![Investigation of hydrothermal synthesis of wollastonite ...jcpr.kbs-lab.co.kr/file/JCPR_vol.11_2010/JCPR11-3/14[1].348-353.pdf · Investigation of hydrothermal synthesis of wollastonite](https://static.fdocuments.us/doc/165x107/5adae79a7f8b9ae1768dce91/investigation-of-hydrothermal-synthesis-of-wollastonite-jcprkbs-labcokrfilejcprvol112010jcpr11-3141348-353pdfinvestigation.jpg)
Mesothelial Cell Proliferation and Biopersistence of Wollastonite
11
FUNDAMENTAL AND APPLIED TOXICOLOGY 38, 173-183 (1997) ARTICLE NO. FA972344 Mesothelial Cell Proliferation and Biopersistence of Wollastonite and Crocidolite Asbestos Fibers J. L. Macdonald and A. B. Kane Department of Pathology and Laboratory Medicine, Brown University School of Medicine, Providence, Rhode Island 02912 Received January 3, 1997; accepted June 17, 1997 Mesothelial Cell Proliferation and Biopersistence of Wollaston- ite and Crocidolite Asbestos Fibers. Macdonald, J. L., and Kane, A. B. (1997). Fundam. Appl. Toxicol. 38, 173-183. The mesothelial lining is a target for the fibrotic and carcino- genic effects of mineralfibers.Fiber geometry, dimensions, chemi- cal composition, surface reactivity, and biopersistence at the target tissue have been proposed to contribute to these toxic endpoints. We established a dose-response relationship between the number of fibers delivered to the parietal peritoneal lining, inflammation, and mesothelial cell proliferation induced by intraperitoneal injec- tion of crocidolite asbestos fibers in mice. Persistence of these in- flammatory and proliferative responses depended on persistence offibersat the target tissue. Intraperitoneal injection of wollaston- itefibersinduced an early inflammatory and proliferative response that subsided after 21 days. Approximately 50% of wollastonite fibers were recovered by bleach digestion after 21 days and only 2% were recovered after 6 months. In contrast, the number of fibers recovered from tissue digests had not declined 6 months after injection of crocidolite asbestos. These results support the hypothesis that biopersistent fibers cause persistent inflammation and chronic mesothelial cell proliferation, o 1997 Sod«y of Toxicology Human exposure to asbestos fibers causes a range of pathologic reactions of the pleura: benign asbestos pleural effusion, diffuse visceral pleural fibrosis, fibrotic plaques of the parietal pleura, and diffuse malignant mesothelioma (reviewed by Churg and Green, 1995). The pathogenesis of these lesions is unknown. Pleural effusion and fibrosis of the visceral pleura may be nonspecific responses to active inflammation in the adjacent pulmonary parenchyma be- cause these pathologic reactions also accompany other acute and chronic inflammatory reactions in the lungs (Adamson et al., 1994). Fibrotic scarring of the parietal pleura and dome of the diaphragm are more specific reactions associated with exposure to asbestos fibers; calcified fibrotic pleural plaques as visualized by chest X ray or computerized axial tomography are considered as markers of prior asbestos ex- posure (Churg and Green, 1995). Diffuse malignant meso- thelioma has been reported to arise in the parietal pleura adjacent to areas with abundant lymphatic drainage. These areas have been described recently as "black spots" because they preferentially accumulate anthracotic pigment (Boutin etal., 1966). Direct intrapleural or intraperitoneal injection of natural or man-made fibers has been used to reproduce these fibrotic and malignant reactions in rodents. The pioneering studies of Pott and Stanton and their colleagues suggested that fiber dimensions are important determinants of fiber carcinogenic- ity (Stanton et al, 1981; Pott etal., 1987). Additional param- eters relevant to the biologic activity of fibers are chemical composition, surface reactivity, and biopersistence in the lungs or pleura (reviewed in McClellan et al, 1992; and Warheit et al., 1995). Although numerous studies have eval- uated biopersistence of various natural and man-made fibers in the lungs of rodents after intratracheal instillation (Muhle et al, 1994) or inhalation (Hesterberg et al, 1996), few studies have evaluated biopersistence in the pleural or perito- neal lining. A recent investigation after short-term inhalation of refractory ceramic fibers by rats confirms that fibers are rapidly translocated to the pleural space (Gelzleichter et al, 1996a). Previous investigations using intrabronchial instilla- tion of amosite asbestos fibers in dogs provide evidence for migration of fibers into lymphatics (Oberdb'rster et al, 1988). One marker of fiber migration to the pleural space is induc- tion of an inflammatory response. Intrabronchial or intratra- cheal instillation of amosite (Oberdorster et al, 1983) or crocidolite asbestos (Li et al, 1994) in rats caused an early acute inflammatory response, followed by a delayed influx of monocytes into the pleural space. Short-term inhalation of RCF-1 fibers by rats caused an increase in pleural macro- phages as well as eosinophils (Gelzleichter et al, 1996b). These investigators developed a novel technique using agar- ose gel casts to quantitate pleural fiber burden (Bermudez, 1994). Significant numbers of fibers were recovered in the pleural cast, although most were thin and short, <5 /-im in length. This recent finding raises questions about the poten- tial pathogenicity of short, thin fibers (Goodglick and Kane, 1990). Other investigators report no apparent translocation of fibers to the visceral pleura after intratracheal instillation (Adamson et al, 1994) or inhalation (Quinlan et al, 1995) 173 0272-0590/97 $25.00 Copyright © 1997 by Ihe Society of Toxicology. All rights of reproduction in any form reserved. Downloaded from https://academic.oup.com/toxsci/article-abstract/38/2/173/1633315 by guest on 04 April 2019
Transcript of Mesothelial Cell Proliferation and Biopersistence of Wollastonite

FUNDAMENTAL AND APPLIED TOXICOLOGY 38, 1 7 3 - 1 8 3 (1997)
ARTICLE NO. FA972344
Mesothelial Cell Proliferation and Biopersistenceof Wollastonite and Crocidolite Asbestos Fibers
J. L. Macdonald and A. B. Kane
Department of Pathology and Laboratory Medicine, Brown University School of Medicine, Providence, Rhode Island 02912
Received January 3, 1997; accepted June 17, 1997
Mesothelial Cell Proliferation and Biopersistence of Wollaston-ite and Crocidolite Asbestos Fibers. Macdonald, J. L., and Kane,A. B. (1997). Fundam. Appl. Toxicol. 38, 173-183.
The mesothelial lining is a target for the fibrotic and carcino-genic effects of mineral fibers. Fiber geometry, dimensions, chemi-cal composition, surface reactivity, and biopersistence at the targettissue have been proposed to contribute to these toxic endpoints.We established a dose-response relationship between the numberof fibers delivered to the parietal peritoneal lining, inflammation,and mesothelial cell proliferation induced by intraperitoneal injec-tion of crocidolite asbestos fibers in mice. Persistence of these in-flammatory and proliferative responses depended on persistenceof fibers at the target tissue. Intraperitoneal injection of wollaston-ite fibers induced an early inflammatory and proliferative responsethat subsided after 21 days. Approximately 50% of wollastonitefibers were recovered by bleach digestion after 21 days and only2% were recovered after 6 months. In contrast, the number offibers recovered from tissue digests had not declined 6 monthsafter injection of crocidolite asbestos. These results support thehypothesis that biopersistent fibers cause persistent inflammationand chronic mesothelial cell proliferation, o 1997 Sod«y of Toxicology
Human exposure to asbestos fibers causes a range ofpathologic reactions of the pleura: benign asbestos pleuraleffusion, diffuse visceral pleural fibrosis, fibrotic plaquesof the parietal pleura, and diffuse malignant mesothelioma(reviewed by Churg and Green, 1995). The pathogenesis ofthese lesions is unknown. Pleural effusion and fibrosis ofthe visceral pleura may be nonspecific responses to activeinflammation in the adjacent pulmonary parenchyma be-cause these pathologic reactions also accompany other acuteand chronic inflammatory reactions in the lungs (Adamsonet al., 1994). Fibrotic scarring of the parietal pleura anddome of the diaphragm are more specific reactions associatedwith exposure to asbestos fibers; calcified fibrotic pleuralplaques as visualized by chest X ray or computerized axialtomography are considered as markers of prior asbestos ex-posure (Churg and Green, 1995). Diffuse malignant meso-thelioma has been reported to arise in the parietal pleuraadjacent to areas with abundant lymphatic drainage. These
areas have been described recently as "black spots" becausethey preferentially accumulate anthracotic pigment (Boutinetal., 1966).
Direct intrapleural or intraperitoneal injection of naturalor man-made fibers has been used to reproduce these fibroticand malignant reactions in rodents. The pioneering studiesof Pott and Stanton and their colleagues suggested that fiberdimensions are important determinants of fiber carcinogenic-ity (Stanton et al, 1981; Pott etal., 1987). Additional param-eters relevant to the biologic activity of fibers are chemicalcomposition, surface reactivity, and biopersistence in thelungs or pleura (reviewed in McClellan et al, 1992; andWarheit et al., 1995). Although numerous studies have eval-uated biopersistence of various natural and man-made fibersin the lungs of rodents after intratracheal instillation (Muhleet al, 1994) or inhalation (Hesterberg et al, 1996), fewstudies have evaluated biopersistence in the pleural or perito-neal lining. A recent investigation after short-term inhalationof refractory ceramic fibers by rats confirms that fibers arerapidly translocated to the pleural space (Gelzleichter et al,1996a). Previous investigations using intrabronchial instilla-tion of amosite asbestos fibers in dogs provide evidence formigration of fibers into lymphatics (Oberdb'rster et al, 1988).One marker of fiber migration to the pleural space is induc-tion of an inflammatory response. Intrabronchial or intratra-cheal instillation of amosite (Oberdorster et al, 1983) orcrocidolite asbestos (Li et al, 1994) in rats caused an earlyacute inflammatory response, followed by a delayed influxof monocytes into the pleural space. Short-term inhalationof RCF-1 fibers by rats caused an increase in pleural macro-phages as well as eosinophils (Gelzleichter et al, 1996b).These investigators developed a novel technique using agar-ose gel casts to quantitate pleural fiber burden (Bermudez,1994). Significant numbers of fibers were recovered in thepleural cast, although most were thin and short, < 5 /-im inlength. This recent finding raises questions about the poten-tial pathogenicity of short, thin fibers (Goodglick and Kane,1990). Other investigators report no apparent translocationof fibers to the visceral pleura after intratracheal instillation(Adamson et al, 1994) or inhalation (Quinlan et al, 1995)
173 0272-0590/97 $25.00Copyright © 1997 by Ihe Society of Toxicology.
All rights of reproduction in any form reserved.
Dow
nloaded from https://academ
ic.oup.com/toxsci/article-abstract/38/2/173/1633315 by guest on 04 April 2019

174 MACDONALD AND KANE
of crocidolite or chrysotile asbestos fibers, although in-flammation in the adjacent lung tissue was induced. How-ever, in these studies, fiber localization was assessed by lightmicroscopy and no tissue digests were performed.
A potential biomarker of response to asbestos fibers isproliferation of mesothelial cells. This response has beenobserved morphologically after intraperitoneal injection ofasbestos fibers in rats (Friemann et al., 1990), by [3H]-thymidine incorporation after intraperitoneal injection ofcrocidolite asbestos in mice (Moalli et al., 1987), and byBrdU incorporation after intrapleural injection of man-madefibers in rats and hamsters (Rutten et al., 1994). The mecha-nism responsible for stimulation of acute mesothelial cellproliferation induced by direct injection of fibers into thepleural or peritoneal space is unknown. Proliferation couldbe a compensatory response to cell injury or apoptosis, anonspecific reaction to inflammation in the pleural or perito-neal space, or a direct mitogenic effect of fibers. No studieshave investigated the relationship between fiber dosimetry,inflammation, and mesothelial cell proliferation.
Pleural fiber dosimetry in humans has been determinedusing tissue samples obtained by fiberoptic thoracoscopy orat autopsy. Direct measurement of fibers obtained by tissuedigestion of samples of the visceral or parietal pleura ob-tained at autopsy of asbestos workers has shown fibers atthese sites in some cases, but not in others (Sebastien et al.,1980; Dodson et al., 1990). Predominantly short chrysotilefibers were found in some cases, although long amphibolefibers were recovered in lung tissue digests from the sameworker (Sebastien et al., 1980). A limitation of these studiesis the sampling technique: random biopsies of the parietalpleura were used. Boutin et al. (1996) described focal areasof high concentrations of fibers and other particulates nearlymphatic vessels in the intercostal spaces of the parietalpleura. In asbestos-exposed workers, 4.1 ± 1.9 X 106 fibers/g of dry tissue were recovered from these locations, com-pared to 0.5 ± 0.2 fibers/g of random biopsies of normalpleural tissue. Amphiboles were recovered more frequentlythan chrysotile fibers in these workers; 22.5% of the fiberswere longer than 5 fim (Boutin et al., 1996). Using fiber-optic thoracoscopy, these investigators detected early stagesof malignant mesothelioma arising near these areas of lym-phatic drainage on the parietal pleural lining (Boutin andRey, 1993). Examination of these areas by scanning electronmicroscopy revealed lymphatic channels surrounded by acti-vated macrophages with ruffled surface membranes (Boutinet al., 1996).
We have developed a murine model to study the acuteand chronic effects of crocidolite asbestos fibers after directintraperitoneal injection in mice (Kane and Macdonald,1993). In this murine model, long asbestos fibers are trappedat sites of lymphatic drainage on the inferior surface of the
diaphragm. The initial inflammatory response to fiberstrapped at this site is focal accumulation of activated macro-phages. This inflammatory response is remarkably similarto the reaction visualized by Boutin et al. (1996) in thehuman parietal pleura. We have shown that this initial in-flammatory reaction is accompanied by a peak of mesothelialcell proliferation 3 days after intraperitoneal injection of 200fig of crocidolite asbestos fibers. These initial reactions aresensitive to fiber size: neither short crocidolite asbestos fibersnor paniculate stimuli such as silica or titanium dioxideparticles elicited inflammation or mesothelial cell prolifera-tion (Branchaud et al., 1993; Moalli et al., 1987). It is hy-pothesized that persistence of long crocidolite asbestos fibersat these sites would lead to persistent inflammation andmesothelial cell proliferation. This hypothesis was testedusing a known carcinogenic fiber, crocidolite asbestos, withprolonged persistence in the lungs of humans and experimen-tal animals. In contrast, wollastonite is a naturally occurringcalcium metasilicate that has a half-life of only 21 days inthe lungs of rats following inhalation (Warheit et al., 1994).Crocidolite asbestos fibers induce malignant mesotheliomasafter direct intraperitoneal injection (Pott et al., 1987) orinhalation in rats (McConnell etal., 1994). However, wollas-tonite did not induce mesotheliomas after intraperitoneal in-jection of up to 100 mg (Pott et al., 1987) or inhalationby rats (McConnell et al., 1991). The dose-response andtemporal relationships between inflammation and mesothe-lial cell proliferation induced by crocidolite and wollastonitefibers were evaluated in a subchronic assay.
MATERIALS AND METHODS
Animals. Male C57BI/6 mice, 2 months old (Charles River Labora-tories, North Wilmington, MA) were injected intraperitoneally with crocido-lite asbestos fibers, wollastonite, or 1 ml of phosphate-buffered saline (PBS).The mice were housed in filter-top plastic cages in a barrier facility main-tained at constant temperature and humidity and a 12-hr light-dark cycle.The mice were handled according to the guidelines described in the N1HGuide for the Care and Use of Laboratory Animals as approved by theBrown University Animal Care and Use Committee.
Fiber samples. Crocidolite asbestos fibers were obtained from stocksoriginally prepared and characterized by the Union Internationale Contrele Cancer (Timbrell, 1971/1972) and purchased from Duke Scientific Corpo-ration (Palo Alto, CA). Crocidolite is a member of the amphibole asbestosfamily: (Na2(Fe3+)2(Fe2+)3Si8O22(OH)2). NYAD 1250 wollastonite (Lot 11-28-94), a fine particle grade of calcium metasilicate (CaSiO3), was a giftfrom NYCO (Willsboro, NY). Stock suspensions of crocidolite were pre-pared and characterized as described previously (Moalli et al, 1987; Good-glick and Kane, 1990). This stock of crocidolite asbestos fibers inducedmalignant mesotheliomas after weekly intraperitoneal injections in 40-50%of mice (Kane and Macdonald, 1993). The size distribution of NYAD 1250was described previously (Bellmann and Muhle, 1994).
Peritoneal lavage. At the times indicated in the text, mice were termi-nated with an overdose of ether and the peritoneum was lavaged with a totalof 15 ml of PBS as described previously (Macdonald and Kane, 1986). Thelavage fluid was centrifuged at 200g for 5 min at 4°C, resuspcnded in Dulbec-
Dow
nloaded from https://academ
ic.oup.com/toxsci/article-abstract/38/2/173/1633315 by guest on 04 April 2019

FIBER PERSISTENCE AND MESOTHELIAL CELL PROLIFERATION 175
FIG. 1. BrdU immunohistochemistry on whole mounts of the diaphragm. (A) Three days after ip injection of 200 /ig of crocidolite asbestos. (B) 21days after ip injection of 200 /jg of crocidolite asbestos. (C) Three days after ip injection of 2 mg of wollastonite. (D) 21 days after ip injection of 2mg of wollastonite. X200.
co's modified Eagle's medium, high-glucose formula supplemented with 20mM L-glutamine, 10 mM sodium pyruvate, 5000 units of penicillin, 5000 mgof streptomycin, and 10% heat-inactivated fetal calf serum (Life Technolo-gies, Inc., Grand Island, NY). The total cell number was counted in a hemacy-tometer. Cells were plated overnight at 1 x 106/ml in 24 multiwell platescontaining glass coverslips 1.2 cm in diameter. The coverslips were fixed,stained with May-Grilnwald-Giemsa solution, and examined by light mi-croscopy as described previously (Branchaud el al., 1993).
Fixation and dissection of tissues. Whole mounts of the diaphragmincluding the falciform ligament, omentum, and intestinal mesenteries weredissected at 20-70X using a dissecting stereomicroscope as described inMoalli et al. (1987). One hour prior to termination, mice were injected
intraperitoneally with 5-bromodeoxycytidine (BrdC; 1 mg/ml, Sigma Chemi-cal Co., St. Louis, MO); BrdC is more soluble than 5-bromodeoxyuridine(BrdU) so the labeling indices are more reproducible (Ntlsse et al, 1985).Whole mounts of the diaphragm were fixed overnight in Omni-fix II (An-ConGenetics, Inc., Melville, NY) and then processed for immunohistochemistry.Cross-sections were prepared from the whole mounts, including the falciformligament, the omentum, and the intestinal mesenteries; embedded in paraffin;and processed for light microscopy as described previously (Moalli et al.,1987). Serial sections were stained with a polyclonal antibody (1:6O0 dilution)directed against a broad spectrum of cytokeratins (A575; Dako Corp., Carpin-teria, CA) that reacts with mature mesothelial cells to confirm the mesothelialorigin of surface proliferating cells (Bolen et al., 1986).
Dow
nloaded from https://academ
ic.oup.com/toxsci/article-abstract/38/2/173/1633315 by guest on 04 April 2019

176 MACDONALD AND KANE
A Asbestos B Wollastonlte B Wollastonlte
20
2
S
10
FIG. 2. Time course of mesothelial cell proliferation after ip injectionof crocidolite asbestos (A) or wollastonite (B).
Immunohistochemistry. Incorporation of BrdU by proliferating meso-thelial cells on the inferior surface of the diaphragm was detected by immu-nohistochemistry using fixed whole mounts. The diaphragms were quenchedin H2O2-methanol, rinsed, and denatured in 0.75 N HCI for I hr at roomtemperature. The diaphragms were neutralized in 0.1 M borate buffer, pH8.5, and then incubated with a rat monoclonal anti-BrdU antibody (AccurateChemical & Scientific Corp., Westbury, NY) at 1:40 dilution in PBS con-taining 0.1 % BSA for 1 hr at room temperature. A secondary biotinylatedanti-rat antibody (Organon Teknika-Cappel, Malvern, PA) at 1:250 dilutionwas applied for 30 min followed by ABC reagent (Vector Laboratories,Inc., Burlingame, CA) and development of a brown precipitate using diami-nobenzidine. Twenty areas were selected at the musculotendinous junctionand over the muscular region of the diaphragm and counted at 20x magni-fication using an eyepiece grid. The mean number of proliferating cells thatincorporated BrdU was calculated per square millimeter of surface area.
Fiber digests. Samples of the central tendinous region of the diaphragm,the musculotendinous junction, and the falciform ligament were dissected,measured, and digested using bleach (5.25% sodium hypochlorite) for 48hr at room temperature. This procedure has been verified using crocidoliteasbestos and wollastonite fibers (Warheit et al., 1994). All solutions usedin the digestion procedure were prefiltered. Recovery of fibers by thisdigestion technique was determined as follows. A series of increasinglydilute solutions of native fibers of known gravimetric quantities was distrib-uted on nucleopore filters using vacuum filtration. One-half of each filterwas prepared for scanning electron microscopy (SEM) and the number offibers/mm2 was counted as described below. A corresponding 1-mm2 area
FIG. 3. Total cell recovery from peritoneal lavage after ip injection ofcrocidolite asbestos (A) or wollastonite (B).
was cut from the remaining half of the filter, dissolved in chloroform, andsubjected to the digestion and filtration procedure. Recovery ranged from78% for the most dilute concentration of fibers (5000/mm2) to 89% for thehighest concentration of fibers tested (25,000/mm2). No fibers were recov-ered following digestion of tissues from mice injected with PBS. Fiberswere collected on 25-mm nucleopore polycarbonate filters (pore size 0.4fim), dehydrated, gold coated, and mounted on an aluminum planchet.Filters were examined using an Hitachi 2700 analytical scanning electronmicroscope equipped with a Link EXL-FQ1 X-ray detector. The WHO/EURO (1985) counting guidelines were followed with these modifications.All fibers and particulates were counted at 4000X. The number of fieldscounted covered at least 0.18 mm2 of the filter surface. Counting stoppedwhen a total of 1000 fibers and nonfibrous particles were reached or when1 mm2 of the filter surface had been examined. Representative photomicro-graphs of original samples and fiber digests were taken. The chemicalcomposition of all fiber samples before injection and after recovery fromtissue digests was verified by energy dispersive X-ray analysis (Macdonaldand Kane, 1986).
Statistical analysis. The mean values ± SEM were calculated for threeto five mice from each treatment group. Statistical significance of differ-ences between control and treated groups was determined by analysis ofvariance (ANOVA) using Fisher's PLSD test A p value « 0.05 was consid-ered as statistically significant.
RESULTS
Mesothelial Cell ProliferationThe time course of mesothelial cell proliferation produced
by intraperitoneal injection of crocidolite asbestos fibers or
TABLE 1Dose Response of Fiber Deposition and Mesothelial Cell Prolif-
eration 3 Days after Intraperitoneal Injection of Crocidolite Asbes-tos or Wollastonite
SalineAsbestos
20 pg200 fig
Wollastonite200/jg
2 mg
No. fibers X
4.04 ± 2.026.20 ± 4.2
4.25 ± 1.619.50 ± 6.1
21 ± 3.1
689 ± 2431231 ± 155
98 ±668 ±
4.082.8
No. labeled cells/mm2 ^
Asbestos B Wollastonlte
100
o
201.11 •
1 X i ft21
DAYS
FIG. 4. Differential cell count recovered from peritoneal lavage afterip injection of 200 pg crocidolite asbestos (A) or 10 mg wollastonite (B).Hatched bars, macrophages; solid bars, neutrophils; open bars, lymphocytes.
Dow
nloaded from https://academ
ic.oup.com/toxsci/article-abstract/38/2/173/1633315 by guest on 04 April 2019
FIBER PERSISTENCE AND MESOTHELIAL CELL PROLIFERATION 177
#••
V '
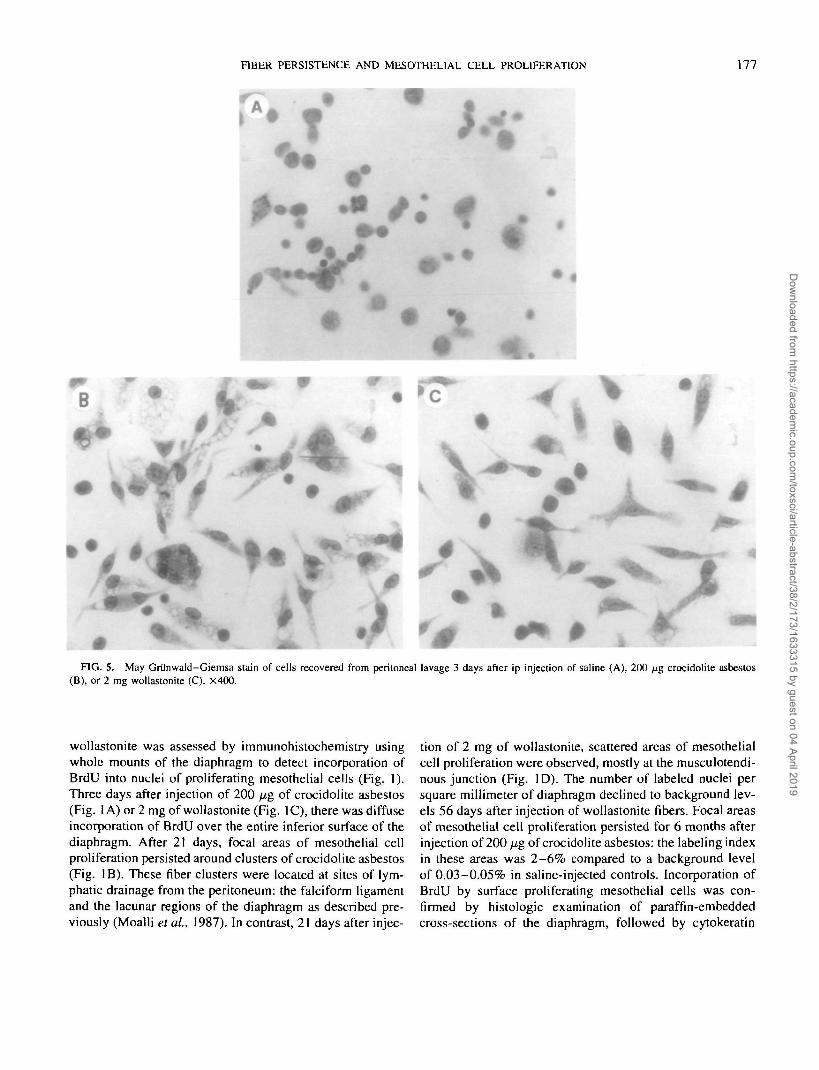
FIG. 5. May Grilnwald-Giemsa stain of cells recovered from peritoneal lavage 3 days after ip injection of saline (A), 200 fj.g crocidolite asbestos(B), or 2 mg wollastonite (C). X400.
wollastonite was assessed by immunohistochemistry usingwhole mounts of the diaphragm to detect incorporation ofBrdU into nuclei of proliferating mesothelial cells (Fig. 1).Three days after injection of 200 fig of crocidolite asbestos(Fig. 1 A) or 2 mg of wollastonite (Fig. 1C), there was diffuseincorporation of BrdU over the entire inferior surface of thediaphragm. After 21 days, focal areas of mesothelial cellproliferation persisted around clusters of crocidolite asbestos(Fig. IB). These fiber clusters were located at sites of lym-phatic drainage from the peritoneum: the falciform ligamentand the lacunar regions of the diaphragm as described pre-viously (Moalli et ai, 1987). In contrast, 21 days after injec-
tion of 2 mg of wollastonite, scattered areas of mesothelialcell proliferation were observed, mostly at the musculotendi-nous junction (Fig. ID). The number of labeled nuclei persquare millimeter of diaphragm declined to background lev-els 56 days after injection of wollastonite fibers. Focal areasof mesothelial cell proliferation persisted for 6 months afterinjection of 200 fig of crocidolite asbestos: the labeling indexin these areas was 2 -6% compared to a background levelof 0.03-0.05% in saline-injected controls. Incorporation ofBrdU by surface proliferating mesothelial cells was con-firmed by histologic examination of paraffin-embeddedcross-sections of the diaphragm, followed by cytokeratin
Dow
nloaded from https://academ
ic.oup.com/toxsci/article-abstract/38/2/173/1633315 by guest on 04 April 2019

178 MACDONALD AND KANE
x -
FIG. 6. Histologic sections of the diaphragm. (A) Three days after ip injection of 200 fig of crocidolite asbestos. X400. (B) Three days after ipinjection of 10 mg of wollastonite. X400. (C) 21 days after ip injection of 10 mg of wollastonite. X200. Hematoxylin and eosin stain.
immunohistochemistry to identify mature mesothelial cells(data not shown).
The initial proliferative response to intraperitoneal injec-tion of crocidolite asbestos or wollastonite fibers was dosedependent (Fig. 2). After 3 days, mesothelial cell prolifera-tion increased after injection of 20 or 200 fig of crocidoliteasbestos; no further increase was noted after injection of 1mg. Mesothelial cell proliferation was also increased afterinjection of 200 fig or 2 mg of wollastonite, although evenat these higher doses, wollastonite fibers were less potentin inducing mesothelial cell proliferation than crocidoliteasbestos fibers after 3 days. At the highest dose of wollaston-ite fibers tested (10 mg), mesothelial cell proliferation waselevated after 21 days and then declined to background levelsafter 56 days. There was a dose-response relationship be-tween the number of fibers deposited on the inferior surfaceof the diaphragm and stimulation of mesothelial cell prolifer-ation 3 days after intraperitoneal injection of crocidolite as-bestos or wollastonite (Table 1).
Inflammatory Responses to Crocidolite Asbestos orWollastonite Fibers
Cellular analysis of peritoneal lavage fluid indicated atransient inflammatory response to intraperitoneal injection
of crocidolite asbestos or wollastonite fibers. The initial in-flammatory response to crocidolite asbestos was dose depen-dent over the range of 20 fig-\ mg after 3 days (Fig. 3A).As reported previously (Branchaud et ai, 1993), this in-flammatory reaction declined after 21 days. However, after56 days, a statistically significant increase in the number ofcells recovered by peritoneal lavage was found at all dosesof crocidolite asbestos fibers. The magnitude of the initialinflammatory response to wollastonite was dose dependentat 200 fig or 2 mg (Fig. 3B). Similar to crocidolite asbestos,the inflammatory response to wollastonite subsided after 21days. At later time points, inflammation persisted only afterinjection of 10 mg of wollastonite.
The initial inflammatory response to 200 fig of crocidoliteasbestos or 10 mg of wollastonite was characterized by acti-vated macrophages. A low percentage of neutrophils wasrecovered 3 and 56 days after injection of crocidolite asbes-tos at 200 fig (Fig. 4A). After injection of 10 mg of wollas-tonite, activated macrophages were recovered after 3 or 21days. At later time points, a statistically significant increasein lymphocytes was found (Fig. 4B). The morphology ofinflammatory cells recovered by peritoneal lavage 3 daysafter injection of saline, crocidolite asbestos, or wollastoniteis compared in Fig. 5. Peritoneal lavage cells from saline-
Dow
nloaded from https://academ
ic.oup.com/toxsci/article-abstract/38/2/173/1633315 by guest on 04 April 2019

FIBER PERSISTENCE AND MESOTHELIAL CELL PROLIFERATION 179
B
•.-•.; W 'r.
FIG. 7. Histologic sections of mesentery. (A) Twenty-one days after ip injection of 200 \i% of crocidolite asbestos. (B) Three days after ip injectionof 2 mg of wollastonite. (C) Three days after ip injection of 10 mg of wollastonite, dark-field illumination. (D) Three days after ip injection of 10 mgof wollastonite, bright-field illumination. XI00. Hematoxylin and eosin stain.
injected mice plated on glass coverslips overnight weresmaller and round in comparison with the large, activatedmacrophages and multinucleated giant cells obtained afterinjection of 200 pig of crocidolite asbestos. After injectionof wollastonite, the peritoneal macrophages were smallerand less adherent in comparison to those obtained after injec-tion of crocidolite asbestos. The morphology and histochem-ical characteristics of activated macrophages obtained 3 daysafter intraperitoneal injection of crocidolite asbestos weredescribed previously (Branchaud et at., 1993).
Histopathology
Long fibers of crocidolite asbestos or wollastonite aretrapped at lymphatic stomata on the inferior surface of thediaphragm. The inflammatory responses to these fiberstrapped on the parietal peritoneal lining reflects the distribu-tion of inflammatory cells recovered by peritoneal lavage.Cross-sections of the diaphragm at sites of fiber depositionwere compared after intraperitoneal injection of 200 /xg ofcrocidolite asbestos fibers or 2 mg of wollastonite (Fig. 6).
Dow
nloaded from https://academ
ic.oup.com/toxsci/article-abstract/38/2/173/1633315 by guest on 04 April 2019

180 MACDONALD AND KANE
These doses were selected because they produced a similarmagnitude of inflammation as reflected by the total cell num-ber recovered by peritoneal lavage. The initial histopatho-logic reaction to crocidolite asbestos fibers trapped on theinferior surface of the diaphragm is accumulation of macro-phages and formation of multinucleated giant cells aroundlong fibers or clusters of short fibers (Fig. 6A). A few lym-phocytes, plasma cells, and neutrophils are intermingledwithin this lesion. A similar, but less intense, inflammatoryresponse was observed at sites of wollastonite deposition onthe inferior surface of the diaphragm (Fig. 6B).
Macrophages and multinucleated giant cells persisted atsites of asbestos fiber deposition on the inferior surface ofthe diaphragm. After 56 days, these lesions became progres-sively more fibrotic as described previously (Kane and Mac-donald, 1993). In contrast, the inflammatory response to in-traperitoneal injection of 200 fig or 2 mg of wollastoniteresolved completely after 21 days. At the highest dose ofwollastonite (10 mg), focal accumulations of mature lym-phocytes were observed on the inferior surface of the dia-phragm after 21 days (Fig. 6C). Serial sections of the dia-phragm confirmed that these focal lesions completely re-solved after 6 months.
The inferior surface of the diaphragm is a major site oflymphatic clearance of any particulates or fibers introducedinto the peritoneal cavity. At high doses, particulates andfibers are also trapped at milky spots (Kampmeier's foci) atthe mesenteric surface. Particles or fibers less than 10 (imin diameter are successfully cleared by the lymphatics(Moalli et ai, 1987) and accumulate in lymph nodes withinthe omentum and mesenteries (Branchaud et al., 1993). Themineral samples used in this study are a mixture of particu-lates, short fibers, and long fibers. Short asbestos fibers arecleared to mesenteric lymph nodes, while long fibers accu-mulate in milky spots on the surface of the mesenteries. Theinflammatory response to asbestos fibers at these sites isaccumulation of activated macrophages and multinucleatedgiant cells which become progressively fibrotic (Fig. 7A).Short fragments of wollastonite also accumulate in milkyspots initially; only a few macrophages and multinucleatedgiant cells are found 3 days after injection of 200 fxg or 2mg of wollastonite (Fig. 7B). At the highest dose of wollas-tonite (10 mg), large aggregates of fibers were found looselyadherent to the mesenteric surface; these are readily apparentby dark field illumination (Fig. 7C). Under bright field illu-mination, few inflammatory cells are found in these largeaggregates of wollastonite (Fig. 7D). After 6 months, serialsections of the mesenteries showed no residual evidenceof wollastonite aggregates or.macrophages, while lymphoidhyperplasia at milky spots persisted.
Fiber BiopersistenceSections of the diaphragm and falciform ligament were
dissected, digested in bleach, and analyzed for fiber content
60-
40-
20-
20 40DAYS
60
FIG. 8. Biopersistence of crocidolite asbestos (closed circles) or wollas-tonite fibers (open circles) on the diaphragm following ip injection.
at various times after intraperitoneal injection of 200 /xg ofcrocidolite asbestos fibers or 2 mg of wollastonite (Fig. 8).After 3 days, approximately 20-25 X 103 fibers/mm2 weredeposited on the diaphragm. After 21 days, the number ofcrocidolite asbestos fibers recovered by bleach digestion in-creased and then leveled off after 56 days. In contrast, thenumber of wollastonite fibers recovered by bleach digestiondecreased by 50% after 21 days. After 6 months, only 2%of wollastonite fibers deposited after 3 days was recovered.No changes in surface morphology or fiber shape were seenin crocidolite asbestos fibers recovered from the diaphragmat any time point (Fig. 9A). After only 3 days, however,surface etching of wollastonite fibers was visible (Figs. 9Band 9C).
DISCUSSION
These results support the hypothesis that persistence ofcrocidolite asbestos fibers at sites of lymphatic drainage inthe peritoneum leads to chronic inflammation and persistentmesothelial cell proliferation. The chronic inflammatory re-sponse to crocidolite is characterized by accumulation ofactivated macrophages and multinucleated giant cells at sitesof fiber deposition. Focal areas of persistent mesothelial cellproliferation are localized to sites of asbestos fiber clusters.Although intraperitoneal injection of wollastonite fibers alsoproduced an early, but less intense, inflammatory response,this resolved as the fibers were cleared from or dissolved attheir initial sites of deposition around lymphatic stomata andat milky spots in the mesenteries. The initial proliferativeresponse to both crocidolite asbestos fibers and wollastonitedepended on the dose of fibers delivered to the diaphragm 3days after intraperitoneal injection. In contrast to crocidoliteasbestos fibers, wollastonite was cleared from the diaphragmafter 21 days and mesothelial cell proliferation returned tocontrol levels.
Dow
nloaded from https://academ
ic.oup.com/toxsci/article-abstract/38/2/173/1633315 by guest on 04 April 2019

FIBER PERSISTENCE AND MESOTHELIAL CELL PROLIFERATION 181
FIG. 9. Scanning electron micrographs of fibers: (A) Native UICC crocidolite asbestos at 3000X. (B) Native wollastonite at 1500X. (C) Wollastoniterecovered from the diaphragm 3 days after ip injection of 2 mg. 5000X.
This is the first study that correlates fiber persistence andinflammation with mesothelial cell proliferation in the peri-toneal lining. Although the kinetics for clearance of wollas-tonite from the peritoneum are slower than reported fromthe lungs after inhalation in rats (Warheit et al., 1994), thesefibers are cleared almost completely from the inferior surfaceof the diaphragm after 6 months. Similar to the results ofintratracheal instillation (Muhle et ai, 1994) or inhalationstudies (Hesterberg et ai, 1996), crocidolite asbestos fibersare not modified or cleared from sites of deposition in theperitoneal lining. Transient inflammatory reactions were alsoreported in the lungs of rats during a short-term inhalationbioassay of wollastonite, while the inflammatory and prolif-erative responses to crocidolite asbestos persisted in the
lungs of rats following inhalation (Warheit et ai, 1994; Be-rub6 et ai, 1996).
In addition to biopersistence, chemical composition andsurface reactivity of asbestos fibers also contribute to thechronic inflammatory, fibrotic, and proliferative reactions ob-served in the lungs and mesothelial lining. Reactive oxygenmetabolites such as hydroxyl radicals generated by the iron-catalyzed Fenton reaction are hypothesized to mediate theinflammatory and carcinogenic effects of asbestos fibers(Moyer et ai, 1994). Amphiboles such as crocidolite asbestoshave a high iron content that is bioavailable (Fubini, 1993).
Modification of the shape of wollastonite fibers was notedin the short-term inhalation bioassay described above (War-heit et al., 1994). Surface etching of wollastonite fibers was
Dow
nloaded from https://academ
ic.oup.com/toxsci/article-abstract/38/2/173/1633315 by guest on 04 April 2019

182 MACDONALD AND KANE
also observed following intraperitoneal injection. This sur-face etching may leave wollastonite fibers more vulnerableto transverse splitting. Although macrophages initially accu-mulated at sites of crocidolite asbestos and wollastonite fiberdeposition in the peritoneal lining, it is unknown whethersurface etching, breaking, and dissolution of wollastoniteoccurred in the peritoneal lining fluid or within macrophagephagolysosomes.
A major caveat in intraperitoneal injection studies is selec-tion of doses that produce a biologic response but do notcause a bolus effect. This was demonstrated by Collier etal. (1994) by injecting a range of doses of glass fibers intothe peritoneal cavity of rats. In their study, doses greaterthan 1.5 mg cause formation of fiber clumps or nodules.This effect did not occur after injection of up to 10 mg ofcrocidolite asbestos fibers in mice; however, large fiberclumps were produced by injection of 10 mg of wollastonite.In the study reported by Collier et al. (1994), the glass fiberswere 2 t̂m in diameter; this may have contributed to theirtendency to form aggregates at lower doses than we observedfor crocidolite asbestos or wollastonite in mice. The in-flammatory response to the highest dose of wollastonite asso-ciated with a bolus effect in mice was different from theresponse to lower doses. Large aggregates of wollastonitefibers were loosely attached to the mesenteries or floatingfree in the peritoneal cavity. In contrast to fibers trapped atlymphatic stomata or in milky spots, very few macrophagesaccumulated on or within these large aggregates. In the Col-lier et al. study (1994) using glass fibers, these aggregateswere the site of a granulomatous inflammatory response.Despite aggregation into clumps, even the high dose of wol-lastonite fibers injected in mice eventually dissolved com-pletely with no evidence of a granulomatous reaction orfibrosis seen on serial sections of the diaphragm or mesenter-ies 6 months after the initial injection.
Although most of the wollastonite fibers disappeared frominitial sites of deposition, a focal chronic inflammatory re-sponse remained 3 weeks after injection of 10 mg on theinferior surface of the diaphragm and 6 months after injec-tion within the mesenteries. This focal chronic inflammatoryresponse to wollastonite was qualitatively different from theinitial response to these fibers characterized by accumulationof macrophages. The initial macrophage response resolvedquite rapidly and was replaced by focal clusters of maturelymphocytes on the inferior surface of the diaphragm andlymphoid hyperplasia within the mesenteries. No wollaston-ite fibers were identified in these lesions using darkfieldillumination. This lymphocytic hyperplasia may be a re-sponse to soluble components released from dissolving wol-lastonite fibers.
This subchronic assay of biopersistence, inflammation, andmesothelial cell proliferation may provide information about
the comparative biologic activities of man-made fibers in themesothelial lining. Although the mechanism responsible forbiopersistence and clearance of fibers from the mesotheliallining may be different from the lungs, recent rodent inhala-tion studies suggest that man-made fibers are translocated tothe pleura following inhalation (Gelzleichter et al., 1996a).There is recent documentation that asbestos fibers also accu-mulate in the parietal pleura of humans. In this murine model,approximately 1.2 X 107 crocidolite asbestos fibers per gramof wet tissue were deposited on the diaphragm and inducedpersistent inflammatory and proliferative responses. This iscomparable to approximately 4.1 X 10* fibers per gram ofdry tissue recovered from "black spots" on the parietal pleuraof asbestos-exposed workers (Boutin et al., 1996). Additionalinformation is required to assess the biopersistence and bio-logical activity of fibers in the mesothelial lining. If precau-tions are taken to avoid a bolus phenomenon, a subchronicintraperitoneal injection assay can rank the biologic effects ofman-made fibers relative to those produced by wollastonite,a noncarcinogenic fiber, and crocidolite asbestos, a biopersis-tent, carcinogenic fiber.
ACKNOWLEDGMENTS
The authors acknowledge the excellent technical assistance of S. Kunz.This research was supported by N1H Grant RO1 ESO5712 from the NationalInstitute of Environmental Health Sciences.
REFERENCES
Adamson, I. Y. R., Bakowski, J., and Bowden D. H. (1994). Mesothelialcell proliferation: A nonspecific response to lung injury associated withfibrosis. Am. J. Respir. Cell Mol. Biol. 10, 253-258.
Bellmann, B., and Muhle, H. (1994). Investigation of the biodurability of wollas-tonite and xonotlite. Environ. Health Perspect 102(Suppl. 5), 191-195.
Bermudez, E. (1994). Recovery of particles from the pleural cavity usingagarose casts: A novel method for the determination of fiber dose to therat pleura. Inhal. Toxicol. 6, 115 — 124.
Be'rube', J. A., Quinlan, T. R., Moulton, G., Hemenway, D., O'Shaughnessy,P., Vacek, P. and Mossman, B. T. (1996). Comparative proliferative andhistopathologic changes in rat lungs after inhalation of chrysotile or cro-cidolite asbestos. Toxicol. Appl. Pharmacol. 137, 67-74.
Bignon, J. (1989). Mineral fibers in the non-occupational environment InNon-Occupational Exposure to Mineral Fibers (J. Bignon, J. Peto, andR. Saraci, Eds.), IARC Scientific Publications No. 90, pp. 3-29. IARC,Lyon, France.
Bolen, J. W., Hammar, S. P., and McNutt, M. A. (1986). Reactive andneoplastic serosal tissue: A light-microscopic, ultrastructural and im-muno-cytochemical study. Am. J. Surg. Pathol. 10, 34-47.
Boutin, C , Dumortier, P., Rey, F., Viallat, J. R., and De Vuyst, P. (1996).Black spots concentrate oncogenic asbestos fibers in the parietal pleura:Thoracoscopic and mineralogic study. Am. J. Respir. Crit. Care Med.153, 444-449.
Boutin, C. and Rey, F. (1993). Thoracoscopy in pleural malignant mcsotheli-oma; A prospective study of 188 consecutive patients. Cancer 72, 389-393.
Branchaud, R. M., Garant, L. J., and Kane, A. B. (1993). Pathogenesis of
Dow
nloaded from https://academ
ic.oup.com/toxsci/article-abstract/38/2/173/1633315 by guest on 04 April 2019

FIBER PERSISTENCE AND MESOTHELIAL CELL PROLIFERATION 183
mesothelial reactions to asbestos fibers: Monocyte recruitment and mac-
rophage activation. Palhobiology 61, 154—163.
Churg, A. M., and Green, F. H. Y. (1995). Occupational lung c'sease. InPathology of the Lung (W. M. Thurlbeck and A. M. Churg, Eds.), 2nd
ed., pp. 851-930. Thieme, New York.
Collier, C. G., Morris, K. J., Launder, K. A., Humphreys, I. A., Morgan,A., Eastes, W., and Townsend, S. (1994). The behavior of glass fibersin the rat following intraperitoneal injection. Regul. Toxicol. Pharmacol.
20, S89-S103.
Davis, J. M. G., Bolton, R. E., Miller, B. G., and Niven, K. (1991). Mesothe-lioma dose response following intraperitoneal injection of mineral fibres.
Int. J. Exp. Pathol. 11, 263-274.
De Vuyst, P., Dumortier, P., Swaen, G. M. H., Pairon, J. C, and Brochard,P. (1995). Respiratory health effects of man-made vitreous (mineral)
fibres. Eur. Respir. J. 8, 2149-2173.
Dodson, R. F., Williams, M. J., Corn, C. J., Brollo, A., and Bianchi, C.(1990). Asbestos content of lung tissue, lymph nodes and pleura] plaquesfrom former shipyard workers. Am. Rev. Respir. Dis. 142, 843-847.
Friemann, J., Pott, F., and MUller, K.-M. (1990). Mesothelial proliferationdue to asbestos and man-made fibres. Experimental studies on rat omen-tum. Path. Res. Pract. 186, 117-123.
Fubini, B. (1993). The possible role of surface chemistry in the toxicityof inhaled fibers. In Fiber Toxicology (D. Warheit, Ed.), pp. 229-258.Academic Press, San Diego.
Gelzleichter, T. R., Bermudez, E., Mangum, J. B , Wong, B. A., Eventt,J. I., and Moss, O. R. (1996a). Pulmonary and pleural responses in Fischer344 rats following short-term inhalation of a synthetic vitreous fiber. I.Quantitation of lung and pleural fiber burdens. Fundam. Appl. Toxicol.30, 31-38.
Gelzleichter, T. R., Bermudez, E., Mangum, J. B., Wong, B. A., Moss,O. R., and Everitt, J. I. (1996b). Pulmonary and pleural responses inFischer 344 rats following short-term inhalation of a synthetic vitreousfiber. II. Pathobiologic responses. Fundam. Appl. Toxicol 30, 39-46.
Goodglick, L. A., and Kane, A. B. (1990). Cytotoxicity of long and shortcrocidolite asbestos fibers in vivo and in vitro. Cancer Res. 50, 5152-5163.
Hesterberg, T. W., Miller, W. C , Mast, R., McConnell, E. E., Berstein,D. M., and Anderson, R. (1994). Relationship between lung biopersis-tence and biological effects of man-made vitreous fibers after chronicinhalation in rats. Environ. Health Perspect. 102(Suppl. 5), 122—137.
Hesterberg, T. W., Miller, W. C , Musselman, R. P., Kamstrup, Q., Hamil-ton, R. D., and Thevenaz, P. (1996). Biopersistence of man-made vitreousfibers and crocidolite asbestos in the rat lung following inhalation. Fun-dam. Appl. Toxicol. 29, 267-279.
Kane, A. B., and Macdonald, J. L. (1993). Mechanisms of mesothelial cellinjury, proliferation and neoplasia induced by asbestos fibers. In FiberToxicology (D. Warheit, Ed.), pp. 323-348. Academic Press, San Diego.
Li, X. Y., Lamb, D., and Donaldson, K. (1994). Mesothelial cell injurycaused by pleural leukocytes from rats treated with intratracheal instilla-tion of crocidolite asbestos or corynebacterium parvum. Environ. Res.64, 181-191.
Macdonald, J. L., and Kane, A. B. (1986). Identification of asbestos fiberswithin single cells. Lab. Invest. 55, 177-185.
McClellan, R. O., Miller, F. J., Hesterberg, T. W., Warheit, D. B., Bunn,W. B., Kane, A. B., Lippmann, M., Mast, R. W., McConnell, E. E., andReinhardt, C. F. (1992). Approaches to evaluating the toxicity and carci-nogenicity of man-made fibers. Regul. Toxicol. Pharmacol. 16, 321-364.
McConnell, E. E., Mast, R. W., Hesterberg, T. W., Chevalier, J., Kotin, P.,Bernstein, D. M., Thevanaz, P., Glass, L. R., and Anderson, R. (1995).
Chronic inhalation toxicity of a kaolin-based refractory ceramic fiber inSyrian golden hamsters. Inhal. Toxicol. 7, 503-532.
McConnell, E. E., Kamstrup, O., Musselman, R., Hesterberg, T. W., Cheva-lier, J., Miller, W. C , and Thevenaz, P. (1994). Chronic inhalation studyof size-separated rock and slag wool insulation fibers in Fischer 344/N
rats. Inhal. Toxicol. 6, 541-614.
McConnell, E. E., Hall, L., and Adkins, B. (1991). Studies on the chronictoxicity (inhalation) of wollastonite in Fischer 344 rats. Inhal. Toxicol.
3, 323-337.
Moalli, P. A., Macdonald, J. L., Goodglick, L. A., and Kane, A. (1987).Acute injury and regeneration of the mesothelium in response to asbestosfibers. Am, J. Pathol. 128, 426-445.
Moyer, V. D., Cistulli, C. A., Vaslet, C. A., and Kane, A. B. (1994). Oxygenradicals and asbestos carcinogenesis. Environ. Health Perspect.
102(Suppl. 10), 131-136.
Muhle, H., Bellman, B., and Pott, F. (1994). Comparative investigations ofthe biodurabihty of mineral fibers in the rat lung. Environ, Health Per-spect. 102(Suppl. 5), 163-168
NUsse, M., Afzal, S. M. J., Carr, B., and Kavanau, K. (1985). Cell cyclekinetic measurements in an irradiated rat rhabdomyosarcoma using a
monoclonal antibody to bromodeoxyuridine, Cytometry 6, 611-619.
Oberdbrster, G., Morrow, P. E., and Spumy, K. (1988). Size dependentlymphatic short-term clearance of amosite fibers in the lung. Ann. Occup.
Hyg. 32, 149-156.
Oberdorster, G., Ferin, I., Marcello, N. C , and Meinhold, S. H. (1983).Effect of intrabronchiallly instilled amosite on lavageable lung and pleuralcells. Environ. Health Perspect. 51, 41-48.
Pott, F., Ziem, U., Reiffer, F.-J., Huth, F., Emst, H., and Mohr, U. (1987).Carcinogenicity studies on fibres, metal compounds and some other dustsin rats. Exp. Pathol. 32, 129-152.
Quinlan, T. R., Berube, K. A., Marsh, J. P., Janssen, Y. M. W., Taishi, P.,Leslie, K. O., Hemenway, D., O'Shaughnessy, P. T., Vacekk, P., andMossman, B. T. (1995). Patterns of inflammation, cell proliferation, andrelated gene expression in lung after inhalation of chrysotile asbestos.Am. J. Pathol. 147, 728-739.
Rutten, A. A. J. J. L., Bermudez, E., Mangum, J. B., Wong, B. A., Moss,O. R., and Everitt, J. I. (1994). Mesothelial cell proliferation induced byintrapleural instillation of man-made fibers in rats and hamsters. Fundam.Appl. Toxicol. 23, 107-116.
Sebastein, P., Janson, X., Gaudiechet, A., Hirsch, A., and Bignon, J. (1980).Asbestos retention in human respiratory tissues: Comparative measuresin lung parenchyma and in parietal pleura. In Biological Effects of MineralFibres (J. C. Wagner, Ed.), Wagner, IARC Scientific Publications No.30, pp. 237-246. IARC, Lyon, France.
Stanton, M. F., Layard, M., Tegeris, A., Miller, E., May, M., Morgan, E.and Smith, A. (1981). Relation of particle dimension to carcinogenicityin amphibole asbestoses and other fibrous minerals, J. Natl. Cancer Inst.67, 965-975.
Timbrell, V., and Rendall, R. E. G. (1991/1992). Preparation of the UICC
standard reference sample of asbestos, Powder Technol. 5, 279-287.
Warheit, D. B., Hartsky, M. A., McHugh, T. A., and Kelley, K. A. (1994).Biopersistence of inhaled organic and inorganic fibers in the lungs ofrats. Environ. Health Perspect. 102(Suppl. 5), 151-157.
Warheit, D. B., Driscoll, K. E., Oberdorster, G., Walter, C , Kuschner, M.,and Hesterberg, T. W. (1995). Contemporary issues in fiber toxicology.Fundam. Appl. Toxicol. 25, 171-183.
World Health Organization (1985). Reference Methods for Measuring Man-Made Mineral Fibers. WHO Regional Office, Copenhagen, Denmark.[Prepared by WHO/EURO Technical Committee for Evaluating MMMF]
Dow
nloaded from https://academ
ic.oup.com/toxsci/article-abstract/38/2/173/1633315 by guest on 04 April 2019


















