Aspang, F., Mazzucco, K., and Christian, I. Der Einfluss von Meth-ylcholanthren auf Synthese, Vernetzung und Abbau des Kollagens derMausedermis und die mögliche Bedeutung dieses Abbaues fürmalignesWachstum. Z. Naturforsch., 24b: 894-901, 1969.
39. Rohrbach, R., Thomas, C., Rumpier, R., and Lau, M. Die Kollagenasenaktivitätund der Kollagengehalt der Haut wahrend der Carcinogenese. Verh.Dlsch. Ges. Pathol., 54: 442-50, 1970.
40. Pierce, G. B., and Wallace, C. Differentiation of malignant to benign cells.Cancer Res., 31: 127-134, 1971.
4865
on June 6, 2020. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1992;52:4858-4865. Cancer Res Nikolaos P. Zilakos, Panagiotis A. Tsonis, Katia Del Rio-Tsonis, et al. of Tumors as Caricatures of Tissue RenewalNewt Squamous Carcinoma Proves Phylogenetic Conservation
Updated version
http://cancerres.aacrjournals.org/content/52/18/4858
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/52/18/4858To request permission to re-use all or part of this article, use this link
on June 6, 2020. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from


















