
In vitro antioxidant activity of a peptide isolated from Nile tilapia (Oreochromis niloticus) scale...
11
In vitro antioxidant activity of a peptide isolated from Nile tilapia (Oreochromis niloticus) scale gelatin in free radical-mediated oxidative systems Dai-Hung Ngo a , Zhong-Ji Qian b , BoMi Ryu a , Jae W. Park c , Se-Kwon Kim a,b, * a Department of Chemistry, Pukyong National University, Busan 608-737, Republic of Korea b Marine Bioprocess Research Center, Pukyong National University, Busan 608-737, Republic of Korea c Food Science and Technology, Seafood Research and Education Center, 2001 Marine Dr., Rm 253 Astoria, OR 97103, USA ARTICLE INFO Article history: Received 18 December 2009 Received in revised form 26 January 2010 Accepted 3 February 2010 Available online 2 March 2010 Keywords: Nile tilapia scale Gelatin hydrolysates Free radical scavenging Antioxidative peptide Electron spin resonance RAW 264.7 cells ABSTRACT In the present study, a peptide possessing antioxidant properties was isolated from Nile tilapia (Oreochromis niloticus) scale gelatin. Gelatin protein was hydrolyzed using alcalase, pronase E, trypsin and pepsin. Antioxidant efficacy of respective hydrolysates were evalu- ated using 1,1-diphenyl-2-picrylhydrazyl (DPPH) radical, hydroxyl radical and superoxide radical anion scavenging activities. Moreover, protective effect on DNA damage caused by hydroxyl radicals generated was determined. Further, the level of reactive oxygen spe- cies (ROS) was determined using a fluorescence probe, 2 0 ,7 0 -dichlorofluorescin diacetate (DCFH-DA), which could be converted to highly fluorescent dichlorofluorescein (DCF) with the presence of intracellular ROS on mouse macrophages, RAW 264.7 cells. Among hydrol- ysates, alcalase-derived hydrolysate exhibited the highest antioxidant activity compared to other enzymatic hydrolysates. Therefore, it was further analyzed and the sequence of an active peptide present in it was identified as Asp-Pro-Ala-Leu-Ala-Thr-Glu-Pro-Asp-Pro- Met-Pro-Phe (1382.57 Da). This peptide showed no cytotoxic effect on mouse macrophages (RAW 264.7) and human lung fibroblasts (MRC-5). In addition, it scavenged hydroxyl, DPPH and superoxide radicals at the IC 50 values of 7.56, 8.82 and 17.83 lM, respectively. These results suggest that the peptide derived from Nile tilapia (O. niloticus) scale gelatin acts as a candidate against oxidative stress and could be used as a potential functional food ingredient. Ó 2010 Elsevier Ltd. All rights reserved. 1. Introduction Oxidative stress and antioxidants are widely discussed in many current research endeavours. Oxidative stress responsi- ble for injuries to all the important cellular components, thus reactive oxygen species (ROS) generated excessively in tissues can cause cell death. Besides, ROS have direct and indirect relationships with oxidation of cellular biomolecules and are involved in many health disorders such as neurodegener- ative, hypertension, inflammation, diabetes, cancer and aging (Calabrese et al., 2005; Je et al., 2004). Accordingly, many stud- ies have been carried out to find new antioxidant compounds from natural resources such as plants (Choi et al., 2002), ani- mals (Kim & Je, 2007), microorganisms (Ke et al., 2009; Kalog- eropoulos et al., 2009) and foods (Borneo et al., 2009) in order to inhibit oxidation with its deleterious effects. 1756-4646/$ - see front matter Ó 2010 Elsevier Ltd. All rights reserved. doi:10.1016/j.jff.2010.02.001 * Corresponding author: Address: Marine Bioprocess Research Center, Pukyong National University, Busan 608-737, Republic of Korea. Tel.: +82 51 629 7094; fax: +82 51 629 7099. E-mail address: [email protected] (S.-K. Kim). JOURNAL OF FUNCTIONAL FOODS 2 (2010) 107 – 117 available at www.sciencedirect.com journal homepage: www.elsevier.com/locate/jff
-
Upload
dai-hung-ngo -
Category
Documents
-
view
217 -
download
14
Transcript of In vitro antioxidant activity of a peptide isolated from Nile tilapia (Oreochromis niloticus) scale...

J O U R N A L O F F U N C T I O N A L F O O D S 2 ( 2 0 1 0 ) 1 0 7 – 1 1 7
. sc iencedi rec t . com
ava i lab le a t wwwjournal homepage: www.elsevier .com/ locate / j f f
In vitro antioxidant activity of a peptide isolated fromNile tilapia (Oreochromis niloticus) scale gelatin in freeradical-mediated oxidative systems
Dai-Hung Ngoa, Zhong-Ji Qianb, BoMi Ryua, Jae W. Parkc, Se-Kwon Kima,b,*
aDepartment of Chemistry, Pukyong National University, Busan 608-737, Republic of KoreabMarine Bioprocess Research Center, Pukyong National University, Busan 608-737, Republic of KoreacFood Science and Technology, Seafood Research and Education Center, 2001 Marine Dr., Rm 253 Astoria, OR 97103, USA
A R T I C L E I N F O
Article history:
Received 18 December 2009
Received in revised form
26 January 2010
Accepted 3 February 2010
Available online 2 March 2010
Keywords:
Nile tilapia scale
Gelatin hydrolysates
Free radical scavenging
Antioxidative peptide
Electron spin resonance
RAW 264.7 cells
1756-4646/$ - see front matter � 2010 Elsevidoi:10.1016/j.jff.2010.02.001
* Corresponding author: Address: Marine BioTel.: +82 51 629 7094; fax: +82 51 629 7099.
E-mail address: [email protected] (S.-K
A B S T R A C T
In the present study, a peptide possessing antioxidant properties was isolated from Nile
tilapia (Oreochromis niloticus) scale gelatin. Gelatin protein was hydrolyzed using alcalase,
pronase E, trypsin and pepsin. Antioxidant efficacy of respective hydrolysates were evalu-
ated using 1,1-diphenyl-2-picrylhydrazyl (DPPH) radical, hydroxyl radical and superoxide
radical anion scavenging activities. Moreover, protective effect on DNA damage caused
by hydroxyl radicals generated was determined. Further, the level of reactive oxygen spe-
cies (ROS) was determined using a fluorescence probe, 2 0,7 0-dichlorofluorescin diacetate
(DCFH-DA), which could be converted to highly fluorescent dichlorofluorescein (DCF) with
the presence of intracellular ROS on mouse macrophages, RAW 264.7 cells. Among hydrol-
ysates, alcalase-derived hydrolysate exhibited the highest antioxidant activity compared to
other enzymatic hydrolysates. Therefore, it was further analyzed and the sequence of an
active peptide present in it was identified as Asp-Pro-Ala-Leu-Ala-Thr-Glu-Pro-Asp-Pro-
Met-Pro-Phe (1382.57 Da). This peptide showed no cytotoxic effect on mouse macrophages
(RAW 264.7) and human lung fibroblasts (MRC-5). In addition, it scavenged hydroxyl, DPPH
and superoxide radicals at the IC50 values of 7.56, 8.82 and 17.83 lM, respectively. These
results suggest that the peptide derived from Nile tilapia (O. niloticus) scale gelatin acts as
a candidate against oxidative stress and could be used as a potential functional food
ingredient.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
Oxidative stress and antioxidants are widely discussed in
many current research endeavours. Oxidative stress responsi-
ble for injuries to all the important cellular components, thus
reactive oxygen species (ROS) generated excessively in tissues
can cause cell death. Besides, ROS have direct and indirect
relationships with oxidation of cellular biomolecules and
er Ltd. All rights reserved
process Research Center
. Kim).
are involved in many health disorders such as neurodegener-
ative, hypertension, inflammation, diabetes, cancer and aging
(Calabrese et al., 2005; Je et al., 2004). Accordingly, many stud-
ies have been carried out to find new antioxidant compounds
from natural resources such as plants (Choi et al., 2002), ani-
mals (Kim & Je, 2007), microorganisms (Ke et al., 2009; Kalog-
eropoulos et al., 2009) and foods (Borneo et al., 2009) in order
to inhibit oxidation with its deleterious effects.
.
, Pukyong National University, Busan 608-737, Republic of Korea.

108 J O U R N A L O F F U N C T I O N A L F O O D S 2 ( 2 0 1 0 ) 1 0 7 – 1 1 7
Some fish gelatin hydrolysates have been found to have
noticeable antioxidant, antihypertensive, anticancer, and
antimicrobial as well as immuno-modulatory and choles-
terol-lowering effects (Kim et al., 2001a,b; Mendis et al.,
2005a,b). Recently, a number of research works have shown
that peptides derived from various fish gelatin hydrolysates
act as potential antioxidants. Among these, fish gelatin ex-
tracted from lumpfish (Osborne et al., 1990), conger eel
(Kim & Cho, 1996), cod (Gudmundsson & Hafsteinsson,
1997), shark (Yoshimura et al., 2000), megrim (Sarabia
et al., 2001), tilapia (Jamilah & Harvinder, 2002), pollack
(Je et al., 2005) and tuna (Cho et al., 2005) have widely been
studied.
Tilapia is an important species in freshwater aquaculture
(Fitzsimmons, 2004). It is the third most widely cultured
fish, after carp and salmonids (El-Sayed, 2006). The produc-
tion of farmed tilapia increased from 383,654 metric tonnes
in 1990 to 1,505,804 metric tonnes in 2002. The value of
farmed tilapia also increased greatly, rising from US$154
million in 1984 to US$1800 million in 2002. The major
farmed-tilapia producing Asian countries are Korea, China,
Indonesia, the Philippines, Taiwan, and Thailand (El-Sayed,
2006). Gelatins are produced on a large scale from skins
and bones of mammalians. However, the skins and bones
of fish with high collagen content are by-products of the fil-
leting process and can also be used to produce gelatin (Na-
gai and Suzuki, 2000). Here, we report the purification and
characterization of an antioxidative peptide derived from
enzymatic hydrolysis of Nile tilapia (Oreochromis niloticus)
scale gelatin and assessment of its antioxidant properties
based on in vitro antioxidant activity in free radical-medi-
ated oxidative systems.
2. Materials and methods
2.1. Materials
Gelatin was obtained from Geltech Co., Busan, South Korea.
The fluorescence probes 2 0,7 0-dichlorofluorescin diacetate
(DCFH-DA) was obtained from Molecular Probes Inc. (Eugene,
OR, USA). MTT (3-(4,5-dimethyl-2-yl)-2,5-diphenyltetrazolium
bromide), agarose, foetal bovine serum (FBS), pronase E, pep-
sin and trypsin were purchased from Sigma Chemical Co. (St.
Louis, MO, USA). Alcalase was purchased from Novozymes
Co. (Bagsvaerd, Denmark).
Radical testing chemicals, including 1,1-diphenyl-2-pic-
rylhydrazyl (DPPH), 5,5-dimethyl-1-pyrroline N-oxide (DMPO),
FeSO4, H2O2, riboflavin and ethylenediaminetetraacetic acid
(EDTA) were purchased from Sigma Chemical Co. (St. Louis,
MO, USA).
Mouse macrophages (RAW 264.7) and human lung fibro-
blast (MRC-5) cell lines were obtained from American Type
Culture Collection (Manassas, VA, USA). Dulbecco’s Modified
Eagle Medium (DMEM), penicillin/streptomycin, and the other
materials required for culturing of cells were purchased from
Gibco BRL, Life Technologies (Grand Island, NY, USA). All
other reagents were of the highest grade available
commercially.
2.2. Preparation of enzymatic hydrolysates
To extract bioactive peptides from Nile tilapia (O. niloticus)
scale gelatin, enzymatic hydrolysis was performed using var-
ious commercial enzymes (alcalase, pronase E, pepsin, and
trypsin) under optimal conditions. At enzyme to substrate ra-
tio of 1/100 (w/w), 1% substrate and enzyme were mixed. The
mixture was incubated for 4 h at each optimal temperature
with stirring and then heated in a boiling water bath for
10 min to inactivate the enzyme. Lyophilized hydrolysates
were stored at �80 �C until used.
2.3. Electron spin resonance (ESR) spectrometric assays
2.3.1. Scavenging effect on DPPH radicalDPPH radical scavenging activity was measured using the
method described by Nanjo et al. (1996). A 30 ll sample solu-
tion (or distilled water itself as control) was added to 30 ll of
DPPH (60 lM) in methanol solution. After mixing vigorously
for 10 s, the solution was then transferred into a 50 ll quartz
capillary tube, and the scavenging activity of hydrolysate on
DPPH radical was measured using a JES-FA ESR spectrometer
(JEOL Ltd., Tokyo, Japan). The spin adduct was measured on
an ESR spectrometer exactly 2 min later. Spectrometer set-
tings were: magnetic field, 336.5 ± 5 mT; power, 5 mW; modu-
lation frequency, 9.41 GHz; amplitude, 1 · 1000; modulation
width, 0.8 mT; sweep width, 10 mT; sweep time, 30 s. DPPH
radical scavenging ability was calculated following equation
in which H and H0 were relative peak height of radical signals
with and without sample, respectively.
Radical scavenging activity ð%Þ ¼ ðH0 �HÞ=H0 � 100%
2.3.2. Hydroxyl radicals scavenging activityHydroxyl radicals were generated by Fenton reaction and
trapped using DMPO nitrone spin trap (Rosen & Rauckman,
1984). The resultant DMPO-OH adducts was detectable with
an ESR spectrometer. The sample solution (15 ll) was mixed
with DMPO (0.3 M, 15 ll), FeSO4 (10 mM, 15 ll) and H2O2
(10 mM, 15 ll) in a phosphate buffer solution (pH 7.4), and
then transferred into a 50 ll quartz capillary tube. After
2.5 min, the ESR spectrum was recorded using an ESR spec-
trometer. Spectrometer settings were: magnetic field,
336.5 ± 5 mT; power, 1 mW; modulation frequency, 9.41 GHz;
amplitude, 1 · 200; modulation width, 0.1 mT; sweep width,
10 mT; sweep time, 30 s. Hydroxyl radical scavenging ability
was calculated following equation in which H and H0 were rel-
ative peak height of radical signals with and without sample,
respectively.
Radical scavenging activity ð%Þ ¼ ðH0 �HÞ=H0 � 100%
2.3.3. Superoxide radical anion scavenging activitySuperoxide radical anions were generated by UV irradiated
riboflavin/EDTA system (Guo et al., 1999). The reaction mix-
ture containing 0.3 mM riboflavin, 1.6 mM EDTA, 800 mM
DMPO and indicated concentration of sample solution was
irradiated for 1 min under UV lamp at 365 nm. The reaction
mixture was transferred to 50 ll quartz capillary tube of the
ESR spectrometer for measurement. Spectrometer settings

J O U R N A L O F F U N C T I O N A L F O O D S 2 ( 2 0 1 0 ) 1 0 7 – 1 1 7 109
were: magnetic field, 336.5 ± 5 mT; power, 10 mW; modulation
frequency, 9.41 GHz; amplitude, 1 · 1000; modulation width,
0.1 mT; sweep width, 10 mT; sweep time, 1 min. Superoxide
radical scavenging ability was calculated following equation
in which H and H0 were relative peak height of radical signals
with and without sample, respectively.
Radical scavenging activity ð%Þ ¼ ðH0 �HÞ=H0 � 100%
2.4. Cell culture and viability determination
MRC-5 cells and RAW 264.7 cells were grown in DMEM media
containing 5% (v/v) FBS, 100 lg/ml penicillin–streptomycin
and 5% CO2 at 37 �C.
Cytotoxicity levels of samples on cells were measured
using the MTT method as described by Hansen et al. (1989).
The cells were grown in 96-well plates at a density of 5 · 103
cells/well. After 24 h, cells were washed with fresh medium
and treated with different concentrations of samples. After
48 h of incubation, cells were rewashed and 50 ll of MTT
(5 mg/ml) were added and incubated for 4 h. Finally, 200 ll
of DMSO were added to solubilize the formazan salt formed
and the amount of formazan salt was determined by measur-
ing the optical density (OD) at 540 nm using an GENios micro-
plate reader (TECAN Austria GmbH, Grodig/Salzburg, Austria).
2.5. Determination of intracellular formation of ROS usingDCFH-DA labeling
Intracellular formation of ROS was assessed according to a
method described previously by employing oxidation sensi-
tive dye DCFH-DA as the substrate (Engelmann et al., 2005).
RAW 264.7 cells were grown in black microtiter 96-well plates
and were labeled with 20 lM DCFH-DA in HBSS (Hanks bal-
anced salt solution) and kept for 20 min in the dark. The
non-fluorescent DCFH-DA dye which freely penetrates into
cells was then hydrolyzed by intracellular esterase to 2 0,7 0-
dichlorodihydrofluorescein (DCFH), and trapped inside the
cells. Cells were then treated with different concentrations
of sample and incubated for 1 h. After washing cells for three
times with PBS, 500 lM H2O2 (in HBSS) were added. The for-
mation of fluorescent dichlorofluorescein (DCF) due to oxida-
tion of DCFH in the presence of various ROS was read after
every 30 min at the excitation wavelength of 485 nm and
the emission wavelength of 528 nm (GENios microplate read-
er, Tecan Austria GmbH, Grodig/Salzburg, Austria). Dose-
dependent and time-dependent effects of treatments were
plotted and compared with fluorescence intensity of the con-
trol group in which samples were not treated.
2.6. Genomic DNA isolation
Genomic high molecular weight DNA was extracted from
RAW 264.7 cells using standard phenol/proteinase K proce-
dure with slight modifications (Sambrook & Russell, 2001).
Briefly, cells cultured in 10 cm culture dishes were washed
twice with PBS and scraped into 1 ml of PBS containing
10 mM EDTA. After centrifugation cells were dissolved in
RNase (0.03 mg/ml), NaOAC (0.175 M), proteinase K (0.25 mg/
ml) and SDS (0.6%). The mixture was then incubated for
30 min at 37 �C and 1 h at 55 �C. Following incubation, phe-
nol/chloroform/isoamyl alcohol was added at a 1:1 (v/v) ratio
and the mixture was centrifuged at 6000g for 5 min at 4 �C.
Following centrifugation, the supernatant was mixed with
100% cold ethanol at a 1:1.5 (v/v) ratio and kept for 15 min
at �20 �C. After centrifugation at 12,000g for 5 min, the pellet
was dissolved in 10 mM TE buffer (pH 7.5) and the purity of
DNA was spectrophotometrically determined at 260/280 nm.
Further, the quality of isolated DNA was evaluated using 1%
agarose gel electrophoresis in 0.04 M Tris–acetate, 0.001 M
EDTA buffer (pH 7.5).
2.7. Determination of radical-mediated DNA damage
Hydrogen peroxide mediated DNA oxidation was determined
according to Milne et al. (1993). Briefly, 40 ll of DNA reaction
mixture were prepared by adding pre-determined concentra-
tions of test sample (or same volume of distilled water as con-
trol), 100 lM final concentration of FeSO4, 0.1 mM final
concentration of H2O2 and 5 ll of isolated genomic DNA in
the same order. Then the mixture was incubated at room
temperature for 10 min and the reaction was terminated by
adding 10 mM final concentration of EDTA. An Aliquot
(20 ll) of the reaction mixture containing about 1 lg of DNA
was electrophoresed on a 1% agarose gel for 40 min at 100 V.
Gels were then stained with 1 mg/ml ethidium bromide and
visualized under UV light using AlphaEase gel image analysis
software (Alpha Innotech, St. San Leandro, CA, USA).
2.8. Purification of antioxidant peptide
2.8.1. Ion exchange chromatographyThe antioxidant peptide was purified from enzymatic hydrol-
ysates using fast protein liquid chromatography (FPLC AKTA,
Amersham Bioscience Co., Uppsala, Sweden) on a HiPrep 16/
10 DEAE FF ion-exchange column (1.6 · 10 cm, Amersham
Biosciences, Piscataway, NJ, USA). The hydrolysate was loaded
onto a HiPrep 16/10 DEAE FF ion-exchange column equili-
brated with a 20 mM sodium acetate buffer (pH 4.0), and
eluted with a linear gradient of NaCl (0–2 M) in the same buf-
fer at a flow rate of 62 ml/h. Each fraction was monitored at
280 nm, collected at a volume of 4 ml and concentrated using
a rotary evaporator; antioxidant activity was also determined.
The strong antioxidant fraction was lyophilized, and ion ex-
change chromatography was used as the next step.
2.8.2. High performance liquid chromatography (HPLC)The fraction exhibiting antioxidant activity was further puri-
fied using reversed-phase high performance liquid chroma-
tography (RP-HPLC, Dionex Korea Ltd., Sunnyvale, CA, USA)
on a Primesphere 10 C18 (10 mm · 250 mm, Phenomenex,
Cheshire, England) column with a linear gradient of acetoni-
trile (0–40% in 40 min) containing 0.1% trifluoroacetic acid
(TFA) at a flow rate of 2 ml/min. Elution peaks were detected
at 215 nm, and active peak was concentrated using a rotary
evaporator. Active peak (target peak, data not shown) was
pooled and lyophilized immediately. The active fraction from
analytical column was further applied onto a SynChropak RP-
P-100 column (4.6 mm · 250 mm, SynChrom, Inc., Lafayette,
Indiana, USA) with a linear gradient of acetonitrile (15%, v/v,

110 J O U R N A L O F F U N C T I O N A L F O O D S 2 ( 2 0 1 0 ) 1 0 7 – 1 1 7
in 20 min) containing 0.1% trifluoroacetic acid (TFA) at flow
rate of 1.2 ml/min. Finally, the purified peptide from alca-
lase-digest of Nile tilapia (O. niloticus) scale gelatin was ana-
lyzed for its amino acid sequence.
2.9. Determination of amino acid sequence
An accurate molecular mass and amino acid sequence of the
purified peptide was determined using a Q-TOF mass spec-
trometer (Micromass, Altrincham, UK) coupled with an elec-
trospray ionization (ESI) source. The purified peptide was
separately infused into the electrospray source after being
dissolved in methanol/water (1:1, v/v), and its molecular mass
was determined by the doubly charged (M + 2H)+2 state in the
mass spectrum. Following molecular mass determination,
the peptide was automatically selected for fragmentation,
and sequence information was obtained by tandem MS
analysis.
3. Results and discussion
3.1. Preparation of gelatin hydrolysates and theirantioxidant properties
Gelatin was sequentially hydrolyzed with various enzymes
such as alcalase, pronase E, pepsin and trypsin. The antiox-
idative activity of a substance may be distinguished exactly
by testing scavenging activities on free radicals in oxidative
systems. Consequently, hydrolysates were evaluated for
their free radical scavenging effects on DPPH, hydroxyl
and superoxide radicals using electron spin resonance tech-
nique. As shown in Table 1, scavenging of hydroxyl radicals
was more efficient than that of DPPH and superoxide radi-
cals. Among hydrolysates, the alcalase-derived hydrolysate
exhibited the highest antioxidant activity compared to
other enzymatic hydrolysates. The IC50 (concentration of
hydrolysate to scavenge 50% radical activity) values of alca-
lase hydrolysate on scavenging DPPH, hydroxyl and super-
oxide radicals were found to be 660 ± 0.40, 263 ± 0.31 and
720 ± 0.35 lg/ml, respectively. The hydrolytic reaction de-
pends on the availability of susceptible peptide bonds on
which the enzyme attack is concentrated, as well as the
physical structure of the protein molecules (Diniz and Mar-
tin, 1998). Alcalase is an alkaline protease industrially used
and produced from Bacillus licheniformis, in which one of the
Table 1 – The IC50 values of hydrolysates to scavengeDPPH, hydroxyl and superoxide radicals.
Hydrolysate IC50 (lg/ml)a
DPPH Hydroxyl Superoxide
Alcalase 660 ± 0.40 263 ± 0.31 720 ± 0.350
Pronase E 1020 ± 0.13 500 ± 0.05 740 ± 0.002
Trypsin 860 ± 0.42 320 ± 0.30 720 ± 0.440
Pepsin 920 ± 0.24 420 ± 0.12 1150 ± 0.500
a Concentration of hydrolysate required to inhibit 50% of the
respective radical ESR signal intensity.
amino acids at the active site is serine. It is capable of pro-
ducing bioactive peptides when it is used to hydrolyze nat-
ural proteins (Hyun and Shin, 2000), with a rather broad
range of specificity for peptide bonds, leading to the pro-
duction of antioxidant peptides (Chand, 2009; Nieto et al.,
2009). Bioactive peptides produced by alcalase are resistant
to digestive enzymes such as pepsin, trypsin and pronase
E, which would be absorbed when present in this sort of
hydrolysate (Park et al., 2001; Saito et al., 1994). Moreover,
related studies have clearly shown that alcalase produces
shorter peptide sequences as well as terminal amino acid
sequences responsible for various bioactives, including anti-
oxidant activity.
3.2. Effect of gelatin hydrolysates on cell viability of MRC-5and RAW 264.7 cells
In this study, MRC-5 and RAW 264.7 cells were treated with
different concentrations of gelatin hydrolysates to deter-
mine non-cytotoxic concentration for further experiments.
The cell viability data confirmed that gelatin hydrolysates
were non-cytotoxic on both RAW 264.7 cells (Fig. 1A) and
MRC-5 cells (Fig. 1B). Therefore, it was determined that
these gelatin hydrolysates could be used in further
experiments.
3.3. Cellular radical scavenging effect and DNA protectionactivity of gelatin hydrolysate
The gelatin hydrolysates from other enzymes (alcalase, pron-
ase E, trypsin and pepsin) exhibited a free radical scavenging
effect even only after 30 min of incubation compared to the
control (H2O2 treatment and non-gelatin hydrolysate) as
shown in Fig. 2A. Among hydrolysates, the alcalase-derived
hydrolysate exhibited the highest free radical scavenging ef-
fect against that of other enzymatic hydrolysates. Our results
showed that their antioxidant activity were different with
various concentrations of gelatin hydrolysate (100, 200,
1000 lg/ml) and other enzymatic hydrolysates. The DNA
damage at all tested concentrations of gelatin hydrolysate
with alcalase were inhibited by about 80% based on the inten-
sity of DNA bands as shown in Fig. 2B. Therefore, we could
confirm that the alcalase-derived hydrolysate scavenged free
radical in a dose- and time-dependent manner in RAW 264.7
cells.
3.4. Purification of antioxidant peptide
Considering the free radical scavenging activities, alcalase-
derived hydrolysate was employed for the purification and
identification of an antioxidative peptide. Firstly, alcalase-
derived hydrolysate was dissolved in sodium acetate buffer
(20 mM, pH 4.0) at 20 mg/ml, and loaded onto a HiPrep 16/
10 DEAE FF ion-exchange column using FPLC with a linear
gradient of NaCl (0–2 M). The elution peaks were monitored
at 280 nm, fractionated into three portions and their antiox-
idant activities measured using their potency to scavenge
free radicals (Fig. 3A). Fraction 3 quenched hydroxyl, DPPH
and superoxide radicals by 72.04%, 42.80% and 18.03%,
respectively. Fraction 3, which had a strong free radical
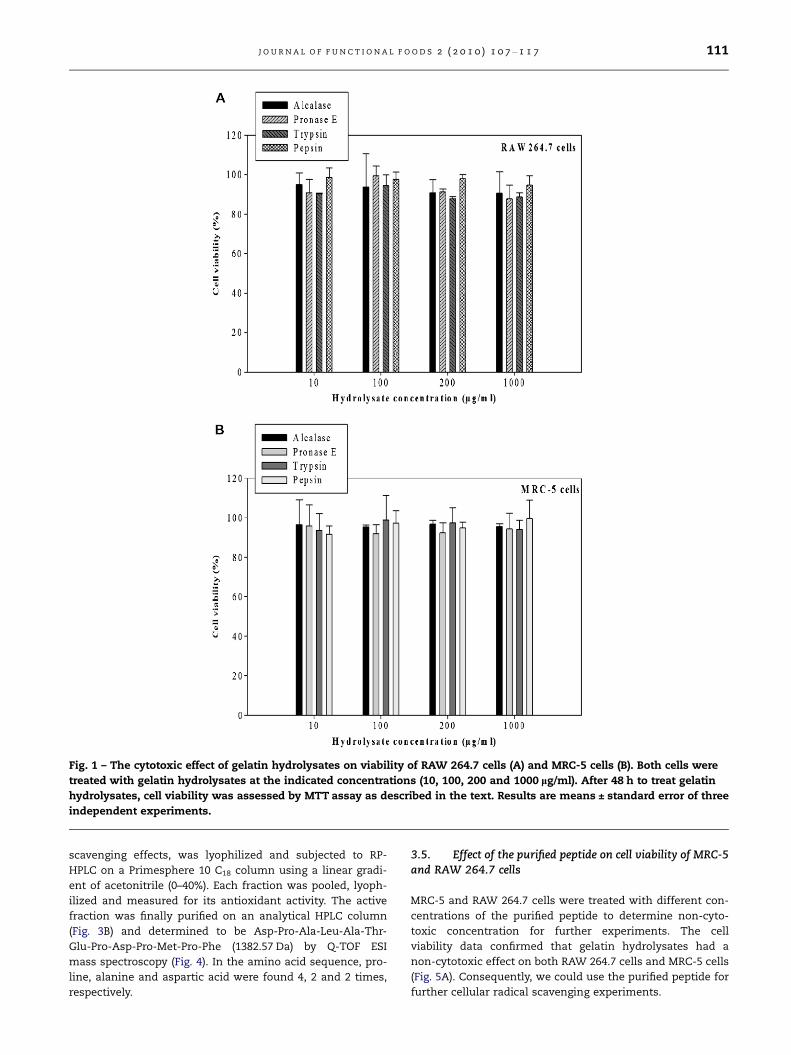
Fig. 1 – The cytotoxic effect of gelatin hydrolysates on viability of RAW 264.7 cells (A) and MRC-5 cells (B). Both cells were
treated with gelatin hydrolysates at the indicated concentrations (10, 100, 200 and 1000 lg/ml). After 48 h to treat gelatin
hydrolysates, cell viability was assessed by MTT assay as described in the text. Results are means ± standard error of three
independent experiments.
J O U R N A L O F F U N C T I O N A L F O O D S 2 ( 2 0 1 0 ) 1 0 7 – 1 1 7 111
scavenging effects, was lyophilized and subjected to RP-
HPLC on a Primesphere 10 C18 column using a linear gradi-
ent of acetonitrile (0–40%). Each fraction was pooled, lyoph-
ilized and measured for its antioxidant activity. The active
fraction was finally purified on an analytical HPLC column
(Fig. 3B) and determined to be Asp-Pro-Ala-Leu-Ala-Thr-
Glu-Pro-Asp-Pro-Met-Pro-Phe (1382.57 Da) by Q-TOF ESI
mass spectroscopy (Fig. 4). In the amino acid sequence, pro-
line, alanine and aspartic acid were found 4, 2 and 2 times,
respectively.
3.5. Effect of the purified peptide on cell viability of MRC-5and RAW 264.7 cells
MRC-5 and RAW 264.7 cells were treated with different con-
centrations of the purified peptide to determine non-cyto-
toxic concentration for further experiments. The cell
viability data confirmed that gelatin hydrolysates had a
non-cytotoxic effect on both RAW 264.7 cells and MRC-5 cells
(Fig. 5A). Consequently, we could use the purified peptide for
further cellular radical scavenging experiments.

Fig. 2 – (A) The intracellular ROS generation level of cells was measured by the DCFH-DA to detect produced H2O2. RAW 264.7
cells were treated with 20 nmol/ml DCFH-DA, and then, the cells were treated with different concentrations of gelatin
hydrolysates for 1 h. Results are means ± standard error of three independent experiment. (B) DNA oxidative damage
protection by the alcalase-derived hydrolysate. Genomic DNA from RAW 264.7 cells were pre-treated with the alcalase-
derived hydrolysate and exposed to *OH using Fenton chemistry. After 10 min, reaction mixture containing about 0.5 lg of
DNA was electrophoresed on a 1% agarose gel for 40 min at 100 V and visualized by UV light after stained with 1 mg/ml
ethidium bromide.
112 J O U R N A L O F F U N C T I O N A L F O O D S 2 ( 2 0 1 0 ) 1 0 7 – 1 1 7
3.6. Direct scavenging effects on free radicals of thepurified peptide
Free radical scavenging effects of the purified peptide
against different free radical sources using ESR technique
was examined. DPPH is a static free radical and accepts
an electron or hydrogen radical to become a static diamag-
netic molecule. Thus, DPPH is often used as a substrate to
evaluate the potency of an antioxidant. Hydroxyl radicals
were generated in a Fenton reaction and were visualized
by ESR spectrometer. The ESR signal was inhibited by the
presence of *OH scavengers, which compete against DMPO
for *OH. Superoxide radicals were generated by UV irradia-
tion of a riboflavin/EDTA solution. As shown in Table 2,
the peptide scavenged hydroxyl, DPPH and superoxide rad-
icals at the IC50 values of 7.56, 8.82 and 17.83 lM, respec-
tively. Antioxidative peptide was more dynamic on
hydroxyl radical than on other radicals. The chemical reac-
tivity of hydroxyl radical was the strongest among the ROS.
It easily responds with biomolecules such as amino acids,

Fig. 3 – (A) A FPLC chromatogram of hydrolysate loaded (2 ml) onto a HiPrep 16/10 DEAE FF ion-exchange column (lower
panel), and their antioxidant activity against free radicals measured by ESR spectroscopy (upper panel). Adsorption fractions
were eluted with a linear gradient of 0–2 M NaCl and antioxidant activity of the eluted peaks were measured by ESR
spectrometer. (B) A chromatogram of RP-HPLC (Primesphere 10 C18 column) for the purification of antioxidant peptides.
Elution was performed with the linear gradient of acetonitrile (0–40%) containing 0.1% TFA.
J O U R N A L O F F U N C T I O N A L F O O D S 2 ( 2 0 1 0 ) 1 0 7 – 1 1 7 113
proteins and DNA (Cacciuttolo et al., 1993). Consequently,
the removal of hydroxyl radical is probably one of the most
efficacious defenses of a living body against different dis-
eases. Several amino acids, such as tyrosine, methionine,
lysine, and tryptophan, are generally accepted as antioxi-
dants despite their pro-oxidative effects in some cases
(Marcuse, 1962; Yamaguchi, 1971). It has been known that
many antioxidative peptides include hydrophobic amino
acid residues, valine, or leucine, at the N-terminus of the
peptides (Chen et al., 1995; Uchida and Kawakishi, 1992).
The peptide from Nile tilapia (O. niloticus) scale gelatin
was rich in hydrophobic amino acids (>69%). In fact, aro-
matic amino acids are less in gelatin peptide sequence
and rich in glycine and proline. Therefore, it may be sug-
gested that, observed potent scavenging activities of Nile
tilapia (O. niloticus) scale gelatin peptide on hydroxyl, DPPH

Fig. 4 – Identification of the molecular mass and amino acid sequence of the purified peptide using TOF-MS/MS with an ESI
source. Sequencing of the active peptide was acquired over the m/z range 50–2500 and sequenced by using the PepSeq
denove sequencing algorithm.
114 J O U R N A L O F F U N C T I O N A L F O O D S 2 ( 2 0 1 0 ) 1 0 7 – 1 1 7
and superoxide radicals could be due to non-aromatic ami-
no acids such as alanine, proline, valine and leucine (Men-
dis et al., 2005a,b).
3.7. Cellular radical scavenging effect of the purifiedpeptide
The direct scavenging effects of peptide on cellular radicals
were studied in order to confirm their ability to scavenge free
radicals in a cellular environment. For that, RAW 264.7 cells
were labeled with DCFH-DA fluorescence probe as described
in section 2.5. DCFH-DA is a specific probe for reactive oxygen
species (ROS). The principle of this assay is that DCFH-DA dif-
fuses through cell membrane and is enzymatically hydro-
lyzed by esterase to become DCFH, which reacts with ROS
to form DCF, a fluorescent product. When cells were labeled
with DCFH-DA to DCFH and trapped inside cells and incu-
bated with H2O2 for 30 min, the rapid increment in DCF fluo-
rescence intensity indicated oxidation of DCFH by
intracellular radicals (Fig. 5B). The monitoring of DCF fluores-
cence intensities in every 30 min for 3 h duration that radical-
mediated oxidation increased with the incubation time. How-
ever, the pre-treatment with the purified peptide decreased
the DCF fluoresce by time dependent. The concentrations of
the purified peptide (0.2, 2, and 20 lg/ml) exhibited the free
radical scavenging effect even only after 30 min of incubation
compared with the control (H2O2 treatment and non-peptide)
as shown in Fig. 5B. Our results showed that its antioxidant
activity varies with the concentration of the purified peptide,
and the free radical scavenging effect of the purified peptide
at 20 lg/ml concentration was higher than other tested con-
centrations. Therefore, we could confirm that the peptide
scavenged free radicals in a dose- and time-dependent man-
ner in RAW 264.7 cells.
3.8. DNA protection activity of the purified peptide
Hydroxyl radicals (*OH) generated by the Fenton reaction are
known to cause oxidative breaks in DNA strands to yield its
open circular or relaxed forms. Genomic DNA was isolated
from RAW 264.7 cells to study protective effects of peptide
against DNA oxidative damage. In this experiment the com-
bined effects of 100 lM FeSO4 and 0.1 mM H2O2 on the integ-
rity of genomic DNA was studied by DNA electrophoresis in
the presence or absence of peptide. After 10 min of reaction,
almost all DNA was degraded in the control group treated
only with Fe (II)–H2O2. It was observed that the peptide signif-
icantly inhibited the oxidative damage of DNA (Fig. 5C). The
DNA damage was inhibited at all tested concentrations of
the peptide by about 70% based on the intensity of DNA
bands. The results of this study clearly explain that peptide
can prevent oxidative damage to DNA when DNA is exposed
to *OH generated by Fe (II)/H2O2. Fe2+ catalyses the conversion
of H2O2, which is a major route to the synthesis of *OH in bio-

Fig. 5 – (A) The cytotoxic effect of the purified peptide on viability of RAW 264.7 cells and MRC-5 cells. Both cells were treated
with peptide at the indicated concentrations (0.02, 0.2, 2, 10 and 20 lg/ml). After 48 h to treat peptide, cell viability was
assessed by MTT assay as described in the text. Results are means ± standard error of three independent experiments. (B)
The intracellular ROS generation level of cells was measured by the DCFH-DA which is compound to detect produced H2O2.
RAW 264.7 cells were treated with 20 nmol/ml DCFH-DA, and then, the cells were treated with different concentrations of
purified peptide for 1 h. Results are means ± standard error of three independent experiments. (C) DNA oxidative damage
protection by purified peptide. Genomic DNA from RAW 264.7 cells were pre-treated with peptide and exposed to *OH using
Fenton chemistry. After 10 min, reaction mixture containing about 0.5 lg of DNA was electrophoresed on a 1% agarose gel for
40 min at 100 V and visualized by UV light after stained with 1 mg/ml ethidium bromide.
J O U R N A L O F F U N C T I O N A L F O O D S 2 ( 2 0 1 0 ) 1 0 7 – 1 1 7 115
logical systems. The *OH reacts with all components of the
DNA molecule, including purine and pyrimidine bases and
also deoxyribose backbone and causes DNA lesion (Valko
et al., 2007). This lesion increases with increment of the free
radical attacking DNA of cells, which involved in mutagene-
sis, cancer and aging. However, this peptide can contribute

Table 2 – The IC50 values of peptide to scavenging DPPH,hydroxyl and superoxide radicals.
Peptideb IC50 (lM)a
DPPH Hydroxyl Superoxide
8.82 7.56 17.83
a Concentration of peptide required to inhibit 50% of the respective
radical ESR signal intensity.b Asp-Pro-Ala-Leu-Ala-Thr-Glu-Pro-Asp-Pro-Met-Pro-Phe.
116 J O U R N A L O F F U N C T I O N A L F O O D S 2 ( 2 0 1 0 ) 1 0 7 – 1 1 7
to preventing these diseases by its inhibitory effect against
DNA oxidation in live cells.
4. Conclusions
According to this study, the antioxidant capacity of Nile
tilapia (O. niloticus) scale gelatin peptide in various aspects
of oxidation and understanding the contribution of hydro-
phobic amino acids for their antioxidant activities relevant
to the sequence. Therefore, this antioxidant peptide derived
from Nile tilapia (O. niloticus) scale gelatin might be useful
as a potential functional ingredient in food and pharmaceu-
tical industries.
Acknowledgement
This research was supported by a grant from Marine Bioproc-
ess Research Center of the Marine Bio 21 Project funded by the
Ministry of Land, Transport and Maritime, Republic of Korea.
R E F E R E N C E S
Borneo, R., Leon, A. E., Aguirre, A., Ribotta, P., & Cantero, J. J. (2009).Antioxidant capacity of medicinal plants from the Province ofCordoba (Argentina) and their in vitro testing in a model foodsystem. Food Chemistry, 112, 664–670.
Cacciuttolo, M. A., Trinh, L., Lumpkin, J. A., & Rao, G. (1993).Hyperoxia induces DNA damage in mammalian cells. FreeRadical Biology and Medicine, 14, 267–276.
Calabrese, V., Lodi, R., Tonon, C., D’Agata, V., Sapienza, M.,Scapagnini, G., Mangiameli, A., Pennisi, G., Giuffrida Stella, A.M., & Butterfield, D. A. (2005). Oxidative stress, mitochondrialdysfunction and cellular stress response in Friedrich’s ataxia.Journal of the Neurological Sciences, 233, 145–162.
Chand, S. (2009). Bioactive peptides isolated from alcalasehydrolysed horse gram (Macrotyloma uniflorum) proteinexhibiting antioxidant activity. New Biotechnology, 25,S167–S168.
Chen, H. M., Muramoto, K., & Yamauchi, F. (1995). Structuralanalysis of antioxidative peptides from soybean b-conglycinin.Journal of Agricultural Food and Chemistry, 43, 574–578.
Cho, S. M., Gu, Y. S., & Kim, S. B. (2005). Extracting optimizationand physical properties of yellowfin tuna (Thunnus albacares)skin gelatin compared to mammalian gelatins. FoodHydrocolloids, 19, 221–229.
Choi, C. W., Kim, S. C., Hwang, S. S., Choi, B. K., Ahn, H. J., Lee, M.Y., Park, S. H., & Kim, S. K. (2002). Radical scavenging capacitybetween Korean medicinal plants and flavonoids by assay-guided comparison. Plant Science, 163, 1161–1168.
Diniz, F. M., & Martin, A. M. (1998). Influence of process variableson the hydrolysis of shark muscle protein. Food Science andTechnology International, 4, 91–98.
El-Sayed, A. M. (2006). Tilapia culture. Oxford: CABI publishing, 1-24.Engelmann, J., Volk, J., Leyhausen, G., & Geurtsen, W. (2005). ROS
formation and glutathione levels in human oral fibroblastsexposed to TEGDMA and camphorquinone. Journal ofBiomedical Materials Research Part B: Applied Biomaterials, 75B,272–276.
Fitzsimmons, K. (2004). Development of new products andmarkets for the global tilapia trade. In Proceeding of the sixthinternational symposium on tilapia in aquaculture. Manila,Philippines, pp. 624–633.
Gudmundsson, M., & Hafsteinsson, H. (1997). Gelatin from codskins as affected by chemical treatments. Journal of FoodScience, 62, 37–39.
Guo, Q., Zhao, B., Shen, S., Hou, J., Hu, J., & Xin, W. (1999). ESRstudy on the structure-antioxidant activity relationship of teacatechins and their epimers. Biochimica et Biophysica Acta, 1427,13–23.
Hansen, M. B., Nielsen, S. E., & Berg, K. (1989). Re-examination andfurther development of a precise and rapid dye method formeasuring cell growth/cell kill. Journal of ImmunologicalMethods, 119, 203–210.
Hyun, C. K., & Shin, H. K. (2000). Utilization of bovine bloodplasma proteins for the production of angiotensin I convertingenzyme inhibitory peptides. Process Biochemistry, 36, 65–71.
Jamilah, B., & Harvinder, K. G. (2002). Properties of gelatins fromskins of fish-black tilapia (Oreochromis mossambicus) and redtilapia (Oreochromis nilotica). Food Chemistry, 77, 81–84.
Je, J. Y., Park, P. J., & Kim, S. K. (2004). Free radical scavengingproperties of hetero-chitooligosaccharides using an ESRspectroscopy. Food and Chemical Toxicology, 42, 381–387.
Je, J. Y., Park, P. J., & Kim, S. K. (2005). Antioxidant activity of apeptide isolated from Alaska pollack (Theragra chalcogramma)frame protein hydrolysate. Food Research International, 38,45–50.
Kalogeropoulos, N., Konteles, S. J., Troullidou, E., Mourtzinos, I., &Karathanos, V. T. (2009). Chemical composition, antioxidantactivity and antimicrobial properties of propolis extracts fromGreece and Cyprus. Food Chemistry, 116, 452–461.
Ke, B., Xu, Z., Ling, Y., Qiu, W., Xu, Y., Higa, T., & Aruoma, O. I.(2009). Modulation of experimental osteoporosis in rats by theantioxidant beverage effective microorganism-X (EM-X).Biomedicine and Pharmacotherapy, 63, 114–119.
Kim, J. S., & Cho, S. Y. (1996). Screening for raw material ofmodified gelatin in marine animal skins caught in coastaloffshore water in Korea. Agricultural and Chemical Biotechnology,39, 134–139.
Kim, S. Y., & Je, J. Y. (2007). Purification and characterization ofantioxidant peptide from hoki (Johnius belengerii) frame proteinby gastrointestinal digestion. The Journal of NutritionalBiochemistry, 18, 31–38.
Kim, S. K., Byun, H. G., Park, P. J., & Shahidi, F. (2001a). AngiotensinI converting enzyme inhibitory peptides purified form bovineskin gelatin hydrolysate. Journal of Agricultural and FoodChemistry, 49, 2992–2997.
Kim, S. K., Kim, Y. T., Byun, H. G., Nam, K. S., Joo, D. S., & Shahidi,F. (2001b). Isolation and characterization of antioxidativepeptides from gelatin hydrolysate of Alaska pollack skin.Journal of Agricultural and Food Chemistry, 49, 1984–1989.
Marcuse, R. (1962). The effect of some amino acids on theoxidation of linoleic acid and its methyl ester. Journal of theAmerican Oil Chemists’ Society, 39, 97–103.
Mendis, E., Rajapakse, N., Byun, H. G., & Kim, S. K. (2005a).Investigation of jumbo squid (Dosidicus gigas) skin gelatinpeptides for their in vitro antioxidant effects. Life Sciences, 77,2166–2178.

J O U R N A L O F F U N C T I O N A L F O O D S 2 ( 2 0 1 0 ) 1 0 7 – 1 1 7 117
Mendis, E., Rajapakse, N., & Kim, S. K. (2005b). Antioxidantproperties of a radical scavenging peptide purified fromenzymatically prepared fish skin gelatin hydrolysate. Journal ofAgricultural and Food Chemistry, 53, 581–587.
Milne, L., Nicotera, P., Orrenius, S., & Burkitt, M. J. (1993). Effects ofglutathione and chelating agents on copper-mediated DNAoxidation: Pro-oxidant and antioxidant properties ofglutathione. Archives of Biochemistry and Biophysics, 304,102–109.
Nagai, T., & Suzuki, N. (2000). Isolation of collagen from fish wastematerial-skin, bone and fins. Food Chemistry, 68, 277–281.
Nanjo, F., Goto, K., Seto, R., Suzuki, M., Sakai, M., & Hara, Y. (1996).Scavenging effects of tea catechins and their derivatives on1,1,-diphenyl-2-picrylhydrazyl radical. Free Radical Biology andMedicine, 21, 895–902.
Nieto, G., Castillo, M., Xiong, Y. L., Alvarez, D., Payne, F. A., &Garrido, M. D. (2009). Antioxidant and emulsifying propertiesof alcalase-hydrolyzed potato proteins in meat emulsions withdifferent fat concentrations. Meat Science, 83, 24–30.
Osborne, K., Voight, M. N., & Hall, D. E. (1990). Utilization oflumpfish (Cyclopterus lumpus) carcasses for production ofgelatin. In M. N. Voight & J. K. Botta (Eds.), Advances infisheries technology and biotechnology for increased profitability(pp. 145–150). Lancaster, PA, USA: Technomic PublishingCo.
Park, P. J., Jung, W. K., Nam, K. S., Shahidi, F., & Kim, S. K. (2001).Purification and characterization of antioxidative peptidesfrom protein hydrolysate of lecithin-free egg yolk. Journal of theAmerican Oil Chemists’ Society, 78, 651–656.
Rosen, G. M., & Rauckman, E. J. (1984). Spin trapping of superoxideand hydroxyl radicals. In Packer, L. Methods in enzymology,105, Academic Press, Orlando, FL, 198–209
Sambrook, J., & Russell, D. (2001). In molecular cloning a laboratorymanual (pp. 84–87). New York: Cold Spring Harbor LaboratoryPress, Sanchez-Reus.
Saito, Y., Wanezaki, K., Kawato, A., & Imayasu, S. (1994). Structureand activity of angiotensin I converting enzyme inhibitorypeptides from sake lees. Bioscience, Biotechnology andBiochemistry, 58, 1767–1771.
Sarabia, A. I., Gomez-Guillen, M. C., Solas, M. T., & Montero, P.(2001). Effect of microbial transglutaminase on the functionalproperties of megrim (Lepidorhombus boscii) skin gelatin. Journalof the Science of Food and Agriculture, 81, 665–673.
Uchida, K., & Kawakishi, S. (1992). Sequence-dependent reactivityof histidine-containing peptides with copper(II)/ascorbate.Journal of Agricultural and Biological Chemistry, 40, 13–16.
Valko, M., Leibfritz, D., Moncol, J., Cronin, M. T. D., Mazur, M., &Telser, J. (2007). Free radicals and antioxidants in normalphysiological functions and human disease. The InternationalJournal of Biochemistry and Cell Biology, 39, 44–84.
Yamaguchi, N. (1971). Studies on antioxidative activities of aminocompounds on fats and oils. Part I. Oxidation of methionineduring course of autoxidation of linoleic acid. Nippon ShokuhinKogyo Gakkaishi, 18, 313–318.
Yoshimura, K., Terashima, M., Hozan, D., Ebato, T., Nomura, Y.,Ishii, Y., & Shirai, K. (2000). Physical properties of shark gelatincompared with pig gelatin. Journal of Agricultural FoodChemistry, 48, 2023–2027.







![Biomarkers in Nile Tilapia Oreochromis niloticus niloticus ... · biomarker measurements in Nile tilapia from the river Nile [2,23]. Therefore as a consequence, in this study I present](https://static.fdocuments.us/doc/165x107/5ebc4a0fc30023287c3a7364/biomarkers-in-nile-tilapia-oreochromis-niloticus-niloticus-biomarker-measurements.jpg)










