
Lactate Metabolism by Veillonella parvula - Journal of Bacteriology
7
JOURNAL OF BACTERIOLOGY, Mar. 1971, p. 999-1005 Copyright © 1971 American Society for Microbiology Vol. 105, No. 3 Printed in U.S.A. Lactate Metabolism by Veillonella parvula STEPHEN K. C. NG AND IAN R. HAMILTON Department of Oral Biology (Biochemistry), Faculty of Dentistry, University of Manitoba, Winnipeg, Canada Received for publication 21 December 1970 A strain of Veillonella parvula M4, which grows readily in lactate broth without a requirement for carbon dioxide, has been isolated from the oral cavity. Anaero- bic, washed cells of this organism fermented sodium lactate to the following prod- ucts (moles/ 100 moles of lactate): propionate, 66; acetate, 40; carbon dioxide, 40; and hydrogen, 14. Cells grew readily in tryptone-yeast extract broth with pyruvate, oxaloacetate, malate, and fumarate, but poorly with succinate. The fermentation of pyruvate, oxaloacetate, or lactate plus oxaloacetate by washed cells resulted in the formation of propionate and acetate in ratios significantly lower than those ob- served with lactate as the sole carbon source. This was primarily due to increased acetate production. Cell-free extracts were unable to degrade lactate but metabo- lized lactate in the presence of oxaloacetate, indicating the presence of malic-lactic transhydrogenase in this organism. Lactic dehydrogenase activity was not observed. Evidence is presented for oxaloacetate decarboxylase and malic dehydrogenase ac- tivities in extracts. Members of the genus Veillonella are unable to utilize carbohydrates but readily metabolize pur- ines and short-chain acids, such as lactate and pyruvate, for energy (6, 11). An extensive review of the characteristics and properties of the Veillo- nella (16, 18, 19) prompted Rogosa (17) to revise the classification of these obligate anaerobes into two main species: V. alcalescens and V. parvula. Foubert and Douglas (6) first reported in detail on the metabolism of members of the Veillonella by showing that Micrococcus lactilyticus, now classified as V. alcalescens (17), fermented lactic acid with the production of hydrogen, carbon dioxide, acetate, and propionate. In the same year, Johns (10) isolated a "new micrococcus" from the rumen of sheep which was unable to ferment carbohydrates but was capable of de- grading pyruvate, lactate, oxaloacetate, malate, and fumarate to the same end products as those of M. lactilyticus. Succinate fermentation, on the other hand, resulted in the formation of only propionate and carbon dioxide. Johns (10, 11) identified the "new micrococcus" as V. gazogenes (V. alcalescens) and showed that the utilization of lactate by this strain had a mandatory require- ment for carbon dioxide. Rogosa (16) confirmed this by demonstrating that strains of both type- species fermented lactate poorly, or not at all, in the absence of carbon dioxide. Furthermore, even in the presence of carbon dioxide, lactate metabo- lism by these strains was limited. The long and confused history of Veillonella taxonomy has made it difficult to make compari- sons between the metabolism of V. parvula and that of V. alcalescens. As pointed out by Rogosa (16), most investigators have generally designated any veillonella isolate as V. alcalescens; conse- quently, the literature contains few reports of studies with V. parvula. With this in mind, a study was undertaken to examine the metabolic properties of V. parvula species isolated from the human oral cavity. During the course of this study, a strain of V. parvula (M4) was obtained which differed from the strains of Veillonella reported previously, in that it grew and metabo- lized sodium lactate readily in the absence of carbon dioxide. This report describes the ability of this organism to degrade lactate and other short-chain acids with the formation of propio- nate, acetate, carbon dioxide, and hydrogen. Data will be presented elsewhere demonstrating the involvement of carbon dioxide fixation in the formation of propionate by this strain (I. R. Hamilton and K. C. S. Ng, manuscript in prepa- ration). MATERIALS AND METHODS Strains of V. parvula were isolated anaerobically and with the aid of antibiotics (15, 20) from human saliva by employing Rogosa's 1% lactate medium (16) supple- mented with phosphate buffer (0.1 M, pH 6.5). The via- bility and purity of the cultures was checked at monthly intervals by the staining procedures and routine taxon- omic tests described by Rogosa (16) and Rogosa and Bishop (18, 19). The Veillonella species isolated were classified by the scheme proposed by Rogosa (17). The strain selected for study, V. parvula M4, had the following characteristics: gram-negative; obligate an- aerobe; small coccoid (0.4 Am in diameter); unable to 999 Downloaded from https://journals.asm.org/journal/jb on 21 December 2021 by 183.80.30.18.
Transcript of Lactate Metabolism by Veillonella parvula - Journal of Bacteriology

JOURNAL OF BACTERIOLOGY, Mar. 1971, p. 999-1005Copyright © 1971 American Society for Microbiology
Vol. 105, No. 3Printed in U.S.A.
Lactate Metabolism by Veillonella parvulaSTEPHEN K. C. NG AND IAN R. HAMILTON
Department of Oral Biology (Biochemistry), Faculty of Dentistry, University of Manitoba, Winnipeg, Canada
Received for publication 21 December 1970
A strain of Veillonella parvula M4, which grows readily in lactate broth withouta requirement for carbon dioxide, has been isolated from the oral cavity. Anaero-bic, washed cells of this organism fermented sodium lactate to the following prod-ucts (moles/ 100 moles of lactate): propionate, 66; acetate, 40; carbon dioxide, 40;and hydrogen, 14. Cells grew readily in tryptone-yeast extract broth with pyruvate,oxaloacetate, malate, and fumarate, but poorly with succinate. The fermentation ofpyruvate, oxaloacetate, or lactate plus oxaloacetate by washed cells resulted in theformation of propionate and acetate in ratios significantly lower than those ob-served with lactate as the sole carbon source. This was primarily due to increasedacetate production. Cell-free extracts were unable to degrade lactate but metabo-lized lactate in the presence of oxaloacetate, indicating the presence of malic-lactictranshydrogenase in this organism. Lactic dehydrogenase activity was not observed.Evidence is presented for oxaloacetate decarboxylase and malic dehydrogenase ac-tivities in extracts.
Members of the genus Veillonella are unable toutilize carbohydrates but readily metabolize pur-ines and short-chain acids, such as lactate andpyruvate, for energy (6, 11). An extensive reviewof the characteristics and properties of the Veillo-nella (16, 18, 19) prompted Rogosa (17) to revisethe classification of these obligate anaerobes intotwo main species: V. alcalescens and V. parvula.
Foubert and Douglas (6) first reported in detailon the metabolism of members of the Veillonellaby showing that Micrococcus lactilyticus, nowclassified as V. alcalescens (17), fermented lacticacid with the production of hydrogen, carbondioxide, acetate, and propionate. In the sameyear, Johns (10) isolated a "new micrococcus"from the rumen of sheep which was unable toferment carbohydrates but was capable of de-grading pyruvate, lactate, oxaloacetate, malate,and fumarate to the same end products as thoseof M. lactilyticus. Succinate fermentation, on theother hand, resulted in the formation of onlypropionate and carbon dioxide. Johns (10, 11)identified the "new micrococcus" as V. gazogenes(V. alcalescens) and showed that the utilization oflactate by this strain had a mandatory require-ment for carbon dioxide. Rogosa (16) confirmedthis by demonstrating that strains of both type-species fermented lactate poorly, or not at all, inthe absence of carbon dioxide. Furthermore, evenin the presence of carbon dioxide, lactate metabo-lism by these strains was limited.The long and confused history of Veillonella
taxonomy has made it difficult to make compari-sons between the metabolism of V. parvula and
that of V. alcalescens. As pointed out by Rogosa(16), most investigators have generally designatedany veillonella isolate as V. alcalescens; conse-quently, the literature contains few reports ofstudies with V. parvula. With this in mind, astudy was undertaken to examine the metabolicproperties of V. parvula species isolated from thehuman oral cavity. During the course of thisstudy, a strain of V. parvula (M4) was obtainedwhich differed from the strains of Veillonellareported previously, in that it grew and metabo-lized sodium lactate readily in the absence ofcarbon dioxide. This report describes the abilityof this organism to degrade lactate and othershort-chain acids with the formation of propio-nate, acetate, carbon dioxide, and hydrogen. Datawill be presented elsewhere demonstrating theinvolvement of carbon dioxide fixation in theformation of propionate by this strain (I. R.Hamilton and K. C. S. Ng, manuscript in prepa-ration).
MATERIALS AND METHODSStrains of V. parvula were isolated anaerobically and
with the aid of antibiotics (15, 20) from human salivaby employing Rogosa's 1% lactate medium (16) supple-mented with phosphate buffer (0.1 M, pH 6.5). The via-bility and purity of the cultures was checked at monthlyintervals by the staining procedures and routine taxon-omic tests described by Rogosa (16) and Rogosa andBishop (18, 19). The Veillonella species isolated wereclassified by the scheme proposed by Rogosa (17).The strain selected for study, V. parvula M4, had the
following characteristics: gram-negative; obligate an-aerobe; small coccoid (0.4 Am in diameter); unable to
999
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Dec
embe
r 20
21 b
y 18
3.80
.30.
18.

NG AND HAMILTON
ferment carbohydrates; grew in tryptone-yeast extractbroth with lactate (growth-rate constant = 0.49), pyru-vate (0.49), oxaloacetate (0.25), malate (0.26), fumarate(0.25), and succinate (0.19); produced H2S; producednitrite from nitrate; was indole-, gelatin-, starch- andcatalase-negative; and was beta-hemolytic on bloodagar. Putrescine and cadaverine were not required forgrowth in semidefined basal medium containing 1% lac-tate. The basal medium employed was that proposed byRogosa and Bishop (18) without hypoxanthine, uracil,and putrescine.
Preparation of resting cells and cell-free extracts.Large quantities of exponential-phase cells were rou-tinely obtained from heavily inoculated anaerobic lac-tate broth cultures of V. parvula M4 grown in 1- to 2-liter flasks. Growth was followed turbidimetrically in aKlett-Summerson colorimeter containing a red filter(640 to 700 nm). Exponential-phase cells were collectedby centrifugation at 13,000 x g for 10 to 15 min in aLRA Lourdes refrigerated centrifuge at 4 C. The cellswere washed once and ultimately suspended in degassedphosphate buffer (0.1 M, pH 6.5) containing 20 mMMgSO4 and 20 mM 2-mercaptoethanol; these conditionswere optimal for lactate fermentation with resting cells.Reaction mixtures routinely contained 2 mg (dryweight) of cells per ml.
Cell-free extracts were obtained from exponential-phase cells [30 mg (dry weight)/ml] either by sonic dis-ruption for 15 min in a stream of nitrogen in a Ray-theon sonic oscillator at 4 C or by passing washed cellsthrough a French pressure cell (American InstrumentsCo.) three to four times at a pressure of 6,000 to 9,000psi. The broken-cell preparations were centrifuged at13,000 x g for 15 min at 4 C, and the dark, reddishbrown supernatant was stored at 0 C under nitrogenuntil used.
All manipulations involving cells or extract prepara-tions were carried out in a stream of nitrogen to main-tain anaerobic conditions.
Experimental procedure. Experiments were normallycarried out at 37 C in 25- to 50-ml Erlenmeyer flasksfitted with tight fitting serum stoppers except when gasproduction was determined. The flasks containingwashed cells or extract preparations in buffer wereevacuated and gassed with nitrogen via a syringe needlethree to four times, and the substrate was added by sy-ringe to start the reaction. To stop the reaction, sam-ples were withdrawn at various times by syringe andadded either to 2 ml of 0.05 N NaOH or to 0.1 ml of25% ZnSO4 in plastic centrifuge tubes. After neutraliza-tion, the supernatant fluid obtained by centrifugationwas recovered and frozen until analyzed. When ketoacids were to be measured, samples were added to anequal volume of 0.1% 2,4-dinitrophenylhydrazine in 2N HCI, and the mixture was incubated at 4 C for 60min. Some of the hydrazones formed precipitatedduring this procedure and were separated from cells orprotein by extraction with ethyl acetate.One unit of metabolic activity is defined as the deg-
radation of I Amole of substrate per mg (dry weight) ofcell material (or mg of protein) per hour.
Analyses. Lactic acid was assayed by the modifiedlactic dehydrogenase method of Cohen and Noell (3).Pyruvic acid was assayed either colorimetrically with
2,4-dinitrophenylhydrazine (9) or enzymatically withlactic dehydrogenase (8). Oxaloacetate was measuredeither colorimetrically with 2,4-dinitrophenylhydrazine,enzymatically with malic dehydrogenase, or by the di-rect spectrophotometric method of Allen (1). The enzy-matic assay contained 0.2 ml of sample (0 to 10 ,ug ofoxaloacetate) and 0.8 ml of buffer-reduced nicotinamideadenine dinucleotide (NADH)-enzyme solution con-sisting of 90 nmoles of NADH and 0.05 gg of malicdehydrogenase (Boehringer Mannheim Corp.) in eitherphosphate or tris (hydroxymethyl)aminomethane buffer(0.05 M, pH 7.5). The remaining NADH was measuredspectrophotometrically at 340 nm after incubation for20 min at room temperature. Freshly prepared sodiumoxaloacetate solutions were used as standards. Thepresence of malic dehydrogenase activity in V. parvulawas determined in the same assay system by substi-tuting cell-free extract, prepared by sonic treatment, forthe commercial malic dehydrogenase. Protein was de-termined by the ultraviolet-adsorption method of Layne(13).
Gaseous products. The extreme oxygen sensitivity ofV. parvula M4 militated against the use of conventionalmanometric methods for the measurement of carbondioxide and hydrogen evolution. For this purpose, amodification of the triple side-arm Warburg method ofHamilton et al. (9) was employed, which gave con-sistent results for measurement of carbon dioxide andhydrogen in the same flask. The modification consistedof adding the cells or extract preparation to the flaskafter the nitrogen-gassing procedure by syringe via thenormal side-arm which had been stoppered with a tightfitting serum cap.
Nongaseous products. Nongaseous end productsformed during metabolism were separated by silicicacid column chromatography employing a 0 to 15%butanol-chloroform gradient (2). All solvents were satu-rated with 0.5 N H2S04. Acidified 15% butanol in chlo-roform was added by peristaltic pump at a rate of 0.5ml/min to 100 ml of acidified chloroform in a 250-mlmixing chamber. The contents of this chamber werethen added to a column (1.3 by 20 cm) of 100-meshsilicic acid which had been equilibrated with 0.5 NH2S04. The preparation of the column and sample hasbeen previously described (9). Fractions (3 ml) werecollected, and the acidity was determined in duplicatel-ml samples by titrating to the phenolphthalein endpoint with standardized NaOH. The radioactivity con-tent of the remaining portion was determined by liquidscintillation counting.
Preparation of lactate-U-'4C. Uniformly labeled "C-lactate was prepared enzymatically from sodium pyru-vate-U-14C by the method of Krebs et al. (12) or iso-lated from the medium after the anaerobic metabolismof glucose-U-'4C by the homofermentative organismStreptococcus salivarius under conditions previouslydescribed (8).
RESULTSGrowth. The growth-rate constant for V. par-
vula M4 in lactate broth was 0.49, calculatedfrom an average generation time of 88 min, andwas the same whether nitrogen or nitrogen plus
1000 J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Dec
embe
r 20
21 b
y 18
3.80
.30.
18.
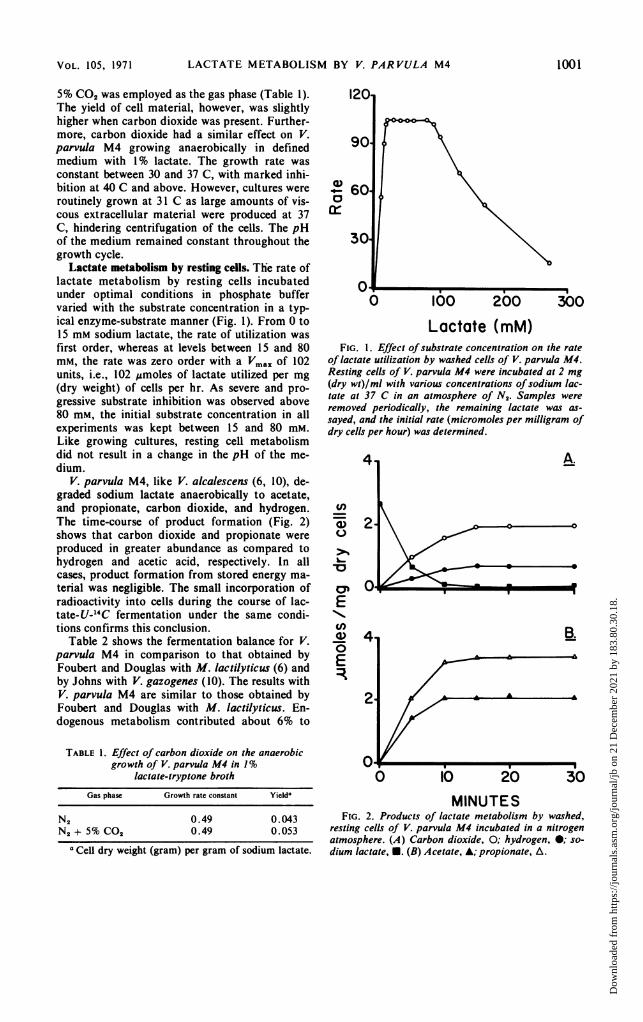
LACTATE METABOLISM BY V. PAR VULA M4
5% CO2 was employed as the gas phase (Table 1).The yield of cell material, however, was slightlyhigher when carbon dioxide was present. Further-more, carbon dioxide had a similar effect on V.parvula M4 growing anaerobically in definedmedium with 1% lactate. The growth rate wasconstant between 30 and 37 C, with marked inhi-bition at 40 C and above. However, cultures wereroutinely grown at 31 C as large amounts of vis-cous extracellular material were produced at 37C, hindering centrifugation of the cells. The pHof the medium remained constant throughout thegrowth cycle.
Lactate metabolism by resting cells. The rate oflactate metabolism by resting cells incubatedunder optimal conditions in phosphate buffervaried with the substrate concentration in a typ-ical enzyme-substrate manner (Fig. 1). From 0 to15 mm sodium lactate, the rate of utilization wasfirst order, whereas at levels between 15 and 80mm, the rate was zero order with a Vmax of 102units, i.e., 102 ,umoles of lactate utilized per mg(dry weight) of cells per hr. As severe and pro-gressive substrate inhibition was observed above80 mM, the initial substrate concentration in allexperiments was kept between 15 and 80 mm.Like growing cultures, resting cell metabolismdid not result in a change in the pH of the me-dium.
V. parvula M4, like V. alcalescens (6, 10), de-graded sodium lactate anaerobically to acetate,and propionate, carbon dioxide, and hydrogen.The time-course of product formation (Fig. 2)shows that carbon dioxide and propionate wereproduced in greater abundance as compared tohydrogen and acetic acid, respectively. In allcases, product formation from stored energy ma-terial was negligible. The small incorporation ofradioactivity into cells during the course of lac-tate-U-_4C fermentation under the same condi-tions confirms this conclusion.
Table 2 shows the fermentation balance for V.parvula M4 in comparison to that obtained byFoubert and Douglas with M. lactilyticus (6) andby Johns with V. gazogenes (10). The results withV. parvula M4 are similar to those obtained byFoubert and Douglas with M. lactilyticus. En-dogenous metabolism contributed about 6% to
TABLE 1. Effect of carbon dioxide on the anaerobicgrowth of V. parvula M4 in 1%
lactate-tryptone broth
Gas phase Growth rate constant YieWda
N2 0.49 0.043N2 + 5% CO2 0.49 0.053
a Cell dry weight (gram) per gram of sodium lactate.
-60
30
0 100 200 300Lactate (mM)
FIG. 1. Effect of substrate concentration on the rateof lactate utilization by washed cells of V. parvula M4.Resting cells of V. parvula M4 were incubated at 2 mg(dry wt)/ml with various concentrations ofsodium lac-tate at 37 C in an atmosphere of N2. Samples wereremoved periodically, the remaining lactate was as-sayed, and the initial rate (micromoles per milligram ofdry cells per hour) was determined.
4 A.
2 \
0 0E
R
30MINUTE S
FIG. 2. Products of lactate metabolism by washed,resting cells of V. parvula M4 incubated in a nitrogenatmosphere. (A) Carbon dioxide, 0; hydrogen, 0; so-dium lactate, U. (B) Acetate, A; propionate, A.
1001VOL. 105, 1971
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Dec
embe
r 20
21 b
y 18
3.80
.30.
18.

NG AND HAMILTON
the end products as observed from the carbonrecovery, i.e., 105.5%.
Related metabolism. Johns (11) proposed apathway for the degradation of lactate by V. ga-zogenes which was similar to that proposed byDelwiche (4) for Propionibacterium pentosa-ceum. In this pathway, lactate was thought to bedegraded initially to pyruvate which is then eitherdecarboxylated to the end products acetate,carbon dioxide, and hydrogen or converted tooxaloacetate by a carbon dioxide fixation reac-tion. Oxaloacetate is then further metabolized tomalate, fumarate, and succinate. The final step isthe decarboxylation of succinate to form pro-pionate and carbon dioxide.To test whether this pathway might be oper-
able in V. parvula M4, the ability of washed cellsto degrade these compounds was determined.Anaerobic cells readily fermented pyruvate andoxaloacetate (Fig. 3A), although the initial ratesof degradation (70 and 48 units, respectively)were considerably lower than that for lactate (102units). Although the degradation of lactate bymalic-lactic transhydrogenase in species of Veil-lonella requires oxaloacetate (E. F. Phares andM. V. Long, 1956, Amer. Chem. Soc. MeetingAbstr., p.626), the incubation of cells with bothoxaloacetate and lactate resulted in a markeddecrease in the rate of utilization of both sub-strates (Fig. 3B). The initial rates of 66 and 26units for lactate and oxaloacetate, respectively,were almost half of the rates obtained when thecells were incubated with each substrate alone,indicating that they possibly compete for en-zymes in the same pathway.
Further insight into the reactions involved inlactate metabolism in V. parvula was obtained byquantitative determination of the acidic end prod-ucts formed during the metabolism of pyruvate,oxaloacetate, and lactate- U-14C plus oxaloace-tate. These substrates, like lactate, were degradedto propionate and acetate (Table 3), as well as to
TABLE 2. Fermentation balance obtained for V.parvula M4 compared to those obtained by Foubert
and Douglas (6) andJohns (10)for V. alcalescens
'Sodium lactate fermented(moles/100 moles)
Product V. alcaiescens
V. parvula M4
Douglas (6) Johns (10)
Propionate ....... 66.0 63.7 58.5Acetate .......... 39.6 39.5 48.7Carbon dioxide .... 38.3 39.1 35.2Hydrogen ........ 13.6 14.2 14.8Carbon recovery ... 105.5 103.6 102.0O/R ratio ........ 0.96 0.9 1.04
A.
0JolE
6Ini B.
0E
3
00 10 20 30
MINUTESFIG. 3. Metabolism of "4C-lactate, oxaloacetate,
pyruvate, and "4C-lactate plus oxaloacetate by restingcells of V. parvula M4 in an atmosphere of nitrogen +5% carbon dioxide. (A) Individual flasks contained 10mm of the following: sodium lactate-U-"4C (1,491dpm/1umole), 0; potassium pyruvate, *; sodium oxal-oacetate, A. (B) Combined incubation of 10 mM so-dium lactate-U-"4C plus 10 mm sodium oxaloacetate:sodium lactate-U-'4C, 0; sodium oxaloacetate, A.
carbon dioxide and hydrogen. However, the pro-pionate to acetate ratios for these substrates wereappreciably lower than that obtained with lactatealone. The lowest ratios were obtained when py-ruvate or oxaloacetate were the substrates (0.46and 0.44, respectively) indicating that the metab-olism of these substrates was primarily in thedirection of acetate formation. The incubation oflactate-_U-4C with oxaloacetate (Table 3) resultedin decreased acetate formation as compared toacetate production when oxaloacetate was thesole carbon source.A comparison of the "4C-specific activities of
the acetate and propionate formed in experimentsI and 4 indicates that the addition of 10 mm unla-beled sodium oxaloacetate to 10 mm sodium lac-tate-U-"1C resulted in a decrease of only 14% inthe propionate specific activity, whereas that ofacetate, on the other hand, was reduced 61 %. Thedata in Table 3 indicate that lactate-U-_4C con-tributed 96% of the carbon in propionate when itwas the sole energy source, with 4% arising from
1002 J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Dec
embe
r 20
21 b
y 18
3.80
.30.
18.

LACTATE METABOLISM BY V. PARVULA M4
TABLE 3. Acidic end products produced by resting cells of V. parvula M4 incubated with 14C-lactate, pyruvate,oxaloacetate, and 14C-lactate plus oxaloacetate in an atmosphere of nitrogen
Propionate Acetate
Expt Substrate Specific Specific Propionate/Concn" activity" Concn activity acetate
I Lactate-U-_4CC 60.7 1,428 40.8 1,402 1.492 Pyruvate 34.8 80.0 0.463 Oxaloacetate 29.0 65.4 0.444 Lactate-U-t4C + oxalo acetate 31.3 1,226 52.0 546 0.60
a Moles/ 100 moles of total substrate.I Distintegrations per minute per micromole.c Lactate-U-_4C specific activity = 1,491 dpm/gmole.
TABLE 4. Distribution of radioactive carbon in pro-pionate and acetate resulting from the utilization of
lactate-U-_4Ca plus oxaloacetate by cell-freeextracts of V. parvula M4
Per centcontribution
Specific OaoProducts Concn° activity Oxalo-acetateLac- + endo-tate genous
Propionate ........ 34.1 538 30.7 69.3Acetate ........... 47.8 776 41.4 58.6Propionate/acetate
ratio. 0.71
a Specific activity = 1791 dpm/Amole.I Moles/ 100 moles of total substrate.c Disintegrations per minute per micromole.
endogenous material. However, with lactate-U-14C plus oxaloacetate, 83% of the propionateoriginated from the lactate and 14% from theunlabeled oxaloacetate. On the other hand, 94%of the acetate carbon originated from lactate-U-14C when it was the sole energy source but only37% when oxaloacetate was the additional sub-strate. In this case, oxaloacetate provided 57% ofthe acetate carbon, indicating the presence of anactive oxaloacetate decarboxylase in cells of V.parvula M4. Endogenous metabolism contributed4 to 6% of the carbon to the respective end prod-ucts.Metabolism of cell-free extracts. The initial
oxidation of lactate by species of Veillonella wasshown to be catalyzed by malic-lactic transhydro-genase, which couples the oxidation of lactate topyruvate with the reduction of oxaloacetate tomalate (1, 5; E. F. Phares and M. V. Long, 1956,Amer. Chem. Soc. Meeting Abstr., p. 626). Thepresence of this enzyme in V. parvula M4 wasindicated by the inability of cell-free extracts todegrade sodium lactate-U-_4C (Fig. 4A). How-ever, when oxaloacetate was incubated with lac-tate in equimolar quantities, both substrates were
degraded at an initial rate of 36 units (Fig. 4B).No lactic dehydrogenase activity was observed incell-free extracts of V. parvula M4 assayedeither with lactate at pH 6.5 and 10.0 or withpyruvate atpH 6.5.
End-product analysis of the 60-min samplefrom the flask containing lactate-U-_4C plus oxal-
6 AO
0
E 0
en 6 B.
E
3
00 20 40 60
MINUTESFIG. 4. Metabolism of pyruvate, oxaloacetate, and
lactate plus oxaloacetate by cell-free extracts of V. par-vula M4 in an atmosphere of nitrogen. (A) Individualflasks contained 5 mg ofprotein/ml and 10 mm of thefollowing: sodium lactate-U-'4C (1,491 dpmhlmole), 0;potassium pyruvate, *; sodium oxaloacetate, A. (B)Combined incubation of 10 mm sodium lactate-U-"4Cplus 10 mm sodium oxaloacetate: sodium lactate-U-_4C,0; sodium oxaloacetate, A.
1003VOL. 105, 1971
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Dec
embe
r 20
21 b
y 18
3.80
.30.
18.

NG AND HAMILTON
oacetate (Fig. 4B) demonstrated that the deg-radation of those two substrates resulted in theformation of radioactive propionate and acetatein a ratio of 0.71 (Table 4), which was somewhathigher than that for resting cells (0.60). The dis-tribution of 14C into the acidic end productsshowed that the radioactive lactate contributed30% of the propionate carbon, whereas the re-maining 70% originated with oxaloacetate plusendogenous sources. As lactate-U-14C was notmetabolized by cell-free extracts, it was not pos-sible to calculate the contribution of carbon fromendogenous sources. Oxaloacetate plus endoge-nous material contributed slightly more to ace-tate production than did the lactate substrate.
Pyruvate and oxaloacetate were also metabo-lized by extracts, although the rates (13.3 and19.5 units, respectively) were lower than that forthe degradation of lactate plus oxaloacetate.Results to be presented elsewhere (in preparation)demonstrate that pyruvate degradation proceedsprincipally via the "phosphoroclastic" reaction(14) to acetate, carbon dioxide, and hydrogenwith only small quantities of propionate beingformed. Acetate and carbon dioxide were also themajor products of oxaloacetate metabolism, indi-cating that this substrate was decarboxylated byoxaloacetate decarboxylase followed by phos-phoroclastic cleavage of the pyruvate formed.Furthermore, as malic dehydrogenase activitywas observed in extracts of V. parvula M4, it isclear that oxaloacetate can also be converted tomalate (Table 5). The enzyme was completelyinhibited by 10 mM malonate.
DISCUSSIONThe stimulation of lactate metabolism in V.
gazogenes (11) by carbon dioxide and bicarbon-ate, as well as in those strains studied by Rogosa(16), is an important characteristic differentiatingthese organisms from our strain of V. parvula.
TABLE 5. Oxidation of reduced nicotinamide adeninedinucleotide (NADH) by cell-free extracts of V. par-
vula M4 in the presence and absence ofoxaloacetate (OAA)
Additions to basic Rate of NADHsystem, oxidation"
NADHC 612NADH + OAAc 9,600NADH + OAAC + malonate 12
a Basic system: extract (2.5 mg/ml) in 0.05 M phos-phate buffer,pH 6.5.
t Rate of NADH oxidation in nanomoles of NADHoxidized per milligram of protein per hour. Oxidationwas measured as the loss ofNADH at 340 nm.
c Component omitted from the blank.
The growth of V. parvula M4 in lactate brothwas not carbon dioxide-dependent (Table 1) norwas the rate of lactate metabolism by washedcells altered by the presence or absence of carbondioxide in the gas phase (in preparation). Fur-thermore, the rate and extent of lactate fermenta-tion as observed by Johns (11) and Rogosa (16)was still limited, even in the presence of carbondioxide. The latter author reported that only 6,umoles of lactate was metabolized by 20 mg (dryweight) of cells in 2 hr. By contrast, Fig. 3 illus-trates the degradation of 5 smoles of lactate permg (dry weight) of V. parvula M4 cells in 5 min.In addition, higher substrate to cell ratios pro-duced higher rates to a maximum of 102 4molesof lactate metabolized per mg .(dry weight) ofcells per hour (Fig. 1).The stoichiometry of lactate metabolism by V.
parvula M4 from the fermentation balance(Table 2) can be depicted in equation 1:
8 lactate = 5 propionate+ 3 acetate + 3 CO2 + H2 (1)
This equation is supported by the fermentationdata obtained with V. parvula M4 and with M.lactilyticus (6). The ratios of the end productsformed by V. gazogenes (1 1), however, varyslightly because equimolar quantities of acetateand carbon dioxide were not obtained. Thepathway of lactate degradation proposed byJohns (11) suggests that every molecule of carbondioxide fixed in the formation of oxaloacetate isrecovered in the subsequent decarboxylation ofsuccinate. Thus, the degradation of pyruvate toacetate, carbon dioxide, and hydrogen would bethe only reaction responsible for net carbon di-oxide evolution, such that acetate, carbon diox-ide, and hydrogen should theoretically appear inequimolar quantities. This is confirmed by morerecent evidence that methylmalonyl-coenzyme Ais ultimately decarboxylated to propionate andcarbon dioxide (7), rather than being coupled tooxaloacetate formation from pyruvate by methyl-malonyl-coenzyme A transcarboxylase, as is thecase with the propionibacteria (21).The validity of equation I can be tested by
examining the hypothetical production and con-sumption of reducing power at various steps oflactate metabolism by V. parvula M4. By ap-proximating the values in Table 2 for this orga-nism, it can be seen that 106 moles of substratelactate (100 moles added plus 5.5 moles of endog-enous "lactate" as calculated from total re-covery) produced the following moles of product:propionate, 66; acetate, 40; carbon dioxide, 40;hydrogen, 14. Accordingly, the initial conversionof lactate by malic-lactic transhydrogenase (equa-tion 2) would require the presence of 106moles of oxaloacetate and would produce 106
1004 J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Dec
embe
r 20
21 b
y 18
3.80
.30.
18.

LACTATE METABOLISM BY V. PAR VULA M4
moles of pyruvate and malate.
lactate + oxaloacetate pyruvate + malate (2)
A reaction producing reducing power duringlactate metabolism is the phosphoroclastic deg-radation of pyruvate. From the fermentationbalance (Table 2), 40 moles of pyruvate was
theoretically degraded to 40 moles of acetate,carbon dioxide, and hydrogen. However, only 14moles of hydrogen was actually produced by hy-drogenase during metabolism, indicating that 26electron pairs (40 - 14 = 26) were utilized toreduce another component of the system.The remaining pyruvate (106 - 40 = 66)
would be converted to 66 moles of oxaloacetateby carbon dioxide fixation, and the oxaloacetatethus formed would be subsequently degraded to66 moles of propionate. Net carbon dioxide con-sumption is not involved in this pathway as suc-
cinate decarboxylation regenerates any carbondioxide fixed with pyruvate. Obviously, the oxal-oacetate formed during this reaction could drivemalic-lactic transhydrogenase with the net forma-tion of 66 moles of malate and pyruvate. How-ever, as 106 moles of lactate was consumed, anadditional 40 moles of oxaloacetate would haveto be generated for this purpose. This could beaccomplished by recycling the malate formed inthe transhydrogenase reaction back to oxaloace-tate either by the combined action of malic en-zyme and pyruvate carboxylase (in preparation)or directly with malic dehydrogenase without dis-turbing the stoichiometry of propionate produc-tion, i.e., 66 moles of oxaloacetate to 66 pro-pionate. The net effect would be the productionof reducing power in the form of 40 moles ofNADH or reduced nicotinamide adenine dinu-cleotide phosphate (NADPH). This NADH or
NADPH, coupled with the 26 electron pairs pro-duced during pyruvate catabolism, would be suf-ficient to drive the reduction of 66 moles of fu-marate to succinate by succinic dehydrogenase.The only requirement for this sequence of eventswould be a suitable electron transport system.
ACKNOWLEDGMENT
This investigation was supported by grant A-4426 from theNational Research Council of Canada.
LITERATURE CITED
1. Allen, S. H. G. 1966. The isolation and characterization ofmalate-lactate transhydrogenase from Micrococcus lac-
1005
tilyticus. J. Biol. Chem. 241:5266-5275.2. Bulen, W. A., J. E. Varner, and R. C. Burrell. 1952. Sepa-
ration of organic acids from plant tissues. Chromato-graphic techniques. Anal. Chem. 24:187-190.
3. Cohen, L. H., and W. K. Noell. 1960. Glucose catabolismof rabbit retina before and after development of visualfunction. J. Neurochem. 5:253-276.
4. Delwiche, E. A. 1948. Mechanism of propionic acid forma-tion by Propionibacterium pentosaceum. J. Bacteriol. 56:811-820.
5. Dolin, M. 1., E. F. Phares, and M. V. Long. 1965. Boundpyridine nucleotide of malic-lactic transhydrogenase.Biochem. Biophys. Res. Commun. 21:303-310.
6. Foubert, E., Jr., and H. C. Douglas. 1948. Studies on theanaerobic Micrococci. 11. The fermentation of lactate byMicrococcus lactilyticus. J. Bacteriol. 56:35-36.
7. Galivan, J. H., and S. H. G. Allen. 1968. Methyl malonylcoenzyme A decarboxylase. Its role in succinate decar-boxylation by Micrococcus lactilyticus. J. Biol. Chem.243:1253-1261.
8. Hamilton, 1. R. 1968. Synthesis and degradation of intra-cellular polyglucose in Streptococcus salivarius. Can. J.Microbiol. 14:65-77.
9. Hamilton, 1. R., R. H. Burris, and P. W. Wilson. 1965.Pyruvate metabolism by nitrogen-fixing bacterium.Biochem. J. 96:383-389.
10. Johns, A. T. 1951. Isolation of a bacterium, producing pro-
pionic acid, from the rumen of sheep. J. Gen. Microbiol.5:317-325.
11. Johns, A. T. 1951. The mechanism of propionic acid for-mation by Veillonella gazogenes. J. Gen. Microbiol. 5:
326-336.12. Krebs, H. A., R. Hems, M. J. Weidemann, and R. N.
Speake. 1966. The fate of isotopic carbon in kidneycortex synthesizing glucose from lactate. Biochem. J.101:242-249.
13. Layne, E. 1951. Spectrophotometric and turbidimetricmethods for measuring proteins, p. 447-454. In S. P.Colowick and N. 0. Kaplan (ed.), Methods in enzymol-ogy, vol 3. Academic Press Inc., New York.
14. McCormick, N. G., E. J. Ordal, and H. R. Whiteley. 1962.Degradation of pyruvate by Micrococcus lactilyticus. 1.General properties of the formate-exchange reaction. J.Bacteriol. 83:887-898.
15. Rogosa, M. 1956. A selective medium for the isolation andenumeration of the Veillonella from the oral cavity. J.Bacteriol. 72:533-536.
16. Rogosa, M. 1964. The genus Veillonella. 1. General, cul-tural, ecological, and biochemical considerations. J. Bac-teriol. 87:162-170.
17. Rogosa, M. 1965. The genus Veillonella. IV. Serologicalgroupings and species emendations. J. Bacteriol. 90:704-709.
18. Rogosa, M., and F. S. Bishop. 1964. The genus Veillonella.11. Nutritional studies. J. Bacteriol. 87:574-580.
19. Rogosa, M., and F. S. Bishop. 1964. The genus Veillonella.Ill. Hydrogen sulphide production by growing cultures.J. Bacteriol. 88:37-41.
20. Rogosa, M., R. J. Fitzgerald, M. E. Mackintosh, and A. J.Beaman. 1958. Improved medium for selective isolationof Veillonella. J. Bacteriol. 76:455-456.
21. Swick, R. W., and H. G. Wood. 1960. The role of transcar-boxylation in propionic acid fermentation. Proc. Nat.Acad. Sci. U.S.A. 46:28-41.
VOL. 105, 1971
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Dec
embe
r 20
21 b
y 18
3.80
.30.
18.


















