
INTEGRATION EFFICIENCY IN DNA-INDUCED TRANSFORMATION … · INTEGRATION EFFICIENCY IN DNA-INDUCED...
14
INTEGRATION EFFICIENCY IN DNA-INDUCED TRANSFORMATION OF PNEUMOCOCCUS. 11. GENETIC STUDIES OF MUTANT INTEGRATING ALL THE MARKERS WITH A HIGH EFFICIENCY GRRARD TIRABY AND MICHEL ARMAND SICARD Laboratoire de Ginitique, Universitk Paul Sabatier, Toulouse, France Manuscript received March 19, 1973 Revised copy received May 23, 1973 ABSTRACT Transformation of the pneumococcus mutant 402 by DNA's bearing the standard reference marker and several other markers belonging to two un- linked loci has shown that differences in the integration efficiencies of these markers were considerably reduced in this strain compared to the wild-type strain CZ,. The sensitivities of mutant 401 to ultraviolet light and to X-ray irradiation are the same as those of CZ,. However, in 402 all the markers tested are more resistant to inactivation as shown by transformation of 402 and Cl, by ultraviolet-irradiated DNA. The increase in resistance is greater for low efficiency (LE) markers than for high efficiency (HE) markers.-The decreased discrimination between LE and HE markers in strain 402 is not due to a mechanism related to modification of markers in the transforming DNA by the recipient cells, nor are the proteins inducing competence of the cells responsible for the differences in the integration efficiencies of various markers. -Genetic studies of the fate of recombinants as well as the measure of the amount of DNA taken up have shown that all the markers are integrated in strain 402 by the same recombination process, that specific to high efficiency markers. HE nature of the events occurring during genetic recombination in bacteria Thas not yet been completely elucidated. With the isolation of recombination- deficient mutants of E. coli, it appears that in this organism, the recombination process involves several pathways (CLARK 1971). A new pathway of recombi- nation might explain what seems to be a unique characteristic of Diplococcus pneumoniae among transformable bacteria: the marker effect observed during transformation. Genetic studies in pneumococcus have shown that some single- site markers are less efficient than others in giving transformants (EPHRUSSI- TAYLOR, SICARD and KAMEN 1965; SICARD and EPHRUSSI-TAYLOR 1965; LACKS 1966; SIROTNAK and HACHTEL 1969). According to their integration efficiency, auxotrophic or antibiotic resistance markers fall into at least two classes: one with high efficiency (HE) and one with low efficiency (LE). Mutants of Diplo- COCCUS prreumoniae that yield the same number of transformants for markers of both classes have been isolated fortuitously (RAVIN 1959; LITMAN 1961) or intentionally (LACKS 1970). We have also isolated a similar mutant and have Genetics 75: 35-48 September, 1973
Transcript of INTEGRATION EFFICIENCY IN DNA-INDUCED TRANSFORMATION … · INTEGRATION EFFICIENCY IN DNA-INDUCED...

INTEGRATION EFFICIENCY IN DNA-INDUCED TRANSFORMATION OF PNEUMOCOCCUS. 11. GENETIC STUDIES OF MUTANT
INTEGRATING ALL THE MARKERS WITH A HIGH EFFICIENCY
GRRARD TIRABY AND MICHEL ARMAND SICARD
Laboratoire de Ginitique, Universitk Paul Sabatier, Toulouse, France
Manuscript received March 19, 1973 Revised copy received May 23, 1973
ABSTRACT
Transformation of the pneumococcus mutant 402 by DNA's bearing the standard reference marker and several other markers belonging to two un- linked loci has shown that differences in the integration efficiencies of these markers were considerably reduced in this strain compared to the wild-type strain CZ,. The sensitivities of mutant 401 to ultraviolet light and to X-ray irradiation are the same as those of CZ,. However, in 402 all the markers tested are more resistant to inactivation as shown by transformation of 402 and Cl, by ultraviolet-irradiated DNA. The increase in resistance is greater for low efficiency (LE) markers than for high efficiency (HE) markers.-The decreased discrimination between LE and HE markers in strain 402 is not due to a mechanism related to modification of markers in the transforming DNA by the recipient cells, nor are the proteins inducing competence of the cells responsible for the differences in the integration efficiencies of various markers. -Genetic studies of the fate of recombinants as well as the measure of the amount of DNA taken up have shown that all the markers are integrated in strain 402 by the same recombination process, that specific to high efficiency markers.
HE nature of the events occurring during genetic recombination in bacteria Thas not yet been completely elucidated. With the isolation of recombination- deficient mutants of E. coli, it appears that in this organism, the recombination process involves several pathways (CLARK 1971). A new pathway of recombi- nation might explain what seems to be a unique characteristic of Diplococcus pneumoniae among transformable bacteria: the marker effect observed during transformation. Genetic studies in pneumococcus have shown that some single- site markers are less efficient than others in giving transformants (EPHRUSSI- TAYLOR, SICARD and KAMEN 1965; SICARD and EPHRUSSI-TAYLOR 1965; LACKS 1966; SIROTNAK and HACHTEL 1969). According to their integration efficiency, auxotrophic or antibiotic resistance markers fall into at least two classes: one with high efficiency (HE) and one with low efficiency (LE). Mutants of Diplo- COCCUS prreumoniae that yield the same number of transformants for markers of both classes have been isolated fortuitously (RAVIN 1959; LITMAN 1961) or intentionally (LACKS 1970). We have also isolated a similar mutant and have Genetics 75: 35-48 September, 1973

36 G . TIRABY A N D M. A. SICARD
introduced the mutation which confers reduction in the differences of integration efficiencies of various markers into the wild-type strain (TIRABY, CLAVERYS and SICARD 1973).
This report deals with the comparative properties of the non-discriminating strain and the wild-type strain. Evidence is presented suggesting that in the mutant, the product of integration of all markers is that characteristic of high efficiency markers.
MATERIALS A N D METHODS
Strains: The standard wild-type strain of DipZococcus pneumoniae referred to as "clone 3" (CZ,) was used as recipient in the transformation experiments. Drug-resistant strains were con- structed by introducing the appropriate markers by transformation into C1,. In addition to strain 69 (TIRBAY, CLAVERYS and SICARD 1973) strain 39 was used for the donor DNA. This latter strain bears the markers: str-r41 (HE) scored at 200 pg streptomycin/ml; opt-r2 (LE) scored at 5 pg optochin/ml; Ami A-rl (HE) scored at 2 x 10-5 M aminopterin/ml; ery-r2 and ery-r6, two double mutations conferring resistance to erythromycin. The double mutant ery-r2, ery-r6 resists up to 4 pg/ml of erythromycin, the single ery-r2 resists up to 1 ug/ml and the single ery-rb resists up to 0.1 pg/ml.
Media, transformation procedure and uptake experiments have been described in the preced- ing paper (TIRABY, CLAVERYS and SICARD 1973).
Isolation of the 401 strain: Using nitrosoguanidine as a mutagenic agent we have isolated one clone transforming the LE marker o p t 4 with half the efficiency of the HE marker str-r42. Since it is likely that under our experimental conditions several mutations have been induced per cell, the new character has been transferred into our standard wild-type strain, Cl,, by transformation. Eesides the integration efficiency, no other difference can be detected between the mutant and the wild type, including generation time, at 25" or 37". To screen for this character we have taken advantage of the ability of pneumococcus to give rise to transformed bacteria on solid medium. Wild-type cells were transformed by DNA extracted from the mutant strain and clones show- ing the same efficiency of transformation for HE and LE markers were selected by the method described in the preceding paper. Only one clone, named 401, isogenic to CZ, except for the selected mutation, was retained. The frequency of transmission of this mutation from the original strain to the wild type suggests that only one gene is involved in the 402 character.
Ultraviolet irradiation of DNA: DNA was isolated from cells of strain 69, as described else- where (MARTIN and EPHRUSSI-TAYLOR 1964). Samples of this DNA at the concentration of 20 pg/ml were placed in petri dishes of 5 cm diameter and irradiated with stirring under a 15 W General Electric germicidal lamp at a dose rate of 14 ergs/mm2/sec. After exposure to ultraviolet light the DNA solutions were kept at 4" in screw cap tubes.
Irradiation of the bacteria LJV irradiation of the cells was done as described in the first paper of this series (TIRABY, CLAVERYS and SICARD 1973).
The source of ionizing irradiation was a Secasi X-ray machine operated at a 100 kv peak and 30 ma. The dose rate was not measured. The cells t o be irradiated were grown until they reached a density corresponding to 5 x 107 colony-forming units/ml. Two-ml aliquots were put into covered plastic petri dishes and exposed with stirring to X-rays. After exposure and before plating, to determine the fractions of surviving colonies, the cells were held in an ice bath to minimize DNA repair.
Preparation of DNA-bacterial complexes: Highly competent cells were treated at 37" for 15 minutes with a saturating concentration of transforming DNA bearing appropriate markers (SICARD 1964). DNA uptake was arrested by the addition of 5 pg/ml of crystalline pancreatic DNase, followed by 2 minutes more incubation at 37". The culture was then chilled, glycerol added to I O % , and distributed in 2 m l aliquots into small tubes. A small sample was reserved for assay of transformation and the remaining aliquots stored at 70".

MARKER DISCRIMINATION IN TRANSFORMATION 37
Phenotypic expression timc and segregation pattern of transformants: 1) A sample of the DNA-bacterial complexes was quickly thawed, diluted tenfold into pre-warmed medium and incubated at 37". Small samples were removed from the dilute culture at 10-minute intervals during a total incubation period of 120 minutes and chilled on ice. Then the bacteria, after appropriate dilution increasing with the time of incubation of each sample, were plated in selective medium. The plates were further incubated for 24 hours. The maximum number of transformants was determined by plating bacteria from the thawed DNA-bacterial complexes in non-selective medium and scoring transformants by the overlay method after 150 minutes' incubation at 37".
2) Another sample of the same DNA-bacterial complexes was quickly thawed and diluted a hundred-fold into fresh medium at 37". Small samples were withdrawn from the dilute culture a t 20 or 30 minute intervals. Then three plates were made from each sample and incubated at 37" immediately after the medium hardened. All the plates were overlayered at the same time with a nutrient agar containing the appropriate antibiotic, 3 hours after the first plates were placed in the incubator. The plates were incubated for another 24 hours to allow for the appear- ance of visible colonies.
RESULTS
Integration Eficiency of Various Markers The property of strain 401 to transform one LE and one HE marker with the
same efficiency was extended to other markers. Tm70 markers belonging to the same locus, one HE (ami A d ) and the other LE (ami A-rP), displayed the same integration efficiency. The integration efficicncy of the opt-r2 marker was never found to be as high as the reference str-r41 marker (Table 1). Other LE markers of spontaneous origin, conferring resistance to 5 yug of optochin, were isolated. Nine different markers tested gave the characteristic integration efficiency of the opt r-2 mutation. Likewise the integration efficiency of 15 LE ami A markers was found to be 0.80 with no more than 10% variation from this value.
Radiation Sensitiuity
Strain 401 has the same sensitivity to UV light as the wild type (Figure 1) . It seems, therefore, that the modification introduced by the mutation 401 on the
TABLE 1
Relatiue efficiencies of integration of various markers in strains 401 and C1,
Markers str-r41 ami A-rl ami A-r9 o p t 4 ery-rb ery-r2
Recipient strains (HE) (HE) (LE) (LE) (HE) (HE)
c1, 1.00 1.10 0.15 0.20 0.60 0.65 1.00 1.22 0.13 0.16 0.60 0.60
401 1 .oo 0.83 0.80 0.45 0.90 0.90 1 .oo 0.85 0.82 0.48 0.95 1 .oo 1 .oo 0.80 0.85 0.50 0.95 0.90
The integration efficiency of a given marker is determined by the ratio of the number of trans- formants for this marker to the number of transformants for the reference marker conferring resistance to streptomycin (str-r41). Strains CZ, and 401 are transformed by DNA from strain 69 and by DNA bearing both markers str-r41 and ery-r2.

38 G. TIRABY AND M. A. SICARD
ULTRAVIOLET DOSE (ergs mm-')
FIGURE 1.-The fraction of colony-forming units surviving as a function of the ultraviolet radiation dose.
The wild-type strain, C1, (O), and the mutant that gives only high integration efficiency, 401 ( A ) , an ultraviolet-sensitive mutant, 402 (e) , and a mutant ultraviolet-sensitive that gives only high integration efficiency, 406 (A), were tested for survival after ultraviolet irradiation as described.
efficiency of tradormation has no effect on sensitivity to radiation. In order to uncover a possible interaction between these properties a gene conferring sensi- tivity to UV was transferred into CZ, (strain 402) and 401 (strain 406). Doses sufficient to reduce survival to 37% were 155 ergs/mm2 for C1, and 401 and 10 ergs/mm2 for 402 and 406. Thus sensitivity to UV light with respect to viability appears to be independent of the effect on transformation efficiency controlled by gene 401.
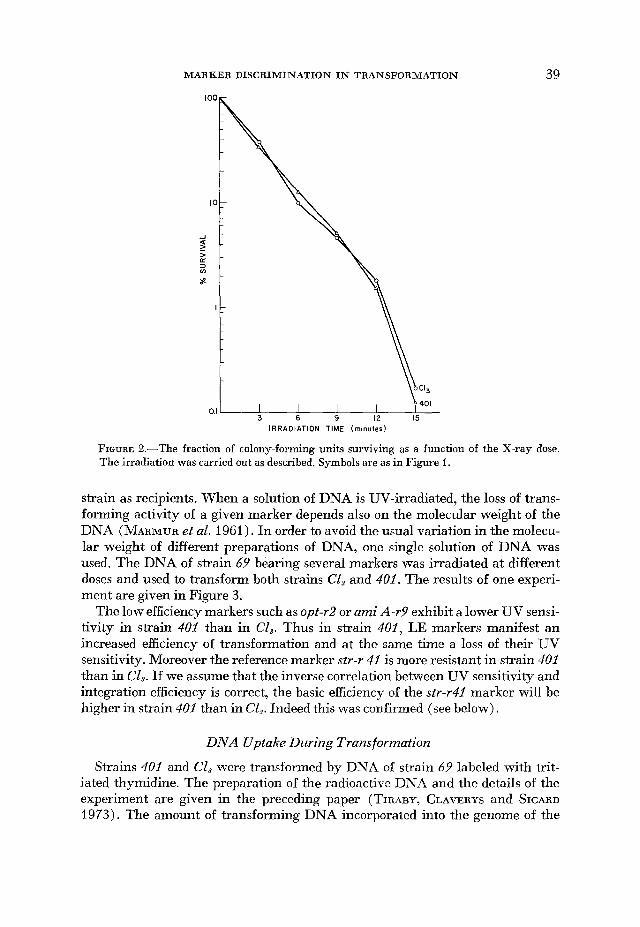
X-ray sensitivity of 401 was compared to the sensitivity of the wild-type strain. It was identical for both strains (Pigure 2).
Genetic markers of transforming DNA are inactivated by UV irradiation. EPHRUSSI-TAYLOR, SICARD and KAMEN (1965) observed that the sensitivity to UV is higher for LE markers than for HE markers belonging to the ami A locus. This inverse carrelation between UV sensitivity afid integration efficiency was confirmed for the different efficiencies of mal- markers (LACKS 1965), and of methicillin resistance markers (HOTCHKISS and GABOR 1971). We thought it of interest to compare the sensitivity of various markers using 401 and wild-type
MARKER DISCRIMINATION IN TRANSFORMATION 39
I I I I \401 0.1
3 6 9 12 15 IRRADIATION TIME (minutes)
FIGURE 2.-The fraction of colony-forming units surviving as a function of the X-ray dose. The irradiation was carried out as described. Symbols are as in Figure 1.
strain as recipients. When a solution of DNA is UV-irradiated, the loss of trans- forming activity of a given marker depends also on the molecular weight of the DNA ( MARMUR et al. 1961 ) . In order to avoid the usual variation in the molecu- lar weight of different preparations of DNA, one single solution of DNA was used. The DNA of strain 69 bearing several markers was irradiated at different doses and used to transform both strains CZ, and 401. The results of one experi- ment are given in Figure 3.
The low efficiency markers such as opt-1-2 or ami A-r9 exhibit a lower UV sensi- tivity in strain 401 than in CZ,. Thus in strain 401, LE markers manifest an increased efficiency of transformation and at the same time a loss of their UV sensitivity. Moreover the reference marker str-r 41 is more resistant in strain 401 than in CZ,. If we assume that the inverse correlation between UV sensitivity and integration efficiency is correct, the basic efficiency of the str-r41 marker will be higher in strain 401 than in CZ,. Indeed this was confirmed (see below).
DNA Uptake During Transformation
Strains 401 and CZ, were transformed by DNA G€ strain 69 labeled with trit- iated thymidine. The preparation of the radioactive DNA and the details of the experiment are given in the preceding paper (TIRABY, CLAVERYS and SICARD 1973). The amount of transforming DNA incorporated into the genome of the

40 G. TIRABY A N D M. A. SICARD
I I I I I I
550 1700 2550 3400 4250 ULTRAVIOLET DOSE (ergs mm-?
FIGURE 3.-Inactivation of markers str-r41, opt-r2, ami A-r9 after ultraviolet irradiation of
The recipient strains are C1, (filled symbols) and 401 (open symbols). Markers: str-r4l the DNA in solution.
(circles), Ami A-r9 (triangles), Opt-r.2 (squares).
TABLE 2
DNA uptake and transformation efficiency in strains C1, (wild type) and 401 (recombination-modified)
~
Recipient strains
~~
( 2 ) Number of streptomycin
(1) DNA
g/ml cpm/ml of culture X 104 sir-r/cpm concentration transformants/ml
~
c4 0.25 0.25 0.05
~
14930 10370 2510
313 243 1 .o 57
401 0.25 13120 4.50 0.25 8750 335 1.5 0.05 2530 83.4
The values of the str-r/cpm are the averages of the values obtained with the three different experiments reported in this table. A new preparation of a competent culture was used in each case.

MARKER DISCRIMINATION IN TRANSFORMATION 41
recipient bacteria was determined by the ratio of the number of transformants str-r41/ml to the number of cpm/ml.
The basic transformation efficiency for the reference marker str-r41 increased by a factor of 1.5 to 1.6 in 401 as compared to the wild-type strain (Table 2). This increase corresponds to the difference existing between the efficiencies of HE markers and markers of very high efficiencies as found by LACKS (1966) and by SIROTNAK and HACHTEL (1969). Mutants in the ami A locus with very high efficiency were also recently isolated in our laboratory (TIRABY and SICARD, in preparation). Thus the same fraction of HE transforinants is probably lost when CZ, is used as recipient. Since we have shown that LE markers are integrated in strain 401 almost as well as HE markers, the basic integration efficiency is appar- ently increased for both types of markers in this strain.
Absence of Modification of the Markers
The increase in the efficiencies of the LE markers in 401 could result from a modification of these markers in such a way that the synapsis between the trans- forming and the recipient strands becomes permissive in this strain. An attempt was made to determine whether LE markers behave like HE markers after their integration into strain 401 and reextraction. For this purpose, strain 401 was transformed by the DNA of strain 69 bearing the mutations opt-r2, ami A-r9, str-r41. Five colonies for each opt-r, str-r, and ami-), str-r double transformants were picked up. The DNA of these ten colonies was prepared and used to trans- €orm CZ,. The characteristic efficiencies of all the LE markers exhibit the initial low value. Thus the LE markers were not modified after transfer through strain 401.
Absence of Extracellular Factors to Account for Marker efficiencies
OTTOLENGHI and HOTCHKISS (1962) observed an increase in the amount of protein in the medium at the competence peak. Besides the competence factor, other proteins are synthesized by competent cells (TOMASZ 1970). TOMASZ and ZANATI (1971) showed that one of these protein factors induces the agglutination of the competent cells in the medium. Experiments were undertaken in an attempt to detect the presence of a protein either in the medium or on the surface of the cells that could account for the difference in transformation efficiency of LE markers in strains Cl, and 401. Competence of a culture of strain 401 bearing an erythromycin-resistance marker (ery-r) (3 x lo6 c.f.u./ml) was induced by competent cells from strain Cl, (3 x I O 7 c.f.u./ml), and on the other hand, the competence of a culture of strain CZ, bearing the same erythromycin marker was induced by competent cells from strain 401. Aiter transformation by DNA bearing the mutations ami A-r28 (HE), opt-r2 (LE) and str-r41, the efficiencies of these markers among the 401 and Cl, cells of the two mixtures were deter- mined. The marker conferring resistance to erythromycin was used to select 401 bacteria among the CZ, bacteria or vice versa. Under these conditions, each of the two strains retained its own properties as far as the efficiencies of the markers are

42 G. TIRABY AND M. A. SICARD
TABLE 3
Integration efieiency of opt-r2 in a C1, culture made competent by 401 cells and in a 4Q1 culture made competent by c1, cells
Number of transformants In minority
Total ( e r y r ) population Ratio Ratio str-r opt-r str-r,eryr opt-r,ery-r opt-r,ery-r/
Recipient cells x 104 x 104 X 1P x 102 opt-r/str-r str-r,ery-r
401 ery-r competent C1, added 93 17.6 100 56 0.19 0.56 401 ery-r 0 0
Cl , ery-r competent 401 added 180 108 210 37.8 0.60 0.18
- -
- - Cl, ery-r 0 0
A fresh culture of 401 ery-r or CZ, ery-r (3.106 colony-forming units/&) are mixed to an equal volume of highly competent cells of CZ, or 401 (3.107 colony-forming units/ml). These mixed cultures are treated with DNA bearing str-r41, opt-r2, ami A-r28 markers, for 30 min. The trans- formants are then scored by the overlay method. As control for the level of competence without induction, cultures of Cl, ery-r or 401 ery-r are mixed with an equal volume of warmed compe- tence medium. DNA is added for 30 min.
concerned (Table 3 ) . The difference in the transformation efficiency of the two classes of markers is therefore not subject to an extracellular factor present in the medium during competence.
Molde of integration of Markers
From measurement of the uptake of radioactive DNA we have shown that the integration efficiency of str-r$l in strain 401 is as high as that of the very high efficiency markers ( W E ) in the wild-type strain. GRAY and EPHRUSSI-TAYLOR (1967) have postulated that the efficiencies of VHE and LE markers could be accounted for by invoking specific excision and repair events. According to the nature of the mismatched bases occurring in the heteroduplex region between the transforming and the recipient strands, the postulated correcting mechanism preferentially excises either ( 1 ) the recipient strand (for VHE markers) or (2) the donor strand (for LE markers) or (3) neither (for HE markers). The possi- bility of a difference in recognitim of various mismatched bases in the discrimi- natir?g strain encouraged us to investigate the mode of integration of markers in strain 401.
1 ) Kinetics of division of the transformed bacteria
In the discriminating strain (CZs), bacteria transformed for an LE marke-. begin to increase in number about one generation sooner than bacteria trans- formed for an HE marker (LOUARN 1970). This difference provides evidence for the proposal that the introduction of an LE marker results in the alteration of both recipient strands while the introduction of an HE marker results in the alteration of only one of the strands of the recipient g- .nome.
M A R K E R D I S C R I M I N A T I O N I N T R A N S F O R M A T I O N 43
I O r-
20 40 60 80 100 120
20 40 60 80 100 120 I50 INCUBATION TIME (minutes)
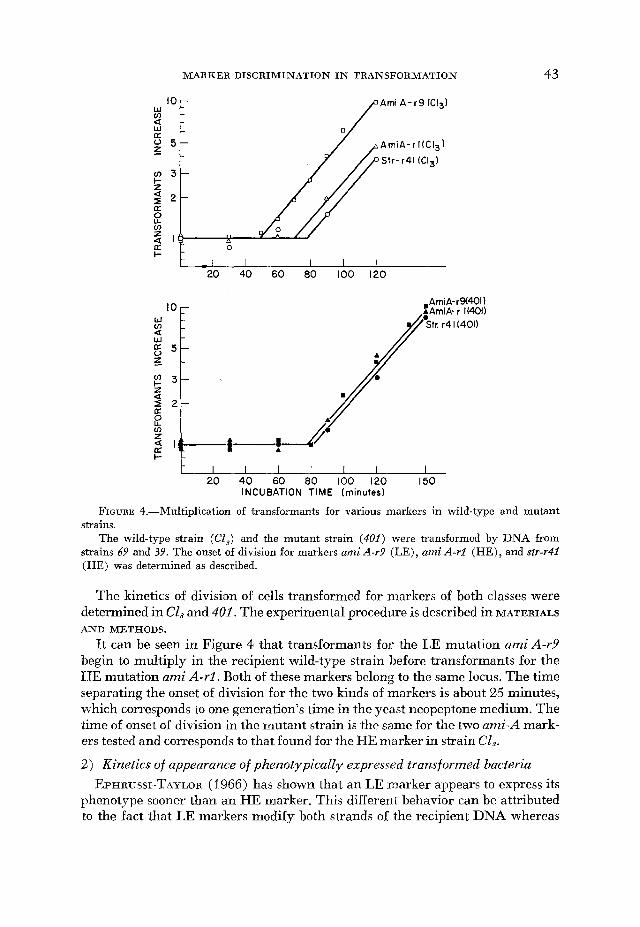
FIGURE 4.-Multiplication of transformants for various markers in wild-type and mutant strains.
The wild-type strain (CL,) and the mutant strain (401) were transformed by DNA from strains 69 and 39. The onset of division for markers ami A-r9 (LE), ami A-rl (HE), and str-r41 (HE) was determined as described.
The kinetics of division oE cells transformed for markers of both classes were determined in Cl, and 401. The experimental procedure is described in MATERIALS
It can be seen in Figure 4 that transformants for the LE mutation amiA-r9 begin to multiply in the recipient wild-type strain before transformants for the HE mutation ami A-r1, Both of these markers belong to the same locus. The time separating the onset of division for the two kinds of markers is about 25 minutes, which corresponds to one generation’s time in the yeast neopeptone medium. The time of onset of division in the mutant strain is the same for the two ami-A mark- ers tested and corresponds to that lound for the HE marker in strain Cl,.
2 ) Kinetics of appearance of phenotypically expresved transformed bacteria EPHRUSSI-TAYLOR (1966) has shown that an LE marker appears to express its
phenotype sooner than an HE marker. This different behavior can be attributed to the fact that LE markers modify both strands of the recipient DNA whereas
A N D M E T H O D S .

44 G. TIRABY A N D M. A. SICARD
106 -
- E . I n - i- z
IL
a
e f lo5- IL + - !A 0 -
a : - W m -
3 z -
IO4 10 20 30 40 50 60 70 80 90 I O 0
INCUBATION TIME (minutes1
FIGURE 5.-Phenotypic expression of a high efficiency marker in wild-type and mutant strains. The kinetics of appearance of transformants str-rll were determined in CIS (open symbols)
and in 401 (closed symbols). Arrows indicate the number of transformants corresponding to 50% of the maximum number of transformants, determined as described in MATERIALS AND METHODS.
HE markers are integrated in either one of the strands but do not modify the complementary strand (GURNEY and Fox 1968). Sime one strand of the DNA is transcribed into messenger RNA, an additional round of DNA replication is required for all of the HE markers to be expressed, whereas all of the LE markers can be expressed without the prerequisite of a round 01 replication.
The kinetics of appearance of the phenotypically-expressed opt-r2 and str-r41 markers was determined in both strains CE, and 401. The experimental method used was very similar to the one used in the experiments above, except that the transformed bacteria were not allowed to express their phenotypes before being challenged with the selective drug.
The time at which a number corresponding to 50% of the transformants str-r41 is reached is approximately the same for both strains (Figure 5) , whereas the transformants opt-r2 are expressed 15 to 20 min later in strain 401 than in strain CZ, (Figure 6). This result suggests that LE markers are expressed at a later time in strain 401 in the same manner as HE markers in strain CZ,. Hence in the mutant strain, LE and HE markers exhibit the integration properties characteristic of HE markers in a wild-type strain.

M A R K E R D I S C R I M I N A T I O N IN T R A N S F O R M A T I O N
- E . ( 0 -
+ z I n a
F !i? lo5- a - n + - L L - 0 n - w
a I3 z -
m -
45
t
ti INCUBATION TIME (minutes)
FIGURE 6.-Phenotypic expression of a low efficiency marker in wild-type and mutant strains. The kinetics of appearance of transformants opt-r2 were determined in C1, (open symbols)
and in 401 (closed symbols). Arrows indicate the number of transformants corresponding to 50% of the maximum number of transformants, determined as described in MATERIALS AND
METHODS.
3 ) Determination of the frequency of double trarrsfwmanfs for strains 401 and C1,
LOUARN and SICARD (1969) showed that the frequency of doubly-transformed bacteria is characteristic of the mode of integration of the markers. When CI, is used as recipient, the genetic information of HE markers is transmitted into either recipient strand at random, whereas LE markers modify both recipient strands. The frequency of double transformants fo r a pair of LE-LE or HE-LE markers will be higher than that for a pair of HE-HE markers. Thus the value R of the ratio R = (D )< N ) / ( A x B ) will depend on the efficiencies of the selected markers (where N is the number of colony-forming units per ml; D, the number of doubly-transformed bactuia for the independent markers A and B ; A and B, the number of singly-transformed bacteria for these markers). These values were determined for several markers in each of the two strains, CZ, and 401.
The DNA-bacterial complexes (described in MATERIALS AND METHODS) giving an efficiency of trans€ormation of at least 10% of colony-forming units were retained for these experiments. This high frequency of transformation can be reached only if all the bacteria in the culture are competent. The double trans-

46 G. TIRABY A N D M. A. SICARD
TABLE 4
R values for uurious pairs of markers in the transformation of struins C1, and 401
Pairs of markers ami A-r l , str-r4f opt-rZ, str-r41 opt-r2, ami A-rl
Recipient strains (HE-HE) (LE-HE) (LE-HE)
c4 0.28 0.34 0.37
Average 0.33
401 0.29 0.35 0.38 0.38
Average 0.35
0.38 0.45 0.46 0.43
0.30 0.35 0.39 0.40 0.36
0.39 0.45 0.48 0.44
0.29 0.33 0.36 0.38 0.34
The reported values of R corresponding to the ratio DN/AB are computed from three inde- pendent experiments for CL, and four f o r 401. DNA from strain 39 bearing str-r41, opt-r2, ami A-r1 markers is used throughout these experiments.
formants were isolated using the same concentration of antibiotics as for selection o i single transformants. The R values in the three different experiments with cl, and the four experiments with 401 are given in Table 4. In strain 401, the values obtained for the three pairs of markers under consideration were identical with one another as well as with the value found for the pair of HE-HE markers: ami A-rl , str-r41 in strain CZ,. By this criterion all markers appear to be trans- formed in strain 401 by the same recombination mechanism as high efficiency markers in strain Cl,.
Thus, the experimental results on the whole rule out the hypothesis of the intervention of a correcting mechanism for VHE markers and are in favor of a single mode of integration of markers in strain 401. The product of integration of markers in mutant strains appears to be identical to that of the HE markers in strain Cl,.
DISCUSSION AND CONCLUSION
Our standard strain of Diplococcus pneumoniae (cl,) which is able to dis- criminate between various markers yields fewer transformants for low efficiency markers (LE) than for high efficiency markers (HE). From their work, EPHRUSSI-TAYLOR (1 966) and LOUARN and SICARD (1 968,1969) have concluded that discriminating strains yield LE transformants that are homozygous while HE transformants are heterozygous. By analysis OI the segregation behavior and the time required for phenotypic expression of both kinds of markers, we have confirmed this behavior for transformants occurring in Cl,. The nature of the mechanism that renders LE markers homozygous is not understood. Several different models might explain the characteristics of this class of markers.
1) LE markers are integrated through a single-strand insertion of the donor DNA, followed by the correction of mismatched bases by a mechanism of excision and repair.

MARKER DISCRIMINATION IN TRANSFORMATION 47
2) LE markers are integrated through a single-strand insertion of the donor DNA just ahead of the replicating fork.
3 ) LE markers are integrated through a double-strand insertion of the donor DNA.
Integration of both classes of markers in strain 401 yields the product charac- teristic of high efficiency markers in CZ,. GURNEY and Fox (1968) have shown that, in a strain similar to CZ,, the recombinant DNA for HE markers contains a single-strand fragment of donor DNA inserted into the DNA of the recipient genome. Since markers ami A-r9 (LE) and ami A-r1 (HE) have been shown to be the result of a single-site mutation (SICARD and EPHRUSSI-TAYLOR 1965), the difference in the integration efficiencies between these two markers in CZ, could be accounted for by the existence of one or several enzymes capable of making a distinction between different mismatched bases. The mutant strain 401 which has lost the property of discriminating between ami A-r9 and ami A-r1 mutations might be deficient for such an enzyme. The behavior of strain 401 does not per- mit us to distinguish among the above hypotheses, since they all involve an en- zyme that recognizes certain types of mismatched bases and somehow eliminates some classes. PETERSON and GUILD (1968) have also shown by a different ap- proach that in strain Rx, similar to 401 with respect to marker efficiencies, the integration of all markers tested occurred by single insertion of either of the two strands of the donor DNA.
The sensitivity of strain 401 to UV and to X-rays is the same as that of CZ,. Furthermore it was shown that the introduction, by trandormation into CZ, and 401, of a mutation in a gene governing the excisioii of pyrimidine dimers from DNA of irradiated cells (TIRABY 1971) has no effect on the discrimination process. Thus the discrimination process appears to be either partially or cam- pletely unrelated to the mechanism of repair of UV damage by excision and syn- thesis. This result is in agreement with the observation of LOUARN and SICARD (1965) showing that inhibition of repair enzymes by caffeine did not modify the characteristic efficieiicy of markers.
This research was supported by a grant from the “Dklkgation Gknkrale 1 la Recherche Scien- tifique et Technique.” We wish to thanlr DR. N. SICARD and MISS M. AHLQUIST for their help in preparing the manuscript.
LITERATURE CITED
CLARK, A. J., 1971
EPHRUSSI-TAYLOR, H., 1966
Toward a metabolic interpretation of genetic recombination of E . coli and its phages. Ann. Rev. Microbiol. 25: 437-464.
Genetic recombination in DNA-induced transformation of pneumo- COCCUS. IV. The pattern of transmission and phenotypic expression of high and low efficiency donor sites in the ami A locus. Genetics 54: 211-222.
EPHRUSSI-TAYLOR, H., A. M. SICARD and R. KAMEN, 1965 Genetic recombination in DNA- induced transformation of pneumococcus. I. T h e problem of relative efficiency of transform- ing factors. Genetics 51: 455-475.
GRAY, T. C. and H. EPHRUSSI-TAYLOR, 1967 Genetic recombination in DNA-induced transfor- mation of pneumococcus. V. The absence of interference, and evidence for the selective elimination of certain donor sites from the final recombinants. Genetics 57: 125-153.

48 G. TIRABY A N D M. A. SICARD
GURNEY, T. snd M. S. Fox, 1968 Physical and genetic hybrids formed in bacterial transfor- mation. J. Molec. Biol. 32: 83-100.
HOTCHKISS, R. D. and M. GABOR, 1970 Bacterial transformation with special reference to recom- bination process. Ann. Rev. Genetics 47: 1355-1368.
LACKS, S., 1965 Genetic recombination in pneumococcus. Pp. 159-164. In: The Physiology of Gene and Mutation Expression. Edited by M. KOHOUTOVA and J. HUBACEK. Academia, Prague. __ , 1966 Integration efficiency and genetic recombination in pneumococcal transformation. Genetics 53: 207-235. - , 1970 Mutants of Diplococcus pneumoniae that lack desoxyribonucleases and activities possibly pertinent to genetic transformation. J. Bacteriol. 101 : 373-383.
LITMAN, R. M., 1961 Genetic and chemical alteration in the transforming DNA of pneumo- COCCUS caused by ultraviolet light and by nitrous acid. J. Chimie Phys. 58: 997-1004.
LOUARN, J. M., 1970 Etude de la recombination ghnktique chez Diplococcus pneumoniae. Thesis, University of Toulouse, France.
LOUARN, J. M. and A. M. SICARD, 1965 Sur la sensibilitk diffkrentielle des marqueurs ghnbtiques aux rayonnements ultraviolets, et les mkcanismes de rkactivation au cours de la transfor- mation chez Diplococcus pneumoniae. C. R. Acad. Sc. Paris 261: 852-855. 1968 Transmission of genetic information during transformation in Diplococcus pneumoniae. Biochem. Biophys. Res. Commun. 30: 683-689. __ , 1969 Identical transformability of both strands of recipient DNA in Diplococcus pneumoniae. Biochem. Biophys. Res. Commun. 36: 101-109.
MARMUR, J., W. F. ANDERSON, L. MATTHEWS, I. BERNS, E. GEJEWSKA, D. LANE and P. DOTY, 1961 The effects of ultraviolet light on the biological and physical chemical properties of desoxyribonucleic acids. J. Cell Comp. Phys. 58: 33-55.
MARTIN, C. and H. EPHRUSSI-TAYLOR, 1964 Effects of partial deuteration on transforming DNA of pneumococcus. Biochim. Biophys. Acta 80: 41 1-421.
OTTOLENGHI, E. and R. D. HOTCHKISS, 1962 Release of genetic transforming agent from pneu- mococcal cultures during growth and disintegration. J. Exp. Med. 116: 491-519.
PETERSON, J. M. and W. R. GUILD, 1968 Fractionated strands of bacterial deoxyribonucleic acid. 111. Transformation efficiencies and rates of phenotypic expression. J. Bacteriol. 96: 1991- 1996.
RAVIN, A., 1959 Reciprocal capsular transformation of pneumococci. J. Bacteriol. 77: 296-309. SICARD, A. M., 1964 A new synthetic medium for Diplococcus pneumoniae, and its use for the
study of reciprocal transformation at the ami A locus. Genetics 50: 3144. SICARD, A. M. and H. EPHRUSSI-TAYLOR, 1965 Genetic recombination in DNA-induced transfor-
mation of pneumococcus. 11. Mapping the ami A region. Genetics 52: 1207-1227. SIROTNAK, F. M. and S. L. HACHTEL, 1969 Increased dihydrofolate reductase synthesis in
Diplococcus pneumonL‘ae following alteration of the structural gene. I . Genotype derivation and recombination analyses. Genetics 61 : 293-312.
Etude des mecanismes responsables des efficaciths de transformation chez Diplo- coccus pneumoniae. Thesis, University of Toulouse, France.
Integration efficiency in DNA-induced transformation of pneumococcus. I. A method of transformation in solid medium and its use for isolation of transformation-deficient and recombination-modified mutants. Genetics 75 : w-33.
Cellular metabolism in genetic transformation of pneumococci requirement for prote4n synthesis during induction of competence. J. Bacteriol. 101 : 860-871.
Appearance of a protein “agglutinin” on the spheroplast membrane of pneumococci during induction of competence. J. Bacteriol. 105: 1213-1215.
Corresponding Editor: R. E. ESPOSITO
- ,
TIRABY, G., 1971
TIRABY, G., J. P. CLAVERYS and A. M. SICARD, 1973
TOMASZ, A., 1970
TOMASZ, A. and E. ZANATI, 1971


















