
Mitochondrial DNA hypervariable region-1 sequence variation and phylogeny of the concolor
22
American Journal of Primatology 69:1285–1306 (2007) RESEARCH ARTICLE Mitochondrial DNA Hypervariable Region-1 Sequence Variation and Phylogeny of the Concolor Gibbons, Nomascus KERI MONDA 1,2 , RACHEL E. SIMMONS 1,3 , PHILIPP KRESSIRER 1,4 , BING SU 1,5 , AND DAVID S. WOODRUFF 1 1 Ecology, Behavior & Evolution Section, Division of Biological Sciences, University of California, San Diego, La Jolla, California 2 Department of Epidemiology, University of North Carolina at Chapel Hill, Chapel Hill, North Carolina 3 Genomic Variation Laboratory, Department of Animal Science, University of California, Davis, Davis, California 4 Medical Center of the Ludwig-Maximilians-University, Munich, Germany 5 Key Laboratory of Cellular and Molecular Evolution, Kunming Institute of Zoology, Chinese Academy of Sciences, Kunming, Yunnan, China The still little known concolor gibbons are represented by 14 taxa (five species, nine subspecies) distributed parapatrically in China, Myanmar, Vietnam, Laos and Cambodia. To set the stage for a phylogeographic study of the genus we examined DNA sequences from the highly variable mitochondrial hypervariable region-1 (HVR-1 or control region) in 51 animals, mostly of unknown geographic provenance. We developed gibbon-specific primers to amplify mtDNA noninvasively and obtained 4477 bp sequences from 38 gibbons in North American and European zoos and 4159 bp sequences from ten Chinese museum skins. In hindsight, we believe these animals represent eight of the nine nominal subspecies and four of the five nominal species. Bayesian, maximum likelihood and maximum parsimony haplotype network analyses gave concordant results and show Nomascus to be monophyletic. Significant intraspecific variation within N. leucogenys (17 haplotypes) is comparable with that reported earlier in Hylobates lar and less than half the known interspecific pairwise distances in gibbons. Sequence data support the recognition of five species (concolor, leucogenys, nasutus, gabriellae and probably hainanus) and suggest that nasutus is the oldest and leucogenys, the youngest taxon. In contrast, the subspecies N. c. furvogaster, N. c. jingdongensis, and N. leucogenys siki, are not recognizable at this otherwise informative genetic locus. These results show that HVR-1 sequence is variable enough to define evolutionarily significant units in Nomascus and, if coupled with multilocus micro- satellite or SNP genotyping, more than adequate to characterize their phylogeographic history. There is an urgent need to obtain DNA from Published online 23 April 2007 in Wiley InterScience (www.interscience.wiley.com). DOI 10.1002/ajp.20439 Received 23 August 2006; revised 10 January 2007; revision accepted 12 March 2007 Correspondence to: Dr. David Woodruff, Ecology, Behavior & Evolution Section, Division of Biological Sciences, University of California, San Diego, UCSD, La Jolla, CA 92093-0116. E-mail: [email protected] r r 2007 Wiley-Liss, Inc.
Transcript of Mitochondrial DNA hypervariable region-1 sequence variation and phylogeny of the concolor

American Journal of Primatology 69:1285–1306 (2007)
RESEARCH ARTICLE
Mitochondrial DNA Hypervariable Region-1 SequenceVariation and Phylogeny of the ConcolorGibbons, Nomascus
KERI MONDA1,2, RACHEL E. SIMMONS1,3, PHILIPP KRESSIRER1,4, BING SU1,5,AND DAVID S. WOODRUFF1�1Ecology, Behavior & Evolution Section, Division of Biological Sciences, University ofCalifornia, San Diego, La Jolla, California2Department of Epidemiology, University of North Carolina at Chapel Hill, Chapel Hill,North Carolina3Genomic Variation Laboratory, Department of Animal Science, University of California,Davis, Davis, California4Medical Center of the Ludwig-Maximilians-University, Munich, Germany5Key Laboratory of Cellular and Molecular Evolution, Kunming Institute of Zoology,Chinese Academy of Sciences, Kunming, Yunnan, China
The still little known concolor gibbons are represented by 14 taxa (fivespecies, nine subspecies) distributed parapatrically in China, Myanmar,Vietnam, Laos and Cambodia. To set the stage for a phylogeographicstudy of the genus we examined DNA sequences from the highly variablemitochondrial hypervariable region-1 (HVR-1 or control region) in 51animals, mostly of unknown geographic provenance. We developedgibbon-specific primers to amplify mtDNA noninvasively and obtained4477 bp sequences from 38 gibbons in North American and Europeanzoos and 4159 bp sequences from ten Chinese museum skins.In hindsight, we believe these animals represent eight of the ninenominal subspecies and four of the five nominal species. Bayesian,maximum likelihood and maximum parsimony haplotype networkanalyses gave concordant results and show Nomascus to be monophyletic.Significant intraspecific variation within N. leucogenys (17 haplotypes) iscomparable with that reported earlier in Hylobates lar and less than halfthe known interspecific pairwise distances in gibbons. Sequence datasupport the recognition of five species (concolor, leucogenys, nasutus,gabriellae and probably hainanus) and suggest that nasutus is theoldest and leucogenys, the youngest taxon. In contrast, the subspeciesN. c. furvogaster, N. c. jingdongensis, and N. leucogenys siki, are notrecognizable at this otherwise informative genetic locus. These resultsshow that HVR-1 sequence is variable enough to define evolutionarilysignificant units in Nomascus and, if coupled with multilocus micro-satellite or SNP genotyping, more than adequate to characterize theirphylogeographic history. There is an urgent need to obtain DNA from
Published online 23 April 2007 in Wiley InterScience (www.interscience.wiley.com).DOI 10.1002/ajp.20439
Received 23 August 2006; revised 10 January 2007; revision accepted 12 March 2007
�Correspondence to: Dr. David Woodruff, Ecology, Behavior & Evolution Section, Division ofBiological Sciences, University of California, San Diego, UCSD, La Jolla, CA 92093-0116.E-mail: [email protected]
rr 2007 Wiley-Liss, Inc.

gibbons of known geographic provenance before they are extirpated tofacilitate the conservation genetic management of the surviving animals.Am. J. Primatol. 69:1285–1306, 2007. �c 2007 Wiley-Liss, Inc.
Key words: Hylobatidae; control region; noninvasive genotyping;species; molecular evolution; taxonomy
INTRODUCTION
Until recently, war and habitat inaccessibility frustrated scientific study ofthe Indochinese concolor gibbons of the genus Nomascus. They are variouslyreferred to as the black or crested gibbons but these names are problematic;although adult males are black, adult females are primarily buff-colored, and thecrest varies from prominent to indistinct, even within a species. The variousspecies and subspecies are poorly defined and, not surprisingly, their taxonomicand phylogenetic relationships are still controversial [Bartlett, 2005]. To put ourignorance of these neglected apes in perspective consider the following report: ‘Onthe morning of 24 October 1999, the survey team recorded the songs of threegibbon pairs in Nang Lu foresty the first scientifically confirmed direct evidenceof living N. c. concolor in Vietnam for about 30 years’ [Geissmann et al., 2000:63].Since then another two species, N. nasustus and N. hainanus, with the distinctionof being among the most critically endangered primate species in the world, havealso been recognized in northeastern Vietnam and Hainan Island, China[Geissmann, 2005a,b, 2006b; Geissmann et al., 2003; Groves, 2004; Roos, inprep.]. Here, we present genetic evidence clarifying the relationships of four of thefive species and nine nominal subspecies (Fig. 1, Appendix 1).
For most of its taxonomic history the genus Nomascus has been treated as asubgenus of Hylobates and thought to contain only one species, Hylobates(Nomascus) concolor [Groves, 1972; Marshall & Sugardjito, 1986; and referencestherein] with six subspecies: concolor, lu, hainanus, leucogenys, siki andgabriellae. Ma and Wang [1986] proposed two new Chinese subspeciesjingdongensis and furvogaster, and, following Dao [1983] divided the formersubgenus into two species: the southern Hylobates leucogenys, characterized bywhite or yellow cheek-whiskers in black (male) individuals, and the northernH. concolor, characterized by the lack of contrasting cheek-whiskers in males.Groves and Wang [1990] supported this species distinction and elevated thesouthern yellow-cheeked gibbon H. gabriellae as a third species. Geissmann[1993] also recognized three species, H. concolor, H. leucogenys, and H. gabriellae,and subsequently recognized a fourth species, Hylobates (Nomascus) sp. cf.nasutus [Geissmann, 1995; Geissmann et al., 2000]. Although Zehr [1999]still considered the concolor group as a single species, most researchers havefavored dividing the subgenus into four or five species and eight or ninesubspecies. Furthermore, with the growing realization of the molecular geneticdistances and antiquity of the four gibbon subgenera relative to the age of otherhominoid genera most recent workers have elevated Nomascus from subgenericto generic status [Bartlett, 2005; Brandon-Jones et al., 2004; Geissmann, 1995;Groves, 2001, 2004; Hall et al., 1998; Mootnick & Groves, 2005; Nadler &Streicher, 2004; Roos & Geissmann, 2001]. We support this as our owngenetic research confirms the monophyly of the concolor group and its relativelydeep divergence from the other three clades [Simmons, 2005, Simmons et al.,in prep].
1286 / Monda et al.
Am. J. Primatol. DOI 10.1002/ajp

Fig. 1. Outline map showing the historical distribution of Nomascus species and subspecies [fromKonrad & Geissmann, 2006, modified]. Ranges are shown as they are thought to have occurred 50years ago; current populations are reduced in extent and highly fragmented. Taxa are N. concolor(con) and its subspecies N. c. lu (lu), N. nasutus (nas), N. hainanus (hai), N. leucogenys (leu) and itssubspecies N. l. siki (siki), and N. gabriellae (gab). The three ?-marks indicate uncertaintiessurrounding the identities and current northern ranges of Chinese gibbons surviving east of theRed River; the identity of gibbons now extirpated from a large area of southwestern Yunnan; andthe identity of gibbons found between the ranges of N. l. siki and N. gabriellae. The map is deceptiveas it does not show either the former 1,000 km northern extension of concolor gibbons into China orthe fact that Hainan Island was broadly connected to both Vietnam and China during Pleistocenelow sea stands. Following tradition, most range limits follow rivers or political boundaries.
Nomascus gibbon phylogeny / 1287
Am. J. Primatol. DOI 10.1002/ajp

Following the work of Prouty et al. [1983a,b] gibbons of the genus Nomascusare defined as having 26 pairs of chromosomes (members of the other genera have2n 5 38, 44 or 50) and their species-level taxonomy has been based on geographicdistribution, pelage and other morphological characters. Unfortunately, historicalrange limits of taxa are not well documented and the occurrence of naturalhybridization, which could effect species-level differentiation, remains conten-tious [Fooden, 1996; Geissmann, 2002, 2006a]. Furthermore, the morphologicalcriteria are unreliable guides to taxonomy as anatomical and morphologicalvariation in Nomascus are limited. Although identification of black adult males isfairly straightforward, it is difficult to determine specific or subspecific status offemales (buff-colored as adults) and immature males [Mootnick, 2006]. Othermorphological traits are even more problematic. For example, species-leveldifferentiation between N. leucogenys and N. gabriellae was based on differencesin the penis bone [Groves, 1993; Groves & Wang, 1990]. Unfortunately, only oneN. gabriellae baculum had been studied at that time and Geissmann and Lim[1994] later demonstrated that there was a significant variability in baculum sizeand the particular bone examined was incomplete and attributed to the wrongspecies [Geissmann, 1995]. So although Corbet and Hill [1992] have tabulatedalleged differences between the various taxa traditionally associated with thename Hylobates concolor it should be emphasized that, apart from geographicrange, these are minor and of limited utility, and/or ignore population variability.Groves [1993] was justified in expressing frustration with the mosaic geographicpatterns of character variation in the genus.
Two other approaches to species recognition promise to contribute to theresolution of the controversies surrounding concolor taxonomy. First, behaviormay provide a more reliable guide to the species-level identification of adults thanmorphology. All species of gibbons produce elaborate patterns of vocalizationoften referred to as ‘‘songs’’ and many workers have shown how they havespecies-specific and sex-specific inherited characteristics [see: Brockelman& Schilling, 1984; Geissmann, 1993, 2002; Geissmann & Nijman, 2006; Konrad& Geissmann, 2006; and references therein]. Unfortunately, song variability inthe wild is only now being documented and the songs of captive animals can bemisleading as there is a learned component in their songs. Nevertheless, we cananticipate a phylogeny that is supported by these species-specific behavioralattributes [Geissmann, 2002].
Second, several researchers have begun to describe genetic markers thatshould serve as unequivocal identifiers of an individual gibbon’s taxonomicidentity and relationships. In this context gibbons have featured significantly inthe development of molecular primatology [Di Fiore & Gagneux, 2006]. In thefirst demonstration of noninvasive genotyping involving a non-human primate,Woodruff [1990, 1993] provided evidence that H. lar was variable at amicrosatellite locus amplified from hair. In a preliminary study of the geneticdifferences between gibbon taxa, Garza and Woodruff [1992] sequenced a 252 bpsegment of the cytochrome b (cyt b) gene of the mitochondrial genome fromN. leucogenys, N. gabriellae and an animal identified as siki. The study revealedgenetic evidence of a closer relationship between N. leucogenys and siki thaneither taxon to N. gabriellae, and provided evidence against the then prevailingview of Nomascus as a monospecific clade. Garza and Woodruff [1994]subsequently showed that taxon-specific cyt b sequence variation could be usedto distinguished individuals of the three concolor taxa then in North Americanzoos (N. leucogenys, siki, and gabriellae). However, this data set included onlywhite-cheeked or buff-cheeked individuals and they were consequently unable to
1288 / Monda et al.
Am. J. Primatol. DOI 10.1002/ajp

assess differences that might exist between these taxa and the other blackgibbons. Moreover, the number of informative sites in the cyt b gene wasrelatively low (5% within the sample) and thus limited the resolving power of thelocus. To answer questions left unresolved by the cyt b study we thereforeexamined the more rapidly evolving mitochondrial hypervariable region (HVR-1,sometimes called the control region and less accurately termed the d-loop). Thispaper combines and updates our results from three unpublished theses and anuntraceable report in Chinese [Kressirer, 1993; Monda, 1995; Simmons, 2005; Suet al., 1996]. To interpret our data we also review other reports on variation ingibbon mtDNA [Chatterjee, 2006; Roos, 2004; Whittaker, 2005; Zhang, 1997] andnuclear loci [Zehr, 1999; Zhang, 1997; Zhang et al., 2004]. The phylogeneticprinciples underpinning these analyses are explained by Wagele [2005].
MATERIALS AND METHODS
Our samples representing ten species and subspecies were derived primarilyfrom gibbons in North American zoos supplemented with specimens of otherwiseunavailable taxa from zoos and museums in Europe and China (Appendix 2). Ourmain analysis was based on 476–513 bp sequences of the mtDNA HVR-1 in33 individuals representing seven taxa. We were also able to incorporate theresults of a 159 bp mtDNA HVR-1 sequence survey for 11 Chinese concolorgibbons representing five taxa, including the only data for N. c. furvogaster[Su et al., 1996]. An additional four published hominoid sequences were used asoutgroups in various analyses.
All genotyping was performed non-invasively and non-destructively based onmtDNA extracted and amplified from 1 to 3 plucked hairs/individual of liveanimals or museum skins. We developed a variety of primers to amplify up toapprox. 500 bp of the HVR-1. Most amplifications were performed usingcombinations of four oligonucleotides:
L15997 50-CACCATTAGCACCCAAAGCT-30 [Monda, 1995]L16205 50-AACACAACATGCTTACAAGC-30 [Kressirer, 1993]H16431 50-GTTGGTGATTTCACGGAGGA-30 [Kressirer, 1993]H16498 50-CCTGAAGTAGGAACCAGATG-30[Monda, 1995]The primers are identified according to the human nucleotide reference
sequence [Anderson et al., 1981] of the 30 end, and L or H to indicate light orheavy strand, respectively. Our laboratory protocols are described elsewhere[Monda, 1995] and are available from DSW. We took several precautions to reducethe risk of amplifying nuclear sequences of mitochondrial origin (numts)including re-extracting and verifying key determinations in a second laboratorywithout potential contamination from other non-human primate DNA [Simmons,2005; Simmons et al., in prep.] and comparing our sequences with those reportedfor two whole mitochondrial sequences of H. lar and a partial HVR-1 sequence ofH. lar. Verifications typically involved amplification of a 325 bp fragment usingthe primers H16431 and L16007 [50-CCCAAAGCTAAAATTCTAA-30; Roos& Geissmann, 2001].
We aligned the sequences using Clustal X Version 1.81 [Thompson et al.,1997] and after verification by eye we constructed maximum likelihood,parsimony, and distance trees using PAUP� Version 4.0b [Swofford, 1998] andBayesian inference trees with MrBayes Version 3.0b [Huelsenbeck & Ronquist,2001]. Parsimony analyses used default settings of PAUP� and treated gap statesas ‘‘missing data.’’ Distance analyses were performed with default settings ofPAUP� to optimize minimum evolution. All trees were then rooted by outgroup.
Nomascus gibbon phylogeny / 1289
Am. J. Primatol. DOI 10.1002/ajp

Bootstraps of parsimony and distance methods were each composed of 1,000replicates. The maximum likelihood phylogeny had four steps: a maximumparsimony tree starting point and then three consecutive heuristic searches usingmaximum likelihood criteria. The first search’s likelihood settings employed ageneral time reversible model with gamma-distributed rate variation acrosssites (GTR1G) and estimated base frequencies, rate matrix, and shape. Nearestneighbor interchange (NNI) perturbation drove the state changes. Two sub-sequent heuristic searches used a tree bisection-reconnection (TBR) swappingand the starting point, rate matrix, base frequencies, and shape of the previoussearch. The maximum likelihood tree was then determined from the saved treesof these three searches. Bayesian trees used the GTR1G model on four MCMCchains. Generation number and sample frequency were determined by running abrief preliminary analysis, checking for probability convergence, and estimatingthe values necessary for a large set of equally likely trees. After the secondanalysis, each Bayesian tree’s probabilities file was examined to verify that thelikelihoods converged around a small value range before determining if anappropriate burn-in value had been chosen, and the analysis was repeated twiceto ensure that the MCMC chains were long enough and had converged on thesame tree. Ultimately, we used MrBayes to searched for 5,000,000 generationsand with a burn-in value of 100,000.
Network (4.111; www.fluxus-engineering.com) was used to constructhaplotype networks using median joining [Bandelt et al., 1999] and maximumparsimony [Polzin & Daneschmand, 2003] algorithms. The program TCS[Clement et al., 2000] was used to construct a statistical parsimony networkjoining all haplotypes that connect with a 95% or greater frequency.
RESULTS
The HVR-1 sequences are more informative than the previously publishedcyt b sequences. Comparing a subset of ten representative gibbons over 477 bp wefound 123 variable and phylogenetically informative sites, involving 104transitions and 38 transversions (TS/TV ratio of 2.7:1). Overall, we found a totalof 123 (26%) variable positions and 26 different haplotypes.
Phylogenetic relationships among the eight Nomascus taxa represented bymore than 476 bp of mtDNA HVR-1 sequence are presented in Figures 2 and 3.All phylogenetic trees show similar patterns of tree topology, with long branchlengths between outgroup species representing other hominoids (Homo, Pan,Pongo) (Fig. 2). The concolor gibbons form a monophyletic group (Fig. 2) withinwhich we see consistent evidence for four clusters of sequences that correspond tothe species-level taxa N. leucogenys, N. concolor, N. gabriellae, and N. nasutus(Figs. 2 and 3). Bootstrap values (62–91%) show low to moderate support for thesespecies-level clades (Fig. 2A).
All phylogenetic methods indicate N. nasutus is the most basal group, distinctfrom all other clades with highly significant scores for distance bootstrap (87%)and Bayesian posterior probability (1.00). Depending on which algorithm is usedeither N. gabriellae or N. concolor branch off next. N. leucogenys appears as theyoungest clade regardless of method used.
Our two N. nasutus sequences are very similar differing by only 2.6%. Suchdifferences are relatively small compared with the divergence seen among theleucogenys and comparable to those found within gabriellae and concolor.Although we initially treated one of our specimens as N. hainanus, on the adviceof Christian Roos we have concluded that the animal was misidentified (Appendix 2).
1290 / Monda et al.
Am. J. Primatol. DOI 10.1002/ajp
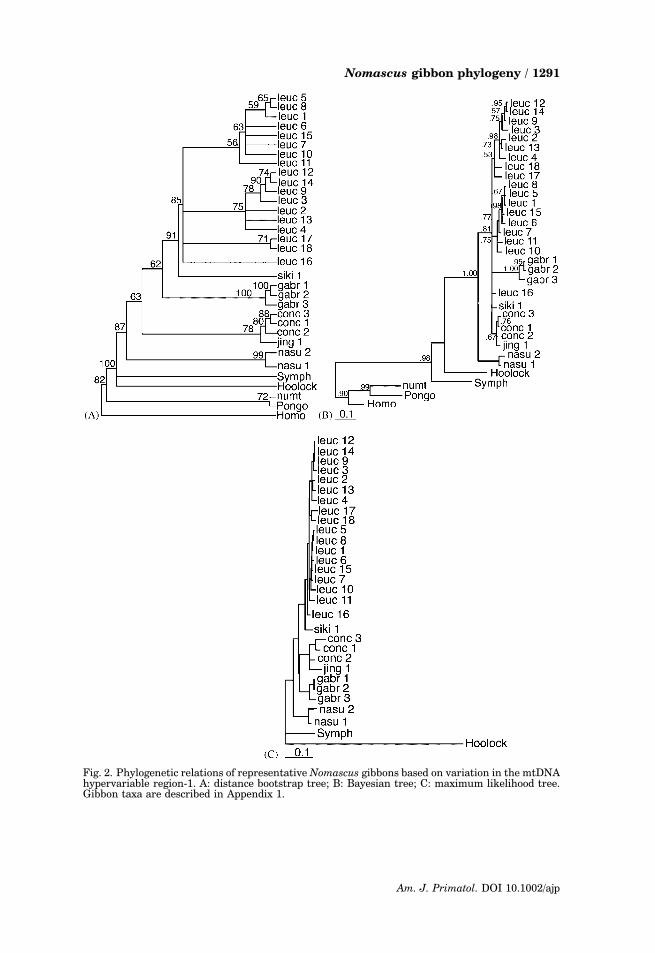
Fig. 2. Phylogenetic relations of representative Nomascus gibbons based on variation in the mtDNAhypervariable region-1. A: distance bootstrap tree; B: Bayesian tree; C: maximum likelihood tree.Gibbon taxa are described in Appendix 1.
Nomascus gibbon phylogeny / 1291
Am. J. Primatol. DOI 10.1002/ajp

Roos (in litt. to DSW, May 2006; in prep.) has now sequenced hainanus of knownprovenance and found them to be much more different from nasutus (cyt b 6.8%,HVR1 14%). This strongly suggests that our Berlin Museum specimen wasmisidentified.
Bayesian (Fig. 2B) and maximum likelihood (Fig. 2C) trees show N. concolorjingdongensis within the concolor cluster; a distance bootstrap (Fig. 2A) shows itas a very closely related outgroup (nine mutations) to the three N. c. concolorgibbons (which vary among themselves by up to 12 mutations).
The 19 gibbons identified as N. leucogenys form a very closely related clusterthat is well separated from the cluster identified as N. gabriellae. N. gabriellaediffers from N. leucogenys by three indels, five transversions and 11 transitions;sites 243–249 are especially variable and may be taxon-specific. The distinctionbetween N. leucogenys and the monophyletic cluster of three N. gabriellae issupported at the 81% level in Bayesian and 62% levels in parsimony and distance.The gibbon identified as siki is seen as an outgroup to the N. leucogenys ratherthan N. gabriellae clade; parsimony, maximum likelihood and distance trees showsiki as a closely related outgroup of the N. leucogenys.
In all tree methods used, 15 N. leucogenys individuals form a monophyleticcluster with two prominent subclusters of closely related individuals we denote asclades A and B (as discovered by Monda, 1995 , and shown most clearly inFig. 2A). We find approximately 2% divergence within each clade and 4%divergence between clades. These clades differ by four transitions at sites 288,294, 307, 348. Two additional subclusters are moderately differentiated from theothers (Fig. 2A shows two individuals of Clade C and one, leuc-16, in Clade D). Thedivision between these subclusters is supported by high posterior probability scores(0.81) in the Bayesian (Fig. 2B) and distance bootstrap methods (85%) (Fig. 2A).There are 26 (5.1%) variable sites within clade A, and 25 (4.9%) within clade B, ofwhich they share only six. Clade A is weakly supported in Bayesian analysis(posterior probability, 0.75) and weakly supported in parsimony (not shown) anddistance bootstraps (Fig. 2A) (62 and 58%, respectively). Clade B is supportedstrongly in Bayesian analysis (0.98) but weakly in bootstraps (parsimony, 56%,distance, 75%).
The maximum parsimony haplotype network (Fig. 3) shows the abovepatterns more clearly and also reveals N. leucogenys clades A and B, and a small
Fig. 3. Maximum parsimony haplotype network showing phylogenetic relations of representativeNomascus gibbons. Gibbons are the same as those described in Fig. 2.
1292 / Monda et al.
Am. J. Primatol. DOI 10.1002/ajp

group clustering with the N. l. siki individual as a closely related outlier. Themedian joining network gave a similar result. The TCS generated statisticalparsimony network (not shown) recognized four clusters concordant with sevenleucogenys A (excluding animal leuc 11) and five leucogenys B (excluding leuc 4),two leucogenys D, and the four concolor1jingdongensis. The remaining tenindividuals (plus outgroups Hoolock hoolock and Symphalangus syndactylus)were not related at our stringent 95% level.
Su et al. [1996] provide some additional information on sequence variation infive Chinese Nomascus taxa (Appendix 2). Unfortunately, and despite the use ofthe same primers as Kressirer [1993], technical problems experienced inamplifying mtDNA from the museum skin samples resulted in only 159 bp ofcomparable sequence for the six taxa compared. Su et al. found no indels and 39(25%) of the sites (not including the outgroup Symphalangus) were variable with8–23 transitions and 1–6 transversions. He used a 6:1 TS/TV ratio. The mostparsimonious tree shown (Fig. 4) is identical to the maximum likelihood tree; treelength 5 78, CI 5 0.73. In agreement with our more comprehensive 477 bpanalyses N. nasutus [misidentified in Su et al. as N. n. hainanus] is seen as theoutgroup. The previously unknown N. c. furvogaster and N. c. jingdongensis areseen as clustering with N. c. concolor (0–2 TV; 0–17 TS). The two N. leucogenysfrom the town of Hekou on the Vietnamese border in southern Yunnan differfrom one another at one transversion and eight transitions.
DISCUSSION
The results reported here were based primarily on DNA extracted fromfreshly plucked hair. Although shed hair is no longer regarded as a reliable sourceof high quality DNA [Morin & Goldberg, 2003; Thalmann et al., 2004; Woodruff,2003] noninvasive genotyping has made it possible to study these apes which areotherwise very difficult to sample. We are confident the sequences presented here
Fig. 4. Relationships among Chinese Nomascus gibbons. Most parsimonious tree based on 159 bpsequences of the HVR-1. Redrawn from Su et al. [1996: Fig. 2b] showing branch lengthsproportional to the number of base substitutions.
Nomascus gibbon phylogeny / 1293
Am. J. Primatol. DOI 10.1002/ajp

are verified mtDNA sequences and not confounded by mitochondrial pseudogenes(numts) or other hominoid contaminants [Bensasson et al., 2001; Simmons et al.,in preparation; Thalmann et al., 2004].
Our results and their interpretation are critically dependent on theidentification of the individuals studied. The specimen data presented inAppendix 2 will facilitate their replication and revision when necessary.
The mtDNA relationships seen are supportive of, but less ambiguous than,the morphology based taxonomy. As this is the first report in which multipleindividuals of each species have been considered, questions presupposingthe availability of information on typical intraspecific genetic variation can nowbe addressed. In particular, the large sample of N. leucogenys (19 haplotypes:the average pairwise distance is 0.03570.014 with a 29.8% percent sequencedifference) can now be compared to the recently published data on comparablevariation in 46 Hylobates lar (0.02470.014 with a 17.9% sequence difference)[Woodruff et al., 2005]. These percent sequence differences are comparable to ourestimates for taxa represented by smaller sample sizes: N. concolor (N 5 3;0.049670.0145), N. gabriellae (N 5 3; 0.024770.0214), and N. nasutus (N 5 2;0.0299). Including the single N. l. siki with the leucogenys changes the averagepairwise distance only slightly: 0.03870.015. Such within taxon variability datawill be invaluable in interpreting the observed between-taxa differences.
At the mtDNA locus studied the concolor gibbons are a monophyletic clade.Their karyotypic distinction (2N 5 52) and mtDNA genetic distance from theother three groups of living gibbons supports their recognition as a separategenus. Divergence between Hylobates and Nomascus is described elsewhere but iscomparable to or greater than that between Nomascus and either Symphalangusor Hoolock, with highly significant scores for distance bootstrap (100%) andBayesian posterior probability (1.00) [Simmons et al., in prep.]. Within the genusare at least four species or species groups (N. concolor, N. leucogenys, N. nasutus,and N. gabriellae) that differ at more than the 5% divergence level; a levelcomparable to that seen in comparably documented species in Hylobates[Simmons et al., in prep]. Below that 5% divergence level it is clear that moregenotyping of individuals of known geographic provenance is required to resolvethe issues and that the following discussion of siki, jingdongensis, and furvogastermust be viewed as preliminary.
Our genetic result showing N. nasutus as the basal group is in agreementwith the findings of Roos [2004] which were based on his survey of cyt b variation.This conclusion is also supported by a preliminary phylogenetic tree based onvocal data for all gibbon species, including the four species of crested gibbons[Konrad & Geissmann, 2006]. Although the relevant bootstrap values in thebehavioral analysis are very low, this tree appears to provide additional supportfor Groves [1993] hypothesis that, within Nomascus, northern species are morebasal than the southern species.
Nomascus nasutus is very different from Nomascus concolor, the northerngibbon with which they have been long confused. The genetic distance betweenour two samples is less than the variation between the A and B clades ofN. leucogenys, or between the three N. gabriellae, or between the fourN. c. concolor. This supports our argument that there is only one speciesrepresented in our sample and not two as reported earlier (see Appendix 1).Christian Roos (in litt. to DSW, April 2006) has recently discovered that Patzi (seeAppendix 2) is genetically closely related to gibbons from the two remaining wildpopulations in Vietnam. His results, when published, should resolve the problemsof the relationships between the surviving northeastern populations of gibbons.
1294 / Monda et al.
Am. J. Primatol. DOI 10.1002/ajp

In reconstructing the evolutionary relations between N. nasutus and N. hainanusit is important to realize that today’s isolated island population (Fig. 1) has beenrepeatedly connected with those on the mainland whenever sea levels fell 425 m(Appendix 1); Neogene hypothermal phase paleogeography could also havebrought hainanus and siki into geographic proximity.
Our data permit the first assessment of the genetic merits of the twosubspecies of Chinese N. concolor named by Ma and Wang [1986]. Our 170 bpanalysis involved four representatives of the central subspecies jingdongensis andthey cluster closely with the three N. c. concolor. N. jingdongensis would notstand out as a distinct taxon if it were a H. lar or N. leucogenys. It is a littledifferent from the southern N. c. concolor but such differences would be expectedover the geographic distances involved. In the case of the western subspecies,furvogaster, we have only one museum specimen. Nevertheless, Su et al. [1996]found this individual’s sequence (Fig. 4) sits even closer to topotypic N. c. concolorthan the four jingdongensis. This suggests that furvogaster has little merit ifmtDNA genetic distances are the metric of subspecific status.
There are two possible explanations for the multiple clades withinN. leucogenys. One is that N. leucogenys contains several distinct haplotypesthat exist in sympatry. This hypothesis can be tested by establishing thegeographic distribution of the various clades, especially clades A and B. It is alsopossible that a cryptic taxon has been absorbed into today’s N. leucogenys. Therange of N. leucogenys is large (Fig. 1), but not sufficiently large or geographicallyheterogeneous to lead one to expect to find multiple allopatric haplotypes. Theaverage distance between A and B group individuals (0.036470.0018) is less thanbetween subspecies in comparable groups, but by a very small margin [macaquesZ0.041; Rosenblum et al., 1997; orangutans Z0.04117.0013; Warren et al.,2001]. Genotyping N. leucogenys individuals of known geographic provenance willbe crucial to interpret the genetic variation. In either situation (sympatricvariants or allopatric subspecies), it is desirable to maintain the full extent of thisspecies’ variability in the captive population.
Our mtDNA data do not support the elevation of siki to species status. Threeof the four individuals that cluster with our H. l. siki are identified asN. leucogenys. Couturier and Lernould [1991] found no reliable morphologicaldistinction between female siki and N. leucogenys; adult males are of courseindistinguishably black with white cheeks. The pairwise differences between oursample identified as siki and the leucogenys are within the range of intraspecificvariation seen in N. leucogenys and H. lar. Although Couturier and Lernould[1991] found their specimens of siki and leucogenys differed by a reciprocaltranslocation between chromosomes 1 and 22, no other workers appear to havekaryotyped their samples and the phylogenetic significance of this discoveryremains unexplored.
Some additional data are provided by Zhang [1997] in a little known reporton cytochrome b variation. Zhang compared five new 252 bp sequences for oneH. hoolock of unknown origin, two northern concolor from the Vietnam–Chinaborder, and two N. leucogenys from northern Vietnam to 11 of the 26 sequencesreported by Garza and Woodruff [1992]. His maximum parsimony analysisshowed that Nomascus was a well-defined sister group (bootstrap value 5 98) tothe other three genera (Hoolock and Syndactylus and Hylobates) and, within itscluster of nine individuals, he found siki as the outgroup to leucogenys, and welldifferentiated from both concolor and gabriellae. Although he argued that thisresult favored the recognition of siki as a separate species in our opinion thebootstrap value is too low (86) to warrant such revision.
Nomascus gibbon phylogeny / 1295
Am. J. Primatol. DOI 10.1002/ajp

Two additional cytochrome b analyses have been reported. Hall et al. [1998]examined the entire 1,040 bp sequence in six species and all four genera but inonly eight individuals. They confirmed the species-level divergence of leucogenysand gabriellae but found insufficient variation to resolve the relationships amongthe genera. Roos [2003, 2004] provides a more recent analysis of the entiresequences of 24 Nomascus. In close agreement with our results, he found fourspecies-level taxa within his samples: nasutus, concolor, leucogenys, gabriellae.N. nasutus [represented by Patzi] had the deepest root (8% interspecificdivergence). Six N. leucogenys and three N. gabriellae show moderateintraspecific divergence (4% each) and more than was found among nineN. concolor. Four siki were examined: two individuals were about 2% differentfrom gabriellae and two were about 1% different from the leucogenys. In theabsence of specimen and locality data it is not possible to interpret this finding ofparaphyly in siki as it could be due to specimen misidentification and/orhybridization. Both natural and artificial (captive) hybrids have been identified[Mootnick, 2006, personal communication, May 2006] and, more generally,hybridization has to be recognized as a normal process in gibbon evolution[Arnold & Meyer, 2006].
If the historical taxonomic problems of the gibbons have arisen in part fromthe practice of defining taxa based on characters of single individuals we shouldbe cautious not to further contribute to such malpractice. Although we have triedto go beyond dependence on a typological approach there are several issues thatstill frustrate the elucidation of concolor gibbon phylogeny. First, taxa should notbe defined solely on the basis of maternally transmitted mtDNA patterns.Comparable genetic studies of nuclear variation (autosomal sequences,Y-chromosome sequences, and microsatellite allele frequencies) and karyotypesare essential components of contemporary species characterization. Althoughfamily-wide surveys have not yet been undertaken, an indication of theirresolving power is illustrated by, for example, Chambers et al. [2004], Tanakaet al. [2004] and Zhang et al. [2004]. Second, vocalization, which may serve as apartial reproductive isolating mechanism must also be considered [Konrad& Geissmann, 2006]. Third, it is important to recognize that today’s geographicranges for some species are recently contracted and fragmented. Subfossil teethand old paintings show that until 1,000–2,000 years ago gibbons ranged 1,600 kmfurther north in China to the level of the Huang (Yellow) River [Geissmann, 1995;Groves, 1972; Marshall & Sugardjito, 1986; van Gulik, 1967]; Hoolock andnorthern Nomascus have been extirpated over most of their historical ranges.
Finally, there is a need to pursue additional karyological studies of gibbons astheir chromosomes are very unusual among the mammals in exhibiting a ten-foldhigher incidence of chromosomal rearrangements (especially translocations),most of which appear to be species-specific [Arnold et al., 1996; Couturier et al.,1992; Bigoni & Stanyon, 2006; Carbone et al., 2006; Hirai et al., 2005; Jauch et al.,1992; Koehler et al., 1995; Mueller et al., 2003; Nie et al., 2001]. As chromosomalrearrangements can function as postmating reproductive isolating mechanismsand contribute to stasipatric speciation [White, 1978; King, 1993] they warrantclose examination in species complexes exhibiting parapatric distributionpatterns like the gibbons. Effort should be made to characterize Nomascuskaryologically before key populations are extirpated. Consideration of theprocesses of karyotypic evolution suggested to Mueller et al. [2003] thatNomascus was the last (youngest) hylobatid genus to diverge [between 10 and5 Mya according to Groves, 2004]; a conclusion supported by our own work[Simmons et al., in prep.] and Chatterjee [2006], and at odds with Roos and
1296 / Monda et al.
Am. J. Primatol. DOI 10.1002/ajp

Geissmann [2001] who concluded that Nomascus was basal within the family.This difference of opinion is based on different data to those described here andwith the increased concordance between data-sets it is clear that within a fewyears the phylogeny of the surviving gibbons will be fully resolved and stable.
The concolor gibbons are variously listed by regional governments and NGOsas globally threatened or, in the cases of N. nasutus and N. hainanus, criticallyendangered [IUCN, 2006]. Gibbons are hunted for food, the alleged medicinalvalue of their parts, and as trophies and pets [Sterling et al., 2006]. In addition,habitat degradation results in the survivors living in small isolated patches offorest and their disappearance across much of their historic range [Chivers, 2005;Konstant et al., 2003; D.S. Woodruff, personal observation in Xishuangbanna,July 2006]. This is vividly illustrated by the widely separated confirmed-occurrence dots on recent maps of Vietnam [Geissmann et al., 2000; Nadler &Streicher, 2004]. Time is fast disappearing to document the genetic variability ofthe remaining animals and provide a foundation for the sound conservationmanagement of both captive and free-ranging gibbons. The real promise ofmultilocus genetic data is that it will permit a partial reconstruction of theoriginal phylogeographic patterns. Multilocus genotyping of all available museumspecimens and noninvasive genotyping of the surviving wild individuals willpermit resolution of the taxonomic ambiguities and the estimation of historicalrates of gene flow and natural hybridization.
ACKNOWLEDGMENTS
We thank Kurt Benirschke, Warren Brockelman, Helen Chatterjee, ThomasGeissmann, Colin Groves, Alan Mootnick and Christian Roos for sharing theirknowledge of gibbons. Ronald Tilson and Cathy Castle facilitated Monda’soriginal study of the AZA gibbons. Francine Frasier, Dawn Field, Hopi Hoekstra,John Huelsenbeck, Pascal Gagneux, Nick Mundy, Sukamol Srikwan and RomelHokanson provided advice or technical assistance in the laboratory or with thephylogenetic analyses. Our studies were supported, in part, by the AZA GibbonTaxon Advisory Group, the US National Science Foundation, the ChineseAcademy of Sciences, the University of Munich, and the University of California.
REFERENCES
Anderson S, Bankier AT, Barrell BG, deBruijin MHL, Coulson AR, Drouin J, EperonIC, Nierlich DP, Roe BA, Sanger F,Schreier PH, Smith AJH, Staden R, YoungIG. 1981. Sequence and organization of thehuman mitochondrial genome. Nature290:457–465.
Arnold ML, Meyer A. 2006. Natural hybridiza-tion in primates: one evolutionary mechan-ism. Zoology 109:261–276.
Arnold N, Stanyon R, Jauch A, O’Brien P,Weinberg J. 1996. Identification of complexchromosome rearrangements in the gibbonby fluorescent in situ hybridization (FISH)of a human chromosome 2q specific micro-library, yeast artificial chromosomes, andreciprocal chromosome painting. CytogenetCell Genet 74:80–85.
Baker LR, Geissmann T, Nadler T, Long B,Walston J. 2002. Cambodia: Primate field
guide. Phnom Penh, Cambodia: Fauna &Flora Cambodia. 8 p.
Bandelt H-J, Forster P, Rohl A. 1999. Median-joining networks for inferring intraspecificphylogenies. Mol Biol Evol 16:37–48.
Bartlett TQ. 2005. The hylobatidae. In: Camp-bell CJ, Fuentes A, MacKinnon KC, PangerM, Bearder SK, editors. Primates in per-spective. New York: Oxford UniversityPress. p 274–289.
Bensasson D, Zhang D-X, Hartl DL, HewittGM. 2001. Mitochondrial pseudogenes: evo-lution’s misplaced witnesses. TREE 16:314–321.
Bigoni F, Stanyon R. 2006. Hylobates concolor.In: O’Brien SJ, Menninger JC, Nash WG,editors. Atlas of mammalian chromosomes.Hoboken, NJ: Wiley. p 160.
Bleisch WV, Jiang X. 2000. Action plan forconservation of the gibbons of the Wuliang
Nomascus gibbon phylogeny / 1297
Am. J. Primatol. DOI 10.1002/ajp

Mountains. Sino-Dutch Forest Conserva-tion and Community Development Project,Kunming, Yunnan, China.
Bleisch W, Zhang Y. 2004. The view across theborder: China and the future of Vietnameseprimates. In: Nadler T, Streicher U, HaThang L, editors. Conservation of primatesin Vietnam. Hanoi: Frankfurt Zool Soc,Vietnam Primate Conserv Progr, CucPhuong National Park, Endangered Pri-mate Rescue Center. p 107–114.
Brandon-Jones D, Eudey AA, Geissmann T,Groves CP, Melnick DJ, Morales JC, She-kelle M, Stewart CB. 2004. Asian primateclassification. Int J Primatol 25:97–164.
Brockelman WY, Schilling D. 1984. Inheri-tance of stereotyped gibbon songs. Nature312:634–636.
Carbone L, Vessere GM, Hallers BFH, Zhu B,Osoegawa K, Mootnick M, Kofler A, Wien-berg J, Rogers J, Humphray S, Scott C,Harris RA, Milosavljevic A, de Jong PA.2006. A high-resolution map of syntenydisruptions in gibbon and human genomes.PLoS Genet 2:2162–2175.
Chambers KE, Reichard UH, Moller A, NowakK, Vigilant L. 2004. Cross-species amplifica-tion of human microsatellite markers usingnoninvasive samples from white-handedgibbons (Hylobates lar). Am J Primatol 64:19–27.
Chan BPL, Fellowes JR, Geissmann T, ZhangJ, editors. 2005. Status survey and conser-vation action plan for the Hainan gibbon—version I (last updated November 2005).Hong Kong: Kadoorie Farm & BotanicGarden Technical Report 3. p 1–33.
Chatterjee HJ. 2006. Phylogeny and biogeo-graphy of gibbons: a dispersal-vicarianceanalysis. Int J Primatol 27:699–712.
Chivers DJ. 2005. Gibbons: the smallapes. In: Caldecott J, Miles L, editors. Worldatlas of great apes and their conservation.Berkeley: University of California Press.p 205–214.
Clement M, Posada D, Crandall KA. 2000.TCS: a computer program to estimate genegenealogies. Mol Ecol 9:1657–1659.
Corbet GB, Hill JE. 1992. The mammals of theIndomalayan Region: a systematic review.Oxford: British Museum (Natural History)& Oxford University Press.
Couturier J, Lernould JM. 1991. Karyotypicstudy of four gibbon forms provisionallyconsidered as subspecies of Hylobates(Nomascus) concolor (Primates, Hylobati-dae). Folia Primatol 56:95–104.
Couturier J, Dutrillaux B, Turleau C, De-Grouchy J. 1992. Comparative karyotypingof four gibbon species or subspecies. AnnGenet 25:5–10.
Dao VT. 1983. On the north Indochinesegibbons (Hylobates concolor) (Primates:
Hylobatidae) in North Vietnam. J HumEvol 12:367–372.
DiFiore A, Gagneux P. 2006. Molecular pri-matology. In: Campbell CJ, Fuentes A,MacKinnon KC, Panger M, Bearder SK,editors. Primates in perspective. New York:Oxford University Press. p 369–393.
Fooden J. 1996. Zoogeography of Vietnameseprimates. Int J Primatol 67:845–899.
Fooden J, Quan G, Luo Y. 1987. Gibbondistribution in China. Acta Theriol Sin 7:161–167.
Garza JC, Woodruff DS. 1992. A phylogeneticstudy of the gibbons (Hylobates) using DNAobtained noninvasively from hair. Mol Phy-logen Evol 1:202–210.
Garza JC, Woodruff DS. 1994. Crested gibbon(Hylobates [Nomascus]) identification usingnoninvasively obtained DNA. Zoo Biol 13:383–387.
Geissmann T. 1989. A female black gibbon,Hylobates concolor subspecies, from north-eastern Vietnam. With appendix: a note onthe Laotian black gibbon, H. concolor lu. IntJ Primatol 10:455–476.
Geissmann T. 1993. Evolution of communica-tion in gibbons (Hylobatidae). Ph.D. disser-tation. Zurich University, Switzerland.
Geissmann T. 1995. Gibbon systematics andspecies identification. Int Zoo News 42:65–77.
Geissmann T. 1997. New sounds from thecrested gibbons (Hylobates concolor group):first results of a systematic revision. In:Zissler D, editor. Verhandlungen derDeutschen Zoologischen Gesellschaft: Kurz-publikationen, 90. Jahresversammlung,Gustav Fischer, Stuttgart. p 170.
Geissmann T. 2002. Taxonomy and evolutionof gibbons. Evol Anthropol 11(Suppl 1):28–31.
Geissmann T. 2005a. Der Hainan-Schopfgib-bon: Der bedrohteste Menschenaffe derWelt. Gibbon J 1:10–12.
Geissmann T. 2005b. Auf der Suche nach denletzten Gibbons von Hainan. Gibbon J 1:18–22.
Geissmann T. 2006a. Gibbon research lab.Available from: http://www. gibbons.de
Geissmann T. 2006b. Schutz des Hainan-Schopfgibbons, des seltensten Menschenaf-fen der Welt: ein Projektbericht. Gibbon J 2:11–13.
Geissmann T, Lim KKP. 1994. Extraction ofbacula from tanned gibbon skins. RafflesBull Zool 42:29–41.
Geissmann T, Nijman V. 2006. Calling in wildsilvery gibbons (Hylobates moloch) in Java(Indonesia): behavior, phylogeny, and con-servation. Am J Primatol 68:1–19.
Geissmann T, Nguyen Xuan D, Lormee N,Momberg F. 2000. Vietnam primate con-servation status review 2000. Part 1:
1298 / Monda et al.
Am. J. Primatol. DOI 10.1002/ajp

Gibbons. Hanoi: Fauna & Flora Interna-tional, Indochina Programme. 139p.
Geissmann T, La Quang T, Trinh Dinh H, VuDinh T, Dang Ngoc C, Pham Duc T. 2003.Rarest ape species rediscovered in Vietnam.Asian Primates 8(3–4):8–9.
Geissmann T, Traber S, von Allmen A. 2006.Das Nangunhe-Naturreservat, ProvinzYunnan, China: Ein Projektbericht [Nan-gunhe Nature Reserve, Yunnan Province,China: a project report]. Gibbon J 2:14–17.
Groves CP. 1972. Systematics and phylogenyof gibbons. In: Rumbaugh, DM, editor.Gibbon and siamang, vol. 1. Basel: Karger.p 1–89.
Groves CP. 1993. Speciation in living hominoidprimates. In: Kimbel WH, Martin LB,editors. Species, species concepts, and primateevolution. New York: Plenum. p 109–121.
Groves CP. 2001. Primate taxonomy. Washing-ton, DC: Smithsonian Institution Press.
Groves CP. 2004. Taxonomy and biogeographyof primates in Vietnam and neighbouringregions. In: Nadler T, Streicher U, HaThang Long, editors. Conservation of pri-mates in Vietnam. Hanoi: Frankfurt ZoolSoc, Vietnam Primate ConservationProgramme, Cuc Phuong National Park,Endangered Primate Rescue Center.p 15–22.
Groves CP, Wang Y. 1990. The gibbons of thesubgenus Nomascus (Primates, Mammalia).Zool Res 11:148–154.
Hall LM, Jones D, Wood B. 1998. Evolution ofthe gibbon subgenera inferred from cyto-chrome b DNA sequence data. Mol Phylo-gen Evol 10:281–286.
Hirai H, Wijayanto H, Tanaka H, MootnickAR, Hayano A, Perwitasari-Farajallah D,Iskandriati D, Sajuthi D. 2005. A whole-armtranslocation (WAT8/9) separating agilegibbons and its evolutionary features. Chro-mosome Res 13:123–133.
Huelsenbeck JP, Ronquist F. 2001.MRBAYES: Bayesian inference of phyloge-netic trees. Bioinformatics 17:754–755.
IUCN. 2006. IUCN Red list of threatened spe-cies. Available from: http://www.iucnredlist.org.
Jauch A, Weinberg J, Stanyon R, Arnold N,Tofanelli S, Ishida T, Cremer T. 1992.Reconstruction of genomic rearrangementsin great apes and gibbons by chromosomepainting. Proc Natl Acad Sci USA 89:8611–8615.
Jiang XL, Wang YX. 1999. Population anddistribution of black-crested gibbons (No-mascus concolor jingdongensis) in WuliangNature Reserve, Jingdong, Yunnan. ZoolRes 20:421–425.
King M. 1993. Species evolution: the role ofchromosome change. Cambridge: Cam-bridge University Press.
Koehler U, Bigoni F, Weinberg J, Stanyon R.1995. Genomic reorganization in the con-color gibbon (Hylobates concolor) revealedby chromosome painting. Genomics 30:287–292.
Konrad R, Geissmann T. 2006. The vocaldiversity and taxonomy of crested gibbons(genus Nomascus) in Cambodia. IntJ Primatol 27:713–745.
Konstant WR, Mittermeier RA, Rylands AB,Butynski TM, Eudey AA, Ganzhorn J,Kormos R. 2003. The world’s top 25 mostendangered primates. Asian Primates 8:29–34.
Kressirer P. 1993. Eine Molekulare Phylogen-ie der Gibbons (Hylobatidae). Munchen:Diplomarbeit der Fakultat fur Biologie derLudwig-Maximilians-Universitat.
Ma S, Wang Y. 1986. The taxonomy anddistribution of gibbons in southern Chinaand its adjacent region, with descriptionof three new subspecies. Zool Res 7:393–410.
Ma S, Wang Y, Poirier FE. 1988. Taxonomy,distribution, and status of gibbons (Hylo-bates) in southern China and adjacentareas. Primates 29:277–286.
Marshall J, Sugardjito J. 1986. Gibbonsystematics. In: Swindler DR, Erwin J,editors. Comparative primate biology,vol. 1: systematics, evolution and anatomy.New York: Liss. p 137–185.
Monda K. 1995. A phylogenetic study of thegibbons (Hylobates) using the controlregion of the mitochondrial genome. SanDiego: Master’s thesis, University ofCalifornia.
Mootnick AR. 2006. Gibbon (Hylobatidae)species identification recommended for res-cue or breeding centers. Primate Conserv21:103–138.
Mootnick AR, Groves CP. 2005. A new genericname for the hoolock gibbon (Hylobatidae).Int J Primatol 26:971–976.
Morin PA, Goldberg TL. 2003. Determinationof genealogical relationships from geneticdata: a review of methods and applications.In: Chapais B, Berman CM, editors. Kinshipand behavior in primates. Oxford: OxfordUniversity Press. p 15–45.
Mueller S, Hollatz M, Weinberg J. 2003.Chromosomal phylogeny and evolution ofgibbons (Hylobatidae). Hum Genet 113:493–501.
Nadler T, Streicher U. 2004. The primates ofVietnam—an overview. In: Nadler T, Strei-cher U, Ha Thang L, editors. Conservationof primates in Vietnam. Hanoi: FrankfurtZool Soc, Vietnam Primate ConservationProgramme, Cuc Phuong National Park,Endangered Primate Rescue Center. p 5–11.
Nie W, Rens W, Wang J, Yang F. 2001.Conserved chromosome segments in
Nomascus gibbon phylogeny / 1299
Am. J. Primatol. DOI 10.1002/ajp

Hylobates hoolook revealed by human andH. leucogenys paint probes. Cytogenet CellGenet 92:248–253.
Polzin T, Daneschmand SV. 2003. On Steinertrees and minimum spanning trees inhypergraphs. Oper Res Lett 31:12–20.
Prouty LA, Buchanan PD, Pollitzer WS,Mootnick AR. 1983a. Bunopithecus: agenus-level taxon for the hoolock gibbon(Hylobates hoolock). Am J Primatol 5:83–87.
Prouty LA, Buchanan PD, Pollitzer WS,Mootnick AR. 1983b. A presumptive newhylobatid subgenus with 38 chromosomes.Cytogen Cell Genet 35:141–142.
Roos C. 2003. Molekulare Phylogenie derHalbaffen, Schlankaffen und Gibbons. PhDthesis. Germany: Technical Univ Munich.
Roos C. 2004. Molecular evolution and sys-tematics of Vietnamese primates. In: NadlerT, Streicher U, Ha Thang L, editors.Conservation of primates in Vietnam. Ha-noi: Frankfurt Zool Soc, Vietnam PrimateConservation Programme, Endangered Pri-mate Rescue Center, Cuc Phuong NationalPark. p 23–28.
Roos C, Geissmann T. 2001. Molecular phylo-geny of the major hylobatid divisions. MolPhylogen Evol 19:486–494.
Rosenblum LL, Supriatna J, Melnick DJ.1997. Phylogeographic analysis of pigtailmacaque populations (Macaca nemestrina)inferred from mitochondrial DNA. Amer JPhys Anthro 104:35–45.
Sheeran, LK, Zhang Y, Poirier FE, Yang D.1998. Preliminary report on the behavior ofthe Jingdong black gibbon (Hylobates con-color furvogaster). Trop Biodiv 5:113–125.
Simmons RE. 2005. A phylogenetic study ofthe gibbons (Hylobatidae). Master’s thesis.San Diego: University of California.
Simmons RE, Monda K, Woodruff DS. Phylo-genetics of gibbons (Hylobatidae): mito-chondrial and nuclear DNA sequence dataresolve long-standing questions. in prep.
Sterling EJ, Hurley MM, Minh LD. 2006.Vietnam: a natural history. New Haven:Yale University Press.
Su B, Kressirer P, Wang W, Jiang X, Wang YX,Woodruff DS, Monda K, Zhang YP. 1996.Molecular phylogeny of Chinese N. c.concolor gibbons. Sci China (C) 26:414–419.
Swofford DL. 1998. Phylogenetic analysisusing parsimony and other methods(PAUP� Version 4.0b). Sunderland, MA:Sinauer.
Takacs Z, Morales JC, Geissmann T, MelnickDJ. 2005. A complete species-level phylo-geny of the Hylobatidae based on mitochon-drial ND3-ND4 gene sequences. MolPhylogen Evol 36:456–467.
Tanaka H, Wijayanto H, Mootnick AR, Iskan-driati D, Perwitasari-Farajallah D, SajuthiD, Hirai H. 2004. Molecular phylogenetic
analyses of subspecific relationships in agilegibbons (Hylobates agilis) using mitochon-drial and TSPY gene sequences. FoliaPrimatol 75(Suppl 1):418.
Thalmann O, Hebler J, Poinar HN, Paabo S,Vigilant L. 2004. Unreliable mtDNA datadue to nuclear insertions: a cautionary talefrom analysis of humans and other greatapes. Mol Ecol 13:321–335.
Thompson JD, Gibson TJ, Plewniak F, Jean-mougin F, Higgins DG. 1997. The CLUS-TAL-X windows interface: Flexiblestrategies for multiple sequence alignmentaided by quality analysis tools. NucleicAcids Res 25:4876–4882.
Trung LQ, Hoang TD. 2004. Status review ofthe Cao Vit black-crested gibbon (Nomascusnasutus nasutus) in Vietnam. In: Nadler T,Streicher U, Ha Thang L, editors. Conserva-tion of primates in Vietnam. Hanoi: Frank-furt Zool Soc, Vietnam Primate ConservationProgr. Cuc Phuong National Park, Endan-gered Primate Rescue Center. p 90–94.
van Gulik RH. 1967. The gibbon in China. Anessay in Chinese animal lore. Leiden: Brill.
Wagele J-W. 2005. Foundations of phyloge-netic systematics. Munich: Pfeil.
Wang Y. 2003. A complete checklist of mam-mal species and subspecies in China—ataxonomic and geographic reference.Beijing: China Forestry Publishing House.394p.
Warren KS, Verschoor EJ, Langenhuijzen S,Swan RA, Vigilant L, Heeney JL. 2001.Speciation and intrasubspecific variation inBornean orangutans, Pongo pygmaeus pyg-maeus. Mol Biol Evol 18:472–480.
White MJD. 1978. Modes of speciation. SanFrancisco: Freeman.
Whittaker DJ. 2005. Evolutionary genetics ofKloss’s gibbons (Hylobates klossii):systematics, phylogeography, and conserva-tion. Ph.D. thesis. The City University ofNew York. 201p.
Woodruff DS. 1990. Genetics and demographyin the conservation of biodiversity. J Sci SocThailand 16:117–132.
Woodruff DS. 1993. Non-invasive genotypingof primates. Primates 34:333–346.
Woodruff DS. 2003. Noninvasive genotypingand field studies of free-ranging nonhumanprimates. In: Chapais B, Berman CM,editors. Kinship and behavior in primates.Oxford: Oxford University Press. p 46–68.
Woodruff DS, Monda K, Simmons RE. 2005.Mitochondrial DNA sequence variation andsubspecific taxonomy in the white-handedgibbon, Hylobates lar. Nat Hist J Chula-longkorn Univ 1:71–78.
Zehr SM. 1999. A nuclear and mitochondrialphylogeny of the lesser apes (Primates,genus Hylobates). Ph.D. thesis. HarvardUniversity. 221p.
1300 / Monda et al.
Am. J. Primatol. DOI 10.1002/ajp

Zhang Y. 1992. Hainan gibbon (Hylobates concolorhainanus) threatened. Asian Primates 2:6.
Zhang Y, Sheeran L. 1994. Current status ofthe Hainan black gibbon (Hylobates con-color hainanus). Asian Primates 3:3.
Zhang YP. 1997. Mitochondrial DNA sequenceevolution and phylogenetic relationships ofgibbons. Acta Genet Sin 24:231–237.
Zhang YW, Luo HR, Ryder OA, Zhang YP.2004. Evolution of the tandem repeats inthymidylate synthase enhancer region(TSER) in primates. Gene 338:47–54.
Zhou J, Wei F, Li M, Zhang J, Wang D, Pan R.2005. Hainan black-crested gibbon isheaded for extinction. Int J Primatol 26:453–465.
APPENDIX 1
Annotated list of currently recognized Nomascus species and subspecies
Nomascus concolor Harlan, 1826. Western black gibbon or black crestedgibbon. The tortuous taxonomic history of N. concolor is discussed by Geissmann[1989]. Our genetic data are in agreement with the cyt b study by C. Roos(unpublished) and support the recognition of this taxon as a full species, and donot support the recognition of three parapatric subspecies—concolor, jingdon-gensis, furvogaster. Endangered [IUCN, 2006] with less than 2,000 individuals inChina, Vietnam and Laos. Recent reports include Bartlett [2005], Bleisch andZhang [2004], Sheeran et al. [1998], Nadler and Streicher [2004].
Nomascus concolor concolor Harlan, 1826. Black gibbon or Tonkin blackcrested gibbon. South China (eastern Yunnan) and northwest Vietnam, betweenthe Black (Song Da) and Red (Song Hong) rivers. Range historically parapatricwith N. c. jingdongensis, N. nasutus, and N. leucogenys [Geissmann et al., 2000;Nadler & Streicher, 2004], but interactions with those taxa are undocumentedand probably no longer occur.
N. concolor jingdongensis Ma and Wang, 1986. Black gibbon or centralYunnan black crested gibbon. South China (central Yunnan) between the Mekongand Black rivers, Wenbu, Jingdong, and Wuliang Mountains [Bleisch & Jiang,2000; Jiang & Wang, 1999; Sheeran et al., 1998]. Extirpated from most of range[Geissmann et al., 2000, 2006]. Our genetic data support subjugation withinN. concolor, a view supported by Brandon-Jones et al. [2004], Geissmann et al.[2000] and C. Roos (unpublished), as the diagnostic features involve only minorcolor differences in females.
Nomascus concolor furvogaster Ma and Wang, 1986. Black gibbon or westYunnan black crested gibbon. South China (western Yunnan), between theMekong and Salween rivers. [Ma & Wang, 1986; Ma et al., 1988]. Geneticdata reported here support subjugation within N. concolor, a view supportedby Geissmann [1995], Geissmann et al. [2000] as the diagnostic features werebased on a subadult female. Geissmann et al. [2006] provide limited recentobservations.
Nomascus concolor lu Delacour, 1951. Loatian black crested gibbon. BokeoProvince, Laos, westernmost Laos at Ban Nam Khueng and Khao Tham Phra onthe Mekong river and Nam Kan valley at about 201N, where Geissmann recentlyconfirmed their presence and identity [Geissmann et al., 2000; see also Mootnick,2006]. This is an isolated allopatric population of a few hundred individualsseparated today from the main population of N. concolor by 250 km occupied byN. leucogenys. Probably a synonym of N. c. concolor with minor differencespossibly a result of limited hybridization with N. leucogenys [Geissmann, 1989].Until genetic data are available to test this hypothesis the subspecies should berecognized; preliminary genetic data support subjugation within N. concolor[C. Roos, unpublished].
Nomascus gibbon phylogeny / 1301
Am. J. Primatol. DOI 10.1002/ajp

Nomascus nasutus Kunckel d’Herculais, 1884. Eastern black crested gibbon.Northeast Vietnam, north and east of the Red River delta and the adjacent coastalChina. Largely extirpated, 20–28 gibbons were re-discovered in Cao Bang and HoaBinh Provinces in 2002 [Bleisch & Zhang, 2004; Geissmann et al., 2000; Trung &Hoang, 2004]. Critically Endangered [IUCN, 2004; Konstant et al., 2003]. Thistaxon was referred to awkwardly for many years as Hylobates (Nomascus) sp. cf.nasutus because of uncertainties over the source of the type specimen (see notesin Appendix 2) and its relation to N. concolor [see Geissmann, 1989, 2002].Genetically, we have shown that it is clearly specifically distinct from N. concolor,a conclusion reached independently by Takacs et al. [2005]. Until recently onlyone captive animal (Patzi, see Appendix 2) had been examined and herrelationship to the remaining wild gibbons had not been established. Patzi wasunusual in looking like a concolor gibbon but having a distinctive hainanus-likevocalization [Geissmann, 1989, 1997] and she may have been a hybrid or anunrecognized taxon [Geissmann et al., 2002; Groves, 2004]. However, Roos (inlitt. April 2006) has just genotyped apes from both surviving Vietnamesepopulations and found them to be very closely related to Patzi at the cyt bsequence studied. Furthermore, as reported in the Results above, he discoveredthe mainland gibbons were significantly different from Hainan Island animals atthis locus and accordingly proposes to elevate nasutus to species rank [Roos, inprep.]; a decision anticipated by Nadler and Streicher [2004].
N. hainanus Thomas 1892. Hainan black crested gibbon. The population isolatedtoday on Hainan Island, China, is now restricted to the Bawangling Nature Reserve[Zhang, 1992; Zhang & Sheeran, 1994]. Dao [1983] thought the island gibbons weresubspecifically distinct from nasutus and several museum specimens in Vietnamesecollections are referred to the subspecies N. n. hainanus. Roos’ (in prep, in litt. toDSW) cyt b genetic sequence divergence data indicate, however, that although relatedto nasutus, hainanus merits full species level status. It is unknown when gibbons firstarrived on the continental island of Hainan but they could have dispersed acrossbroad (50–70 km wide) dry land connections during one or more Pleistocenehypothermal phase(s). Today’s population has been physically isolated for less than10,000 years, since the sea rose above –25 m and flooded the narrow Qiongzhou Strait.Their genetic divergence from nasutus suggests they have been separated for muchlonger and Chatterjee [2006] estimates the species antiquity to be 0.3–1.8 My. Theidentity of mainland gibbons referred to this species cannot be accepted withoutgenetic confirmation. Groves [2001] and Wang [2003] treat hainanus as a full species,without comment. Now critically endangered, this population fell from an estimated2,000 animals in the 1950s, to 23 in 1998, and 13 in 2004 [Bleisch & Zhang, 2004;Chan et al., 2005; Chivers, 2005; Geissmann et al., 2000; Zhou et al., 2005].
N. leucogenys Ogilby, 1840. Northern white-cheeked crested gibbon.Historical range: China (Mengla county, Xishuangbanna, southern Yunnan),northern Laos, and northwestern Vietnam [Fooden et al., 1987; Geissmann et al.,2000]. Corbet and Hill [1992] reported it is sympatric with N. concolor but thisseems unlikely as the historical interactions with other taxa are undocumented.IUCN [2006]: data deficient. Endangered in Vietnam and o100 individuals inChina [Bleisch & Zhang, 2004; Geissmann et al., 2000; Nadler & Streicher, 2004].The southern populations are referred to the subspecies N. l. siki.
N. leucogenys siki Delacour, 1951. Southern white-cheeked crested gibbon.Southern Laos, central Vietnam. During the last 15 years various authors haveincluded siki as either a subspecies of N. gabriellae [Corbet & Hill, 1992; Groves &Wang, 1990], or N. leucogenys [Geissmann, 1995], or treated it as a separatespecies [Groves, 2001, 2004; Zhang, 1997]. The size and shape of the white cheek
1302 / Monda et al.
Am. J. Primatol. DOI 10.1002/ajp

patches of adult males and juveniles are diagnostic features for siki, but adultfemales of N. leucogenys and siki are morphologically indistinguishable althoughboth differ from females of N. gabriellae [Geissmann, 1995; Geissmann et al.,2000; Mootnick, 2006]. The songs of the three taxa are different [Konrad &Geissmann, 2006] and siki resembles N. leucogenys more than that of any otherform of crested gibbon including N. gabriellae [Konrad & Geissmann, 2006]. Theboundary between N. leucogenys and siki lies near the lower Ca River south of thetown of Vinh and east of the Annamite Mountains in Nghe An Province, Vietnam.Further south, N. l. siki is replaced by N. gabriellae, but the southern range limitsof N. l. siki are undocumented. Konrad and Geissmann [2006] have describedvocalizations in northeast Cambodia (Rattanakiri) and provisionally assign thoseapes to N. l. siki because of their resemblance to calls of topotypic siki from nearbyBach Ma, Vietnam. They postulate that a taxon boundary exists somewherebetween Rattanakiri and southern Mondulkiri in eastern Cambodia, and discussthe roles of the Srepok river and dry dipterocarp forest as possible distributionbarriers. Our mitochondrial DNA sequences suggest that siki is more closelyrelated to N. leucogenys than to N. gabriellae [Garza & Woodruff, 1992, 1994;Zhang, 1997; herein] but we recognize that a couple of specimens reported byothers have the opposite affinity; we regard the problem as unresolved untilanimals of known geographic provenance are genotyped. Konrad and Geissmann[2006] discuss the possibility that siki and gabriellae are separated by a broad zoneof intergradation (hybridization) or, alternatively, that a currently unrecognizedtaxon occupies a large area of south central Vietnam between their ranges.
N. gabriellae Thomas, 1909. Yellow-cheeked (or buff-cheeked) crestedgibbon. Eastern Cambodia, southern Laos, and southern Vietnam, south of151300. Historically thought to be parapatric or hybridize with siki, but theirinteractions are poorly documented and the two species are allopatric inCambodia today [Baker et al., 2002; Geissmann et al., 2000; Konrad &Geissmann, 2006]. Endangered.
APPENDIX 2
Specimens examined
North American zoo gibbons used in this study
All taxonomic identifications are those provided by the owners and are thesame as those recorded in the AZA concolor studbook [1990] and ISIS. For eachindividual the following data are provided: [the ID number used in Figures in thispaper in square brackets], Studbook number/ISIS number, Sex, House name,Holding Zoo, Origin, GenBank accession number, and no. of base pairs (bp) ofmtDNA sequenced.
Notes: Studbook number: two individuals had no studbook number (]nsb).Sex and House name: male or female and name (or not available, na). HoldingZoo: at the time hair sample was provided, typically 1984–5. Origin: captive born(cb) or wild born (wb); country or zoo of origin or unknown.
Nomascus leucogenys (24 individuals)[leuc 1] 11/32055C, F Betsy, National, wb, unknown, EF203867, 497 bp.
[leuc 2] 18/92, F na, Minnesota, wb unknown, EF203868, 530 bp. [leuc 3]21/1345, F Muneca, Gladys Porter, wb unknown, EF203869, 486 bp. [leuc 4]24/36233, M Joe, National, wb unknown, EF203870, 510 bp. [leuc 5] 28/36336, F
Nomascus gibbon phylogeny / 1303
Am. J. Primatol. DOI 10.1002/ajp

Beryl National, wb unknown, EF203871, 497 bp. [leuc 6] 30/92, F China, GladysPorter, wb unknown, EF203872, 485 bp. [leuc 7] 31/93, F Goldie, Gladys PorterZoo, wb unknown, EF203873, 480 bp. [leuc 8] 42/1318, F Phyllis, WashingtonPark, wb unknown, EF203874, 285 bp. [leuc 9] 43/101675, F Mae, National, wbunknown, EF203875, 521 bp. [leuc 10] 53/101519, M Gilly, Cheyenne, wbunknown, EF203876, 300 bp. [leuc 11] 55/91, M Archie, Minnesota, wb unknown,EF203877, 300 bp. [leuc 12] 196/107858, M Mekong, National, cb National,EF203878, 485 bp. [leuc 13] 213/5926, F Minnesota, cb Minnesota, EF203879,513 bp. [leuc 14] 229/109732, F Melaka, National, cb National, EF203880, 176 bp.
Comparable sequences were obtained from the following individuals butare not shown here as they are subsumed within the clusters revealed bythe above animals [Monda, 1995; Woodruff et al., in prep.]. Some are thedescendants of females listed above and were found to have identical mtDNA,as expected.
1 nsb/110265, M, Beryl’s baby, National, unknown, 474 bp. 2 nsb/91200, ?F,Hue, Washington Park, cb Washington Park 285 bp. 14/1325, M, Gunther,Washington Park, wb unknown. 79/100739, M, Bert, National, cb, National,521 bp. 134/105020, M, Ralph, National, cb National, son of 28 and sib of 1 nsb.143/107869, F, Siam, National, cb, National, 499 bp. 146/2044, F, Deborah, GladysPorter, cb unknown, 536 bp. 207/108291, F, Burma, National, cb National,daughter of 28 and sib of 1 nsb. 209/88071, M, Tanh Linh, Washington Park, cbWashington Park, 291 bp. 231/109740, F, Sena, National, cb National, 474 bp.
Nomascus gabriellae (2)[gabr 1], 122/94241, F Robin, Los Angeles, cb Los Angeles, daughter of 65/
94111, F Bahmetoo, Los Angeles, wb S. Vietnam and 64/94110, M Koo, LosAngeles, wb S. Vietnam, EF203886, 492 bp. [gabr 2], 180/95141, M YangMenggangu, Los Angeles, cb Los Angeles, sib of 122, EF203887, 492 bp.
Symphalangus syndactylus (1)[Symphalangus], 267/870020, F, Juice, Cheyene Mountain Zoo, cb Cheyene,
EF203866, 385 bp.
Samples studied by Kressirer [1993]
Gibbons were adults unless noted and all identifications were provided by theowners. For each individual the following data are provided: [the ID number usedin this paper in square brackets], Kressirer [1993] specimen no., Sex, Housename, Holding Zoo or Museum, mtDNA source, GenBank accession number.
N. l. leucogenys[leuc 15] 4 M juvenile (parents: Jack & Jacqueline), Mulhouse Zoo, tissue
sample 1986, EF203883, 476 bp. [leuc 16] 280 M Claude, Mulhouse Zoo, bloodsample 1986, EF203881, 476 bp. Used by Kressirer [1993] to represent this taxonand identified as P3leuc in some analyses by him, and subsequently by Monda[1995] and Simmons [2005]. [leuc 17] 281 M Jack, Mulhouse Zoo, blood sample1986, EF203882, 476 bp. [leuc 18] 342 M Dodo, Budapest Zoo, arrived fromsoutheast Asia in 1968, hair sample 1991, EF203884, 476 bp.
N. l. siki[siki 1] 20 M Charly, Hellabrunn Zoo, Munich, hair sample in 1991,
EF203885, 495 bp. Used by Kressirer [1993] to represent this taxon and identified
1304 / Monda et al.
Am. J. Primatol. DOI 10.1002/ajp

as by him as P1siki in some of his analyses, and subsequently by Monda [1995]and Simmons [2005].
Identical but shorter sequences were obtained from the following individualsbut are not used here: 21 F Charlotte, Hellabrunn Zoo, Munich, hair sample in1991; 22 F Mimi, Hellabrunn Zoo, Munich, hair sample in 1991.
N. gabriellae[gabr 3] 340 M Tschico, Budapest Zoo, arrived from Laos in 1987, hair
sample 1991, EF203888, 407 bp. Used by Kressirer [1993] to represent this taxonand identified as by him as P2gabriellae in some of his analyses, and subsequentlyby Monda [1995] and Simmons [2005].
An identical but shorter sequence was obtained from the following individualbut is not used here: 341 F Juschka, Budapest Zoo, transferred from Moscow Zoo1991, born ca. 1968 in Vietnam, hair sample 1991.
N. nasutus[nasu 1] 409/410 F Patzi, Humboldt University Museum, ZMB 70036, from
Tierpark Berlin (1962–1986), reported to have been shipped from Hon Gai(north-eastern Vietnam) but geographic origin unknown, skin sample, EF203889,446 bp. This individual is discussed in detail by Geissmann [1989] who regardsis as sufficiently different in coloration from the other known northeasternVietnamese black gibbons to warrant subspecific status. Used by Kressirer[1993] to represent this taxon and identified as by him as P4nasutus in someof his analyses, and subsequently by Monda [1995], Su et al. [1996] and Simmons[2005]. [nasu 2] Kressirer ]0.2, M, Museum Naturkunde, Berlin, 85357, captiveanimal labeled as N. n. hainanus from ‘‘Houchow (?), China’’, EF203890,519 bp. Our efforts to establish the source locality of this specimen havefailed; there is no Houchou on Hainan Island and there is a reasonable possibilitythat this animal was actually from the mainland as an anonymous reviewer ofthis manuscript has kindly drawn our attention to a locality named Hou Chau (orKou Chau) at 221380 N, 104150 E in Vietnam. Used by Kressirer [1993] torepresent this taxon and identified as by him as P5hainanus in some of hisanalyses, and subsequently by Monda [1995], Su et al. [1996] and Simmons[2005]. We now believe this specimen was misidentified and report it hereinas N. nasutus.
Partial sequences obtained by Su et al. [1996]
For each individual the following data are provided: [ID number in thispaper] specimen identification ] in Su et al. [1996], sex, collection locality, mtDNAsource, collection year, GenBank accession number. Skins are in the Museum ofthe Kunming Institute of Zoology unless noted. Comparable sequences were143 bp, and all shared a 16 bp deletion relative to Symphalangus.
[leuc 19] N. leucogenys-1, M, Hekou, Yunnan, fresh hair sampled, 1994,EF212884. [leuc 20] N. leucogenys-2, M, Hekou, Yunnan, fresh hair sampled,1994, EF212885.
[jing 1] N. concolor jingdongensis-1, F, Jingdong, Yunnan, collected 1964,old skin, EF203891. [jing 2] N. c. jingdongensis-2, M, Jingdong, Yunnan,collected 1964 old skin, EF212886. [jing 3] N. c. jingdongensis-3, M, Jingdong,Yunnan, collected 1964 old skin, EF212887. [jing 4] N. c. jingdongensis-4, F,Jingdong, Yunnan, old skin, details unknown, EF212888.
Nomascus gibbon phylogeny / 1305
Am. J. Primatol. DOI 10.1002/ajp

[furv 1] N. c. furvogaster, M, Cangyuan, Yunnan, collected 1983, old skin,EF212889.
[conc 1] N. c. concolor-1, sex unknown, Jianshui, Yunnan, collected 1987 oldskin, EF203893. [conc 2] N. c. concolor-2, F, Luchum, Yunnan, collected 1972, oldskin, EF203894. [conc 3] N. c. concolor-3, F Jianshui, Yunnan, collected 1989, oldskin, EF203892.
[nasu 2] N. nasutus hainanus is Kressirer [1993] P5hainanus: ]0.2, M,Museum Naturkunde, Berlin, 85357, captive animal from ‘‘Houchow (?), China’’,and now thought to be N. nasutus from Vietnam (see above).
[Symphalangus] Symphalangus syndactylus is Monda’s ]267/870020, F,Juice, Cheyene Mountain Zoo, cb Cheyene.
Outgroup sequences from GenBank
An additional four sequences were used as outgroups in some analyses:[Hoolock], Hoolock (formerly Bunopithecus) hoolock (AF311725)[Roos andGeissmann]; [numt] a S. syndactylus nuclear insertion (numt) of the HVR-1(AF035467); [Pongo] Pongo pygmaeus (D38115); [Homo] Homo sapiens(NC001807).
1306 / Monda et al.
Am. J. Primatol. DOI 10.1002/ajp


















