
The Effectiveness of a Mnemonic-Type Startle and Surprise ...
Upload
laura-fermanCategory
view
218download
0description

1 23
Brain Structure and Function ISSN 1863-2653 Brain Struct FunctDOI 10.1007/s00429-013-0585-8
A fast cholinergic modulation of theprimary acoustic startle circuit in rats
Ricardo Gómez-Nieto, Donal G. Sinex,José de Anchieta C. Horta-Júnior,Orlando Castellano, Javier M. Herrero-Turrión, et al.

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag Berlin Heidelberg. This e-offprint is
for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

ORIGINAL ARTICLE
A fast cholinergic modulation of the primary acoustic startlecircuit in rats
Ricardo Gomez-Nieto • Donal G. Sinex •
Jose de Anchieta C. Horta-Junior • Orlando Castellano •
Javier M. Herrero-Turrion • Dolores E. Lopez
Received: 15 February 2013 / Accepted: 16 May 2013
� Springer-Verlag Berlin Heidelberg 2013
Abstract Cochlear root neurons (CRNs) are the first
brainstem neurons which initiate and participate in the full
expression of the acoustic startle reflex. Although it has
been suggested that a cholinergic pathway from the ventral
nucleus of the trapezoid body (VNTB) conveys auditory
prepulses to the CRNs, the neuronal origin of the VNTB–
CRNs projection and the role it may play in the cochlear
root nucleus remain uncertain. To determine the VNTB
neuronal type which projects to CRNs, we performed tract-
tracing experiments combined with mechanical lesions,
and morphometric analyses. Our results indicate that a
subpopulation of non-olivocochlear neurons projects
directly and bilaterally to CRNs via the trapezoid body. We
also performed a gene expression analysis of muscarinic
and nicotinic receptors which indicates that CRNs contain
a cholinergic receptor profile sufficient to mediate the
modulation of CRN responses. Consequently, we investi-
gated the effects of auditory prepulses on the neuronal
activity of CRNs using extracellular recordings in vivo.
Our results show that CRN responses are strongly inhibited
by auditory prepulses. Unlike other neurons of the cochlear
nucleus, the CRNs exhibited inhibition that depended on
parameters of the auditory prepulse such as intensity and
interstimulus interval, showing their strongest inhibition at
short interstimulus intervals. In sum, our study supports the
idea that CRNs are involved in the auditory prepulse
inhibition of the acoustic startle reflex, and confirms the
existence of multiple cholinergic pathways that modulate
the primary acoustic startle circuit.
Keywords Biotinylated dextran amine � Cochlear root
neurons � Extracellular recordings � Olivocochlear bundle �Prepulse inhibition � Ventral nucleus of the trapezoid body
Introduction
In rodents, cochlear root neurons (CRNs) have large cell
bodies and thick dendrites oriented parallel or perpendic-
ular to the eighth nerve fibers among which they are dis-
tributed (Merchan et al. 1988; Lopez et al. 1993). They are
recipients of numerous cholinergic inputs from the ventral
nucleus of the trapezoid body (VNTB) (Gomez-Nieto et al.
2008a), converging glutamatergic inputs from auditory
nerve fibers (Harrison et al. 1962; Merchan et al. 1988;
Gomez-Nieto et al. 2008b), noradrenergic inputs from
the locus coeruleus (Gomez-Nieto et al. 2008b), as well
as GABA and glycinergic inputs of unknown origin
Electronic supplementary material The online version of thisarticle (doi:10.1007/s00429-013-0585-8) contains supplementarymaterial, which is available to authorized users.
R. Gomez-Nieto � J. A. C. Horta-Junior � O. Castellano �J. M. Herrero-Turrion � D. E. Lopez
Neuroscience Institute of Castilla y Leon (INCyL), University of
Salamanca, 37007 Salamanca, Spain
R. Gomez-Nieto � O. Castellano � D. E. Lopez (&)
Department of Cell Biology and Pathology, Medical School,
University of Salamanca, 37007 Salamanca, Spain
e-mail: [email protected]
R. Gomez-Nieto � O. Castellano � D. E. Lopez
Institute of Biomedical Research of Salamanca (IBSAL),
University of Salamanca, 37007 Salamanca, Spain
D. G. Sinex
Departments of Psychology and Biology, Utah State University,
Logan, UT 84322-2810, USA
J. A. C. Horta-Junior
Department of Anatomy, Biosciences Institute, Sao Paulo State
University (UNESP), Botucatu, Sao Paulo 18618-970, Brazil
123
Brain Struct Funct
DOI 10.1007/s00429-013-0585-8
Author's personal copy

(Osen et al. 1991; Gomez-Nieto et al. 2008b). The primary
neuronal circuit underlying the mediation of acoustic
startle reflex encompasses three central synaptic relay sta-
tions: CRNs, giant PnC neurons, and cranial and spinal
motoneurons (Lingenhohl and Friauf 1994; Lee et al. 1996;
Lopez et al. 1999; Nodal and Lopez 2003). Thus, CRNs are
the first brainstem neurons which initiate and participate in
the full expression of the acoustic startle reflex (Lopez
et al. 1999; Horta-Junior et al. 2008). CRNs contain all the
necessary neurotransmitter receptors to provide short
latency acoustic input to neurons in the caudal pontine
reticular nucleus (PnC); mediating alert and escape
behaviors elicited by a sudden and intense acoustic stim-
ulus (Lee et al. 1996; Lopez et al. 1999; Sinex et al. 2001;
Gomez-Nieto et al. 2008b). The acoustic startle response
can be reduced when a strong acoustic startling stimulus
(pulse) is shortly preceded by a weaker sound (prepulse), a
paradigm called auditory prepulse inhibition (PPI). Inves-
tigation of PPI mediating pathways is critical for estab-
lishing new animal models for studying both cognitive
features and neural bases of some neuropsychiatric disor-
ders, which are characterized by PPI deficits (Swerdlow
and Geyer 1998; Swerdlow et al. 2001; Li et al. 2009;
Molina et al. 2009). It is widely accepted that the inferior
colliculus (IC) relays auditory information to the primary
acoustic startle circuit at the level of the PnC (Leitner and
Cohen 1985; Li et al. 1998a, b; Fendt et al. 2001; Yeomans
et al. 2006), through a slow multimodal pathway which
serially connects the IC, the superior colliculus, and the
inhibitory cholinergic projection from the pedunculopon-
tine tegmental nucleus to the PnC (Koch et al. 1993; Fendt
and Koch 1999; Fendt et al. 2001; Li et al. 2009). Never-
theless, this neuronal circuit does not explain all of the
singularities of PPI such as the effectiveness of interstim-
ulus intervals as short as 20 ms (Hoffman and Ison 1980).
This leads us to presume the existence of another short
auditory projection to the primary acoustic startle circuit.
Previous studies suggested that the descending auditory
pathway IC–VNTB–CRNs conveys auditory prepulses at
the level of the cochlear root nucleus to mediate the
auditory PPI (Gomez-Nieto et al. 2008a). However, the
VNTB neuronal type which projects to the CRNs is not
known and no electrophysiological data is available to
support the inhibition of CRNs by auditory prepulses. This
study focuses on determining the neuronal type that is the
source of the cholinergic projection to the cochlear root
nucleus. In agreement with our morphological findings
(current study, Gomez-Nieto et al. 2008a, b), we also show
a gene expression analysis of cholinergic receptors and
electrophysiological data which supports that CRNs par-
ticipate in the auditory PPI of the acoustic startle reflex via
the VNTB–CRNs projection.
Materials and methods
Experimental animals
In total, 21 adult female Sprague–Dawley rats (Charles
River Laboratories) weighing 290–320 g were used in this
study. The experiments were conducted according to the
guidelines for the use and care of laboratory animals of the
European Communities Council Directive (DOCE L 222;
24-08-1999) and with those established by the United
States (NIH publication No. 80–23). All efforts were made
to minimize the number of animals used and their suffer-
ing. For the surgical procedures, the animals were deeply
anesthetized with a mixture of ketamine (40 mg/kg body
weight) and xylazine (7 mg/kg body weight), and main-
tained in this state by supplementary doses, as required,
throughout the duration of the experiment.
Neuroanatomical experiments: surgery, tissue
processing and image analysis
A total of 9 animals were used to study the source of the
VNTB inputs on the CRNs. The rats received unilaterally
injections of the bidirectional tracer, biotinylated dextran
amine (BDA, 10,000 MW; #D-1956; Molecular Probes,
Eugene, OR) into brain structures that are well known to
contain axonal tracts of VNTB neurons (bundles) or its
efferent innervations (Warr and Beck 1996). Specifically,
the tracer experiments included BDA injections into the
following areas: the VNTB, the medial olivocochlear
bundle (OCB), the ventral cochlear nucleus (VCN), and the
trapezoid body (TB). All surgical and stereotaxic proce-
dures for injecting the tracer were identical to that used in
our previous studies (Gomez-Nieto et al. 2008a, b; Horta-
Junior et al. 2008). BDA (10 % in distilled water) was
injected iontophoretically via a glass micropipette (25 lm
tip diameter), with 3 lA positive current pulses (7 s on/7 s
off) for a period of 10 min. The coordinates for the left
VNTB were precisely the same as those devised by
Gomez-Nieto et al. (2008a). The coordinates for the medial
OCB, the TB and the VCN were obtained from the atlas of
the rat brain (Paxinos and Watson 1998), using an electrode
angle calibrator (David Kopf Instruments). In another set of
experiments, two animals were injected with BDA into the
VNTB followed by a mechanical lesion of the OCB. The
lesion was made with a surgical needle which was aimed
with the stereotaxic apparatus to target the medial OCB.
The lesion was attempted contralateral to the injection site,
and placed off of midline at a point where olivocochlear
fibers travel. The needle was moved several times along the
rostro-caudal plane to ensure the surgical transection of
medial olivocochlear fibers.
Brain Struct Funct
123
Author's personal copy

Tissue preparation for histology including perfusion of
the animals, Nissl staining, visualization of the BDA neu-
rotracer, and calbindin protein-D28 K (CaBP) immuno-
histochemistry was identical to those used in our previous
studies (Lopez et al. 1993, 1999; Gomez-Nieto et al.
2008a). Nickel-intensified peroxidase reaction was devel-
oped for the BDA visualization to distinguish from the
CaBP immunohistochemistry (Hancock 1982; Gomez-Ni-
eto et al. 2008a). Cochleae of animals with BDA injections
were also processed for the tracer visualization and were
used to analyze the distribution of medial olivocochlear
fibers in the cochlea, as in our previous study (Gomez-
Nieto et al. 2008a).
All sections were examined on an upright brightfield
microscope (#BX5; Olympus, Center Valley, PA, USA)
equipped with a digital camera (SpotRt�; Diagnostic
Instruments, Sterling Heights, MI, USA). Low-magnifica-
tion images were taken with the 49, 109 or 209 objective
lens, and high magnification images were taken with a 409
or 1009 objective lens (oil immersion) for morphometric
analysis of BDA-labeled structures.
The morphometric analysis of labeled structures was
carried out with ImageJ (version 1.42; Rasband, N.S.,
National Institutes of Health, Bethesda, Maryland, USA;
http://rsb.info.nih.gov/ij). Photomicrographs shown in the
figures were processed with minor modifications in
brightness, contrast and to remove the tissue-free back-
ground using Adobe Photoshop� (version 9.0; Adobe
Systems Incorporated, San Jose, CA, USA) and assembled
in Canvas 7.0 software.
Isolation of RNA and RT-PCR
The cochlear nerve roots of 8 rats were collected bilaterally
to study the gene expression of acetylcholine receptors,
particularly the muscarinic (M1-5) and nicotinic (Na4,
Na7, Nb2 and Nb3) receptors subunits. Prefrontal cortex
(PFC) and hippocampus (H) were also sampled to serve as
positive controls. Isolation of RNA and reverse transcrip-
tion-polymerase chain reaction (RT-PCR) were then per-
formed by standard procedures as described elsewhere
(Gomez-Nieto et al. 2008b). Total RNA was immediately
isolated from the individual tissue samples using the
TRIZOL� reagent (Gibco BRL, Gaithersburg, MD, USA)
in accordance with the manufacturer’s procedure. The
quantity of purified RNA was assessed by spectropho-
tometry Nano Photometer (Implen GmbH) and the integ-
rity subsequently confirmed with RNA 6000 Nano
LabChip (Agilent Technologies, Palo Alto, CA, USA),
obtaining the RNA integrity number (RIN, 0 corresponding
to fully degraded RNA and 10 corresponding to intact
RNA). For all PCRs, only RNA samples with RIN of at
least 7.5 were used, with the vast majority of samples
having a RIN of at least 8.0.
Total RNA (2 lg), primed with oligo-dT, was reverse-
transcribed into complementary DNA (cDNA) at 37 �C for
2 h using the first-strand cDNA synthesis kit (Promega
Corporation, Madison, WI, USA) in a 20 ll volume and
stored at -20 �C until use, according to manufacturer’s
instructions. In all cases, a reverse transcriptase negative
control was used for testing genomic DNA contamination.
Also, DNase treatment (Turbo DNA-free Kit, Applied
Biosystems) was performed at 37 �C for 2 h to remove
DNA contamination, followed by inactivation of the
DNases at 75 �C for 15 min.
A 25 ll PCR mixture contained 100 ng of cDNA tem-
plate, 20 pmol of each primer, 0.2 mM dNTPs, 1.5 mM
MgCl2 and 5 units of GoTaq Flexi DNA polymerase
(Promega Corp.). Supplemental figure 1 lists primers spe-
cific for the genes examined in the present study. PCR
reactions were performed in an ABI7000 PCR system
(Applied Biosystems, Europe). Amplification of rat b-actin
(GenBank accession no. NM_031144) was used as an
internal and loading control. PCRs amplification were as
follows: 1 cycle at 95 �C for 5 min as an initial denatur-
ation step, denaturation at 95 �C for 30 s, annealing at
57–59 �C for 30 s, and extension at 72 �C for 45 s (30
cycles), followed by further incubation for 10 min at 72 �C
(1 cycle). PCR conditions were shown to be at the linear
phase of amplification to assess a semiquantitative analysis.
PCR products were electrophoresed on 2.5 % agarose gels
in 1 9 40 mM Tris–acetate, 1 mM ethylenediamine tetra-
acetic acid pH 8.0, and visualized by ethidium bromide
staining.
The primers were designed in such a way that RT-PCR
products spanned two identified introns. Moreover, the
amplification of b-actin was performed with equal amounts
of RNA and demonstrated an identical expression pattern
in all cerebral regions used. Finally, in all RT-PCRs, an
RNA-free (negative) control sample was used which did
not produce any amplified bands.
Electrophysiology experiments: acoustic stimulation,
data collection and analysis
Four rats were used for the extracellular recording of CRNs
neuronal activity after auditory prepulse stimulation. The
rats were mounted in a stereotaxic apparatus using hollow
ear bars coupled to an earphone (#1310B, Radio Shack
Super Tweeter, Korea) for stimuli delivery. The recording
microelectrode was aimed to pass through the rat’s
cochlear root nucleus, using identical coordinates and
surgical procedures to those used in our previous studies
(Lopez et al. 1999; Sinex et al. 2001; Nodal and Lopez
Brain Struct Funct
123
Author's personal copy

2003; Gomez-Nieto et al. 2008a; Horta-Junior et al. 2008).
Acoustic stimulation was performed using almost identical
devices and method to those used by Sinex et al. (2001).
The frequency response of the acoustic system was mea-
sured in a coupler with a calibrated microphone (#4136,
B&K, GA, USA). This standard calibration curve was used
to set the levels of tones in all experiments. To record the
neuronal activity, we used carbon fiber electrodes (#E1011,
Carbostar-1, Kation Scientific, MN, USA) with impedance
at 1 kHz of 0.4–0.8 MX and tip length of 15 lm. The
electrode was fixed to the stereotaxic frame by a Kopf
carrier, positioned as described by Lopez et al. (1999), and
then advanced using a microdrive (Trent-Wells). After that,
we followed the procedure described by Sinex et al. (2001)
to isolate and identify single units. Single units were
identified by the constant shape and magnitude of the
spikes, which were monitored throughout the experiment.
Up to seven electrode passes with slight variations in the
point of entry were needed to encounter single units in the
cochlear root nucleus. As a control, auditory brainstem
evoked potentials were recorded prior to and after each
experiment to check the status of nerve conduction in the
auditory nerve during the experiment (Supplemental
Fig. 2). Changes in wave amplitude of auditory brainstem
evoked potentials were taken into account when analyzing
the data to assess possible auditory nerve damage due to
multiple electrodes passes.
Stimulus presentation paradigms and data collection
were controlled by a PC-compatible computer and custom
software written for MATLAB (MathWorks, Natick MA,
USA). Stimulus waveforms were digitally synthesized
online. Search stimuli were tone bursts, and records were
obtained only from well-isolated single units. At the
beginning of the experiment, characteristic frequency and
threshold were measured from the frequency tuning curves
which were acquired with an automated procedure that
estimated threshold at many closely spaced frequencies.
Frequency response maps (combination of frequencies and
intensities capable of evoking a response) were obtained
automatically using a randomized paradigm that presented
a set of frequency/intensity pairings specifically chosen to
define the complete details of a unit’s frequency selectivity.
The response map consisted of a set of spike counts
obtained at 6 sound pressure levels (SPLs) and 63 fre-
quencies. After the collection of tuning curve and response
maps, pure tones were used as experimental stimuli to
perform more detailed measurements of unit responses to
assess the modifications by prepulse presentation at dif-
ferent conditions (duration, intensity and interstimulus
intervals).
In the probes designed to test the auditory prepulse
inhibition, two consecutive pure tones of the characteristic
frequency (CF) for the units investigated were presented
ipsilaterally to the recording site. The spikes times evoked
by the different stimuli were stored and were used to cal-
culate the response magnitude (spikes per trial) and first-
spike latencies. In each probe, we obtained peristimulus
time (PST) histograms and dot rasters to display the
responses to 50–100 repetitions of the tones. Intensity,
duration and interstimulus intervals (ISI) of the pulse and
prepulse tones were shown in each PST histogram. The bin
size for the PST histograms was 1.4 ms, and was reduced
to 0.3 ms in the expanded views.
Results
VNTB efferent pathways toward the cochlear root
nucleus
To determine the course and termination of descending
axons from the ventral nucleus of the trapezoid body
(VNTB) to the cochlear root nucleus, we injected BDA
into the rostral end of the VNTB. BDA injections into the
VNTB were small (0.3–0.6 mm in diameter), round in
shape, and were located in the rostral end of the nucleus
without spreading to adjacent superior olivary nuclei
(Fig. 1a). Following BDA injections in the VNTB, we
found two axonal pathways projecting toward the cochlear
root nucleus (Fig. 1). The first pathway consisted of thin
fibers coursing bilaterally through the trapezoid body
(TB). These BDA-labeled fibers have a diameter of
approximately 2 lm and projected to the cochlear root
nucleus of both sides. VNTB axons in the TB were clearly
distinguishable from thicker axons (6–8 lm in diameter)
of CRNs which were immunostained for CaBP (Fig. 1b).
The second track of VNTB axons was identified as the
medial olivocochlear bundle (OCB). We observed that
smooth-labeled fibers (*1.3 lm in diameter) projected
bilaterally from the injection site in the left VNTB. The
contralateral labeled fibers cross beneath the floor of the
fourth ventricle (Fig. 1c), running through the cochlear
nerve to project predominantly to the contralateral cochlea
and terminate on the outer hair cells (Fig. 1d). As
described in our previous study (Gomez-Nieto et al.
2008a), we found that VNTB neurons massively innervate
CRNs of both sides (Fig. 1e, f). To visualize the mor-
phology of the CRNs including cell bodies, dendrites and
axons, we immunostained brainstem sections for CaBP.
CRNs showed strong positive immunolabeling for CaBP
(Fig. 1e, f) as previously described by Lopez et al. (1993).
BDA-labeled axons from the VNTB enter along the
cochlear root nucleus, where they terminate in numerous
endings onto the cell bodies and dendrites of CRNs in a
bead-like pattern (Fig. 1e). We can infer from these
results that two possible populations of VNTB neurons
Brain Struct Funct
123
Author's personal copy
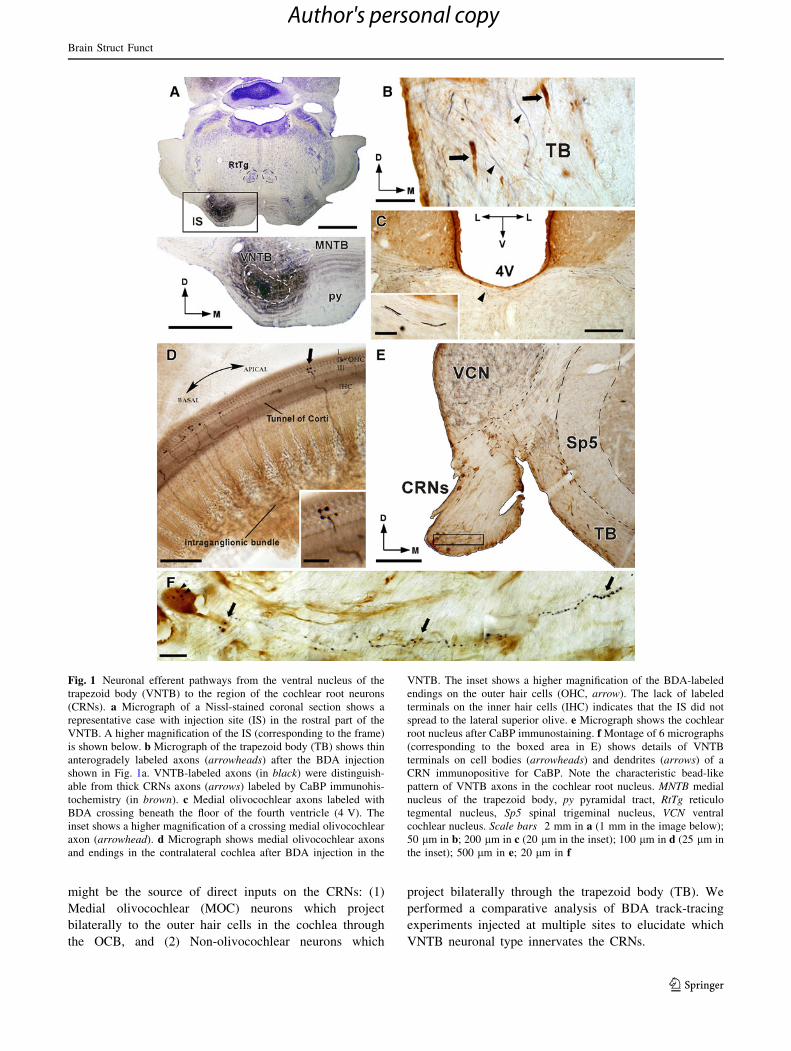
might be the source of direct inputs on the CRNs: (1)
Medial olivocochlear (MOC) neurons which project
bilaterally to the outer hair cells in the cochlea through
the OCB, and (2) Non-olivocochlear neurons which
project bilaterally through the trapezoid body (TB). We
performed a comparative analysis of BDA track-tracing
experiments injected at multiple sites to elucidate which
VNTB neuronal type innervates the CRNs.
Fig. 1 Neuronal efferent pathways from the ventral nucleus of the
trapezoid body (VNTB) to the region of the cochlear root neurons
(CRNs). a Micrograph of a Nissl-stained coronal section shows a
representative case with injection site (IS) in the rostral part of the
VNTB. A higher magnification of the IS (corresponding to the frame)
is shown below. b Micrograph of the trapezoid body (TB) shows thin
anterogradely labeled axons (arrowheads) after the BDA injection
shown in Fig. 1a. VNTB-labeled axons (in black) were distinguish-
able from thick CRNs axons (arrows) labeled by CaBP immunohis-
tochemistry (in brown). c Medial olivocochlear axons labeled with
BDA crossing beneath the floor of the fourth ventricle (4 V). The
inset shows a higher magnification of a crossing medial olivocochlear
axon (arrowhead). d Micrograph shows medial olivocochlear axons
and endings in the contralateral cochlea after BDA injection in the
VNTB. The inset shows a higher magnification of the BDA-labeled
endings on the outer hair cells (OHC, arrow). The lack of labeled
terminals on the inner hair cells (IHC) indicates that the IS did not
spread to the lateral superior olive. e Micrograph shows the cochlear
root nucleus after CaBP immunostaining. f Montage of 6 micrographs
(corresponding to the boxed area in E) shows details of VNTB
terminals on cell bodies (arrowheads) and dendrites (arrows) of a
CRN immunopositive for CaBP. Note the characteristic bead-like
pattern of VNTB axons in the cochlear root nucleus. MNTB medial
nucleus of the trapezoid body, py pyramidal tract, RtTg reticulo
tegmental nucleus, Sp5 spinal trigeminal nucleus, VCN ventral
cochlear nucleus. Scale bars 2 mm in a (1 mm in the image below);
50 lm in b; 200 lm in c (20 lm in the inset); 100 lm in d (25 lm in
the inset); 500 lm in e; 20 lm in f
Brain Struct Funct
123
Author's personal copy

VNTB neurons projecting to CRNs do not send axon
collaterals into the VCN
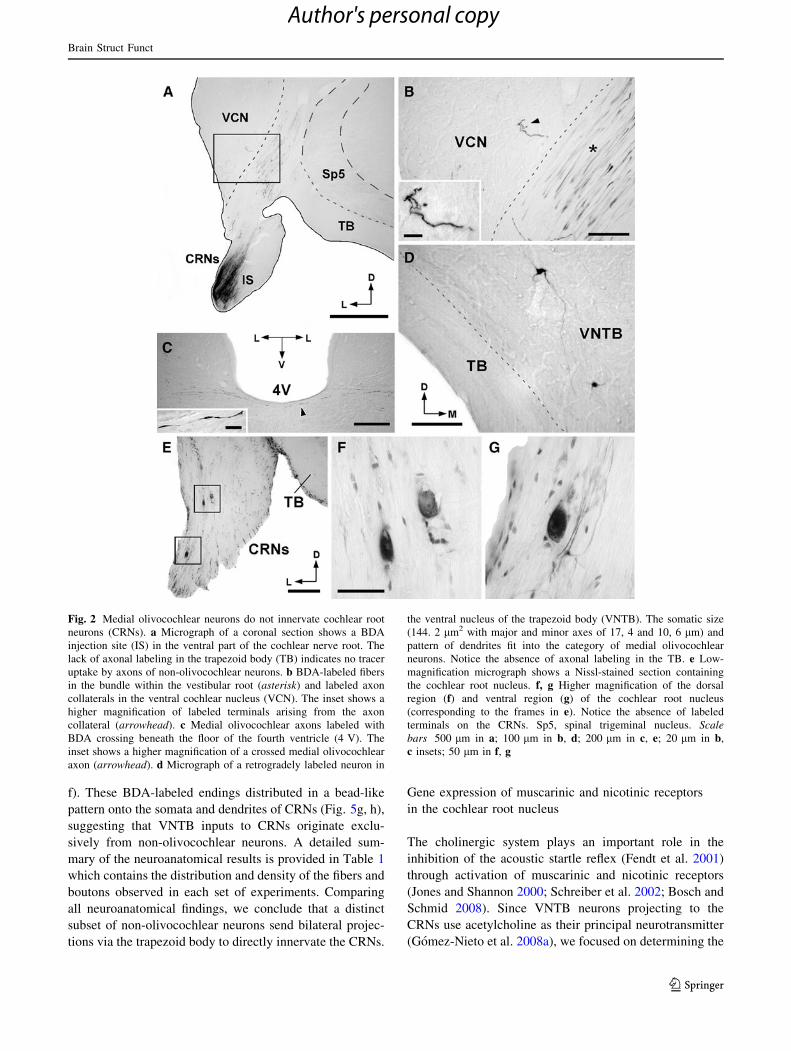
To determine whether MOC neurons innervate the CRNs,
two animals received BDA injections into the medial OCB
at two different locations. In the first animal, the injection
had a small, elongated shape (0.2–0.3 mm in diameter) and
was located deep in the cochlear nerve just where medial
OCB fibers run into the periphery toward the cochlea
(Fig. 2a). BDA-labeled fibers were located in a bundle
within the vestibular root and axon collaterals entered the
ventral cochlear nucleus (VCN, Fig. 2b). OCB fibers
labeled with BDA were also observed crossing the mid-line
at the floor of the fourth ventricle (Fig. 2c). The tracer
uptake by these axons resulted in retrograde labeling of
MOC neurons (e. g. in Fig. 2d). The non-olivocochlear
efferent pathway of the VNTB was not filled with tracer as
we did not observe any labeled structure in the TB (Fig. 2a,
d, e). In the second animal, a small injection (0.3 mm in
diameter) was made, without spreading to the adjacent
VCN, in the course of OCB axons just before they enter the
cochlear nerve root. As in the first case, we found the
typical distribution of medial OCB fibers (see below
Fig. 5a–c). In both cases with BDA injections into the
OCB, there was an absence of labeled boutons on the
CRNs (Fig. 2e–g). Thus, there is a strong likelihood that
the MOC neurons do not innervate the CRNs.
In addition to the MOC projection, VNTB non-olivo-
cochlear neurons project to almost the entire rostrocaudal
extent of the VCN by way of the trapezoid body of both
sides (Warr and Beck 1996). To determine if the beaded
endings on the CRNs from the VNTB (Fig. 1e) arise as
terminal branches from axons of these neurons, we injected
BDA into the left VCN. The BDA injection site was round
(0.4 mm in diameter) and included both posteroventral and
anteroventral areas of the VCN (Fig. 3a). Due to the
bidirectional nature of the transport of the BDA, we found
anterograde labeling in the form of a thin band of axons
and swellings in the ipsilateral lateral superior olive as well
as retrogradely labeled neurons in the VNTB (Fig. 3b;
Supplemental Fig. 3). Neurons that were retrogradely
labeled in the VNTB varied in size and dendritic pattern
(Fig. 3b, d) suggesting that both medial olivocochlear and
non-olivocochlear neurons send axonal terminals into the
VCN. Accordingly, we observed labeled axons in the TB
(Supplemental Fig. 3) and in the medial OCB (Fig. 3c). In
the ipsilateral cochlear root nucleus, our material showed
large synaptic terminals (4–7 lm in diameter) that pre-
sumably correspond to collaterals of auditory primary
afferents which followed a straight course toward the
center of the VCN (Fig. 3e, f). These large terminals were
entirely different in size and distribution to those present in
the cochlear root nucleus after BDA injection in the
VNTB. We did not find the typical en passant and bead-
like pattern of VNTB axons in the cochlear root nucleus
(Fig. 3e, f; Supplemental Fig. 3), suggesting that VNTB
inputs onto the CRNs do not arise from VNTB neurons that
also target the VCN.
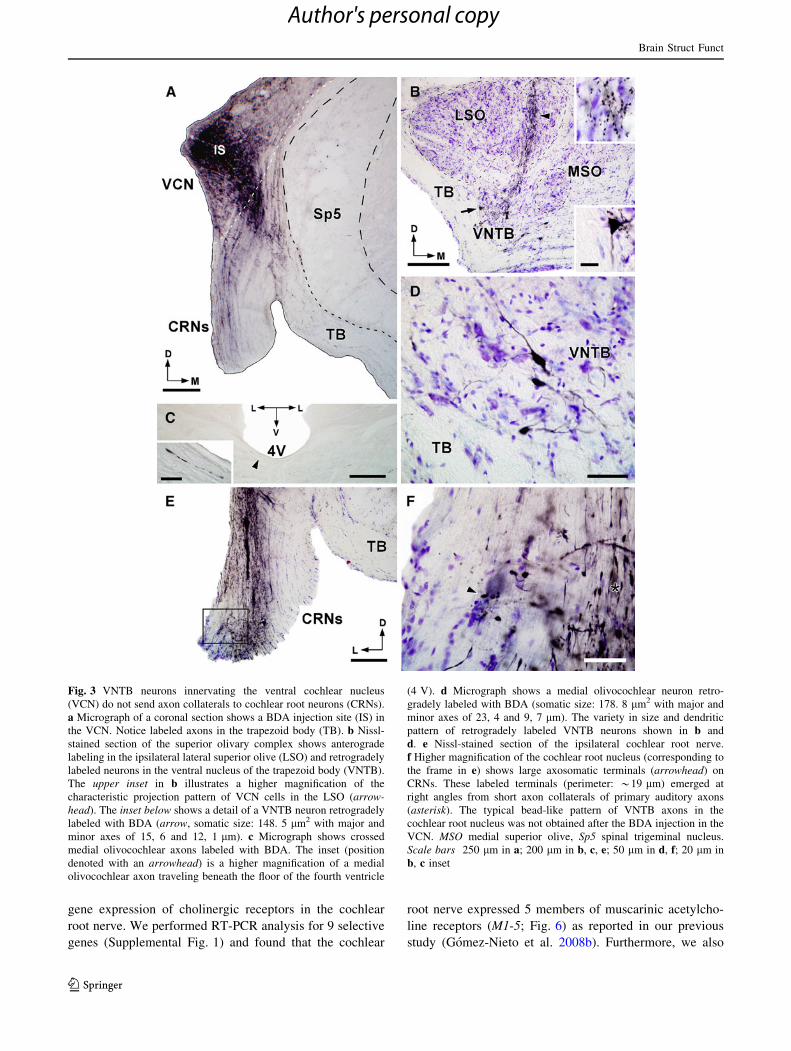
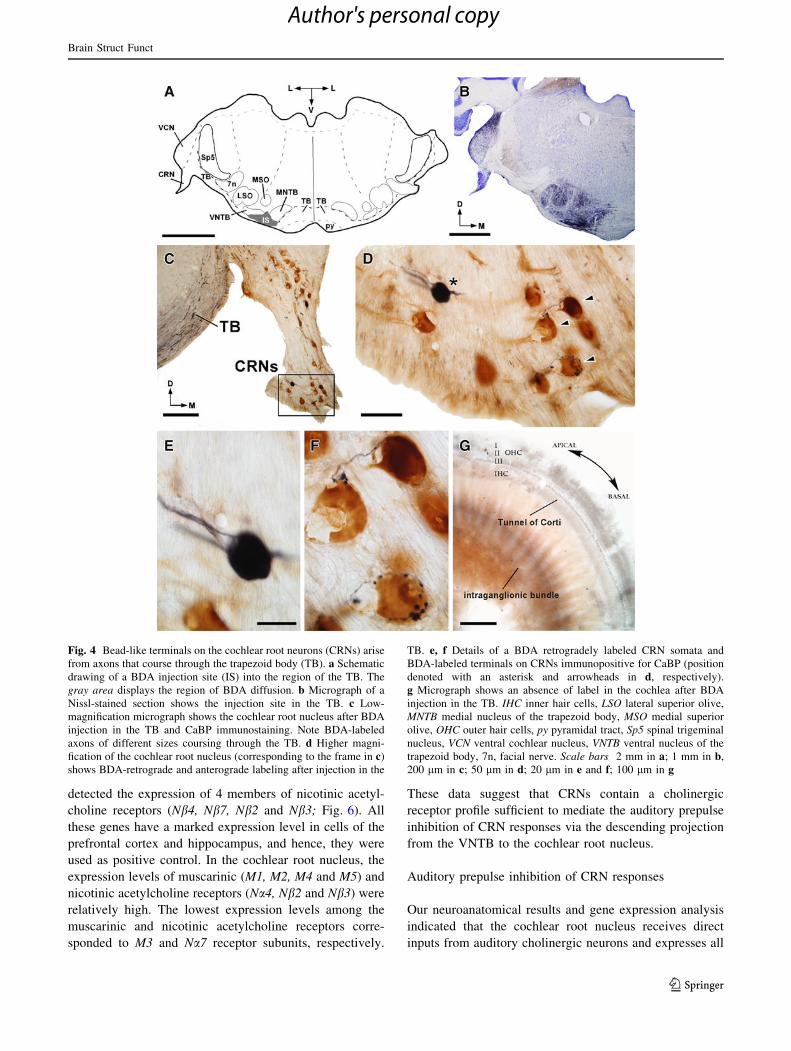
Non-olivocochlear neurons send direct projections
to CRNs via the trapezoid body
The results described above suggest that a subpopulation of
non-olivocochlear cells are the source of VNTB inputs on
the CRNs. To verify the pathway that VNTB axons follow
before entering the cochlear root nucleus, we injected BDA
into the TB. The injection sites varied in size (0.4–1 mm in
diameter) and were confined to the TB in an area which
was beneath and in close proximity to the superior olivary
complex (Fig. 4a, b). As expected, we observed BDA-
labeled axons in the TB (Fig. 4c; Supplemental Fig. 4).
Many of these BDA-labeled fibers were thick and resem-
bled those axons of CRNs immunostained for CaBP,
whereas other axons were thinner with the same diameter
as those observed in the TB after BDA injections in the
VNTB. We followed the directions of both types of axons
and found that they turn ventrally to enter the cochlear root
nucleus. The thicker axons emerged from retrogradely
labeled CRNs somata (Fig. 4d, e; Supplemental Fig. 4),
and the thinner fibers terminate as numerous endings onto
the cell bodies and dendrites of CRNs in a bead-like pattern
(Fig. 4d, f; Supplemental Fig. 4). Thus, VNTB axons
innervating the CRNs followed the same course as CRNs
axons, running through the TB. As in cases with BDA
injections in the VNTB, the labeled en passant boutons
distributed along the entire course of the fibers from the
dorsal to the most ventral divisions of the cochlear root
nucleus (Supplemental Fig. 4). In this set of experiments,
we did not encounter any BDA-labeled structures in the
medial OCB or terminals on the outer hair cells of the
cochlea (Fig. 4g).
To fully confirm the source of VNTB inputs to CRNs, a
mechanical lesion of the OCB was made in the rostro-
caudal plane after injecting BDA in the left VNTB. To
assess the effectiveness of the lesion, the cochleae were
analyzed histologically to detect any BDA labeling. Fig-
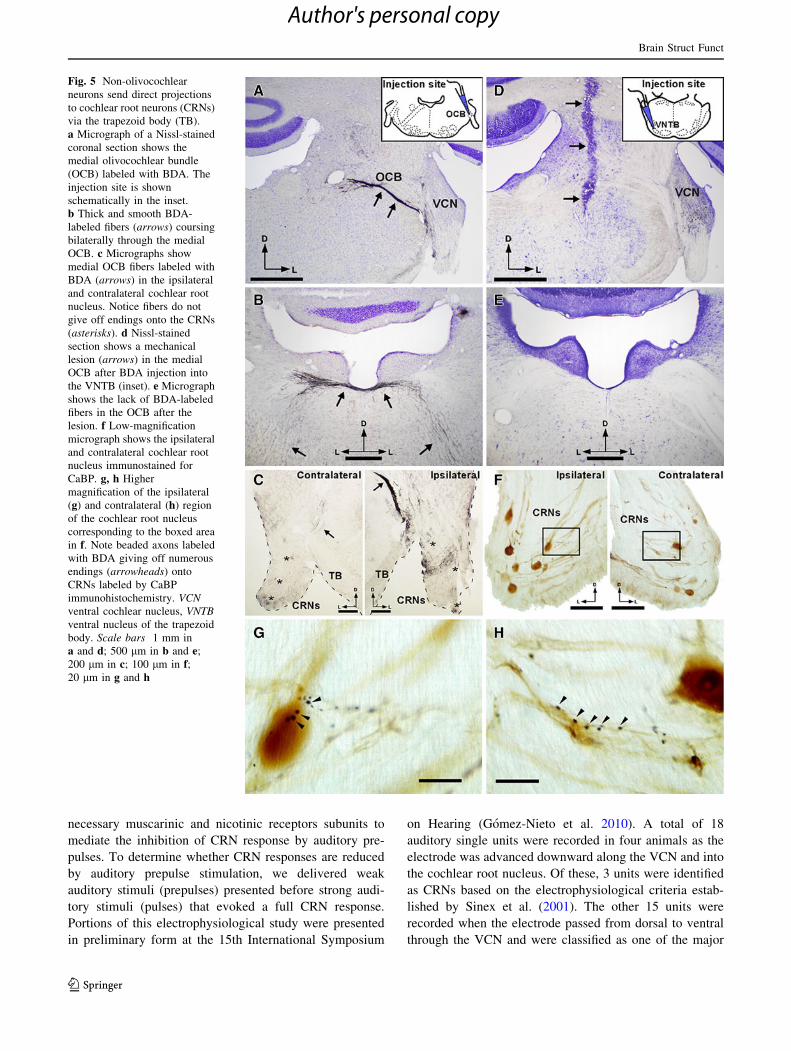
ure 5 illustrates an animal with BDA injection in the OCB
(Fig. 5a–c) and a representative case with OCB transection
after BDA injection in the VNTB (Fig. 5d–h). In the
lesioned cases, the cut was placed off-midline and con-
tralaterally to the injection site (Fig. 5d). As compared to
the animal with OCB labeling, the lesioned cases presented
a lack of BDA-labeled axons in the crossed OCB (Fig. 5b,
e). We did not find boutons on CRNs after OCB labeling,
however, the lesioned animals showed VNTB-labeled ax-
ons that give off boutons on CRNs of both sides (Fig. 5c,
Brain Struct Funct
123
Author's personal copy
f). These BDA-labeled endings distributed in a bead-like
pattern onto the somata and dendrites of CRNs (Fig. 5g, h),
suggesting that VNTB inputs to CRNs originate exclu-
sively from non-olivocochlear neurons. A detailed sum-
mary of the neuroanatomical results is provided in Table 1
which contains the distribution and density of the fibers and
boutons observed in each set of experiments. Comparing
all neuroanatomical findings, we conclude that a distinct
subset of non-olivocochlear neurons send bilateral projec-
tions via the trapezoid body to directly innervate the CRNs.
Gene expression of muscarinic and nicotinic receptors
in the cochlear root nucleus
The cholinergic system plays an important role in the
inhibition of the acoustic startle reflex (Fendt et al. 2001)
through activation of muscarinic and nicotinic receptors
(Jones and Shannon 2000; Schreiber et al. 2002; Bosch and
Schmid 2008). Since VNTB neurons projecting to the
CRNs use acetylcholine as their principal neurotransmitter
(Gomez-Nieto et al. 2008a), we focused on determining the
Fig. 2 Medial olivocochlear neurons do not innervate cochlear root
neurons (CRNs). a Micrograph of a coronal section shows a BDA
injection site (IS) in the ventral part of the cochlear nerve root. The
lack of axonal labeling in the trapezoid body (TB) indicates no tracer
uptake by axons of non-olivocochlear neurons. b BDA-labeled fibers
in the bundle within the vestibular root (asterisk) and labeled axon
collaterals in the ventral cochlear nucleus (VCN). The inset shows a
higher magnification of labeled terminals arising from the axon
collateral (arrowhead). c Medial olivocochlear axons labeled with
BDA crossing beneath the floor of the fourth ventricle (4 V). The
inset shows a higher magnification of a crossed medial olivocochlear
axon (arrowhead). d Micrograph of a retrogradely labeled neuron in
the ventral nucleus of the trapezoid body (VNTB). The somatic size
(144. 2 lm2 with major and minor axes of 17, 4 and 10, 6 lm) and
pattern of dendrites fit into the category of medial olivocochlear
neurons. Notice the absence of axonal labeling in the TB. e Low-
magnification micrograph shows a Nissl-stained section containing
the cochlear root nucleus. f, g Higher magnification of the dorsal
region (f) and ventral region (g) of the cochlear root nucleus
(corresponding to the frames in e). Notice the absence of labeled
terminals on the CRNs. Sp5, spinal trigeminal nucleus. Scalebars 500 lm in a; 100 lm in b, d; 200 lm in c, e; 20 lm in b,
c insets; 50 lm in f, g
Brain Struct Funct
123
Author's personal copy
gene expression of cholinergic receptors in the cochlear
root nerve. We performed RT-PCR analysis for 9 selective
genes (Supplemental Fig. 1) and found that the cochlear
root nerve expressed 5 members of muscarinic acetylcho-
line receptors (M1-5; Fig. 6) as reported in our previous
study (Gomez-Nieto et al. 2008b). Furthermore, we also
Fig. 3 VNTB neurons innervating the ventral cochlear nucleus
(VCN) do not send axon collaterals to cochlear root neurons (CRNs).
a Micrograph of a coronal section shows a BDA injection site (IS) in
the VCN. Notice labeled axons in the trapezoid body (TB). b Nissl-
stained section of the superior olivary complex shows anterograde
labeling in the ipsilateral lateral superior olive (LSO) and retrogradely
labeled neurons in the ventral nucleus of the trapezoid body (VNTB).
The upper inset in b illustrates a higher magnification of the
characteristic projection pattern of VCN cells in the LSO (arrow-head). The inset below shows a detail of a VNTB neuron retrogradely
labeled with BDA (arrow, somatic size: 148. 5 lm2 with major and
minor axes of 15, 6 and 12, 1 lm). c Micrograph shows crossed
medial olivocochlear axons labeled with BDA. The inset (position
denoted with an arrowhead) is a higher magnification of a medial
olivocochlear axon traveling beneath the floor of the fourth ventricle
(4 V). d Micrograph shows a medial olivocochlear neuron retro-
gradely labeled with BDA (somatic size: 178. 8 lm2 with major and
minor axes of 23, 4 and 9, 7 lm). The variety in size and dendritic
pattern of retrogradely labeled VNTB neurons shown in b and
d. e Nissl-stained section of the ipsilateral cochlear root nerve.
f Higher magnification of the cochlear root nucleus (corresponding to
the frame in e) shows large axosomatic terminals (arrowhead) on
CRNs. These labeled terminals (perimeter: *19 lm) emerged at
right angles from short axon collaterals of primary auditory axons
(asterisk). The typical bead-like pattern of VNTB axons in the
cochlear root nucleus was not obtained after the BDA injection in the
VCN. MSO medial superior olive, Sp5 spinal trigeminal nucleus.
Scale bars 250 lm in a; 200 lm in b, c, e; 50 lm in d, f; 20 lm in
b, c inset
Brain Struct Funct
123
Author's personal copy
detected the expression of 4 members of nicotinic acetyl-
choline receptors (Nb4, Nb7, Nb2 and Nb3; Fig. 6). All
these genes have a marked expression level in cells of the
prefrontal cortex and hippocampus, and hence, they were
used as positive control. In the cochlear root nucleus, the
expression levels of muscarinic (M1, M2, M4 and M5) and
nicotinic acetylcholine receptors (Na4, Nb2 and Nb3) were
relatively high. The lowest expression levels among the
muscarinic and nicotinic acetylcholine receptors corre-
sponded to M3 and Na7 receptor subunits, respectively.
These data suggest that CRNs contain a cholinergic
receptor profile sufficient to mediate the auditory prepulse
inhibition of CRN responses via the descending projection
from the VNTB to the cochlear root nucleus.
Auditory prepulse inhibition of CRN responses
Our neuroanatomical results and gene expression analysis
indicated that the cochlear root nucleus receives direct
inputs from auditory cholinergic neurons and expresses all
Fig. 4 Bead-like terminals on the cochlear root neurons (CRNs) arise
from axons that course through the trapezoid body (TB). a Schematic
drawing of a BDA injection site (IS) into the region of the TB. The
gray area displays the region of BDA diffusion. b Micrograph of a
Nissl-stained section shows the injection site in the TB. c Low-
magnification micrograph shows the cochlear root nucleus after BDA
injection in the TB and CaBP immunostaining. Note BDA-labeled
axons of different sizes coursing through the TB. d Higher magni-
fication of the cochlear root nucleus (corresponding to the frame in c)
shows BDA-retrograde and anterograde labeling after injection in the
TB. e, f Details of a BDA retrogradely labeled CRN somata and
BDA-labeled terminals on CRNs immunopositive for CaBP (position
denoted with an asterisk and arrowheads in d, respectively).
g Micrograph shows an absence of label in the cochlea after BDA
injection in the TB. IHC inner hair cells, LSO lateral superior olive,
MNTB medial nucleus of the trapezoid body, MSO medial superior
olive, OHC outer hair cells, py pyramidal tract, Sp5 spinal trigeminal
nucleus, VCN ventral cochlear nucleus, VNTB ventral nucleus of the
trapezoid body, 7n, facial nerve. Scale bars 2 mm in a; 1 mm in b,
200 lm in c; 50 lm in d; 20 lm in e and f; 100 lm in g
Brain Struct Funct
123
Author's personal copy
necessary muscarinic and nicotinic receptors subunits to
mediate the inhibition of CRN response by auditory pre-
pulses. To determine whether CRN responses are reduced
by auditory prepulse stimulation, we delivered weak
auditory stimuli (prepulses) presented before strong audi-
tory stimuli (pulses) that evoked a full CRN response.
Portions of this electrophysiological study were presented
in preliminary form at the 15th International Symposium
on Hearing (Gomez-Nieto et al. 2010). A total of 18
auditory single units were recorded in four animals as the
electrode was advanced downward along the VCN and into
the cochlear root nucleus. Of these, 3 units were identified
as CRNs based on the electrophysiological criteria estab-
lished by Sinex et al. (2001). The other 15 units were
recorded when the electrode passed from dorsal to ventral
through the VCN and were classified as one of the major
Fig. 5 Non-olivocochlear
neurons send direct projections
to cochlear root neurons (CRNs)
via the trapezoid body (TB).
a Micrograph of a Nissl-stained
coronal section shows the
medial olivocochlear bundle
(OCB) labeled with BDA. The
injection site is shown
schematically in the inset.
b Thick and smooth BDA-
labeled fibers (arrows) coursing
bilaterally through the medial
OCB. c Micrographs show
medial OCB fibers labeled with
BDA (arrows) in the ipsilateral
and contralateral cochlear root
nucleus. Notice fibers do not
give off endings onto the CRNs
(asterisks). d Nissl-stained
section shows a mechanical
lesion (arrows) in the medial
OCB after BDA injection into
the VNTB (inset). e Micrograph
shows the lack of BDA-labeled
fibers in the OCB after the
lesion. f Low-magnification
micrograph shows the ipsilateral
and contralateral cochlear root
nucleus immunostained for
CaBP. g, h Higher
magnification of the ipsilateral
(g) and contralateral (h) region
of the cochlear root nucleus
corresponding to the boxed area
in f. Note beaded axons labeled
with BDA giving off numerous
endings (arrowheads) onto
CRNs labeled by CaBP
immunohistochemistry. VCNventral cochlear nucleus, VNTBventral nucleus of the trapezoid
body. Scale bars 1 mm in
a and d; 500 lm in b and e;
200 lm in c; 100 lm in f;20 lm in g and h
Brain Struct Funct
123
Author's personal copy

cell types of the VCN (primary-like, chopper, pauser, and
on-sustained neurons) according to their temporal
response patterns in peristimulus time (PST) histograms.
CRNs exhibited a secure response with first-spike laten-
cies of approximately 2.2 ms (Fig. 7a, b). Figure 7b shows
a representative example of spike waveforms from an
isolated single unit. The characteristic frequencies of the
recorded CRNs were about 30 kHz, and the tuning curves
were sharp with a threshold of 10 dB sound pressure level
(SPL) (Fig. 7c, e, f). In the same animal, we found that
first-spike latencies and characteristic frequencies of
CRNs deviated markedly from units in the overlying
portions of the VCN (Fig. 7c–f). After electrophysiologi-
cal identification of the isolated units, we tested the effects
of auditory prepulses. The frequency of the prepulse and
the main pulse always matched the characteristic fre-
quency of the recorded unit, and the time between both
stimuli (interstimulus intervals: ISIs) was varied. In all
cases, the responses of CRNs were strongly inhibited by
auditory prepulse stimuli (Fig. 8). We systematically
observed a manifest decrease in the first-spike discharge
which yielded a reduction in the CRN overall response.
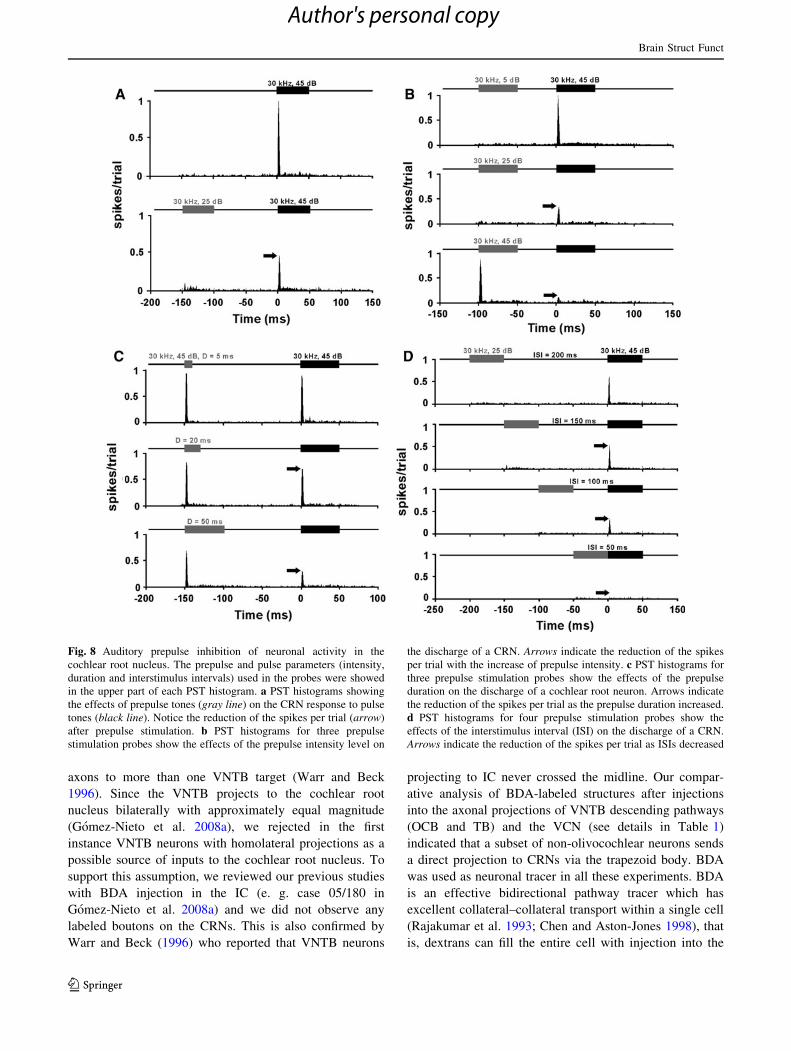
For example, the response of the CRN shown in Fig. 8a
was drastically reduced by prepulses which were presented
at 150 ms of ISI and with an intensity of just 15 dB SPL
over the threshold. Although this unit responded weakly
with a few spike discharges to the onset of the prepulse,
the response to the pulse was decreased by 50 %. This
result highlighted the very strong influence of the prepulse
on the CRN’s response. In a different probe, we tested the
effect of the prepulse intensity by increasing the level of
the prepulse in steps of 20 dB up to a maximum of the
pulse intensity (Fig. 8b). We kept prepulse duration
Ta
ble
1S
um
mar
yta
ble
of
BD
A-l
abel
edst
ruct
ure
so
bse
rved
inea
chex
per
imen
tse
t
VN
TB
retr
og
rad
ely
lab
eled
neu
ron
s
Lab
eled
fib
ers
inth
e
OC
B
Lab
eled
bo
uto
ns
on
the
ou
ter
hai
rce
lls
Lab
eled
fib
ers
inth
eT
BB
ead
edla
bel
edb
ou
ton
so
n
the
CR
Ns
Ipsi
late
ral
Co
ntr
alat
eral
Ipsi
late
ral
Co
ntr
alat
eral
Ipsi
late
ral
Co
ntr
alat
eral
Ipsi
late
ral
Co
ntr
alat
eral
Ipsi
late
ral
Co
ntr
alat
eral
BD
Ain
ject
ion
into
the
VN
TB
??
-?
??
??
??
??
??
??
?
BD
Ain
ject
ion
into
the
OC
B1
11
11
11
11
--
--
BD
Ain
ject
ion
into
the
VC
N1
11
11
11
11
11
1-
-
BD
Ain
ject
ion
into
the
TB
11
11
--
--
11
11
11
11
BD
Ain
ject
ion
into
the
VN
TB
?O
CB
lesi
on
11
-1
1-
11
-1
11
11
11
1
Th
ed
ata
issh
ow
nas
pre
sen
ce(?
)o
rab
sen
ce(-
)o
fB
DA
-lab
eled
fib
ers
and
bo
uto
ns
inea
chb
rain
area
.D
ensi
tyo
ffi
ber
san
db
ou
ton
sw
asev
alu
ated
qu
alit
ativ
ely
intw
og
rad
es(?
?an
d?
)
wh
enco
mp
ared
the
ipsi
late
ral
wit
hth
eco
ntr
alat
eral
sid
eo
fth
ein
ject
ion
site
.T
he
anal
ysi
so
fth
eco
ntr
alat
eral
OC
Bin
clu
des
the
area
un
der
the
fourt
hv
entr
icle
(4V
)
BD
Ab
ioti
ny
late
dd
extr
anam
ine,
OC
Bo
liv
oco
chle
arb
un
dle
,V
NT
Bv
entr
aln
ucl
eus
of
the
trap
ezo
idb
od
y,
TB
trap
ezo
idb
od
y
Fig. 6 Different gene expression levels of muscarinic acetylcholine
receptors (M1-5) and nicotinic acetylcholine receptors (Na4, Na7,Nb2 and Nb3) in the cochlear root nerve (CRN), prefrontal cortex
(PFC), hippocampus (H) and negative control (C-) in the exponential
phase of different PCRs (30 cycles). Primer sequences, sizes of the
distinct fragments and conditions of PCRs are detailed in supple-
mental figure 1. b-actin was used as control for equal amounts in RT-
PCR reactions
Brain Struct Funct
123
Author's personal copy

(50 ms) and interstimulus interval (100 ms) fixed during
this probe. As the prepulse intensity was increased, we
observed a reduction of the CRN response to the onset of
the pulse (Fig. 8b). The spike discharges to the pulse were
maximally inhibited (nearly 90 %) with a prepulse deliv-
ered at the same intensity as the pulse (45 dB SPL). In the
probe to test the effect of the prepulse duration, we
increased the durations of the prepulse while an intensity of
45 dB SPL and an ISI of 150 ms were kept fixed (Fig. 8c).
The loss of spikes to the onset of the pulse was produced by
increasing durations of the prepulse. The strongest inhibi-
tory effects (80 %) were observed with a prepulse duration
equal to the pulse duration (50 ms). We also examined the
effect of the ISI by decreasing the time between the pre-
pulse and the pulse (Fig. 8d). In this probe, the pulse that
evoked a full CRN response was preceded by a low-
intensity prepulse (25 dB SPL). It is shown in PST histo-
grams of Fig. 8d that CRN responses were significantly
reduced as the ISI decreased, and spikes to the onset of the
pulse were totally abolished when the prepulse was deliv-
ered just before the pulse (50 ms of ISI). Thus, the
responses of CRNs were dependent on the prepulse
intensity, the prepulse duration and the interstimulus
intervals.
CRNs differ from VCN neurons in their response
to auditory prepulse stimulation
Our neuroanatomical experiments showed that non-olivo-
cochlear neurons of the VNTB project to CRNs without
sending axons collaterals into the VCN. If only this direct
projection is involved in auditory prepulse inhibition of
CRN responses, it might be expected that VCN neurons
would not have the same sensitivity. Therefore, a total of
15 single units of the VCN were subjected to the same
probes previously performed on CRNs. None of the VCN
units exhibited reduction of spike discharges comparable to
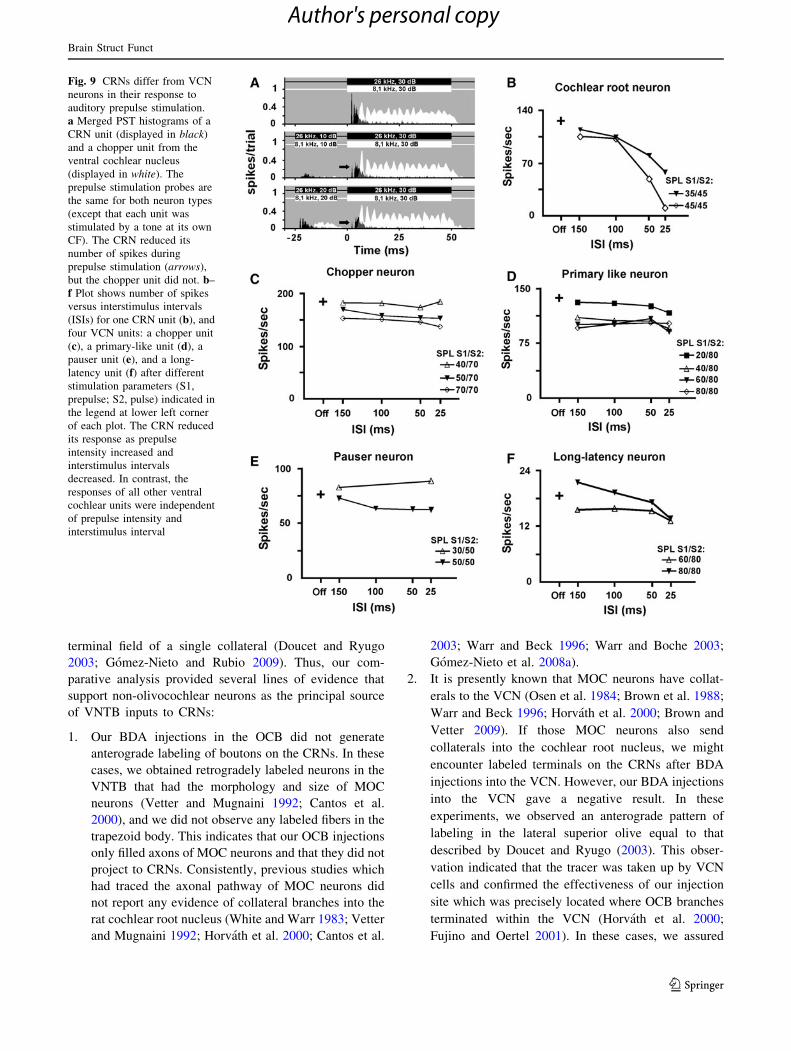
those observed in CRNs. As a reference, Fig. 9a shows a
PST histogram in which the responses of a CRN unit and a
VCN chopper unit were merged. Those units were recorded
in the same animal along the same electrode track, and
were subjected to auditory prepulse stimulation using
identical prepulse and pulse parameters (intensity, dura-
tion, and ISI) presented at the respective neuron’s charac-
teristic frequency. The VCN chopper unit maintained the
number of spikes per trial and the PST histogram-shape as
the prepulse intensity increased. On the contrary, the
response of the CRN unit was drastically reduced by
auditory prepulse stimulation (Fig. 9a). For comparison,
we further recorded the response of CRN and VCN units by
presenting auditory prepulse stimuli at many prepulse
intensities and ISIs (Figs. 9b–f). The number of spikes of
CRN units to the onset of the pulse was reduced as prepulse
intensity increased and ISIs decreased. This inhibitory
effect was maximal when prepulses were presented at high-
intensity level and short ISI (Fig. 9b). Conversely, VCN
units that received auditory prepulse stimulation did not
show such loss in the ratio of the number of spikes. Various
classes of VCN neurons including chopper, primary-like,
pauser, and long-latency neurons showed no evidence of
the inhibition observed in CRNs (Fig. 9c–f). This suggests
that CRNs, unlike any other type of neuron in the VCN, are
evolved to participate in auditory prepulse inhibition.
Discussion
In the present study, we provide insights into the structure
and function of the cholinergic VNTB–CRNs projection
which was described in our previous report (Gomez-Nieto
et al. 2008a). We determined the type of VNTB neuron
which innervates the CRNs and the pathway followed by
the axons to the cochlear root nucleus. Thus, we showed
that non-olivocochlear neurons from the VNTB send direct
and bilateral protections to the cochlear root nucleus via
the trapezoid body. From this result, we can infer that
CRNs receive cholinergic auditory inputs from a system
which is not involved in the descending control of the
cochlea. This led us to question the functional role of
cholinergic efferent innervation in the cochlear root
nucleus, an essential element of the primary acoustic startle
pathway (Lopez et al. 1999). Our results also showed that
CRNs possess muscarinic and nicotinic receptors which are
involved in the inhibition of startle mediating PnC neurons
(Jones and Shannon 2000; Fendt et al. 2001; Bosch and
Schmid 2006). Finally, our electrophysiological study
showed that the neuronal activity of CRNs is strongly
inhibited by auditory prepulses, suggesting the existence of
at least two cholinergic pathways that modulate the pri-
mary acoustic startle circuit.
The VNTB neuronal type which project to the cochlear
root nucleus
The neuronal heterogeneity of the VNTB is consistent
with its multiple efferent projections to a large variety of
structures. Besides its projection to each cochlea, VNTB
has axonal terminations in the dorsal and ventral cochlear
nucleus of both sides, the contralateral lateral superior
olive and the ipsilateral IC (Warr and Beck 1996). Our
previous report demonstrated that VNTB also sends
bilateral projections to the cochlear root nucleus (Gomez-
Nieto et al. 2008a). One of the main goals of the present
study was to elucidate the VNTB neuronal type which
projects to CRNs. This was a complex issue considering
that some types of VNTB neurons project with branching
Brain Struct Funct
123
Author's personal copy
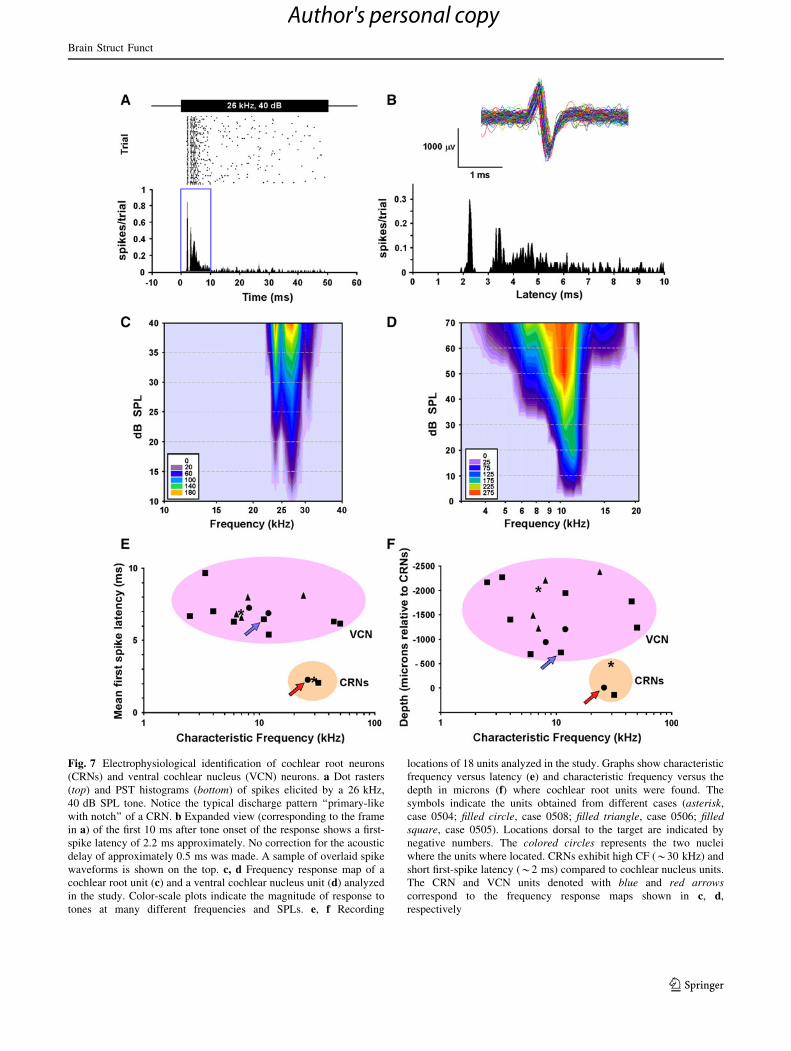
Fig. 7 Electrophysiological identification of cochlear root neurons
(CRNs) and ventral cochlear nucleus (VCN) neurons. a Dot rasters
(top) and PST histograms (bottom) of spikes elicited by a 26 kHz,
40 dB SPL tone. Notice the typical discharge pattern ‘‘primary-like
with notch’’ of a CRN. b Expanded view (corresponding to the frame
in a) of the first 10 ms after tone onset of the response shows a first-
spike latency of 2.2 ms approximately. No correction for the acoustic
delay of approximately 0.5 ms was made. A sample of overlaid spike
waveforms is shown on the top. c, d Frequency response map of a
cochlear root unit (c) and a ventral cochlear nucleus unit (d) analyzed
in the study. Color-scale plots indicate the magnitude of response to
tones at many different frequencies and SPLs. e, f Recording
locations of 18 units analyzed in the study. Graphs show characteristic
frequency versus latency (e) and characteristic frequency versus the
depth in microns (f) where cochlear root units were found. The
symbols indicate the units obtained from different cases (asterisk,
case 0504; filled circle, case 0508; filled triangle, case 0506; filledsquare, case 0505). Locations dorsal to the target are indicated by
negative numbers. The colored circles represents the two nuclei
where the units where located. CRNs exhibit high CF (*30 kHz) and
short first-spike latency (*2 ms) compared to cochlear nucleus units.
The CRN and VCN units denoted with blue and red arrowscorrespond to the frequency response maps shown in c, d,
respectively
Brain Struct Funct
123
Author's personal copy
axons to more than one VNTB target (Warr and Beck
1996). Since the VNTB projects to the cochlear root
nucleus bilaterally with approximately equal magnitude
(Gomez-Nieto et al. 2008a), we rejected in the first
instance VNTB neurons with homolateral projections as a
possible source of inputs to the cochlear root nucleus. To
support this assumption, we reviewed our previous studies
with BDA injection in the IC (e. g. case 05/180 in
Gomez-Nieto et al. 2008a) and we did not observe any
labeled boutons on the CRNs. This is also confirmed by
Warr and Beck (1996) who reported that VNTB neurons
projecting to IC never crossed the midline. Our compar-
ative analysis of BDA-labeled structures after injections
into the axonal projections of VNTB descending pathways
(OCB and TB) and the VCN (see details in Table 1)
indicated that a subset of non-olivocochlear neurons sends
a direct projection to CRNs via the trapezoid body. BDA
was used as neuronal tracer in all these experiments. BDA
is an effective bidirectional pathway tracer which has
excellent collateral–collateral transport within a single cell
(Rajakumar et al. 1993; Chen and Aston-Jones 1998), that
is, dextrans can fill the entire cell with injection into the
Fig. 8 Auditory prepulse inhibition of neuronal activity in the
cochlear root nucleus. The prepulse and pulse parameters (intensity,
duration and interstimulus intervals) used in the probes were showed
in the upper part of each PST histogram. a PST histograms showing
the effects of prepulse tones (gray line) on the CRN response to pulse
tones (black line). Notice the reduction of the spikes per trial (arrow)
after prepulse stimulation. b PST histograms for three prepulse
stimulation probes show the effects of the prepulse intensity level on
the discharge of a CRN. Arrows indicate the reduction of the spikes
per trial with the increase of prepulse intensity. c PST histograms for
three prepulse stimulation probes show the effects of the prepulse
duration on the discharge of a cochlear root neuron. Arrows indicate
the reduction of the spikes per trial as the prepulse duration increased.
d PST histograms for four prepulse stimulation probes show the
effects of the interstimulus interval (ISI) on the discharge of a CRN.
Arrows indicate the reduction of the spikes per trial as ISIs decreased
Brain Struct Funct
123
Author's personal copy
terminal field of a single collateral (Doucet and Ryugo
2003; Gomez-Nieto and Rubio 2009). Thus, our com-
parative analysis provided several lines of evidence that
support non-olivocochlear neurons as the principal source
of VNTB inputs to CRNs:
1. Our BDA injections in the OCB did not generate
anterograde labeling of boutons on the CRNs. In these
cases, we obtained retrogradely labeled neurons in the
VNTB that had the morphology and size of MOC
neurons (Vetter and Mugnaini 1992; Cantos et al.
2000), and we did not observe any labeled fibers in the
trapezoid body. This indicates that our OCB injections
only filled axons of MOC neurons and that they did not
project to CRNs. Consistently, previous studies which
had traced the axonal pathway of MOC neurons did
not report any evidence of collateral branches into the
rat cochlear root nucleus (White and Warr 1983; Vetter
and Mugnaini 1992; Horvath et al. 2000; Cantos et al.
2003; Warr and Beck 1996; Warr and Boche 2003;
Gomez-Nieto et al. 2008a).
2. It is presently known that MOC neurons have collat-
erals to the VCN (Osen et al. 1984; Brown et al. 1988;
Warr and Beck 1996; Horvath et al. 2000; Brown and
Vetter 2009). If those MOC neurons also send
collaterals into the cochlear root nucleus, we might
encounter labeled terminals on the CRNs after BDA
injections into the VCN. However, our BDA injections
into the VCN gave a negative result. In these
experiments, we observed an anterograde pattern of
labeling in the lateral superior olive equal to that
described by Doucet and Ryugo (2003). This obser-
vation indicated that the tracer was taken up by VCN
cells and confirmed the effectiveness of our injection
site which was precisely located where OCB branches
terminated within the VCN (Horvath et al. 2000;
Fujino and Oertel 2001). In these cases, we assured
Fig. 9 CRNs differ from VCN
neurons in their response to
auditory prepulse stimulation.
a Merged PST histograms of a
CRN unit (displayed in black)
and a chopper unit from the
ventral cochlear nucleus
(displayed in white). The
prepulse stimulation probes are
the same for both neuron types
(except that each unit was
stimulated by a tone at its own
CF). The CRN reduced its
number of spikes during
prepulse stimulation (arrows),
but the chopper unit did not. b–
f Plot shows number of spikes
versus interstimulus intervals
(ISIs) for one CRN unit (b), and
four VCN units: a chopper unit
(c), a primary-like unit (d), a
pauser unit (e), and a long-
latency unit (f) after different
stimulation parameters (S1,
prepulse; S2, pulse) indicated in
the legend at lower left corner
of each plot. The CRN reduced
its response as prepulse
intensity increased and
interstimulus intervals
decreased. In contrast, the
responses of all other ventral
cochlear units were independent
of prepulse intensity and
interstimulus interval
Brain Struct Funct
123
Author's personal copy

that MOC neurons were completely filled with the
tracer as our results showed labeled crossed OCB
fibers under the fourth ventricle and retrogradely
labeled neurons which fulfilled the morphological
criteria established for MOC neurons (Vetter and
Mugnaini 1992; Cantos et al. 2000). Following our
BDA injections into the VCN, we did find labeled
terminals on the somata of CRNs. These labeled
axosomatic terminals were only observed ipsilateral to
the injection site and differed from VNTB terminals in
size and axonal branching pattern (Gomez-Nieto et al.
2008a). As reported in earlier studies, their large size
and axonal branching pattern resembled that of
terminals immunolabeled for the vesicular glutamate
transporter 1 which arise from collaterals of auditory
primary axons (Osen et al. 1991; Lopez et al. 1993;
Gomez-Nieto et al. 2008b).
3. The lesion of the OCB after BDA injection in the
VNTB confirmed the non-olivocochlear origin of the
VNTB–CRNs projection. Previous studies demon-
strated that mechanical lesions of the OCB near the
floor of the fourth ventricle were effective in elimi-
nating efferent terminals of MOC neurons (Liberman
1990; Bledsoe et al. 2009). In our material, the
transection of crossed olivocochlear fibers was made
at this particular location and the cochlea was analyzed
histologically to assess the completeness of the lesion.
Our results showed VNTB bead-like axons on CRNs
after the OCB lesion suggesting that the VNTB–CRNs
projection followed a non-olivocochlear pathway. The
same conclusion arises from our injections in the TB
which indicates that VNTB axons innervate CRNs
course via the trapezoid body. This course appears to
be similar to the initial trajectory of CRN axons in the
trapezoid body (Lopez et al. 1993). We verified this,
since the injections in the TB resulted in retrograde
labeling of CRNs somata as well as anterograde
labeling of VNTB terminals in the cochlear root
nucleus.
In sum, our morphology study indicates that non-oli-
vocochlear neurons from the VNTB are a significant source
of axonal terminals in the cochlear root nucleus. These
non-olivocochlear neurons appeared not to send collaterals
to brain structures receiving projections from other VNTB
neuronal types such as the VCN or the cochlea. Therefore,
it is reasonable to conclude that VNTB neurons projecting
to CRNs belong to a distinct neuronal population which do
not participate in controlling the cochlea. This makes it
likely that the VNTB–CRNs projection is involved in the
modulation of the acoustic startle reflex rather than the
medial olivocochlear reflex.
The role of the cholinergic system within the cochlear
root nucleus
Previous neuroanatomical studies have shown strong evi-
dence of acetylcholine release in the cochlear root nucleus
(Vetter et al. 1993a; Yao and Godfrey 1999; Gomez-Nieto
et al. 2008a). In the superior olivary complex, the VNTB is
the largest source of descending cholinergic inputs (Sher-
riff and Henderson 1994; Yao and Godfrey 1998). Our
group has recently demonstrated that CRNs receive their
cholinergic input from VNTB neurons (Gomez-Nieto et al.
2008a). The fact that our RT-PCR analysis showed a sig-
nificant gene expression of muscarinic and nicotinic ace-
tylcholine receptors in the cochlear root nucleus also
supported the release of acetylcholine on CRNs. These
cholinergic receptors are likely to be specific for projec-
tions from the VNTB given that injections of neuronal
tracers in the cochlear root nucleus do not generate retro-
grade labeling in any other cholinergic nuclei (Gomez-
Nieto et al. 2008a). It is presently known that acetylcholine
plays a crucial role in mediating PPI of the acoustic startle
response (reviewed in Fendt et al. 2001). Several studies
have shown that muscarinic receptors, particularly the
subtypes M2 and M4, are involved in PPI mediation at the
level of PnC (Koch et al. 1993; Fendt and Koch 1999;
Jones and Shannon 2000; Bosch and Schmid 2006). Thus,
activation of M2 and M4 receptors subtypes mediates a
strong inhibition of PnC giant neurons (Bosch and Schmid
2006, 2008). Our data showed that CRNs also expressed
M2 and M4 subtypes as confirmed by immunolabeling in
Gomez-Nieto et al. (2008b), and hence, acetylcholine
might have also an inhibitory effect on CRNs. Although the
role of nicotinic receptors in PPI is less studied, several
pharmacological experiments have shown, with contro-
versial results, that stimulation of nicotinic receptors
affects sensorimotor gating (Acri et al. 1994; Schreiber
et al. 2002). In fact, the investigation of the effects of
different nicotine receptors agonist and antagonist on PPI
supports the involvement of the nicotinic receptor subtypes
a4 and b2, and possibly a7 in the control of PPI (Schreiber
et al. 2002; Suemaru et al. 2004). Our result showed that
CRNs contain both a and b subunits, and more specifically,
the a4 and b2 subtypes which were expressed in a rela-
tively high level. Some evidence also suggests that these
nicotinic receptors play a role in a faster PPI pathway
acting at short interstimulus intervals (Bosch and Schmid
2008). This idea together with the fact that CRNs exhibit
all the molecular cholinergic machinery involved in PPI
mediation conforms well with our initial hypothesis: the
VNTB projection might inhibit startle signaling in the
cochlear root nucleus through a cholinergic mechanism.
Brain Struct Funct
123
Author's personal copy

The observed inhibition of CRN responses after auditory
prepulse stimulation, particularly strong when short inter-
stimulus intervals were used, is also consistent with that
suggestion.
Multiple pathways for the auditory inhibition
of the primary acoustic startle circuit
One of the most accepted anatomical models for explaining
the circuitry mediating auditory PPI includes three serially
connected midbrain structures: the IC, the superior col-
liculus, and the pedunculopontine tegmental nucleus which
sends inhibitory cholinergic inputs to the PnC (reviewed in
Fendt et al. 2001). Yet, it is important to note that not all
properties of the PPI such as the shorter ISIs in which this
paradigm occurs (20 ms, Hoffman and Ison 1980) are
explained by the proposed circuit. Recent studies support
the existence of additional pathways that bypass some
components of this long multimodal circuit to mediate fast
auditory PPI (Yeomans et al. 2006; Gomez-Nieto et al.
2008a; reviewed in Li et al. 2009). The cochlear root
nucleus is an obligatory relay station in the primary
acoustic startle pathway (Lee et al. 1996; Lopez et al.
1999), and its neurons possess a tremendous similarity with
giant neurons of the reticular formation. Both CRNs and
giant PnC neurons appear to be similar in their electro-
physiological properties (Lingenhohl and Friauf 1992;
Sinex et al. 2001), direct cholinergic afferent inputs (Fendt
et al. 2001; Gomez-Nieto et al. 2008a), and neurotrans-
mitter machinery (Bosch and Schmid 2006; Gomez-Nieto
et al. 2008b). The resemblance supports the idea that the
acoustic startle reflex might be inhibited in both nuclei of
the primary acoustic startle circuit. Our electrophysiologi-
cal results showed that CRNs responses were strongly
inhibited by auditory prepulse stimulation, particularly at
short ISIs. The reduction in the discharge pattern of CRN
responses was observed after presentation of two consec-
utive pure tones, a prepulse followed by the testing pulse,
separated by an ISI. It could be argued that the observed
inhibition was due to forward masking or the medial oli-
vocochlear reflex which might reduce cochlear gain, and
therefore, the response of primary auditory neurons.
However, our results showed that CRN responses were
drastically reduced at ISIs long enough to avoid forward
masking. Physiological studies concerning forward mask-
ing in auditory nerve and VCN units have shown that there
is no significant inhibition at ISIs longer than 100 ms
(Harris and Dallos 1979; Shore 1995). In our results, we
showed auditory prepulse inhibition of CRNs’ response at
ISIs that were varied up to 200 ms. Furthermore, if forward
masking might contribute to the response reduction at the
shortest ISIs, we should have found a similar reduction in
VCN neurons which also receive primary auditory
afferents. Our results showed that VCN units exhibited
minimal reduction of spike discharges, indicating that the
drastic reduction observed in CRNs is likely to have a more
complex basis. The number of CRN units was small
compared with the VCN units that were recorded in each
animal. This is undoubtedly a consequence of the small
size of the cochlear root nucleus, no more than 40–50
neurons (Merchan et al. 1988), and its relative inaccessi-
bility inside the internal auditory meatus. It is important to
notice that such strong inhibition in the CRNs response was
obtained after stimulations with prepulses of weak inten-
sities and short durations. This suggests that the prepulse
signals were processed by a mechanism that enhances the
effect of the prepulses on the CRNs’ response. A musca-
rinic and nicotinic cholinergic mechanism activated by the
descending cholinergic projection from the VNTB to the
cochlear root nucleus could provide that kind of inhibition.
The VNTB contains at least two distinct groups of cho-
linergic cells with descending pathways, MOC neurons and
non-olivocochlear neurons, which provide cholinergic
inputs to the cochlea and the VCN (Vetter et al. 1991;
Sherriff and Henderson 1994; Warr and Boche 2003). Our
tracer experiments indicate that a subpopulation of non-
olivocochlear neurons project solely to the cochlear root
nucleus. The fact that non-olivocochlear neurons innervate
CRNs without sending collaterals to VCN is consistent
with the electrophysiological data showing that CRNs
differ from VCN neurons in their response to auditory
prepulse stimulation. In sum, our study supports the exis-
tence of multiple neuronal pathways for mediating PPI, and
suggests that the cochlear root nucleus might be involved
in the primary circuit of the auditory PPI. In agreement
with the current models (reviewed in Fendt et al. 2001; Li
et al. 2009) and accounting for the above results, we pro-
pose an alternative neuronal pathway for the mediation of
the auditory PPI (Fig. 10). In this proposed circuit, the
CRNs might inhibit the acoustic startle signaling when the
prepulse is presented at short ISIs. Such fast inhibition
might be mediated by the VNTB–CRNs cholinergic pro-
jection which activates a muscarinic and nicotinic inhibi-
tion mechanism in the cochlear root nucleus. Similarly,
PnC neurons might receive auditory prepulse information
at long ISIs through a slow multi-modal circuit which
involves the IC, the superior colliculus, and finally the
cholinergic projection from the pedunculopontine teg-
mental nucleus to PnC (Fendt et al. 2001, Li et al. 2009).
Both pathways might be interconnected by the reciprocal
projections between the IC and the VNTB (Beyerl 1978;
Faye-Lund 1986; Coleman and Clerici 1987; Vetter et al.
1993b; Warr and Beck 1996; Gomez-Nieto et al. 2008a).
The IC–VNTB–CRNs auditory pathway and other con-
verging inputs to the CRNs like those from the locus
coeruleus, a noradrenergic attention system, might be
Brain Struct Funct
123
Author's personal copy

important to understand the binaural processing observed
in both baseline PPI and attentional modulation of PPI
(Gomez-Nieto et al. 2008a, Gomez-Nieto et al. 2008b).
Acknowledgments We gratefully acknowledge Dr. Sebastian
Hormigo for critically reading an early version of the manuscript. We
thank the reviewers for their valuable comments which helped to
considerably improve the quality of the manuscript. This research was
supported by grants from the Spanish Ministry of Science and Inno-
vation (MICINN, #BFU2010-17754) to Dr. Dolores E. Lopez; the
National Institute on Deafness and Other Communication Disorders
(NIDCD, #DC00341) to Dr. Donal E. Sinex; Fundacion ‘‘Samuel
Solorzano Barruso’’ (FS/6-2012) to Dr. Ricardo Gomez-Nieto; and
the Sao Paulo State Research Foundation (FAPESP, #2008/02771-6)
to Dr. Jose de Anchieta C. Horta-Junior.
References
Acri JB, Morse DE, Popke EJ, Grunberg NE (1994) Nicotine
increases sensory gating measured as inhibition of the acoustic
startle reflex in rats. Psychopharmacology 114(2):369–374
Beyerl BD (1978) Afferent projections to the central nucleus of the
inferior colliculus in the rat. Brain Res 145:209–223
Bledsoe SC Jr, Koehler S, Tucci DL, Zhou J, Le Prell C, Shore SE
(2009) Ventral cochlear nucleus responses to contralateral sound
are mediated by commissural and olivocochlear pathways.
J Neurophysiol 102(2):886–900
Bosch D, Schmid S (2006) Activation of muscarinic cholinergic
receptors inhibits giant neurons in the caudal pontine reticular
nucleus. Eur J Neurosci 24:1967–1975
Bosch D, Schmid S (2008) Cholinergic mechanism underlying
prepulse inhibition of the startle response in rats. Neuroscience
155(1):326–335
Brown MC, Vetter DE (2009) Olivocochlear neuron central anatomy
is normal in alpha 9 knockout mice. J Assoc Res Otolaryngol
10(1):64–75
Brown MC, Liberman MC, Benson TE, Ryugo DK (1988) Brain stem
branches from olivocochlear axons in cats and rodents. J Comp
Neurol 278:591–603
Cantos R, Lopez DE, Sala ML, Rueda J (2000) Study of the
olivocochlear neurons using two different tracers, fast blue and
cholera toxin, in hypothyroid rats. Anat Embryol (Berl)
201:245–257
Cantos R, Lopez DE, Merchan JA, Rueda J (2003) Olivocochlear
efferent innervation of the organ of corti in hypothyroid rats.
J Comp Neurol 459(4):454–467
Chen S, Aston-Jones G (1998) Axonal collateral–collateral transport
of tract tracers in brain neurons: false anterograde labelling and
useful tool. Neuroscience 82:1151–1163
Coleman JR, Clerici WJ (1987) Sources of projections to subdivisions
of the inferior colliculus in the rat. J Comp Neurol 262:215–226
Doucet JR, Ryugo DK (2003) Axonal pathways to the lateral superior
olive labeled with biotinylated dextran amine injections in the
dorsal cochlear nucleus of rats. J Comp Neurol 461(4):452–465
Faye-Lund H (1986) Projection from the inferior colliculus to the
superior olivary complex in the albino rat. Anat Embryol
175:35–52
Fendt M, Koch M (1999) Cholinergic modulation of the acoustic
startle response in the caudal pontine reticular nucleus of the rat.
Eur J Pharmacol 370(2):101–107
Fendt M, Li L, Yeomans JS (2001) Brain stem circuits mediating
prepulse inhibition of the startle reflex. Psychopharmacology
156:216–224
Fujino K, Oertel D (2001) Cholinergic modulation of stellate cells in
the mammalian ventral cochlear nucleus. J Neurosci
21:7372–7383
Gomez-Nieto R, Rubio ME (2009) A bushy cell network in the rat
ventral cochlear nucleus. J Comp Neurol 516:241–263
Gomez-Nieto R, Rubio ME, Lopez DE (2008a) Cholinergic input
from the ventral nucleus of the trapezoid body to cochlear root
neurons in rats. J Comp Neurol 506:452–468
Gomez-Nieto R, Horta-Junior JA, Castellano O, Herrero-Turrion MJ,
Rubio ME, Lopez DE (2008b) Neurochemistry of the afferents
to the rat cochlear root nucleus: possible synaptic modulation of
the acoustic startle. Neuroscience 154:51–64
Gomez-Nieto R, Horta-Junior JA, Castellano O, Sinex DG, Lopez DE
(2010) Auditory prepulse inhibition of neuronal activity in the
rat cochlear root nucleus. In: Lopez Poveda EA, Palmer AR,
Meddis R (eds) The neurophysiological bases of auditory
perception. Springer, New York, pp 79–90
Hancock MB (1982) DAB-nickel substrate for the differential
immunoperoxidase staining of nerve fibers and fiber terminals.
J Histochem Cytochem 30(6):578
Harris DM, Dallos P (1979) Forward masking of auditory nerve fiber
responses. J Neurophysiol 42:1083–1107
Harrison JM, Warr WB, Irving R (1962) Second order neurons in the
acoustic nerve. Science 138:893–895
Hoffman HS, Ison JR (1980) Reflex modification in the domain of
startle: I. Some empirical findings and their implications for how
the nervous system processes sensory input. Psychol Rev
87:175–189
Horta-Junior J de A, Lopez DE, Alvarez-Morujo AJ, Bittencourt JC
(2008) Direct and indirect connections between cochlear root
Fig. 10 Multiple pathways for the auditory inhibition of the primary
acoustic startle circuit. The VNTB-to-CRN pathway mediates fast
acoustic inputs to reduce CRN responses at short ISIs (black dashedarrows), while the IC, SC, PPTg to PnC pathway mediates a slower
pathway for auditory prepulse inhibition at long ISIs (gray dashedarrows). Both CRNs and PnC neurons contain muscarinic and
nicotinic acetylcholine receptors (mAChRs and nAChRs) to mediate
inhibition of neuronal activity in the primary acoustic startle circuit.
CRNs receive direct cholinergic projections from non-olivocochlear
cells via trapezoid body. CN cochlear nucleus, CRNs cochlear root
neurons; IC inferior colliculus, PnC caudal pontine reticular forma-
tion, PPTg pedunculopontine tegmental nucleus, SC superior collicu-
lus, VNTB ventral nucleus of the trapezoid body
Brain Struct Funct
123
Author's personal copy

neurons and facial motor neurons: pathways underlying the
acoustic pinna reflex in the albino rat. J Comp Neurol
507:1763–1779
Horvath M, Kraus KS, Illing RB (2000) Olivocochlear neurons
sending axon collaterals into the ventral cochlear nucleus of the
rat. J Comp Neurol 422(1):95–105
Jones CK, Shannon HE (2000) Effects of scopolamine in comparison
with apomorphine and phencyclidine on prepulse inhibition in
rats. Eur J Pharmacol 391:105–112
Koch M, Kungel M, Herbert H (1993) Cholinergic neurons in the
pedunculopontine tegmental nucleus are involved in the medi-
ation of prepulse inhibition of the acoustic startle response in the
rat. Exp Brain Res 97:71–82
Lee Y, Lopez DE, Meloni EG, Davis M (1996) A primary acoustic
startle pathway: obligatory role of cochlear root neurons and the
nucleus reticularis pontis caudalis. J Neurosci 16:3775–3789
Leitner DS, Cohen ME (1985) Role of the inferior colliculus in the
inhibition of acoustic startle in the rat. Physiol Behav 34:65–70
Li L, Korngut LM, Frost BJ, Beninger RJ (1998a) Prepulse inhibition
following lesions of the inferior colliculus: prepulse intensity
functions. Physiol Behav 65:133–139
Li L, Priebe RPM, Yeomans JS (1998b) Prepulse inhibition of
acoustic or trigeminal startle of rats by unilateral electrical
stimulation of the inferior colliculus. Behav Neurosci
112:1187–1198
Li L, Du Y, Li N, Wu X, Wu Y (2009) Top-down modulation of
prepulse inhibition of the startle reflex in humans and rats.
Neurosci Biobehav Rev 33(8):1157–1167
Liberman MC (1990) Effects of chronic cochlear de-efferentation on
auditory-nerve response. Hear Res 49(1–3):209–223
Lingenhohl K, Friauf E (1992) Giant neurons in the caudal pontine
reticular formation receive short latency acoustic input: an
intracellular recording and HRP-study in the rat. J Comp Neurol
325:473–492
Lingenhohl K, Friauf E (1994) Giant neurons in the rat reticular
formation: a sensorimotor interface in the elementary acoustic
startle circuit? J Neurosci 14:1176–1194
Lopez DE, Merchan MA, Bajo VM, Saldana E (1993) The cochlear
root neurons in the rat, mouse and gerbil. In: Merchan MA, Juiz
JM, Godfrey DA, Mugnaini E (eds) The mammalian cochlear
nuclei: organization and function. Plenum Press, New York,
pp 291–301
Lopez DE, Saldana E, Nodal FR, Merchan MA, Warr WB (1999)
Projections of cochlear root neurons, sentinels of the auditory
pathway in the rat. J Comp Neurol 415:160–174
Merchan MA, Collıa F, Lopez DE, Saldana E (1988) Morphology of
cochlear root neurons in the rat. J Neurocytol 17:711–725
Molina V, Montes C, Tamayo P, Villa R, Isabel Osuna M, Perez J,
Sancho C, Lopez-Albuquerque T, Cardoso A, Castellano O,
Lopez DE (2009) Correlation between prepulse inhibition and
cortical perfusion during an attentional test in schizophrenia. A
pilot study. Prog Neuropsychopharmacol Biol Psychiatry
33(1):53–61
Nodal FR, Lopez DE (2003) Direct input from cochlear root neurons
to pontine reticulospinal neurons in albino rat. J Comp Neurol
460:80–93
Osen KK, Mugnaini E, Dahl AL, Christiansen AH (1984) Histo-
chemical localization of acetylcholinesterase in the cochlear and
superior olivary nuclei. A reappraisal with emphasis on the
cochlear granule cell system. Arch Ital Biol 122(3):169–212
Osen KK, Lopez DE, Slyngstad TA, Ottersen OP, Storm-Mathisen J
(1991) GABA-like and glycine-like immunoreactivities of the
cochlear root nucleus in rat. J Neurocytol 20:17–25
Paxinos G, Watson C (1998) The rat brain in stereotaxic coordinates.
Academic Press, New York
Rajakumar N, Elisevich K, Flumerfelt BA (1993) Biotinylated
dextran: a versatile anterograde and retrograde neuronal tracer.
Brain Res 607:47–53
Schreiber R, Dalmus M, DeVry J (2002) Effects of alpha 4/beta 2-
and alpha 7-nicotine acetylcholine receptor agonists on prepulse
inhibition of the acoustic startle response in rats and mice.
Psychopharmacology 159(3):248–257
Sherriff FE, Henderson Z (1994) Cholinergic neurons in the ventral
trapezoid nucleus project to the cochlear nuclei in the rat.
Neuroscience 58(3):627–633
Shore SE (1995) Recovery of forward-masked responses in ventral
cochlear nucleus neurons. Hear Res 82:31–43
Sinex DG, Lopez DE, Warr WB (2001) Electrophysiological
responses of cochlear root neurons. Hear Res 158:28–38
Suemaru K, Yasuda K, Umeda K, Araki H, Shibata K, Choshi T,
Hibino S, Gomita Y (2004) Nicotine blocks apomorphine-
induced disruption of prepulse inhibition of the acoustic startle in
rats: possible involvement of central nicotinic alpha7 receptors.
Br J Pharmacol 142(5):843–850
Swerdlow NR, Geyer MA (1998) Using an animal model of deficient
sensorimotor gating to study the pathophysiology and new
treatments of schizophrenia. Schizophr Bull 24:285–301
Swerdlow NR, Geyer MA, Braff DL (2001) Neural circuit regulation
of prepulse inhibition of startle in the rat: current knowledge and
future challenges. Psychopharmacology 156:194–215
Vetter DE, Mugnaini E (1992) Distribution and dendritic features of
three groups of rat olivocochlear neurons. A study with two
retrograde cholera toxin tracers. Anat Embryol 185:1–16
Vetter DE, Adams JC, Mugnaini E (1991) Chemically distinct rat
olivocochlear neurons. Synapse 7:21–43
Vetter DE, Cozzari C, Hartman BK, Mugnaini E (1993a) Choline
acetyltransferase in the rat cochlear nuclei: immunolocalization
with a monoclonal antybody. In: Merchan MA, Juiz JM, Godfrey
DA, Mugnaini E (eds) The mammalian cochlear nuclei:
organization and function. Plenum, New York, pp 279–290
Vetter DE, Saldana E, Mugnaini E (1993b) Input from the inferior
colliculus to medial olivocochlear neurons in the rat: a double
label study with PHA-L and cholera toxin. Hear Res 70:173–186
Warr WB, Beck JE (1996) Multiple projections from the ventral
nucleus of the trapezoid body in the rat. Hear Res 93:83–101
Warr WB, Boche JE (2003) Diversity of axonal ramifications
belonging to single lateral and medial olivocochlear neurons.
Exp Brain Res 153:499–513
White JS, Warr WB (1983) The dual origins of the olivocochlear
bundle in the albino rat. J Comp Neurol 219:203–214
Yao W, Godfrey DA (1998) Immunohistochemical evaluation of
cholinergic neurons in the rat superior olivary complex. Microsc
Res Tech 41:270–283
Yao W, Godfrey DA (1999) Vesicular acetylcholine transporter in the
rat cochlear nucleus: an immunohistochemical study. J Histo-
chem Cytochem 47:83–90
Yeomans JS, Lee J, Yeomans MH, Steidl S, Li L (2006) Midbrain
pathways for prepulse inhibition and startle activation in rat.
Neuroscience 142:921–929
Brain Struct Funct
123
Author's personal copy


















