
Latent Inhibition Experiments With Goldfish (Carassius ... · ive 7.5-cm goldfish (Carassius...
12
Journal of Comparative Psychology Copyright 1985 by the American Psychological Association, Inc. 1985, Vol. 99, No. 3,316-327 0735-7036/85/$00.75 Latent Inhibition Experiments With Goldfish (Carassius auratus) Akira Shishimi Bekesy Laboratory of Neurobiology University of Hawaii Evidence of latent inhibition was sought in a series of experiments with goldfish. In Experiment 1, goldfish were given nonreinforced preexposure to a color that subsequently predicted shock in an activity conditioning situation; their performance did not differ from that of control animals preexposed to a markedly different color. In Experiment 2, a group of goldfish given nonrein- forced preexposure to a tone and an unstimulated control group were trained in an appetitive situation, with the tone serving either as a conditioned excitor or as a conditioned inhibitor. Preexposure had no significant effect in the conditioned excitation training, but it reduced the level of responding both to the positive stimulus and to the negative compound in the conditioned inhi- bition training. In Experiments 3 and 4, classical aversive conditioning was studied in the shuttle box. In Experiment 3, excitatory conditioning to a color was found to be impaired (relative to the performance of nonpreexposed control animals) as much by nonreinforced preexposure to the training color as by nonreinforced preexposure to a markedly different color; substantial variation in amount of preexposure was without significant effect. In the conditioned inhibition training of Experiment 4, animals with nonreinforced preexposure responded less than did unstimulated control animals both to the positive stimulus and to the negative compound. The results for goldfish can be understood on the assumption that the effect of preexposure in these animals is simply to reduce general responsiveness or level of arousal. Nonreinforced experience with a stimu- ulus becomes inhibitory (Pavlov, 1927); re- lus may interfere with conditioning when sistance to conditioning is, of course, acon- the stimulus subsequently is paired with ventional index of inhibition, although not reinforcement. This phenomenon, known an infallible one (Rescorla, 1971). There is, as "latent inhibition" (Lubow & Moore, however, much contemporary evidence to 1959), has been found in a variety of ani- suggest that inhibition is not produced by mals and conditioning situations (Carlton nonreinforcement per se but by nonrein- & Vogel, 1967; Channell & Hall, 1983; forcement in an otherwise reinforcing con- Crowell & Anderson, 1972; Domjan, 1972; text—that is, by negative correlation with Feldman, 1977; Halgren, 1974; Lantz, 1973; reinforcement (Rescorla & LoLordo, 1965). Lubow, 1965; Lubow, Alek, & Arzy, 1975; An alternative to the Pavlovian explana- Siegel, 1969a). tion of latent inhibition is that the "asso- Latent inhibition may be explained on ciability" of the stimulus—that is, the read- the assumption that the preexposed stim- iness with which it will enter into associa- tive relations with new stimuli—is reduced This article is adapted from a doctoral dissertation by nonreinforced preexposure. Perhaps the submitted to the University of Hawaii. The research simplest and most plausible idea, put for- was supported by Grant MH 33077 from the National wan j in Qne fom Qr another by different Institute of Mental Health. The author is indebted to . ,. , T , ,,, . o n i M. E. Bitterman for advice and to the Center for investigators (Lubow, Werner, & Schnur, Cultural and Technical Interchange between East and 1981; Mackintosh, 1975; Rescorla, 1971), is West for an open grant in support of his graduate that associability is a function of attention stud y- _, and that attention is reduced by nonrein- J^'Z^vS^rt'fSK. forced Preexposur but there are others. Hiroshima Shudo University, Hiroshima 731-31, Ja- Wagner (1976, 1978) Suggested, lor exam- pan: pie, that the preexposed stimulus is less 316
Transcript of Latent Inhibition Experiments With Goldfish (Carassius ... · ive 7.5-cm goldfish (Carassius...

Journal of Comparative Psychology Copyright 1985 by the American Psychological Association, Inc.1985, Vol. 99, No. 3,316-327 0735-7036/85/$00.75
Latent Inhibition Experiments With Goldfish(Carassius auratus)
Akira ShishimiBekesy Laboratory of Neurobiology
University of Hawaii
Evidence of latent inhibition was sought in a series of experiments withgoldfish. In Experiment 1, goldfish were given nonreinforced preexposure to acolor that subsequently predicted shock in an activity conditioning situation;their performance did not differ from that of control animals preexposed to amarkedly different color. In Experiment 2, a group of goldfish given nonrein-forced preexposure to a tone and an unstimulated control group were trainedin an appetitive situation, with the tone serving either as a conditioned excitoror as a conditioned inhibitor. Preexposure had no significant effect in theconditioned excitation training, but it reduced the level of responding both tothe positive stimulus and to the negative compound in the conditioned inhi-bition training. In Experiments 3 and 4, classical aversive conditioning wasstudied in the shuttle box. In Experiment 3, excitatory conditioning to a colorwas found to be impaired (relative to the performance of nonpreexposedcontrol animals) as much by nonreinforced preexposure to the training coloras by nonreinforced preexposure to a markedly different color; substantialvariation in amount of preexposure was without significant effect. In theconditioned inhibition training of Experiment 4, animals with nonreinforcedpreexposure responded less than did unstimulated control animals both to thepositive stimulus and to the negative compound. The results for goldfish canbe understood on the assumption that the effect of preexposure in theseanimals is simply to reduce general responsiveness or level of arousal.
Nonreinforced experience with a stimu- ulus becomes inhibitory (Pavlov, 1927); re-lus may interfere with conditioning when sistance to conditioning is, of course, acon-the stimulus subsequently is paired with ventional index of inhibition, although notreinforcement. This phenomenon, known an infallible one (Rescorla, 1971). There is,as "latent inhibition" (Lubow & Moore, however, much contemporary evidence to1959), has been found in a variety of ani- suggest that inhibition is not produced bymals and conditioning situations (Carlton nonreinforcement per se but by nonrein-& Vogel, 1967; Channell & Hall, 1983; forcement in an otherwise reinforcing con-Crowell & Anderson, 1972; Domjan, 1972; text—that is, by negative correlation withFeldman, 1977; Halgren, 1974; Lantz, 1973; reinforcement (Rescorla & LoLordo, 1965).Lubow, 1965; Lubow, Alek, & Arzy, 1975; An alternative to the Pavlovian explana-Siegel, 1969a). tion of latent inhibition is that the "asso-
Latent inhibition may be explained on ciability" of the stimulus—that is, the read-the assumption that the preexposed stim- iness with which it will enter into associa-
tive relations with new stimuli—is reducedThis article is adapted from a doctoral dissertation by nonreinforced preexposure. Perhaps the
submitted to the University of Hawaii. The research simplest and most plausible idea, put for-was supported by Grant MH 33077 from the National wanj in Qne fom Qr another by differentInstitute of Mental Health. The author is indebted to . ,. ,T , ,,, . o n iM. E. Bitterman for advice and to the Center for investigators (Lubow, Werner, & Schnur,Cultural and Technical Interchange between East and 1981; Mackintosh, 1975; Rescorla, 1971), isWest for an open grant in support of his graduate that associability is a function of attentionstudy- _, and that attention is reduced by nonrein-J^'Z^vS^rt'fSK. forced Preexposur but there are others.Hiroshima Shudo University, Hiroshima 731-31, Ja- Wagner (1976, 1978) Suggested, lor exam-pan: pie, that the preexposed stimulus is less
316

LATENT INHIBITION EXPERIMENTS IN GOLDFISH 317
readily associated with new stimuli becauseof its association with contextual stimuliduring preexposure, an idea supported (al-though not exclusively so) by the contex-tual specificity of the latent inhibition ef-fect (Channell & Hall, 1983; Lubow, Rifkin,& Alek, 1976). The explanation of latentinhibition in terms of reduced associabilityis confirmed by several experiments inwhich nonreinforced preexposure wasfound to retard not only the acquisition ofexcitatory properties when the stimulussubsequently was used to signal reinforce-ment but also when the stimulus was usedto signal nonreinforcement—the acquisi-tion of inhibitory properties (Feldman,1977; Halgren, 1974; Reiss & Wagner, 1972;Rescorla, 1971).
Interest in the phyletic generality ofthese results, all of which were obtained inwork with mammals, is prompted by workwith other animals. In three studies of pi-geons—one on conditioned heart rate(Cohen & MacDonald, 1971) and two onconditioned key pecking (Mackintosh,1973; Tomie, Murphy, Fath, & Jackson,1980)—no evidence of latent inhibition wasfound. The positive results obtained in afourth (autoshaping) study (Tranberg &Rilling, 1978) are inconclusive becausemagazine training was given in advance ofpreexposure; the interference may havebeen due not to unreinforced preexposureper se but to unreinforced preexposure in areinforcing context. There is a report oflatent inhibition in a shuttle box avoidanceexperiment with goldfish (Braud, 1971), butit is not convincing for several reasons. Oneis that the course of acquisition was notshown. A second is that the test perform-ance of the nonpreexposed animals was sogood, given the small amount of trainingand the size of the animals in relation tothe space through which they were requiredto shuttle. A third is that the test perform-ance of animals for which the conditionedstimulus had previously been paired eitherwith food or with long trains of inescapableshock was even better than that of thenonpreexposed animals. It is worth notingthat negative results (i.e., absence of latentinhibition) were obtained in a recent exper-iment on classically conditioned proboscisextension in honeybees (Bitterman, Men-
zel, Fietz, & Schafer, 1983); although fewpreexposures were given, only a few pair-ings were required for conditioning.
If latent inhibition is an attentional phe-nomenon, the question of whether it can befound in goldfish is of particular interest inrelation to previous efforts to find evidenceof learned modification of attention infishes. Goldfish have failed in several ex-periments (Couvillon, Tennant, & Bitter-man, 1976; Tennant & Bitterman, 1973) toshow better intradimensional (ID) than ex-tradimensional (ED) transfer of discrimi-native training, which has been found inrats (Shepp & Eimas, 1964) and monkeys(Shepp & Schrier, 1969) and which hasbeen taken as strong indication of learnedchange in attention (Mackintosh, 1975;Sutherland & Mackintosh, 1971). In viewof the negative outcome of experiments onlatent inhibition in pigeons, it may be wellto note that the ID-ED results for pigeonsare unclear; Mackintosh and Little (1969)found better ID than ED transfer in pi-geons, but Couvillon, Tennant, and Bitter-man (1976) did not. Goldfish and carp (likemany other animals) do show overshadow-ing and blocking (Tennant & Bitterman,1975a), which have been explained in atten-tional terms (Sutherland & Mackintosh,1971), but these phenomena can be ex-plained without reference to attention(Rescorla & Wagner, 1972). Goldfish alsoshow better ED discrimination after truediscrimination (TD) than after pseudodis-crimination (PD) training—that is, train-ing in which the discriminanda are nondif-ferentially reinforced (Tennant & Bitter-man, 1975b). Similar results for pigeonshave been explained on the assumptionthat "general attentiveness" is enhanced byTD training and reduced by PD training(Thomas, Freeman, Svinicki, Burr, &Lyons, 1970), but they can be understood,as well, in terms of blocking by backgroundstimuli. Experiments on latent inhibitionprovide another avenue of approach to theproblem of attentional learning in goldfish.
Experiment 1
Because the meaning of the conventionallatent inhibition experiment is ambiguous,slower excitatory conditioning of the preex-

318 AKIRA SHISHIMI
posed stimulus being interpretable either interms of inhibition or in terms of reducedassociability, it seemed expedient in thissearch for a preexposure effect in goldfishto go at once to a conditioned inhibitionparadigm: After nonreinforced experiencewith a light in the first stage of the experi-ment, the animals were trained in the sec-ond stage to discriminate between a tonethat was paired with shock and a nonrein-forced compound of the tone and the light.If the effect of the preexposure is inhibi-tory, the subsequent discrimination shouldbe facilitated, but if preexposure reducesthe associability of the light, the subse-quent discrimination should be impaired.The control procedure employed in thisexperiment also deserves particular atten-tion in that the control animals were notsimply nonexposed to the light in the firststage but exposed to a light markedly dif-ferent in color from that to be used in thesecond stage. The purpose, as in the origi-nal experiment of Lubow and Moore(1959), was to look not for a general effectof preexposure but for an effect specific toexperience with the preexposed stimulus.The technique employed was activity con-ditioning, with brief shock as the uncondi-tioned stimulus.
Method
Subjects. The subjects were 24 experimentally na-ive 7.5-cm goldfish (Carassius auratus) obtained fromOzark Fisheries in Missouri. They were maintainedindividually on a 24-hr feeding schedule in compart-ments that were formed by partitioning a large tank.The tank was located in a temperature-controlledlaboratory room. The water in the tank was continu-ously filtered and circulated.
Apparatus. The apparatus used in this experimentwas designed to study the classical conditioning ofgeneral activity in fishes (Homer, Longo, & Bitter-man, 1960). Situated in a large sound-attenuatingenclosure, it consisted of six small Plexiglas compart-ments (12.7 x 7.6 X 17.5 cm) through which freshfiltered water flowed continuously. The depth of thewater in the compartments, each of which containedan animal, was 8.5 cm. One long wall of each com-partment was slatted vertically to permit the watercurrent produced by the animal's movement to dis-place a paddle outside. The paddle was fixed to a rod,the other end of which was attached to a strain gaugemounted above the compartment. Resistance changesin the transducer caused by the animal's movementwere transmitted to an integrator described by Wood-ard (1972). The output of the integrator drove a
mercury relay, the contact duration of which wasadjusted to 10 ms. One of the conditioned stimuli(CSs) was general illumination of each compartmentwith a 7.5-W Christmas-tree bulb, red in some casesand blue in others. (All the walls of the compartmentwere of opaque white Plexiglas except for the long wallopposite the slatted wall which was of translucentPlexiglas to permit illumination by the CS light.)Another CS was a 400-Hz tone (35 dB above re 0.1/jN/m2 as calibrated with a hydrophone) deliveredfrom a 7-cm speaker in the ceiling of each compart-ment. The unconditioned stimulus (US) was a single0.45-V/cm, 0.4-s pulse of ac shock delivered throughstainless steel electrodes on the long walls of eachcompartment. All events of the experiment were pro-grammed automatically, and the outputs of the inte-grator relays were recorded by a PDP-8/S minicom-puter.
Procedure. At the outset of the experiment, theanimals were randomly assigned to one of two groupsof 12 subjects each. Group PES was to be preexposedto the same color and Group FED to a different colorfrom that subsequently used in the conditioned inhi-bition training. The two colors, blue and red, werebalanced over conditions. In the first stage of theexperiment, there were 200 preexposures per sessionto one of the colors in five daily sessions. The durationof the CS was 10 s, and the mean intertrial intervalwas 30 s. In the second stage, all animals receivedconditioned inhibition training with tone as S+ andtone plus light as S—. On each S+ trial, a 10-s tone(T) was followed by the US (T+). On each S- trial, a10-s presentation of the same tone in compound withlight ended without shock (TL-). For the PES ani-mals, the color of the light was the same in both stagesof the experiment; for the PED animals, it was differ-ent. There also were blank trials on which no stimuliwere presented. In each of 10 daily sessions of condi-tioned inhibition training, there were five S+, five S—,and five blank trials in quasi-random order, with amean intertrial interval of 60 s. On each trial, generalactivity in the first 10 s was measured. One PESanimal that was preexposed to the red light and onePED animal that was preexposed to the blue wereinjured during the training and were eliminated fromthe analysis.
Results
In Figure 1, the responses of the twogroups to each of the two stimuli and onblank trials are plotted in terms of meanactivity per trial on each day of training inthe second stage of the experiment. Bothgroups developed good discrimination be-tween T+ and TL—, responding more toT+ than on blank trials and less to TL-than on blank trials. Analysis of varianceyields a significant effect of stimuli, F(2,40) = 70.19, p < .0001, and of days, F(9,180) = 5.82, p < .0001, and a significantStimuli X Days interaction, F(IS, 360) =
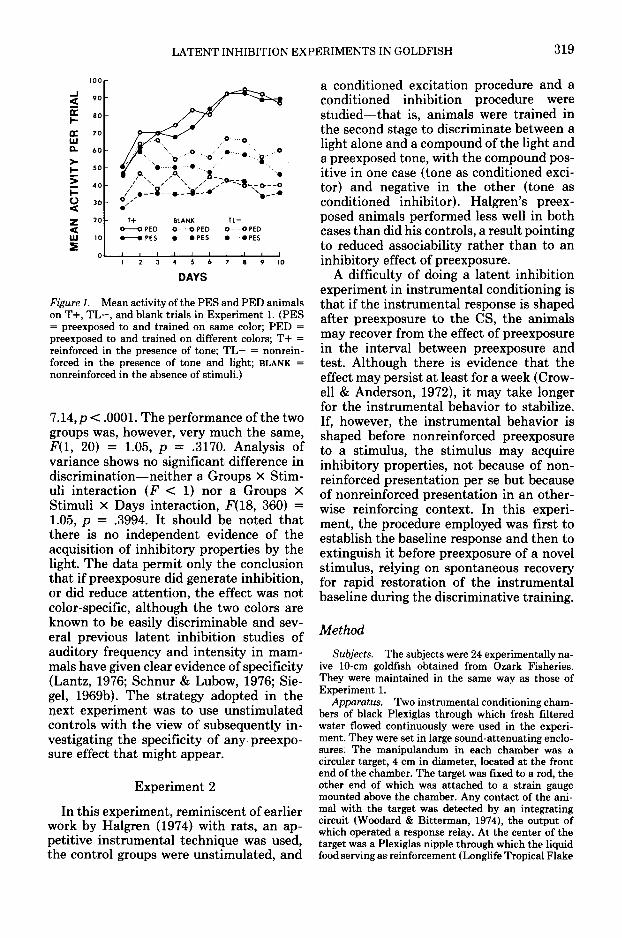
LATENT INHIBITION EXPERIMENTS IN GOLDFISH 319
ccUlQ.
o<
UJ5
100
90
eo
70
60
so
40
30
20
10
0
T+ BtANK Tl-O OPED O -OPED 0--OPED
• »PES • »PES •--•PES
1 2 3 4 5 6 7 8 9 1 0
DAYS
Figure 1. Mean activity of the PES and PED animalson T+, TL-, and blank trials in Experiment 1. (PES= preexposed to and trained on same color; PED =preexposed to and trained on different colors; T+ =reinforced in the presence of tone; TL— = nonrein-forced in the presence of tone and light; BLANK =nonreinforced in the absence of stimuli.)
7.14,p < .0001. The performance of the twogroups was, however, very much the same,F(l, 20) = 1.05, p = .3170. Analysis ofvariance shows no significant difference indiscrimination—neither a Groups X Stim-uli interaction (F < 1) nor a Groups XStimuli X Days interaction, F(18, 360) =1.05, p = .3994. It should be noted thatthere is no independent evidence of theacquisition of inhibitory properties by thelight. The data permit only the conclusionthat if preexposure did generate inhibition,or did reduce attention, the effect was notcolor-specific, although the two colors areknown to be easily discriminable and sev-eral previous latent inhibition studies ofauditory frequency and intensity in mam-mals have given clear evidence of specificity(Lantz, 1976; Schnur & Lubow, 1976; Sie-gel, 1969b). The strategy adopted in thenext experiment was to use unstimulatedcontrols with the view of subsequently in-vestigating the specificity of any preexpo-sure effect that might appear.
Experiment 2
In this experiment, reminiscent of earlierwork by Halgren (1974) with rats, an ap-petitive instrumental technique was used,the control groups were unstimulated, and
a conditioned excitation procedure and aconditioned inhibition procedure werestudied—that is, animals were trained inthe second stage to discriminate between alight alone and a compound of the light anda preexposed tone, with the compound pos-itive in one case (tone as conditioned exci-tor) and negative in the other (tone asconditioned inhibitor). Halgren's preex-posed animals performed less well in bothcases than did his controls, a result pointingto reduced associability rather than to aninhibitory effect of preexposure.
A difficulty of doing a latent inhibitionexperiment in instrumental conditioning isthat if the instrumental response is shapedafter preexposure to the CS, the animalsmay recover from the effect of preexposurein the interval between preexposure andtest. Although there is evidence that theeffect may persist at least for a week (Crow-ell & Anderson, 1972), it may take longerfor the instrumental behavior to stabilize.If, however, the instrumental behavior isshaped before nonreinforced preexposureto a stimulus, the stimulus may acquireinhibitory properties, not because of non-reinforced presentation per se but becauseof nonreinforced presentation in an other-wise reinforcing context. In this experi-ment, the procedure employed was first toestablish the baseline response and then toextinguish it before preexposure of a novelstimulus, relying on spontaneous recoveryfor rapid restoration of the instrumentalbaseline during the discriminative training.
Method
Subjects. The subjects were 24 experimentally na-ive 10-cm goldfish obtained from Ozark Fisheries.They were maintained in the same way as those ofExperiment 1.
Apparatus. Two instrumental conditioning cham-bers of black Plexiglas through which fresh filteredwater flowed continuously were used in the experi-ment. They were set in large sound-attenuating enclo-sures. The manipulandum in each chamber was acirculer target, 4 cm in diameter, located at the frontend of the chamber. The target was fixed to a rod, theother end of which was attached to a strain gaugemounted above the chamber. Any contact of the ani-mal with the target was detected by an integratingcircuit (Woodard & Bitterman, 1974), the output ofwhich operated a response relay. At the center of thetarget was a Plexiglas nipple through which the liquidfood serving as reinforcement (Longlife Tropical Flake
320 AKIRA SHISHIMI
Food blended with tragacanth and water) was deliv-ered with a 10-ml syringe by a Peti-Pump (HarvardApparatus Co.). The target could be illuminated by a7.5-W white Christmas-tree bulb in a light-box behindit, and a 400-Hz tone (28 dB above re 0.1 jiN/m2,hydrophone calibration) could be presented through a18-cm speaker mounted in the ceiling of the sound-attenuating enclosure. All events of the experimentwere programmed automatically, and responses wererecorded with printing counters.
Procedure. In the first stage of the experiment,there were daily 30-min pretraining sessions with thetarget illuminated by the white light. To begin with,each response to the target was rewarded with a dropof food, but in subsequent sessions the reinforcementschedule was shifted to a variable interval averaging15 s (VI15 s), then to VI 30 s, and finally to VI1 min.On the basis of the animals' performance in the last 3days of the VI 1-min training, they were assigned tofour equated groups of 6 subjects each. In the secondstage of the experiment, there were three extinctionsessions in which response to the white light declinedsharply (from an initial rate of about 35 per minute toa rate of about 3 per minute). In the third stage of theexperiment, the animals in two preexposure (PE)groups had six daily sessions in each of which therewere five 3-min preexposures of the tone; the animalsin two nonpreexposure (NPE) groups remained in thechamber without any stimulus presentations. In thefourth stage of the experiment, all animals had 10daily sessions of discrimination training, with the toneserving as a conditioned exciter for one PE and oneNPE group and as a conditioned inhibitor for theother two groups. In the first case, response to thewhite target was reinforced on a VI 1-min schedule inthe presence of the tone (WT+) but not in the absenceof the tone (W—); in the second case, response wasreinforced in the absence (W+) but not in the presenceof the tone (WT—). Each session consisted of ten 3-min white-light segments separated by a brief time-out in darkness, half of the segments with and halfwithout the accompanying tone, in Gellermann orders.The number of responses to the target in each segmentwas recorded with the exception of a 5-s period follow-ing each reinforcement, the purpose of the exclusionbeing to eliminate responses to the food itself. The 3-min segment timer was stopped during these 5-s pe-riods to equate opportunity for response in S+ andS-.
Results
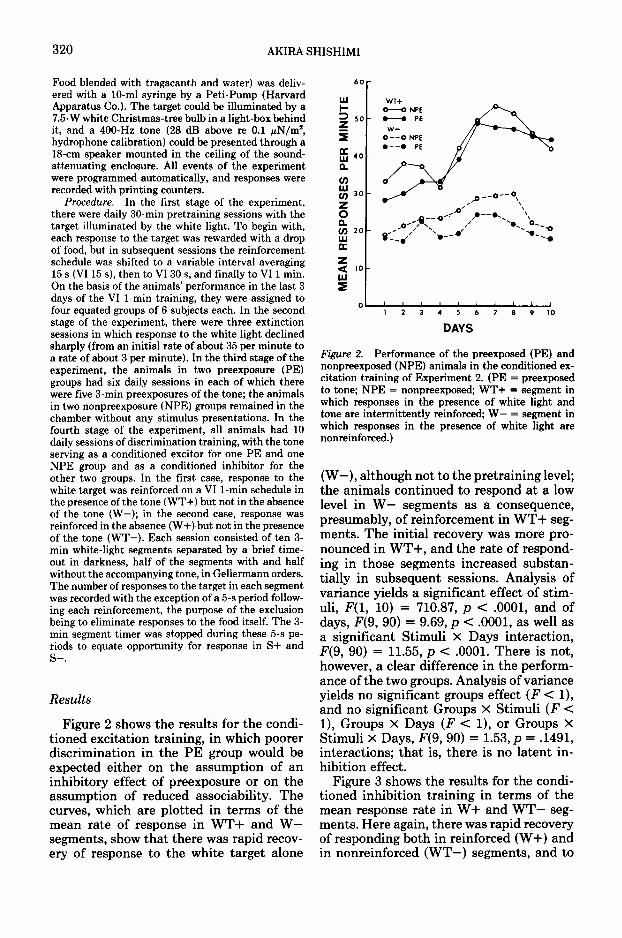
Figure 2 shows the results for the condi-tioned excitation training, in which poorerdiscrimination in the PE group would beexpected either on the assumption of aninhibitory effect of preexposure or on theassumption of reduced associability. Thecurves, which are plotted in terms of themean rate of response in WT+ and W—segments, show that there was rapid recov-ery of response to the white target alone
UJ
£j 400.
W 30
zoa.tn 20
<iii
1 2 3 4 5 6 7 8 9 1 0
DAYS
Figure 2. Performance of the preexposed (PE) andnonpreexposed (NPE) animals in the conditioned ex-citation training of Experiment 2. (PE = preexposedto tone; NPE = nonpreexposed; WT+ = segment inwhich responses in the presence of white light andtone are intermittently reinforced; W— = segment inwhich responses in the presence of white light arenonreinforced.)
(W—), although not to the pretraining level;the animals continued to respond at a lowlevel in W— segments as a consequence,presumably, of reinforcement in WT+ seg-ments. The initial recovery was more pro-nounced in WT+, and the rate of respond-ing in those segments increased substan-tially in subsequent sessions. Analysis ofvariance yields a significant effect of stim-uli, F(l, 10) = 710.87, p < .0001, and ofdays, F(9, 90) = 9.69, p < .0001, as well asa significant Stimuli X Days interaction,F(9, 90) = 11.55, p < .0001. There is not,however, a clear difference in the perform-ance of the two groups. Analysis of varianceyields no significant groups effect (F < 1),and no significant Groups X Stimuli (F <1), Groups X Days (F < 1), or Groups XStimuli x Days, F(9, 90) = 1.53, p = .1491,interactions; that is, there is no latent in-hibition effect.
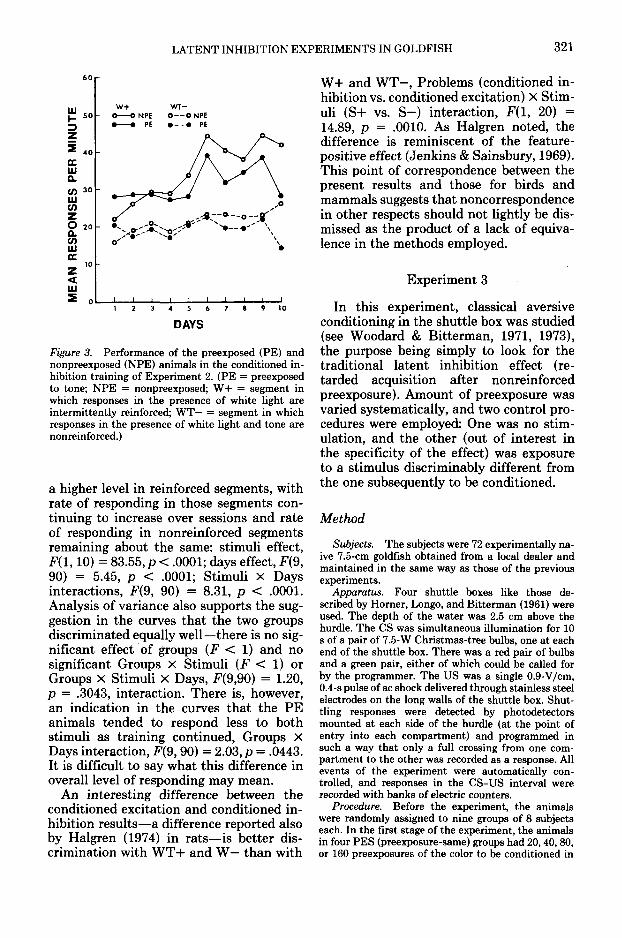
Figure 3 shows the results for the condi-tioned inhibition training in terms of themean response rate in W+ and WT— seg-ments. Here again, there was rapid recoveryof responding both in reinforced (W+) andin nonreinforced (WT—) segments, and to
LATENT INHIBITION EXPERIMENTS IN GOLDFISH 321
UJ
ccUJQ.
COUJCO
oa.COUJcc
UJ
W+ WT->NPE O—ONPE
PE • — • PE
1 2 3 4 5 6 7 8 9 1 0
DAYS
Figure 3. Performance of the preexposed (PE) andnonpreexposed (NPE) animals in the conditioned in-hibition training of Experiment 2. (PE = preexposedto tone; NPE = nonpreexposed; W+ = segment inwhich responses in the presence of white light areintermittently reinforced; WT— = segment in whichresponses in the presence of white light and tone arenonreinforced.)
a higher level in reinforced segments, withrate of responding in those segments con-tinuing to increase over sessions and rateof responding in nonreinforced segmentsremaining about the same: stimuli effect,F(l, 10) = 83.55, p < .0001; days effect, F(9,90) = 5.45, p < .0001; Stimuli X Daysinteractions, F(9, 90) = 8.31, p < .0001.Analysis of variance also supports the sug-gestion in the curves that the two groupsdiscriminated equally well—there is no sig-nificant effect of groups (F < 1) and nosignificant Groups X Stimuli (F < 1) orGroups X Stimuli X Days, F(9,90) = 1.20,p = .3043, interaction. There is, however,an indication in the curves that the PEanimals tended to respond less to bothstimuli as training continued, Groups XDays interaction, F(9,90) = 2.03, p = .0443.It is difficult to say what this difference inoverall level of responding may mean.
An interesting difference between theconditioned excitation and conditioned in-hibition results—a difference reported alsoby Halgren (1974) in rats—is better dis-crimination with WT+ and W— than with
W+ and WT-, Problems (conditioned in-hibition vs. conditioned excitation) X Stim-uli (S+ vs. S-) interaction, F(l, 20) =14.89, p = .0010. As Halgren noted, thedifference is reminiscent of the feature-positive effect (Jenkins & Sainsbury, 1969).This point of correspondence between thepresent results and those for birds andmammals suggests that noncorrespondencein other respects should not lightly be dis-missed as the product of a lack of equiva-lence in the methods employed.
Experiment 3
In this experiment, classical aversiveconditioning in the shuttle box was studied(see Woodard & Bitterman, 1971, 1973),the purpose being simply to look for thetraditional latent inhibition effect (re-tarded acquisition after nonreinforcedpreexposure). Amount of preexposure wasvaried systematically, and two control pro-cedures were employed: One was no stim-ulation, and the other (out of interest inthe specificity of the effect) was exposureto a stimulus discriminably different fromthe one subsequently to be conditioned.
Method
Subjects. The subjects were 72 experimentally na-ive 7.5-cm goldfish obtained from a local dealer andmaintained in the same way as those of the previousexperiments.
Apparatus. Four shuttle boxes like those de-scribed by Homer, Longo, and Bitterman (1961) wereused. The depth of the water was 2.5 cm above thehurdle. The CS was simultaneous illumination for 10s of a pair of 7.5-W Christmas-tree bulbs, one at eachend of the shuttle box. There was a red pair of bulbsand a green pair, either of which could be called forby the programmer. The US was a single 0.9-V/cm,0.4-s pulse of ac shock delivered through stainless steelelectrodes on the long walls of the shuttle box. Shut-tling responses were detected by photodetectorsmounted at each side of the hurdle (at the point ofentry into each compartment) and programmed insuch a way that only a full crossing from one com-partment to the other was recorded as a response. Allevents of the experiment were automatically con-trolled, and responses in the CS-US interval wererecorded with banks of electric counters.
Procedure. Before the experiment, the animalswere randomly assigned to nine groups of 8 subjectseach. In the first stage of the experiment, the animalsin four PES (preexposure-same) groups had 20,40,80,or 160 preexposures of the color to be conditioned in
322 AKIRA SHISHIMI
the second stage. For half of the animals in each group,the color was red, and for the rest it was green. Theanimals in four PED (preexposure-different) groupshad corresponding numbers of preexposures of thecolors, but the color experienced by each animal wasdifferent from that to be conditioned in the secondstage. The animals in the NPE (nonpreexposed) grouphad only blank trials in the first stage; half of themwere conditioned to red in the second satage, and therest to green. In the first stage, there were eight dailysessions, each consisting of 20 trials with a meanintertrial interval of 60 s. For Group NPE, all trialswere blank—that is, the lights were not turned on.For Groups 20 PES and 20 PED, there were blanktrials in the first seven sessions and trials with lightin the eighth; for Groups 40 PES and 40 PED, blanktrials in the first six sessions and trials with light inthe last two; for Groups 80 PES and 80 PED, blanktrials in the first four sessions and trials with light inthe last four; for Groups 160 PES and 160 PED, trialswith light in all eight sessions. The second stage ofthe experiment consisted of 10 daily sessions in eachof which there were 10 (classical) color-shock pairingswith a 10-s CS-US interval and an intertrial intervalaveraging 60 s. On every trial, the number of responsesmade by each animal in the CS-US interval wasrecorded.
Results
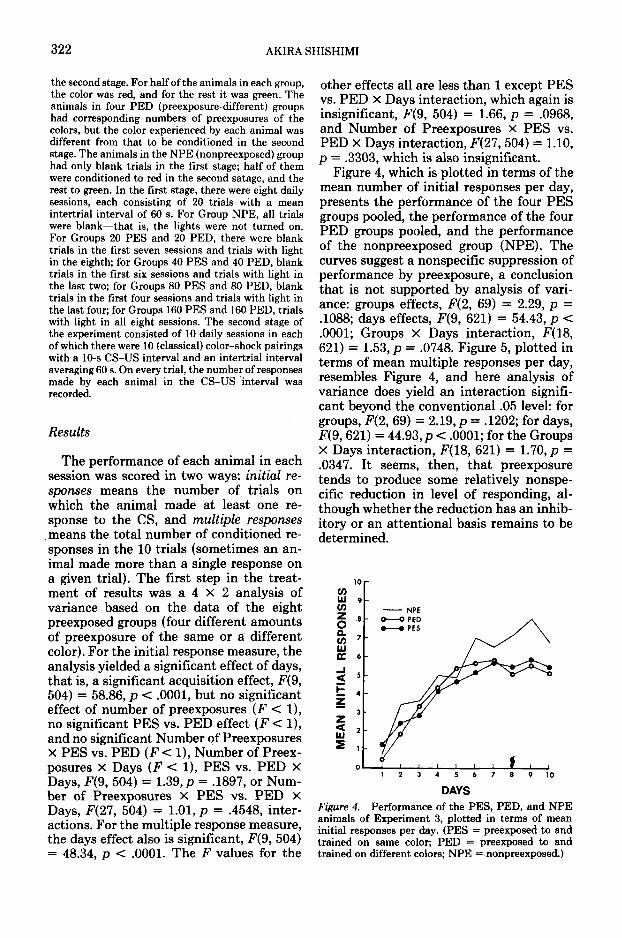
The performance of each animal in eachsession was scored in two ways: initial re-sponses means the number of trials onwhich the animal made at least one re-sponse to the CS, and multiple responsesmeans the total number of conditioned re-sponses in the 10 trials (sometimes an an-imal made more than a single response ona given trial). The first step in the treat-ment of results was a 4 X 2 analysis ofvariance based on the data of the eightpreexposed groups (four different amountsof preexposure of the same or a differentcolor). For the initial response measure, theanalysis yielded a significant effect of days,that is, a significant acquisition effect, F(9,504) = 58.86, p < .0001, but no significanteffect of number of preexposures (F < I),no significant PES vs. PED effect (F < 1),and no significant Number of PreexposuresX PES vs. PED (F< 1), Number of Preex-posures x Days (F < 1), PES vs. PED XDays, F(9, 504) = 1.39, p = .1897, or Num-ber of Preexposures X PES vs. PED XDays, F(21, 504) = 1.01, p = .4548, inter-actions. For the multiple response measure,the days effect also is significant, F(9, 504)= 48.34, p < .0001. The F values for the
other effects all are less than 1 except PESvs. PED X Days interaction, which again isinsignificant, ,F(9, 504) = 1.66, p = .0968,and Number of Preexposures X PES vs.PED X Days interaction, F(27, 504) = 1.10,p = .3303, which is also insignificant.
Figure 4, which is plotted in terms of themean number of initial responses per day,presents the performance of the four PESgroups pooled, the performance of the fourPED groups pooled, and the performanceof the nonpreexposed group (NPE). Thecurves suggest a nonspecific suppression ofperformance by preexposure, a conclusionthat is not supported by analysis of vari-ance: groups effects, F(2, 69) = 2.29, p =.1088; days effects, F(9, 621) = 54.43, p <.0001; Groups X Days interaction, F(18,621) = 1.53, p = .0748. Figure 5, plotted interms of mean multiple responses per day,resembles Figure 4, and here analysis ofvariance does yield an interaction signifi-cant beyond the conventional .05 level: forgroups, F(2, 69) = 2.19, p = .1202; for days,F(9, 621) = 44.93, p < .0001; for the GroupsX Days interaction, F(18, 621) = 1.70, p =.0347. It seems, then, that preexposuretends to produce some relatively nonspe-cific reduction in level of responding, al-though whether the reduction has an inhib-itory or an attentional basis remains to bedetermined.
10
UJ 9
I 'a(A
L. 4
z"" 3
< 2
DAYSFigure 4, Performance of the PES, PED, and NPEanimals of Experiment 3, plotted in terms of meaninitial responses per day. (PES = preexposed to andtrained on same color; PED = preexposed to andtrained on different colors; NPE = nonpreexposed.)

LATENT INHIBITION EXPERIMENTS IN GOLDFISH 323
« '0UJto 9
o 8Q.V)UlDC
35
UJ5
—— NPEO OPED
• PES
1 2 3 4 5 6 7 8 9 1 0
DAYS
Figure 5. Performance of the PES, PED, and NPEanimals of Experiment 3, plotted in terms of meanmultiple responses per day. (PES = preexposed to andtrained on same color; PED = preexposed to andtrained on different colors; NPE = nonpreexposed.)
Experiment 4
The appearance in the classical shuttlebox conditioning situation used in Experi-ment 3 of what might be thought of as anonspecific latent inhibition effectprompted a conditioned inhibition experi-ment in the same situation. Several differ-ent outcomes could be anticipated: One wasthe better discrimination in preexposed an-imals predicted by inhibition theory; a sec-ond was the poorer discrimination in preex-posed animals predicted on the theory ofreduced associability; and a third was anoverall suppression of responding in preex-posed animals such as found in Experiment2. The third expectation would rest, ofcourse, on the assumption that the factorresponsible for the suppression found inExperiment 2 operates in aversive as wellas appetitive training.
MethodSubjects. The subjects were 24 experimentally na-
ive, 7.5-cm goldfish obtained from a local dealer. Theywere maintained in the same way as those of theprevious experiments.
Apparatus. The four shuttle boxes used in Exper-iment 3 were used again in this experiment, but withdifferent CSs. One CS — designated as W — was simul-taneous illumination of a pair of 7.5-W white Christ-mas-tree bulbs, one at each end of the shuttle box.The other — designated as R — was a flickering red light
(0.4 s on, 0.1 s off) from a pair of red bulbs above theshuttle box, one over each compartment. The US wasagain a single 0.9-V/cm, 0.4-s pulse of ac shock deliv-ered through stainless steel electrodes on the longwalls of the shuttle box. The shuttling response, reg-istered by a pair of photodetectors, was defined in theusual way as a complete crossing from one compart-ment to the other. All events of the experiment wereautomatically controlled, and responses were recordedwith banks of electric counters.
Procedure. Before the experiment, the animalswere randomly assigned either to a preexposure (PE)group or to a nonpreexposure (NPE) group, 12 subjectsin each. In the first (preexposure) stage of the exper-iment, the animals in the PE group had eight dailysessions in each of which there were 20 presentationsof the flickering red light. The duration of each stim-ulus was 10 s, and the mean intertrial interval was 60s. For the animals in the NPE group, there were eightsessions in which blank trials (trials without the light)were programmed. In the second (training) stage ofthe experiment, all animals were given 18 days ofconditioned inhibition training with the white lightalone as S+ (W+) and the white light plus red flick-ering light as S— (WR—). In each session, there were10 W+ and 10 WR— trials in Gellermann orders, witha mean intertrial interval of 60 s. On reinforced trials,the 10-s CS was always followed by the US (theconditioning was classical); on nonreinforced trials,the compound ended after 10 s without shock. On eachtrial, all responses in the first 10 s were recorded. OnePE animal was injured during the training and waseliminated from the analysis.
Results
Figure 6 presents the performance of thetwo groups in terms of the mean number of
PREEXPOSURE
O ONPE• • PE
CONDITIONING
DAYS
Figure 6. Performance of the preexposed (PE) andnonpreexposed (NPE) animals in both stages of Ex-periment 4, plotted in terms of mean initial responsesper day. (PE = preexposed to flickering red light; NPE= nonpreexposed; W+ = trial on which white light isreinforced: WR— = trial on which a compound ofwhite and flickering red lights is nonreinforced.)
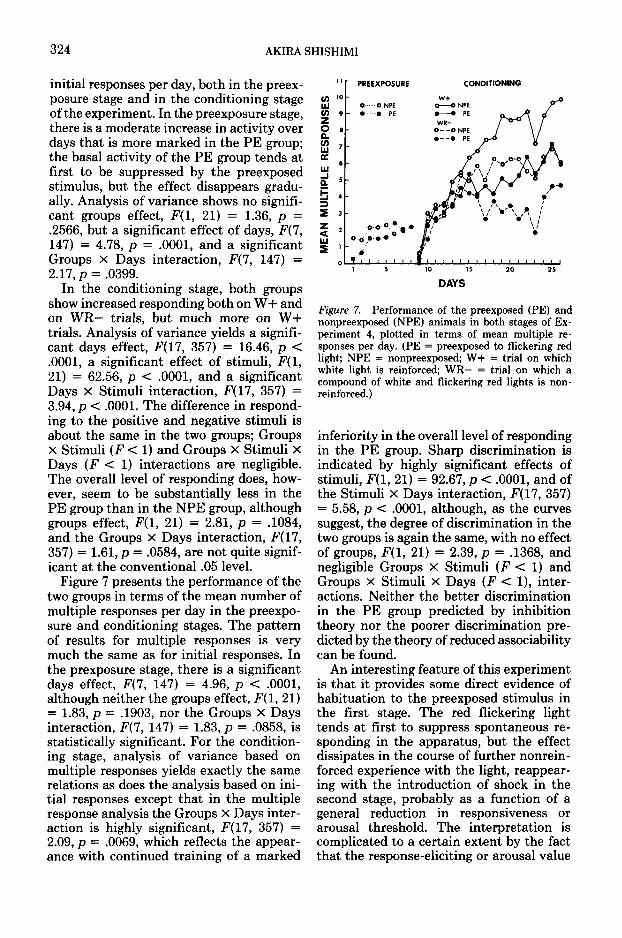
324 AKIRA SHISHIMI
initial responses per day, both in the preex-posure stage and in the conditioning stageof the experiment. In the preexposure stage,there is a moderate increase in activity overdays that is more marked in the PE group;the basal activity of the PE group tends atfirst to be suppressed by the preexposedstimulus, but the effect disappears gradu-ally. Analysis of variance shows no signifi-cant groups effect, F(l, 21) = 1.36, p =.2566, but a significant effect of days, F(7,147) = 4.78, p = .0001, and a significantGroups X Days interaction, F(7, 147) =2.17, p = .0399.
In the conditioning stage, both groupsshow increased responding both on W+ andon WR— trials, but much more on W+trials. Analysis of variance yields a signifi-cant days effect, F(17, 357) = 16.46, p <.0001, a significant effect of stimuli, F(l,21) = 62.56, p < .0001, and a significantDays X Stimuli interaction, F(17, 357) =3.94, p < .0001. The difference in respond-ing to the positive and negative stimuli isabout the same in the two groups; GroupsX Stimuli (F < 1) and Groups X Stimuli XDays (F < 1) interactions are negligible.The overall level of responding does, how-ever, seem to be substantially less in thePE group than in the NPE group, althoughgroups effect, F(l, 21) = 2.81, p = .1084,and the Groups X Days interaction, F(17,357) = 1.61, p = .0584, are not quite signif-icant at the conventional .05 level.
Figure 7 presents the performance of thetwo groups in terms of the mean number ofmultiple responses per day in the preexpo-sure and conditioning stages. The patternof results for multiple responses is verymuch the same as for initial responses. Inthe prexposure stage, there is a significantdays effect, F(7, 147) = 4.96, p < .0001,although neither the groups effect, F(l, 21)= 1.83, p = .1903, nor the Groups X Daysinteraction, F(7, 147) = 1.83, p = .0858, isstatistically significant. For the condition-ing stage, analysis of variance based onmultiple responses yields exactly the samerelations as does the analysis based on ini-tial responses except that in the multipleresponse analysis the Groups X Days inter-action is highly significant, F(17, 357) =2.09, p = .0069, which reflects the appear-ance with continued training of a marked
v>inwoav>UJtc111
PREEXPOSURE
O NPE• PE
CONDITIONING
1 5 1 0 1 5 2 0 2 5
DAYS
Figure 1. Performance of the preexposed (PE) andnonpreexposed (NPE) animals in both stages of Ex-periment 4, plotted in terms of mean multiple re-sponses per day. (PE = preexposed to flickering redlight; NPE = nonpreexposed; W+ = trial on whichwhite light is reinforced; WR— = trial on which acompound of white and flickering red lights is non-reinforced.)
inferiority in the overall level of respondingin the PE group. Sharp discrimination isindicated by highly significant effects ofstimuli, F(l, 21) = 92.67, p < .0001, and ofthe Stimuli X Days interaction, F(17, 357)= 5.58, p < .0001, although, as the curvessuggest, the degree of discrimination in thetwo groups is again the same, with no effectof groups, F(l, 21) = 2.39, p = .1368, andnegligible Groups x Stimuli (F < 1) andGroups X Stimuli X Days (F < I), inter-actions. Neither the better discriminationin the PE group predicted by inhibitiontheory nor the poorer discrimination pre-dicted by the theory of reduced associabilitycan be found.
An interesting feature of this experimentis that it provides some direct evidence ofhabituation to the preexposed stimulus inthe first stage. The red flickering lighttends at first to suppress spontaneous re-sponding in the apparatus, but the effectdissipates in the course of further nonrein-forced experience with the light, reappear-ing with the introduction of shock in thesecond stage, probably as a function of ageneral reduction in responsiveness orarousal threshold. The interpretation iscomplicated to a certain extent by the factthat the response-eliciting or arousal value

LATENT INHIBITION EXPERIMENTS IN GOLDFISH 325
of a stimulus is indexed by reduced re-sponding (relative to interval responding)in the preconditioning phase but by en-hanced responding as conditioning pro-ceeds. Nevertheless, the simplest way toaccount for the results of the second stageis in terms of a nonspecific reduction inresponsiveness or arousal by preexposuresuch as was seen in Experiment 3.
Discussion
The first noteworthy feature of these re-sults is that no preexposure effects specificto the preexposed stimulus were found.There was no significant difference be-tween experimental animals and differentlystimulated controls either in Experiment 3,in which the stimulus was paired withshock and the acquisition of excitatoryproperties was studied, or in Experiment 1,in which the stimulus predicted the non-occurrence of shock and the acquisition ofinhibitory properties was studied. It is con-ceivable that the animals were attending inthe preexposure period to some commonproperty of the stimuli used rather than totheir distinctive properties, but that is un-likely because the stimuli were lights ofdifferent color, and color is a highly salientstimulus dimension for goldfish.
Some evidence of a preexposure influ-ence did, however, appear when unstimu-lated control groups (preexposed only tothe training situation) were used. Althoughthe results for conditioned excitation inExperiment 2, in which the animals wererewarded with food for striking a target,were negative, some indication of a preex-posure effect was obtained in Experiment3, in which a classical aversive conditioningprocedure was employed. There were sig-nificant preexposure effects also in condi-tioned inhibition, both for instrumental re-ward training in Experiment 2 and for clas-sical aversive conditioning in Experiment4, although they were not what would bepredicted on the assumption either thatpreexposure produces inhibition or that itreduces associability. The former assump-tion suggests better discrimination in thepreexposed animals (for which the preex-posed stimulus should already have ac-
quired inhibitory properties), and the lattersuggests poorer performance in the preex-posed animals (which should be slower tolearn about the new significance of thepreexposed stimulus). Instead, the preex-posed animals responded less in each casethan did the control animals, both to thereinforced stimulus and to the nonrein-forced compound consisting of the preex-posed stimulus and the otherwise rein-forced stimulus, with no evidence of a dif-ference in the degree of differentiation orin the rate at which it developed. Nor canthe results be explained in terms of thegeneralization of latent inhibition, whichmight have been expected to impair dis-crimination in the preexposed groups ofExperiments 2 and 4. What these results,which are quite different from those forrats, seem to show is that a general declinein responsiveness or level of arousal is pro-duced by preexposure.
How is the difference between the resultsof the present experiments with goldfishand those of previous experiments withmammals to be understood? In the interestof parsimony, one may be reluctant to thinkin terms of some evolutionary divergencein learning mechanisms, preferring insteadto look for an explanation in terms of dif-ferences in contextual variables or in termsof the nonequivalence of the training situ-ations employed (Bitterman, 1975). Thereare, however, two reasons why the failureof these experiments with goldfish to du-plicate the mammalian results cannot soeasily be dismissed. One has to do with thepower of the goldfish training techniquesand the many strong resemblances betweenthe results obtained with them and thosefor vertebrates of other classes—not onlygeneral resemblances but much more spe-cific ones, such as the feature-positive ef-fect found here and the phenomena of con-ditioned excitation and conditioned inhi-bition themselves. A second concerns theconsistency of the present results with theresults of experiments of quite differentdesign in which evidence of learned modi-fication of attention in goldfish has beensought unsuccessfully. The new results arecompatible with the view that the perform-ance of mammals in latent inhibition andrelated experiments is influenced by mech-

326 AKIRA SHISHIMI
anisms of attentional modification that donot operate in goldfish.
References
Bitterman, M. E. (1975). The comparative analysis oflearning. Science, 188, 699-709.
Bitterman, M. E., Menzel, R., Fietz, A., & Schafer, S.(1983). Classical conditioning of proboscis extensionin honeybees (Apis mellifera). Journal of Compara-tive Psychology, 97, 107-119.
Braud, W. G. (1971). Effectiveness of "neutral," ha-bituated, shock-related, and food-related stimuli asCSs for avoidance learning in goldfish. ConditionalReflex, 6, 153-156.
Carlton, P. L., & Vogel, J. R. (1967). Habituation andconditioning. Journal of Comparative and Physiolog-ical Psychology, 63, 348-351.
Channell, S., & Hall, G. (1983). Contextual effects inlatent inhibition with an appetitive conditioningprocedure. Animal Learning & Behavior, 11, 67-74.
Cohen, D. H., & MacDonald, R. L. (1971). Somevariables affecting orienting and conditioned heart-rate responses in the pigeon. Journal of Comparativeand Physiological Psychology, 74, 123-133.
Couvillon, P. A., Tennant, W. A., & Bitterman, M. E.(1976). Intradimensional vs. extradimensionaltransfer in the discriminative learning of goldfishand pigeons. Animal Learning & Behavior, 4, 197-203.
Crowell, C. R., & Anderson, D. C. (1972). Variationsin intensity, interstimulus interval, and interval be-tween preconditioning CS exposures and condition-ing with rats. Journal of Comparative and Physio-logical Psychology, 79, 291-298.
Domjan, M. (1972). CS preexposure in taste-aversionlearning: Effects of deprivation and preexposureduration. Learning and Motivation, 3, 389-402.
Feldman, M. A. (1977). The effects of preexposure toa warning or a safety signal on the acquisition of atwo-way avoidance response in rats. Animal Learn-ing & Behavior, 5, 21-24.
Halgren, C. R. (1974). Latent inhibition in rats: As-sociative or non-associative? Journal of Compara-tive and Physiological Psychology, 86, 74-78.
Homer, J. L., Longo, N., & Bitterman, M. E. (1960).A classical conditioning technique for small aquaticanimals. American Journal of Psychology, 73, 623-626.
Horner, J. L., Longo, N., & Bitterman, M. E. (1961).A shuttlebox for the fish and a control circuit ofgeneral applicability. American Journal of Psychol-ogy, 74, 114-120.
Jenkins, H. M., & Sainsbury, R. S. (1969). The devel-opment of stimulus control through differential re-inforcement. In N. J. Mackintosh & W. K. Honig(Eds.), Fundamental issues in associative learning(pp. 123-161). Halifax, Nova Scotia: Dalhousie Uni-versity Press.
Lantz, A. E. (1973). Effect of number of trials, inter-stimulus interval, and dishabituation during CS ha-bituation on subsequent conditioning in a CER par-adigm. Animal Learning & Behavior, 1, 273-277.
Lantz, A. E. (1976). The effect of intensity of the
preexposed stimulus on subsequent conditioning.Bulletin of the Psychonomic Society, 7, 381-383.
Lubow, R. E. (1965). Latent inhibition: Effects offrequency of nonreinforced preexposure of the CS.Journal of Comparative and Physiological Psychol-ogy, 60, 454-457.
Lubow, R. E., Alek, M., & Arzy, J. (1975). Behavioraldecrement following stimulus preexposure: Effectsof number of preexposures, presence of a secondstimulus, and interstimulus interval in children andadults. Journal of Experimental Psychology: AnimalBehavior Processes, 104, 178-188.
Lubow, R. E., & Moore, A. U. (1959). Latent inhibi-tion: The effect of nonreinforced pre-exposure tothe conditional stimulus. Journal of Comparativeand Physiological Psychology, 52, 415-419.
Lubow, R. E., Rifkin, B., & Alek, M. (1976). Thecontext effect: The relationship between stimuluspreexposure and environmental preexposure deter-mines subsequent learning. Journal of ExperimentalPsychology: Animal Behavior Processes, 2, 38-47.
Lubow, R. E., Weiner, I., & Schnur, P. (1981). Con-ditioned attention theory. In G. H. Bower (Ed.), Thepsychology of learning and motivation (Vol. 15, pp.1-49). New York: Academic Press.
Mackintosh, N. J. (1973). Stimulus selection: Learn-ing to ignore stimuli that predict no change inreinforcement. In R. A. Hinde & J. S. Hinde (Eds.),Constraints on learning (pp. 75-100). New York:Academic Press.
Mackintosh, N. J. (1975). A theory of attention: Var-iations in the associability of stimuli with reinforce-ment. Psychological Review, 82, 276-298.
Mackintosh, N. J., & Little, L. (1969). Intradimen-sional and extradimensional shift learning in pi-geons. Psychonomic Science, 14, 5-6.
Pavlov, I. P. (1927). Conditioned reflexes (G. V. Anrep,Trans.). London: Oxford University Press.
Reiss, S., & Wagner, A. R. (1972). CS habituationproduces a "latent inhibition effect" but no active"conditioned inhibition." Learning and Motivation,3,237-245.
Rescorla, R. A. (1971). Summation and retardationtests of latent inhibition. Journal of Comparativeand Physiological Psychology, 75, 77-81.
Rescorla, R. A., & LoLordo, V. M. (1965). Inhibitionof avoidance behavior. Journal of Comparative andPhysiological Psychology, 59, 406-412.
Rescorla, R. A., & Wagner, A. R. (1972). A theory ofPavlovian conditioning: Variations in the effective-ness of reinforcement and nonreinforcement. In A.H. Black & W. F. Prokasy (Eds.), Classical condi-tioning II: Current research and theory (pp. 64-99).New York: Appleton-Century-Crofts.
Schnur, P., & Lubow, R. E. (1976). Latent inhibition:The effects of ITI and CS intensity during preex-posure. Learning and Motivation, 7, 540-550.
Shepp, B. E., & Eimas, P. D. (1964). Intradimensionaland extradimensional shifts in the rat. Journal ofComparative and Physiological Psychology, 57, 357-361.
Shepp, B. E., & Schrier, A. M. (1969). ConsecutiveIntradimensional and extradimensional shifts inmonkeys. Journal of Comparative and PhysiologicalPsychology, 67, 199-203.

LATENT INHIBITION EXPERIMENTS IN GOLDFISH 327
Siegel, S. (1969a). Effects of CS habituation on eyelidconditioning. Journal of Comparative and Physiolog-ical Psychology, 69, 245-248.
Siegel, S. (1969b). Generalization of latent inhibition.Journal of Comparative and Physiological Psychol-ogy, 69, 157-159.
Sutherland, N. S., & Mackintosh, N. J. (1971). Mech-anisms of animal discrimination learning. New York:Academic Press.
Tennant, W. A., & Bitterman, M. E. (1973). Somecomparisons of intra- and extradimensional transferin the discriminative learning of goldfish. Journalof Comparative and Physiological Psychology, 83,134-139.
Tennant, W. A., & Bitterma, M. E. (1975a). Blockingand overshadowing in two species of fish. Journalof Experimental Psychology: Animal Behavior Proc-esses, 104, 22-29.
Tennant, W. A., & Bitterman, M. E. (1975b). Extra-dimensional transfer in the discriminative learningof goldfish. Animal Learning & Behavior, 3, 201-204.
Thomas, D. R., Freeman, F., Svinicki, J. G., Burr, D.E. S., & Lyons, J. (1970). Effects of extradimen-sional training on stimulus generalization. Journalof Experimental Psychology Monographs 83(1, Pt.2).
Tomie, A., Murphy, A. L., Path, S., & Jackson, R. L.(1980). Retardation of autoshaping following pre-training with unpredictable food: Effects of chang-ing the context between pretraining and testing.Learning and Motivation, 11, 117-134.
Tranberg, D. K., & Rilling, M. (1978). Latent inhibi-tion in the autoshaping paradigm. Bulletin of thePsychonomic Society, 11, 273-276.
Wagner, A. R. (1976). Priming in STM: An informa-tion processing mechanism for self-generated or re-trieval-generated depression in performance. In T.J. Tighe & R. N. Leaton (Eds.), Habituation: Per-spectives from child development, animal behavior,and neurophysiology (pp. 95-128). Hillsdale, NJ:Erlbaum.
Wagner, A. R. (1978). Expectancies and the primingof STM. In S. H. Hulse, H. Fowler, & W. K. Honig(Eds.), Cognitive processes in animal behavior (pp.177-209). Hillsdale, NJ: Erlbaum.
Woodard, W. T. (1972). A logarithmic electronic in-tegrator for activity transducers. Behavior ResearchMethods & Instrumentation, 4, 149-150.
Woodard, W. T., & Bitterman, M. E. (1971). Classicalconditioning of goldfish in the shuttlebox. BehaviorResearch Methods & Instrumentation, 3, 193-194.
Woodard, W. T., & Bitterman, M. E. (1973). Pavlov-ian analysis of avoidance conditioning in the gold-fish (Carassius auratus). Journal of Comparativeand Physiological Psychology, 82, 123-129.
Woodard, W. T., & Bitterman, M. E. (1974). Improvedtechniques for the measurement of consummatorybehavior in fishes. Behavior Research Methods &Instrumentation, 6, 321-324.
Received January 28, 1984Revision received April 18,1985
Improved Reproduction of Photomicrographsin Behavioral Neuroscience
Behavioral Neuroscience is pleased to announce new and improved photomicro-graph reproduction. Previously, photomicrographs lacked the high resolutionneeded for detailed study. Beginning in 1985, the photomicrograph will appeartwice: once in the text, as usual, and again in a special added signature of betterquality, coated paper stock that will yield substantially more detail.


















