
MARKERY STRESU OKSYDACYJNEGO I PARAMETRY …arch.apsl.edu.pl/spb/pliki/nr10/01.pdf ·...
22
5 Słupskie Prace Biologiczne 10 • 2013 MARKERY STRESU OKSYDACYJNEGO I PARAMETRY BIOCHEMICZNE WE KRWI KONI BIORĄCYCH UDZIAŁ WE WSZECHSTRONNYM KONKURSIE KONIA WIERZCHOWEGO W DYNAMICE TRENINGU OXIDATIVE STRESS BIOMARKERS AND BIOCHEMICAL PARAMETERS IN THE BLOOD OF EVENTING HORSES IN THE DYNAMICS OF TRAINING Anastasiia Andriichuk 2 Halyna Tkachenko 1 Natalia Kurhaluk 1 Iryna Tkachova 2 Mykhaylo Vartovnyk 3 1 Akademia Pomorska w Słupsku Instytut Biologii i Ochrony Środowiska Zakład Zoologii i Fizjologii Zwierząt ul. Arciszewskigo 22b, 76-200 Słupsk e-mail: [email protected] [email protected] 2 National Academy of Agrarian Sciences of Ukraine Institute of Animal Science 7 Gvardeyskoi Armii Str. 3 p.d. Kulynychi, Kharkiv Region Kharkiv District, Ukraine e-mail: [email protected] 3 Dziecięco-Młodzieżowa Szkoła Sportu Konnego „BUREVISNYK” „Burevisnyk”, ul. Stryjska 119, 79000 Lwów ABSTRACT The physical exercise in athletic horses induces a series of normal physiological and biochemical adaptations. However, strenuous exercise is a potent inducer of ox- idative stress associated with disturbances in muscle homeostasis, fatigue and injury of sport horses. The aim of the current study was to determine the level of oxidative stress biomarkers, aspartate aminotransferase (AST), alanine aminotransferase (ALT), and lactate dehydrogenase (LDH) activity, as well as lactate and pyruvate concentrations in sport horses involved in eventing before and after training. All
Transcript of MARKERY STRESU OKSYDACYJNEGO I PARAMETRY …arch.apsl.edu.pl/spb/pliki/nr10/01.pdf ·...

5
S ł u p s k i e P r a c e B i o l o g i c z n e 10 •••• 2013
MARKERY STRESU OKSYDACYJNEGO I PARAMETRY BIOCHEMICZNE WE KRWI KONI BIORĄCYCH UDZIAŁ
WE WSZECHSTRONNYM KONKURSIE KONIA WIERZCHOWEGO W DYNAMICE TRENINGU
OXIDATIVE STRESS BIOMARKERS AND BIOCHEMICAL PARAMETERS IN THE BLOOD OF EVENTING HORSES
IN THE DYNAMICS OF TRAINING
Anastasiia Andriichuk2 Halyna Tkachenko1 Natalia Kurhaluk1 Iryna Tkachova2
Mykhaylo Vartovnyk3
1 Akademia Pomorska w Słupsku Instytut Biologii i Ochrony Środowiska Zakład Zoologii i Fizjologii Zwierząt ul. Arciszewskigo 22b, 76-200 Słupsk e-mail: [email protected] [email protected] 2 National Academy of Agrarian Sciences of Ukraine Institute of Animal Science 7 Gvardeyskoi Armii Str. 3 p.d. Kulynychi, Kharkiv Region Kharkiv District, Ukraine e-mail: [email protected] 3 Dziecięco-Młodzieżowa Szkoła Sportu Konnego „BUREVISNYK” „Burevisnyk”, ul. Stryjska 119, 79000 Lwów
ABSTRACT The physical exercise in athletic horses induces a series of normal physiological
and biochemical adaptations. However, strenuous exercise is a potent inducer of ox-idative stress associated with disturbances in muscle homeostasis, fatigue and injury of sport horses. The aim of the current study was to determine the level of oxidative stress biomarkers, aspartate aminotransferase (AST), alanine aminotransferase (ALT), and lactate dehydrogenase (LDH) activity, as well as lactate and pyruvate concentrations in sport horses involved in eventing before and after training. All

6
horses were in a regular systematic training and had the same diet. Significant in-creases in the thiobarbituric acid reactive substrates (TBARS) level in the blood of horses after exercise was observed. In the present study, the increased lipid peroxi-dation, in horses after exercise may indicate the development of oxidative stress caused strenuous exercises. TBARS level in the blood and the plasma of sport hors-es immediately after training showed an increase by 67% and 45% (p<0.05), respec-tively. There were no significant differences in erythrocyte TBARS level between resting period and after exercises. Significant decreases by 8% (р<0.05) in the aldehyde derivatives of protein oxidation in the plasma after training was noted. Ex-ercise can induce the activity of the proteasome complex, which is significantly in-volved in the degradation of oxidatively modified proteins. ALT and AST activity was 4% and 15% (p<0.05) lower in horses after training compared to the rest peri-od. In post-exercise horses, lactate level was higher by 12% (p<0.05) compared to rest period. No statistically significant differences in pyruvate level between the rest period and after training were observed. Significant increase of lactate concentration after training indicates the level of contribution of anaerobic glycolysis to the total energy supply of muscle activity in eventing horses. The preventive effect of regular exercise leads to adaptation to prolonged exercises, which is accompanied by in-crease of oxidative stress-induced adaptation and changes in redox homeostasis, increased antioxidant defenses, lower oxidative damage, and increased resistance to oxidative stress. Regularly performed exercise might induce an adaptive enhance-ment in skeletal muscle and erythrocytes of the defense mechanisms that protect them against oxidative stress. The level of oxidative stress markers and activity of aminotransferases, lactate dehydrogenase and concentration of lactate and pyruvate in the blood of sport horses can be sensitive and informative parameters for the as-sessment of equine athletic performance.
Słowa kluczowe: stres oksydacyjny, aminotransferazy, dehydrogenaza mleczanowa, mleczan, pirogronian, trening systematyczny, Wszechstronny Konkurs Konia Wierz-chowego (WKKW) Key words: oxidative stress, aminotransferases, lactate dehydrogenase, lactate, pyruvate, systematic training, eventing
WPROWADZENIE
Wszechstronny Konkurs Konia Wierzchowego (WKKW) to jeden z konkursów
klasycznej dyscypliny hippicznej (Banasiak i Modzelewska 2003). Składa się z trzech dyscyplin: ujeżdżania, próby terenowej oraz skoków (Williams i Burk 2012). Konkursy WKKW rozgrywane są w trzech klasach (C, N i P) i trwają trzy dni (Marlin i Nankervis 2002). Odbywają się również tzw. konkursy WKKW kom-binowane. Trwają one jeden, dwa lub trzy dni. Próba terenowa w konkursach WKKW odbywa się na czterech odcinkach. Pierwszy to tzw. drogi i ścieżki. Drugi to tzw. steepplechase, czyli bieg z przeszkodami. Długość tego odcinka wynosi ok. 3,2 km. Później znowu drogi i ścieżki. Czwartym etapem jest cross, którego długość wynosi ok. 7,2 km i zawiera 32 przeszkody. Etapy pierwszy i trzeci liczą razem ok.

7
15,2 km. Cała próba terenowa to trasa o długości 25,6 km do pokonania przez konia (Marlin i Nankervis 2002).
Wiadomo, że podczas długotrwałej adaptacji organizmu konia do wysiłku fi-zycznego o różnej intensywności dochodzi do specyficznych zmian reakcji metabo-licznych skierowanych na obniżenie uwalniania energii z ciepłem, a jednocześnie kumulowania jej w postaci związków makroergicznych (ATP) (Anderson 1975, Muñoz i in. 2002). Zmiana metabolizmu energetycznego podczas treningu konia wywołuje zwiększenie w mięśniach szkieletowych stężenia takich źródeł energii, jak fosforan kreatyniny i glikogen, powoduje wzrost aktywności enzymów glikolizy i cyklu Krebsa oraz β-oksydacji kwasów tłuszczowych etc. (Marlin i Nankervis 2002). Metabolity odpowiedzialne za dostarczanie energii do mięśni szkieletowych oraz enzymy, które odzwierciedlają tendencję zmian biochemicznych podczas wy-siłku fizycznego, mogą służyć jako indykatory poziomu kondycji fizycznej koni sportowych (Harris i in. 1998, McGowan i in. 2002, Padalino i in. 2007). Do takich markerów należą mleczan i pirogronian, aktywność dehydrogenazy mleczanowej, aminotransferazy alaninowej i asparaginianowej (Szarska 1994, Harris i in. 1998, Kedzierski i in. 2009). Zmiany aktywności wyżej wymienionych enzymów w dy-namice treningu koni sportowych odzwierciedlają specyficzne zmiany w reakcjach metabolicznych (Davies i Pethick 1983, Harris i in. 1998, McGowan i in. 2002, Padalino i in. 2007).
Wraz z procesem utleniania substratów energetycznych w mitochondriach pod-czas treningu zachodzi również proces powstawania reaktywnych form tlenu (RFT) (Deaton i Marlin 2003). Przy dużej ilości RFT, a zmniejszonej skuteczności działa-nia układów antyoksydacyjnych i proteolitycznych dochodzi do kumulacji utlenio-nych produktów białkowych (Deaton i Marlin 2003, Radak i in. 2008). Zmodyfiko-wane oksydacyjnie białka wykryto w licznych tkankach i wykazano, że stres oksy-dacyjny i modyfikacja białek zachodząca pod wpływem RFT odgrywają rolę w wielu patogenetycznych mechanizmach chorób (Stadtman i Levine 2000). Jednym z waż-niejszych procesów biologicznych związanych z działaniem RFT jest peroksydacja lipidów (Yagi 1992). Kaskadowy proces utleniania obecnych w lipidach nienasyco-nych kwasów tłuszczowych, w którym powstają nadtlenki tych związków, zapewnia również ciągłą dostawę wolnych rodników, inicjujących kolejne reakcje peroksyda-cji (Yagi 1992, Deaton i Marlin 2003). Peroksydacji ulegają przede wszystkim resz-ty wielonienasyconych kwasów tłuszczowych, wchodzące w skład fosfolipidów, które są podstawowym składnikiem budulcowym błon biologicznych. Zaburzenia metabolizmu komórkowego mogą prowadzić do trwałych zmian w strukturze DNA, RNA, białek, lipidów i cukrów, czego konsekwencją jest najczęściej utrata ich bio-logicznych funkcji i w dalszej kolejności rozwój procesów chorobowych (Deaton i Marlin 2003, Kirschvink i in. 2008).
Produkcja RFT w nadmiarze stanowi powód obniżenia sportowej kondycji konia, zmęczenia, uszkodzenia mięśni i stawów, zaburzenia funkcjonowania układu serco- wo-naczyniowego i oddechowego, uszkodzeń neurologicznych etc. (Chiaradia i in. 1998, Art i Lekeux 2005). Nadmiar RFT i obniżenie obrony antyoksydacyjnej w wyniku intensywnego wysiłku fizycznego jest powodem inicjacji stresu oksyda-cyjnego (Deaton i Marlin 2003, Art i Lekeux 2005, Kinnunen i in. 2005, Kirschvink i in. 2008).

8
Stres oksydacyjny zależy od intensywności wysiłku, a także od czasu trwania ak-tywności oraz poziomu wytrenowania koni (Andriichuk i in. 2012, 2013). Zmiany wskaźników stresu oksydacyjnego pojawiają się też w różnym czasie, nie zawsze bezpośrednio po wysiłku fizycznym (Kirschvink i in. 2008). Poprzednie badania wykazały, że u koni sportowych, które znajdują się w długotrwałym treningu, po-ziom markerów stresu oksydacyjnego wraz z aktywacją enzymów antyoksydacyj-nych jest znacznie niższy po wysiłku fizycznym w porównaniu ze stanem w spo-czynku (Tkachenko i in. 2011, Andriichuk i in. 2012, 2013).
Systematyczny trening koni biorących udział w WKKW ukierunkowany jest na rozwój wytrzymałości, udoskonalenia nerwowo-regulatorowych mechanizmów funk-cjonowania układu sercowo-naczyniowego oraz oddechowego, a także wytrzymało-ści mięśni i in. WKKW to najtrudniejszy typ sportu hippicznego, który wymaga od konia wysokiego poziomu kondycji i ogromnych funkcjonalnych zdolności organi-zmu oraz dużej wytrzymałości (Art i Lekeux 2005, Williams i Burk 2012).
Analiza zmian markerów stresu oksydacyjnego i zmian metabolicznych pod wpływem wysiłku fizycznego jest bardzo ważną predyspozycją do badania adapta-cyjnych możliwości i poziomu kondycji koni sportowych (Szarska 1994, Deaton i Marlin 2003, Art i Lekeux 2005, Kirschvink i in. 2008, Kedzierski i in. 2009, So-ares i in. 2011, Williams i Burk 2012). Za pomocą analizy dynamiki zmian marke-rów stresu oksydacyjnego i zmian metabolicznych można ocenić stopień wytreno-wania konia oraz uzasadnić czas i tryb treningu. Zwracając uwagę na aktualność podjętego problemu, celem naszych badań było oznaczenie poziomu markerów stre-su oksydacyjnego, aktywności aminotransferaz i dehydrogenazy mleczanowej, jak również zmian ilościowych mleczanu i pirogronianu jako wskaźników wykorzysty-wanych w WKKW w dynamice treningu dla oceny funkcjonalnych zdolności orga-nizmu koni sportowych.
MATERIAŁ I METODY BADAŃ Obiekt badań stanowiło 9 koni sportowych (klacze, ogiery i wałachy) rasy ukra-
ińskiej wierzchowej, w wieku 6-12 lat, które czynnie biorą udział w WKKW. Wszystkie osobniki były zdrowe, bez objawów patologicznych. Konie pochodziły ze stadniny Dziecięco-Młodzieżowej Szkoły Sportu Konnego „BUREVISNYK” (Lwów, Ukraina). Brały one czynny udział w zawodach miejscowych, krajowych i międzynarodowych. Warunki hodowli koni były jednakowe, ponadto zwierzęta poddawano ciągłemu, systematycznemu treningowi sportowemu. Codzienny trening zawierał: jazdę stępem (5 min), jazdę kłusem (10 min), jazdę stępem (10 min), jazdę kłusem (10 min) z różnymi chodami, zwrotami, zatrzymaniem i zmianą kierunku jazdy (10 min), jazdę stępem (5 min), jazdę galopem ze skokami przez przeszkody (wys. 90-120 cm i szer. 100-120 cm) 30 min, jazdę stępem (20 min). Całkowity czas treningu wynosił 1,5 godziny (Nerodenko 2009).
Krew zwierząt pobrano z żyły szyjnej zewnętrznej do próbówek z EDTA w sta-nie spokoju, przed treningiem i natychmiast po wysiłku fizycznym. Krew wirowano przy prędkości 3000 obr./min w czasie 10 min. Suspensję erytrocytów otrzymano trzykrotnym płukaniem osadu schłodzonym roztworem fizjologicznym. Intensyw-

9
ność peroksydacji lipidów określano według stężenia produktów reagujących z kwa-sem 2-tiobarbiturowym (TBARS) w pełnej krwi, osoczu i suspensji erytrocytów (Ka- myshnikov 2004). Poziom aldehydowych i ketonowych pochodnych oksydacyjnej modyfikacji białek oznaczono w suspensji erytrocytów i osoczu w reakcji z 2,4-dini- trofenylohydrazyną (Levine i in. 1990, Dubinina i in. 1995), a aktywność amino-transferaz w surowicy krwi w reakcji z 2,4-dinitrofenylohydrazyną (Kamyshnikov 2004). Aktywność dehydrogenazy mleczanowej w surowicy określono w reakcji oksydacji L-mleczanu do pirogronianu z NAD+ w środowisku zasadowym (Ka-myshnikov 2004). Stężenia mleczanu i pirogronianu oznaczono w nieenzymatycznej metodzie w każdej próbie pełnej krwi: stężenia mleczanu w reakcji z hydrochino-nem, a pirogronianu – z p-dimetylaminobenzaldehydem (Herasimov i Plaksina 2000). Całkowitą zdolność antyoksydacyjną (CZA) osocza i erytrocytów określono w reakcji inhibicji askorbat- i żelazo-indukowanej oksydacji Tween 80 do dialdehy-du malonowego (Galaktionova i in. 1998). Otrzymane wyniki przeanalizowano sta-tystycznie programem STATISTICA 8.0 (StatsSoft, Polska). Normalność rozkładu danych sprawdzono za pomocą kryteriów Shapiro-Wilka i Lillieforsa. Obliczono średnią i błąd statystyczny średniej. Istotność różnic pomiędzy grupami zwierząt w stanie spokoju przed treningiem i po treningu oznaczono według kryterium Wil-koxona. Korelacyjne zależności między badanymi parametrami określono według rang Spearmana (Zar 1999).
WYNIKI BADAŃ I ICH OMÓWIENIE Pomiary stężenia nadtlenków lipidów, a przede wszystkim stężenia związków,
które powstają na skutek degradacji utlenionych lipidów, są często wykorzystywane jako wskaźniki stresu oksydacyjnego (Deaton i Marlin 2003, Kinnunen i in. 2005, Kirschvink i in. 2008). W większości prac do oceny stresu oksydacyjnego stosuje się pomiar stężenia TBARS (Jagrič-Munih i in. 2012). Poziom peroksydacji lipidów we krwi koni w stanie spokoju był najwyższy w erytrocytach i pełnej krwi – odpowied-nio (14,59±0,77) mkmol/L i (11,71±0,50) mkmol/L (ryc. 1). Natomiast najmniejszy poziom TBARS zanotowano w surowicy – (4,87±0,33) mkmol/L (ryc. 1).
Stres oksydacyjny można określić jako zaburzenie równowagi między natężeniem procesów oksydacyjnych, które indukują powstawanie RFT, a przeciwdziałającym systemem obronnym antyoksydacyjnym (Deaton i Marlin 2003). Wzrost wytwarzania wolnych rodników, zmniejszenie aktywności systemów enzymatycznych odpowie-dzialnych za ich usuwanie i spadek aktywności antyoksydacyjnej powodują, że nie są one skutecznie usuwane przez mechanizmy obronne organizmu (Yagi 1992). Nasilony lub długo utrzymujący się stres oksydacyjny jest bardzo szkodliwy dla komórek, po-nieważ może prowadzić do trwałych zmian w strukturze ważnych biologicznie ma-krocząsteczek (DNA, białek, cukrów i in.) (Deaton i Marlin 2003). Zmiany te prowa-dzą do zaburzeń ich funkcji biologicznych, co z kolei stanowi przyczynę nieprawi-dłowości w metabolizmie komórkowym. Integralność i stabilność struktur biologicz-nych są warunkiem prawidłowego funkcjonowania komórek. Uszkodzenia lipidów błon komórkowych mogą prowadzić do zaburzenia procesów metabolicznych w wy-niku rozwoju peroksydacji lipidów (Yagi 1992, Kulbacka i in. 2009).

10
Ryc. 1. Zmiany poziomu produktów reagujących z kwasem 2-tiobarbiturowym (TBARS) ja-ko markerów peroksydacji lipidów w pełnej krwi, osoczu i suspensji erytrocytów koni w dy-namice treningu (n=9) * – zmiany statystycznie istotne między wartościami w stanie spokoju i po treningu (p<0,05) Fig. 1. TBARS content in the blood, plasma and erythrocytes of horses at the rest and after training period (n=9) * – statistically significant differences between TBARS level at the rest and after training pe-riod (p<0.05)
Peroksydacja lipidów to lawinowy, wolnorodnikowy proces utleniania obecnych w lipidach nienasyconych kwasów tłuszczowych, w którym powstają nadtlenki tych związków (Yagi 1992). Peroksydacji ulegają przede wszystkim reszty wielonienasy-conych kwasów tłuszczowych, wchodzące w skład fosfolipidów, które są głównym składnikiem budulcowym błon komórkowych (Alessio i Goldfarb 1988). Jednym z wielu związków wytwarzanych w procesie peroksydacji wielonienasyconych kwa-sów tłuszczowych jest dialdehyd malonowy (malondialdehyde, MDA) (Nair i in. 2008). Powstaje on między innymi w organizmach żywych podczas procesów pe-roksydacji lipidów komórkowych (Gaweł i in. 2004). MDA może stanowić ważny wskaźnik intensywności procesów peroksydacji lipidów zachodzących w organi-zmie (Kulbacka i in. 2009, Gaweł i in. 2004).
Istotne okazały się doniesienia dotyczące aktywności MDA i jego roli w organi-zmie. Udowodniono bowiem dużą reaktywność MDA względem związków biolo-gicznie ważnych. Z miejsc, w których powstaje, może przedostawać się do odle-głych tkanek i tam, dzięki możliwości tworzenia wiązań kowalencyjnych z czą-steczkami molekuł, modyfikować ich strukturę, a w konsekwencji zmieniać ich wła-ściwości. W badaniach in vitro stwierdzono, że MDA uczestniczy w inaktywacji en-zymów oraz pośrednio w syntezach białek, DNA i RNA, co leży u podstaw patoge-nezy wielu schorzeń (Gaweł i in. 2004). MDA modyfikuje właściwości fizyczne błon komórkowych, powodując zakłócenie hydrofobowości lipidowego wnętrza

11
i naruszenie dwuwarstwowej struktury błony. Wpływa zatem na struktury komór-kowe, co prowadzi do zaburzenia ich prawidłowych funkcji, a w efekcie – do dys-funkcji poszczególnych narządów (Gaweł i in. 2004). Zarówno endogenny, jak i eg-zogenny MDA ulega w organizmie przemianom pod wpływem dehydrogenazy alde-hydowej do semialdehydu malonowego (MSA), z którego po spontanicznej dekar-boksylacji powstają aldehyd octowy i dwutlenek węgla. Aldehyd octowy jest utle-niany przez dehydrogenazę aldehydową do kwasu octowego, a następnie przekształ-cany przez tiokinazę do acetylo-CoA. Niewielka ilość MDA może być metabolizo-wana do kwasu malonowego. Ten z kolei z udziałem tiokinazy przekształcany jest do malonylo-CoA. W kolejnej reakcji malonylo-CoA ulega dekarboksylacji do ace-tylo-CoA (Gaweł i in. 2004).
Z literatury wiadomo, że najbardziej wrażliwe na stres oksydacyjny są erytrocyty, głównie dlatego, iż biorą udział w transporcie tlenu, przez co są bezpośrednio narażo-ne na jego działanie (Çimen Burak 2008). Zawarta w erytrocycie hemoglobina zawie-ra jony żelaza (II), które umożliwiają zachodzenie reakcji Fentona oraz katalizują pe-roksydację lipidów (Zapora i Jarocka 2013). RFT w erytrocycie mogą być również tworzone w wyniku nieenzymatycznej i enzymatycznej degradacji hemu. Proces nie-enzymatycznej degradacji hemu inicjuje m.in. nadtlenek wodoru, natomiast proces enzymatyczny zachodzi pod wpływem oksygenazy hemowej. W obu przypadkach w wyniku degradacji hemu tworzy się biliwerdyna, tlenek węgla (II) oraz uwalniane są jony żelaza Fe2+. Powstałe produkty degradacji hemu mogą inicjować procesy oksydacyjne w erytrocycie. Zasadnicze znaczenie w tworzeniu RFT w erytrocytach ma proces degradacji hemu, który może zachodzić zarówno enzymatycznie, jak i nie-enzymatycznie. Anionorodnik ponadtlenkowy O2-, pierwszy i podstawowy spośród RFT, powstaje w erytrocycie głównie w wyniku autooksydacji hemoglobiny, tj. spon-tanicznego przejścia oksyhemoglobiny w methemoglobinę (Zapora i Jarocka 2013).
Poza procesami autooksydacji hemoglobiny białko to narażone jest na oddziały-wanie anionorodników ponadtlenkowych, wytwarzanych w wyniku aktywacji in-nych morfotycznych elementów krwi, głównie fagocytów. Na powierzchni komórek fagocytarnych, m.in. neutrofilów, eozynofilów czy makrofagów, znajduje się system zależny od oksydazy NADPH, tworzący anionorodnik ponadtlenkowy (Bylund i in. 2010). Akumulacja methemoglobiny prowadzi do powstania hemichromów będą-cych nieaktywną postacią hemoglobiny. W początkowym etapie ich tworzenia do-chodzi do utlenienia grup tiolowych hemoglobiny, czego konsekwencją są zmiany konformacyjne łańcuchów globiny (Sugawara i in. 2003, Telen i Kaufman 2004). Wykazano, iż przemianom hemoglobiny towarzyszy degradacja hemu i utrata żelaza hemowego, co w konsekwencji prowadzi do precypitacji zdenaturowanych cząste-czek hemoglobiny i powstawania złogów, tzw. ciałek Heinza. Powstałe produkty de-gradacji hemu inicjują procesy oksydacyjne w erytrocycie (Dani i in. 2004, Pradines i in. 2005). Obecność zarówno O2-, jak i H2O2 przyczynia się do wytworzenia rodni-ków hydroksylowych w reakcji Habera-Weissa, katalizowanej przez uwolnione pod-czas degradacji hemu jony żelaza (Nagababu i in. 2003). Ponadto uwolniony hem hamuje aktywność enzymów cytosolowych oraz nieswoiście reaguje z białkami bło-nowymi erytrocytu, powodując utlenienie ich grup sulfhydrylowych (Nagababu i in. 2010). W efekcie tworzą się niepożądane wiązania krzyżowe, co prowadzi do utraty funkcji białek i ich agregacji (Zapora i Jarocka 2013).

12
Niezależnie od procesów nieenzymatycznych hem ulega również degradacji en-zymatycznej. Najważniejszą rolę w tym procesie odgrywa oksygenaza hemowa (HO) [EC 1.14.99.3] (Zapora i Jarocka 2013). Głównym efektem działania oksyge-nazy hemowej HO jest proces degradacji wolnego hemu, w którym HO z udziałem tlenu cząsteczkowego i NADPH rozkłada hem do tlenku węgla (II) [CO], jonów że-laza Fe2+ oraz rozpuszczalnej biliwerdyny, przekształcanej następnie przez reduktazę biliwerdyny w nierozpuszczalną bilirubinę (Ryter i in. 2006, Florczyk i in. 2008). Produkty rozkładu hemu w wysokich stężeniach są toksyczne. Zarówno jony Fe2+, jak i wolny hem, uczestnicząc w reakcji Fentona, katalizują powstawanie RFT (Wag- ner i in. 2003, Florczyk i in. 2008, Zapora i Jarocka 2013). Powstała z rozkładu he-mu biliwerdyna zostaje natychmiast przekształcona w bilirubinę. Związek ten w po-staci wolnej (niezwiązany z glukuronianem) jest toksyczny. Po przekroczeniu barie-ry krew-mózg wywołuje żółtaczkę jąder podkorowych mózgu, co skutkuje uszko-dzeniem komórek nerwowych (Wagner i in. 2003, Zapora i Jarocka 2013).
Nawet w znikomym stężeniu oksyhemoglobiny nieznaczna autooksydacja he-moglobiny powoduje nagły wzrost RFT w erytrocytach. Produkcja anionorodnika ponadtlenkowego wywołuje oksydację hemoglobiny do methemoglobiny (Zapora i Jarocka 2013). Stąd zanotowany przez nas najwyższy poziom TBARS w erytrocy-tach krwi koni w stanie spokoju wynika ze strukturalnych i funkcjonalnych osobli-wości tych morfotycznych elementów krwi.
Jednym z procesów prowadzących do podwyższenia stężenia oksydantów w or-ganizmie jest wysiłek fizyczny. Zwiększone podczas wysiłku fizycznego zużycie tlenu pociąga za sobą wzrost produkcji anionorodnika ponadtlenkowego i innych RFT (Art i Lekeux 2005, Williams i Burk 2012). Konsekwencją ubytku naturalnych antyoksydantów i nadmiaru oksydantów w wyniku wysiłku fizycznego jest podwyż-szony poziom peroksydacji lipidów komórek mięśni (Kirschvink i in. 2008). Po-wstające podczas wysiłku fizycznego RFT powodują nie tylko uszkodzenie mięśni, ale także oksydacyjne uszkodzenia innych struktur komórkowych (Deaton i Marlin 2003, Kirschvink i in. 2008). W wyniku naszych badań po treningu zanotowano istotne podwyższenie poziomu TBARS w pełnej krwi o 67% (p=0,001), w osoczu – o 45% (p=0,000), natomiast w suspensji erytrocytów nie stwierdzono istotnych zmian (ryc. 1). Nasze wyniki pokrywają się z danymi literaturowymi, według któ-rych u koni trenowanych na wytrzymałość pokazano podwyższenie stężenia marke-rów stresu oksydacyjnego we krwi po treningu (Kirschvink i in. 2008, Gondim i in. 2009). Długodystansowe wyścigi (210 km) powodują podwyższenie poziomu TBARS we krwi koni, zaś po trzech dobach od zakończenia wyścigów poziom ten wraca on do normy (Gondim i in. 2009). Podsumowując nasze wyniki, zaznaczamy, że podwyższenie poziomu TBARS we krwi koni świadczy o stresie oksydacyjnym wywołanym wysiłkiem fizycznym.
Jednak największym zagrożeniem dla życia komórek w warunkach stresu oksy-dacyjnego jest utlenianie grup tiolowych w błonach komórkowych, co może powo-dować ich dezintegrację i zwiększenie przepuszczalności (Yagi 1992, Levine 2002). Reakcje RFT z białkami prowadzą nie tylko do utleniania białek, ale także do po-wstawania w białkach grup redukujących, mających zdolność do redukcji cytochro-mu c i metali (Levine 2002, Radak i in. 2008). Patogeneza powstania takich grup zachodzi w wyniku uszkodzenia aminokwasów aromatycznych (Radak i in. 2008).
13
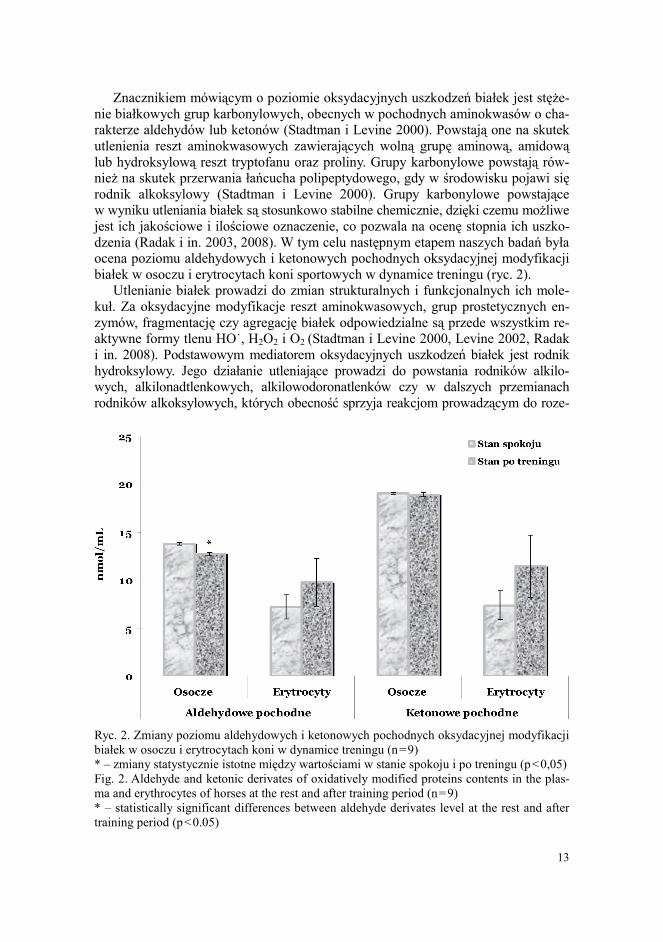
Znacznikiem mówiącym o poziomie oksydacyjnych uszkodzeń białek jest stęże-nie białkowych grup karbonylowych, obecnych w pochodnych aminokwasów o cha- rakterze aldehydów lub ketonów (Stadtman i Levine 2000). Powstają one na skutek utlenienia reszt aminokwasowych zawierających wolną grupę aminową, amidową lub hydroksylową reszt tryptofanu oraz proliny. Grupy karbonylowe powstają rów-nież na skutek przerwania łańcucha polipeptydowego, gdy w środowisku pojawi się rodnik alkoksylowy (Stadtman i Levine 2000). Grupy karbonylowe powstające w wyniku utleniania białek są stosunkowo stabilne chemicznie, dzięki czemu możliwe jest ich jakościowe i ilościowe oznaczenie, co pozwala na ocenę stopnia ich uszko-dzenia (Radak i in. 2003, 2008). W tym celu następnym etapem naszych badań była ocena poziomu aldehydowych i ketonowych pochodnych oksydacyjnej modyfikacji białek w osoczu i erytrocytach koni sportowych w dynamice treningu (ryc. 2).
Utlenianie białek prowadzi do zmian strukturalnych i funkcjonalnych ich mole-kuł. Za oksydacyjne modyfikacje reszt aminokwasowych, grup prostetycznych en-zymów, fragmentację czy agregację białek odpowiedzialne są przede wszystkim re-aktywne formy tlenu HO˙, H2O2 i O2 (Stadtman i Levine 2000, Levine 2002, Radak i in. 2008). Podstawowym mediatorem oksydacyjnych uszkodzeń białek jest rodnik hydroksylowy. Jego działanie utleniające prowadzi do powstania rodników alkilo-wych, alkilonadtlenkowych, alkilowodoronatlenków czy w dalszych przemianach rodników alkoksylowych, których obecność sprzyja reakcjom prowadzącym do roze-
Ryc. 2. Zmiany poziomu aldehydowych i ketonowych pochodnych oksydacyjnej modyfikacji białek w osoczu i erytrocytach koni w dynamice treningu (n=9) * – zmiany statystycznie istotne między wartościami w stanie spokoju i po treningu (p<0,05) Fig. 2. Aldehyde and ketonic derivates of oxidatively modified proteins contents in the plas-ma and erythrocytes of horses at the rest and after training period (n=9) * – statistically significant differences between aldehyde derivates level at the rest and after training period (p<0.05)

14
rwania łańcucha polipeptydowego (Stadtman i Levine 2000). Najbardziej podatne na działanie RFT są reszty aminokwasów aromatycznych i siarkowych. Szczególną wraż-liwość wykazują tyrozyna, tryptofan, cysteina i metionina (Stadtman i Levine 2000, Levine 2002). Utlenianie przez RFT aminokwasów z wolną grupą aminową, amidową lub hydroksylową prowadzi do powstania pochodnych karbonylowych. Pochodne karbonylowe mają zdolność do reagowania z wolnymi grupami aminowymi reszt li-zyny. Reakcja ta prowadzi do powstawania w białku wiązań krzyżowych (Stadtman i Levine 2000). RFT mogą indukować peroksydację białek, która powoduje powsta-wanie nadtlenków białek i nadtlenków aminokwasów (Radak i in. 2008). Wykazują one również utleniające działanie w stosunku do grup prostetycznych w enzymach. Mogą one utleniać np. węglowodany czy jony metali zawarte w białkach, co często prowadzi do zaburzenia ich funkcji biologicznych (Stadtman i Levine 2000). Wykaza-no, że pod wpływem RFT dochodzi do utraty aktywności niektórych enzymów, m.in. dehydrogenazy gliceroaldehydofosforanowej i dehydrogenazy glukozo-6-fosforano- wej (Stadtman i Levine 2000, Levine 2002, Radak i in. 2008).
W wyniku naszych badań stwierdzono wyższy poziom aldehydowych i ketono-wych pochodnych oksydacyjnej modyfikacji białek w osoczu w porównaniu z su-spensją erytrocytów (ryc. 2). Wiadomo, że poziom oksydacyjnej modyfikacji białek nie zawsze jest związany z intensywnością peroksydacji lipidów i/lub oksydacyjne-go uszkodzenia DNA, a może być determinowany aktywnością obrony antyoksyda-cyjnej i reparacją struktur biologicznych (Radak i in. 2003). Zanotowany przez nas wysoki poziom oksydacyjnej modyfikacji białek w osoczu wskazuje na zmiany oksydacyjne w innych tkankach i narządach, a nie tylko w krwinkach czerwonych. Jednakże po treningu stwierdzono zmniejszenie poziomu aldehydowych pochod-nych OMB w osoczu (o 8%, p<0,05) (ryc. 2). Zanotowano brak istotnych zmian w stężeniu ketonowych pochodnych OMB osocza i erytrocytów. Poziom aldehydo-wych i ketonowych pochodnych oksydacyjnej modyfikacji białek w erytrocytach koni po treningu wzrósł odpowiednio o 35 i 55% (p>0,05).
Zmiany oksydacyjne białek są nieodłącznym elementem metabolizmu tlenowego komórki. Za oksydacyjne modyfikacje białek odpowiadają przede wszystkim: nad-tlenek wodoru (H2O2), rodnik hydroksylowy (·OH) oraz anionorodnik ponadtlenko-wy (O2·–). Mimo że wszystkie reszty aminokwasowe obecne w białkach są podatne na utlenianie, największą wrażliwość na działanie RFT wykazują: cysteina, metioni-na, tyrozyna i tryptofan (Rutkowski i in. 2007). W wyniku utlenienia reszt amino-kwasowych w łańcuchu polipeptydowym może dojść do jego rozerwania, utworze-nia wiązań krzyżowych w obrębie tego samego lub kilku łańcuchów polipeptydo-wych, a także do modyfikacji reszt aminokwasowych. W wyniku tego procesu wiele białek może stracić lub zwiększyć swoją aktywność biologiczną. Utlenione białka łatwo tworzą agregaty, które z kolei mogą hamować układy enzymatyczne odpowie-dzialne za ich degradację (Radak i in. 2003, 2008).
Działanie RFT może przyczynić się do utlenienia białek, tłuszczów i DNA, w kon-sekwencji prowadząc do uszkodzenia tkanek (Deaton i Marlin 2003). Toksyczne produkty reakcji utleniania działają cytostatycznie na komórkę, powodując uszko-dzenie błon komórkowych oraz prowadząc ją do śmierci na drodze apoptozy lub ne-krozy (Rutkowski i in. 2007). Wielu autorów podaje, że oksydacyjne uszkodzenie białek, które polega m.in. na modyfikacji reszt aminokwasowych, agregacji lub

15
fragmentacji cząsteczki i oksydacyjnej modyfikacji grup prostetycznych enzymów, prowadzi w głównej mierze do zaburzenia aktywności biologicznej komórki (Stadt- man i Levine 2000, Levine 2002, Radak i in. 2003, 2008, Rutkowski i in. 2007). Lecz mając na uwadze wszystkie ważne funkcje, jakie białka spełniają w każdym żywym organizmie, ich modyfikacje mogą spowodować wystąpienie reakcji patolo-gicznych prowadzących do dezorganizowania funkcjonowania biologicznego (Rut-kowski i in. 2007).
Powstające w wyniku modyfikacji oksydacyjnych agregaty są oporne na degra-dację, co przy zmniejszonej wydajności działania mechanizmów naprawczych sprzyja gromadzeniu się zmienionych białek w komórkach i prowadzi do stopniowej utraty ich biochemicznych i fizjologicznych funkcji (Rutkowski i in. 2007). Przy dużej ilości RFT, a zmniejszonej skuteczności działania układów antyoksydacyjnych i proteolitycznych, dochodzi do akumulacji utlenionych produktów białkowych. Zmodyfikowane oksydacyjnie białka wykryto w licznych tkankach i wykazano, że stres oksydacyjny i modyfikacja białek zachodząca pod wpływem RFT odgrywają rolę w patogenezie wielu schorzeń (Radak i in. 2003, 2008).
Badania wykazały ponadto, iż w reakcji H2O2 i methemoglobiny powstają rów-nież rodniki białkowe, które mogą następnie reagować z tlenem, w wyniku czego powstają rodniki nadtlenkowe białek, a także mogą się przyczynić do rozkładu lipo-filowych pochodnych choleglobiny, które wbudowane między lipidy błony komór-kowej indukują dalsze uszkodzenia oksydacyjne (Sztiller i in. 2006, Zapora i Jaroc-ka 2013). Z kolei uwodornienie anionorodnika znajdującego się wewnątrz „kiesze-ni” hemowych prowadzi do wytworzenia rodnika wodoronadtlenkowego (·OOH), będącego inicjatorem procesu peroksydacji lipidów w błonie erytrocytarnej (Naga-babu i Rifkind 2000, Zapora i Jarocka 2013). Ponieważ głównym białkiem erytrocy-tów jest hemoglobina, zanotowane przez nas zwiększenie poziomu pochodnych oksydacyjnej modyfikacji białek wiąże się z oksydacją reszt aminokwasowych łań-cuchów globiny w wyniku działania RFT pod wpływem wysiłku fizycznego. Zało-żenie to jest zgodne z wynikami poprzednich naszych badań, w których ustalono, że średnio intensywne ćwiczenia fizyczne u koni rasy holsztyńskiej powodowały znaczne zwiększenie poziomu oksydacyjnej modyfikacji białek (Andriichuk i in. 2013).
Jednakże utlenione reszty aminokwasowe białek pod wpływem działania RTF pełnią również funkcję adaptacyjną do aktywności fizycznej. Oksydacyjna mody- fikacja białek towarzyszy aktywacji kompleksów proteasomowych, które selektyw-nie degradują utlenione białka (Radak i in. 2008). Zanotowane przez nas znaczne zmniejszenie poziomu aldehydowych pochodnych w osoczu koni po treningu wska-zuje na wyraźną aktywację procesu degradacji proteolitycznej reszt aminokwaso-wych utlenionych białek krwi.
Mechanizm funkcjonowania zrównoważonego systemu obrony przed wolnymi rodnikami organizmu opiera się na wspólnym i synergistycznym działaniu jego składowych (Mrowicka i in. 2009). Skutecznie działającą grupę czynników ochron-nych i mechanizmów naprawczych organizmu stanowi antyoksydacyjny układ ochronny, który zabezpiecza komórki przed działaniem RFT. Ma on rozbudowany system naprawczy. Jego funkcja polega na uniemożliwieniu inicjacji reakcji utlenia-nia oraz naprawie już powstałych uszkodzeń (Yagi 1992). W skład tego systemu

16
wchodzą enzymy rozkładające RFT, a także nieenzymatyczne związki niskoczą-steczkowe, które podlegając działaniu reaktywnego tlenu, stanowią tym samym tar-czę obronną dla cząsteczek ważnych dla komórki. Związki te nazywa się antyoksy-dantami (Yagi 1992). Występują one w małych stężeniach i mogą znacznie opóźniać lub zapobiegać utlenianiu substratu. Organizm broni się przed wolnymi rodnikami także w sposób pośredni, poprzez naprawę bądź eliminację tych składników komór-ki, które zostały uszkodzone (Halliwell 1994). Sugeruje się, iż uzupełnianie pozio-mu egzogennych antyoksydantów niskocząsteczkowych, m.in. takich jak witamina A, askorbinian czy witamina E, może być sposobem na kompensowanie osłabionego działania systemu antyoksydacyjnego (Halliwell 1994). System obrony antyoksyda-cyjnej zachodzi w trzech etapach. Pierwszy polega na niedopuszczeniu do powsta-wania wolnych rodników tlenowych, za co odpowiadają enzymy antyoksydacyjne. Drugi etap stanowią „zmiatacze” przerywające łańcuchowe reakcje wolnorodniko- we. Trzeci etap odpowiada za usuwanie skutków reakcji RFT ze składnikami komó-rek i polega na odtwarzaniu prawidłowej struktury uszkodzonych cząsteczek (Yagi 1992, Halliwell 1994, Mrowicka i in. 2009).
W wyniku naszych badań zanotowano również zmiany całkowitej zdolności an-tyoksydacyjnej osocza i erytrocytów koni sportowych w dynamice treningu (ryc. 3). Całkowita zdolność antyoksydacyjna osocza zmniejszyła się o 6% (p>0,05), nato-miast w suspensji erytrocytów – zwiększyła się o 1,1% (p>0,05).
Ryc. 3. Zmiany całkowitej zdolności antyoksydacyjnej w osoczu i erytrocytach koni w dy-namice treningu (n=9) * – zmiany statystycznie istotne między wartościami w stanie spokoju i po treningu (p<0,05) Fig. 3. Total antioxidant capacity in the plasma and erythrocytes of horses at the rest and after training period (n=9) * – statistically significant differences between total antioxidant capacity at the rest and after training period (p<0.05)

17
Dynamika aktywności enzymów uczestniczących w zmianach metabolicznych podczas treningu koni ujawnia procesy mobilizacji i wykorzystania głównych sub-stratów energetycznych w organizmie. W związku z tym kolejnym etapem naszych badań było określenie aktywności aminotransferaz i dehydrogenazy mleczanowej (ryc. 4), jak również stężenia metabolitów metabolizmu energetycznego – mleczanu i pirogronianu we krwi koni w dynamice treningu (ryc. 5).
Aminotransferaza alaninowa (ALT) jest enzymem należącym do grupy transfe-raz, przenoszącym w sposób odwracalny grupę aminową wraz z protonem i parą elektronów z aminokwasu na węgiel karbonylowy 2-oksokwasu. W wyniku reakcji substrat aminokwasowy staje się oksokwasem, a wchodzący w reakcję oksokwas – aminokwasem. W przypadku tego enzymu katalizuje ona reakcję przeniesienia grupy aminowej z L-alaniny na 2-oksoglutaran oraz odwrotną do niej, z L-glutami- nianu na pirogronian (Berg i in. 2002).
Aminotransferaza asparaginianowa (AST) normalnie występuje w surowicy krwi oraz w niektórych tkankach, zwłaszcza serca i wątroby. Odpowiada za przeniesienie grupy aminowej z cząsteczki kwasu asparaginowego do kwasu alfa-ketoglutarowe- go, tworząc kwas glutaminowy i kwas szczawianowy. Reakcja jest odwracalna (Berg i in. 2002). Enzym ten zostaje uwolniony do surowicy z powodu uszkodzenia tkanek. Aminotransferaza asparaginowa należy do grupy enzymów o znaczeniu dia-gnostycznym, bowiem jest enzymem wskaźnikowym pochodzenia cytoplazmatycz-no-mitochondrialnego. Wzrost aktywności tego enzymu powoduje obumarcie komó-
Ryc. 4. Zmiany aktywności aminotransferazy alaninowej (ALT), aminotransferazy asparagi-nianowej (AST) oraz dehydrogenazy mleczanowej (LDH) w osoczu koni sportowych w dy-namice treningu (n=9) * – zmiany statystycznie istotne między wartościami w stanie spokoju i po treningu (p<0,05) Fig. 4. Alanine aminotransferase (ALT), aspartate aminotransferase (AST), and lactate dehydro-genase (LDH) activities in the plasma of sport horses at the rest and after training period (n=9) * – statistically significant differences between ALT and AST activities at the rest and after training period (p<0.05)
18
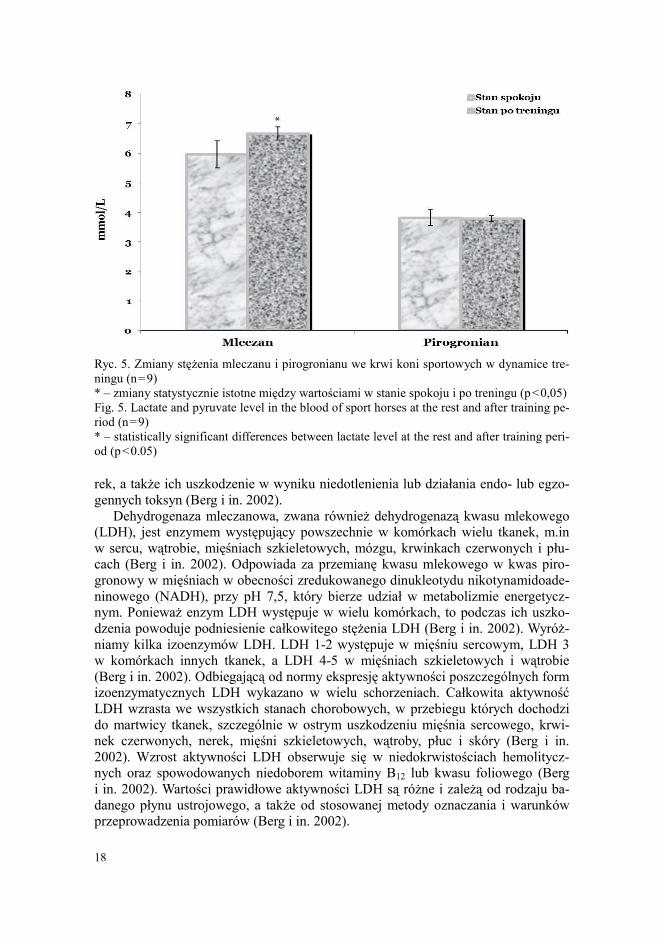
Ryc. 5. Zmiany stężenia mleczanu i pirogronianu we krwi koni sportowych w dynamice tre-ningu (n=9) * – zmiany statystycznie istotne między wartościami w stanie spokoju i po treningu (p<0,05) Fig. 5. Lactate and pyruvate level in the blood of sport horses at the rest and after training pe-riod (n=9) * – statistically significant differences between lactate level at the rest and after training peri-od (p<0.05) rek, a także ich uszkodzenie w wyniku niedotlenienia lub działania endo- lub egzo-gennych toksyn (Berg i in. 2002).
Dehydrogenaza mleczanowa, zwana również dehydrogenazą kwasu mlekowego (LDH), jest enzymem występujący powszechnie w komórkach wielu tkanek, m.in w sercu, wątrobie, mięśniach szkieletowych, mózgu, krwinkach czerwonych i płu-cach (Berg i in. 2002). Odpowiada za przemianę kwasu mlekowego w kwas piro-gronowy w mięśniach w obecności zredukowanego dinukleotydu nikotynamidoade-ninowego (NADH), przy pH 7,5, który bierze udział w metabolizmie energetycz-nym. Ponieważ enzym LDH występuje w wielu komórkach, to podczas ich uszko-dzenia powoduje podniesienie całkowitego stężenia LDH (Berg i in. 2002). Wyróż-niamy kilka izoenzymów LDH. LDH 1-2 występuje w mięśniu sercowym, LDH 3 w komórkach innych tkanek, a LDH 4-5 w mięśniach szkieletowych i wątrobie (Berg i in. 2002). Odbiegającą od normy ekspresję aktywności poszczególnych form izoenzymatycznych LDH wykazano w wielu schorzeniach. Całkowita aktywność LDH wzrasta we wszystkich stanach chorobowych, w przebiegu których dochodzi do martwicy tkanek, szczególnie w ostrym uszkodzeniu mięśnia sercowego, krwi-nek czerwonych, nerek, mięśni szkieletowych, wątroby, płuc i skóry (Berg i in. 2002). Wzrost aktywności LDH obserwuje się w niedokrwistościach hemolitycz-nych oraz spowodowanych niedoborem witaminy B12 lub kwasu foliowego (Berg i in. 2002). Wartości prawidłowe aktywności LDH są różne i zależą od rodzaju ba-danego płynu ustrojowego, a także od stosowanej metody oznaczania i warunków przeprowadzenia pomiarów (Berg i in. 2002).

19
W wyniku naszych badań zanotowano zmniejszenie aktywności aminotransferaz w osoczu koni sportowych po wysiłku fizycznym (ryc. 4). W szczególności aktyw-ność ALT zmniejszyła się o 4% (p<0,05), a AST – o 15% (p<0,05). W medycynie weterynaryjnej aktywność aminotransferaz oraz dehydrogenazy mleczanowej jest często wykorzystywana do oceny stanu funkcjonalnego układu mięśniowo-szkiele- towego oraz odpowiedniej objętości stosowanych treningów fizycznych dla koni sportowych (Harris i in. 1998, Padalino i in. 2007, Kedzierski i in. 2009). Znaczny wzrost aktywności AST w wyniku wyczerpujących ćwiczeń u koni sportowych mo-że wskazywać na uszkodzenie mięśni oraz rozwój zespołu przetrenowania (Padalino i in. 2007). Jednakże w procesie adaptacji do regularnego wysiłku aktywności ami-notransferaz w dynamice treningu mogą podlegać zmianom od niskich do wysokich wartości. W szczególności w badaniach Fazio i in. (2011) stwierdzono znaczny wzrost aktywności AST we krwi koni ras czystej krwi w 60 dniu intensywnego tre-ningu. Jednak pod koniec makrocyklu treningu, w 80 dniu, aktywność AST znacz- nie spadła, co towarzyszyło zwiększeniu sprawności fizycznej i adaptacji koni do regularnych ćwiczeń (Fazio i in. 2011). Wyniki naszych badań są więc zgodne z da-nymi literatury, według których niższa aktywność aminotransferaz po treningu wskazuje na wysoki poziom kondycji fizycznej i sprawności koni (Szarska 1994, Hosoya i in. 2004, Fazio i in. 2011).
Kwas mlekowy (mleczan) jest produktem beztlenowego metabolizmu glukozy. Mleczan powstaje z pirogronianu w reakcji enzymatycznej z LDH (Berg i in. 2002). W warunkach wystarczającego stężenia tlenu pirogronian włącza się w cykl Krebsa i ulega zmianom metabolicznym w mitochondriach do końcowego produktu – wody i dwutlenku węgla. W warunkach intensywnego wysiłku z niedotlenieniem w mięś- niach szkieletowych reakcja enzymatyczna z udziałem LDH przebiega głównie w kierunku powstawania mleczanu z pirogronianu (Berg i in. 2002). Kwas mleko-wy, który gromadzi się w mięśniach szkieletowych podczas intensywnego wysiłku fizycznego, przenika do krwiobiegu i wchłania się przez hepatocyty. W wątrobie mleczan utlenia się do pirogronianu w odwrotnej reakcji enzymatycznej z udziałem LDH (cykl Cori). Część pirogronianu wykorzystuje się do uzupełniania glukozy w glukoneogenezie, a reszta zostaje utleniona w cyklu Krebsa z następną syntezą ATP (Berg i in. 2002). Powstała w glukoneogenezie glukoza dostarczana jest przez krew do mięśni w celu odnowienia zapasów glikogenu (Berg i in. 2002). W związku z tym, poprzez odnowienie pirogronianu do mleczanu z następnym wykorzystaniem jego w glukoneogenezie w wątrobie (cykl Cori), mięśnie szkieletowe nie tylko po-zbywają się „dodatkowego” kwasu mlekowego, który tworzy się w wyjątkowo du-żych ilościach podczas intensywnego wysiłku, ale zapewniają wysoki stosunek NAD+/NADH+ wymagany dla aktywnego funkcjonowania glikolizy (Berg i in. 2002). Z naszych badań wynika, że stężenie mleczanu we krwi koni sportowych zwiększa się po wysiłku o 12% (p<0,05), przy niewielkim wzroście aktywności LDH o 3% (p>0,05) (ryc. 4 i 5). Brak istotnych zmian zaobserwowano w stężeniu pirogronianu we krwi koni sportowych w dynamice treningu. Zanotowany przez nas znaczny wzrost stężenia mleczanu po ćwiczeniach fizycznych odzwierciedla dyna-mikę reakcji tlenowych-beztlenowych i wskazuje na wpływ anaerobowej glikolizy w całkowitej podaży energetycznej dla aktywności mięśniowej.
Podsumowując, można stwierdzić, że systematyczny wysiłek fizyczny koni, któ-

CZ
A (
erytr
ocy
ty):
mle
czan
y=
9,1
8-0
,05
*x;
r=-0
,84
7;
p=
0,0
04;
r2=
0,7
17
, n
=9
CZ
A (
erytr
ocy
ty):
pir
ogro
nia
n y
=5
,58
-0,0
3*x;
r=-0
,77
7;
p=
0,0
14;
r2=
0,6
03
, n
=9
20
A

OM
B370 (
oso
cze)
: T
BA
RS
(er
ytr
ocy
ty)
y=
44
,75
-2,4
9*x;
r=-0
,84
9;
p=
0,0
04;
r2=
0,7
22
, n
=9
OM
B370 (
oso
cze)
: C
ZA
(er
ytr
ocy
ty)
y=
25
5,1
7-1
5,0
4*x;
r=-0
,74
1;
p=
0,0
22
; r2
=0
,54
9, n
=9
Ryc.
6.
Ko
rela
cyjn
e za
leżn
ośc
i m
ięd
zy z
awart
ośc
ią m
etab
oli
tów
gli
ko
lizy i
cał
ko
wit
ą zd
oln
ośc
ią a
nty
oksy
dac
yjn
ą (C
ZA
) er
ytr
ocytó
w w
sp
o-
czyn
ku
prz
ed t
ren
ingie
m (
A)
ora
z m
ięd
zy p
ozi
om
em m
arkeró
w s
tres
u o
ksy
dac
yjn
ego
i c
ałko
wit
ą zd
oln
ośc
ią a
nty
oksy
dac
yjn
ą er
ytr
ocytó
w k
on
i
spo
rto
wych
po
wysi
łku
(B
)
Fig
. 6
. C
orr
elat
ive
lin
ks
bet
wee
n g
lico
lysi
s m
etab
oli
ts l
evel
an
d e
ryth
rocyte
s to
tal
anti
oxid
ant
cap
acit
y o
f sp
ort
ho
rses
at
the
rest
(A
), a
s w
ell
as
bet
wee
n l
evel
of
oxid
ativ
e st
ress
bio
mar
ker
s an
d e
ryth
rocyte
s to
tal
anti
oxid
ant
cap
acit
y o
f sp
ort
ho
rses
aft
er t
rain
ing (
B)
21
B

22
re czynnie biorą udział w WKKW, mającego na celu kształcenie siły i wytrzymało-
ści ich mięśni, powoduje specyficzne zmiany biochemiczne we krwi.
Przeanalizowaliśmy również zależności pomiędzy markerami stresu oksyda-
cyjnego, metabolitami glikolizy i całkowitą zdolnością antyoksydacyjną (CZA)
erytrocytów w dynamice treningu (ryc. 6). Korelacyjna analiza potwierdziła nasze
przypuszczenia. W szczególności w stanie spoczynku markery beztlenowej glikoli-
zy korelują z całkowitą zdolnością antyoksydacyjną erytrocytów (rys. 6A). Ponadto
rosnąca całkowita zdolność antyoksydacyjna po intensywnych ćwiczeniach ograni-
cza pojawienie się stresu oksydacyjnego i tworzenia markerów oksydacyjnej mody-
fikacji białek, powstanie których najwyraźniej jest ograniczone przez dostosowanie
się koni do intensywnego treningu fizycznego. Zanotowana przez nas znaczna re-
dukcja aldehydowych pochodnych zmodyfikowanych białek (OMB370) w osoczu po
wysiłku fizycznym koreluje z całkowitą zdolnością antyoksydacyjną erytrocytów
(ryc. 6B). Jednakże poziom aldehydowych pochodnych zmodyfikowanych białek
osocza po treningu koni sportowych jest odwrotnie skorelowany z zawartością mar-
kerów peroksydacji lipidów w erytrocytach.
PODSUMOWANIE
Znaczny wzrost stężenia markerów peroksydacji lipidów we krwi koni biorących
czynny udział we Wszechstronnym Konkursie Konia Wierzchowego wskazuje na
rozwój stresu oksydacyjnego spowodowanego przez intensywne ćwiczenia fizyczne.
Znaczne zmniejszenie poziomu aldehydowych pochodnych oksydacyjnie zmodyfi-
kowanych białek osocza u koni sportowych po wysiłku fizycznym świadczy o sku-
tecznych adaptacyjnych mechanizmach obrony antyoksydacyjnej organizmu koni do
systematycznego treningu. Aktywacja proteolitycznej degradacji zmodyfikowanych
reszt aminokwasowych może być jedną z przyczyn redukcji oksydacyjnie zmodyfi-
kowanych pochodnych w wyniku adaptacji do wysiłku fizycznego u koni wytreno-
wanych. Znaczny spadek aktywności aminotransferazy asparaginianowej wskazuje
na wysoki poziom kondycji fizycznej koni sportowych oraz ich zdolności adapta-
cyjne do wykonywania intensywnych ćwiczeń fizycznych. Zanotowany przez nas
znaczny wzrost stężenia mleczanu we krwi koni sportowych po wysiłku fizycznym
odzwierciedla wkład beztlenowej glikolizy do całkowitej puli podaży energii dla ak-
tywności mięśniowej. Przeprowadzona analiza korelacyjna pomiędzy stężeniem
markerów stresu oksydacyjnego po treningu wykazała ograniczenie stresu oksyda-
cyjnego przez aktywację obrony antyoksydacyjnej u koni wytrenowanych.
Acknowledgments
This study was carried out during Scholarship Program of Anastasiia Andriichuk
supported by The The Polish National Commission for UNESCO in the Department
of Animal Physiology, Institute of Biology and Environmental Protection, Pomera-
nian University (Słupsk, Poland). We thank to The Polish National Commission for
UNESCO for the support of our study.

23
LITERATURA
Alessio H.M., Goldfarb A.H. 1988. Lipid peroxidation and scavenger enzymes during
exercise: adaptive response to training. J. Appl. Physiol., 64(4): 1333-1334.
Anderson M.G. 1975. The influence of exercises on serum enzyme levels in the horses.
Equine Vet. J., 7: 1605.
Andriichuk A., Tkachova I., Tkachenko H., Kurhaluk N. 2012. Oxidative stress markers in
training dressage horses. Natural Almanac. Life Sciences, a collection of scientific papers,
17(3): 32-43 (in Ukrainian).
Andriichuk A., Tkachova I., Tkachenko H., Kurhaluk N., Matiukha I. 2013. Influence of
training on oxidative stress markers in sport horses of Holstein breed. The Animal Biolo-
gy, 15(1): 9-18 (in Ukrainian).
Art T., Lekeux P. 2005. Exercise-induced physiological adjustments to stressful conditions in
sports horses. Livestock Production Science, 92: 101-111.
Banasiak T., Modzelewska E. 2003. Jazda konna. Pascal sp. z o.o., Bielsko-Biała.
Berg J.M., Tymoczko J.L., Stryer L. 2002. Biochemistry, 5th
edition. W.H. Freeman, New
York.
Bylund J., Brown K.L., Movitz C., Dahlgren C., Karlsson A. 2010. Intracellular generation of
superoxide by the phagocyte NADPH oxidase: how, where, and what for? Free Radic.
Biol. Med., 49: 1834-1845.
Chiaradia E., Avellini L., Rueca F., Spaterna A., Porciello F., Antonioni M.T., Gaiti A. 1998.
Physical exercise, oxidative stress and muscle damage in racehorses. Comparative Bio-
chemistry and Physiology, Part B, Biochemistry and Molecular Biology, 119: 833-836.
Çimen Burak M.Y. 2008. Free radical metabolism in human erythrocytes. Clinica Chimica
Acta, 390: 1-11.
Dani C., Masini E., Bertini G., di Felice A.M., Pezzati M., Ciofini S., Rossetti M., Gallori G.,
Ciuti R., Buonocore G., Paffetti P., Mannaioni P.F., Rubaltelli F.F. 2004. Role of heme
oxygenase and bilirubin in oxidative stress in preterm infants. Pediatr. Res., 56: 873-877.
Davies R., Pethick D.W. 1983. Blood lactate as a measure of work intensity in Standardbred
horses in training. Australian Veterinary Journal, 60: 280-281.
Deaton C.M, Marlin D.J. 2003. Exercise-associated oxidative stress. Clin. Tech. in Equine
Pract., 2: 278-291.
Dubinina E.E., Burmistrov S.O., Khodov D.A., Porotov I.G. 1995. Oxidative modification of
human serum proteins. A method of determining it. Vopr. Med. Khim., 41: 24-26.
Fazio F., Assenza A., Tosto F., Cassela S., Piccione G., Caola G. 2011. Training and haemato-
logical profile in Thoroghbreds and Standardbreds: a longitudinal study. Livestock Jour-
nal, 141: 221-226.
Florczyk U., Józkowicz A.A., Dulak J.J. 2008. Biliverdin reductase: new features of an old
enzyme and its potential therapeutic significance. Pharmacol. Rep., 60: 38-48.
Galaktionova L.P., Molchanova A.V., Elchaninova S.A., Varshavskiy B.Ya. 1998. The lipid
peroxidation processes in at patients with ulcerous illness of stomach and duodenum.
Clin. Lab. Diagnostics 6: 10-14 (in Russian).
Gaweł S., Wardas M., Niedworok E., Wardas P. 2004. Dialdehyd malonowy (MDA) jako
wskaźnik procesów peroksydacji lipidów w organizmie. Wiad. Lek., 57(9-10): 453-455.
Gondim F.J., Zoppi C.C., Silveira L.R., Pereira-da-Silva L., Vaz de Macedo D. 2009. Possi-
ble relationship between performance and oxidative stress in endurance horses. Journal of
Equine Veterinary Science, 29: 206-212.
Halliwell B. 1994. Free radicals, antioxidants and human disease: Curiosity, cause or conse-
quence. Lancet, 344: 721-724.
Harris P.A., Marlin D.J., Gray J. 1998. Plasma aspartate aminotransferase and creatine kinase

24
acnivities in throughbred racehorses in relation to age, sex, exercise and training. The
Veterinary Journal, 155(3): 295-304.
Herasimov I., Plaksina O., 2000. Non-enzymatic determination of lactate and pyruvate con-
centrations in blood simple. Laboratorna Diagnostyka, 2: 46-48 (in Ukrainian).
Hosoya M., Inoue A., Kimura N., Arai T. 2004. Enzyme activities in some types of peripheral
leukocytes of thoroughbred race horses before and after the race. Research in Veterinary
Science, 77: 101-104.
Jagrič-Munih S., Nemec-Svete A., Zrimšek P., Kramarič P., Kos-Kadunc V., Vovk T., Kobal
S. 2012. Plasma malondialdehyde, biochemical and haematological parameters in
standardbred horses during a selected field exercise test. Acta Veterinaria, 62(1): 53-65.
Kamyshnikov V.S. 2004. Reference book on clinic and biochemical researches and laboratory
diagnostics. MEDpress-inform, Moscow (in Russian).
Kedzierski W., Bergero D., Assenza A. 2009. Trends of hematological and biochemical
values in the blood of young race horses during standardized field exercise. Aсta
Veterinaria (Beograd), 59(5-6): 457-446.
Kinnunen S., Atalay M., Hyppä S., Lehmuskero A., Hänninen O., Oksala N. 2005. Effects of
prolonged exercise on oxidative stress and antioxidant defense in endurance horse. Jour-
nal of Sport Science and Medicine, 4: 415-421.
Kirschvink N., de Moffarts B., Lekeux P. 2008. The oxidant/antioxidant equilibrium in hors-
es. The Veterinary Journal, 177: 178-191.
Kulbacka J., Saczko J., Chwiłkowska A. 2009. Stres oksydacyjny w procesach uszkodzenia
komórek. Pol. Merk. Lek., 27(157): 44-47.
Levine R.L., Garland D., Oliver C.N., Amici A., Climent I., Lenz A.G., Ahn B.W., Shaltiel
S., Stadtman E.R. 1990. Determination of carbonyl content in oxidatively modified pro-
teins. Methods in Enzymology, 186: 464-478.
Levine R.L. 2002. Carbonyl modified proteins in cellular regulation, aging, and disease. Free
Radical Biology and Medicine, 32: 790-796.
Marlin D., Nankervis K. 2002. Equine Exercise Physiology, Iowa State Press, Ames, Iowa, USA.
McGowan C.M., Golland L.C., Evans D.L., Hodgson D.R., Rose R.J. 2002. Effects of
prolonged training, overtraining and detraining on skeletal muscle metabolites and
enzymes. Equine Vet. J., 34: 257-263.
Mrowicka M., Bortnik K., Malinowska K., Kędziora J., Mrowicki J. 2009. Całkowity poten-
cjał antyoksydacyjny w osoczu sportowców po dozowanym wysiłku fizycznym. Pol.
Merk. Lek., 27(157): 22-25.
Muñoz A., Riber C., Santisteban R., Lucas R.G., Castejón F.M. 2002. Effect of training duration
and exercise on blood-borne substrates, plasma lactate and enzyme concentrations in
Andalusian, Anglo-Arabian and Arabian horses. Equine Vet. J., 34: 245-251.
Nagababu E., Chrest F.J., Rifkind J.M. 2003. Hydrogen-peroxide-induced heme degradation
in red blood cells: the protective roles of catalase and glutathione peroxidase. Biochim.
Biophys. Acta, 1620: 211-217.
Nagababu E., Mohanty J.G., Bhamidipaty S., Ostera G.R., Rifkind J.M. 2010. Role of the
membrane in the formation of heme degradation products in red blood cells. Life Sci., 86:
133-138.
Nagababu E., Rifkind J.M. 2000. Heme degradation during autoxidation of oxyhemoglobin.
Biochem. Biophys. Res. Comm., 273: 839-845.
Nair V., O’Neil C.L., Wang P.G. 2008. Malondialdehyde. Encyclopedia of Reagents for Or-
ganic Synthesis. John Wiley & Sons, New York.
Nerodenko V.V. 2009. Biological basis of sports training in equestrian sports. Cherkassy,
“Jantar’-Cherkassy” (in Russian).
Padalino B., Rubino G., Centoducati P., Petazzi F. 2007. Training versus Overtraining:
evaluation of protocols. Journal of Equine Veterinary Science, 27(1): 28-31.

25
Pradines B.F., Ramiandrasoa T.F., Hammadi A., Henry M. 2005. Generation of free radicals
and enhancement of hemin-induced membrane damage by a catechol iron chelator in
Plasmodium falciparum. J. Biological Sci., 5: 463-471.
Radák Z., Ogonovsky H., Dubecz J., Pavlik G., Sasvari M., Pucsok J., Berkes I., Csont T.
2003. Super-marathon rice increases serum and urinary nitrotyrosine and carbonyl level.
European Journal of Clinical Investigation, 33: 726-730.
Radák Z., Young Chung H., Goto S. 2008. Systematic adaptation to oxidative challenge in-
duced by regular exercise. Free Radical Biology & Medicine, 44: 153-159.
Rutkowski R., Pancewicz S.A., Rutkowski K., Rutkowska J. 2007. Znaczenie reaktywnych
form tlenu i azotu w patomechanizmie procesu zapalnego. Pol. Merk. Lek., 23(134):
131-136.
Ryter S.W., Alam J., Choi A.M. 2006. Heme oxygenase-1/carbon monoxide: from basic sci-
ence to therapeutic applications. Physiol. Rev., 86: 583-650.
Soares J.C.M., Zanella R., Bondan C., Alves L.P., de Lima M.R., da Motta A.C., Zanella
E.L. 2011. Biochemical and antioxidant changes in plasma, serum, and erythrocytes of
horses before and after a jumping competition. Journal of Equine Veterinary Science, 31:
357-360.
Stadtman E.R., Levine R.L. 2000. Protein oxidation. Ann. N.Y. Acad. Sci., 899: 191-208.
Sugawara Y., Kadono E., Suzuki A., Yukuta Y., Shibasaki Y., Nishimura N., Kameyama Y.,
Hirota M., Ishida C., Higuchi N., Haramoto K., Sakai Y., Soda H. 2003. Hemichrome
formation observed in human haemoglobin A under various buffer conditions. Acta
Physiol. Scand., 179: 49-59.
Szarska E. 1994. Evaluation the fitness of horses during three-day competition. Med Wet.,
50: 274-276.
Sztiller M., Puchała M., Kowalczyk A., Bartosz G. 2006. The influence of ferrylhemoglobin
and methemoglobin on the human erythrocyte membrane. Redox Rep., 11: 263-271.
Telen M.J., Kaufman R.E. 2004. The mature erythrocyte. W: Wintrobe’s clinical hematology,
11th ed., J.P. Greer, J. Forester i in. (red.). Williams and Wilkins, Lippincot.
Tkachenko H., Andriichuk A., Kurhalyuk N., Zalewska K., Tkachova I. 2011. Impact of
training on blood pro- and antioxidant balance of trained horses. Abstracts of Internation-
al Conference “Horse welfare”, December, Wrocław University of Environmental and
Life Sciences, Poland.
Wagner K.R., Sharp F.R., Ardizzone T.D. 2003. Heme and iron metabolism: role in cerebral
hemorrhage. J. Cereb. Blood Flow. Metab., 23: 629-652.
Williams C.A., Burk A.O. 2012. Antioxidant status in elite three-day event horses during
competition. Oxidative Medicine and Cellular Longevity: 1-8.
Yagi K. 1992. Lipid peroxides and exercise. Med. Sci. Sport Sci., 21: 37-40.
Zapora E., Jarocka I. 2013. Hemoglobina – źródłem reaktywnych form tlenu. Postępy Hig.
Med. Dośw., 67: 214-220.
Zar J.H. 1999. Biostatistical Analysis, 4th ed., Prentice Hall Inc., New Jersey.

26


















