
MutationsofanArylalkylamine-N-acetyltransferase, Bm-iAANAT ... ·...
9
Mutations of an Arylalkylamine- N -acetyltransferase, Bm-iAANAT , Are Responsible for Silkworm Melanism Mutant * □ S Received for publication, December 18, 2009, and in revised form, March 18, 2010 Published, JBC Papers in Press, March 23, 2010, DOI 10.1074/jbc.M109.096743 Fang-yin Dai ‡1 , Liang Qiao ‡1 , Xiao-ling Tong ‡§1 , Cun Cao ‡ , Peng Chen ‡ , Jun Chen ‡ , Cheng Lu ‡2 , and Zhong-huai Xiang ‡ From the ‡ College of Biotechnology, Institute of Sericulture and Systems Biology, Southwest University, Chongqing 400715, China and the § Institute of Agriculture and Life Science, Chongqing University, Chongqing 400030, China Coloration is one of the most variable characters in animals and provides rich material for studying the developmental genetic basis of pigment patterns. In the silkworm, more than 100 gene mutation systems are related to aberrant color patterns. The mel- anism (mln) is a rare body color mutant that exhibits an easily dis- tinguishable phenotype in both larval and adult silkworms. By positional cloning, we identified the candidate gene of the mln locus, Bm-iAANAT, whose homologous gene (Dat) converts dopa- mine into N-acetyldopamine, a precursor for N-acetyldopamine sclerotin in Drosophila. In the mln mutant, two types of abnormal Bm-iAANAT transcripts were identified, whose expression levels are markedly lower than the wild type (WT). Moreover, dopamine content was approximately twice as high in the sclerified tissues (head, thoracic legs, and anal plate) of the mutant as in WT, result- ing in phenotypic differences between the two. Quantitative reverse transcription PCR analyses showed that other genes involved in the melanin metabolism pathway were regulated by the aberrant Bm-iAANAT activity in mln mutant in different ways and degrees. We therefore propose that greater accumula- tion of dopamine results from the functional deficiency of Bm- iAANAT in the mutant, causing a darker pattern in the sclerified regions than in the WT. In summary, our results indicate that Bm-iAANAT is responsible for the color pattern of the silkworm mutant, mln. To our knowledge, this is the first report showing a role for arylalkylamine-N-acetyltransferases in color pattern mutation in Lepidoptera. Coloration is one of the most variable traits in insects, and pigmentation is known to play a role in mimicry, sexual selec- tion, thermoregulation, and other adaptive processes in many insect groups (1). More recently, there has been much interest among biologists in the molecular mechanisms underlying the great diversity of insect colors and color patterns. Melanin, the dark pigment found in melanophores, is an important class of insect pigments (2– 4). The genes respon- sible for pigment metabolism have been systematically stud- ied in many insects, including Drosophila melanogaster, Manduca sexta, and Papilio xuthus (2, 3, 5– 8). The melanin is derived from tyrosine (10, 11). Tyrosine hydroxylase and dopa decarboxylase convert tyrosine into dopa and dopa into dopamine, respectively (2). Dopa serves as the precursor of dopa melanin (which is black) and is converted to dopa mel- anin by the activities of the Yellow, Yellow-f 1 and Yellow-f 2 proteins (12). Furthermore, dopamine serves as the precur- sor of dopamine melanin (black or brown in color and a major pigment of the insect cuticle), the change being caused by the catalytic action of phenol oxidases (1, 2, 3, 5, 6, 12–14). The current study confirmed that the dopamine content was elevated in melanized insects or in the melanized regions of insects (5, 15). Alternatively, dopamine can reversibly convert to N--alanyldopamine (NBAD, 3 yellowish) by NBAD synthe- tase (EBONY) and NBAD hydrolase. NBAD deposited in the cuticle of an insect causes a yellow coloration. Finally, a family of arylalkylamine-N-acetyltransferases (AANATs) convert dopamine into N-acetyldopamine, a precursor of the colorless N-acetyldopamine sclerotin (2, 3). In insects, AANATs are essen- tial for sclerotization of the cuticle and catabolism of monoamines (16, 17). However, they might also be related to melanism in insects (15). In addition, the activities of AANAT and melatonin display day/night fluctuations in the head tissues of the silkworm (Bombyx mori) and in cockroach (Periplaneta americana) (18– 20). Tsugehara et al. (21) cloned and characterized full-length cDNA of Bm-iAANAT in silkworm. At present, nothing is known about the relationship between Bm-iAANAT and pigmentation metabolism or pigment mutants. Silkworm, B. mori, is an important economic insect and lep- idopteran molecular model (22). More than 600 mutant sys- tems are maintained in the silkworm gene bank at Southwest University (Chongqing, China). More than 100 of these are related to body color and body markings (23), and they are distributed over all of the silkworm linkage groups except the 27th and 28th. Therefore, silkworm provides an excellent model for studying the genetic and molecular mechanisms that generate varied pigment patterns. Recently, the candidate genes of some color pattern mutants such as ch, so (7), lem (24), and rb (25) * This work was supported by National Basic Research 973 Program of China Grant 2005CB121000, Hi-Tech Research and Development 863 Program of China Grant 2006AA10A117, National Natural Science Foundation of China Grant 30671591, Science Foundation of Ministry of Education of China Grant 20060635016, and Technological Innovative Foundation Project for postgraduates of Southwest University of China Grant 2006052. □ S The on-line version of this article (available at http://www.jbc.org) contains supplemental Tables 1– 6 and Figs. 1 and 2. The nucleotide sequences reported in this paper have been submitted to the Gen- Bank TM /EBI Data Bank with accession number(s) GQ169236 and GQ169237. 1 These authors contributed equally to this work. 2 To whom correspondence should be addressed: Key Sericultural Laboratory of Agricultural Ministry, Institute of Sericulture and Systems Biology, Southwest University, Chongqing 400715, China. Tel.: 86-23-68250346; Fax: 86-23-68251128; E-mail: [email protected]. 3 The abbreviations used are: NBAD, N--alanyldopamine; EBONY, N--ala- nyldopamine synthetase; AANAT, arylalkylamine-N-acetyltransferase; WT, wild-type; SSR, simple sequence repeat; RT, reverse transcription; HPLC, high performance liquid chromatography. THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 285, NO. 25, pp. 19553–19560, June 18, 2010 © 2010 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. JUNE 18, 2010 • VOLUME 285 • NUMBER 25 JOURNAL OF BIOLOGICAL CHEMISTRY 19553 by guest on July 27, 2018 http://www.jbc.org/ Downloaded from
-
Upload
truongthuan -
Category
Documents
-
view
216 -
download
0
Transcript of MutationsofanArylalkylamine-N-acetyltransferase, Bm-iAANAT ... ·...

Mutations of an Arylalkylamine-N-acetyltransferase, Bm-iAANAT,Are Responsible for Silkworm Melanism Mutant*□S
Received for publication, December 18, 2009, and in revised form, March 18, 2010 Published, JBC Papers in Press, March 23, 2010, DOI 10.1074/jbc.M109.096743
Fang-yin Dai‡1, Liang Qiao‡1, Xiao-ling Tong‡§1, Cun Cao‡, Peng Chen‡, Jun Chen‡, Cheng Lu‡2, and Zhong-huai Xiang‡
From the ‡College of Biotechnology, Institute of Sericulture and Systems Biology, Southwest University, Chongqing 400715, Chinaand the §Institute of Agriculture and Life Science, Chongqing University, Chongqing 400030, China
Coloration is one of the most variable characters in animalsand provides richmaterial for studying the developmental geneticbasis of pigment patterns. In the silkworm, more than 100 genemutation systems are related to aberrant color patterns. The mel-anism (mln) is a rare body colormutant that exhibits an easily dis-tinguishable phenotype in both larval and adult silkworms. Bypositional cloning, we identified the candidate gene of the mlnlocus,Bm-iAANAT, whose homologous gene (Dat) converts dopa-mine into N-acetyldopamine, a precursor for N-acetyldopaminesclerotin inDrosophila. In themlnmutant, two types of abnormalBm-iAANAT transcripts were identified, whose expression levelsaremarkedly lower than thewild type (WT).Moreover, dopaminecontent was approximately twice as high in the sclerified tissues(head, thoracic legs, and anal plate) of themutant as inWT, result-ing in phenotypic differences between the two. Quantitativereverse transcription PCR analyses showed that other genesinvolved in the melanin metabolism pathway were regulated bythe aberrant Bm-iAANAT activity in mln mutant in differentways and degrees.We therefore propose that greater accumula-tion of dopamine results from the functional deficiency of Bm-iAANAT in themutant, causing a darker pattern in the sclerifiedregions than in the WT. In summary, our results indicate thatBm-iAANAT is responsible for the color pattern of the silkwormmutant,mln. To our knowledge, this is the first report showing arole for arylalkylamine-N-acetyltransferases in color patternmutation in Lepidoptera.
Coloration is one of the most variable traits in insects, andpigmentation is known to play a role in mimicry, sexual selec-tion, thermoregulation, and other adaptive processes in manyinsect groups (1). More recently, there has been much interestamong biologists in the molecular mechanisms underlying thegreat diversity of insect colors and color patterns.
Melanin, the dark pigment found in melanophores, is animportant class of insect pigments (2–4). The genes respon-sible for pigment metabolism have been systematically stud-ied in many insects, including Drosophila melanogaster,Manduca sexta, and Papilio xuthus (2, 3, 5–8). The melaninis derived from tyrosine (10, 11). Tyrosine hydroxylase anddopa decarboxylase convert tyrosine into dopa and dopa intodopamine, respectively (2). Dopa serves as the precursor ofdopa melanin (which is black) and is converted to dopa mel-anin by the activities of the Yellow, Yellow-f 1 and Yellow-f 2proteins (12). Furthermore, dopamine serves as the precur-sor of dopamine melanin (black or brown in color and amajor pigment of the insect cuticle), the change being causedby the catalytic action of phenol oxidases (1, 2, 3, 5, 6, 12–14).The current study confirmed that the dopamine content waselevated in melanized insects or in the melanized regions ofinsects (5, 15). Alternatively, dopamine can reversibly convertto N-�-alanyldopamine (NBAD,3 yellowish) by NBAD synthe-tase (EBONY) and NBAD hydrolase. NBAD deposited in thecuticle of an insect causes a yellow coloration. Finally, a familyof arylalkylamine-N-acetyltransferases (AANATs) convertdopamine into N-acetyldopamine, a precursor of the colorlessN-acetyldopamine sclerotin (2, 3). In insects, AANATs are essen-tial for sclerotization of the cuticle and catabolismofmonoamines(16, 17). However, they might also be related to melanism ininsects (15). In addition, the activities of AANAT and melatonindisplay day/night fluctuations in the head tissues of the silkworm(Bombyx mori) and in cockroach (Periplaneta americana) (18–20). Tsugehara et al. (21) cloned and characterized full-lengthcDNAofBm-iAANAT in silkworm. At present, nothing is knownabout the relationship between Bm-iAANAT and pigmentationmetabolism or pigment mutants.Silkworm, B. mori, is an important economic insect and lep-
idopteran molecular model (22). More than 600 mutant sys-tems are maintained in the silkworm gene bank at SouthwestUniversity (Chongqing, China). More than 100 of these arerelated to body color and body markings (23), and they aredistributed over all of the silkworm linkage groups except the27th and 28th. Therefore, silkworm provides an excellentmodel for studying the genetic andmolecular mechanisms thatgenerate variedpigment patterns. Recently, the candidate genes ofsome color patternmutants such as ch, so (7), lem (24), and rb (25)
* This work was supported by National Basic Research 973 Program of ChinaGrant 2005CB121000, Hi-Tech Research and Development 863 Program ofChina Grant 2006AA10A117, National Natural Science Foundation of ChinaGrant 30671591, Science Foundation of Ministry of Education of ChinaGrant 20060635016, and Technological Innovative Foundation Project forpostgraduates of Southwest University of China Grant 2006052.
□S The on-line version of this article (available at http://www.jbc.org) containssupplemental Tables 1– 6 and Figs. 1 and 2.
The nucleotide sequences reported in this paper have been submitted to the Gen-BankTM/EBI Data Bank with accession number(s) GQ169236 and GQ169237.
1 These authors contributed equally to this work.2 To whom correspondence should be addressed: Key Sericultural Laboratory
of Agricultural Ministry, Institute of Sericulture and Systems Biology,Southwest University, Chongqing 400715, China. Tel.: 86-23-68250346;Fax: 86-23-68251128; E-mail: [email protected].
3 The abbreviations used are: NBAD, N-�-alanyldopamine; EBONY, N-�-ala-nyldopamine synthetase; AANAT, arylalkylamine-N-acetyltransferase; WT,wild-type; SSR, simple sequence repeat; RT, reverse transcription; HPLC,high performance liquid chromatography.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 285, NO. 25, pp. 19553–19560, June 18, 2010© 2010 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
JUNE 18, 2010 • VOLUME 285 • NUMBER 25 JOURNAL OF BIOLOGICAL CHEMISTRY 19553
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from

have been cloned. However, the molecular bases of many othermutations are still unknown.Themln (melanism) is a rare body colormutant that exhibits
a clearly recognizable phenotype both in larval and adult B.
mori. It is regulated by a single recessive gene and has beenmapped at 41.5 cM on the 18th linkage in the silkworm geneticlinkage map. In the recessive homozygote of the mln mutant,the larval head, anal plate, thoracic legs, and sieve plates of thespiracles are black. In the adult, the body, wings, and integu-ment are also black instead of the normal color pattern (Fig. 1).Excessmelanin appears to accumulate in themlnmutant.How-ever, the candidate gene responsible has not been identifiedpreviously.In the present study, we finemapped themln locus and iden-
tified the candidate gene responsible, Bm-iAANAT. In addi-tion, we found differences between Dazao and Dazao-mln intheir gene sequences and transcription levels of Bm-iAANTAand investigated the differences in their expression patterns ofother key genes involved in the melanin metabolism pathway.Based on our experimental results, we conclude that the Bm-iAANAT gene plays an essential role in the pigmentmetabolismin silkworm and that the abnormal Bm-iAANAT is responsiblefor themlnmutant.
EXPERIMENTAL PROCEDURES
Silkworm Strains—The melanism mutant strain Dazao-mln(near isogenic line of mln), wild-type (WT) strain Dazao andC108were obtained from the silkworm gene bank in SouthwestUniversity. The silkworms were reared on mulberry leavesunder a photoperiod of 12 h/12 h light/dark at 25 °C during theexperiments.Fine Mapping of mln Locus and Annotation Analysis of Can-
didate Gene—Two silkworm strains, C108 (�mln/�mln) andDazao-mln (mln/mln) were used for preliminary mapping. Asingle-pair cross between a female (C108) and a male (Daza-
mln) produced the F1 offspring. Forlinkage analysis, 20 BC1F progeny(10 wild-type and 10 melanism off-spring) from the cross (C108 �Dazao-mln) � �Dazao-mlnZwereused, although another backcross,Dazao-mln� � (C108 � Dazao-mln) Z was used for recombinationanalysis. SSR (simple sequence re-peat) markers were obtained fromthe published SSRs linkage map(26). Segregation patterns were ana-lyzed usingMapmaker/exp (27) andtheKosambimapping function (28).Based on the results of the pre-
liminary SSR mapping, we analyzedthe upstream and downstream se-quences close to the tightly linkedSSR marker of the mln locus. Thisprogram requires the silkworm 9�assembly genome database (29),BLAST (30), and primer 5.0. Weused the predicted gene fragmentsas query sequences to search theNCBI database with the BLASTpprogram and found the homologand ortholog genes that have been
FIGURE 1. Phenotype of WT and mln mutant strain. A, WT (Dazao,�mln/�mln) and mln mutant strain (Dazao-mln, mln/mln) larvae are shown onday 1 of the 5th instar. In the mln mutant, the head, anal plate, thoracic legs,sieve plates of the spiracle, and claw hook of prolegs of larvae are blacker ormore buffy compared with the WT. B, body, scales, dorsal plate, vein, thoraciclegs, and tentacle of the mln adult are also blacker than the WT. Scale bar,1 cm. Magnification, indicated in the panels.
FIGURE 2. Preliminary and fine mapping of the mln locus. A, marker S1807 was tightly linked with the mlnlocus. The upstream and downstream sequences (�300 kb on each side) of S1807 were analyzed. BGIB-MGA008538 (Bm-iAANAT) and BGIBMGA008539 were selected as the candidate genes. B, marker of Bm-iAANAT,P3C, shows polymorphism between Dazao-mln and C108. Short lines indicate the markers. C, fine mapping ofthe mln locus was done using P3C. There is no recombination between P3C and the mln locus. The resultsindicate that P3C was also tightly linked with the mln locus.
Functional Defect of Bm-iAANAT in Silkworm Melanism Mutant
19554 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 25 • JUNE 18, 2010
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from

shown to be involved in the pigmentation pathway in otherspecies.We then designed primers as newmarkers based on thegenome sequences of the predicted candidate gene. Thegenome fragment of candidate gene that exhibited polymor-phism between the parents was used for fine mapping. Theprimers are shown in supplemental Tables 1 and 2.
Cloning Full cDNA Sequence of mln Candidate Gene—Thetotal RNAswere extracted fromDazao andDazao-mln on day 4of the 5th instar. The primers (supplemental Table 3) for thefull-length cDNA of the candidate gene were designed accord-ing to the sequence in GenBank (21). The PCR products werecloned into the pMD19-T vector (Takara) and sequenced.
FIGURE 3. Schematic diagram of Bm-iAANAT sequences of WT (Dazao) and mutant (Dazao-mln). A, amplification of genome using P3C primers betweenDazao and Dazao-mln. Double horizontal lines indicate the last 67 bp of exon 4 of Bm-iAANAT in Dazao. Arrows indicate splicing sites of intron 4 in Dazao andDazao-mln, respectively. The box indicates a 29-bp-long insertion, the first 15 bp being included in the aberrant open reading frame of exon 4 in Dazao-mln.B, identification of the full-length cDNA (no poly(A) tag) of Bm-iAANAT in Dazao-mln (mutant) and Dazao (WT). Two abnormal transcripts were detected in themutant strain. C, comparison of the structure of Bm-iAANAT between Dazao and Dazao-mln. White boxes indicate open reading frames. Coarse horizontal linesindicate untranslated regions (3�-UTR). Diagonal lines indicate introns. Gray region indicates the deletion. Gray solid box indicates the insertion of 15 bp. Blackbox indicates abnormal exon 5. Vertical arrows indicate the location of the termination codon. The termination codon of type 1 is located at �34 bp of exon 5.The termination codon of type 2 is located at �74 bp of 3� UTR. D, alignments of the speculative Bm-iAANAT amino acid sequences among Dazao, Dazao-mlntype 1 and type 2. Horizontal line indicates the acetyltransferase domain of the WT strain, Dazao.
Functional Defect of Bm-iAANAT in Silkworm Melanism Mutant
JUNE 18, 2010 • VOLUME 285 • NUMBER 25 JOURNAL OF BIOLOGICAL CHEMISTRY 19555
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from
Expression Profile Analysis of mln Candidate Gene—Toinvestigate the expression patterns, we used semiquantitativeRT-PCR to analyze total RNAs isolated from the whole body ofseveral developmental stages (fromday 3 of the 4th instar larvaeto eclosion of the adult). Total RNAswere purified usingTRIzol(Invitrogen) according to themanufacturer’s protocol and sub-jected to cDNA synthesis using oligo(dT) primers and a Molo-ney murine leukemia virus reverse transcriptase (Promega)according to themanufacturer’s instructions. Ten silkworm tis-sues were sampled on day 4 of the 5th instar larvae of Dazao,including the silk gland, midgut, malpighian tubule, fat body,testis, ovary, head, integument, hemocyte, and trachea andusedto investigate the pattern of spatial expression. The primersdesigned for RT-PCR are shown in supplemental Tables 4. TheBmActin3 gene was used as an internal control. The expressionlevels of the candidate gene in sclerified (anal plate, thoraciclegs, and head) and less sclerified tissues (epidermis) of theDazao larvae were detected using quantitative RT-PCR.Transcriptional Level Analysis of Candidate Gene between
WT and Mutant Strains by Quantitative RT-PCR—We per-formed the quantitative RT-PCR using the ABI Prism 7000sequence detection system (Applied Biosystems) with a SYBRPremix EX Taq kit (Takara) to detect the expression levels ofthe Bm-iAANAT gene in both Dazao (WT) and Dazao-mln
(mutant) at the same developmentalstages and in the same sampled tis-sues (head, thoracic legs, and analplate). The quantitative RT-PCRprimers used are shown in supple-mental Tables 5. Eukaryotic transla-tion initiation factor 4A (silkwormmicroarray probe ID: sw22934) wasused as an internal control.Quantitative RT-PCR Detection
of OtherMelaninMetabolismGenesin Dazao and Dazao-mln—Refer-ring to the melanin metabolismpathway in D. melanogaster (2, 3)and the full-length cDNA se-quences of Pale (AB439286), Yellow(AB438999), Ddc (AF372836), andebony (AB439000) in silkworm, wedesigned primers for use in quanti-tative RT-PCR (supplemental Ta-ble 6). The sw22934 was used as aninternal control.Analysis of Dopamine Content in
WT and Mutant—Dopamine wasextracted from the strain-specificphenotype tissues (head, thoraciclegs, and anal plate: total quantity,200 mg.) on day 2 of 4th instar mlnandWT. The tissues were homoge-nized in 0.5ml of 1.2 MHCl contain-ing 5 mM ascorbic acid in a centri-fuge tube and centrifuged at 14,000rpm, 4 °C for 10 min. Supernatantswere incubated at 100 °C for 10 min
and then added to 0.5ml chloroform and shaken before furthercentrifugation at 12,000 rpm at 4 °C for 10 min. Supernatantswere analyzed by HPLC. Chromatography was performed on aWaters 1525 binary HPLC equipped with aWaters 2487 dual �absorbance detector and a SymmetryShield RP18 (5 �m, 4.6 �150-mm) column. The flow velocity of the mobile phases(69.5 mM KH2PO4, 5.5 mM K2HPO4 (pH 5.8):methanol(Sigma; chromatography pure) 98:2) was 1 ml/min, isocraticelution. The column elution was monitored at 280 nm. Thestandard sample was dopamine hydrochloride (Sigma,supplemental Fig. 2).
RESULTS
Mapping of mln Locus and Identification of Candidate Gene—To determine the chromosomal location of the mln locus, weperformed genetic linkage analysis. First, using the microsatel-lite (SSR) markers on the 18th linkage in the silkworm SSRmolecular linkage map, we roughly mapped the mln mutationusing 372 BC1M individuals. The preliminarymapping showedthat the marker S1807 was tightly linked with the mln locus(Fig. 2). We analyzed the upstream and downstream sequences(�300 kb on each side) of S1807 in the silkworm database (31).Within �100 kb upstream of the S1807 marker, we found twogenes (BGIBMGA008538 and BGIBMGA008539) encoding
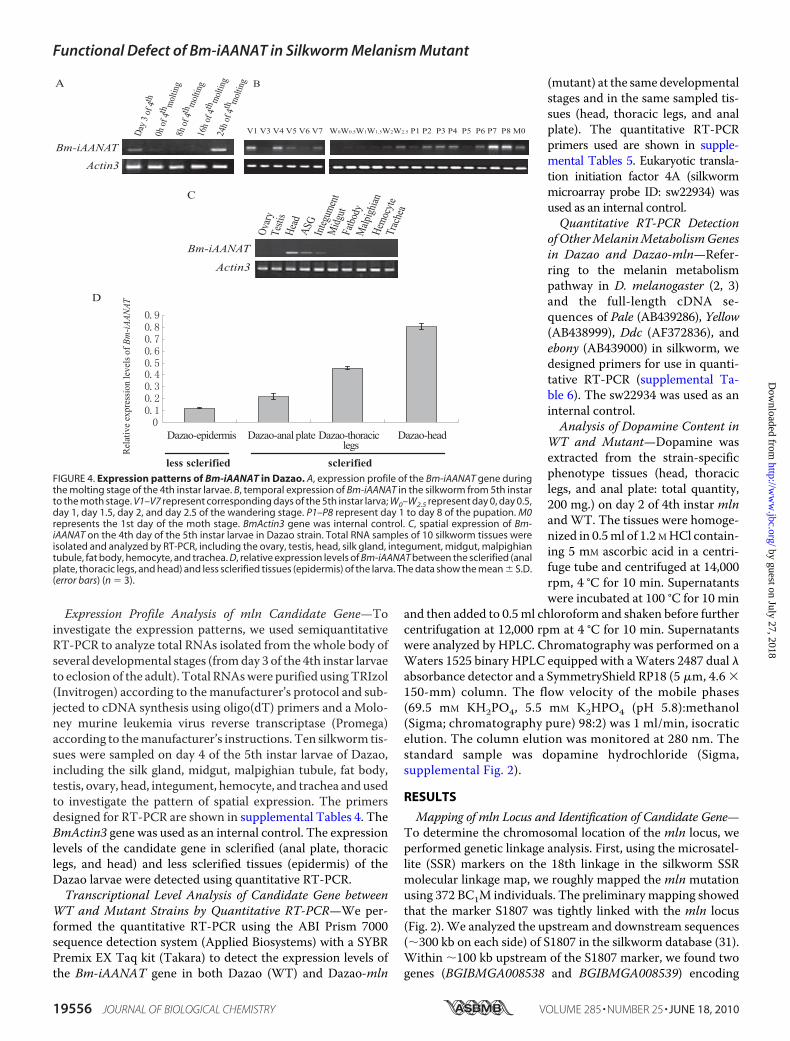
FIGURE 4. Expression patterns of Bm-iAANAT in Dazao. A, expression profile of the Bm-iAANAT gene duringthe molting stage of the 4th instar larvae. B, temporal expression of Bm-iAANAT in the silkworm from 5th instarto the moth stage. V1–V7 represent corresponding days of the 5th instar larva; W0–W2.5 represent day 0, day 0.5,day 1, day 1.5, day 2, and day 2.5 of the wandering stage. P1–P8 represent day 1 to day 8 of the pupation. M0represents the 1st day of the moth stage. BmActin3 gene was internal control. C, spatial expression of Bm-iAANAT on the 4th day of the 5th instar larvae in Dazao strain. Total RNA samples of 10 silkworm tissues wereisolated and analyzed by RT-PCR, including the ovary, testis, head, silk gland, integument, midgut, malpighiantubule, fat body, hemocyte, and trachea. D, relative expression levels of Bm-iAANAT between the sclerified (analplate, thoracic legs, and head) and less sclerified tissues (epidermis) of the larva. The data show the mean � S.D.(error bars) (n � 3).
Functional Defect of Bm-iAANAT in Silkworm Melanism Mutant
19556 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 25 • JUNE 18, 2010
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from

AANAT which plays an important role in the synthesis ofN-acetyldopamine in the pigmentation pathway of D. melano-gaster. We designed primer sets based on the sequences of thetwo genes to detect the genomes of Dazao-mln and C108. Onepair of primers, named P3C, fromBGIBMGA008538, displayeda smaller amplification product in Dazao-mln than in C108and showed the same polymorphism between Dazao-mlnand its recurrent parent, Dazao. We used P3C as a newmarker for fine mapping. The result suggested that there wasno recombination between P3C and the mln locus (Fig. 2).Furthermore, the sequences amplified by the primers of P3Care partly located on the exon of the BGIBMGA008538,which has been previously identified as Bm-iAANAT (GenBankaccession number NM_001079654.1). Therefore, we con-cluded that the Bm-iAANAT gene was the candidate gene forthe melanism (mln) mutant.Polymorphisms in Bm-iAANAT—Cloning and comparing
the sequences of the P3C-amplified region in Dazao, C108, andDazao-mln, we found that it was 410 bp and 506 bp in theDazao-mln and wild-type strains (C108, Dazao), respectively.The mutation deleted 96 bp and inserted another 29-bp frag-ment compared with the WT (Fig. 3A). In addition, we clonedthe complete Bm-iAANAT cDNA sequences of the WT andmln mutant. A significant amount of a 1460-bp fragment wasamplified from the WT (Fig. 3B). Compared with the genomeassembly data, it was apparent that Bm-iAANAT has 5 exonsencoding 261 amino acids. However, two types of abnormaltranscript, type 1 cDNA (1253 bp) and type 2 cDNA (1407 bp),were amplified in the Dazao-mln mutant (Fig. 3B). As shownschematically in Fig. 3C, the type 1 cDNA lacked almost thewhole 4th exon except for the first 2 bp and causes a prematureTAG stop codon not found in the WT. Type 2 lacked the last67 bp of the 4th exon, and recruited a 15-bp length intronsequence. The termination codon of type 2 was located on the3�-untranslated region of the normal transcript (Fig. 3C). Thus,type 1 and type 2 both encoded for potentially aberrant proteinsand destroyed the acetyltransferase 1 domain compared withthe WT (Fig. 3D).Expression Profile Analysis of Bm-iAANAT Gene in Normal
Strain—Temporal expression patterns of the Bm-iAANATgene were detected in the whole body of Dazao during thedevelopmental period from the 3rd day of the 4th instar larvaeto eclosion of the adult (Fig. 4, A and B). The semi-RT-PCRanalysis showed that the expression levels of Bm-iAANAT fluc-tuated during the development period. It was expressed nor-mally during the 3rd day of the 4th instar larvae (intermolt),turned off at the beginning of the fourth molting stage (from 0to 16 h), and then reappeared in the postmolt stage (24 h later)(Fig. 4A). Relatively high expression levels were maintainedduring most of the 5th instar larval period, and no expressionwas detected at the commencement of the wandering stage. Asharp increase followed at the end of thewandering stage (W2.5day), which is opposite to the change of a 20E titer at the sametime (32). The greatest expression appeared on the seventh (P7)and eighth (P8) day following pupation (Fig. 4B).To investigate the spatial expression patterns of the
Bm-iAANAT gene, we also used semi-RT-PCR to analyzetotal RNA in samples of 10 silkworm tissues from 4th day 5th
instar larvae of Dazao. The results showed that theBm-iAANAT gene is expressed in the head, silk gland, andintegument. The highest level of expression was found in thehead. In other tissues, its expression was very low or nearlyundetectable (Fig. 4C).To examine the relationship between the expression of
Bm-iAANAT and the body regions where the mln strainexhibited a different color compared with the WT, we per-formed a quantitative RT-PCR analysis using cDNA pre-pared from the less sclerified epidermis and the more scleri-fied head, thoracic legs, and anal plate of 5th instar Dazaolarvae. The Bm-iAANATwas found to be expressed at higherlevels in sclerified tissues (head, thoracic legs, and anal plate)than in less sclerified tissue (epidermis) (Fig. 4D). In otherwords, high expression of Bm-iAANAT correlated with theblack-colored regions but not with the non—black-coloredregion in the mln mutant.Expression Level of Bm-iAANAT Gene in Melanism (mln)
Mutant Compared with Normal Strain Dazao—We comparedthe expression of Bm-iAANAT between Dazao-mln and Dazaoat several developmental stages, including the 4th day of the 5thinstar larva (V4), the 2nd day of pupation (P2), and at the eclo-sion of the adult (M0), when Bm-iAANAT normally showed its
FIGURE 5. Comparison of the expression levels of Bm-iAANAT betweenDazao and Dazao-mln in the several developmental stages and strain-specific phenotype tissues (head, thoracic legs, and anal plate). A, semi-quantitative RT-PCR detection, BmActin3 as inner control. Lane 1, Dazao-mln.Lane 2, Dazao. B, quantitative PCR analysis. Expression levels were normalizedto the expression of the eukaryotic translation initiation factor 4A gene(microarray probe ID, sw22934). C, relative expression levels of Bm-iAANAT inthree strain-specific phenotype tissues of the larva in Dazao and Dazao-mln.The data indicate the mean � S.D. (error bars) (n � 3).
Functional Defect of Bm-iAANAT in Silkworm Melanism Mutant
JUNE 18, 2010 • VOLUME 285 • NUMBER 25 JOURNAL OF BIOLOGICAL CHEMISTRY 19557
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from

highest activity. The amplified sequences were shared by thethree types of Bm-iAANAT cDNA. The result of the semiquan-titative RT-PCR showed that the level of expression of the twotranscripts in the mln mutant was lower than that of the nor-mal transcript in Dazao (Fig. 5A). As a more rigorous test, weperformed a quantitative RT-PCR analysis with the samemate-rials and showed that the level of expression of normal Bm-iAANAT in WT is about 10–20 times higher than that of thetwo transcripts in the mln mutant (Fig. 5B). In addition, the
expression level of Bm-iAANAT inDazao was also higher than that inthe mln mutant in the regionswhich exhibit obvious phenotypicdifferences between WT and mln(head, thoracic legs, and analplate) (Fig. 5C).Expression Diversification of Key
Genes Involved in Melanin Metabo-lism Pathway between Dazao andDazao-mln—To test how othergenes involved in the melaninmetabolism pathway respond toBm-iAANAT level changes, we per-formed a quantitative RT-PCR anal-ysis to detect the expression levels ofseveral genes (Pale, DdC, Yellow,and ebony) in the regions with dif-ferent Dazao and Dazao-mln col-oration (head, thoracic legs, andanal plate). The results show thatexpression of Pale,DdC, and Yellowin these regions were all higher inDazao than in Dazao-mln, whereasthe expression of ebonywas lower inDazao than in Dazao-mln, except inthe head (Fig. 6A). In the adultmoth, the expression of all four ofthe above genes was higher inDazao than in Dazao-mln, theratios of relative expression levels(Dazao-mln/Dazao) of DdC andebony being 0.471 and 0.573,respectively (Fig. 6B).Contents of Dopamine inWT and
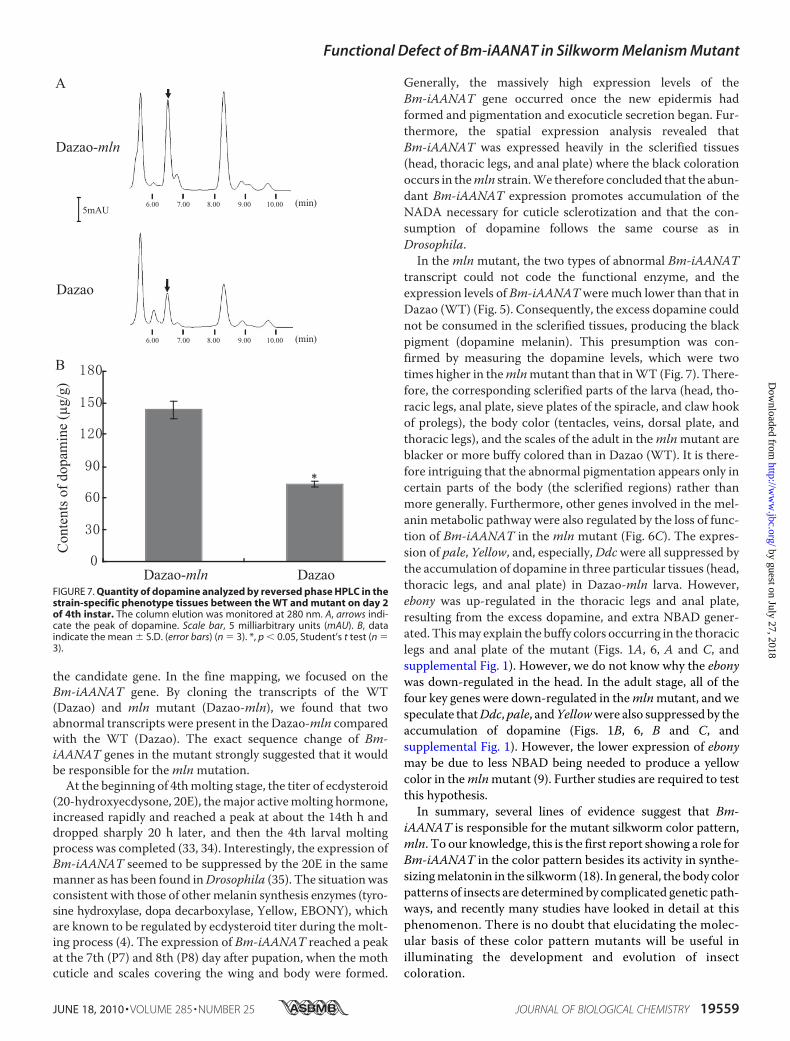
mln Mutant—The result of a re-versed phase HPLC revealed thatthe content of dopamine in themlnmutant (143.233 � 11.821 �g/g)was �2 times higher than that inthe WT (72.953 � 2.022 �g/g)(Fig. 7). Therefore, we concludedthat a greater accumulation ofdopamine resulted from the func-tional deficiency of Bm-iAANATin the mutant and that the exces-sive dopamine was converted intodopamine melanin, causing thedarker color pattern of the scleri-
fied regions in the mln mutant compared with the WT.
DISCUSSION
In this study, we fine map the candidate gene of the mlnmutant using SSR markers and material from the silkwormgenome database. We confirmed the results using the Dazaoand the near isogenic line Dazao-mln obtained by backcrossingto a Dazao strain for more than 20 generations. This provided apowerful combination of genetic material with which to map
FIGURE 6. Investigation of the expression differences of key melanin metabolism genes betweenDazao and Dazao-mln. A, quantitative RT-PCR detection of melanin metabolism key genes in three larvastrain-specific phenotype tissues between Dazao and Dazao-mln on V1 stage. Error bars, S.D. B, result ofinvestigations on M0 stage. **, p � 0.002, Student’s t test. n � 3. Ratio � (mean of quantitative PCR forDazao-mln)/(mean of quantitative PCR for Dazao). C, diagram of the melanin metabolism pathway (seeRef. ref. 3). Black arrows indicate the expression pattern in WT. Green color indicates expression pattern inmln mutant. Solid green arrows indicate the up-regulated expression of ebony in thoracic legs (TL) and analplate (AP) of the larva compared with WT. Dotted line arrows indicate the down-regulated expression of themoth compared with WT. Frame indicates the accumulation of dopamine in mln mutant. Fork indicatesthe loss of function of Bm-iAANAT in mln mutant. POs, phenol oxidases. The genes are shown in blue. Thepigment precursors are shown in red.
Functional Defect of Bm-iAANAT in Silkworm Melanism Mutant
19558 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 25 • JUNE 18, 2010
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from
the candidate gene. In the fine mapping, we focused on theBm-iAANAT gene. By cloning the transcripts of the WT(Dazao) and mln mutant (Dazao-mln), we found that twoabnormal transcripts were present in the Dazao-mln comparedwith the WT (Dazao). The exact sequence change of Bm-iAANAT genes in the mutant strongly suggested that it wouldbe responsible for themlnmutation.
At the beginning of 4thmolting stage, the titer of ecdysteroid(20-hydroxyecdysone, 20E), themajor activemolting hormone,increased rapidly and reached a peak at about the 14th h anddropped sharply 20 h later, and then the 4th larval moltingprocess was completed (33, 34). Interestingly, the expression ofBm-iAANAT seemed to be suppressed by the 20E in the samemanner as has been found inDrosophila (35). The situationwasconsistent with those of othermelanin synthesis enzymes (tyro-sine hydroxylase, dopa decarboxylase, Yellow, EBONY), whichare known to be regulated by ecdysteroid titer during the molt-ing process (4). The expression of Bm-iAANAT reached a peakat the 7th (P7) and 8th (P8) day after pupation, when the mothcuticle and scales covering the wing and body were formed.
Generally, the massively high expression levels of theBm-iAANAT gene occurred once the new epidermis hadformed and pigmentation and exocuticle secretion began. Fur-thermore, the spatial expression analysis revealed thatBm-iAANAT was expressed heavily in the sclerified tissues(head, thoracic legs, and anal plate) where the black colorationoccurs in themln strain.We therefore concluded that the abun-dant Bm-iAANAT expression promotes accumulation of theNADA necessary for cuticle sclerotization and that the con-sumption of dopamine follows the same course as inDrosophila.In the mln mutant, the two types of abnormal Bm-iAANAT
transcript could not code the functional enzyme, and theexpression levels of Bm-iAANATweremuch lower than that inDazao (WT) (Fig. 5). Consequently, the excess dopamine couldnot be consumed in the sclerified tissues, producing the blackpigment (dopamine melanin). This presumption was con-firmed by measuring the dopamine levels, which were twotimes higher in themlnmutant than that inWT (Fig. 7). There-fore, the corresponding sclerified parts of the larva (head, tho-racic legs, anal plate, sieve plates of the spiracle, and claw hookof prolegs), the body color (tentacles, veins, dorsal plate, andthoracic legs), and the scales of the adult in themlnmutant areblacker or more buffy colored than in Dazao (WT). It is there-fore intriguing that the abnormal pigmentation appears only incertain parts of the body (the sclerified regions) rather thanmore generally. Furthermore, other genes involved in the mel-aninmetabolic pathway were also regulated by the loss of func-tion of Bm-iAANAT in the mln mutant (Fig. 6C). The expres-sion of pale, Yellow, and, especially,Ddcwere all suppressed bythe accumulation of dopamine in three particular tissues (head,thoracic legs, and anal plate) in Dazao-mln larva. However,ebony was up-regulated in the thoracic legs and anal plate,resulting from the excess dopamine, and extra NBAD gener-ated. Thismay explain the buffy colors occurring in the thoraciclegs and anal plate of the mutant (Figs. 1A, 6, A and C, andsupplemental Fig. 1). However, we do not know why the ebonywas down-regulated in the head. In the adult stage, all of thefour key genes were down-regulated in themlnmutant, and wespeculate thatDdc,pale, andYellowwere also suppressed by theaccumulation of dopamine (Figs. 1B, 6, B and C, andsupplemental Fig. 1). However, the lower expression of ebonymay be due to less NBAD being needed to produce a yellowcolor in themlnmutant (9). Further studies are required to testthis hypothesis.In summary, several lines of evidence suggest that Bm-
iAANAT is responsible for the mutant silkworm color pattern,mln. To our knowledge, this is the first report showing a role forBm-iAANAT in the color pattern besides its activity in synthe-sizingmelatonin in the silkworm (18). In general, the body colorpatterns of insects are determined by complicated genetic path-ways, and recently many studies have looked in detail at thisphenomenon. There is no doubt that elucidating the molec-ular basis of these color pattern mutants will be useful inilluminating the development and evolution of insectcoloration.
FIGURE 7. Quantity of dopamine analyzed by reversed phase HPLC in thestrain-specific phenotype tissues between the WT and mutant on day 2of 4th instar. The column elution was monitored at 280 nm. A, arrows indi-cate the peak of dopamine. Scale bar, 5 milliarbitrary units (mAU). B, dataindicate the mean � S.D. (error bars) (n � 3). *, p � 0.05, Student’s t test (n �3).
Functional Defect of Bm-iAANAT in Silkworm Melanism Mutant
JUNE 18, 2010 • VOLUME 285 • NUMBER 25 JOURNAL OF BIOLOGICAL CHEMISTRY 19559
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from

REFERENCES1. True, J. R. (2003) Trends Ecol. Evol. 18, 640–6472. Wright, T. R. (1987) Adv. Genet. 24, 127–2223. Wittkopp, P. J., Carroll, S. B., and Kopp, A. (2003) Trends Genet. 19,
495–5044. Hiruma, K., and Riddiford, L. M. (2009) Insect Biochem. Mol. Biol. 39,
245–2535. Futahashi, R., and Fujiwara, H. (2005) Dev. Genes Evol. 215, 519–5296. Futahashi, R., and Fujiwara, H. (2007) Insect Biochem. Mol. Biol. 37,
855–8647. Futahashi, R., Sato, J., Meng, Y., Okamoto, S., Daimon, T., Yamamoto, K.,
Suetsugu, Y., Narukawa, J., Takahashi, H., Banno, Y., Katsuma, S., Shi-mada, T., Mita, K., and Fujiwara, H. (2008) Genetics 180, 1995–2005
8. Gorman,M. J., An, C., and Kanost, M. R. (2007) Insect Biochem.Mol. Biol.37, 1327–1337
9. Koch, P. B., Behnecke, B., Weigmann-Lenz, M., and Ffrench-Constant,R. H. (2000) Pigment Cell Res. 13, Suppl. 8, 54–58
10. Barsh, G. S. (1996) Trends Genet. 12, 299–30511. Kramer, K. J., and Hopkins, T. L. (1987) Arch. Insect Biochem. Physiol. 6,
279–30112. Han, Q., Fang, J., Ding, H., Johnson, J. K., Christensen, B. M., and Li, J.
(2002) Biochem. J. 368, 333–34013. Hiruma, K., and Riddiford, L. M. (1984) J. Exp. Zool. 230, 393–40314. Hiruma, K., and Riddiford, L. M. (1985) Dev. Biol. 110, 509–51315. Czapla, T. H., Hopkins, T. L., and Kramer, K. J. (1990) J. Insect Physiol. 36,
647–65416. Sasaki, K., and Nagao, T. (2001) J. Insect. Physiol. 47, 1205–121617. Smith, T. J. (1990) Bioessays 12, 30–3318. Itoh, M. T., Hattori, A., Nomura, T., Sumi, Y., and Suzuki, T. (1995)Mol.
Cell. Endocrinol. 115, 59–6419. Ichihara, N., Okada, M., Nakagawa, H., and Takeda, M. (1997) Insect Bio-
chem. Mol. Biol. 27, 241–24620. Ichihara, N., Okada, M., and Takeda, M. (2001) Insect Biochem. Mol. Biol.
32, 15–2221. Tsugehara, T., Iwai, S., Fujiwara, Y., Mita, K., and Takeda, M. (2007)
Comp. Biochem. Physiol. B Biochem. Mol. Biol. 147, 358–36622. Goldsmith, M. R., Shimada, T., and Abe, H. (2005) Annu. Rev. Entomol.
50, 71–10023. Lu, C., Dai, F., and Xiang, Z. (2003) Sci. Agric. Sin. 36, 968–97524. Meng, Y., Katsuma, S., Daimon, T., Banno, Y., Uchino, K., Sezutsu, H.,
Tamura, T., Mita, K., and Shimada, T. (2009) J. Biol. Chem. 284,11698–11705
25. Meng, Y., Katsuma, S., Mita, K., and Shimada, T. (2009) Genes Cells 14,129–140
26. Miao, X. X., Xub, S. J., Li, M. H., Li, M.W., Huang, J. H., Dai, F. Y., Marino,S.W.,Mills, D. R., Zeng, P., Mita, K., Jia, S. H., Zhang, Y., Liu,W. B., Xiang,H., Guo, Q. H., Xu, A. Y., Kong, X. Y., Lin, H. X., Shi, Y. Z., Lu, G., Zhang,X., Huang, W., Yasukochi, Y., Sugasaki, T., Shimada, T., Nagaraju, J.,Xiang, Z.H.,Wang, S. Y., Goldsmith,M. R., Lu, C., Zhao,G. P., andHuang,Y. P. (2005) Proc. Natl. Acad. Sci. U.S.A. 102, 16303–16308
27. Lander, E. S., Green, P., Abrahamson, J., Barlow, A., Daly, M. J., Lincoln,S. E., Newberg, L. A., and Newburg, L. (1987) Genomics 1, 174–181
28. Kosambi, D. D. (1944) Annu. Eugenics 12, 172–17529. Consortium, I. S. G. (2008) Insect Biochem. Mol. Biol. 38, 1036–104530. Altschul, S. F., Madden, T. L., Schaffer, A. A., Zhang, J., Zhang, Z., Miller,
W., and Lipman, D. J. (1997) Nucleic Acids Res. 25, 3389–340231. Duan, J., Li, R., Cheng, D., Fan, W., Zha, X., Cheng, T., Wu, Y., Wang, J.,
Mita, K., Xiang, Z., and Xia, Q. (2010) Nucleic Acids Res. 38, D453–D45632. Mizoguchi, A., Dedos, S. G., Fugo, H., and Kataoka, H. (2002)Gen. Comp.
Endocrinol. 127, 181–18933. Kiguchi, K., and Agui, N. (1981) J. Insect Physiol. 27, 805–81234. Suzuki, Y., Matsuoka, T., Iimura, Y., and Fujiwara, H. (2002) Insect Bio-
chem. Mol. Biol. 32, 599–60735. Rauschenbach, I. Y., Adonyeva, N. V., Alekseev, A. A., Chentsova, N. A.,
and Gruntenko, N. E. (2008) J. Comp. Physiol. B 178, 315–320
Functional Defect of Bm-iAANAT in Silkworm Melanism Mutant
19560 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 25 • JUNE 18, 2010
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from

and Zhong-huai XiangFang-yin Dai, Liang Qiao, Xiao-ling Tong, Cun Cao, Peng Chen, Jun Chen, Cheng Lu
Responsible for Silkworm Melanism Mutant, AreBm-iAANAT-acetyltransferase, NMutations of an Arylalkylamine-
doi: 10.1074/jbc.M109.096743 originally published online March 23, 20102010, 285:19553-19560.J. Biol. Chem.
10.1074/jbc.M109.096743Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2010/03/23/M109.096743.DC1
http://www.jbc.org/content/285/25/19553.full.html#ref-list-1
This article cites 35 references, 2 of which can be accessed free at
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from


















