
Molecular Basis of Spectrin and Ankyrin Deficiencies in Severe
10
Molecular Basis of Spectrin and Ankyrin Deficiencies in Severe Hereditary Spherocytosis: Evidence Implicating a Primary Defect of Ankyrin By Manjit Hanspal, Sa-Hyun Yoon, Huilan Yu, Jatinder S. Hanspal, Stephen Lambert, Jiri Palek, and Josef T. Prchal While varying degrees of spectrin deficiency have been found in the majority of patients with hereditary spherocytosis (HS), a combined severe deficiency of both spectrin and the spectrin-binding protein, ankyrin, has been reported only in two patients with severe HS. To elucidate the molecular basis of these protein deficiencies, we have studied the synthesis, assembly, and the mRNA levels of spectrin and ankyrin in peripheral blood reticulocytes in one of the previ- ously reported probands. Pulse-labeling studies showed that in HS reticulocytes, the synthesis of a-spectrin was compara- ble with control reticulocytes while that of p-spectrin was increased about fourfold, presumably reflecting increased erythropoietic drive. On the HS reticulocyte membrane, the amount of newly assembled spectrin was reduced to about half of the control values, presumablyreflecting a decrease in EREDITARY spherocytosis (HS) is a common inher- H ited hemolytic anemia that is heterogenous in terms of its clinical presentation, molecular basis, and inheritance.' The primary defect is thought to reside in the red blood cell (RBC) membrane skeleton. This submembranous protein network is organized as a hexagonal lattice of fibers of spectrin tetramers that are interconnected by junctional complexes of oligomeric actin, with the aid of proteins 4.1, adducin, and possibly other proteins including myosin, tropomyosin, and band 4.9?-5 Each of the spectrin tetramers is composed of the ap-spectrin heterodimers that self- assemble at their head region into tetramers. The skeleton is attached to the plasma membrane via the association of P-spectrin with ankynn (protein 2.1) and protein 4.1 which, in turn, bind to several integral membrane proteins."' The assembly of the membrane skeleton that occurs during erythroid development has been previously studied in rat and chicken erythroid cells and, to some extent, in erythroleukemic cell lines. In both rat and chicken erythro- blasts, a spectrin is synthesized in a twofold to threefold excess over P-spectrin. The two polypeptides assemble in equimolar amounts on the membrane forming a stable skeletal network, while the excess cytosolic a- and P-spec- trins are rapidly degraded."." In the final stages of erythro- poiesis, the synthesis of ankyrin and protein 4.1 and the incorporation of these proteins into the membrane contin- ues well after the synthesis of other components of the membrane skeleton has been downregulated.'O,'z In the majority of patients with HS, a partial deficiency of spectrin has been observed." In two subsets of HS patients, this spectrin deficiency may represent a primary lesion: (1) in rare HS kindred with an autosomal recessive form of the disease, where HS has been linked to an abnormality involving a-I1 domain of ~pectrin,'~-'~ and (2) in a small subset of patients with dominantly inherited HS, where the primary defect involves abnormal P-spectrin that binds poorly to the protein 4.1.13'7~'8~19 In contrast, in many HS patients, particularly those with the autosomal dominant form of the disease, spectrin deficiency does not seem to represent the primary molecular defect as shown by recent preliminary studies failing to link HS with polymorphism of the synthesis of the spectrin binding protein, ankyrin: the ankyrin synthesis was nearly absent in the cytosol and the amounts of membrane-associated ankyrin were reduced to about half of the normal values. The changes in the amounts of spectrin and ankyrin mRNAs quantitated by slot blot and Northern blot analyses were comparablewith changes in the synthesis of these proteins: The a spectrin mRNA was within a control range and the p-spectrin mRNA was slightly in- creased, while the amounts of ankyrin mRNA were reduced to about 50% of control values. We concludethat the primary defect underlying the combined spectrin and ankyrin defi- ciency is a deficiency of ankyrin mRNA leading to a reduced synthesis of ankyrin which, in turn, underlies the decreased assembly of spectrin on the membrane. 8 1991 by The American Society of Hematology. both a- and P-spectrin.m'21 Several recent studies have raised a possibility that a deficiency or dysfunction of ankyrin may represent a common molecular defect in autosomal dominant HS. For example, HS has been found in association with either interstitial deletion of chromo- some 8 or its translocation to chromosomes 3 and 12.22-z Chromosome 8 has now been shown to contain the gene for ankyrin.&%In a large kindred, the transmission of HS was linked to a polymorphism of ankyrin Furthermore, in one HS patient with interstitial deletion of the eighth chromosome, both the ankyrin DNA and the membrane- associated ankyrin were reduced.z6 We have previously reported a combined severe defi- ciency of both spectrin and ankyrin in two patients with severe atypical HS.M We showed that the residual spectrin and ankyrin present in the patients' RBC membranes were structurally and functionally normal. In our further attempt to determine the molecular basis of these protein deficien- cies in one of the HS patients, we have now examined the transcriptional and translational aspects of the synthesis and assembly of spectrin and ankyrin. Peripheral blood reticulocytes could be obtained because of persisting hemo- lysis after splenectomy. We find that in HS reticulocytes, the synthesis of spectrin was the same as in control From the Department of Biomedical Research, St Elizabeth's Hospital of Boston, TUBS University School of Medicine, Boston, MA; and Division of Hematology, Universityof Alabama in Birmingham. Submitted April 17,1990; acceptedAugust 31,1990. Supported by the National Institutes of Health Grant Nos. POI-HL- 37462 and ROI-HL-27215. Presented in part in abstract form at the 29th annual meeting of the American Society of Hematology in Washington,DC, 1988. Address reprint requests to Manjit Hanspal, PhD, Depament of Bwmedical Research, St Elizabeth's Hospital of Boston, 736 Cam- bridge St, Boston, MA 02135. The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. section I734 soleiy to indicate this fact. 8 1991 by TheAmerican Society of Hematology. 0006-4971 l91l7701-0022$3. OOJO Blood, Vol77, No 1 (January I), 1991: pp 165-173 165 For personal use only. on April 13, 2019. by guest www.bloodjournal.org From
Transcript of Molecular Basis of Spectrin and Ankyrin Deficiencies in Severe

Molecular Basis of Spectrin and Ankyrin Deficiencies in Severe Hereditary Spherocytosis: Evidence Implicating a Primary Defect of Ankyrin
By Manjit Hanspal, Sa-Hyun Yoon, Huilan Yu, Jatinder S. Hanspal, Stephen Lambert, Jiri Palek, and Josef T. Prchal
While varying degrees of spectrin deficiency have been found in the majority of patients with hereditary spherocytosis (HS), a combined severe deficiency of both spectrin and the spectrin-binding protein, ankyrin, has been reported only in two patients with severe HS. To elucidate the molecular basis of these protein deficiencies, we have studied the synthesis, assembly, and the mRNA levels of spectrin and ankyrin in peripheral blood reticulocytes in one of the previ- ously reported probands. Pulse-labeling studies showed that in HS reticulocytes, the synthesis of a-spectrin was compara- ble with control reticulocytes while that of p-spectrin was increased about fourfold, presumably reflecting increased erythropoietic drive. On the HS reticulocyte membrane, the amount of newly assembled spectrin was reduced to about half of the control values, presumably reflecting a decrease in
EREDITARY spherocytosis (HS) is a common inher- H ited hemolytic anemia that is heterogenous in terms of its clinical presentation, molecular basis, and inheritance.' The primary defect is thought to reside in the red blood cell (RBC) membrane skeleton. This submembranous protein network is organized as a hexagonal lattice of fibers of spectrin tetramers that are interconnected by junctional complexes of oligomeric actin, with the aid of proteins 4.1, adducin, and possibly other proteins including myosin, tropomyosin, and band 4.9?-5 Each of the spectrin tetramers is composed of the ap-spectrin heterodimers that self- assemble at their head region into tetramers. The skeleton is attached to the plasma membrane via the association of P-spectrin with ankynn (protein 2.1) and protein 4.1 which, in turn, bind to several integral membrane proteins."'
The assembly of the membrane skeleton that occurs during erythroid development has been previously studied in rat and chicken erythroid cells and, to some extent, in erythroleukemic cell lines. In both rat and chicken erythro- blasts, a spectrin is synthesized in a twofold to threefold excess over P-spectrin. The two polypeptides assemble in equimolar amounts on the membrane forming a stable skeletal network, while the excess cytosolic a- and P-spec- trins are rapidly degraded."." In the final stages of erythro- poiesis, the synthesis of ankyrin and protein 4.1 and the incorporation of these proteins into the membrane contin- ues well after the synthesis of other components of the membrane skeleton has been downregulated.'O,'z
In the majority of patients with HS, a partial deficiency of spectrin has been observed." In two subsets of HS patients, this spectrin deficiency may represent a primary lesion: (1) in rare HS kindred with an autosomal recessive form of the disease, where HS has been linked to an abnormality involving a-I1 domain of ~pectrin, '~- '~ and (2) in a small subset of patients with dominantly inherited HS, where the primary defect involves abnormal P-spectrin that binds poorly to the protein 4.1.13'7~'8~19 In contrast, in many HS patients, particularly those with the autosomal dominant form of the disease, spectrin deficiency does not seem to represent the primary molecular defect as shown by recent preliminary studies failing to link HS with polymorphism of
the synthesis of the spectrin binding protein, ankyrin: the ankyrin synthesis was nearly absent in the cytosol and the amounts of membrane-associated ankyrin were reduced to about half of the normal values. The changes in the amounts of spectrin and ankyrin mRNAs quantitated by slot blot and Northern blot analyses were comparable with changes in the synthesis of these proteins: The a spectrin mRNA was within a control range and the p-spectrin mRNA was slightly in- creased, while the amounts of ankyrin mRNA were reduced to about 50% of control values. We conclude that the primary defect underlying the combined spectrin and ankyrin defi- ciency is a deficiency of ankyrin mRNA leading to a reduced synthesis of ankyrin which, in turn, underlies the decreased assembly of spectrin on the membrane. 8 1991 by The American Society of Hematology.
both a- and P-spectrin.m'21 Several recent studies have raised a possibility that a deficiency or dysfunction of ankyrin may represent a common molecular defect in autosomal dominant HS. For example, HS has been found in association with either interstitial deletion of chromo- some 8 or its translocation to chromosomes 3 and 12.22-z Chromosome 8 has now been shown to contain the gene for ankyrin.&% In a large kindred, the transmission of HS was linked to a polymorphism of ankyrin Furthermore, in one HS patient with interstitial deletion of the eighth chromosome, both the ankyrin DNA and the membrane- associated ankyrin were reduced.z6
We have previously reported a combined severe defi- ciency of both spectrin and ankyrin in two patients with severe atypical HS.M We showed that the residual spectrin and ankyrin present in the patients' RBC membranes were structurally and functionally normal. In our further attempt to determine the molecular basis of these protein deficien- cies in one of the HS patients, we have now examined the transcriptional and translational aspects of the synthesis and assembly of spectrin and ankyrin. Peripheral blood reticulocytes could be obtained because of persisting hemo- lysis after splenectomy. We find that in HS reticulocytes, the synthesis of spectrin was the same as in control
From the Department of Biomedical Research, St Elizabeth's Hospital of Boston, TUBS University School of Medicine, Boston, MA; and Division of Hematology, University of Alabama in Birmingham.
Submitted April 17,1990; accepted August 31,1990. Supported by the National Institutes of Health Grant Nos. POI-HL-
37462 and ROI-HL-27215. Presented in part in abstract form at the 29th annual meeting of the
American Society of Hematology in Washington, DC, 1988. Address reprint requests to Manjit Hanspal, PhD, Depament of
Bwmedical Research, St Elizabeth's Hospital of Boston, 736 Cam- bridge St, Boston, M A 02135.
The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. section I734 soleiy to indicate this fact.
8 1991 by The American Society of Hematology. 0006-4971 l91l7701-0022$3. OOJO
Blood, Vol77, No 1 (January I ) , 1991: pp 165-173 165
For personal use only.on April 13, 2019. by guest www.bloodjournal.orgFrom

166 HANSPAL ET AL
reticulocytes (a-spectrin) or even increased (P-spectrin) as were the amounts of reticulocyte mRNA for both spectrin polypeptides. In a striking contrast, the assembly of spectrin on the membrane was markedly reduced. We further show that the underlying molecular defect involves a reduced synthesis of ankyrin, as indicated by marked decrease of ankyrin synthesis in the cytosol and its reduced assembly on the membrane together with a marked decrease in the amount of the reticulocyte mRNA. Furthermore, the resid- ual ankyrin was unstable as evidenced by the presence of ankyrin degradation products in the cytosol that were not detected in control reticulocytes. Thus, the primary defect involves a reduced synthesis of ankyrin resulting in a reduced incorporation of ankyrin into the RBC membrane which, in turn, leads to a reduced assembly of spectrin on the membrane.
MATERIALS AND METHODS
Subjects. Normal reticulocytes were isolated from the periph- eral blood of patients with sickle cell anemia or other hemolytic disorders and from patients with hemochromatosis undergoing therapeutic phlebotomies.
Clinical data on subject CC has been summarized by Coetzer et aLM In brief, the subject is a 41-year-old white man with chronic hemolytic anemia since infancy, who underwent a splenectomy at 3 months of age, The peripheral blood smear showed marked microspherocytosis, poikilocytosis, and polychromasia. The proband’s deceased father suffered from alcoholic hepatic cirrhosis and anemia; however, he was recorded to have normal hemoglobin concentration and erythrocyte indices at the time of military training, whereas his deceased mother was hematologically normal as are two living siblings.
Venous blood was collected in tubes containing acid citrate dextrose as anticoagulant at the University of Alabama, Birming- ham. Blood was immediately processed for reticulocyte RNA preparation. For synthetic studies, the samples were cooled and transported to Boston, MA overnight on ice in an insulated container and analyzed immediately.
Fractionation of reticulocytes and nucleated RBC precursors. Reticulocytes of sickle cell and pyruvate kinase deficiency patients were used as control reticulocytes in the present study. Reticulo- cytes were isolated from the peripheral blood of control and HS individuals. Blood was centrifuged at 1,OOOg for 5 minutes and the cells were washed three times with Hank’s balanced salt solution (HBSS) without Ca2+ and M g + (M.A. Bioproducts, Walkersville, MD). Four milliliters of this cell suspension was then mixed with 18 mL 75% Percoll (Pharmacia Fine Chemicals, Uppsala, Sweden) in HBSS resulting in a starting density of 1.09 g/mL, and centrifuged in a fixed angle rotor at 45,OoOg for 30 minutes at 4°C. After the run, 1-mL fractions were collected from the top of the gradient and diluted with HBSS. The cells were harvested by centrifugation and reticulocytes were identified by staining with New Methylene Blue (J.T. Baker Chemical Co, Phillipsburg, NJ). RNA content of reticulocytes was determined by means of the orcinol technique.” Reticulocytes in fraction numbers 9 to 12 corresponding to Percoll density of 1.07 g/mL contained 40 f 5 pg of RNA per lo7 cells. These fractions were pooled and used in the present studies.
Human nucleated RBC precursors were isolated from the sternal or rib marrow using a combination of Ficoll-Paque and a discontinuous Percoll gradient, as described earlier.” The sternal and rib marrow represented a discarded marrow during sternot- omy and rib resection, respectively.
Pulse labeling of cells with (”S) methionine. Human reticulocytes or nucleated RBC precursors, 2 to 3 X lo’, were washed twice in minimum essential medium (MEM) without methionine (Flow Laboratories, McLean, VA), resuspended in 10 mL of the same medium containing 10% fetal calf serum prewarmed to 37T , and incubated at 37°C for 15 minutes. They were then labeled for different lengths of time with 300 pCi (35S) methionine (1,000 Ci/mmol; New England Nuclear, Boston, MA). For the pulse-chase experiment, further incorporation of (%) methionine was stopped by the addition of unlabeled methionine (0.4 mmoUL) and the incubation was then continued for different time periods. At the end of the labeling period, 10 vol of 155 mmoUL choline chloride, 5 mmol/L HEPES, pH 7.1, were added and the cells were harvested by centrifugation.
(”S) methionine-labeled cells were treated with Diisopropyl fluorophosphate (Sigma Chemical Co, St Louis, MO) before isolating plasma membranes. Plasma membranes were isolated by using the procedure of Chan?’ The cells were suspended in hypotonic buffer (10 mmol/L Tris HCl, pH 7.5 10 mmoW KCl, and 1.5 mmol/L MgCl,) and then disrupted with 40 strokes of a tight-fitting Dounce homogenizer. An appropri- ate volume of 2 mol/L sucrose was added immediately to the homogenate to restore isotonicity. The homogenate was then layered over a sucrose step gradient (3 ~0128% [wthol] sucrose, 1 vol 50% [wthol] sucrose; sucrose solutions made in 140 mmol/L NaC1, 2.4 mmoW MgCI,, and 5 mmol/L Tris-HC1, pH 7.4) and centrifuged in an SW 41 rotor (Beckman Instruments, Inc, Fuller- ton, CA) at 117,OoOg for 40 minutes. The membrane fraction, at the 28%/50% sucrose interface, and the soluble fraction on top of the sucrose gradient were collected. The membrane fraction was diluted with 20 mmoliL Tris-HC1, pH 7.4, and centrifuged at 22,0008 for 10 minutes. The resulting membrane pellet was washed once more before it was solubilized in sodium dodecyl sulfate (SDS) sample buffer.
Membranes and cell lysates isolated from (’%) methionine-labeled cells were used for immunoprecipitation of spectrin and ankyrin. Spectrin immuno- precipitation was performed as described earlier,” whereas for ankyrin immunoprecipitation, the method used was a modification of Marcantonio and Hynes.” The samples in 0.1 mol/L Tris-HC1, pH 8.5,0.15 mol/L NaC1,5 mmol/L EDTA, 0.5% NP-40 (buffer A) containing 2% SDS were placed in a boiling water bath for 2 minutes and then diluted with an equal volume of buffer A. Triton X-100 was then added to 5% final concentration. The fivefold excess of Triton X-100 (Sigma) forms mixed micelles with SDS, thereby preventing antibody from being denatured by SDS. Antihu- man ankyrin IgG, 70 pL, was then added and the samples incubated at room temperature for 90 minutes with gentle shaking. Thereafter, 100 pL of protein A-Sepharose CL-4B (Pharmacia) (50 mg sepharose beads per milliliter of buffer A) was added and incubation continued for 1 hour at room temperature with gentle shaking. The protein A-sepharose beads were then collected and washed four times with buffer A. The final pellet was resuspended in 70 pL SDS sample buffer and boiled for 2 minutes. Beads were removed by centrifugation and the supernatant was directly loaded on SDS-polyacrylamide gels.
SDS-polyacrylamide gel electrophoresis (SDS-PAGE). Proteins were separated by SDS-PAGE according to the buffer systems of Fairbanks et a134 and Laemmli.” The gels were processed for fluorography with En’Hance (New England Nuclear), dried, and exposed to Kodak XAR-5 x-ray film (Eastman Kodak, Rochester, NY). The autoradiograms were scanned at 570 nm with a Quick- Scann (Helena Laboratories, Beaumont, TX), and the area under each peak was integrated.
Isolation of plasma membranes.
Immunoprecipitation of spectrin and ankyrin.
For personal use only.on April 13, 2019. by guest www.bloodjournal.orgFrom

ANKYRIN DEFICIENCY IN HEREDITARY SPHEROCYTOSIS 167
Preparation of reticulocyte RNA. Blood from the spectrin- and ankyrin-deficient patient-'" and from a patient with reticulocytosis undergoing therapeutic phlebotomies, were used as a source of human reticulocyte RNA. Total reticulocyte RNA was isolated by the method of Goossens and Kan,36 and modified by additional purification of the acid RNA precipitate by the guanidine HCl- phenol method?' Poly (A)' RNA was isolated by oligo(dT)- cellulose chromatography," and by one to three passages through oligo-(dT) resin. All human tissues were obtained from the Tissue Procurement Facility of the University of Alabama at Birmingham (UAB) using protocols previously approved by the UAB Human Use Committee.
Preparation of p-spectrin, a-spectrin, ankyrin, and a-globin cDNA probes. The human p-spectrin cDNA probe was isolated from the plasmid pGemp3.7 previously characterized in our lab~ratory.'~ The human a-spectrin cDNA probe" was prepared from the plasmid pHaSp5 (kindly provided by Dr Peter Curtis, The Wistar Institute of Anatomy and Biology, Philadelphia, PA) by poly- merase chain reaction with two primers flanking the PstI site of pBR 322:1,42 The human ankyrin cDNA probe" was prepared from the plasmid pHankl9A that contains a 2-kb insert spanning part of the spectrin binding domain of ankyrin. The human %-globin cDNA probe (kindly provided by Dr Steven Liebhaber, School of Medicine, University of Pennsylvania) was obtained from plasmid p H d A , and a,-globin c D N A ~ ~ was separated from the plasmid DNA by PstI digestion using standard technique^.'^
Slot blotting and quantitation of spectrin and ankyrin transcripts. Poly (A)' RNA samples were applied on nitrocellulose paper by using a slot blot system (Schleicher & Schuell, Keene, NH). The quantity of each poly (A)' RNA sample was assayed spectrophotometrically.38 The filter was hybridized with an appro- priate cDNA probe labeled with -"P by nick translation. In addition, each nitrocellulose filter was analyzed by %-globin cDNA probe:-' Thus, the amount of globin mRNA in each reticulocyte mRNA sample served as an internal standard of poly (A)+ RNA purity and quantity. The hybridization and washing was performed under stringent conditions as described e l ~ e w h e r e . - ' ~ ~ ~ Autoradio- grams were scanned using an LKB 2400 Ultrascan XL laser densitometer (LKB Produkter, Broma, Sweden) and the amount of message was calculated by comparison of the relative intensity of hybridization of the sample with that of an %-globin message in the same sample. In these studies, the control was a subject of Anglo-Saxon background in whom a possible thalassemia was ruled out by normal globin chain synthesis.
Poly (A)' RNAs were fractionated on 1.4% agarose formaldehyde gels followed by transfer to nitrocel- lulose filters and hybridation to cDNA probe. Conditions for hybridation and washing were performed as previously d e ~ c r i b e d ? ~ . ~ The Northern blot membrane was subsequently rehybridized with a,-globin cDNA probe4-'; thus, the globin mRNA in each sample served as an internal standard of poly (A)+ RNA quantity and purity.
Chromosomal analysis and Southem blotting of ankyrin DNA. Cytogenetic analysis of the spectrin- and ankyrin-deficient patient was performed from a bone marrow aspirate, The chromosomal spreads were examined using a modification of Hozier and Lindquist's techniq~e!~ All metaphases were GTG banded.
Southern blot analyses were performed using standard Two different cDNA human ankyrin probes were
used." Probe 13A is a 1.5-kb probe spanning both spectrin and band 3 binding domains, while probe 25A is a 1.58-kb cDNA probe covering the 5' coding region of ankyrin molecule and extending downstream to the band 3 binding domain. In each Southern blot analysis, at least five control subjects were used. These were
Northern blot hybridization.
patients with hereditaxy spherocytosis and their uninvolved rela- tives. The enzymes used with probe 13A were HindIII, PvuII, EcoRI, BamHI, PstI, Q n l , Sst I, Xba I, TuqI, BglII, and BglI; those used with probe 25A were Q n I , BglI, and BamHI. Similar studies were performed using the a-spectrin genomic and cDNA probes'"," and two p-spectrin cDNA probes covering p7 to p14 repetitive
RESULTS Synthesis, assembb, and tumover of spectrin in reticulocytes.
The synthesis of spectrin was studied in reticulocytes after a pulse-label with ("S) methionine. In the cytosol of control reticulocytes, the synthesis of a- and P-spectrins was compa- rable with that previously reported in nucleated RBCs from the rat and human,10347 with a-spectrin synthesis being twofold to threefold greater than that of P-spectrin. In contrast, on the membrane the two chains of spectrin assembled in equimolar amounts as shown previously for rat, human, and chicken erythroid In the cytosol of HS reticulocytes, the synthesis of a-spectrin was identical to control while the synthesis of P-spectrin was increased about fourfold (Fig 1A). On the HS membrane, the two polypeptides were assembled stoichiometrically but their amounts were markedly reduced to about half the control values. We have obtained identical results by two indepen- dent methods: (1) direct quantitation of spectrin synthesis and assembly from the autoradiogram of SDS-PAGE of plasma membranes and cell lysates, and (2) spectrin immu- noprecipitation (data not shown). As suggested earlier in our previous report,"' the extent of spectrin synthesis in reticulocytes is dependent on the age of these cells as determined by their RNA contents. Early reticulocytes synthesize significant amounts of both a- and P-spectrins, while at the late reticulocyte stage spectrin synthesis is markedly diminished (data not shown). Therefore, early reticulocytes were used throughout in the present study for both the control and patient samples.
To ascertain that the synthesis of spectrin in reticulocytes is comparable with that in their nucleated RBC precursors, we have compared the synthesis and assembly of spectrin in nucleated RBC precursors and reticulocytes. The synthetic ratios of spectrin a- and @-subunits in reticulocytes and in their nucleated RBC precursors were similar, implying that reticulocytes represent a useful system to study the synthe- sis and assembly of spectrin in patients with hereditary hemolytic anemias associated with deficiencies of mem- brane proteins (data not shown).
The stability of the newly synthesized a- and P-spectrins in the membrane and cytosol was examined by a pulse- chase of control and HS reticulocytes labeled with (35S) methionine and chased in a medium containing unlabeled methionine. Both in control and HS reticulocytes (Fig lB), approximately equimolar proportions of labeled a- and P-spectrin entered the membrane fraction and reached a plateau by 1 hour following the commencement of the chase. As previously reported by us for the rat erythroid precursors," there was no detectable turnover of either control or HS spectrin in the membrane during the 3-hour chase period. However, in the soluble fraction of both
For personal use only.on April 13, 2019. by guest www.bloodjournal.orgFrom

168 HANSPAL ET AL
Lysates 120r
Membranes 6or
5 30 2 20
10
A 2o
, I l l
30 40 50 60 time(min)
1 20 , I l l 30 40 50 60
time(min1
Membranes Lysates r120
time (hr)
Fig 1. Representative synthesis, assembly, and turnover of spec- trin in HS and control reticulocytes. An equal number (2 x lo7) of age-matched HS (C.C) and control (N) reticulocytes were labeled with ("S) methionine for different time periods. For pulse-chase in (B), following a 15-minute labeling, the cells were incubated in a medium containing cold methionine for 0, 1, 2, and 3 hours. At the end of pulse-label or pulse-chase incubations, the cells were lysed to obtain their plasma membranes and cell lysates, which were then examined by SDS-PAGE. The autoradiograms generated from these gels were scanned at 570 nm and the area under each peak was integrated. (A) The synthesis of a-spectrin in the cytosol of HS reticulocytes is similar to control while the synthesis of p-spectrin is increased about fourfold. On the HS membrane, the amount of the two polypeptides is reduced to about half the control values. (B) In the membranes of both control and HS reticulocytes, there is no detectable turnover during the 3-hour chase period. In the cytosols, however, a- and P-spectrins are gradually turning over with similar half-lives in HS and control reticulocytes. These experiments were performed four times with similar results.
control and HS reticulocytes, the amount of labeling increased during the first hour after the onset of the chase and subsequently declined. The slopes of the curves for soluble HS and control a- and p-spectrins were identical, suggesting that they turn over with similar half-lives. The initial increase in radioactivity is probably due to the completion of the synthesis of polypeptides labeled during the pulse period.
Our find- ings of normal synthesis and turnover of spectrin in HS reticulocytes were in a striking contrast to a reduced assembly of spectrin on the HS RBC membrane. Therefore, we have asked whether or not the reduced assembly of spectrin on the HS membrane is caused by a defective synthesis of the spectrin-binding protein ankyrin. As in the studies of spectrin synthesis, an equal number of (35S) methionine-labeled reticulocytes matched for their RNA content were used, and their plasma membranes and cell lysates isolated as described in Materials and Methods. Ankyrin was immunoprecipitated from both the plasma
Synthesis, assembly, and turnover of ankyrin.
membranes and cell lysates, and the resulting immunopre- cipitates were analyzed on SDS-polyacrylamide gels (Fig 2A). In control reticulocytes, the amounts of ankyrin synthesized in the cytosol exceed those assembled on the membrane, as has also been shown in avian erythroid cells.48 In contrast, no intact ankyrin was detectable in the cytosol of HS reticulocytes and only immunoreactive degradation products (indicated by arrows in Fig 2A) were seen that were absent in the nonimmune serum controls (data not shown). While we were unable to detect newly synthesized intact ankyrin in the HS cytosol, some newly synthesized ankyrin was found in the membrane preparations of HS reticulocytes; however, it was reduced to about half the control values (Fig 2B).
To determine the turnover of the newly synthesized ankyrin, (35S) methionine-labeled control and HS reticulo- cytes were chased for various lengths of time in a medium containing unlabeled methionine. Ankyrin was then immu- noprecipitated from both the plasma membranes and cell lysates isolated from an equal number of cells for each time point. In membranes of both the control and HS reticulo- cytes (Fig 2C), the total amount of radioactivity increased for some time after the beginning of the chase and then reached a plateau that remained constant for up to 120 minutes of the chase, indicating that the membrane- associated ankyrin was stable. The turnover of membrane- associated HS ankyrin was the same as in the control samples. In contrast to membrane-associated ankyrin, the amount of radiolabeled ankyrin in the soluble fraction of control reticulocytes declines gradually, implying that the soluble ankyrin is gradually degraded. Because there was no detectable intact ankyrin in the HS reticulocyte cytosol, the pulse-chase could not be performed.
Quantitation of a-, p-spectrin, and ankyrin mRNAs in reticulocytes. We have previously shown that human retic- ulocytes have a low abundance of both a and p spectrin I ~ R N A . ~ ~ , " The relative abundance of the spectrin and ankyrin mRNAs in control and HS reticulocytes was quanti- tated by using a slot-blot analysis (Fig 3), and the amounts were expressed as a ratio to the amount in a,-globin mRNA used as an internal standard (Table 1). While the amount of a-spectrin mRNA was identical to the control sample, the amount of p-spectrin appeared slightly greater as judged by relative intensity of p-spectrin signal. This increase in p-spectrin mRNA was less apparent when the results were adjusted to the relative amounts of a,-globin mRNA, used as an internal standard for each sample. However, the difference was relatively minor, with the mean ratio of relative values of p-spectrin and a,-globin mRNA signal .705 as compared with control value of .64. This apparent mild increase of P-spectrin message of HS patients was not statistically significant with the P value of 305. In a striking contrast, the amount of ankyrin mRNA was markedly reduced as compared with a control sample. While the data presented in Fig 3A suggest that the propositus had less than 50% of ankyrin mRNA as compared with the control, the correction for the amount and purity of poly (A)' RNA by a,-globin internal standard showed that the deficiency of ankyrin mRNA is about 50% of normal (Table 1).
For personal use only.on April 13, 2019. by guest www.bloodjournal.orgFrom

ANKYRIN DEFICIENCY IN HEREDITARY SPHEROCYTOSIS 169
Fig 2. Representative synthe- sis, assembly, and turnover of ankyrin in HS and control reticu- locytes. (A) HS and control retic- ulocytes (3 x 10' each) were la- beled with (%) methionine for 30,60, and 120 minutes and their plasma membranes and lysates isolated. Ankyrin was immuno- precipitated from both the mem- branes and cell lysates using an- t ihuman ankyr in IgG and analyzed by SDS-PAGE. The gels were processed for fluorogra- phy. In the cytosol of HS reticulo- cytes, intact ankyrin is not detect- able, only immunoreactive degradation products (arrows) are detected. Some of the darker bands not labeled by arrows were seen with nonimmune se- rum in both control and patient samples. (6) The autoradiogram shown in (A) was scanned and the area under each peak inte- grated. The amount of newly syn- thesized ankyrin assembled on the membrane of HS reticulo- cytes is only about half the con- trol values. (C) Cells were la- beled with ('%) methionine for 30 minutes and then chased with cold methioninefor different time periods (0 to 120 minutes). There- after, the cells were lysed and their plasma membranes and ly- sates isolated. Ankyrin was im- munoprecipitated and analyzed as described above. The mem- brane-associated ankyrin is sta- ble in both HS and control reticu- locytes, while the soluble ankyrin in control reticulocytes is gradu- ally degraded. Note that in HS reticulocyte cytosol, no detect- able intact ankyrin is present. These experiments were per- formed three times with similar results.
Control (N) HS (C.C)
30' 60' 120' 30' 60' 120' 30' 60' 120' 30' 60' 120'
2.1-
I
---- Membranes Lysates Membranes Lysates
A
Membranes Lysates 2oor
4Or Membranes Lysates
(! 20 40 60 80 100 120 0 20 40 60 80 100 120 I 1 I 1 I I I I 1 I I I I
C time ( m i d time (min)
Northern blot analysis of reticulocyte poly (A)' RNA hybridized with a human ankyrin cDNA probe is shown in Fig 4. As previously shown,27.'x ankyrin mRNA was present as two distinct species of 7.2 and 9.0 kb. The amounts of both the species was dramatically reduced in the HS reticulocytes as compared to a control sample, consistent with our slot-blot analysis.
To more accurately quantitate the ankyrin message in
reticulocytes, the filter from Fig 4 was rehybridized with human %-globin cDNA probe. Because of the marked differences in abundance of ankyrin and human qglobin message in reticulocytes, the rehybridized membrane was exposed for only 30 minutes at room temperature, and the globin signal and the major ankyrin 7.2-kb signal were quantitated by laser densitometry. The ratio of globin to ankyrin signal was 10.5 as compared with the same measure-
For personal use only.on April 13, 2019. by guest www.bloodjournal.orgFrom
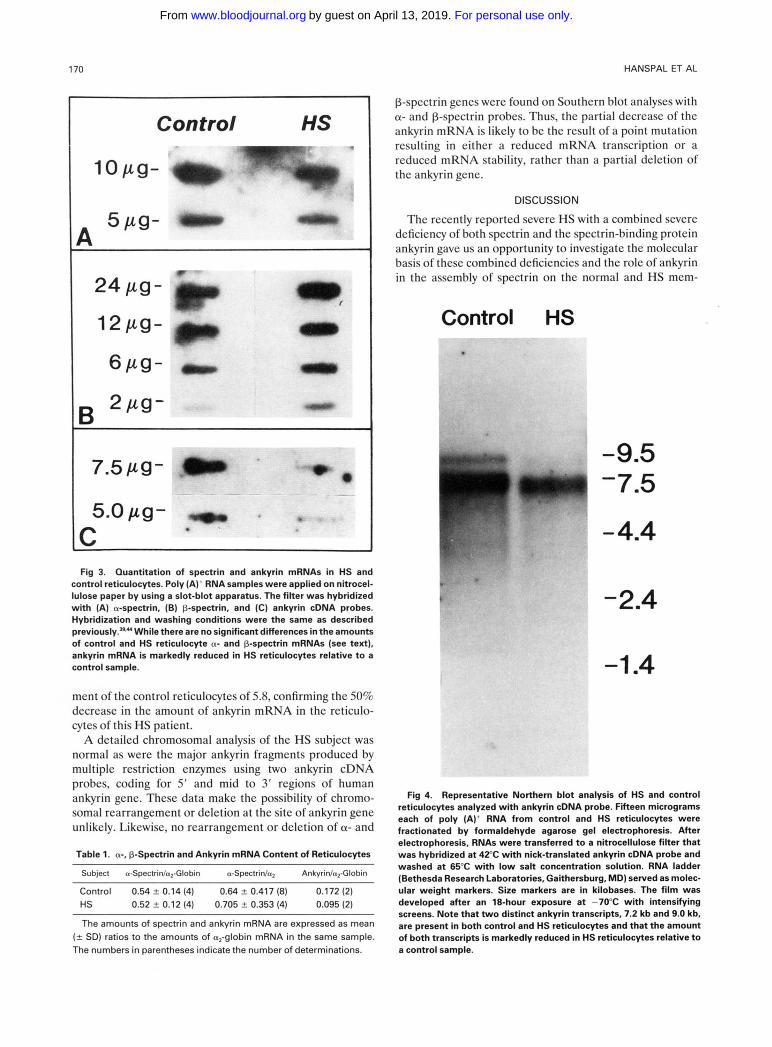
1 70 HANSPAL ET AL
Control HS
7 . 5 p g -
5.0 pg- C Fig 3. Ouantitation of spectrin and ankyrin mRNAs in HS and
control reticulocytes. Poly (A) * RNA samples were applied on nitrocel- lulose paper by using a slot-blot apparatus. The filter was hybridized with (A) a-spectrin, (B) p-spectrin, and (C) ankyrin cDNA probes. Hybridization and washing conditions were the same as described previously.=." While there are no significant differences in the amounts of control and HS reticulocyte a- and p-spectrin mRNAs (see text), ankyrin mRNA is markedly reduced in HS reticulocytes relative to a control sample.
ment of the control reticulocytes of 5.8, confirming the 50% decrease in the amount of ankyrin mRNA in the reticulo- cytes of this HS patient.
A detailed chromosomal analysis of the HS subject was normal as were the major ankyrin fragments produced by multiple restriction enzymes using two ankyrin cDNA probes, coding for 5' and mid to 3' regions of human ankyrin gene. These data make the possibility of chromo- somal rearrangement or deletion at the site of ankyrin gene unlikely. Likewise, no rearrangement or deletion of a- and
Table 1. a; P-Spectrin and Ankyrin mRNA Content of Reticulocytes
Subject a-Spectrinla,-Globin a-Spectrinla, Ankyrinla,Globin
Control 0.54 -+ 0.14 (4) 0.64 f 0.417 (8) 0.172 (2) HS 0.52 ? 0.12 (4) 0.705 f 0.353 (4) 0.095 (2)
The amounts of spectrin and ankyrin mRNA are expressed as mean ( 2 SD) ratios to the amounts of a,-globin mRNA in the same sample. The numbers in parentheses indicate the number of determinations.
P-spectrin genes were found on Southern blot analyses with a- and P-spectrin probes. Thus, the partial decrease of the ankyrin mRNA is likely to be the result of a point mutation resulting in either a reduced mRNA transcription or a reduced mRNA stability, rather than a partial deletion of the ankyrin gene.
DISCUSSION
The recently reported severe HS with a combined severe deficiency of both spectrin and the spectrin-binding protein ankyrin gave us an opportunity to investigate the molecular basis of these combined deficiencies and the role of ankyrin in the assembly of spectrin on the normal and HS mem-
Control HS
-9.5
- 4.4
-2.4
-1.4
Fig 4. Representative Northern blot analysis of HS and control reticulocytes analyzed with ankyrin cDNA probe. Fifteen micrograms each of poly (A). RNA from control and HS reticulocytes were fractionated by formaldehyde agarose gel electrophoresis. After electrophoresis, RNAs were transferred t o a nitrocellulose filter that was hybridized at 42°C with nick-translated ankyrin cDNA probe and washed at 65°C with low salt concentration solution. RNA ladder (Bethesda Research Laboratories, Gaithersburg, MD) sewed as molec- ular weight markers. Size markers are in kilobases. The film was developed after an 18-hour exposure at -70°C with intensifying screens. Note that two distinct ankyrin transcripts, 7.2 kb and 9.0 kb. are present in both control and HS reticulocytes and that the amount of both transcripts is markedly reduced in HS reticulocytes relative to a control sample.
For personal use only.on April 13, 2019. by guest www.bloodjournal.orgFrom

ANKYRIN DEFICIENCY IN HEREDITARY SPHEROCYTOSIS 171
brane. As previously rep~r ted ,~ ' the amounts of spectrin and ankyrin in the RBC membranes of the HS proband were about half the normal amounts. Furthermore, previ- ously reported structural and functional studies of the residual spectrin and ankyrin, including the binding of ankyrin to ankyrin-depleted inside-out vesicles, showed no abnormalities, raising a possibility that the underlying defect involves a reduced synthesis or increased turnover of either spectrin or ar~kyrin.~' Because the synthesis and assembly of spectrin a- and P-subunits in human reticulo- cytes and the nucleated RBC precursors are very similar, we have used the former system to evaluate the molecular basis of a combined spectrin and ankyrin deficiency in this patient.
As previously reported for nucleated RBC precursor^,'"^^' we have found that in control reticulocytes, a-spectrin is synthesized in excess of P-spectrin. We further found that in the HS reticulocytes the a-spectrin synthesis in the cytosol was normal while p-spectrin synthesis was increased about fourfold. In contrast, the incorporation of newly synthesized a- and P-spectrins into the HS membrane was reduced to about half the control values while the turnover of both cytosolic and membrane-associated spectrin was normal. Thus, despite normal (a-spectrin) or even in- creased (P-spectrin) synthesis, the assembly of the newly synthesized spectrin chains into the HS membrane was markedly reduced. The reason for the increased P-spectrin synthesis is likely related to a highly stimulated erythropoie- sis: this is suggested by recent data showing that in erythroblasts isolated from the spleens of mice infected with the anemia-inducing strain of Friend virus (FVA cells) induced with erythropoietin5' or murine erythroleukemia (MEL) cells induced with dimethyl sulfoxide (DMSO) ,52,53 the synthesis of p-spectrin is selectively increased. There- fore, it is possible that the HS reticulocytes represent a similar system of stress erythropoiesis with increased synthe- sis of p-spectrin. The finding of normal a- and increased P-spectrin synthesis is consistent with our findings of normal amounts of a-spectrin mRNA, although the in- crease of p-spectrin synthesis was more striking than the increase in mRNA level, presumably due to a more efficient translation process.
We further found that the marked decrease of the assembly of the newly synthesized spectrin on the mem- brane appears to be related to abnormalities in the synthe- sis of ankyrin. In control reticulocytes, the amounts of ankyrin synthesized in the cytosol are in excess of those assembled on the membrane (Fig 2) . In contrast, no intact ankyrin was detected in the cytosol of HS reticulocytes, and only immunoreactive ankyrin degradation products were present. Furthermore, the amounts of newly synthesized ankyrin which were stably assembled on the HS membrane were markedly reduced to about half the control value. Immunoprecipitations of other proteins like 4.1 or band 3
were not performed because of the limited amounts of samples available. However, plasma membranes derived from (35S) methionine-labeled HS and control reticulocytes, as examined by SDS-PAGE followed by autoradiography, showed that in HS reticulocyte membranes the incorpora- tion of all other newly synthesized proteins except spectrin and ankyrin was similar to control reticulocyte membrane, suggesting that the synthesis of other proteins was normal.
Our findings of reduced ankyrin synthesis were consis- tent with a marked decrease in ankyrin mRNA. The normal chromosomal studies as well as an absence of abnormalities on Southern blots using multiple restriction enzymes and two ankyrin probes excluded a deletion of a major segment of the ankyrin gene. Thus, it is likely, but remains to be proven, that the partial ankyrin mRNA deficiency is a result of a point mutation leading to a reduced expression or a reduced stability of the ankyrin mRNA.
Although the above data indicate that a deficiency of ankyrin mRNA and, consequently, a reduced synthesis of ankyrin constitutes the primary molecular defect, the exact inheritance pattern cannot be unequivocally established in this particular proband because of unavailability of his parents for study. However, several points suggest that the patient is either a homozygote or a compound heterozygote for a defect involving reduced synthesis of ankyrin: (1) All relatives of this patient were hematologically normal with the exception of the father whose anemia was nonhemolytic and likely acq~ired.~" ( 2 ) In our studies of ankyrin synthesis in normal reticulocytes, we detected large amounts of ankyrin synthesized in the cytosol with only a fraction of this newly synthesized ankyrin being assembled on the mem- brane. In contrast, there was no detectable ankyrin synthe- sis in the cytosol of the patient's reticulocytes, although some ankyrin (about 50% of normal) was found assembled on the membrane. (3) Furthermore, the deficiency of both spectrin and ankyrin in our proband as originally reported in reference 30 appears more profound than in the recently studied cases of HS associated with a deletion of the ankyrin gene in one of the two chromosome pairs.26
Our results further show that the decrease in spectrin assembly on the HS reticulocyte membrane is of a similar magnitude as the decrease in ankyrin incorporation into the membrane, suggesting that the assembly of spectrin on the membrane is limited by the amount of membrane- associated ankyrin. Hence, the underlying defect seems to be similar to that of the nb/nb spectrin-deficient mutant mice54s5 involving a reduced synthesis of ankyrin, which in part is also partially degraded before its attachment on the membrane. We conclude that the resulting partial defi- ciency of membrane-associated ankyrin leads to a second- ary deficiency of spectrin, that, in turn, leads to a weakened interaction of the membrane skeleton with the lipid bilayer and integral membrane proteins, facilitating loss of surface area and spherocyte f ~ r m a t i o n . ~ ~
REFERENCES 1. Becker PS, Lux SE: Hereditary spherocytosis and related
2. Shen BW, Josephs R, Steck T L Ultrastructure of unit
fragments of the skeleton of the human erythrocyte membrane. J Cell Biol99:810, 1984
3. Byers TJ, Branton D: Visualization of the protein associations disorders. Clin Haematol14:15,1985
For personal use only.on April 13, 2019. by guest www.bloodjournal.orgFrom

172 HANSPAL ET AL
in the erythrocyte membrane skeleton. Proc Natl Acad Sci USA 82:6153,1985
4. Liu SC, Derick LH, Palek J: Visualization of the hexagonal lattice in the erythrocyte membrane skeleton. J Cell Biol 104527, 1987
5. Mische SM, Mooseker MS, Morrow JS: Erythrocyte adducin: A calmodulin-regulated actin bundling protein that stimulates spectrin-actin binding. J Cell Biol105:2837, 1987
6. Bennett V, Stenbuck P: Identification and partial purification of ankyrin, the high affinity membrane attachment site for human erythrocyte spectrin. J Biol Chem 254:2533,1979
7. Bennett V, Stenbuck P: Association between ankyrin and the cytoplasmic domain of band 3 isolated from the human erythrocyte membrane. J Biol Chem 255:6424,1980
8. Anderson RA, Lovrien R E Glycophorin is linked by band 4.1 protein to the human erythrocyte membrane skeleton. Nature 307:655,1984
9. Pasternack GR, Anderson RA, Leto TL, Marchesi VT: Interactions between protein 4.1 and band 3. An alternative binding site for an element of the membrane skeleton. J Biol Chem 260:3676,1985
10. Hanspal M, Palek J: Synthesis and assembly of membrane skeletal proteins in mammalian red cell precursors. J Cell Biol 105:1417,1987
11. Blikstad I, Nelson NJ, Moon RT, Lazarides E: Synthesis and assembly of spectrin during avian erythropoiesis: Stoichiometric assembly but unequal synthesis of a and p spectrin. Cell 32:1081, 1983
12. Cox JV, Stack JH, Lazarides E: Erythroid anion transporter assembly is mediated by a developmentally regulated recruitment onto a preassembled membrane cytoskeleton. J Cell Biol 105:1405, 1987
13. Agre P, Casella JF, Zinkham WH, McMillan C, Bennett V: Partial deficiency of erythrocyte spectrin in hereditary spherocyto- sis. Nature 314:380,1985
14. Winkelman JC, Marchesi SL, Watkins P, Linnenbach AJ, Agre P, Forget B: Recessive hereditary spherocytosis is associated with an abnormal a spectrin subunit. Clinical Research 34:474a, 1986 (abstr)
15. Marchesi SL, Agre PA, Spiecher DW, Tse WT, Forget BG: Mutant spectrin a I1 domain in recessively inherited spherocytosis. Blood 74:182a, 1989 (abstr)
16. Marchesi S L The erythrocyte cytoskeleton in hereditary elliptocytosis and spherocytosis, in Agre P, Parker JC (eds): Red Blood Cell Membranes: Structure, Function, Clinical Implications, ch 4. New York, NY, Dekker, 1989, p 77
17. Becker PS, Morrow JS, Lux SE: Abnormal oxidant sensitiv- ity and p-chain structure of spectrin in hereditary spherocytosis associated with defective spectrin-protein 4.1 binding. J Clin Invest 80:557,1987
18. Goodman SR, Shiffer KA, Casoria LA, Eyster ME: Identifi- cation of the molecular defect in the erythrocyte membrane skeleton of some kindreds with hereditary spherocytosis. Blood 60:772, 1982
19. Wolfe LC, John KM, Falcone JC, Byrne AM, Lux SE: A genetic defect in the binding of protein 4.1 to spectrin in a kindred with hereditary spherocytosis. N Engl J Med 307:1367,1982
20. Winkelmann J, Marchesi SL, Gillespie F, Agre P, Forget B: Dominant hereditary spherocytosis is not closely linked to the gene for a spectrin in three kindreds. Blood 68:42a, 1986 (abstr)
21. Forget BG, Chang JG, Coupal E, Stanislovitis P, Costa FF, Winkelmann JC, Agre PC, Marchesi VT, Watkins P C Molecular genetics of the human p-spectrin gene. Trans Assoc Am Phys C1:149,1988
22. Kimberling WJ, Fulbeck T, Dixon L, Lubs HA: Localization
of spherocytosis to chromosome 8 or 12 and report of a family with spherocytosis and a reciprocal translocation. Am J Human Genet 27586, 1975
23. Kimberling WJ, Taylor RA, Chapman RG, Lubs H A Linkage and gene localization of hereditary spherocytosis (HS). Blood 52859,1978
24. Bass EB, Smith SW, Stevenson RE, Rose W F Further evidence of the location of the spherocytosis gene on chromosome 8. Ann Int Med 99:192,1983
25. Chilcote RR, LeBeau MM, Dampier C, Pergament E, Verlinsky Y, Mohandas N, Frischer H, Rowley JD: Association of red cell spherocytosis with deletion of the short arm of chromo- some 8. Blood 69:156,1987
26. Lux SE, Tse WT, Menninger JC, John KM, Harris P, Shalev 0, Chilcote RR, Marchesi SL, Watkins PC, Bennett V, McIntosh S, Collins FS, Francke U, Ward DC, Forget BG: Hereditary spherocytosis associated with deletion of human erythrocyte ankyrin gene on chromosome 8. Nature 345:736,1990
27. Lambert S, Yu H, Prchal JT, Lawler J, Ruff P, Speicher D, Cheung MC, Kan YW, Palek J: The cDNA sequence for human erythrocyte ankyrin. Proc Natl Acad Sci USA 87:1730,1990
28. Lux SE, John KM, Bennett V: Analysis of cDNA of human erythrocyte ankyrin indicates a repeated structure with homology to tissue differentiation and cell cycle control proteins. Nature 344:36,1990
29. Costa FF, Lux SE, Agre P, Watkins P, John K, Forget BG: Linkage of dominant hereditary spherocytosis to the gene for the erythrocyte membrane-skeleton protein ankyrin. N Engl J Med 323:1046,1990
30. Coetzer TL, Lawler J, Liu SC, Prchal JT, Gualtieri RJ, Brain MC, Dacie JV, Palek J: Partial ankyrin and spectrin deficiency in severe atypical hereditary spherocytosis. N Engl J Med 318:230, 1988
31. Shatkin AJ: Colorimetric reactions for DNA, RNA, and protein determinations, in Habel K, Salzman NP (eds): Fundamen- tal Techniques in Virology. San Diego, CA, Academic, 1969, p 231
32. Chan L-NL: Changes in the composition of plasma mem- brane proteins during differentiation of embryonic chick erythroid cell. Proc Natl Acad Sci USA 74:1062, 1977
33. Marcantonio EE, Hynes RO: Antibodies to the conserved cytoplasmic domain of the integrin beta 1 subunit react with proteins in vertebrates, invertebrates, and fungi. J Cell Biol 106:1765,1988
34. Fairbanks G, Steck TL, Wallach DFM: Electrophoretic analysis of the major polypeptides of the human erythrocyte membrane. Biochemistry 10:2606,1971
35. Laemmli U K Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 27:680,1970
36. Goossens M, Kan YW: DNA analysis in the diagnosis of hemoglobin disorder. Methods Enzymol76:805,1981
37. Chomczynski P, Sacchi N: Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform ex- traction. Anal Biochem 162:156, 1987
38. Maniatis T, Fritsch EF, Sambrook J: Molecular cloning: A laboratory manual. Cold Spring Harbor, NY, Cold Spring Harbor Laboratory, 1982
39. Prchal JT, Morley BJ, Yoon SH, Coetzer TL, Palek J, Conboy JG, Kan YW: Isolation and characterization of cDNA clones for human erythrocyte p-spectrin. Proc Natl Acad Sci USA 84:7468,1987
40. Curtis PJ, Palumbo A, Ming J, Fraser P, Cioe L, Meo P, Shane S, Rovera J: Sequence comparison of human and murine erythrocyte a-spectrin cDNA. Gene 36:357,1985
41. Erlich HA, Gelfand DH, Saiki R K Specific DNA amplifica- tion. Nature 331:461,1988
For personal use only.on April 13, 2019. by guest www.bloodjournal.orgFrom

ANKYRIN DEFICIENCY IN HEREDITARY SPHEROCYTOSIS 173
42. Saiki RK, Gelfand DH, Stoffel S, Scharf SJ, Higuchi R, Horn GT, Mullis KB, Erlich HA: Primer-directed enzymatic amplifica- tion of DNA with a thermostable DNA polymerase. Science 239:487,1988
43. Liebhaber S, Cash FE: Locus assignment of a-globin struc- tural mutations by hybrid-selected translation. J Clin Invest 75:64, 1985
44. Thomas PS: Hybridization of denatured RNA and small DNA fragments transferred to nitrocellulose. Proc Natl Acad Sci USA 77:5201,1980
45. Hozier JC, Lindquist LL Banded karyotypes from bone marrow: A clinical useful approach. Hum Genet 53:205,1980
46. Yoon S-H, Kentros CG, Prchal J T Identification of an unusual deletion within homologous repeats of human reticulocyte p-spectrin and probable peptide polymorphism. Gene 91:297,1990
47. Hanspal M, Dainiak N, Palek J: Synthesis and assembly of spectrin in human erythroblasts. Blood 68:35a, 1986 (abstr)
48. Moon RT, Lazarides E: Biogenesis of the avian erythroid membrane skeleton: Receptor-mediated assembly and stabiliza- tion of ankyrin (Goblin) and spectrin. J Cell Biol98:1899,1984
49. Prchal JT, Papayannopoulou T, Yoon S-H Patterns of spectrin transcripts in erythroid and non-erythroid cells. J Cell Physiol144:287,1990
50. Yoon S-H, Papayannopoulou T, Prchal JT: Characterization and quantitation of p-spectrin mRNA in erythroid precursors. Blood 72333a, 1988 (abstr)
51. Hanspal M, Kalraiya R, Hanspal J, Palek J: Effect of erythropoietin on the OJp spectrin synthetic ratio in Friend virus infected murine erythroblasts. Blood 7461a, 1989 (abstr)
52. Lehnert ME, Lodish HF: Unequal synthesis and differential degradation of alpha and beta spectrin during murine erythroid differentiation. J Cell Biol107:413,1988
53. Pfeffer SR, Huima T, Redman CM: Biosynthesis of spectrin and its assembly into the cytoskeletal system of Friend erythroleu- kemia cells. J Cell Bioi 103:103, 1986
54. Bodine DM IV, Birkenmieier CS, Barker JE: Spectrin deficient inherited hemolytic anemias in the mouse: Characteriza- tion by spectrin synthesis and mRNA activity in reticulocytes. Cell 37:721,1984
55. White RA, Biokenmeier CS, Lux SE, Barker JE: Ankyrin and the hemolytic anemia mutation, nb, map to mouse chromo- some 8: Presence of the nb allele is associated with a truncated erythrocyte ankyrin. Proc Natl Acad Sci USA 87:3117,1990
56. Palek J: Hereditary elliptocytosis, spherocytosis and related disorders. Blood Rev 1:147,1987
For personal use only.on April 13, 2019. by guest www.bloodjournal.orgFrom

1991 77: 165-173
M Hanspal, SH Yoon, H Yu, JS Hanspal, S Lambert, J Palek and JT Prchal ankyrinhereditary spherocytosis: evidence implicating a primary defect of Molecular basis of spectrin and ankyrin deficiencies in severe
http://www.bloodjournal.org/content/77/1/165.full.htmlUpdated information and services can be found at:
Articles on similar topics can be found in the following Blood collections
http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://www.bloodjournal.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://www.bloodjournal.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.Society of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American
For personal use only.on April 13, 2019. by guest www.bloodjournal.orgFrom


















