
Identification of CommonCyanobacterial Symbiont Associated ... · A COMMON SYMBIONT ASSOCIATED WITH...
6
Vol. 57, No. 8 APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Aug. 1991, p. 2141-2146 0099-2240/91/082141-06$02.00/0 Copyright 33 1991, American Society for Microbiology Identification of a Common Cyanobacterial Symbiont Associated with Azolla spp. through Molecular and Morphological Characterization of Free-Living and Symbiotic Cyanobacteria JOAN S. GEBHARDT AND SANDRA A. NIERZWICKI BAUER* Plant Research Group, Department of Biology, Rensselaer Polytechnic Institute, Troy, New York 12180-3590 Received 12 February 1991/Accepted 13 May 1991 Symbiotically associated cyanobacteria from Azolla mexicana and Azolla pinnata were isolated and cultured in a free-living state. Morphological analyses revealed differences between the free-living isolates and their symbiotic counterparts, as did restriction fragment length polymorphism (RFLP) analyses with both single- copy glnA and rbcS gene probes and a multicopy psbA gene probe. RFLP analyses with Anabaena sp. strain PCC 7120 nipj excision element probes, including an xisA gene probe, detected homologous sequences in DNA extracted from the free-living isolates. Sequences homologous to these probes were not detected in DNA from the symbiotically associated cyanobacteria. These analyses indicated that the isolates were not identical to the major cyanobacterial symbiont species residing in leaf cavities of Azolla spp. Nevertheless, striking similarities between several free-living isolates were observed. In every instance, the isolate from A. pinnata displayed banding patterns virtually identical to those of free-living cultures previously isolated from Azolla caroliniana and Azolla filiculoides. These results suggest the ubiquitous presence of a culturable minor cyanobacterial symbiont in at least three species of Azolla. Anabaena azollae is a prokaryotic, filamentous, nitrogen- fixing cyanobacterium that is symbiotically associated with the small, eukaryotic water fern Azolla spp. In the Azolla- Anabaena association, the endosymbiont is contained within specialized leaf cavities of the fern, and under natural conditions, the two partners remain associated throughout vegetative and reproductive development (see reference 2 for a review). Researchers studying the Azolla-Anabaena symbiosis have long been interested in separating the two organisms in order to assess more accurately the relative roles played by each partner. Cyanobacteria-free Azolla spp. are generally not found in nature, but cultures have been generated by a variety of methods for use in laboratory studies. One of the more frequently used methods is antibiotic treatment (19), while perhaps the most direct method is germination of fertile megasporocarps, from which the apical cap, containing the cyanobacterial inoculum, is removed (12). Cyanobacteria from the symbiosis have also been isolated and cultured independently of the fern (1, 7, 16, 25, 26). Methods used to accomplish this have included microdissec- tion or homogenization of surface-sterilized Azolla megaspo- rocarps (25), Azolla fronds (1, 26), or isolated algal packets (7, 16). Once they are liberated from the fern, the cyanobac- teria are cultured autotrophically in a dilute mineral medium lacking a combined nitrogen source. Researchers have noted that although the free-living isolates of Anabaena azollae closely resemble Anabaena or Nostoc spp. in their general morphologies (16, 28), the in vitro-cultured filaments are morphologically distinct from filaments freshly removed from the fern cavity (16). In recent years, there has been interest in determining whether the free-living Anabaena azollae cultures isolated from Azolla spp. are identical to the cyanobacterial sym- * Corresponding author. bionts residing within leaf cavities of the fern. Immunologi- cal and lectin hemagglutination studies have shown that the free-living cultures are immunologically distinct and express different surface proteins than their freshly separated sym- biotic counterparts (1, 7, 10, 14). Most recently, restriction fragment length polymorphism (RFLP) analyses have been used to compare free-living cyanobacteria isolated from Azolla spp. with their symbiotic counterparts. In two studies, a free-living isolate from Azolla filiculoides did not share any common hybridization bands with nine different freshly separated cyanobacterial sym- bionts from Azolla spp., including A. filiculoides (5, 6). A similar study demonstrated that the ginA, psbA, rbcS, and nifH genes are contained on DNA fragments of different sizes in a free-living cyanobacterial culture isolated from Azolla caroliniana and its symbiotic counterpart (18). Dif- ferent hybridization banding patterns were also exhibited by free-living cyanobacteria isolated from A. caroliniana via subculture through symbiotic association with Anthoceros spp. and freshly separated cyanobacterial symbionts when the organization of nif genes was investigated (15). We are primarily interested in the regulation of transcrip- tion in the Azolla symbiosis. The physical compartmental- ization of the cyanobacterial symbionts within the leaf cavities makes direct measurements and manipulations of environmental and nutritional factors difficult, if not impos- sible. For this reason, it was of interest to isolate and culture cyanobacterial symbionts for transcriptional studies to allow comparisons to be made between free-living and symbiotic growth. Here we report on the isolation and culture of cyanobacterial symbionts from the Azolla mexicana and Azolla pinnata associations. These free-living isolates were characterized by RFLP analyses and compared with freshly separated cyanobacterial symbionts. Results of studies on the morphological properties of these isolates and their symbiotic counterparts are also presented. 2141 on April 9, 2020 by guest http://aem.asm.org/ Downloaded from
Transcript of Identification of CommonCyanobacterial Symbiont Associated ... · A COMMON SYMBIONT ASSOCIATED WITH...

Vol. 57, No. 8APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Aug. 1991, p. 2141-21460099-2240/91/082141-06$02.00/0Copyright 33 1991, American Society for Microbiology
Identification of a Common Cyanobacterial Symbiont Associatedwith Azolla spp. through Molecular and Morphological
Characterization of Free-Living and Symbiotic CyanobacteriaJOAN S. GEBHARDT AND SANDRA A. NIERZWICKI BAUER*
Plant Research Group, Department of Biology, Rensselaer Polytechnic Institute, Troy, New York 12180-3590
Received 12 February 1991/Accepted 13 May 1991
Symbiotically associated cyanobacteria from Azolla mexicana and Azolla pinnata were isolated and culturedin a free-living state. Morphological analyses revealed differences between the free-living isolates and theirsymbiotic counterparts, as did restriction fragment length polymorphism (RFLP) analyses with both single-copy glnA and rbcS gene probes and a multicopy psbA gene probe. RFLP analyses with Anabaena sp. strainPCC 7120 nipj excision element probes, including an xisA gene probe, detected homologous sequences in DNAextracted from the free-living isolates. Sequences homologous to these probes were not detected in DNA fromthe symbiotically associated cyanobacteria. These analyses indicated that the isolates were not identical to themajor cyanobacterial symbiont species residing in leaf cavities of Azolla spp. Nevertheless, striking similaritiesbetween several free-living isolates were observed. In every instance, the isolate from A. pinnata displayedbanding patterns virtually identical to those of free-living cultures previously isolated from Azolla carolinianaand Azolla filiculoides. These results suggest the ubiquitous presence of a culturable minor cyanobacterialsymbiont in at least three species of Azolla.
Anabaena azollae is a prokaryotic, filamentous, nitrogen-fixing cyanobacterium that is symbiotically associated withthe small, eukaryotic water fern Azolla spp. In the Azolla-Anabaena association, the endosymbiont is contained withinspecialized leaf cavities of the fern, and under naturalconditions, the two partners remain associated throughoutvegetative and reproductive development (see reference 2for a review). Researchers studying the Azolla-Anabaenasymbiosis have long been interested in separating the twoorganisms in order to assess more accurately the relativeroles played by each partner.
Cyanobacteria-free Azolla spp. are generally not found innature, but cultures have been generated by a variety ofmethods for use in laboratory studies. One of the morefrequently used methods is antibiotic treatment (19), whileperhaps the most direct method is germination of fertilemegasporocarps, from which the apical cap, containing thecyanobacterial inoculum, is removed (12).
Cyanobacteria from the symbiosis have also been isolatedand cultured independently of the fern (1, 7, 16, 25, 26).Methods used to accomplish this have included microdissec-tion or homogenization of surface-sterilized Azolla megaspo-rocarps (25), Azolla fronds (1, 26), or isolated algal packets(7, 16). Once they are liberated from the fern, the cyanobac-teria are cultured autotrophically in a dilute mineral mediumlacking a combined nitrogen source. Researchers have notedthat although the free-living isolates of Anabaena azollaeclosely resemble Anabaena or Nostoc spp. in their generalmorphologies (16, 28), the in vitro-cultured filaments aremorphologically distinct from filaments freshly removedfrom the fern cavity (16).
In recent years, there has been interest in determiningwhether the free-living Anabaena azollae cultures isolatedfrom Azolla spp. are identical to the cyanobacterial sym-
* Corresponding author.
bionts residing within leaf cavities of the fern. Immunologi-cal and lectin hemagglutination studies have shown that thefree-living cultures are immunologically distinct and expressdifferent surface proteins than their freshly separated sym-biotic counterparts (1, 7, 10, 14).Most recently, restriction fragment length polymorphism
(RFLP) analyses have been used to compare free-livingcyanobacteria isolated from Azolla spp. with their symbioticcounterparts. In two studies, a free-living isolate from Azollafiliculoides did not share any common hybridization bandswith nine different freshly separated cyanobacterial sym-bionts from Azolla spp., including A. filiculoides (5, 6). Asimilar study demonstrated that the ginA, psbA, rbcS, andnifH genes are contained on DNA fragments of differentsizes in a free-living cyanobacterial culture isolated fromAzolla caroliniana and its symbiotic counterpart (18). Dif-ferent hybridization banding patterns were also exhibited byfree-living cyanobacteria isolated from A. caroliniana viasubculture through symbiotic association with Anthocerosspp. and freshly separated cyanobacterial symbionts whenthe organization of nif genes was investigated (15).We are primarily interested in the regulation of transcrip-
tion in the Azolla symbiosis. The physical compartmental-ization of the cyanobacterial symbionts within the leafcavities makes direct measurements and manipulations ofenvironmental and nutritional factors difficult, if not impos-sible. For this reason, it was of interest to isolate and culturecyanobacterial symbionts for transcriptional studies to allowcomparisons to be made between free-living and symbioticgrowth. Here we report on the isolation and culture ofcyanobacterial symbionts from the Azolla mexicana andAzolla pinnata associations. These free-living isolates werecharacterized by RFLP analyses and compared with freshlyseparated cyanobacterial symbionts. Results of studies onthe morphological properties of these isolates and theirsymbiotic counterparts are also presented.
2141
on April 9, 2020 by guest
http://aem.asm
.org/D
ownloaded from

2142 GEBHARDT AND NIERZWICKI-BAUER
MATERIALS AND METHODS
Azolla cultures. The symbiotically associated water fernsA. mexicana Presl (International Rice Research Institute[IRRI] collection no. 2001) and A. pinnata R. Brown (IRRIcollection no. 2) were used in this study. Both organismswere grown in 2.8-liter Fernbach flasks floating on 1 liter ofIRRI medium (27) lacking combined nitrogen. Cultures weremaintained in a controlled-environment room under a 16 h-8h light-dark photoperiod at temperatures ranging from 20 to25°C. Illumination was provided by cool white fluorescentlights and Agro-Lite lights (7:1) with an intensity of 110 to140 ,umol m2 S-.
Free-living cyanobacterial symbiont cultures. The free-living cyanobacterium Anabaena azollae la was isolated byNewton and Herman (16) from A. caroliniana (IRRI collec-tion no. 3001) and supplied by Jack Newton of the RegionalLaboratory, U.S. Department of Agriculture, Peoria, Ill.This isolate was grown in BG-11 medium (24) lacking com-bined nitrogen, as were the free-living cyanobacteria isolatedfrom A. mexicana and A. pinnata. Cultures were incubatedon a Gyrotory shaker (New Brunswick Scientific Co., Inc.,Edison, N.J.) at 200 rpm under constant illumination fromcool white fluorescent lights (50 to 60 Lmol m-2 S-1)under room temperatures ranging from 20 to 25°C.
Isolation and cultivation of cyanobacterial symbionts in afree-living state. A modification of the procedure devised byWhite (27a) was used to isolate and culture cyanobacterialsymbionts from A. mexicana and A. pinnata. Fern tissue (10to 15 g [fresh weight]) was surface sterilized in 10% Cloroxfor 2 min. This was followed by a 1-min rinse in sterile 0.01N HCI and two 1-min rinses in sterile distilled water. Thetissue was homogenized with a mortar and pestle in 50 ml ofone-eighth-strength BG-11 (BG-11/8) medium and was fil-tered through four layers of cheesecloth and then througheight layers of cheesecloth. The filtrate was centrifuged at500 x g for 5 min to pellet the cells. The cell pellet waswashed three times with BG-11/8 medium and was then usedto inoculate 50 ml of BG-11/8 medium. Cultures were incu-bated under laboratory light conditions (10 Lmol m-2. s-)at temperatures ranging from 20 to 25°C and transferred tofresh medium every 2 weeks. After 4 weeks, the cultureswere incubated on a Gyrotory shaker at 200 rpm in full-strength BG-11 medium under the conditions describedabove. Growth of cultures was evident after 4 to 6 weeks.
Preparation of "freshly separated" cyanobacterial sym-bionts. A modification of the "gentle roller" proceduredevised by Peters and Mayne (19) as described by Ni-erzwicki-Bauer and Haselkorn (18) was used to freshlyseparate the symbiotic cyanobacteria from the ferns foranalyses.
Preparation and analysis of DNA. Total DNA from vege-tative cells, heterocysts, and akinetes (if present) of bothfree-living and symbiotic cyanobacterial cultures was pre-pared as described previously (8) by using 0.45-mm-diameterglass beads and vigorous vortexing to achieve cell lysis.Because of the presence of a thick outer sheath surroundingthe filaments of the free-living cyanobacterium isolated fromA. mexicana, pretreatment with lysozyme, several freeze-thaw cycles, and extensive vortexing were required toachieve cell lysis of this cyanobacterial culture. Chromo-somal DNA (approximately 4 ,ug) was digested with therestriction endonuclease EcoRI or HindIll (Bethesda Re-search Laboratories, Gaithersburg, Md.), according to therecommendations of the manufacturer. These enzymes areroutinely used in conjunction with cyanobacterial DNA. In
some instances, chromosomal DNA was subjected to Gene-clean (Bio 101, La Jolla, Calif.) treatment or was passedthrough an Elutip-D column (Schleicher & Schuell, Keene,N.H.) prior to endonuclease digestion. Restricted DNA wasfractionated by agarose gel electrophoresis in 0.7% agarosegels. Size-fractionated DNA was denatured and neutralizedas described previously (13) and transferred to Nytranmembranes (Schleicher & Schuell) by the technique ofSouthern (23). DNA was immobilized on the solid supportby baking in vacuo for 2 h at 80°C. Growth of Escherichiacoli strains and subsequent isolation and digestion of plasmidDNA from these strains were done as described previously(13).
Probes. Southern blots were probed with the followingcloned genes from Anabaena sp. strain PCC 7120: (i) the 1.4-kb EcoRI fragment of pAn5O3 containing the majority of theginA gene along with some 3'-flanking sequences (4), (ii) the1.4-kb EcoRI-HindIII fragment of pAn625 containing thecomplete psbAI gene along with 5'- and 3'-flanking se-quences (3), and (iii) the 980-bp HindIII-XbaI fragment ofpAn6O6 containing the complete rbcS gene along with 5'- and3'-flanking sequences (17). Blots were also probed with twocloned fragments from within the 11-kb nifD excision ele-ment of Anabaena sp. strain PCC 7120, the 1.8-kb HindIIIfragment of pAn2O7.3 (8, 11), and the 1.2-kb HindlIl frag-ment of pAn2O7.5 containing the majority of the xisA genealong with 3'-flanking sequences (11, 21). Agarose gel-purified fragments (25 ng) were labeled with 50 ,uCi of [a-32P]dCTP (3,000 Ci/mmol; Amersham Corp., Arlington Heights,Ill.) by using the multiprime DNA labeling system (Amer-sham Corp.). The labeled DNA fragments were separatedfrom unincorporated nucleotides by passage over SephadexG-50 columns (NICK-Column; Pharmacia LKB Biotechnol-ogy, Piscataway, N.J.) and heat denatured immediatelybefore addition to the hybridization solution.
Hybridization conditions. Nytran membranes were prehy-bridized for a minimum of 4 h, in the solution recommendedby the manufacturer, at 50'C (pAn2O7 probes), 52°C (rbcS),55°C (ginA), or 62°C (psbA). The membranes were hybrid-ized overnight (for a minimum of 12 h) at the same temper-atures in the recommended solution without formamide.Following hybridization, the membranes were washed asrecommended by the manufacturer, including a stringentwash at the hybridization temperature. The filters were air-dried, and autoradiographs were obtained by exposingKodak X-OMAT AR film at -70°C with Cronex LightningPlus intensifying screens (DuPont NEN Research Products,Boston, Mass.).
Light microscopy. Wet mounts of free-living cyanobacte-rial cultures were viewed with an Olympus BH light micro-scope, and light micrographs were obtained by using IlfordPanF film. Symbiotic cyanobacteria were released from leafcavities by micromanipulation, separated from plant mate-rial, and visualized and photographed in the same manner asthe free-living cultures.
RESULTS
RFLP analyses. (i) A. mexicana cyanobacterial cultures.The restriction fragment patterns obtained by hybridizationofDNA extracted from the free-living isolate of A. mexicanaand its freshly separated symbiotic counterpart with thesingle-copy ginA (Fig. 1A) and rbcS (Fig. 1B) probes indi-cated differences in the distributions of restriction siteswithin and around these structural genes in the two cultures.The banding patterns obtained with both the ginA and rbcS
APPL. ENVIRON. MICROBIOL.
on April 9, 2020 by guest
http://aem.asm
.org/D
ownloaded from
A COMMON SYMBIONT ASSOCIATED WITH AZOLLA SPP.
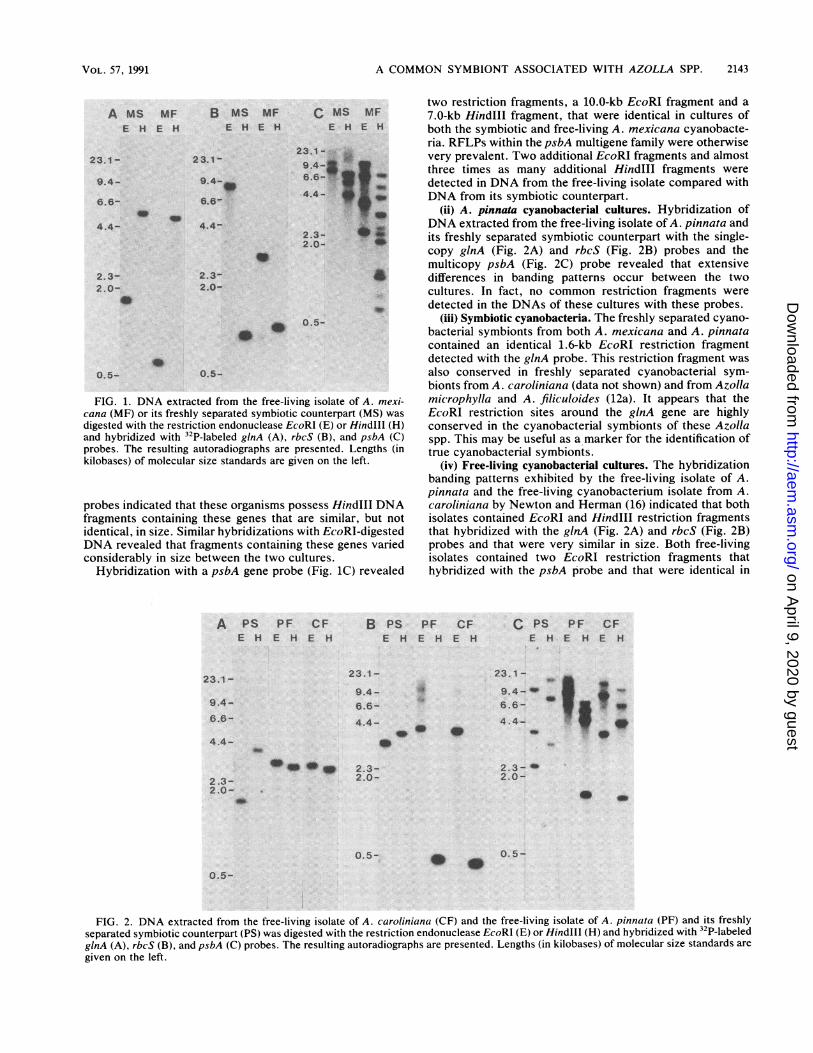
FIG. 1. DNA extracted from the free-living isolate of A. mexi-cana (MF) or its freshly separated symbiotic counterpart (MS) wasdigested with the restriction endonuclease EcoRI (E) or Hindlll (H)and hybridized with 32P-labeled ginA (A), rbcS (B), and psbA (C)probes. The resulting autoradiographs are presented. Lengths (inkilobases) of molecular size standards are given on the left.
probes indicated that these organisms possess HindIII DNAfragments containing these genes that are similar, but notidentical, in size. Similar hybridizations with EcoRI-digestedDNA revealed that fragments containing these genes variedconsiderably in size between the two cultures.
Hybridization with a psbA gene probe (Fig. 1C) revealed
two restriction fragments, a 10.0-kb EcoRI fragment and a7.0-kb Hindlll fragment, that were identical in cultures ofboth the symbiotic and free-living A. mexicana cyanobacte-ria. RFLPs within the psbA multigene family were otherwisevery prevalent. Two additional EcoRI fragments and almostthree times as many additional HindIll fragments weredetected in DNA from the free-living isolate compared withDNA from its symbiotic counterpart.
(ii) A. pinnata cyanobacterial cultures. Hybridization ofDNA extracted from the free-living isolate of A. pinnata andits freshly separated symbiotic counterpart with the single-copy ginA (Fig. 2A) and rbcS (Fig. 2B) probes and themulticopy psbA (Fig. 2C) probe revealed that extensivedifferences in banding patterns occur between the twocultures. In fact, no common restriction fragments weredetected in the DNAs of these cultures with these probes.
(iii) Symbiotic cyanobacteria. The freshly separated cyano-bacterial symbionts from both A. mexicana and A. pinnatacontained an identical 1.6-kb EcoRI restriction fragmentdetected with the ginA probe. This restriction fragment wasalso conserved in freshly separated cyanobacterial sym-bionts from A. caroliniana (data not shown) and from Azollamicrophylla and A. filiculoides (12a). It appears that theEcoRI restriction sites around the ginA gene are highlyconserved in the cyanobacterial symbionts of these Azollaspp. This may be useful as a marker for the identification oftrue cyanobacterial symbionts.
(iv) Free-living cyanobacterial cultures. The hybridizationbanding patterns exhibited by the free-living isolate of A.pinnata and the free-living cyanobacterium isolate from A.caroliniana by Newton and Herman (16) indicated that bothisolates contained EcoRI and HindlIl restriction fragmentsthat hybridized with the ginA (Fig. 2A) and rbcS (Fig. 2B)probes and that were very similar in size. Both free-livingisolates contained two EcoRI restriction fragments thathybridized with the psbA probe and that were identical in
FIG. 2. DNA extracted from the free-living isolate of A. caroliniana (CF) and the free-living isolate of A. pinnata (PF) and its freshlyseparated symbiotic counterpart (PS) was digested with the restriction endonuclease EcoRI (E) or HindIll (H) and hybridized with 32P-labeledginA (A), rbcS (B), and psbA (C) probes. The resulting autoradiographs are presented. Lengths (in kilobases) of molecular size standards aregiven on the left.
VOL. 57, 1991 2143
on April 9, 2020 by guest
http://aem.asm
.org/D
ownloaded from

2144 GEBHARDT AND NIERZWICKI-BAUER
TABLE 1. Sizes of restriction endonuclease fragments ofcyanobacterial DNA that hybridized with the nifD
excision element probes
DNA restriction endonucleaseRestriction fragment size (kb) of the followingProbe endonuclease cyanobacterial isolates":
MF PF CF
pAn207.3 EcoRI 4.6 10.0 10.0
HindIll 0.5 2.2 2.2
xisA EcoRl 5.0b 7.0 7.01.1
HindIll 3.5 4.2 4.20.6 1.4'7 1.4"
"DNA was extracted from the free-living isolates of A. mexic ana (MF), A.pinnata (PF), and A. caroliniana (CF).
b Weakly hybridizing bands.
size and three HindIII restriction fragments that were verysimilar in size. Additional hybridizations with a 23S rRNAprobe from Anabaena sp. strain PCC 7120 (18) and a secondglnA probe containing 5'-flanking and -coding sequehcesrevealed very similar banding patterns, which suggests thepresence of highly conserved restriction sites within andaround the ginA gene in these two organisms (data notshown). The similarities in the hybridization banding pat-terns of these two free-living isolates even occurred withinthe nijD excision element. The pAn2O7.3 and xisA probeshybridized to both EcoRI and HindIll restriction fragmentsthat were very similar in size (Table 1). These probes alsohybridized to DNA extracted from the free-living isolate ofA. mexicana, although the sizes of the restriction fragmentsvaried considerably (Table 1).
Morphological analyses. Wet mounts of cyanobacterialfilaments were examined by light microscopy to establish themorphological characteristics of the free-living and symbioticcyanobacteria. Three cell types were observed in cyanobac-terial filaments found in association with A. mexicana, A.pinnata, and A. caroliniana (Fig. 3). Filaments composedalmost entirely of vegetative cells were predominantly foundin association with the apical meristem and young leaves ofAzolla plants. Vegetative cells comprising a single filamentwere very similar in size and shape; however, these charac-teristics varied considerably among different filaments (Fig.3A). Cyanobacterial filaments typically found in middle-aged(approximately leaf 5 to leaf 9) or older (leaf 10 or greater)leaves contained numerous heterocysts (up to 30%) thatwere morphologically distinct from vegetative cells becauseof their increased size, rounder shape, and increased refrac-tiveness (Fig. 3B). Cells within filaments present in olderAzolla leaf cavities (10 or more leaves) typically possessedlarge cyanophycin accumulations (Fig. 3C). Cyanophycingranules were also frequently observed at the cross walls ofvegetative cells, unlike what is observed for free-livingAnabaena and Nostoc spp. In addition, akinetes (spores)were frequently observed within filaments or as single cellsin older cavities.
Cyanobacterial filaments in association with Azolla spp.were quite diverse in cell type, size, shape, and the inclusionbodies they contained. In contrast, free-living cyanobacteriaisolated from Azolia spp. were similar in size and exhibitedvery uniform morphological characteristics (Fig. 3D and E).
Young filaments made up entirely of vegetative cells areshown in Fig. 3D. The free-living isolate from A. mexicanawas similar in size to the symbiotically associated cyanobac-teria and, on the basis of the pattern of akinete developmentand the formation of motile homogonia, can be classified asa Nostoc sp. In contrast, the free-living isolates from A.pinnata and A. caroliniana were smaller in size than thesymbiotically associated cyanobacteria and resembled Ana-baena spp. The free-living isolates exhibited a heterocystfrequency of only 10% or less (Fig. 3E). Terminal hetero-cysts were predominant in the isolate from A. mexicana,although intercalary heterocysts were also observed. Iso-iates from A. pinnata and A. caroliniana primarily containedintercalary heterocysts. Under the growth conditions used,akinetes were observed only in the A. mexicana isolate (datanot shown). The akinetes displayed an apoheterocytic pat-tern of development in which akinete differentiation began inthe middle of the interval between two heterocysts andproceeded outward to the heterocysts on either side.
DISCUSSION
On the basis of RFLP analyses and morphological char-acteristics, it appears that the free-living cyanobacteriaisolated and cultured in vitro in this study are not the majorcyanobacterial symbionts present within symbiotically asso-ciated Azolla spp. This phenomenon has been noted previ-ously on the basis of differences in morphology (16), surfaceantigenicity (1, 7, 10, 14), and molecular organization (5, 6,15, 18) between free-living isolates of Azolla and theirsymbiotic counterparts.
Several explanations have been offered to account for theaforementioned differences. It has been postulated thatdrastic environmental changes encountered during isolationand subsequent culturing of the cyanobacterial symbiontshave resulted in changes in morphology and surface antige-nicity (1, 7, 10). However, extensive changes in molecularorganization at the DNA level between free-living isolatesand their symbiotic counterparts (5, 6, 15, 18) are unlikely tohave resulted from such drastic environmental changes. Infact, no large-scale genomic rearrangements were observedin Nostoc sp. strain 7801 because of the transition fromsymbiotic association with Anthoceros spp. to a free-livingstate (15).
It has also been postulated that the major cyanobacterialconstituent of Azolla cavities is an obligate endosymbiontincapable of in vitro growth (7, 16, 20). Whether this isindeed the case, the question that still remains is, what isactually being cultured?
It has been suggested that isolation and culturing mayselect for a mutant strain capable of in vitro growth (7). Thispossibility seems unlikely, given the results of this studydemonstrating that free-living cultures isolated from A.caroliniana and A. pinnata display nearly identical bandingpatterns with the ginA, rbcS, and psbA (Fig. 2) and the nifD(Table 1) excision element probes. These isolates also dis-played nearly identical banding patterns when they wereprobed with a 23S rRNA gene (data not shown). Thepossibility that many different mutations would result inalmost identical banding patterns for two unique isolatesfrom different species of Azolla seems very unlikely.
It has also been suggested that free-living isolates may notbe true isolates from within the dorsal leaf cavities (10) but,in faq4, are contaminating epiphytic organisms associatedwith the outer surface of the ferns (5). This seems improba-ble, because the ferns are extensively surface sterilized prior
APPL. ENVIRON. MICROBIOL.
on April 9, 2020 by guest
http://aem.asm
.org/D
ownloaded from

A COMMON SYMBIONT ASSOCIATED WITH AZOLLA SPP. 2145
Symbiotic
A B
Azollamexicana.
Azollapinnata
Free- Living;:;t t0;; d;0 lk 0 . ... .. -. f: S}y .. ; ::.t.j;7S . . S : i . i . . ..... ,.: ..... : S . ..
L _: .fG.: :0 C::tC
0:
A:00 t040f'VAV:X00000 0:,,,,,_,A._ ......................... ,.,_o_ ............ j. C,___=_
7 vf w: v -<X.-i-z/t,7-.. +Me 10\' s,,; ,. -t-.,: ,- X, ts ,w, .; e64s,,, '' :R ;,._!
; 000 t; 0: *,s.;5f0'V.-' 7'; 0 ' , H W i | # M ! _! X X i X i i |_|: f; u iM | S | |W: :0fff::00
W^L',00iX) i. fffU;00 -Ff.-,,..,00X0. .0.:S: :0\::f
Azolla k :;caroliniana
FIG. 3. Light micrographs of cyanobacterial filaments symbiotically associated with or isolated and cultured in a free-living state from A.mexicana, A. pinnata, and A. caroliniana. Examples of cyanobacterial filaments representative of those commonly found in symbioticassociation with the apical meristem and young leaves, middle-aged leaves (approximately 5 to 9), and older leaves (10 or greater) of Azollaspp. are shown in panels A, B, and C, respectively. Unless otherwise noted, cells making up the filaments present in young leaves (A) are
all vegetative. Cyanobacterial filaments of the free-living isolates from Azolla spp. are shown in panels D and E. Filaments made up entirelyof vegetative cells are presented in panel D. Filaments which contain heterocysts and vegetative cells are shown in panel E. A, akinete; H,heterocyst; PA, proakinete; PH, proheterocyst; large arrowheads, cyanophycin granules. Bars, 5.0 p.m.
to the isolation of the cyanobacterial symbionts, or isolatesare obtained from purified algal packets.The third, and most likely, possibility is that more than
one species of Anabaena, Nostoc, or both is living insymbiotic association with a single Azolla sp. (2, 5, 15, 20,28). RFLP analyses have shown that free-living isolates fromA. caroliniana and A. filiculoides are not the major endo-symbiotic species residing in the fern cavities (5, 6, 15, 18).Therefore, the work presented here extends these findings tothe free-living isolates of A. mexicana and A. pinnata. Thefree-living isolates most likely exist as minor constituents ofthe cyanobacterial population within the cavity.RFLP analyses comparing two isolates from A. carolini-
ana with their symbiotic counterparts lend further support to
the existence of minor constituents. Although one isolatewas obtained through an association with an Anthoceros sp.(15) and the other was obtained from purified algal packets(16), neither isolate was identical to the major cyanobacterialspecies residing within Azolla leaf tips (15, 18). Theseisolates were also genetically different from each other onthe basis of RFLP analyses (15, 18). This raises the possi-bility that three cyanobacterial populations may residewithin the leaf cavities of A. caroliniana.The free-living isolates from A. caroliniana and A. pinnata
displayed very similar RFLP banding patterns and, in fact,could only be distinguished from each other by hybridizationwith the psbA probe. These results are surprising, given thefact that these Azolla spp. belong to two separate sections
VOL. 57, 1991
on April 9, 2020 by guest
http://aem.asm
.org/D
ownloaded from

2146 GEBHARDT AND NIERZWICKI-BAUER
(subgenera) (2). It is even more surprising that a free-livingisolate from A. filiculoides (26) displayed banding patternsidentical to those of both of these isolates when they werehybridized with the glnA probe and a pattern identical to thatof the free-living isolate from A. caroliniana when they werehybridized with the psbA probe (data not shown). Addition-ally, these three isolates were virtually indistinguishable onthe basis of morphological assessments (28; this study) andextensive allozyme analyses (28). The remarkable similar-ities between these isolates from three Azolla spp. also lendsfurther support to the existence of culturable minor constit-uents within Azolla leaf cavities.On the basis of morphological assessments (28; this study)
and allozyme analyses (28), the free-living isolate from A.pinnata has been classified as an Anabaena sp., whereas thefree-living isolate from A. mexicana has been classified as aNostoc sp. Such assignments have been based on previouslypublished classification systems (9, 22). Restriction mappingof conserved sites in the nif region has also been suggestedas a means of distinguishing the genus Nostoc from thegenus Anabaena (15).While it should be possible to detect the free-living iso-
lates, if they exist as minor constituents of the cyanobacte-rial symbiont population, DNA fingerprints of these isolateshave not yet been detected in RFLP analyses with DNAextracted from freshly separated cyanobacterial symbionts,even given extensive exposure times for autoradiography. Inaddition, comparative morphological analyses have been oflittle value because of the morphological diversity of thecyanobacterial symbionts residing in leaf cavities of Azollaspp. (19; this study). Thus, the polymerase chain reaction isbeing used in our laboratory to elucidate whether DNA ofthe minor constituents is a component of total DNA ex-tracted from the cyanobacterial symbiont population ofAzolla spp.
ACKNOWLEDGMENTS
This work was supported by grant 1ROlGM3705-01 awarded bythe National Institute of General Medical Sciences (to S.A.N.-B.).We thank Mary Ann Sessions for typing the manuscript.
REFERENCES1. Arad, H., A. Keysari, E. Tel-Or, and D. Kobiler. 1985. A
comparison between cell antigens in different isolates of Ana-baena azollae. Symbiosis 1:195-204.
2. Braun-Howland, E., and S. A. Nierzwicki-Bauer. 1990. Azolla-Anabaena symbiosis: biochemistry, ultrastructure, and molec-ular biology, p. 65-117. In A. N. Rai (ed.), CRC handbook ofsymbiotic cyanobacteria. CRC Press, Inc., Boca Raton, Fla.
3. Curtis, S. E., and R. Haselkorn. 1984. Isolation, sequence andexpression of two members of the 32 kd thylakoid membraneprotein gene family from the cyanobacterium Anabaena 7120.Plant Mol. Biol. 3:249-258.
4. Fisher, R., R. Tuli, and R. Haselkorn. 1981. A clone cyanobac-terial gene for glutamine synthetase functions in Escherichiacoli, but the enzyme is not adenylated. Proc. Natl. Acad. Sci.USA 78:3393-3397.
5. Franche, C., and G. Cohen-Bazire. 1985. The structural nifgenes of four symbiotic Anabaena azollae show a highly con-served physical arrangement. Plant Sci. 39:125-131.
6. Franche, C., and G. Cohen-Bazire. 1987. Evolutionary diver-gence in the nifH.D.K. gene region among nine symbioticAnabaena azollae and between Anabaena azollae and somefree-living heterocystous cyanobacteria. Symbiosis 3:159-178.
7. Gates, J. E., R. W. Fisher, T. W. Goggin, and N. I. Azrolan.1980. Antigenic differences between Anabaena azollae freshfrom the Azolla fern leaf cavity and free-living cyanobacteria.Arch. Microbiol. 128:126-129.
8. Golden, J. W., S. J. Robinson, and R. Haselkorn. 1985. Rear-rangement of nitrogen fixation genes during heterocyst differen-tiation in the cyanobacterium Anabaena. Nature (London)314:419-423.
9. Komarek, J., and K. Anagnostidis. 1989. Modem approach tothe classification system of Cyanophytes 4-Nostocales. Arch.Hydrobiol. Suppl. 82:247-345.
10. Ladha, J. K., and I. Watanabe. 1982. Antigenic similarity amongAnabaena azollae separated from different species of Azolla.Biochem. Biophys. Res. Commun. 109:675-682.
11. Lammers, P. J., J. W. Golden, and R. Haselkorn. 1986. Identi-fication and sequence of a gene required for a developmentallyregulated DNA excision in Anabaena. Cell 44:905-911.
12. Lin, C., and I. Watanabe. 1988. A new method for obtainingAnabaena-free Azolla. New Phytol. 108:341-344.
12a.Madan, A. P. 1990. Ph.D. thesis. Rensselaer Polytechnic Insti-tute, Troy, N.Y.
13. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecularcloning: a laboratory manual. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y.
14. McCowen, S. M., L. MacArthur, and J. E. Gates. 1987. Azollafern lectins that specifically recognize endosymbiotic cyanobac-teria. Curr. Microbiol. 14:329-333.
15. Meeks, J. C., C. M. Joseph, and R. Haselkorn. 1988. Organiza-tion of the nif genes in cyanobacteria in symbiotic associationwith Azolla and Anthoceros. Arch. Microbiol. 150:61-71.
16. Newton, J. W., and A. I. Herman. 1979. Isolation of cyanobac-teria from the aquatic fern, Azolla. Arch. Microbiol. 120:161-165.
17. Nierzwicki-Bauer, S. A., S. E. Curtis, and R. Haselkorn. 1984.Cotranscription of genes encoding the small and large subunitsof ribulose-1,5-bisphosphate carboxylase in the cyanobacteriumAnabaena 7120. Proc. Natl. Acad. Sci. USA 81:5961-5965.
18. Nierzwicki-Bauer, S. A., and R. Haselkorn. 1986. Differences inmRNA levels in Anabaena living freely or in symbiotic associ-ation with Azolla. EMBO J. 5:29-35.
19. Peters, G. A., and B. C. Mayne. 1974. The Azolla, AnabaenaAzollae relationship. I. Initial characterization of the associa-tion. Plant Physiol. 53:813-819.
20. Peters, G. A., and J. C. Meeks. 1989. The Azolla-Anabaenasymbiosis: basic biology. Annu. Rev. Plant Physiol. Plant Mol.Biol. 40:193-210.
21. Rice, D., B. J. Mazur, and R. Haselkorn. 1982. Isolation andphysical mapping of nitrogen fixation genes from the cyanobac-terium Anabaena 7120. J. Biol. Chem. 257:13157-13163.
22. Rippka, R., J. Deruelles, J. B. Waterbury, M. Herdman, andR. Y. Stanier. 1979. Genomic assignments, strain histories, andproperties of pure cultures of cyanobacteria. J. Gen. Microbiol.111:1-61.
23. Southern, E. M. 1975. Detection of specific sequences amongDNA fragments separated by gel electrophoresis. J. Mol. Biol.98:503-517.
24. Stanier, R. Y., R. Kunisawa, M. Mandel, and G. Cohen-Bazire.1971. Purification and properties of unicellular blue-green algae(order Chroococcaeles). Bacteriol. Rev. 35:171-205.
25. Subramanian, G., and P. Malliga. 1988. Isolation of Anabaenaazollae from the megasporocarp of Azolla. Curr. Sci. 57:1352-1353.
26. Tel-Or, E., T. Sandovsky, D. Kobiler, C. Arad, and R. Wein-berg. 1983. The unique symbiotic properties of Anabaena in thewater fern Azolla, p. 303-314. In G. C. Papageorgiou and L.Packer (ed.), Photosynthetic prokaryotes: cell differentiationand function. Elsevier Science, New York.
27. Watanabe, I., C. R. Espinas, N. S. Berja, and B. V. Alimagno.1977. Utilization of the Azolla-Anabaena complex as a nitrogenfertilizer for rice. Int. Rice Res. Inst. Res. Paper Ser. No.11:1-15.
27a.White, W. B. 1983. M.S. thesis. Virginia Commonwealth Uni-versity, Richmond.
28. Zimmerman, W. J., B. H. Rosen, and T. A. Lumpkin. 1989.Enzymatic, lectin, and morphological characterization and clas-sification of presumptive cyanobionts from Azolla Lam. NewPhytol. 113:497-503.
APPL. ENVIRON. MICROBIOL.
on April 9, 2020 by guest
http://aem.asm
.org/D
ownloaded from


















