
Inhibition of high-mobility group box 1 expression by ...
9
ORIGINAL ARTICLE Inhibition of high-mobility group box 1 expression by siRNA in rat hepatic stellate cells Wen-Song Ge, Jian-Xin Wu, Jian-Gao Fan, Yao-Jun Wang, Ying-Wei Chen Wen-Song Ge, Jian-Xin Wu, Jian-Gao Fan, Ying-Wei Chen, Department of Gastroenterology, Shanghai Xinhua Hospital Af- filiated to Shanghai Jiao Tong University School of Medicine, Shanghai 200092, China Yao-Jun Wang, Department of Gastroenterology, General Hos- pital of Jinan Military Region, Jinan 250031, Shandong Prov- ince, China Author contributions: Ge WS, Wu JX, Fan JG, Wang YJ and Chen YW designed research; Ge WS, Wang YJ and Chen YW performed research; Wang YJ and Chen YW contributed new reagents/analytic tools; Ge WS and Chen YW analyzed data; and Ge WS, Wang YJ and Chen YW wrote the paper. Supported by The Select and Train Outstanding Young Teach- ers Foundation of Shanghai, No. jdy08086 and WUJieping Ex- perimental Diagnosis of Liver Disease Medical Foundation, No. LDWMF-SY-2011B009 Correspondence to: Ying-Wei Chen, Vice-professor, Depart- ment of Gastroenterology, Shanghai Xinhua Hospital Affiliated to Shanghai Jiao Tong University School of Medicine, Shanghai 200092, China. [email protected]m Telephone: +86-21-25076431 Fax: +86-21-25071316 Received: July 5, 2011 Revised: September 5, 2011 Accepted: September 12, 2011 Published online: September 28, 2011 Abstract AIM: To explore the role of high-mobility group box 1 (HMGB1) protein during liver fibrogenesis and investi- gate the functional effects of HMGB1 gene silencing in hepatic stellate cells (HSCs) using siRNA. METHODS: Hepatic fibrosis in rats was induced throu- gh serial subcutaneous injections of dimethylnitrosamine, and expression of HMGB1 was detected by immunohis- tochemistry. HMGB1 siRNAs were developed and tran- siently transfected into HSC-T6 cells using Lipofectamine 2000. HMGB1 expression was evaluated by real-time polymerase chain reaction (PCR) and Western blotting analysis. Expression of α-smooth muscle actin (α-SMA) and collagen types Ⅰ and Ⅲ was evaluated by real-time PCR. Cell proliferation and the cell cycle were determined using the methyl thiazolyl tetrazolium method. Finally, collagen content in HSC supernatant was evaluated by an enzyme-linked immunosorbent assay. RESULTS: The results showed that HMGB1 was up- regulated during liver fibrosis and that its expression was closely correlated with the deposition of collagen. siRNA molecules were successfully transfected into HSCs and induced inhibition of HMGB1 expression in a time-dependent manner. Moreover, HMGB1 siRNA treatment inhibited synthesis of α-SMA and collagen types Ⅰ and Ⅲ in transfected HSCs. CONCLUSION: This study suggests a significant fun- ctional role for HMGB1 in the development of liver fibrosis. It also demonstrates that downregulation of HMGB1 expression might be a potential strategy to treat liver fibrosis. © 2011 Baishideng. All rights reserved. Key words: Hepatic fibrosis; High-mobility group box 1; Hepatic stellate cells; RNA interference Peer reviewers: Ekihiro Seki, MD, PhD, Department of Med- icine, University of California San Diego, Leichag Biomedical Research Building Rm 349H, 9500 Gilman Drive MC#0702, La Jolla, CA 92093-0702, United States; London Lucien Ooi, Professor, Chairman, Division of Surgery, Singapore General Hospital, 1 Hospital Drive, 169608, Singapore Ge WS, Wu JX, Fan JG, Wang YJ, Chen YW. Inhibition of high- mobility group box 1 expression by siRNA in rat hepatic stellate cells. World J Gastroenterol 2011; 17(36): 4090-4098 Available from: URL: http://www.wjgnet.com/1007-9327/full/v17/ i36/4090.htm DOI: http://dx.doi.org/10.3748/wjg.v17.i36.4090 INTRODUCTION Hepatic fibrosis is a major medical problem associated with significant morbidity and mortality. Regardless of 4090 World J Gastroenterol 2011 September 28; 17(36): 4090-4098 ISSN 1007-9327 (print) ISSN 2219-2840 (online) © 2011 Baishideng. All rights reserved. Online Submissions: http://www.wjgnet.com/1007-9327office [email protected] doi:10.3748/wjg.v17.i36.4090 September 28, 2011|Volume 17|Issue 36| WJG|www.wjgnet.com
Transcript of Inhibition of high-mobility group box 1 expression by ...

ORIGINAL ARTICLE
Inhibition of high-mobility group box 1 expression by siRNA in rat hepatic stellate cells
Wen-Song Ge, Jian-Xin Wu, Jian-Gao Fan, Yao-Jun Wang, Ying-Wei Chen
Wen-Song Ge, Jian-Xin Wu, Jian-Gao Fan, Ying-Wei Chen, Department of Gastroenterology, Shanghai Xinhua Hospital Af-filiated to Shanghai Jiao Tong University School of Medicine, Shanghai 200092, ChinaYao-Jun Wang, Department of Gastroenterology, General Hos-pital of Jinan Military Region, Jinan 250031, Shandong Prov-ince, China Author contributions: Ge WS, Wu JX, Fan JG, Wang YJ and Chen YW designed research; Ge WS, Wang YJ and Chen YW performed research; Wang YJ and Chen YW contributed new reagents/analytic tools; Ge WS and Chen YW analyzed data; and Ge WS, Wang YJ and Chen YW wrote the paper.Supported by The Select and Train Outstanding Young Teach-ers Foundation of Shanghai, No. jdy08086 and WUJieping Ex-perimental Diagnosis of Liver Disease Medical Foundation, No. LDWMF-SY-2011B009Correspondence to: Ying-Wei Chen, Vice-professor, Depart-ment of Gastroenterology, Shanghai Xinhua Hospital Affiliated to Shanghai Jiao Tong University School of Medicine, Shanghai 200092, China. [email protected]: +86-21-25076431 Fax: +86-21-25071316 Received: July 5, 2011 Revised: September 5, 2011Accepted: September 12, 2011Published online: September 28, 2011
AbstractAIM: To explore the role of high-mobility group box 1 (HMGB1) protein during liver fibrogenesis and investi-gate the functional effects of HMGB1 gene silencing in hepatic stellate cells (HSCs) using siRNA.
METHODS: Hepatic fibrosis in rats was induced throu-gh serial subcutaneous injections of dimethylnitrosamine, and expression of HMGB1 was detected by immunohis-tochemistry. HMGB1 siRNAs were developed and tran-siently transfected into HSC-T6 cells using Lipofectamine 2000. HMGB1 expression was evaluated by real-time polymerase chain reaction (PCR) and Western blotting analysis. Expression of α-smooth muscle actin (α-SMA) and collagen types Ⅰ and Ⅲ was evaluated by real-time PCR. Cell proliferation and the cell cycle were determined
using the methyl thiazolyl tetrazolium method. Finally, collagen content in HSC supernatant was evaluated by an enzyme-linked immunosorbent assay.
RESULTS: The results showed that HMGB1 was up-regulated during liver fibrosis and that its expression was closely correlated with the deposition of collagen. siRNA molecules were successfully transfected into HSCs and induced inhibition of HMGB1 expression in a time-dependent manner. Moreover, HMGB1 siRNA treatment inhibited synthesis of α-SMA and collagen types Ⅰ and Ⅲ in transfected HSCs.
CONCLUSION: This study suggests a significant fun-ctional role for HMGB1 in the development of liver fibrosis. It also demonstrates that downregulation of HMGB1 expression might be a potential strategy to treat liver fibrosis.
© 2011 Baishideng. All rights reserved.
Key words: Hepatic fibrosis; High-mobility group box 1; Hepatic stellate cells; RNA interference
Peer reviewers: Ekihiro Seki, MD, PhD, Department of Med-icine, University of California San Diego, Leichag Biomedical Research Building Rm 349H, 9500 Gilman Drive MC#0702, La Jolla, CA 92093-0702, United States; London Lucien Ooi, Professor, Chairman, Division of Surgery, Singapore General Hospital, 1 Hospital Drive, 169608, Singapore
Ge WS, Wu JX, Fan JG, Wang YJ, Chen YW. Inhibition of high-mobility group box 1 expression by siRNA in rat hepatic stellate cells. World J Gastroenterol 2011; 17(36): 4090-4098 Available from: URL: http://www.wjgnet.com/1007-9327/full/v17/i36/4090.htm DOI: http://dx.doi.org/10.3748/wjg.v17.i36.4090
INTRODUCTIONHepatic fibrosis is a major medical problem associated with significant morbidity and mortality. Regardless of
4090
World J Gastroenterol 2011 September 28; 17(36): 4090-4098 ISSN 1007-9327 (print) ISSN 2219-2840 (online)
© 2011 Baishideng. All rights reserved.
Online Submissions: http://www.wjgnet.com/[email protected]:10.3748/wjg.v17.i36.4090
September 28, 2011|Volume 17|Issue 36|WJG|www.wjgnet.com

Ge WS et al. Inhibition of HMGB1 expression in HSC
the underlying aetiology[1], hepatic fibrosis is character-ized by the accumulation of excess extracellular matrix (ECM). The amount of matrix deposition depends on the balance between its synthesis and degradation. When synthesis of ECM exceeds its degradation, the patholog-ical accumulation of ECM leads to liver fibrosis. There-fore, a critical balance must be achieved between main-taining the proper amount of ECM for homeostasis, while at the same time, providing a means of ensuring that excess or improper accumulation does not occur. High-mobility group box 1 (HMGB1) protein was originally identified as a nuclear nonhistone protein with DNA-binding domains, and it has been implicated as an important endogenous danger signaling molecule. In addition, it can be secreted from cells and exert ex-tracellular functions as a proinflammatory cytokine[2,3]. Increasing evidence now points to multiple functions of HMGB1 in infection, tissue injury, inflammation, apoptosis, and the immune response[4]. HMGB1 can be released both through active secretion from various cells, including activated monocytes/macrophages, neutro-phils, and endothelial cells, and through passive release from necrotic cells[3-7]. HMGB1 can directly promote the secretion of proinflammatory cytokines [tumor necrosis factor (TNF), interleukin (IL)-1A/B, IL-6 and IL-8] and chemokines (macrophage inflammatory protein-1A/B) by peripheral blood mononuclear cells (PBMCs)[8,9]. In turn, PBMCs also produce different cytokines that are potentially involved in virus-induced liver damage. HMGB1 acts as a chemoattractant for fibroblasts and endothelial and smooth muscle cells, which are cell types that significantly contribute to wound repair[9,10]. Consequently, HMGB1 can directly stimulate fibroblast proliferation and participate in fibrogenesis[4]. Addition-ally, inhibitors of HMGB1 significantly reduce tissue damage[5,6]. Moreover, Hamada et al[4] have reported that inhibition of HMGB1 may be beneficial in pulmonary fibrosis. Therefore, we postulated that inhibiting the up-regulation of HMGB1 during liver fibrogenesis could be a potential strategy for treating liver fibrosis.
RNA interference is known as a powerful tool for post-transcriptional gene silencing[11] and has opened new avenues in gene therapy. In this study, we induced hepatic fibrosis in rats through serial subcutaneous injections of dimethylnitrosamine (DMN) for 4 wk and evaluated the expression of HMGB1 during the process of hepatic fibrogenesis. Additionally, siRNA molecules targeting the sequences within the rat HMGB1 gene were transfected into hepatic stellate cell (HSC)-T6 cells. The results show that the expression of HMGB1 was correlated with col-lagen deposition during hepatic fibrosis and that down-regulating HMGB1 expression could prohibit collagen production and enhance collagen degradation.
MATERIALS AND METHODSAnimal modelsThirty-two 6-wk-old male Sprague-Dawley rats (230-260 g)
were purchased from the Shanghai Laboratory Animal Centre of Chinese Academy of Sciences and fed ad libitum with standard laboratory chow. All rats received humane care according to the Guide for the Care and Use of Laboratory Animals by the Chinese Academy of Sciences. Hepatic fibrosis was induced by intraperitoneal injections of 1% DMN (1 mL/kg body weight) for three consecu-tive days per week for up to 4 wk[11]. Rats were sacrificed at 1, 2 and 3 wk from the first DMN injection. Liver tis-sues were either snap-frozen in liquid nitrogen or fixed in 10% formalin for histology and immunostaining.
Histological and immunohistochemical examinationLiver tissue sections were stained with hematoxylin-eosin (HE) for histopathological examination. Immunohis-tochemical examination was performed to detect the expression of HMGB1 and collagen types Ⅰ and Ⅲ in liver tissues. Briefly, the paraffin sections of left median hepatic lobes were incubated with 3% H2O2 in methanol at 37 ℃ for 10 min to quench endogenous peroxidase ac-tivity. After blocking at room temperature for 20 min, the sections were incubated with antibodies against HMGB1 (R and D Systems, Germany), collagen type Ⅰ or col-lagen type Ⅲ (Boster, Wuhan, China) overnight at 4 ℃ followed by incubation with horseradish-peroxidase-conjugated secondary antibody (Dako, Kyoto, Japan) at 37 ℃ for 20 min. Finally, the signals were detected using the Diaminobenzidine Substrate Kit (Vector Laborato-ries, Burlingame, CA, United States), and a positive out-come was indicated by brown staining in the cytoplasm or nucleus. For the semiquantitative analysis of HMGB1 and collagen expression, the brown-stained tissues in immunohistostaining sections were measured on an im-age analyzer by a technician blinded to the samples. Five fields were selected randomly from each of two sections, and six rats from each group were examined.
Double immunostaining of HMGB1 and α-smooth mus-cle actinLiver sections were blocked with 5% normal goat serum after fixing and then simultaneously incubated with both monoclonal anti-HMGB1 (R and D Systems, Germany) and polyclonal α-smooth muscle actin (α-SMA) (Fre-mont, CA, United States) antibodies prepared in phos-phate-buffered saline (PBS). The sections were incubated overnight at 4 ℃ or 1 h at room temperature and then washed with PBS. Sections were then simultaneously in-cubated with fluorescein-isothiocyanate-conjugated sec-ondary antibody and rhodamine-conjugated secondary antibody for 30 min at 37 ℃ in the dark. Both primary antibodies were produced in different species. Antibody labeling was examined under a Zeiss LSM-510 laser scanning confocal microscope.
Cell cultureThe HSC-T6 cell line, an immortalized rat HSC line, which has a stable phenotype and biochemical charac-teristics, was kindly provided by Dr. SL Friedman (Divi-
4091 September 28, 2011|Volume 17|Issue 36|WJG|www.wjgnet.com

sion of Liver Diseases, Mount Sinai School of Medicine, New York, NY, United States). All cells were cultured in RPMI1640 medium supplemented with 10% fetal bovine serum and 5% antibiotics and incubated at 37 ℃ in a hu-midified atmosphere of 5% CO2. Cells were seeded at 2 × 105 per well in six-well plates 24 h before transfection. The amount of siRNA and transfection reagent was calculated according to the manufacturer’s instructions.
Immunofluorescence studyHSC-T6 cells were cultured for 24 h on glass coverslips and fixed in 4% formaldehyde for 30 min at room tem-perature prior to detergent extraction with 0.1% Triton X-100 for 10 min at 4 ℃. Coverslips were saturated with PBS containing 2% bovine serum albumin (BSA) for 1 h at room temperature. Next, cells were incubated with the specific primary antibody for HMGB1 (R and D Sys-tems, Germany) in 1% BSA for 1 h, washed, and incu-bated with secondary antibody (TRITC AffiniPure Goat Anti-Rabbit IgG, EarthOx, LLC, United States). Finally, cells were stained for 30 min at room temperature with 4,6-diamidino-2-phenylindole. Slides were viewed with a Zeiss LSM-510 laser scanning confocal microscope.
Preparation of siRNA, construction of siRNA expression vector and transfection assayThe siRNAs for rat HMGB1 mRNA were designed and synthesised by Invitrogen Life Technologies. We pre-pared three siRNAs, and the most effective one was se-lected for construction of the siRNA expression vector. The siRNA sequences used are shown in Table 1. Nega-tive control siRNAs were used to assess non-specific gene silencing effects, and the mock group was the non-transfection group. Cells were transfected with a mixture of plasmid DNA and Lipofectamine 2000 (Invitrogen) in Opti-MEM I medium without serum as recommend-ed by the manufacturer. The medium was then replaced with standard RPMI medium (containing 10% FBS and gentamicin) 24 h post-transfection.
Real-time quantitative polymerase chain reactionTotal RNA was extracted at different time points after siRNA transfection using the Trizol kit (Gibco/Life Technologies) according to the manufacturer’s protocol. The mixture of RNA and primers was loaded into the polymerase chain reaction (PCR) amplifier. The PCR protocol was as follows: predenaturate setting at 95 ℃ for 5 min, 94 ℃ for 45 s, annealing at 50 ℃ for 1 min, and extension at 72 ℃ for 1 min. The PCR was per-formed for 40 cycles followed by a final extension at 72 ℃ for 10 min. We then visualized the PCR product by running it on a 1.5% agarose gel and quantitatively analysed it with Lab Works 4.5 analysis software.
Western blottingThe same quantities of cells were collected from the four groups, and the protein was extracted from the cells at the 24, 48 and 72 h after transfection. The pro-
tein content in the supernatant was detected using the bicinchoninic acid method. An equal amount of protein was used for sodium dodecyl sulfate polyacrylamide gel electrophoresis electrophoresis and transferred onto a polyvinylidene fluoride (PVDF) membrane. The PVDF membrane was incubated overnight at 4 ℃ with mono-clonal anti-human HMGB1 (1:300) and was then incu-bated for 2 h with a secondary antibody (1:5000). Finally, after staining and fixing, the film was analyzed using the Image Analysis System.
Enzyme-linked immunosorbent assay Commercial kits (Sigma, St. Louis, MO, United States) were used to quantitate the amount of collagen types Ⅰ and Ⅲ in the culture supernatant of HSCs at different time points after siRNA transfection.
Methyl thiazolyl tetrazolium used for observing cell pro-liferationThe cell suspension was inoculated into 96-well plates at 1000 cells per well with eight ambi-wells and incubated for 1, 2, 3, 4 and 5 d after transfection. Cells were incubated with 20 μL methyl thiazolyl tetrazolium for 4 h. After centrifugation, 150 μL dimethyl sulfoxide was added to the precipitate, and the absorbance of the enzyme was measured at 490 nm. Cell growth rates (average absor-bance of each transfected group/non-transfected group) were then calculated.
Statistical analysisContinuous data were expressed as the mean ± SD and were analyzed using the Student’s t test. Correlations among the study variables were tested using Pearson’s correlation coefficients. P < 0.05 were considered sta-tistically significant. All calculations were performed using SPSS version 13.0 (SPSS Inc., Chicago, IL, United States).
RESULTSHistological and immunohistochemical assessmentTo investigate the expression of HMGB1 during liver fibrosis, liver sections were analysed by HE staining and immunohistochemistry. We localized HMGB1 and col-lagen types Ⅰ and Ⅲ in liver specimens by immunohisto-chemistry. None of these proteins were observed in con-trol rat livers. In fibrotic rat livers, HMGB1 was markedly increased during liver fibrogenesis and was correlated with
4092 September 28, 2011|Volume 17|Issue 36|WJG|www.wjgnet.com
Plasmid constructs Target sequence in mRNA(5’-3’)
HMGB1-1 (shRNAH1) GCAAATGACTCAATCTGATT HMGB1-2 (shRNAH2) AATAGGAAAAGGATATTGCT HMGB1-3 (shRNAH3) ACCCGGATGCTTCTGTCAAC
Table 1 Design of small interfering RNA sequences for high-mobility group box 1
HMGB1: High-mobility group box 1.
Ge WS et al. Inhibition of HMGB1 expression in HSC
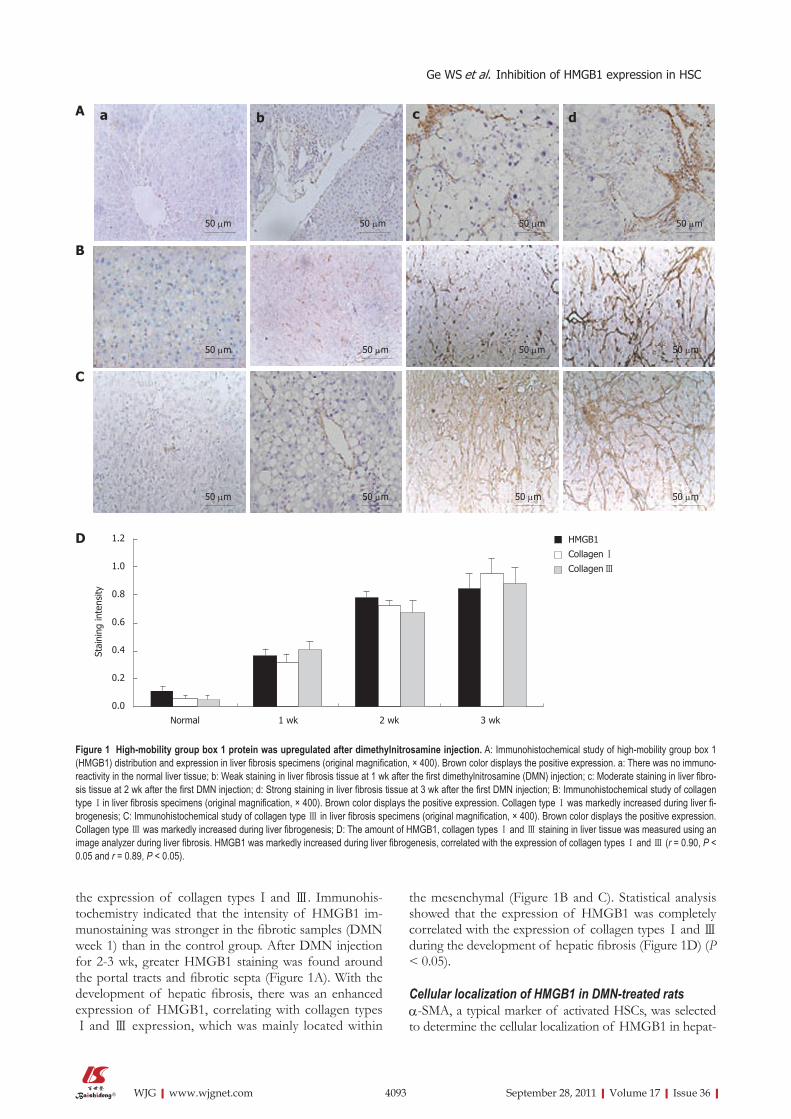
the expression of collagen types Ⅰ and Ⅲ. Immunohis-tochemistry indicated that the intensity of HMGB1 im-munostaining was stronger in the fibrotic samples (DMN week 1) than in the control group. After DMN injection for 2-3 wk, greater HMGB1 staining was found around the portal tracts and fibrotic septa (Figure 1A). With the development of hepatic fibrosis, there was an enhanced expression of HMGB1, correlating with collagen typesⅠ and Ⅲ expression, which was mainly located within
the mesenchymal (Figure 1B and C). Statistical analysis showed that the expression of HMGB1 was completely correlated with the expression of collagen types Ⅰ and Ⅲ during the development of hepatic fibrosis (Figure 1D) (P < 0.05).
Cellular localization of HMGB1 in DMN-treated ratsα-SMA, a typical marker of activated HSCs, was selected to determine the cellular localization of HMGB1 in hepat-
4093 September 28, 2011|Volume 17|Issue 36|WJG|www.wjgnet.com
Figure 1 High-mobility group box 1 protein was upregulated after dimethylnitrosamine injection. A: Immunohistochemical study of high-mobility group box 1 (HMGB1) distribution and expression in liver fibrosis specimens (original magnification, × 400). Brown color displays the positive expression. a: There was no immuno-reactivity in the normal liver tissue; b: Weak staining in liver fibrosis tissue at 1 wk after the first dimethylnitrosamine (DMN) injection; c: Moderate staining in liver fibro-sis tissue at 2 wk after the first DMN injection; d: Strong staining in liver fibrosis tissue at 3 wk after the first DMN injection; B: Immunohistochemical study of collagen type Ⅰin liver fibrosis specimens (original magnification, × 400). Brown color displays the positive expression. Collagen type Ⅰ was markedly increased during liver fi-brogenesis; C: Immunohistochemical study of collagen type Ⅲ in liver fibrosis specimens (original magnification, × 400). Brown color displays the positive expression. Collagen type Ⅲ was markedly increased during liver fibrogenesis; D: The amount of HMGB1, collagen types Ⅰ and Ⅲ staining in liver tissue was measured using an image analyzer during liver fibrosis. HMGB1 was markedly increased during liver fibrogenesis, correlated with the expression of collagen types Ⅰ and Ⅲ (r = 0.90, P < 0.05 and r = 0.89, P < 0.05).
A
B
C
50 μm 50 μm50 μm50 μm
50 μm 50 μm50 μm50 μm
50 μm 50 μm50 μm50 μm
Normal 1 wk 2 wk 3 wk
1.2
1.0
0.8
0.6
0.4
0.2
0.0
Stai
ning
inte
nsity
HMGB1
Collagen ⅠCollagen Ⅲ
D
a b c d
Ge WS et al. Inhibition of HMGB1 expression in HSC

ic fibrosis tissue. The localization of HMGB1 and α-SMA was visualized by immunofluorescent double labeling and laser scanning confocal microscopy. The image analysis showed a diffused distribution of HMGB1 throughout the hepatic fibrosis tissue (Figure 2A), and a similar distri-bution was observed for α-SMA (Figure 2B). When the two images were merged, there was a very high degree of co-localization of HMGB1 with α-SMA throughout the hepatic fibrosis tissue (Figure 2C).
Intracellular localization of HMGB1 in activated HSC-T6 cellsAn immunofluorescence study of HSC-T6 cells after 24 h of culture demonstrated the intracellular localization of HMGB1. We evaluated the subcellular localization of HMGB1 by separating bulk nuclei and cytosolic fractions, and HMGB1 was detected primarily within the cytosol of activated HSC-T6 cells (Figure 3).
Selection of HMGB1 mRNA sequence targetAs shown in Table 1, a total of three candidate siRNA sequences were chosen to be complementary to various regions of the rat HMGB1 gene. In a set of preliminary experiments designed to identify the most appropriate se-quence for further study, these sequences were transfected
into HSC-T6 using Lipofectamine. Forty-eight hours after transfection, HMGB1 transcript and protein levels were reduced in transfected cells. This HMGB1 gene-silencing effect was reproducible and was specific in that it failed to knock down the expression of an unrelated protein, β-actin. All three HMGB1 shRNAs tested in this study were able to reduce the HMGB1 expression in HSC-T6 cells compared with the negative control (NC) siRNA transfectants. Although all three HMGB1 shRNA con-structs were effective, shRNAH3 was more efficient in reducing the HMGB1 transcript levels than shRNAH2 and shRNAH1 (Figure 4A). Western blotting analysis (Fig-ure 4C) further confirmed the shRNAH3 silencing of the HMGB1 protein in HSC-T6 cells. Semiquantitative analysis of the real-time (RT)-PCR and Western blot results (Fig-ure 4B and D) also showed that HMGB1 shRNAH3 de-creased the expression of HMGB1 in HSC-T6 cells more efficiently than shRNAH2 and shRNAH1. Accordingly, we chose shRNAH3 for the subsequent experiments.
HMGB1 siRNA downregulated mRNA expression of α-SMA and types Ⅰ and Ⅲ collagen in HSC-T6To investigate the effect of HMGB1 siRNA on HSCs and its potential molecular mechanisms, we detected the
4094 September 28, 2011|Volume 17|Issue 36|WJG|www.wjgnet.com
Figure 2 Double immunostaining was used to analyze the cellular localization of high-mobility group box 1 protein and α-smooth muscle actin in hepatic fibrosis tissue (original magnification, × 200). A: α-smooth muscle actin (α-SMA) was stained with polyclonal α-SMA antibody and secondarily by rhodamine -conju-gated anti-rabbit antibody (green); B: High-mobility group box 1 (HMGB1) was stained with monoclonal anti-HMGB1 antibody and secondarily by fluoresceinisothiocya-nate-conjugated anti-rabbit antibody (red); C: The yellow areas on the merged image show co-localization of α-SMA and HMGB1.
A B C
Figure 3 High-mobility group box 1 protein expression in hepatic stellate cell-T6 cells by immunofluorescence staining (original magnification, × 200). A: High-mobility group box 1 (HMGB1) protein was stained with monoclonal anti-HMGB1 antibody and secondarily by fluoresceinisothiocyanate-conjugated anti-rabbit antibody (red); B: Nuclei were labelled with 4’,6-diamidino-2-phenylindole (blue); C: The merge picture.
A B C
Ge WS et al. Inhibition of HMGB1 expression in HSC

mRNA expression of some profibrogenic markers, includ-ing α-SMA and collagen types Ⅰ and Ⅲ, in transfected HSC-T6. As shown in Figure 5, HMGB1 siRNA reduced the mRNA levels of α-SMA and collagen types Ⅰ and Ⅲ. HMGB1 siRNA reduced the collagen content in the HSC-T6 supernatantTo confirm the effect of HMGB1 siRNA on collagen se-cretion and degradation, we examined the amount of colla-gen types Ⅰ and Ⅲ in HSCs 48 and 72 h after transfection with shRNAH3 using an ELISA. The results reveal that the content of both collagen types Ⅰ and Ⅲ was decreased af-ter transfection with HMGB1 siRNA. Compared with the NC group, the content of collagen types Ⅰ and Ⅲ was re-duced to 63% and 61%, respectively, 72 h after shRNAH3 transfection (Figure 6).
HMGB1 siRNA inhibited HSC-T6 cells proliferation The trypan blue dye test showed that there were no sig-
nificant differences in the number of cells in the three-groups 2 d after transfection (P > 0.05), but the prolif-eration in the shRNAH3 group was less than that in the NC group and non-transfection group (Mock group) 3, 4 and 5 d after transfection (P < 0.05, Figure 7). A cell cycle study also indicated that cells were arrested in the G0/G1 phase and that the proportion of cells in the S phase was significantly reduced after downregulation of HMGB1 in HSCs (Table 2).
DISCUSSIONLiver fibrosis is highly associated with chronic hepatocel-lular injury and the subsequent inflammatory response that produces inflammatory cytokines and recruits in-flammatory leukocytes to the injured site. This inflam-matory circumstance in the liver drives the activation of HSCs through various fibrogenic mediators[12,13]. Acti-vated HSCs transdifferentiate into myofibroblasts, which then produce excessive amounts of ECM proteins, including collagen types Ⅰ, Ⅲ and Ⅳ. This leads to irre-versible collagen deposition, resulting in liver fibrosis[12,13]. Many studies have suggested that enhancement of matrix degradation may prove particularly valuable in response to injury caused by matrix deposition[14-17]. Some studies have shown that HMGB1 can stimulate proinflammatory cytokine synthesis and directly stimulate fibroblast prolif-eration and participate in fibrogenesis[8-10].
4095 September 28, 2011|Volume 17|Issue 36|WJG|www.wjgnet.com
Figure 4 Screening the most effective high-mobility group box 1 siRNA sequence. Total RNA and protein were obtained from hepatic stellate cell-T6 transfected with negative control (NC), mock and three different high-mobility group box 1 (HMGB1) siRNA molecules (shRNAH1, shRNAH2 and shRNAH3). A: Real-time poly-merase chain reaction (RT-PCR) for the effect of three different HMGB1 siRNA molecules on HMGB1 mRNA level 48 h after transfection. The expression was normal-ized against β-actin; B: Semiquantitative analysis of the RT-PCR result; C: Western blotting analyzed HMGB1 protein expression 48 h after transfection; D: Semiquan-titative analysis of the western blotting results. Data represent results from one of three similar experiments. Results show that all three HMGB1 shRNA constructs were effective, but shRNAH3 was more efficient in reducing the HMGB1 mRNA and protein levels than shRNAH2 and shRNAH1.
Figure 5 High-mobility group box 1 siRNA inhibited α-smooth muscle actin, collagen typesⅠand Ⅲ mRNA expression in hepatic stellate cell-T6 cells. A: Real-time polymerase chain reaction (RT-PCR) analysis for α-smooth muscle actin (α-SMA), collagen typesⅠand Ⅲ mRNA expression in hepatic stellate cell-T6 cells after siRNA high-mobility group box 1 transfection. β-actin was used as the internal loading control; B-D: Semiquantitative analysis of the RT-PCR result. aP < 0.05 vs negative controls (NC) or mock.
B C DA
α-SMA
Collagen 1
Collagen 3
β-actin
M NC Mock siRNA 1.41.21.00.80.60.40.20.0
NC Mock siRNA
α-S
MA
1.2
1.0
0.8
0.6
0.4
0.2
0.0 NC Mock siRNA
Colla
gen Ⅲ
1.41.21.00.80.60.40.20.0
NC Mock siRNA
Colla
gen Ⅰ
aa a
Cell cycle phases(%) ShRNAH3 group NC group
G0/G1 phase 58.31% ± 0.48%a 44.25% ± 0.63% S phase 29.12% ± 1.26%a 41.32% ± 1.58% G2/M phase 12.57% ± 1.04% 14.53% ± 1.28%
Table 2 Effect of high-mobility group box 1 siRNA on the cell cycle
aP < 0.05 vs negataive controls (NC) group.
Ge WS et al. Inhibition of HMGB1 expression in HSC
A
1000750500
250
100
RNAH M NC Mock 1 2 3
H
MG
B1 β
-act
in
C
β-actin
HMGB1
RNAH M NC Mock 1 2 34.5
4.03.53.02.52.01.51.00.50.0
B
M NC Mock 1 2 3 ShRNAH
Expr
essi
on o
f H
MG
B1 m
RN
A
4.03.53.02.52.01.51.00.50.0
D
M NC Mock 1 2 3 ShRNAH
Expr
essi
on o
f H
MG
B1 p
rote
in

Increased expression of HMGB1 has been reported in several liver diseases, including Con A-induced hepati-tis[18], hepatic ischemia[2], and orthotopic liver transplanta-tion (OLT)[19]. In the present study, we evaluated HMGB1 expression in the DMN rat model. We found that the level of HMGB1 was upregulated during DMN injec-tion. Moreover, the expression of HMGB1 was closely correlated with the expression of collagen types Ⅰ and Ⅲ and was mainly localized to the nonparenchymal cells, especially HSCs. These results suggest that HMGB1 is involved in hepatic fibrogenesis and may play a critical role in the reversal process of liver fibrosis.
HMGB1 was originally identified as a nuclear non-histone protein with DNA-binding domains and was implicated as an important endogenous danger signal-ing molecule. Although predominantly located in the nucleus of quiescent cells, HMGB1 can be actively se-creted in response to exogenous and endogenous infla-mmatory stimuli such as endotoxin, TNF-α, IL-1, and interferon-γ[20,21]. In addition, extracellular HMGB1 mediates a wide range of inflammatory responses and promotes cell proliferation, migration, and differentia-tion[10,22]. The cytoplasmic localization of HMGB1 in our
study may suggest that HMGB1 plays extra nuclear roles in liver fibrosis and that HSC-T6 cells may even secrete HMGB1 to promote extracellular functions. The subcel-lular location of HMGB1 in monocytic cells is known to be dependent on the acetylation status of the nuclear localization signal (NLS) of the HMGB1 protein[23]. Inflammatory signals promote acetylation of the NLS, leading to cytoplasmic accumulation of HMGB1 in se-cretory lysosomes in the monocytic cells[24]. These secre-tory lysosomes are subsequently exocytosed when the monocytic cells are triggered by a second inflammatory stimulus. Whether the subcellular location of HMGB1 in HSC-T6 cells is regulated in a similar way remains to be investigated.
It has become apparent in recent years that HMGB1 is instrumental in mediating a response to tissue dam-age and infection. HMGB1 released from necrotic or damaged cells not only triggers inflammation as a non-specific proinflammatory cytokine but also triggers the adaptive immune response[25,26]. Extracellular HMGB1 functions as a damage-associated molecular pattern mol-ecule and activates proinflammatory signaling pathways by activating pattern-recognition receptors including toll-like receptor 4 (TLR4) and the receptor for advanced glycation end-products (RAGE)[27,28]. A previous report showed that RAGE expression in fibrotic livers is re-stricted to HSCs; its expression is up regulated during cellular activation and transition to myofibroblasts[29], strongly suggesting that HMGB1 is involved in the pathogenesis of liver fibrosis. TLR4 has been suggested to be a receptor for extracellular HMGB1[30,31], and previous studies have indicated that the interaction of HMGB1 with TLR4 plays a critical role in hepatic fibro-sis[32]. To date, little has been reported about the patho-genic interactions between HMGB1 and HSCs in terms of profibrogenic propensity. Kao provided evidence that HMGB1 up regulates α-SMA expression and suppresses the activity of the collagen-degrading enzyme matrix metalloproteinase-2[33]. That study also implied that HMGB1, once it is released during rejection of OLT, activates HSCs and exhibits profibrogenic effects either by increasing the HSC population and ECM deposition
4096 September 28, 2011|Volume 17|Issue 36|WJG|www.wjgnet.com
Figure 6 Determination of content of collagen typesⅠand Ⅲ after shRNAH3 transfection. Enzyme-linked immunosorbent assays were used for quantitative de-termination of collagen typesⅠand Ⅲ content hepatic stellate cells (HSCs) culture supernatant at 48 and 72 h after shRNAH3 transfection using Lipofectamine 2000. Values are presented as mean ± SD. aP < 0.05 vs negative controls (NC) and HSC group.
Figure 7 High-mobility group box 1 siRNA suppressed hepatic stellate cell-T6 proliferation. Cell growth curves of hepatic stellate cell-T6 transfected with shRNAH3 were analyzed by methyl thiazolyl tetrazolium conversion. Each sample was tested in triplicate and error bars were included. Compared with negative controls (NC) group and non-transfection group, proliferation of shR-NAH3 group was less at the 3-5 d after transfection (P < 0.05).
9
8
7
6
5
4
3
2
1
0
Colla
gen Ⅰ
cont
ent
(pg/
mL)
8
7
6
5
4
3
2
1
0
aa
HSC
NC
ShRNAH
48 h 72 h
HSC
NC
ShRNAH
aa
48 h 72 h
Colla
gen Ⅲ c
onte
nt (
pg/m
L)
1 2 3 4 5 (d)
0.9
0.8
0.7
0.6
0.5
0.4
0.3
0.2
0.1
0.0
Mock
NC
ShRNAH3
Ge WS et al. Inhibition of HMGB1 expression in HSCA
490

sion of HMGB1 could be a potential mechanism for mediating collagen expres-sion and downregulating HMGB1 expression might present as a potential strategy to treat liver fibrosis.Innovations and breakthroughsStudies of targeting in vitro and in vivo over expressed genes in hepatic fibrosis by RNA interference, including transforming growth factor-β, connective tissue growth factor and p90RSK, have been reported. However, there has been still no report about targeting HMGB1 by siRNA in hepatic fibrosis. In the pres-ent study, the authors used siRNA approach to block HMGB1 expression in HSC-T6 cells, to determine the role of constitutively activated HMGB1 during hepatic fibrosis pathogenesis, and to explore the role and molecular mecha-nism of targeting HMGB1 in hepatic fibrosis therapy.ApplicationsBy investigating the effect of silencing HMGB1 expression by siRNA on the collagen synthesis and proliferation of HSC-T6 cells, this study may provides a new strategy for biological therapy of liver fibrosis by targeting HMGB1.TerminologyHMGB1 was originally identified as a nuclear nonhistone protein with DNA-binding domains and implicated as an important endogenous danger signaling molecule. But it can also be secreted from cells and exert extracellular functions as a proinflammatory cytokine. HSCs are a minor and quiescent cell type in the liver that usually reside in the space of Disse, but which undergo activation after hepatic injury to produce large quantities of fibrillar collagens. Peer reviewThe authors demonstrated the increase of HMGB1 expression in fibrotic livers. Then, they investigated the effect of HMGB1 silencing by siRNA on stellate cell activation and proliferation. The results show that siRNA for HMGB1 signifi-cantly inhibits collagen expression and stellate cell proliferation.
REFERENCES1 Moreira RK. Hepatic stellate cells and liver fibrosis. Arch
Pathol Lab Med 2007; 131: 1728-1734 2 Tsung A, Sahai R, Tanaka H, Nakao A, Fink MP, Lotze MT,
Yang H, Li J, Tracey KJ, Geller DA, Billiar TR. The nuclear factor HMGB1 mediates hepatic injury after murine liver ischemia-reperfusion. J Exp Med 2005; 201: 1135-1143
3 Wang H, Bloom O, Zhang M, Vishnubhakat JM, Ombrel-lino M, Che J, Frazier A, Yang H, Ivanova S, Borovikova L, Manogue KR, Faist E, Abraham E, Andersson J, Andersson U, Molina PE, Abumrad NN, Sama A, Tracey KJ. HMG-1 as a late mediator of endotoxin lethality in mice. Science 1999; 285: 248-251
4 Hamada N, Maeyama T, Kawaguchi T, Yoshimi M, Fuku-moto J, Yamada M, Yamada S, Kuwano K, Nakanishi Y. The role of high mobility group box1 in pulmonary fibrosis. Am J Respir Cell Mol Biol 2008; 39: 440-447
5 Ilmakunnas M, Tukiainen EM, Rouhiainen A, Rauvala H, Arola J, Nordin A, Mäkisalo H, Höckerstedt K, Isoniemi H. High mobility group box 1 protein as a marker of hepato-cellular injury in human liver transplantation. Liver Transpl 2008; 14: 1517-1525
6 Scaffidi P, Misteli T, Bianchi ME. Release of chromatin pro-tein HMGB1 by necrotic cells triggers inflammation. Nature 2002; 418: 191-195
7 Ito I, Fukazawa J, Yoshida M. Post-translational methyla-tion of high mobility group box 1 (HMGB1) causes its cy-toplasmic localization in neutrophils. J Biol Chem 2007; 282: 16336-16344
8 Andersson U, Wang H, Palmblad K, Aveberger AC, Bloom O, Erlandsson-Harris H, Janson A, Kokkola R, Zhang M, Yang H, Tracey KJ. High mobility group 1 protein (HMG-1) stimulates proinflammatory cytokine synthesis in human monocytes. J Exp Med 2000; 192: 565-570
9 Raucci A, Palumbo R, Bianchi ME. HMGB1: a signal of ne-crosis. Autoimmunity 2007; 40: 285-289
10 Mitola S, Belleri M, Urbinati C, Coltrini D, Sparatore B, Pe-drazzi M, Melloni E, Presta M. Cutting edge: extracellular high mobility group box-1 protein is a proangiogenic cyto-
4097 September 28, 2011|Volume 17|Issue 36|WJG|www.wjgnet.com
in liver grafts or by transforming HSCs into myofibro-blasts. In contrast, neutralization with an anti-HMGB1 antibody may be a therapeutic modality to prevent fibro-genesis in post-OLT liver grafts[33].
siRNA has become a powerful tool for functional ge-netic studies and gene therapy in mammals[34,35]. Although gene knockdown by siRNA is highly effective, the off-target effect of siRNA may represent a major obstacle for therapeutic applications. However, the potential off-target effects could be minimized by choosing an siRNA with maximal sequence divergence from the list of genes with partial sequence identity to the intended mRNA tar-get[36]. Software was used to choose a maximal sequence identity of HMGB1 siRNA, and three siRNA sequences were designed. In preliminary experiments, we identified the fact that shRNAH3 had certain interference effects. Our results show that this sequence was more efficient in reducing the HMGB1 transcript levels.
In the present study, we found that after HMGB1 was downregulated in HSCs by siRNA, there was an inhibi-tory effect on the mRNA levels of α-SMA and collagen types Ⅰ and Ⅲ, suggesting that inhibition of HMGB1 could directly result in suppression of HSC activation and collagen production. We also discovered that HMGB1 siRNA prohibited HSC proliferation, and a cell cycle analysis revealed that downregulation of HMGB1 arrest-ed cells at the G0/G1 phase, which confirmed the effect of HMGB1 on cell proliferation; however, the definitive mechanism responsible is still uncertain because HMGB1 is multifunctional and has multiple molecular interactions.
In conclusion, HMGB1 was upregulated during liver fibrogenesis, and downregulating HMGB1 expression in HSCs by siRNA prohibited the activity of HSCs and collagen synthesis and enhanced collagen degradation. The results of our study indicate a significant functional role for HMGB1 in the development of liver fibrosis, and downregulating HMGB1 expression with siRNA could be an effective way to treat liver fibrosis.
COMMENTSBackgroundHepatic fibrosis is a response to injury in the liver. It is characterized by both a quantitative and qualitative change in the extracellular matrix (ECM). The activated hepatic stellate cell (HSC) is primarily responsible for excessive ECM deposition during liver fibrosis. It has been shown that high-mobility group box 1 (HMGB1) expression is up regulated during myofibroblast cellular activation and involved in the pathogenesis of hepatic fibrosis. This suggests that HMGB1 is a promising molecular target for hepatic fibrosis gene therapy. Inhibition of abnor-mal expression of HMGB1 may be an effective strategy for biological therapy of hepatic fibrosis.Research frontiersHMGB1 is a major component of mammalian chromatin endowed with an architectural function. Increasing evidence now points to multiple functions of HMGB1 in infection, tissue injury, inflammation, apoptosis and the immune response. It has been reported in several liver diseases, including hepatitis, hepatic ischemia, and orthotopic liver transplantation. HMGB1 has been impli-cated in the pathogenesis of several liver diseases, including Con-A-induced hepatitis, hepatic ischemia, and orthotopic liver transplantation. However, the role of HMGB1 and how to inhibit its expressiong in hepatic fibrosis has yet to be fully elucidated. In this study, the authors demonstrate that the overexpres-
COMMENTS
Ge WS et al. Inhibition of HMGB1 expression in HSC

kine. J Immunol 2006; 176: 12-1511 Chen SW, Chen YX, Zhang XR, Qian H, Chen WZ, Xie WF.
Targeted inhibition of platelet-derived growth factor recep-tor-beta subunit in hepatic stellate cells ameliorates hepatic fibrosis in rats. Gene Ther 2008; 15: 1424-1435
12 Bataller R, Brenner DA. Liver fibrosis. J Clin Invest 2005; 115: 209-218
13 Friedman SL. Mechanisms of hepatic fibrogenesis. Gastroen-terology 2008; 134: 1655-1669
14 Hu YB, Li DG, Lu HM. Modified synthetic siRNA targeting tissue inhibitor of metalloproteinase-2 inhibits hepatic fibro-genesis in rats. J Gene Med 2007; 9: 217-229
15 Uchinami H, Seki E, Brenner DA, D’Armiento J. Loss of MMP 13 attenuates murine hepatic injury and fibrosis dur-ing cholestasis. Hepatology 2006; 44: 420-429
16 Roderfeld M, Weiskirchen R, Wagner S, Berres ML, Henkel C, Grötzinger J, Gressner AM, Matern S, Roeb E. Inhibition of hepatic fibrogenesis by matrix metalloproteinase-9 mu-tants in mice. FASEB J 2006; 20: 444-454
17 González-Cuevas J, Bueno-Topete M, Armendariz-Borunda J. Urokinase plasminogen activator stimulates function of active forms of stromelysin and gelatinases (MMP-2 and MMP-9) in cirrhotic tissue. J Gastroenterol Hepatol 2006; 21: 1544-1554
18 Gong Q, Zhang H, Li JH, Duan LH, Zhong S, Kong XL, Zheng F, Tan Z, Xiong P, Chen G, Fang M, Gong FL. High-mobility group box 1 exacerbates concanavalin A-induced hepatic injury in mice. J Mol Med (Berl) 2010; 88: 1289-1298
19 Nakano T, Goto S, Lai CY, Hsu LW, Kao YH, Lin YC, Kawamoto S, Chiang KC, Ohmori N, Goto T, Sato S, Jawan B, Cheng YF, Ono K, Chen CL. Experimental and clinical sig-nificance of antinuclear antibodies in liver transplantation. Transplantation 2007; 83: 1122-1125
20 Rendon-Mitchell B, Ochani M, Li J, Han J, Wang H, Yang H, Susarla S, Czura C, Mitchell RA, Chen G, Sama AE, Tracey KJ, Wang H. IFN-gamma induces high mobility group box 1 protein release partly through a TNF-dependent mecha-nism. J Immunol 2003; 170: 3890-3897
21 Wang H, Vishnubhakat JM, Bloom O, Zhang M, Ombrellino M, Sama A, Tracey KJ. Proinflammatory cytokines (tumor necrosis factor and interleukin 1) stimulate release of high mobility group protein-1 by pituicytes. Surgery 1999; 126: 389-392
22 Palumbo R, Sampaolesi M, De Marchis F, Tonlorenzi R, Co-lombetti S, Mondino A, Cossu G, Bianchi ME. Extracellular HMGB1, a signal of tissue damage, induces mesoangioblast migration and proliferation. J Cell Biol 2004; 164: 441-449
23 Bonaldi T, Talamo F, Scaffidi P, Ferrera D, Porto A, Bachi A, Rubartelli A, Agresti A, Bianchi ME. Monocytic cells hyper-acetylate chromatin protein HMGB1 to redirect it towards secretion. EMBO J 2003; 22: 5551-5560
24 Gardella S, Andrei C, Ferrera D, Lotti LV, Torrisi MR, Bianchi ME, Rubartelli A. The nuclear protein HMGB1 is secreted by monocytes via a non-classical, vesicle-mediated secretory pathway. EMBO Rep 2002; 3: 995-1001
25 Dumitriu IE, Baruah P, Valentinis B, Voll RE, Herrmann M, Nawroth PP, Arnold B, Bianchi ME, Manfredi AA, Rovere-Querini P. Release of high mobility group box 1 by dendritic cells controls T cell activation via the receptor for advanced glycation end products. J Immunol 2005; 174: 7506-7515
26 Yang D, Chen Q, Yang H, Tracey KJ, Bustin M, Oppenheim JJ. High mobility group box-1 protein induces the migration and activation of human dendritic cells and acts as an alar-min. J Leukoc Biol 2007; 81: 59-66
27 Hori O, Brett J, Slattery T, Cao R, Zhang J, Chen JX, Na-gashima M, Lundh ER, Vijay S, Nitecki D. The receptor for advanced glycation end products (RAGE) is a cellular bind-ing site for amphoterin. Mediation of neurite outgrowth and co-expression of rage and amphoterin in the develop-ing nervous system. J Biol Chem 1995; 270: 25752-25761
28 Park JS, Svetkauskaite D, He Q, Kim JY, Strassheim D, Ishizaka A, Abraham E. Involvement of toll-like receptors 2 and 4 in cellular activation by high mobility group box 1 protein. J Biol Chem 2004; 279: 7370-7377
29 Fehrenbach H, Weiskirchen R, Kasper M, Gressner AM. Up-regulated expression of the receptor for advanced glyca-tion end products in cultured rat hepatic stellate cells dur-ing transdifferentiation to myofibroblasts. Hepatology 2001; 34: 943-952
30 Yang H, Hreggvidsdottir HS, Palmblad K, Wang H, Ochani M, Li J, Lu B, Chavan S, Rosas-Ballina M, Al-Abed Y, Akira S, Bierhaus A, Erlandsson-Harris H, Andersson U, Tracey KJ. A critical cysteine is required for HMGB1 binding to Toll-like receptor 4 and activation of macrophage cytokine release. Proc Natl Acad Sci USA 2010; 107: 11942-11947
31 Han J, Zhong J, Wei W, Wang Y, Huang Y, Yang P, Purohit S, Dong Z, Wang MH, She JX, Gong F, Stern DM, Wang CY. Extracellular high-mobility group box 1 acts as an innate immune mediator to enhance autoimmune progression and diabetes onset in NOD mice. Diabetes 2008; 57: 2118-2127
32 Tsung A, Klune JR, Zhang X, Jeyabalan G, Cao Z, Peng X, Stolz DB, Geller DA, Rosengart MR, Billiar TR. HMGB1 re-lease induced by liver ischemia involves Toll-like receptor 4 dependent reactive oxygen species production and calcium-mediated signaling. J Exp Med 2007; 204: 2913-2923
33 Kao YH, Jawan B, Goto S, Hung CT, Lin YC, Nakano T, Hsu LW, Lai CY, Tai MH, Chen CL. High-mobility group box 1 protein activates hepatic stellate cells in vitro. Transplant Proc 2008; 40: 2704-2705
34 Jackson AL, Burchard J, Schelter J, Chau BN, Cleary M, Lim L, Linsley PS. Widespread siRNA “off-target” transcript silencing mediated by seed region sequence complementar-ity. RNA 2006; 12: 1179-1187
35 Tschuch C, Schulz A, Pscherer A, Werft W, Benner A, Hotz-Wagenblatt A, Barrionuevo LS, Lichter P, Mertens D. Off-target effects of siRNA specific for GFP. BMC Mol Biol 2008; 9: 60
36 De Paula D, Bentley MV, Mahato RI. Hydrophobization and bioconjugation for enhanced siRNA delivery and tar-geting. RNA 2007; 13: 431-456
S- Editor Tian L L- Editor Kerr C E- Editor Li JY
4098 September 28, 2011|Volume 17|Issue 36|WJG|www.wjgnet.com
Ge WS et al. Inhibition of HMGB1 expression in HSC


















