
Microfilaments in the cells trabecular meshwork - bjo.bmj.com · fromthe perinuclear zoneto the...
6
British Journal of Ophthalmology, 1979, 63. 3-8 Microfilaments in the cells of the human trabecular meshwork I. GRIERSON AND A. H. S. RAHI From the Department of Pathology, Institute of Ophthalmology, London SUMMARY In this present study the results are presented of a combined ultrastructural and immunofluorescent investigation of 'smooth muscle' elements within the cytoplasm of human trabecular meshwork cells. The cytoplasm of human meshwork cells both in vivo and in vitro is replete with 10 nm intermediate filaments and also contains smaller 6 nm filaments which are particularly prominent in the cell processes. By immunofluorescence using sera rich in antibodies to contractile proteins, particularly actin, cultured meshwork cells showed strong cytoplasmic fluorescence. On occasion the cytoplasmic fluorescence was diffuse, but more often recognisable bundles (stress fibres) or a loose fibrillar framework was found. The possible role of structural and contractile cellular proteins in trabecular function was discussed. It has been shown that trabecular meshwork cells are extremely pliable, and in experimentally induced acute increase of intraocular pressure (Grierson and Lee, 1975; Kayes, 1975; Svedbergh, 1976) or after the application of topical pilocarpine (Grierson et al., 1978) these cells are capable of pronounced attenuation without loss of cell-to-cell contact. Grierson and Lee (1975) suggested that cytoskeletal elements and, in particular, the cytoplasmic microfilaments could be expected to have a key role in cellular pliability, and thus they will have an important function in the maintenance of tissue integrity. Further, it is known that the structural cytoplasmic proteins of many non-muscle cells, in addition to having cytoskeletal functions, can play a part in cellular mobility and contraction (Pollard and Weihing, 1974). If contractile proteins are a com- ponent of the cytoplasm of trabecular meshwork cells, it could be expected that filament-dependent cellular contraction may have an influence on outflow dynamics (Inomata et al., 1972). Indeed it has been shown that the intraocular infusion of cytochalasin B, which disrupts contractile cyto- plasmic microfilament systems, produces a marked increase in the facility of aqueous outflow (Kaugh- man and Barany, 1977). Clearly structural and contractile cytoplasmic proteins may be of some importance to trabecular Address for reprints: Dr I. Grierson, Department of Pathology, Institute of Ophthalmology, Judd Street, London WCIH 9QS 3 function. Therefore the present investigation was conducted to determine the nature and distribution of cytoplasmic microfilaments in the cells of the normal human trabecular meshwork, and an attempt was made to demonstrate the presence of contractile protein antigens within cultured human trabecular meshwork cells by means of an immuno- fluorescent technique. Materials and methods NORMAL TISSUE Normal angular tissue for electron microscopy was obtained from 11 human eyes enucleated in treat- ment of malignant melanoma of the posterior globe. The patients' ages varied between 46 and 76 years. The eyes showed no clinical evidence of abnormalities in the anterior segment. After enucleation they were immersed in 3% phosphate buffered glutaraldehyde and opened by equatorial section; the lenses were removed, and slices of limbal tissue were taken for further processing. After a wash in phosphate buffer the limbal segments were post-fixed in phosphate buffered osmium tetroxide, rewashed in buffer, dehydrated through graded alcohols, cleared with propylene oxide, and embedded in Araldite. TISSUE CULTURE The globes for tissue culture were provided by the eye bank and were used from 6 to 48 hours post mortem. The anterior segment of each eye was divided into quadrants, and the iris and ciliary body were removed under the dissecting microscope. Two on 9 February 2019 by guest. Protected by copyright. http://bjo.bmj.com/ Br J Ophthalmol: first published as 10.1136/bjo.63.1.3 on 1 January 1979. Downloaded from
Transcript of Microfilaments in the cells trabecular meshwork - bjo.bmj.com · fromthe perinuclear zoneto the...

British Journal of Ophthalmology, 1979, 63. 3-8
Microfilaments in the cells of the humantrabecular meshworkI. GRIERSON AND A. H. S. RAHIFrom the Department of Pathology, Institute of Ophthalmology, London
SUMMARY In this present study the results are presented of a combined ultrastructural andimmunofluorescent investigation of 'smooth muscle' elements within the cytoplasm of humantrabecular meshwork cells. The cytoplasm of human meshwork cells both in vivo and in vitro isreplete with 10 nm intermediate filaments and also contains smaller 6 nm filaments which are
particularly prominent in the cell processes. By immunofluorescence using sera rich in antibodiesto contractile proteins, particularly actin, cultured meshwork cells showed strong cytoplasmicfluorescence. On occasion the cytoplasmic fluorescence was diffuse, but more often recognisablebundles (stress fibres) or a loose fibrillar framework was found. The possible role of structuraland contractile cellular proteins in trabecular function was discussed.
It has been shown that trabecular meshwork cellsare extremely pliable, and in experimentally inducedacute increase of intraocular pressure (Griersonand Lee, 1975; Kayes, 1975; Svedbergh, 1976) orafter the application of topical pilocarpine (Griersonet al., 1978) these cells are capable of pronouncedattenuation without loss of cell-to-cell contact.Grierson and Lee (1975) suggested that cytoskeletalelements and, in particular, the cytoplasmicmicrofilaments could be expected to have a keyrole in cellular pliability, and thus they will havean important function in the maintenance of tissueintegrity.
Further, it is known that the structural cytoplasmicproteins of many non-muscle cells, in addition tohaving cytoskeletal functions, can play a part incellular mobility and contraction (Pollard andWeihing, 1974). If contractile proteins are a com-ponent of the cytoplasm of trabecular meshworkcells, it could be expected that filament-dependentcellular contraction may have an influence onoutflow dynamics (Inomata et al., 1972). Indeed ithas been shown that the intraocular infusion ofcytochalasin B, which disrupts contractile cyto-plasmic microfilament systems, produces a markedincrease in the facility of aqueous outflow (Kaugh-man and Barany, 1977).
Clearly structural and contractile cytoplasmicproteins may be of some importance to trabecular
Address for reprints: Dr I. Grierson, Department ofPathology, Institute of Ophthalmology, Judd Street, LondonWCIH 9QS
3
function. Therefore the present investigation wasconducted to determine the nature and distributionof cytoplasmic microfilaments in the cells of thenormal human trabecular meshwork, and anattempt was made to demonstrate the presence ofcontractile protein antigens within cultured humantrabecular meshwork cells by means of an immuno-fluorescent technique.
Materials and methods
NORMAL TISSUENormal angular tissue for electron microscopy wasobtained from 11 human eyes enucleated in treat-ment of malignant melanoma of the posterior globe.The patients' ages varied between 46 and 76 years.The eyes showed no clinical evidence of abnormalitiesin the anterior segment. After enucleation they wereimmersed in 3% phosphate buffered glutaraldehydeand opened by equatorial section; the lenses wereremoved, and slices of limbal tissue were taken forfurther processing. After a wash in phosphate bufferthe limbal segments were post-fixed in phosphatebuffered osmium tetroxide, rewashed in buffer,dehydrated through graded alcohols, cleared withpropylene oxide, and embedded in Araldite.
TISSUE CULTUREThe globes for tissue culture were provided by theeye bank and were used from 6 to 48 hours postmortem. The anterior segment of each eye wasdivided into quadrants, and the iris and ciliary bodywere removed under the dissecting microscope. Two
on 9 February 2019 by guest. P
rotected by copyright.http://bjo.bm
j.com/
Br J O
phthalmol: first published as 10.1136/bjo.63.1.3 on 1 January 1979. D
ownloaded from

l. Grierson and A. H. S. Rahi
parallel cuts were made in the trabecular region,and the strip of drainage tissue was carefullyremoved with minimal handling. The trabecularmeshes were transferred to the cover glass of a tissueculture chamber and embedded in an embryonicplasma clot. The chamber was then sealed andfilled with a tissue culture medium. For furtherdetails see Tripathi et al. (1973).Outgrowths from the explant spread over the
surface of the coverslip, and after 6 weeks to 3 monthsthe primary culture was sufficiently well establishedfor either electron microscopy or immunofluorescentstudies. Preparation of the trabecular meshwork cellson the coverslips for electron microscopy was bythe technique described by Tripathi et al. (1973) andRahi and Ashton (1978).
IMMUNOFLUORESCENT STUDIESFor this part of the investigation the coverslipscontaining monolayers of cultured trabecularmeshwork cells were washed in isotonic saline andfixed in acetone.Blood from patients with chronic uveitis or
intraocular melanoma containing high titres (greaterthan 1/160) of antibodies to 'smooth muscle'proteins, particularly actin (Rahi et al., 1976; Rahiand Ashton, 1978) were used as the source ofantiserum to localised contractile cytoplasmicproteins. The reactivity and specificity of the serawere judged by examination of frozen sections fromcomposite blocks of rat liver, kidney, skeletalmuscle, stomach, and salivary gland which had beentreated with the sera (Trenchev and Holborow, 1976).
After treatment with the appropriate sera themethod outlined by Rahi et al. (1976) and Rahi and
Ashton (1978) using fluorescein labelled antihumanimmunoglobulins was adopted. A Zeiss epifluores-cence microscope equipped with a fluoresceinisothiocyanate interference filter was used forroutine examination of the tissues.
Results
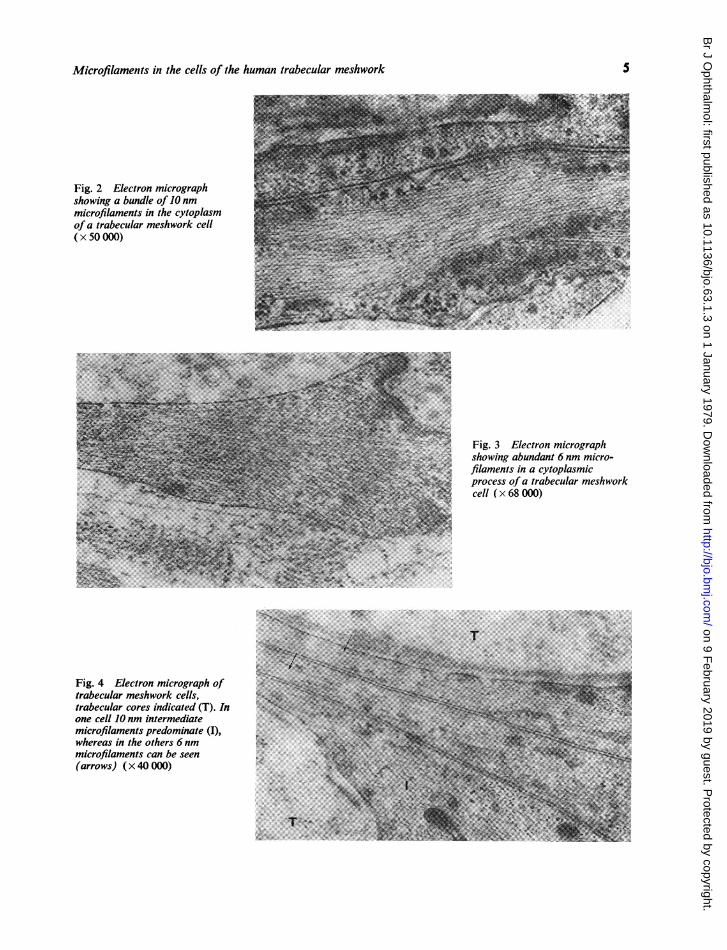
ULTRASTRUCTUREThe trabecular meshwork cells of the normalhuman outflow apparatus contained 2 types ofcytoplasmic microfilaments of indeterminate lengthbut of different diameters. The larger filamentsmeasured 10 nm in diameter and were most abun-dant in the main cell body and in the perinuclearzone (Fig. 1). The 10 nm filaments were formed intoeither sporadically distributed bundles (Fig. 2) or,more commonly, a subcortical network, so that inany one section the filaments were cut both in crosssection and in longitudinal section (Fig. 1). Thesefilaments did not make contact with the plasmamembrane but did come into close association withthe limiting membranes of some cytoplasmicorganelles (Fig. 1).The finer, more delicate filaments measured
between 5 and 6 nm in diameter and were foundin loose aggregates throughout the cell cytoplasmfrom the perinuclear zone to the cell periphery. Thefine filaments were particularly prevalent in the cellprocesses, where they were orientated so that theyformed an array which ran parallel with the longaxis of the cytoplasmic projection (Fig. 3). Unlikethe larger filaments the fine filaments came intoclose proximity with the plasma membrane (Figs.3 and 4).
Fig. 1 Electron micrographshowing 10 nm microfilaments inthe cytoplasm of a trabecularmeshwork cell. The micro-filaments form a network and canbe seen both in cross and inlongitudinal section ( x 60 000)
4
on 9 February 2019 by guest. P
rotected by copyright.http://bjo.bm
j.com/
Br J O
phthalmol: first published as 10.1136/bjo.63.1.3 on 1 January 1979. D
ownloaded from
Microfilaments in the cells of the human trabecular meshwork
Fig. 2 Electron micrographshowing a bundle of 10 nmmicrofilaments in the cytoplasmofa trabecular meshwork cell(x 50 000)
Fig. 3 Electron micrographshowing abundant 6 nm micro-filaments in a cytoplasmicprocess of a trabecular meshworkcell ( x 68 000)
...24|i '-.......As>,..... ..Z.,|:. . . .. ,r .X N.k ~~~~~~...... ......
Fig. 4 Electron micrograph oftrabecular meshwork cells,trabecular cores indicated (T). Inone cell 10 nm intermediatemicrofilaments predominate (I),whereas in the others 6 nmmicrofilaments can be seen(arrows) ( x 40 000)
5
on 9 February 2019 by guest. P
rotected by copyright.http://bjo.bm
j.com/
Br J O
phthalmol: first published as 10.1136/bjo.63.1.3 on 1 January 1979. D
ownloaded from
I. Grierson and A. H. S. Rahi
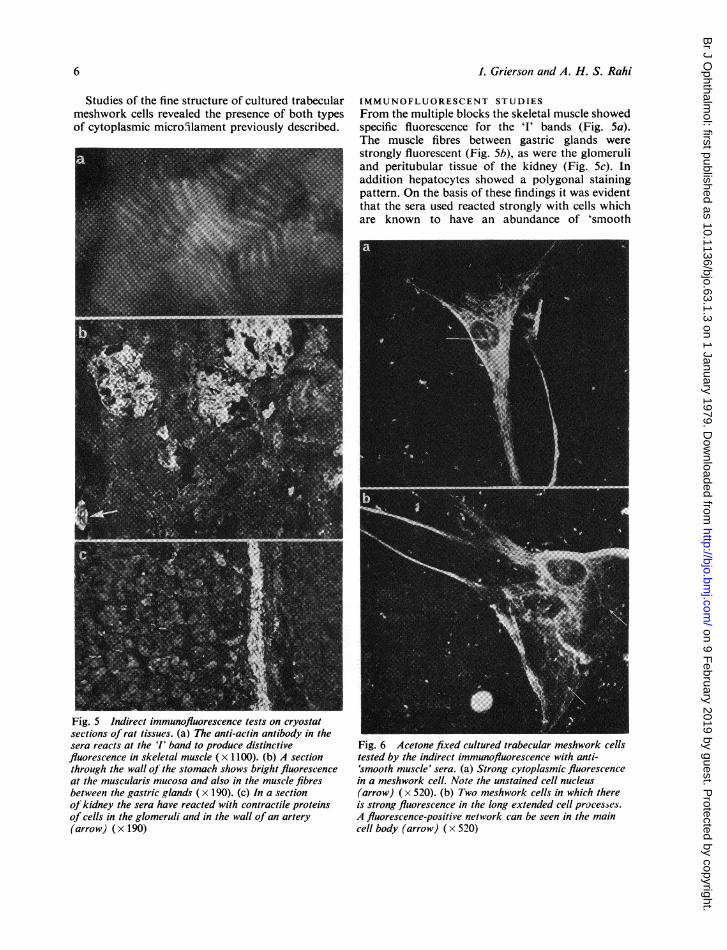
Studies of the fine structure of cultured trabecularmeshwork cells revealed the presence of both typesof cytoplasmic micro.ilament previously described.
Fig. 5 Indirect immunofluorescence tests on cryostatsections of rat tissues. (a) The anti-actin antibody in thesera reacts at the 'I' band to produce distinctivefluorescence in skeletal muscle ( x 1100). (b) A sectionthrough the wall oJ the stomach shows bright fluorescenceat the muscularis mucosa and also in the muscle fibresbetween the gastric glands (x 190). (c) In a sectionof kidney the sera have reacted with contractile proteinsof cells in the glomeruli and in the wall of an artery(arrow) (x 190)
IMMUNOFLUORESCENT STUDIESFrom the multiple blocks the skeletal muscle showedspecific fluorescence for the '1' bands (Fig. 5a).The muscle fibres between gastric glands werestrongly fluorescent (Fig. 5b), as were the glomeruliand peritubular tissue of the kidney (Fig. 5c). Inaddition hepatocytes showed a polygonal stainingpattern. On the basis of these findings it was evidentthat the sera used reacted strongly with cells whichare known to have an abundance of 'smooth
Fig. 6 Acetone fixed cultured trabecular meshwork cellstested by the indirect immunofluorescence with anti-'smooth muscle' sera. (a) Strong cytoplasmic fluorescencein a meshwork cell. Note the unstained cell nucleus(arrow) (x 520). (b) Two meshwork cells in which thereis strong fluorescence in the long extended cell processes.A fluorescence-positive network can be seen in the maincell body (arrow) ( x 520)
6
on 9 February 2019 by guest. P
rotected by copyright.http://bjo.bm
j.com/
Br J O
phthalmol: first published as 10.1136/bjo.63.1.3 on 1 January 1979. D
ownloaded from
Microfilaments in the cells of the human trabecular meshwork
muscle' proteins and to be particularly rich in actin.When cultured trabecular meshwork cells were
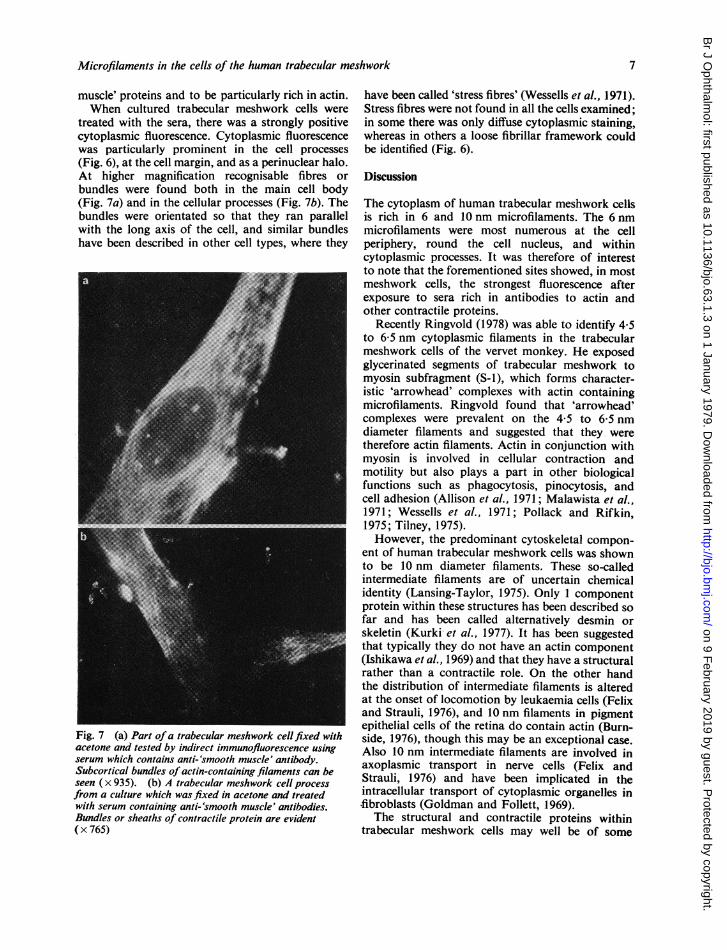
treated with the sera, there was a strongly positivecytoplasmic fluorescence. Cytoplasmic fluorescencewas particularly prominent in the cell processes(Fig. 6), at the cell margin, and as a perinuclear halo.At higher magnification recognisable fibres orbundles were found both in the main cell body(Fig. 7a) and in the cellular processes (Fig. 7b). Thebundles were orientated so that they ran parallelwith the long axis of the cell, and similar bundleshave been described in other cell types, where they
Fig. 7 (a) Part ofa trabecular meshwork cellfixed withacetone and tested by indirect immunofluorescence usingserum which contains anti-'smooth muscle' antibody.Subcortical bundles of actin-containing filaments can beseen ( x 935). (b) A trabecular meshwork cell processfrom a culture which was fixed in acetone and treatedwith serum containing anti-'smooth muscle' antibodies.Bundles or sheaths of contractile protein are evident( x 765)
have been called 'stress fibres' (Wessells et al., 1971).Stress fibres were not found in all the cells examined;in some there was only diffuse cytoplasmic staining,whereas in others a loose fibrillar framework couldbe identified (Fig. 6).
Discussion
The cytoplasm of human trabecular meshwork cellsis rich in 6 and 10 nm microfilaments. The 6 nmmicrofilaments were most numerous at the cellperiphery, round the cell nucleus, and withincytoplasmic processes. It was therefore of interestto note that the forementioned sites showed, in mostmeshwork cells, the strongest fluorescence afterexposure to sera rich in antibodies to actin andother contractile proteins.
Recently Ringvold (1978) was able to identify 4-5to 6 5 nm cytoplasmic filaments in the trabecularmeshwork cells of the vervet monkey. He exposedglycerinated segments of trabecular meshwork tomyosin subfragment (S-1), which forms character-istic 'arrowhead' complexes with actin containingmicrofilaments. Ringvold found that 'arrowhead'complexes were prevalent on the 4-5 to 6-5 nmdiameter filaments and suggested that they weretherefore actin filaments. Actin in conjunction withmyosin is involved in cellular contraction andmotility but also plays a part in other biologicalfunctions such as phagocytosis, pinocytosis, andcell adhesion (Allison et al., 1971; Malawista et al.,1971; Wessells et al., 1971; Pollack and Rifkin,1975; Tilney, 1975).However, the predominant cytoskeletal compon-
ent of human trabecular meshwork cells was shownto be 10 nm diameter filaments. These so-calledintermediate filaments are of uncertain chemicalidentity (Lansing-Taylor, 1975). Only 1 componentprotein within these structures has been described sofar and has been called alternatively desmin orskeletin (Kurki et al., 1977). It has been suggestedthat typically they do not have an actin component(Ishikawa et al., 1969) and that they have a structuralrather than a contractile role. On the other handthe distribution of intermediate filaments is alteredat the onset of locomotion by leukaemia cells (Felixand Strauli, 1976), and 10 nm filaments in pigmentepithelial cells of the retina do contain actin (Burn-side, 1976), though this may be an exceptional case.Also 10 nm intermediate filaments are involved inaxoplasmic transport in nerve cells (Felix andStrauli, 1976) and have been implicated in theintracellular transport of cytoplasmic organelles in-fibroblasts (Goldman and Follett, 1969).
The structural and contractile proteins withintrabecular meshwork cells may well be of some
7
on 9 February 2019 by guest. P
rotected by copyright.http://bjo.bm
j.com/
Br J O
phthalmol: first published as 10.1136/bjo.63.1.3 on 1 January 1979. D
ownloaded from

1. Grierson and A. H. S. Rahi
importance to trabecular function, since trabecularcell pliability and contraction may influence thewidth of intertrabecular spaces and hence affect therate of aqueous outflow (Kaughman, 1977). How-ever, it is also known that trabecular meshwork cellsare latent phagocytes, and in various forms ofsecondary open-angle glaucoma the native cells ofthe drainage system engulf considerable quantitiesof debris (Grierson and Lee, 1973; Van Buskirk andLeure-Dupree, 1978). Recently the suggestion hasbeen put forward that primary open-angle glaucomamay arise as a result of an inability of meshworkcells to phagocytose and get rid of the debris whichmay accumulate in the outflow system of the oldereye (Bill, 1975). If this interesting hypothesis is to betested, clearly it would be worthwhile to embark onfurther studies of the filament systems within thesecells, which, as has been stated previously, are knownto have a key role in the phagocytic process.Although it has been established in the study
reported here that contractile proteins are present inhuman trabecular meshwork cells, more detailedinvestigations need to be performed to determinewhich smooth muscle contractile proteins apart fromactin are present in these cells and what role theyplay in cell contraction, anchorage, and phagocytosisin the normal and diseased outflow system.
We are grateful to Professor N. Ashton for helpful criticism;Mr J. Prasad and Miss E. Robins for technical assistance;and Mrs A. Wood for secretarial help.
References
Allison, A. C., Davies, P., and de Petris, S. (1971). Role ofcontractile microfilaments in macrophage movement andendocytosis. Nature, 232, 153-155.
Bill, A. (1975). The drainage of aqueous humor. InvestigativeOphthalmology, 14, 1-3.
Burnside, M. B. (1976). Possible roles of microtubules andactin filaments in retinal pigmented epithelium. Experi-mental Eye Research, 23, 257-275.
Felix, H., and Strauli, P. (1976). Different distribution patternof 100-A filaments in resting and locomotive leukaemiacells. Nature, 261, 604-605.
Goldman, R. D., and Follet, E. A. C. (1969). The structureof the major cell processes of isolated BHK21 fibroblasts.Experimental Cell Research, 57, 263-276.
Grierson, I., and Lee, W. R. (1973). Erythrocyte phago-cytosis in the human trabecular meshwork. British Journalof Ophthalmology, 57, 400-415.
Grierson, I., and Lee, W. R. (1975). The fine structure of thetrabecular meshwork at graded levels of intraocularpressure. (1) Pressure effects within the near physiologicalrange. Experimental Eye Research, 20, 505-522.
Grierson, I., Lee, W. R., and Abraham, S. (1978). Effects ofpilocarpine on the morphology of the human outflowapparatus. British Journal of Ophthalmology, 62, 302-313.
Inomata, H., Bill, A., and Smelser, G. K. (1972). Aqueous
humor pathways through the trabecular meshwork andinto Schlemm's canal in the cynomolgus monkey (Macacairus). An electron microscopic study. American Journal ofOphthalmology, 73, 760-789.
Ishikawa, H., Brischoff, R., and Holtzer, H. (1969). Forma-tion of arrowhead complexes with heavy meromyosin in avariety of cell types. Journal of Cell Biology, 43, 312-328.
Kaughman, P. L., and Barany, E. H. (1977). Cytochalasin Breversibly increases outflow facility in the eye of thecynomolgus monkey. Investigative Ophthalmology, 16,47-53.
Kaughman, P. L. (1977). Drugs and the trabecular mesh-work. Investigative Ophthalmology, 16, 475-477.
Kayes, J. (1975). Pressure gradient changes in the trabecularmeshwork of monkeys. American Journal of Ophthal-mology, 79, 549.
Kurki, P., Linder, E., Virtanen, I., and Stowman, S. (1977).Human smooth muscle autoantibodies reacting withintermediate (100 A) filaments. Nature, 268, 240-241.
Lansing-Taylor, D. (1975). Dynamics of cytoplasmicstructure and contractility. In Molecules and Cell Move-ment. Edited by S. Inoue and R. E. Stephens. Raven Press:New York.
Malawista, S. E., Gee, J. B. L., and Bensch, K. G. (1971).Cytochalasin B reversibly inhibits phagocytosis. Functional,metabolic, and ultrastructural effects in human bloodleukocytes and rabbit alveolar macrophages. Yale Journalof Biology and Medicine, 44, 286-300.
Pollack, R., and Rifkin, D. (1975). Actin-containing cablewithin anchorage-dependent rat embryo cells are dis-sociated by plasma and trypsin. Cell, 6, 495-506.
Pollard, T. D., and Weihing, R. R. (1974). Actin and myosinand cell movements. Critical Reviews in Biochemistry, 2,1-65.
Rahi, A., and Ashton, N. (1978). Contractile proteins inretinal endothelium and other non-muscle tissues of theeye. British Journal of Ophthalmology, 62, 627-643.
Rahi, A. H. S., Holborow, E. J., Perkins, E. S., Gungen, Y. Y.,and Dinning, W. J. (1976). Immunological investigationsin uveitis. Transactions of the Ophthalmological Societiesof the United Kingdom, 96, 113-122.
Ringvold, A. (1978). Actin filaments in trabecular endothelialcells in eyes of the vervet monkey. Acta Ophthalmologica,56, 217-225.
Svedbergh, B. (1976). Aspects of the aqueous humor drain-age. Functional ultrastructure of Schlemm's canal, thetrabecular meshwork and the corneal endothelium atdifferent intraocular pressure. Acta Universitatis Uppsalien-sis, 256, 1-71.
Tilney, L. G. (1975). Actin: Its association with membranesand the regulation of its polymerization. In Molecules andCell Movement. Edited by S. Inoue and R. E. Stephens.Raven Press: New York.
Trenchev, P., and Holborow, E. J. (1976). The specificity ofanti-actin serum. Immunology, 16, 125-136.
Tripathi, B., Ashton, N., and Knight, G. (1973). Effect ofoxygen on the developing retinal vessels of the rabbit.III. Mode of growth of rabbit retinal vessels in tissueculture. Experimental Eye Research, 15, 321-351.
Van Buskirk, E. M., and Leure-Dupree, A. E. (1978).Pathophysiology and electron microscopy of melano-malytic glaucoma. American Journal of Ophthalmology,85, 160-166.
Wessells, N. K., Spooner, B. S., Ash, J. F., Bradley, M. O.,Luduena, M. A., Taylor, E. C., Wrenn, J. T., and Yamada,K. M. (1971). Microfilaments in cellular and developmentalprocesses. Science, 171, 135-143.
8
on 9 February 2019 by guest. P
rotected by copyright.http://bjo.bm
j.com/
Br J O
phthalmol: first published as 10.1136/bjo.63.1.3 on 1 January 1979. D
ownloaded from


















