
Journal of Controlled Release - cpu.edu.cn
14
ROS-triggered and regenerating anticancer nanosystem: An effective strategy to subdue tumor's multidrug resistance Zhigui Su a , Minglei Chen a , Yanyu Xiao a , Minjie Sun a , Li Zong a, ⁎, Sajid Asghar a,c , Mei Dong b , Huipeng Li a , Qineng Ping a, ⁎, Can Zhang a a State Key Laboratory of Natural Medicines, China Pharmaceutical University, 24 Tong Jia Xiang, Nanjing 210009, PR China b Key Laboratory of Human Functional Genomics of Jiangsu Province, School of Basic Medical Science, Nanjing Medical University, 140 Hanzhong Road, Nanjing 210029, PR China c College of Pharmacy, Government College University Faisalabad, Faisalabad, Pakistan abstract article info Article history: Received 15 June 2014 Accepted 21 September 2014 Available online 30 September 2014 Keywords: Tumor microenvironment Reactive oxygen species Targeted drug delivery nanosystem Multidrug resistance ROS-triggered and regenerating Drug delivery strategies utilizing tumor microenvironment are recognized as a critical doorway to overcome multidrug resistance (MDR). However, the variability of tumor microenvironment at different disease stages would definitely minimize stimuli generation and eventually the therapeutic effects of these stimuli sensitive systems. Herein, we report a unique reactive oxygen species (ROS) triggered nanosystem that can replenish the ROS upon disassembly to maintain its high level. This was accomplished by a new amphiphilic polymer (TBH) composed of D-α-tocopherol polyethylene glycol 1000 succinate (TPGS), hyaluronic acid (HA) and arylboronic ester. As a linker of TPGS to HA, arylboronic ester could efficiently degrade in response to ROS resulting in dismantling of nanosystem followed by rapid release of TPGS. Owing to ROS inducing activity of TPGS with mitochondrial respiratory complex II, ROS regeneration was observed for TBH nanosystem both in MCF-7/ADR cells and tumor tissues xenografted with MCF-7/ADR cells. Furthermore, doxorubicin-loaded TBH nanosystem (DOX-TBH) revealed higher drug cytotoxicity due to enhanced retention effect on account of ROS triggered DOX release and P-gp inhibitory mechanism of TPGS. Moreover, HA significantly improved tumor targeting capability of DOX-TBH, while ROS based triggering and regenerating mechanism lead to marked inhi- bition of the tumor growth in the xenograft MCF-7/ADR tumor-bearing nude mice. Crown Copyright © 2014 Published by Elsevier B.V. All rights reserved. 1. Introduction Cancer, one of the most challenging and complex global maladies, is affecting the life of people around the world. Despite multiple anti- cancer strategies have been used, chemotherapy continues to be an important therapeutic option for different malignancies, especially for primary advanced and metastatic tumors. However, the efficacy of chemotherapy is substantially limited by the intrinsic and acquired re- sistance of cancer cells to the anticancer drugs, which contributes to therapeutic failure and tumor relapses in over 90% of patients [1]. Multidrug resistance (MDR) in cancer cells is a complex phenomenon that may result from numerous mechanisms. The overexpression of pumps of the ATP-binding cassette superfamily (ABCs) present within the cancer cell membrane is one of the most well-known mechanisms, leading to reduced drug uptake. This requires excessive drug doses to kill tumor cells but that can induce severe adverse side effects on healthy tissues [2,3]. With the advancement of drug delivery technologies, a series of new drug delivery systems (DDS) has been developed to provide many alternative strategies to circumvent MDR based on overexpression of ABCs. But, the poor cellular uptake and insufficient intracellular drug re- lease remain rate-limiting steps in reaching the drug concentration level within the therapeutic window [4]. Therefore, a site-dependent drug release character triggered by intracellular pH [5,6], redox property [7] or enzyme levels [8,9] could be vital to obtain a sufficiently high intra- cellular level of drugs for overcoming MDR. Recently, one of the most popular approaches is to target redox property of cancer cells, utilizing disulfide or thiolysis bonds as triggered linkers owing to their tendency to be cleaved by elevated intracellular glutathione (GSH) [10–13]. How- ever, redox property of cancer cells is a dynamic milieu [14], because cancer cells frequently develop multiple genetic alterations. Redox property changes during cancer development and disease progression, even one tumor cell at different stages may have different redox signal levels [15]. To counter the varying stimuli level, some studies designed the nanocarriers by using double stimuli responsive strategy: sensitive to both GSH and ROS [16] or GSH and pH [17]. Therefore, a delivery sys- tem that aims to amplify levels of triggered signal to satisfy the signal response needs would have important implications for potential antican- cer therapeutics. Xiong et al. revealed that a lipase-sensitive polymeric nanogel selectively degraded through fabrication of a special lipase accumulated tumor environment [18]. Maltzahn et al. constructed the Journal of Controlled Release 196 (2014) 370–383 ⁎ Corresponding authors. Tel./fax: +86 25 83271092. E-mail addresses: [email protected] (L. Zong), [email protected] (Q. Ping). http://dx.doi.org/10.1016/j.jconrel.2014.09.020 0168-3659/Crown Copyright © 2014 Published by Elsevier B.V. All rights reserved. Contents lists available at ScienceDirect Journal of Controlled Release journal homepage: www.elsevier.com/locate/jconrel
Transcript of Journal of Controlled Release - cpu.edu.cn

Journal of Controlled Release 196 (2014) 370–383
Contents lists available at ScienceDirect
Journal of Controlled Release
j ourna l homepage: www.e lsev ie r .com/ locate / j conre l
ROS-triggered and regenerating anticancer nanosystem: An effectivestrategy to subdue tumor's multidrug resistance
Zhigui Su a, Minglei Chen a, Yanyu Xiao a, Minjie Sun a, Li Zong a,⁎, Sajid Asghar a,c, Mei Dong b, Huipeng Li a,Qineng Ping a,⁎, Can Zhang a
a State Key Laboratory of Natural Medicines, China Pharmaceutical University, 24 Tong Jia Xiang, Nanjing 210009, PR Chinab Key Laboratory of Human Functional Genomics of Jiangsu Province, School of Basic Medical Science, Nanjing Medical University, 140 Hanzhong Road, Nanjing 210029, PR Chinac College of Pharmacy, Government College University Faisalabad, Faisalabad, Pakistan
⁎ Corresponding authors. Tel./fax: +86 25 83271092.E-mail addresses: [email protected] (L. Zong), pin
http://dx.doi.org/10.1016/j.jconrel.2014.09.0200168-3659/Crown Copyright © 2014 Published by Elsevie
a b s t r a c t
a r t i c l e i n f oArticle history:Received 15 June 2014Accepted 21 September 2014Available online 30 September 2014
Keywords:Tumor microenvironmentReactive oxygen speciesTargeted drug delivery nanosystemMultidrug resistanceROS-triggered and regenerating
Drug delivery strategies utilizing tumor microenvironment are recognized as a critical doorway to overcomemultidrug resistance (MDR). However, the variability of tumor microenvironment at different disease stageswould definitely minimize stimuli generation and eventually the therapeutic effects of these stimuli sensitivesystems. Herein, we report a unique reactive oxygen species (ROS) triggered nanosystem that can replenishthe ROS upon disassembly to maintain its high level. This was accomplished by a new amphiphilic polymer(TBH) composed of D-α-tocopherol polyethylene glycol 1000 succinate (TPGS), hyaluronic acid (HA) andarylboronic ester. As a linker of TPGS to HA, arylboronic ester could efficiently degrade in response to ROSresulting in dismantling of nanosystem followed by rapid release of TPGS. Owing to ROS inducing activity ofTPGS with mitochondrial respiratory complex II, ROS regeneration was observed for TBH nanosystem both inMCF-7/ADR cells and tumor tissues xenografted with MCF-7/ADR cells. Furthermore, doxorubicin-loaded TBHnanosystem (DOX-TBH) revealed higher drug cytotoxicity due to enhanced retention effect on account of ROStriggered DOX release and P-gp inhibitory mechanism of TPGS. Moreover, HA significantly improved tumortargeting capability of DOX-TBH, while ROS based triggering and regenerating mechanism lead to marked inhi-bition of the tumor growth in the xenograft MCF-7/ADR tumor-bearing nude mice.
Crown Copyright © 2014 Published by Elsevier B.V. All rights reserved.
1. Introduction
Cancer, one of the most challenging and complex global maladies,is affecting the life of people around the world. Despite multiple anti-cancer strategies have been used, chemotherapy continues to be animportant therapeutic option for different malignancies, especially forprimary advanced and metastatic tumors. However, the efficacy ofchemotherapy is substantially limited by the intrinsic and acquired re-sistance of cancer cells to the anticancer drugs, which contributes totherapeutic failure and tumor relapses in over 90% of patients [1].Multidrug resistance (MDR) in cancer cells is a complex phenomenonthat may result from numerous mechanisms. The overexpression ofpumps of the ATP-binding cassette superfamily (ABCs) present withinthe cancer cell membrane is one of the most well-known mechanisms,leading to reduced drug uptake. This requires excessive drug dosesto kill tumor cells but that can induce severe adverse side effects onhealthy tissues [2,3].
With the advancement of drug delivery technologies, a series of newdrug delivery systems (DDS) has been developed to provide many
[email protected] (Q. Ping).
r B.V. All rights reserved.
alternative strategies to circumvent MDR based on overexpression ofABCs. But, the poor cellular uptake and insufficient intracellular drug re-lease remain rate-limiting steps in reaching thedrug concentration levelwithin the therapeutic window [4]. Therefore, a site-dependent drugrelease character triggered by intracellular pH [5,6], redox property [7]or enzyme levels [8,9] could be vital to obtain a sufficiently high intra-cellular level of drugs for overcoming MDR. Recently, one of the mostpopular approaches is to target redox property of cancer cells, utilizingdisulfide or thiolysis bonds as triggered linkers owing to their tendencyto be cleaved by elevated intracellular glutathione (GSH) [10–13]. How-ever, redox property of cancer cells is a dynamic milieu [14], becausecancer cells frequently develop multiple genetic alterations. Redoxproperty changes during cancer development and disease progression,even one tumor cell at different stages may have different redox signallevels [15]. To counter the varying stimuli level, some studies designedthe nanocarriers by using double stimuli responsive strategy: sensitiveto both GSH and ROS [16] or GSH and pH [17]. Therefore, a delivery sys-tem that aims to amplify levels of triggered signal to satisfy the signalresponse needswouldhave important implications for potential antican-cer therapeutics. Xiong et al. revealed that a lipase-sensitive polymericnanogel selectively degraded through fabrication of a special lipaseaccumulated tumor environment [18]. Maltzahn et al. constructed the

371Z. Su et al. / Journal of Controlled Release 196 (2014) 370–383
communicating nanoparticle systems composed of signaling and receiv-ing modules that activate the coagulation cascade in target tumors [19].Inspired by the fact that magnifying diseased tissue signal could signifi-cantly improve targeting in biological systems in vivo, we speculatethat constructing a constant high level of ROS in cytoplasm as amplifyingsignaling modules would facilitate the action of ROS signal sensitivenanocarriers.
Herein, we report a novel ROS triggering and positive feedbacknanosystem assembled from a new amphiphilic polymer composed ofD-α-tocopherol polyethylene glycol 1000 succinate (TPGS), hyaluronicacid (HA) and arylboronic esters to effectively overcome multidrug re-sistance. As shown in Fig. 1, to achieve a positive feedback ROS signalin cytoplasm, we selected α-tocopheryl polyethylene glycol succinate(TPGS), a polyethylene glycol (PEG) conjugate of α-tocopheryl succi-nate (α-TOS). TPGS is not only an effective P-gp efflux inhibitor [20]and selective cancer cell killing agent [21], but is also a powerful ROSinducer upon its interaction with mitochondrial respiratory complex II[22,23]. In addition, the arylboronic ester was chosen as ROS sensinglinker because of its facile ROS mediated degradation at physiologicalpH and temperature, which has been studied for imaging and targetingof tumor cells [24]. Amphiphilic material (TBH)was prepared by linkinghyaluronic acid (HA) as hydrophilic chains to the hydrophobic TPGSvia arylboronic esters. Besides its biocompatibility, biodegradabilityand ease of modifications [25], HA is an active target ligand for CD44positive malignant cells [26], which would be beneficial to increasethe nanocarrier uptake.
As displayed in Scheme 1, to check the effectiveness of the nanosys-tem, doxorubicin (DOX) as an anticancer agent that is resisted by cancer
Fig. 1. ROS-triggered degradation of TBH. (a) Scheme of TBH disassembly under H2O2. (b) The Gratio of TBH in time coursewith 1mMH2O2. (d) The change of particle size of TPGS, TBH and THand TH in the presence of different concentrations of H2O2 after 4 h of incubation.
cells was loaded in TBH micelles (DOX-TBH). After CD44 mediated en-docytosis, the arylboronic ester in DOX-TBH was expected to be trig-gered by intracellular ROS leading to the disassembly of DOX-TBH andburst release of TPGS and DOX. Upon interaction with mitochondrialrespiratory complex II, TPGS induces the generation of ROS whichmay create a kind of ROS positive feedback microenvironment withpersisting high levels of ROS generation necessary for degradation ofTBH. In addition, TPGS acts as a P-gp inhibitor to hinder the efflux ofDOX, which might ensure high intracellular levels of drugs, sufficientfor overcoming MDR and inducing programmed cell death.
2. Materials and methods
2.1. Materials
D-α-Tocopherol polyethylene glycol 1000 succinate (TPGS) andDCFH-DA were purchased from Sigma-Aldrich Co. LLC. (Shanghai,China). DCFH-DA Reactive Oxygen Species Assay Kit, DiO, DiI, andAnnexin V-FITC/PI Apoptosis Detection Kit were purchased fromBeyotime Institute of Biotechnology Co. LLC. (Nantong, China).Hyaluronic acid (molecular weights 100 kDa) was purchased fromFreda Biochem Co., Ltd. (Shandong, China). Doxorubicin HCl (DOX)was purchased from Internet Aladdin Reagent Database Inc. (Shanghai,China). DMEM (Hyclone®), fetal bovine serum (FBS, Hyclone®),penicillin–streptomycin solution (Hyclone®), trypsin (Hyclone®),phosphate buffered saline (PBS, Hyclone®) and MTT were provided bySunshine Biotechnology Co., Ltd. (Nanjing, China). All other chemicalsand reagents were HPLC or analytical grade.
PC image of TBH at different times when incubatedwith 1 mMH2O2. (c) The degradationin different incubation timeswith 1mMH2O2. (e) The change of particle size of TPGS, TBH

Scheme 1. The schematic presentation of ROS-triggered and regenerating anticancer nanosystem. A: TBH self-assembling nanosystem (DOX-TBH) and endocytosis mediated by HAthrough the overexpressed CD44 receptors; B: the breakage of arylboronic linkers by ROS, followed byDOX-TBH rapid disassembly and released DOX and TPGS; C: By acting onmitochon-drial respiratory complex II, the released TPGS induced ROS generation for complete disassembly of DOX-TBH; D: TPGS as the inhibitor of P-gp efflux to overcomeMDR; E: DOX as well asROS induced cell death in nucleus.
372 Z. Su et al. / Journal of Controlled Release 196 (2014) 370–383
2.2. Cell culture
P-gp overexpressing and DOX-resistant human breast adenocarcino-ma cells (MCF-7/ADR) were used for the cell studies. MCF-7/ADR werecultured in DMEM medium, supplemented with 10% FBS, 100 U/mL ofpenicillin and 100 μg/mL of streptomycin at 37 °C using a humidified5% CO2 incubator.
2.3. Animals
The female athymic nude mice were obtained from the ShanghaiSilaike Laboratory Animal Limited Liability Company and housed onstandard laboratory diet at an ambient temperature and humidity inair-conditioned chambers. All the animals were pathogen-free andallowed to access food and water freely. All animal experiments wereconducted in full compliancewith local, national, ethical and regulatoryprinciples with the approval of the Institutional Animal Care and UseCommittee at China Pharmaceutical University. To construct xenograftMCF-7/ADR tumor-bearing nude mice, approximately 107 of MCF-7/ADR cells were incubated subcutaneously in the flank region of femaleathymic nude mice. Tumor volume (V) was determined by measuringlength (L) and width (W), and calculated as Eq. (1):
V ¼ 1=2� L�W2: ð1Þ
2.4. Synthesis and characterization of TBH
The synthesis of TBH was described in the Supplementary data. Forcomparison, composed of HA and TPGS another amphiphilic polymerwithout ROS sensitivity was synthesized and named as TH.
Gel-permeation chromatography (GPC) was used to evaluate thedegradation of TBH in 1 mM H2O2 at different incubation times. GPCwas performed using a Shimadzu LC20A HPLC system (Kyoto, Japan),a refractive index detector (RID-10A, Shimadzu Co.), LC solution soft-ware (Shimadzu Co.), and a SB-803 HQ column (300 mm × 8 mm,Shodex, ShokoAmerica, Inc, Japan). Themobile phasewas 0.1M sodium
nitrate at a flow rate of 1 mL/min, and the analyses were performed at40 °C. The injection volume was 20 μL. TBH was dissolved in deionizedwater with 1 mM H2O2. After 0.25, 0.5, 1, 2, 4 and 8 h incubation,20 μL of different samples was withdrawn and analyzed by GPC. Thecontent of degradation products of TPGSwas calculated by the standardcurve of TPGS. The degradation ratio of TBH was calculated as Eq. (2):
Dregadation ratio of TBH %ð Þ ¼ WD =WT � 100%: ð2Þ
whereWDwas the content of degradation products of TPGS andWTwasthe total content of TPGS which was determined by colorimetric meth-od as described by Peracchia et al. [27]. In addition, after 4 h incubationTBH solution was ultra-filtered with a molecular weight cutoff 10 kDaand then the ultrafiltrate was lyophilized and analyzed by 1H NMR inCDCl3. For comparison, HA, TPGS, mixture of HA and TPGS or TH wasalso analyzed by GPC.
Next, the dynamic light scattering (DLS)was also utilized tomonitorthe change of particle size when TPGS, TBH and TH were treated withH2O2. To evaluate time-dependent ROS degradation, TPGS, TBH andTH solution were firstly mixed with H2O2 in which the concentrationof H2O2 was fixed at 1 mM and incubated for 0.25, 0.5, 1, 2, 4 and 8 h.To investigate concentration-dependent ROS degradation, TPGS, TBHand TH solution were incubated with different concentrations of H2O2
in a range of 0, 0.05, 0.2, 0.5, 1 and 2 mM for 4 h. After incubation,3 mL of TPGS, TBH and TH solution was withdrawn and particle sizewas monitored with DLS (Brookhaven Instruments-ZetaPlus, USA).
2.5. Intracellular ROS detection
ROS generation inside cells was detected using DCFH-DA ReactiveOxygen Species Assay Kit. For quantitative evaluation of ROS generationefficacy, MCF-7/ADR cells were seeded in 24-well plates at a density of105 cells/well and incubated for 24 h. Then, the medium was removedand the cells were treated with HBSS for 20 min. To investigate the ef-fect of time-dependent ROS generation, cells were incubated with200 μL of TPGS, TBH or TH solution and 200 μL of DMEM without FBSin which the concentration of TPGS was fixed at 200 μg/mL for 0.5, 1,

373Z. Su et al. / Journal of Controlled Release 196 (2014) 370–383
2, 3, 4 and 8 h at 37 °C. To investigate the effect of concentration-dependent ROS generation, the medium was replaced with 200 μL ofTPGS, TBH or TH solution and 200 μL of DMEM without FBS in whichthe concentration of TPGS was in the range of 10, 20, 50, 100 and200 μg/mL, and incubated for 4 h at 37 °C. At the end of the incubationperiod, the supernatant was removed and the wells were washed threetimes with ice-cold PBS. After treatment with DCFH-DA for 30 min, thecells were washed three times with ice-cold PBS, harvested and resus-pended in PBS. The fluorescence was measured by flow cytometry(FACS-Calibur, BD Biosciences).
For qualitative analysis of ROS generation efficacy, MCF-7/ADR cellswere seeded in confocal dishes at a density of 3× 105 cells/dish. Follow-ing incubation with TPGS, TBH and TH (200 μg/mL calculated by TPGS)for 4 h, DCFH-DAwas loaded into the cells. After 30min incubation, cellswere washed three times with ice-cold PBS and fluorescence images oftreated cells were acquired using Confocal Laser Scanning Microscopy(CLSM) (Olympus FV1100, Japan).
2.6. Measurement of mitochondrial respiratory complex II activity
MCF-7/ADR cells were seeded in 24-well plates at a density of105 cells/well. Following incubation with TPGS, TBH and TH(200 μg/mL calculated by TPGS) for different times (0, 0.5, 1, 2, 4 and8 h), at the end of the incubation period the supernatant was removedand thewellswerewashed three timeswith ice-cold PBS. Subsequently,the cells were incubated with 200 μL of ice-cold cell lysis buffer for10 min. Then, samples were centrifuged to remove the cellular debrisat 4 °C. 20 μL aliquot of each cell lysate was used to determine the cellprotein content using BCA Protein Assay Kit. The activity of mitochon-drial respiratory complex II was measured according to the Mitochon-drial Complex II Activity Assay Kit (Germany, Novagen). The relativeactivity of mitochondrial respiratory complex II was calculated by theEq. (2):
Relative activity %ð Þ ¼ A tð Þ=A cð Þ � 100%: ð2Þ
where A(t) is the activity of mitochondrial respiratory complex II atdifferent times and A(c) is the activity ofmitochondrial respiratory com-plex II before incubation.
2.7. Ex vivo imaging studies for ROS generation in xenograft MCF-7/ADRtumor-bearing nude mice
When the tumor volume reached 0.2–0.3 cm3, mice were firstinjected with free DCFH-DA (2.5 mg/kg) by intratumoral injection,then intravenous injection of saline as control, TPGS, TBH or TH at adose of 50mg of TPGS/kg. Micewere anesthetized using intraperitonealinjection of chloral hydrate (10 mg/kg). Ex vivo fluorescence imagingexperimentswere performed at 1, 4, 8, 12 and 24 h post-injection utiliz-ing a Kodak multimodal-imaging system IS2000MM (Kodak, USA)equipped with an excitation bandpass filter at 460 nm and an emissionat 535 nm. Exposure timewas 30 s per image. Images were analyzed byCarestreamMolecular Imaging Software V 5.3.5 (Kodak ID Image Anal-ysis Software; Kodak). To confirm the biodistribution of DCFH-DAprobes in different organs, the mice were sacrificed at 24 h post-injection. Different organs were separated and washed by saline andassembled for ex vivo fluorescence imaging. In addition, the tumor sam-ples underwent frozen section examination with 7 μm thickness,stained with DAPI and observed by CLSM.
2.8. Preparation and characteristics of DOX-TBH and DOX-TH
DOX base was obtained by the overnight reaction of DOX.HCl withdouble mol ratio of triethylamine in formamide. DOX loading TBH(DOX-TBH) and TH (DOX-TH) micelles were assembled in a simpledialysis technique. 40 mg TBH or TH was dissolved in 2 mL of distilled
water. 1 mL of DOX formamide solution (4 mg/mL) was added intothe polymers solution under stirring for 1 h. The mixture solution wasdialyzed against distilled water using a dialysis bag (MWCO of3.5 kDa) for 24 h to remove free DOX. Dialyzed products were filteredthrough a 0.22 μmmicrofiltrationmembrane. Particle size, polydispersityindex and zeta potential of DOX-TBH and DOX-TH were determined byBrookhaven Instruments-ZetaPlus (Brookhaven, USA). The morphologi-cal observations were performed under TEM (H-600, Hitachi, Japan)following negative staining with 0.1 wt.% sodium phosphotungstatesolution. Particle degradation study was performed with 1 mMH2O2 toconfirm oxidation sensitive characteristic.
The concentration of DOX in DOX-TH andDOX-TBHwas determinedby Shimadzu 10-A vpHPLC system (Kyoto, Japan). The stationary phase,Diamonsil C18 column (150mm×4.6mm, 5 μm),was kept at 40 °C. Themobile phase consisted of 5 mM ammonium acetate (pH 3.5, 0.05%triethylamine): methanol (30:70) and was delivered at a rate of1.0 mL/min. The injection volume was 20 μL and effluent was moni-tored by a fluorescence detector (Shimadzu, Japan) with excitationwavelength of 480 nm and emission wavelength of 550 nm. DOX-TBHand DOX-THwere filtrated through the 0.22 μm cellulose nitrate mem-brane. The 50 μL of DOX-TBH or DOX-TH was dissolved in a mixturewith 250 μL formamide and 700 μL methanol, and then vortexed for30 s and centrifuged at 12000 rpm for 10 min. The drug concentrationsbefore dialysis and after filtration were determined. The encapsulationefficiency (EE) in DOX-TBH and DOX-TH was calculated as Eq. (3):
EE %ð Þ ¼ Ca � Vað Þ= Cb � Vbð Þ � 100% ð3Þ
where Ca is the concentration of the drug after filtration and Va isthe volume of DOX-TBH or DOX-TH solutions after filtration. Cb isthe concentration of the drug before dialysis and Vb is the volume ofDOX-TBH or DOX-TH solutions before dialysis.
2.9. In vitro drug release of DOX-TBH and DOX-TH
In vitro drug release of DOX-TBH and DOX-TH was analyzed by amembrane dialysis technique. In order to investigate ROS-triggered re-lease the study of drug release in vitro was done in 0.9% saline with orwithout 1 mM H2O2. 1 mL of DOX-TBH or DOX-TH (containing 200 μgof DOX) was placed into a dialysis bag (MWCO of 3.5 kDa). Meanwhile,DOX solution was prepared as control. The dialysis bag was tightenedand soaked in 50 mL of 0.9% saline with or without 1 mM H2O2. Theexperiments were carried out at 37 °C for 8 h in 3 replicates. Atpredetermined intervals, 1 mL of the dissolution medium waswithdrawn and replaced with the same amount of pre-warmed freshmedium. The samples of the replicates were analyzed by HPLC. Theconcentration of H2O2 was also determined by the method of ferrousoxidation in xylenol orange (FOX) assay [28].
To investigate the influence of different concentrations of H2O2 in0.9% saline on drug release, H2O2 was added into DOX-TBH and DOX-TH solution with different final concentrations of H2O2 (0, 0.05, 0.2,0.5, 1 and 2 mM). Free DOX was obtained by ultrafiltration methodwith molecular weight cutoff 10 kDa. The ratio of drug release fromDOX-TBH and DOX-TH was calculated as Eq. (4):
Drug release %ð Þ ¼ Cu=Co � 100% ð4Þ
where Cu is the concentration of DOX in the ultrafiltrate and C0 is thetotal concentration of DOX before ultrafiltration.
2.10. FRET analysis of TBH disassembly in vitro
Fluorescence resonance energy transfer (FRET) analysis was con-ducted as previously reported [29,30]. A FRET pair of hydrophobicdyes, DiO as donor and DiI as acceptor, was physically loaded into themicelles cores. To verify the occurrence or disappearance of FRET,

374 Z. Su et al. / Journal of Controlled Release 196 (2014) 370–383
fluorescence spectra ofDiO andDiI double loadedmicelles diluted in de-ionized water or deionized water with 1 mM H2O2 were measuredusing a fluorospectrophotometer (Shimadzu, RF-5301, Japan) with theexcitation at 484 nm and the emission scan from 400 nm to 700 nm.The FRET ratio (FR) was calculated as Eq. (5):
FR %ð Þ ¼ IR= IRþ IGð Þ � 100% ð5Þ
where IR and IG are fluorescence intensities at 565 nm and 501 nm,respectively. DiO and DiI double loaded TBH (FRET-TBH) and TH(FRET-TH)micelles were prepared by a simple dialysis technique. Brief-ly, 40mg TBH or THwas dissolved in 2mL of distilledwater. DiO andDiIwere dissolved in 1 mL DMSO and added into the polymer solutionunder stirring for 1 h. The mixture solution was dialyzed against dis-tilled water using a dialysis bag (MWCO of 3.5 kDa) for 24 h. Dialyzedproducts were filtered through a 0.22 μm microfiltration membrane.
MCF-7/ADR cells were incubated with FRET-TBH and FRET-TH for2 h at 37 °C andwashedwith ice-cold PBS, and then incubatedwith cul-ture medium at 37 °C for an additional 2, 3 and 4 h. CLSM images wereacquired with the excitation at 484 nm, and the emission between555 nm and 655 nm for DiI detection, as well as the emission between500 nm and 530 nm for DiO detection. The fluorescence intensity ofthe hold cells was analyzed by Leica Application Suite Advanced Fluo-rescence Version: 2.3.0 build 5131. The fluorescence intensity of DiOwas named as IG while the fluorescence intensity of DiI was named asIR. The FR was calculated as described in Eq. (5).
2.11. Determination of the intracellular accumulation, retention of DOX
MCF-7/ADR cells were seeded in 24-well plates at a density of105 cells/well and incubated for 24 h. Then, the medium was removedand the cells were treated with HBSS for 20 min. To investigate time-dependent uptake, cells were incubated with 200 μL of free DOX,DOX-TBH or DOX-TH suspension and 200 μL of DMEM without FBS inwhich the concentration of DOX was fixed at 10 μg/mL and incubatedfor 1, 2, 3 and 4 h at 37 °C. To investigate concentration-dependent up-take, the medium was replaced with 200 μL of free DOX, DOX-TBH orDOX-TH suspension and 200 μL of DMEMwithout FBS inwhich the con-centration of DOXwas in the range of 2–20 μg/mL and incubated for 4 hat 37 °C. At the end of the incubation period, the supernatant wasremoved and thewellswerewashed three timeswith ice-cold PBS. Sub-sequently, the cells were incubated with 200 μL of cell lysis buffer for10 min. Then, samples were centrifuged to remove the cellular debrisat 4 °C. 20 μL aliquot of each cell lysate was used to determine the cellprotein content using BCA Protein Assay Kit. The concentration of DOXin cell lysate was determined by HPLC as described above. The uptakeindex (UI) was calculated as Eq. (6) [31]:
UI ¼ C=P ð6Þ
where C is the concentration of DOX in cell lysate and P is the concentra-tion of protein in cell lysate.
To determine the intracellular retention of DOX, MCF-7/ADR cellswere cultured with either free DOX, DOX-TBH or DOX-TH for 4 h,washed with ice-cold PBS, and then incubated with culture medium at37 °C for an additional 0.5, 1, 2 and 4 h. Cells were lysed and the concen-trations of DOX in cell lysate were measured as described above. Theintracellular retention ratio was calculated as Eq. (7):
Relativeresidualamounts %ð Þ ¼ UI tð Þ=UI 0ð Þ � 100% ð7Þ
whereUI(t) is the value ofUI at different additional incubation times andUI(0) is the value of UI before additional incubation.
2.12. The cytotoxicity and apoptosis of free DOX, DOX-TBH and DOX-TH inMCF-7/ADR cells
MCF-7/ADR cells were seeded in 96-well plates at a density of5 × 103 cells/well and incubated for 24 h. MCF-7/ADR cells were incu-bated with free DOX, DOX-TBH and DOX-TH containing the sameconcentration of DOX (0.2–20 μg/mL) for 24 h and cytotoxicity wasmeasured with MTT assay. After incubation, 10 μL of MTT (5 mg/mL inpH 7.4 PBS) was added and incubated for another 4 h; the mediumwas then replaced with 150 μL DMSO. The UV absorbance intensity ofcells was measured by Microplate Reader (Thermo Electron Corpora-tion, USA) at 570 nm. Each point was performed in triplicates.
Apoptosis of MCF-7/ADR was detected using the Annexin V-FITC/PIApoptosis Detection Kit, the cells were incubated with free DOX,DOX-TBH and DOX-TH containing the same concentration of DOX(5 μg/mL) for 12 h. The following procedures were performed in accor-dance with the manufacturers' protocols. Finally, apoptosis was ana-lyzed by flow cytometry (FACS-Calibur, BD Biosciences).
2.13. CLSM Study for uptake of free DOX, DOX-TBH or DOX-TH
MCF-7/ADR cells were placed in the confocal dishes at a density of3 × 105 cells/dish. After 24 h of cell attachment, 500 μL of free DOX,DOX-TBH or DOX-TH (containing 10 μg/mL DOX)was added in the cul-ture medium and incubated for 2 h at 37 °C. After incubation, the cellswere washed thrice by ice-cold PBS and stained with Hoechst 33342for 30 min at 37 °C. Then, the cells were again washed three timeswith ice-cold PBS and observed by CLSM.
2.14. Ex vivo imaging studies for tumor target ability of TBH and THmicelles
To investigate in vivo dynamic distribution and tumor-targeting,DiR, a hydrophobic near infrared dye, was loaded into TBH and THmicelles (DiR-TBH and DiR-TH). The xenograft MCF-7/ADR tumor-bearing nude mice were intravenously administrated with DiR-TBH orDiR-TH (2.5 mg/kg) and anesthetized using intraperitoneal injectionof chloral hydrate (10 mg/kg). Ex vivo fluorescence imaging experi-ments were performed asmentioned above except that thewavelengthwas fixed at 720 nm for excitation and 790 nm for emission. To confirmthe biodistribution of DiR probes in different organs, the mice weresacrificed at 24 h post-injection. Different organs were separated andwashed by saline, assembled for fluorescence imaging and analyzedby Carestream Molecular Imaging Software V 5.3.5. In addition, thetumor samples underwent frozen section examinationwith 7 μm thick-ness, stained with DAPI and observed by CLSM.
2.15. In vivo biodistribution, inhibition of xenograft MCF-7/ADRtumor growth and analysis of apoptosis by TUNEL histology for free DOX,DOX-TBH and DOX-TH
The xenograft MCF-7/ADR tumor-bearing nude mice were intrave-nously injected with the free DOX, DOX-TBH and DOX-TH at DOXdose of 2.5 mg/kg. At indicated times (0.5, 4, 12 and 24 h) after injec-tion, blood samples were collected and mice were then sacrificed bycervical dislocation. Tissues were excised and lightly rinsed to removeany excess blood, weighed and stored at −20 °C until assay. The con-centration of DOX in plasma and homogenized tissues was determinedby HPLC.
For in vivo antitumor evaluation, the xenograft MCF-7/ADR tumor-bearing nude mice were weighed and randomly divided into fourgroups: 1) saline; 2) free DOX (2.5 mg/kg); 3) DOX-TBH (2.5 mg/kg);4) DOX-TH (2.5 mg/kg). The different formulationswere administratedvia tail vein at days 0, 2, 4, 6 and 8. Tumor size was measured everyother day. The survival time of 4 groups treated as above was alsomonitored. To evaluate the apoptotic response in tumor tissues, the ter-minal deoxynucleotidyl transferase (TdT)-mediated dUTP-digoxigenin

Fig. 2.Detection of amplifying ROS signal in vitro and suppressing the activity ofmitochondrial respiratory complex II. (a) The change of ROS level inMCF-7/ADR cells after incubationwithTPGS, TBH or TH for different periods of time when comparingwith basic ROS level (n= 3). The concentrations of TPGS, TBH and THwere fixed at 200 μg/mL calculated by TPGS. (b) Thechange of ROS level inMCF-7/ADR cells after incubationwith TPGS, TBHor TH for different concentrations of TPGS at 4 hwhen comparingwith basic ROS level (n=3). (c) The inhibition ofTPGS, TBH or TH on the activity of mitochondrial respiratory complex II. (d) Observation of intracellular ROS generation by DCFH-DA staining in MCF-7/ADR cells incubated with DMEM(as control), TPGS, TBH and TH for 4 h. The concentrations of TPGS, TBH and TH were fixed at 200 μg/mL calculated by TPGS. (Green for DCF, blue for Hoechst and scale bar = 50 μm).
375Z. Su et al. / Journal of Controlled Release 196 (2014) 370–383
nick-end labeling technique (TUNEL) and hematoxylin and eosin stain-ing (H&E) were applied to frozen section tumor samples (drawn atday 14).
2.16. Statistical analysis
Results are given as Mean ± S.D. Statistical significance was testedby two-tailed Student's t-test or one-way ANOVA. Statistical signifi-cancewas set at⁎P b 0.05 and extreme significance was set at ⁎⁎P b 0.01.
3. Results and discussion
3.1. Synthesis and characterization of TBH
TBH synthesis was confirmed by 1H NMR (Fig. S1a) and showed inFig. 1a. The graft ratio of TPGS (per 100 HA units) linked by arylboronicester under optimal conditionswas 10.9±1.3% (n=3). In addition, thecontent of boron determined by ICP-MS was 4.0% in TBH. Both surfacetension measurement and the steady-state fluorescence probe tech-nique were applied for determination of critical micelle concentration(CMC). The results revealed that the CMC value of TBH was about100 μg/mL. Todetermine theROS response of TBH in a cancerousmicro-environment, GPCwas used to evaluate the degradation of TBH in 1mMH2O2. After 4 h, a newpeak belonging to TPGSwas observed in the chro-matogramof TBH (Fig. 1b). Thismeant that ROS labile arylboronic estersin the TBH were sensitive to H2O2-mediated degradation [32]. 1H NMRimage of degradation product confirmed that the proton chemical shiftsbelong to TPGS and arylboronic groups (Fig. S3). Composed of HA andTPGS, another amphiphilic polymer (TH) without ROS sensitivity wassynthesized and characterized (Fig. S1b). The similar graft ratio ofTPGS (per 100 HA units) and CMC suggested the similar micellizationability and micellar stability toward dilution in vitro and in vivo. Inaddition, no TPGS peak was observed after first 4 h in GPC due to thelack of sensitivity to ROS microenvironment (Fig. S2).
The quantitative study of ROS triggered TBH degradation revealedthat the degradation characteristics of TBH also exhibited time-dependent course. As the incubation time increased to 4 h the degrada-tion ratio of TBH decreased more than 80% and reached a plateau(Fig. 1c).
The DLS results indicated that the change of particle sizewhen TPGS,TBH and TH were treated with H2O2, was time- and concentration-dependent. In Fig. 1d, even after 0.25 h incubation, the particle size ofTBH remarkably decreased from 42.3 nm to 28.1 nm. As the incubationtime increased to 1 h, the particle size of TBH decreased more than50%, which was similar to the particle size of TPGS. Besides, theconcentration-dependent ROS degradation revealed that the particlesize of TBH declined with the elevation of concentration of H2O2 andreached equilibrium stage when the concentration of H2O2 was higherthan 0.5 mM after 4 h incubation (Fig. 1e). But little changes in particlesize were found for TPGS and TH. The results of GPC and changes in theparticle size suggested that an efficient ROS triggered degradationoccurred in TBH.
3.2. Amplifying ROS signals in vitro
To demonstrate the capacity of signaling modules to induce ROS re-generation in MCF-7/ADR cells, non-fluorescent DCFH-DA was chosenas ROSmarker. After entering into the cell DCFH-DAwould behydrolyzedto DCF and yield a highly green fluorescent signal in the presence ofintracellular ROS such as H2O2 [33]. Therefore, the fluorescence intensitywas found to beproportional to the amount of ROS formed intracellularly.Quantitative analysis found that TBH could rapidly increase the level ofROS in MCF-7/ADR cells (Fig. 2a) due to fast release of TPGS. Even inthe initial 0.5 h, the level of ROS increased 3.0-fold for TBH when com-pared with base ROS level, which was 2.3-fold and 1.8-fold higher thanthat for TPGS and TH, respectively (P b 0.05). TBH micelles also showedstronger ability to heighten ROS level at each concentration point(Fig. 2b). Although TPGS, TBH and TH had the same content of TPGS,

Fig. 3. Ex vivo imaging studies for ROS generation in xenograft MCF-7/ADR tumor-bearing nudemice. (a) Fluorescence images of ROS generation in xenograft MCF-7/ADR tumor-bearingnude mice at 1, 4, 8, 12 and 24 h after intravenous injection of saline (I), TPGS (II), TBH (III) and TH (IV). (b) Fluorescence images of isolated organs separated from MCF-7/ADR tumor-bearing mice in different groups at 24 h. The numeric label for each organ is as follows: 1, tumor; 2, heart; 3, liver; 4, spleen; 5, lung; 6, kidney. (c) Quantitative analysis of fluorescentsignals from the tumors and normal tissues (n = 3) ⁎P b 0.05, ⁎⁎P b 0.01 (two-tailed Student's t-test). (d) CLSM images of ROS generation in tumor tissues in different groups (greenfor DCF, blue for DAPI in nucleus and scale bars = 250 μm).
376 Z. Su et al. / Journal of Controlled Release 196 (2014) 370–383
difference in ROS generation might be due to the internalization amountof TPGS in cells. Both TBH and TH could be internalized into MCF-7/ADRby CD44 mediated endocytosis with the help of HA [34]. The more theTPGS was internalized into cells the higher the ROS level was. Further-more, TBH possessed stronger ability of generating ROS than TH did.ROS response of TBH would release TPGS groups quickly, which could
Fig. 4. Preparation and characteristics of DOX-TBH and DOX-TH. (a) Characteristics of DOX-TBHand DOX-TH when incubated with plasma.
induce ROS generation and keep an elevated ROS signal modules in cyto-plasm. CLSM results revealed that TBH could inducemore strong fluores-cence intensity in MCF-7/ADR cells when compared to TPGS and TH,which was similar to the results of quantitative analysis (Fig. 2d).
Next, to confirm the mechanism of ROS intracellular generation inMCF-7/ADR cells, TPGS, TBH or TH was demonstrated to inhibit the
and DOX-TH by DLS and TEMwith or without 1 mMH2O2. (b) The stability of DOX-TBH

Fig. 5. ROS-triggered DOX release from DOX-TBH and DOX-TH. (a) The release of DOX at37 °C for free DOX,DOX-TBH andDOX-TH in saline. (b) The release of DOX at 37 °C for freeDOX, DOX-TBH and DOX-TH in salinewith 1mMH2O2. (c) The change of H2O2 concentra-tion in saline with 1 mM H2O2. (d) The influence of H2O2 concentration on drug release.
377Z. Su et al. / Journal of Controlled Release 196 (2014) 370–383
activity of mitochondrial respiratory complex II significantly (Fig. 2c),which would cause the electron leak from the respiratory chain torecombine with molecular oxygen to yield ROS [23]. This effect createda kind of intracellular microenvironment with persisting high levels ofROS for TBH degradation, making a cycle of ROS consumption and re-generation with positive feedback, hence, facilitating ROS responsivenanocarrier to deliver efficiently.
3.3. Ex vivo imaging studies for ROS generation in xenograft MCF-7/ADRTumor-bearing nude mice
Having probed the ROS generating potential of TBH at the cellularlevel, we next investigated the ability of TBH to generate ROS in thetumor site. The xenograft MCF-7/ADR tumor-bearing nude mice werefirst injected with free DCFH-DA (2.5 mg/kg) by intratumoral injection,then intravenous injection of saline as control, TPGS, TBH or TH. Asshowed in Fig. S4, for free DCFH-DA no fluorescence was foundwithoutoxidation in vitro. In Fig. 3a, after 1 h post-injection TBH and TH pro-duced tumor-specific fluorescence signal. As time increased, elevatedfluorescent signals were found at the tumor site for TBH and TH groupsas compared with saline and TPGS groups within 8 h post-injection,highlighting TBH and TH accumulation in tumor by EPR and CD44mediated active targeting. After 24 h post-injection, tumors as well asnormal tissues were harvested for ex vivo imaging. A high level of fluo-rescence was located in the tumor, while low signals were observed invital organs (Fig. 3b). The DCF was eliminated mainly by the kidney.For TPGS group, no obvious tumor-specific fluorescence occurred incomparisonwith TBH and THgroups, suggesting that TPGS could not ac-cumulate in the tumor or internalize by tumor cells effectively becauseof smallmolecule size and leaking ability of targeting to tumor. Thefluo-rescence signal of TBH at the tumor site was significantly higher thanthat of TPGS and TH, which was 4-fold and 2-fold using quantitativeanalysis, respectively (Fig. 3c). Histopathologic analysis further verifiedthat TBH strongly induced ROS generation and accumulation in tumorcells than TPGS and TH (Fig. 3d).
Giving a thought to safety, in Fig. S5, TBH and TH were non-toxic orlow-toxic to normal, i.e. primary brain capillary endothelial cells(BCBCs) and normal hepatocytes (L02). As it was reported previously,due to the generally higher levels of esterase in normal cells such as he-patocytes, colonocytes, fibroblasts or cardiomyocytes, the succinateester structure of TPGS would be more easily cleaved to produce vita-min E in these tissues [21]. In addition, the biological compatibility eval-uation exhibited that no significant difference was found for the bloodroutine examination after i.v. administration of different materials(Fig. S6a). Both of TBH and TH did not alter the level of IL2, IL6 andINF-λ in serum as well as in spleen, suggesting that TBH and TH didnot induce acute inflammation (Fig. S6b). The level of ALT/GPT,AST/GOT and BUN was the same as the saline group, suggesting thatno liver and kidney damage occurred after consecutive injection ofTBH and TH in healthy mice (Fig. S6c). Moreover, total antioxidantcapacity assay revealed that after continuous injection of TBH and THit still did not change their total antioxidant potential and ROS accumu-lation was found after treatment in normal tissues, further confirmingthe biological compatibility of TBH and TH (Fig. S7).
3.4. Preparation and characteristics of DOX-TBH and DOX-TH
Experiments were conducted to demonstrate whether and how thedrug delivery nanosystem could help the loaded drug accumulation andretention in MCF-7/ADR cells induced by extra ROS generation andTPGS release after the micelle degradation. DOX-TBH and DOX-THwere prepared by a simple dialysismethod. The entrapment efficienciesof DOX-TBH and DOX-TH were 66.35 ± 5.42% and 63.76 ± 6.33% withparticle sizes of 83 ± 2.6 nm and 107 ± 3.9 nm and zeta potential of−14.2± 1.7mV and−10.9± 0.8mV, respectively (n=3). DLS resultsshowed that in the present of 1 mM H2O2 the particle size of DOX-TBH
sharply decreased;meanwhile, TEM image confirmed that after incuba-tion in 1 mM H2O2, the particle size of DOX-TBH changed sharply from80 nm to 30 nm and the structure became loose and coarse due to ROSlabile nature of the DOX-TBH (Fig. 4a). On the other hand, no significantdifferencewas found for DOX-THwhen incubatedwith orwithout H2O2
(Fig. 4a). Asmention above, the change of particle size for DOX-TBHwasdue to the degradation of arylboronic ester in TBH and released TPGS,which would result in smaller particle size.
In addition, in Fig. 4b the stability of DOX-TBH and DOX-TH in plas-ma revealed that particle size could remain stable during the first 2 h,suggesting that particles could survive in the plasma and prolongblood circulation at first period. According to the “Vroman effect” theplasma protein adsorption was time dependent [35]. Proteins withhigh concentrations but lower affinity in plasma would initially occupythe surface of the nanoparticles, and then theywould be replaced by theproteins with low concentration but higher affinity in plasma, whichwould cause a rapid and irreversible change of particle size [36]. Asthe incubation time increases the particle size of DOX-TBH and DOX-TH increases dramatically.
3.5. In vitro drug release of DOX-TBH and DOX-TH
Both DOX-TBH and non-ROS-responsive DOX-TH, which consistedof the same amount of DOX, exhibited a sustained DOX release com-pared with the DOX solution in saline (Fig. 5a). More importantly, therelease rate of DOX-TBH increased observably in the presence of1 mM H2O2, while there was a slight change in the release rate ofDOX-TH (Fig. 5b). The faster release might be due to the degradationof arylboronic esters under H2O2, which would undergo a triggered dis-assembly of DOX-TBH leading to expulsion of hydrophobic core (TPGSand DOX). Furthermore, a fast decline in the concentration of H2O2
in vitro was observed in the case of DOX-TBHwhich could be attributedto consumption by oxidative degradation of arylboronic esters. Whilefor free DOX and DOX-TH, the concentration of H2O2 decreased slightlyover 8 h (Fig. 5c). Besides, the burst release demonstrated by DOX-TBHwas dependent on the concentration of H2O2 (Fig. 5d). With the in-crease in concentration of H2O2, more DOX-TBHwould degrade leadingto complete drug release. It can be safely assumed that the disassemblyof ROS-triggered nanocarrier is also dependent on the ROS levels intumor cells, the lower ROS level would result in the hampered degrada-tion of nanocarrier and slower release of cargo. It is obvious that ROS

Fig. 6. Intracellular FRET analysis of TBH disassembly. (a) Scheme of FRET-TBH assembly in water and disassembly in 1 mMH2O2. (b) Fluorescence emission spectra of FRET-TBH dilutedwith water (red curve) and 1mMH2O2 (green curve) observed at 484 nm excitation wavelength. (c) Fluorescence emission spectra of FRET-TH dilutedwith water (red curve) and 1mMH2O2 (green curve) observed at 484 nmexcitation wavelength. (d) Confocal images of MCF-7/ADR cells incubatedwith FRET-TBH and FRET-THmicelles for additional 2, 3 and 4 h. (greenfor DiO, red for DiI and scale bar = 75 μm).
378 Z. Su et al. / Journal of Controlled Release 196 (2014) 370–383
regeneration in cancer cells would favor the triggered disassembly andsubsequent burst delivery of the payload.
3.6. Intracellular ROS-triggered release of TBH
To illustrate the ROS-triggered release of the core-loadeddrug in realtime in cells, we physically loaded a FRET pair of hydrophobic dyes, DiOas donor and DiI as acceptor, into TBH and TH micelles (referred to asFRET-TBH and FRET-TH) and showed in Fig. 6a. Fluorescence spectraat 484 nm showed that for micelles in deionized water, a strong DiI sig-nal was observed (red curve in Fig. 6b and c) due to the close proximityof DiO and DiI in the micelle core (0–10 nm), with a FRET ratio (FR) of0.95 and 0.96 for FRET-TBH and FRET-TH, respectively. After decompo-sition by 1 mMH2O2, FRET signal plummeted for FRET-TBH groups andthe FR declined from 0.96 to 0.13 during 2 h period (green curve inFig. 6b), indicating the release of core-loaded probes tomediumanddis-assembly of micelles. On the contrary, the FRET signal decreased gradu-ally for FRET-TH and the FR of FRET-TH was 0.74 after 2 h incubation,which was significantly higher than that of FRET-TBH (green curve inFig. 6c). These results suggested that FRET pair release from TBHnanocarrier was more sensitive to H2O2 than that of TH nanocarrier.The degradation of arylboronic esters triggered by H2O2 could be themost likely cause of rapid disassembly of FRET-TBH leading to therelease of FRET pair.
CLSM image revealed that FRET signal of FRET-TBH disappearedfrom MCF-7/ADR cells after an additional 2 h of incubation (Fig. 6d).The fluorescent signal analyzed from the hold cells showed that the FR
of FRET-TBH decreased to 0.2 and only DiO signal (green) was observedclearly, indicating that fast release of FRET pair or micelles disassemblyoccurred intracellularly. Further monitoring FRET signal of FRET-TBHup to 4 h showed a constantly higher disassembly behavior and no sig-nificant change was found for the FR from 2 to 4 h. As anticipated forFRET-TH, owing to its poor ROS sensitivity and subsequently delayeddisassembly, the reduction of FRET was much slower andweaker as ad-ditional incubation time increased from 2 to 4 h (from red to green).With a FR near to 0.2, most of FRET-TH disassembled after an additional4 h of incubation.
To confirm the ROS-triggered and regenerating mechanism,TPGS was replaced by glycerin monostearate to obtain another ROS-triggereddegradation but non-ROS-regenerating amphiphilicmaterials,named as GBH. GBH synthesis was confirmed by 1HNMR (Fig. S8). Afterloading FRET pair, the particle size of FRET-GBHwas 125± 2.8 nmwith−16.7 ± 2.1 mV zeta potential. Moreover, as showed in Fig. S9, FRET-GBH was also capable of ROS-triggered release. Fluorescence spectraat 484 nm showed that a remarkable fluorescence switch from DiIto DiO occurred after incubation with 1 mM H2O2, suggesting thatFRET-GBH was responsive to ROS and the release of FRET pair was dra-matically accelerated. Further, the effect of GBH on ROS level in MCF-7/ADR cells revealed that ROS level decreased gradually as the concentra-tion of GBH increased, indicating that the degradation of arylboronicester in GBH would consume ROS in cytoplasm (Fig. S10a).
As described above, most of FRET-TBH disassembled after additionalincubation for 2 h. To illustrate the degradation of ROS triggered releasein time course MCF-7/ADR cells were incubated with FRET-TBH and

Fig. 7. Study of ROS-triggered and regenerative mechanism. Intracellular disassemble of FRET-GBH, FRET-TBH, FRET-GBH with TPGS, FRET-GBH with GSH and FRET-TBH with GSH onMCF-7/ADR cells after an additional 0.5, 1 and 2 h incubation observed by CLSM (green for DiO, red for DiI and scale bar = 10 μm).
379Z. Su et al. / Journal of Controlled Release 196 (2014) 370–383
FRET-GBH for 2 h at 37 °C and washed with ice-cold PBS, and subse-quently monitored with CLSM after an additional 0.5, 1 and 2 h of incu-bation. As showed in Fig. 7, both of FRET-TBH and FRET-GBH presenteda time-dependent degradation and a decrease of FRET signal was ob-served inside the cells. The FRET signal in FRET-TBH disappeared morequickly than FRET-GBH and after an additional 0.5 h of incubation FRof FRET-TBH decreased to 0.30, which was 2.3-fold lower than that ofFRET-GBH (FR = 0.69, P b 0.01), suggesting that ROS regeneratingmanner of TBH would facilitate the release of cargo modules intocytoplasm. After an additional 1 h of incubation most of FRET-TBHdisassembled and the FR remained unchanged for additional 1 to 2 h.
To further confirm the intracellular ROS concentration-dependentrelease characteristic, co-incubating FRET-GBH with TPGS (200 μg/mL)to elevate intracellular ROS level could dramatically accelerate releaseof FRET pair from FRET-GBH, which was comparable to that ofFRET-TBH. In contrast, for additional 2 h incubation no remarkable re-lease of FRET pair was observed by inhibiting intracellular ROS levelwith glutathione (GSH, 500 μg/mL), an antioxidant and free radicalsscavenger [37] (Fig. S10b), further demonstrating intracellular ROSconcentration-dependent drug release of ROS-triggered drug deliverynanosystem. Interestingly, as TBH could efficiently induce ROS genera-tion, the inhibitory effect of GSH to FRET-TBH was much weaker thanthat to FRET-GBH. Most of FRET pair release from FRET-TBH for addi-tional 2 h incubation. As it was reported, redox property changes duringcancer development and disease progression [15]. In this study, TBH
could create a kind of ROS positive feedback microenvironment withpersisting high levels of ROS generation necessary for degradationof ROS-triggered nanosystem no matter on reduced or oxidized intra-cellular microenvironment.
3.7. Enhanced accumulation, retention, cytotoxicity and apoptosis of DOXin MCF-7/ADR cells by DOX-TBH
In time-dependent uptake study, owing to the efflux of DOX byP-gp, the uptake index (UI) of free DOX in MCF-7/ADR cells was only0.803 ± 0.085 μg per mg protein after 4 h of incubation. Incubationwith DOX-TBH and DOX-TH for 4 h, the UI in MCF-7/ADR cells signifi-cantly increased, which reached up to 11-fold and 7-fold higher thanthat of free DOX, respectively (Fig. 8a). Except for free DOX, the uptakeof DOX presented concentration-dependent and reached a plateau at20 μg/mL of DOX (Fig. 8b). Comparedwith DOX-TH, DOX-TBH revealedhigher UI, indicating that ROS-triggered DOX-TBH was an effectiveintracellular nanocarrier delivery system for overcoming MDR.
In another experiment, we determined how DOX was retained incells. A fast decline in intracellular DOX level was observed in MCF-7/ADR cells pre-incubated with free DOX as a result of drug efflux byP-gp, only 19.67 ± 4.69% DOX was retained in the cells after 4 h(Fig. 8c). However, the amount and the efflux rate of DOX were signifi-cantly lower when pre-incubated with DOX-TBH and DOX-TH. The re-markable retention effect for DOX-TBH was noticed when compared

Fig. 8. Enhanced accumulation, retention, cytotoxicity and apoptosis of DOX inMCF-7/ADR cells by DOX-TBH. (a) The influence of concentration of DOX on intracellular uptake inMCF-7/ADR cells. (b) The influence of time on intracellular uptake inMCF-7/ADR cells. (c) Retention of DOX inMCF 7/ADR cells after pre-incubation with free DOX, DOX-TBH or DOX-TH for 4 h(The concentration of total DOX in the free DOX pre-incubationwas 40 μg/mL, while it was 10 μg/mL for DOX-TBH and DOX-TH). (d) Cell survival ofMCF-7/ADR cells after chemotherapyfor 24 h. (e) CLSM study ofMCF-7/ADR cells incubatedwith freeDOX,DOX-TBH andDOX-TH (The concentration of DOXwas 10 μg/mL; scale bar=50 μm). (f) Flow cytometric analysis ofMCF-7/ADR cells apoptosis induced by free DOX, DOX-TBH or DOX-TH for 12 h using the Annexin V-FITC/PI staining (The concentration of DOX was 10 μg/mL). ⁎P b 0.05; ⁎⁎P b 0.01.
380 Z. Su et al. / Journal of Controlled Release 196 (2014) 370–383
with DOX-TH (69.72 ± 3.29% vs 46.75 ± 1.79%, P b 0.05) after 4 h ofincubation. As over-expression of the drug efflux transporters is rele-vant to the major type of drug resistance in tumor cells, the propertyto evade and preferably inhibit drug pump efflux activity has beenexpected to overcome MDR [38]. TPGS exhibits concentration-dependent inhibition of efflux pump ATPase without inducingsignificant ATPase activity on its own [39]. As demonstrated above,DOX-TBH in MCF-7/ADR cells could quickly degrade, suggesting thatmore amounts of TPGS would dissociate into cytoplasm, which wouldfunction more quickly and effectively than DOX-TH to hinder DOXefflux. A site-dependent drug release character is also vital for drugmolecules to reach their target, leading to higher intracellular drug con-centration necessary for therapeutic efficacy [40].
Furthermore, qualitative analysis by CLSM indicated that faint redfluorescence signals in cytoplasm region were observed for free DOXafter 4 h incubation (Fig. 8e) due to efflux by P-gp. Similar to the quan-titative analysis strong fluorescence signals were observed in MCF-7/ADR cells when incubated with DOX-TBH and DOX-TH. For DOX-THthe fluorescence signals dispersed in the peri-nuclear region. Itmust be mentioned that DOX-TH did not have the ROS responsiveDOX release characteristic and the insufficient intracellular drugrelease remained rate-limiting steps for reaching the therapeutic drugconcentration level in nuclei. More importantly, with the function ofROS-triggered release and ROS regeneration, the released DOX fromDOX-TBH quickly accumulated into the nuclei, suggesting that en-hanced cytotoxicity would be occurred when comparing to DOX-TH.
Furthermore, to confirm the enhanced cytotoxicity and apoptosis byeffectively releasing DOX from DOX-TBH, MTT assay was first used to
evaluate the cytotoxicity in MCF-7/ADR cells. In the research scopefree DOX did not affect the cell viability. But, treatment of DOX-TBH ex-hibited elevated cytotoxicity when compared with DOX-TH in MCF-7/ADR cells (IC50 = 2.75 ± 0.4 μg/mL for DOX-TBH and IC50 = 10.39 ±1.91 μg/mL for DOX-TH, P b 0.05) (Fig. 8d). This approves thesupremacy of continuous ROS-triggered DOX release over conventionaldrug delivery approaches in resistant cancer cells. Annexin V-FITC/PIdetection was conducted to compare the apoptosis-inducing effect ofDOX-TBH and DOX-TH. According to the mechanisms of AnnexinV-FITC/PI detection, cells under initiation of apoptosis would be stainedby Annexin V-FITC while late apoptotic cells would be labeled by PI. InFig. 8f, both DOX-TBH and DOX-TH had similar total apoptotic ratio(66.8% of DOX-TBH vs 63.2% of DOX-TH). While compose of apoptoticratio revealed that DOX-TBH could induce much more late apoptosisin comparisonwith DOX-TH (50.8% vs 34.1%, P b 0.05), further verifyingthe enhanced cytotoxicity and apoptosis-inducing activity of ROS-triggered and regenerating nanosystem in resistant cancer cells.
3.8. In vivo biodistribution and therapeutic efficacy
To verify the feasibility of DOX-TBH and DOX-TH for cancer therapyin vivo, the biodistribution and antitumor efficacy were estimated inxenograft MCF-7/ADR cells tumor-bearing mice. DiR, a near infrareddye, was loaded into TBH and TH micelles (DiR-TBH and DiR-TH) andthe in vivo biodistribution at specific time points was recorded withNIR imaging system (Fig. 9a). It could be seen from the fluorescenceintensity in tumors after 24 h post-injection that DiR-TBH exhibitedstronger tumor targeting, judging from 5.41-fold higher than DiR-TH

Fig. 9. In vivo tumor-targeting of DiR-TBH and DiR-TH. (a) In vivo fluorescence imaging of the xenograft MCF-7/ADR tumor-bearing nude mice at 1, 4, 8, 12 and 24 h after intravenousinjection of DiR-TBH (I) and DiR-TH (II). (b) Ex vivo fluorescence imaging of tumor and tissues harvested from the xenograft MCF-7/ADR tumor-bearing nudemice at 24 h post-injection.The numeric label for each organ is as follows: 1, tumor; 2, heart; 3, liver; 4, spleen; 5, lung; 6, kidney; 7, pancreas; 8, brain. (c) Semi-quantification of DiR-TBH and DiR-TH in the isolatedtumor and tissues of xenograft MCF-7/ADR tumor-bearing nude mice (n = 3). (d). CLSM images of DiR in tumor tissues in different groups (red = DiR in MCF-7/ADR cells; blue = cellnucleus stained by DAPI; scale bar = 250 μm). ⁎⁎P b 0.01.
381Z. Su et al. / Journal of Controlled Release 196 (2014) 370–383
in tumor site (Fig. 9b and c). Histopathological analysis of tumor tissuesby CLSM also indicated similar results to quantitative analysis (Fig. 9d).TBH and TH possessed the potential for efficiently inducing ROSgeneration in tumor tissue. The amplified drug delivery by TBH couldbe ascribed to the augmented ROS signal which would favor thedegradation of ROS-triggered nanomaterial to inhibit MDR tumor cellsefflux and enhance drug retention. All the in vitro and in vivo resultssuggested the significance of ROS triggered and regenerating mecha-nism of this novel ROS sensitive system.
Moreover, the in vivo biodistribution of DOX solution, DOX-TBH andDOX-TH after intravenous administrationwas also evaluated by quanti-tatively detecting the DOX amounts in different tissues. Both TBH andTH micelles exhibited the same pharmacokinetic behavior and anenhanced blood circulation compared to the free DOX (Fig. 10a),which suggested that higher concentrations of DOX for DOX-TBH andDOX-TH would remain in the systemic circulation for improving thera-peutic index and accumulate in solid tumors through enhanced perme-ability and retention (EPR) effect. In addition, DOX-TBH exhibitedhigher DOX accumulation in the tumor than the DOX solution andDOX-TH. The DOX amount in the tumor tissues delivered by DOX-TBHwas 5.3 and 2.1 fold higher than that of those delivered by the DOXsolution and DOX-TH, respectively, at 4 h post-injection. Accordingto Table S1, the AUC0~ t of DOX-TBH in tumor showed 4.3-fold and2.1-fold higher than that of DOX solution and DOX-TH. Hence, it wasproved that DOX-TBH had higher tumor targeting potential because ofboth passive and active targeting mechanisms.
No evident difference was found in the tumor volumes after succes-sive intravenous administration of free DOX due to MDR. However, theefficiency of DOX-TBH and DOX-TH for cancer therapy showed thatDOX-TBH and DOX-TH significantly reduced the tumor volumes whencompared to the free DOX (Fig. 10b). Moreover, DOX-TBH displayeda prominent effect on tumor-size reduction and delivered the moredistinguishable effect by extending the survival period of the tumorbearing mice when compared to DOX-TH (Fig. 10c), suggesting thatthe efficient intracellular delivery and incessant ROS-triggered andregenerating nanosystemwere of great importance to enhanced antitu-mor activity. No notable change in body weights of the mice during the
treatment was found for DOX-TBH in comparison with the saline group(Fig. 10d). Tumor tissues' histological TUNEL analysis of different treat-ment groups also revealed that no obvious apoptosis was found in tu-mors of control group (apoptosis ratio b 5%) (Fig. 10e and f). However,markedly increased percentage of apoptotic and necrotic tumor cellswas observed in treatment groups especially for DOX-TBH groups;nearly 90% of tumor cells underwent apoptosis or necrosis while the ap-optosis ratio of DOX-TH and DOX was only 50% and 18%, respectively(Fig. 10f). In addition, the histologic images using H&E staining showedthat treatment with DOX-TBH, a massive cancer cell underwent necro-sis in the tumor site (Fig. 10e) while no significant pathological abnor-malities in the heart, liver, spleen, lung and kidney were observed(Fig. S11). These results verified that due to ROS responsiveness andregeneration attribute of DOX-TBH a high therapeutic effect on drugresistant human breast cancer could be expected.
4. Conclusion
In summary, we developed a novel targeted drug delivery nanosys-tem based on TPGS, arylboronic esters and HA for anticancer drugdelivery, which was not only responding to ROS signal for drug releasebut also self-regenerating ROS signal in cytoplasm, simultaneously.Our results suggested that EPR effect and targeting moiety do help theformulations to concentrate into the tumor cells; nonetheless, adequatedrug release in the tumors also has a significant role in the successof therapy. We achieved this goal by developing a ROS-triggeredand regenerating nanosystem that keeps high levels of ROS in tumormicroenvironment, rapidly releasing drug and retaining it effectivelyto overcome the drug resistance of tumor.
Acknowledgments
This work was supported by the National Natural Science Founda-tion of China (No.81273467), the Fundamental Research Funds for theCentral Universities (JKY2011072) and the National Basic ResearchProgram of China (2015CB755500).
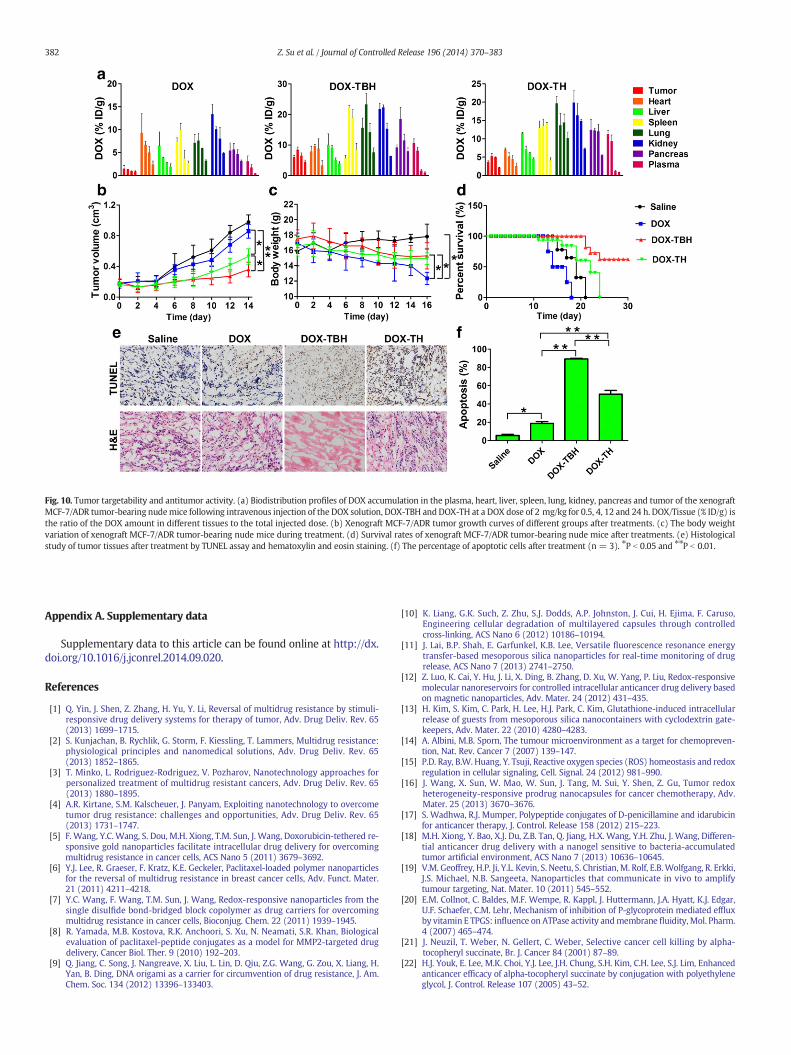
Fig. 10. Tumor targetability and antitumor activity. (a) Biodistribution profiles of DOX accumulation in the plasma, heart, liver, spleen, lung, kidney, pancreas and tumor of the xenograftMCF-7/ADR tumor-bearing nudemice following intravenous injection of the DOX solution, DOX-TBH and DOX-TH at a DOX dose of 2 mg/kg for 0.5, 4, 12 and 24 h. DOX/Tissue (% ID/g) isthe ratio of the DOX amount in different tissues to the total injected dose. (b) Xenograft MCF-7/ADR tumor growth curves of different groups after treatments. (c) The body weightvariation of xenograft MCF-7/ADR tumor-bearing nude mice during treatment. (d) Survival rates of xenograft MCF-7/ADR tumor-bearing nude mice after treatments. (e) Histologicalstudy of tumor tissues after treatment by TUNEL assay and hematoxylin and eosin staining. (f) The percentage of apoptotic cells after treatment (n = 3). ⁎P b 0.05 and ⁎⁎P b 0.01.
382 Z. Su et al. / Journal of Controlled Release 196 (2014) 370–383
Appendix A. Supplementary data
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.jconrel.2014.09.020.
References
[1] Q. Yin, J. Shen, Z. Zhang, H. Yu, Y. Li, Reversal of multidrug resistance by stimuli-responsive drug delivery systems for therapy of tumor, Adv. Drug Deliv. Rev. 65(2013) 1699–1715.
[2] S. Kunjachan, B. Rychlik, G. Storm, F. Kiessling, T. Lammers, Multidrug resistance:physiological principles and nanomedical solutions, Adv. Drug Deliv. Rev. 65(2013) 1852–1865.
[3] T. Minko, L. Rodriguez-Rodriguez, V. Pozharov, Nanotechnology approaches forpersonalized treatment of multidrug resistant cancers, Adv. Drug Deliv. Rev. 65(2013) 1880–1895.
[4] A.R. Kirtane, S.M. Kalscheuer, J. Panyam, Exploiting nanotechnology to overcometumor drug resistance: challenges and opportunities, Adv. Drug Deliv. Rev. 65(2013) 1731–1747.
[5] F. Wang, Y.C. Wang, S. Dou, M.H. Xiong, T.M. Sun, J. Wang, Doxorubicin-tethered re-sponsive gold nanoparticles facilitate intracellular drug delivery for overcomingmultidrug resistance in cancer cells, ACS Nano 5 (2011) 3679–3692.
[6] Y.J. Lee, R. Graeser, F. Kratz, K.E. Geckeler, Paclitaxel-loaded polymer nanoparticlesfor the reversal of multidrug resistance in breast cancer cells, Adv. Funct. Mater.21 (2011) 4211–4218.
[7] Y.C. Wang, F. Wang, T.M. Sun, J. Wang, Redox-responsive nanoparticles from thesingle disulfide bond-bridged block copolymer as drug carriers for overcomingmultidrug resistance in cancer cells, Bioconjug. Chem. 22 (2011) 1939–1945.
[8] R. Yamada, M.B. Kostova, R.K. Anchoori, S. Xu, N. Neamati, S.R. Khan, Biologicalevaluation of paclitaxel-peptide conjugates as a model for MMP2-targeted drugdelivery, Cancer Biol. Ther. 9 (2010) 192–203.
[9] Q. Jiang, C. Song, J. Nangreave, X. Liu, L. Lin, D. Qiu, Z.G. Wang, G. Zou, X. Liang, H.Yan, B. Ding, DNA origami as a carrier for circumvention of drug resistance, J. Am.Chem. Soc. 134 (2012) 13396–133403.
[10] K. Liang, G.K. Such, Z. Zhu, S.J. Dodds, A.P. Johnston, J. Cui, H. Ejima, F. Caruso,Engineering cellular degradation of multilayered capsules through controlledcross-linking, ACS Nano 6 (2012) 10186–10194.
[11] J. Lai, B.P. Shah, E. Garfunkel, K.B. Lee, Versatile fluorescence resonance energytransfer-based mesoporous silica nanoparticles for real-time monitoring of drugrelease, ACS Nano 7 (2013) 2741–2750.
[12] Z. Luo, K. Cai, Y. Hu, J. Li, X. Ding, B. Zhang, D. Xu, W. Yang, P. Liu, Redox-responsivemolecular nanoreservoirs for controlled intracellular anticancer drug delivery basedon magnetic nanoparticles, Adv. Mater. 24 (2012) 431–435.
[13] H. Kim, S. Kim, C. Park, H. Lee, H.J. Park, C. Kim, Glutathione-induced intracellularrelease of guests from mesoporous silica nanocontainers with cyclodextrin gate-keepers, Adv. Mater. 22 (2010) 4280–4283.
[14] A. Albini, M.B. Sporn, The tumour microenvironment as a target for chemopreven-tion, Nat. Rev. Cancer 7 (2007) 139–147.
[15] P.D. Ray, B.W. Huang, Y. Tsuji, Reactive oxygen species (ROS) homeostasis and redoxregulation in cellular signaling, Cell. Signal. 24 (2012) 981–990.
[16] J. Wang, X. Sun, W. Mao, W. Sun, J. Tang, M. Sui, Y. Shen, Z. Gu, Tumor redoxheterogeneity-responsive prodrug nanocapsules for cancer chemotherapy, Adv.Mater. 25 (2013) 3670–3676.
[17] S. Wadhwa, R.J. Mumper, Polypeptide conjugates of D-penicillamine and idarubicinfor anticancer therapy, J. Control. Release 158 (2012) 215–223.
[18] M.H. Xiong, Y. Bao, X.J. Du, Z.B. Tan, Q. Jiang, H.X. Wang, Y.H. Zhu, J. Wang, Differen-tial anticancer drug delivery with a nanogel sensitive to bacteria-accumulatedtumor artificial environment, ACS Nano 7 (2013) 10636–10645.
[19] V.M. Geoffrey, H.P. Ji, Y.L. Kevin, S. Neetu, S. Christian,M. Rolf, E.B.Wolfgang, R. Erkki,J.S. Michael, N.B. Sangeeta, Nanoparticles that communicate in vivo to amplifytumour targeting, Nat. Mater. 10 (2011) 545–552.
[20] E.M. Collnot, C. Baldes, M.F. Wempe, R. Kappl, J. Huttermann, J.A. Hyatt, K.J. Edgar,U.F. Schaefer, C.M. Lehr, Mechanism of inhibition of P-glycoprotein mediated effluxby vitamin E TPGS: influence on ATPase activity andmembrane fluidity,Mol. Pharm.4 (2007) 465–474.
[21] J. Neuzil, T. Weber, N. Gellert, C. Weber, Selective cancer cell killing by alpha-tocopheryl succinate, Br. J. Cancer 84 (2001) 87–89.
[22] H.J. Youk, E. Lee, M.K. Choi, Y.J. Lee, J.H. Chung, S.H. Kim, C.H. Lee, S.J. Lim, Enhancedanticancer efficacy of alpha-tocopheryl succinate by conjugation with polyethyleneglycol, J. Control. Release 107 (2005) 43–52.

383Z. Su et al. / Journal of Controlled Release 196 (2014) 370–383
[23] L.F. Dong, P. Low, J.C. Dyason, X.F. Wang, L. Prochazka, P.K. Witting, R. Freeman, E.Swettenham, K. Valis, J. Liu, R. Zobalova, J. Turanek, D.R. Spitz, F.E. Domann, I.E.Scheffler, S.J. Ralph, J. Neuzil, Alpha-tocopheryl succinate induces apoptosis bytargeting ubiquinone-binding sites in mitochondrial respiratory complex II, Onco-gene 27 (2008) 4324–4335.
[24] A.R. Lippert, G.C. Van de Bittner, C.J. Chang, Boronate oxidation as a bioorthogonalreaction approach for studying the chemistry of hydrogen peroxide in living sys-tems, Acc. Chem. Res. 44 (2011) 793–804.
[25] J.A. Burdick, G.D. Prestwich, Hyaluronic acid hydrogels for biomedical applications,Adv. Mater. 23 (2011) 41–56.
[26] M. Han, Q. Lv, X.J. Tang, Y.L. Hu, D.H. Xu, F.Z. Li, W.Q. Liang, J.Q. Gao, Overcomingdrug resistance of MCF-7/ADR cells by altering intracellular distribution of doxoru-bicin via MVP knockdown with a novel siRNA polyamidoamine–hyaluronic acidcomplex, J. Control. Release 163 (2012) 136–144.
[27] M.T. Peracchia, C. Vauthier, C. Passirani, P. Couvreur, D. Labarre, Complement con-sumption by poly(ethylene glycol) in different conformations chemically coupledto poly(isobutyl 2-cyanoacrylate) nanoparticles, Life Sci. 61 (1997) 749–761.
[28] C.A. Gay, J.M. Gebicki, Perchloric acid enhances sensitivity and reproducibility of theferric-xylenol orange peroxide assay, Anal. Biochem. 304 (2002) 42–46.
[29] H. Chen, S. Kim, L. Li, S. Wang, K. Park, J.X. Cheng, Release of hydrophobic moleculesfrom polymer micelles into cell membranes revealed by Forster resonance energytransfer imaging, Proc. Natl. Acad. Sci. U. S. A. 105 (2008) 6596–6601.
[30] S. Zhao, W. Dai, B. He, J. Wang, Z. He, X. Zhang, Q. Zhang, Monitoring the transport ofpolymeric micelles across MDCK cell monolayer and exploring related mechanisms,J. Control. Release 158 (2012) 413–423.
[31] Y. Xu, X.F. Jin, Q.N. Ping, J. Cheng, M.J. Sun, F. Cao, W.L. You, D.F. Yuan, A novellipoprotein-mimic nanocarrier composed of the modified protein and lipid fortumor cell targeting delivery, J. Control. Release 146 (2010) 299–308.
[32] K.E. Broaders, S. Grandhe, J.M. Frechet, A biocompatible oxidation-triggered carrierpolymer with potential in therapeutics, J. Am. Chem. Soc. 133 (2011) 756–758.
[33] Y. Cui, Y. Zhao, Y. Tian, W. Zhang, X. Lu, X. Jiang, The molecular mechanism of actionof bactericidal gold nanoparticles on Escherichia coli, Biomaterials 33 (2012)2327–2333.
[34] J.H. Park, H.J. Cho, H.Y. Yoon, I.S. Yoon, S.H. Ko, J.S. Shim, J.H. Cho, K. Kim, I.C. Kwon,D.D. Kim, Hyaluronic acid derivative-coated nanohybrid liposomes for cancer imag-ing and drug delivery, J. Control. Release 174 (2014) 98–108.
[35] S. Harnisch, R.H. Muller, Adsorption kinetics of plasma proteins on oil-in-wateremulsions for parenteral nutrition, Eur. J. Pharm. Biopharm. 49 (2000) 41–46.
[36] Z.G. Su, J.X. Niu, Y.Y. Xiao, Q.N. Ping, M.J. Sun, A.W. Huang, W.L. You, X.Y. Sang, D.F.Yuan, Effect of octreotide-polyethylene glycol(100) monostearate modification onthe pharmacokinetics and cellular uptake of nanostructured lipid carrier loadedwith hydroxycamptothecine, Mol. Pharm. 8 (2011) 1641–1651.
[37] R. Cascella, E. Evangelisti, M. Zampagni, M. Becatti, G. D'Adamio, A. Goti, G. Liguri, C.Fiorillo, C. Cecchi, S-linolenoyl glutathione intake extends life-span and stress resis-tance via Sir-2.1 upregulation in Caenorhabditis elegans, Free Radic. Biol. Med. 73C(2014) 127–135.
[38] B. Al-Lazikani, U. Banerji, P. Workman, Combinatorial drug therapy for cancer in thepost-genomic era, Nat. Biotechnol. 30 (2012) 679–692.
[39] E.M. Collnot, C. Baldes, U.F. Schaefer, K.J. Edgar, M.F. Wempe, C.M. Lehr, Vitamin ETPGS P-glycoprotein inhibition mechanism: influence on conformational flexibility,intracellular ATP levels, and role of time and site of access, Mol. Pharm. 7 (2010)642–651.
[40] S. Febvay, D.M. Marini, A.M. Belcher, D.E. Clapham, Targeted cytosolic delivery ofcell-impermeable compounds by nanoparticle-mediated, light-triggered endosomedisruption, Nano Lett. 10 (2010) 2211–2219.


















